Back to Journals » Journal of Multidisciplinary Healthcare » Volume 18
Potential Causal Relationship Between Plasma and Cerebrospinal Fluid Metabolites and Meningioma: Two-Sample Mendelian Randomization Study
Authors Du G ![]() , Xu D
, Xu D ![]() , Zhao H
, Zhao H ![]() , Yan Z, Wang J
, Yan Z, Wang J
Received 11 March 2025
Accepted for publication 23 July 2025
Published 31 July 2025 Volume 2025:18 Pages 4391—4409
DOI https://doi.org/10.2147/JMDH.S527449
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr David C. Mohr
Guangyu Du, Daikang Xu, Hai Zhao, Zhiyong Yan, Jianpeng Wang
Department of Neurosurgery, The Affiliated Hospital of Qingdao University, Qingdao, Shandong, 266000, People’s Republic of China
Correspondence: Zhiyong Yan, Department of Neurosurgery, The Affiliated Hospital of Qingdao University, No. 59 Haier Road, Qingdao, Shandong, 266000, People’s Republic of China, Email [email protected] Jianpeng Wang, Department of Neurosurgery, The Affiliated Hospital of Qingdao University, No. 59 Haier Road, Qingdao, Shandong, 266000, People’s Republic of China, Email [email protected]
Background: Meningioma (MGM) is the most common benign intracranial tumor and ranks as the second most frequent intracranial tumor in terms of incidence, following malignant gliomas. Studying the metabolites in the serum and cerebrospinal fluid (CSF) of patients with MGM is crucial for understanding the underlying biological mechanisms, identifying new biomarkers, and developing novel therapeutic strategies.
Methods: Mendelian randomization (MR) is a powerful analytical approach that leverages genetic variants to assess potential causal relationships between exposures and outcomes. In this study, MR analysis was used to investigate the causal relationships between 486 serum metabolites, 338 CSF metabolites, and MGM. Our MGM data was derived from the genome wide association studies (GWASs) dataset in the FinnGen database, comprising 1,835 cases of European ancestry and 377,674 controls. The data for 486 plasma metabolites was obtained from the GWAS catalog, and the data for 338 CSF metabolites was obtained from the Wisconsin Alzheimer’s Disease Research Center (WADRC) and Wisconsin Alzheimer’s Disease Prevention Registry (WRAP) study collections. We mainly utilized the Inverse Variance Weighted (IVW) approach to evaluate the causal association between metabolites and MGMs, supplemented by four additional methods to further validate and strengthen our findings. False discovery rate (FDR) correction was applied (q< 0.05) to control the false-positive rate.
Results: Through MR analysis, the study identified 19 plasma metabolites and 17 CSF metabolites demonstrating potential causal associations with MGMs. Among these, 14 metabolites indicated positive causality with MGMs, while 22 metabolites displayed a remarkable negative causality. In particular, plasma levels of Glycerol 3−phosphate (G3P) (OR=4.76, 95% CI=1.02– 22.12, P=0.047) and Valine (OR=0.025, 95% CI=0.0020– 0.42, P=0.010) were found to exhibit the optimal efficacy. Multiple sensitivity analyses confirm the robustness of the results. The study found no evidence of a reverse causality between MGMs and the plasma levels of Glycerol 3−phosphate (G3P) and Valine.
Conclusion: This study identified 36 metabolites associated with the incidence of MGMs, among which Glycerol 3-phosphate (G3P) and Valine are the most notable findings.
Keywords: plasma, cerebrospinal fluid, meningioma, Mendelian randomization, metabolites
Introduction
Meningiomas (MGMs) are among the most common primary central nervous system (CNS) tumor adults.1 The incidence rate is approximately 36.4%, accounting for more than 50% of all benign primary central nervous system tumors. The incidence is higher in females (F:M ratio = 3.38:1) and increases with age, with a significant rise in individuals aged 65 and older. According to the statistical report from the Central Brain Tumor Registry of the United States (CBTRUS, 2009–2010), the prevalence of meningiomas is approximately 97.5 per 100,000. The age-standardized incidence rate of meningiomas (per 100,000) is estimated to be between 1.6 and 5.8 in Europe.2 Therefore, the genome-wide association study (GWAS) data in this study were derived from cohorts of European ancestry to ensure homogeneity in genetic background. In 2021, the World Health Organization (WHO) classified MGMs into three grades based on histopathological features, genetic characteristics, and the risk of progression or recurrence: Grade 1 (benign), Grade 2 (atypical),s in and Grade 3 (malignant or anaplastic). Among these, Grade 1 MGMs are the most prevalent, accounting for over 80% of all cases.3 The primary treatment modalities for MGMs include surgery and radiotherapy, with craniotomy being the standard therapeutic approach. However, complete surgical resection is often unachievable, and post-surgical adverse effects are frequently observed. In cases where cranial nerves are involved, neurological dysfunction is a common complication, significantly exacerbating patient morbidity.
With the rapid advancement of metabolomics, new avenues have emerged for elucidating the pathogenesis of meningiomas and identifying potential biomarkers.4 Previous studies have demonstrated that specific high-abundance MGM metabolites, including Arginyl-Proline, can effectively distinguish metabolic alterations between low-grade and high-grade meningiomas.5 Additionally, GATA-4 has been proposed as a potential therapeutic target through its regulation of miR-497, while miR-497 itself may serve as a promising circulating biomarker for high-grade MGMs.6 Furthermore, a research team has identified unique DNA methylation biomarkers in the plasma of 155 MGM patients, distinguishing them from other CNS tumors.7 Notably, the concentration of extracellular vesicles in plasma has also been found to be useful for meningioma grading and the evaluation of postoperative treatment responses.8 However, due to limited sample sizes and heterogeneity in research methodologies, no clinically validated biomarkers for primary CNS tumors, including MGMs, have been established to date.9 These findings highlight the need to elucidate the causal relationship between cerebrospinal fluid (CSF) and plasma metabolites and MGM development.
CSF and plasma are both critical biological compartments that reflect systemic and localized metabolic changes, providing complementary insights into tumor biology. CSF is particularly valuable for exploring molecular alterations at the tumor site, while plasma serves as a readily accessible source for investigating systemic metabolic disruptions. By integrating data from both compartments, we can achieve a more comprehensive understanding of MGM pathophysiology, which is crucial for identifying reliable biomarkers and potential therapeutic targets.
Mendelian randomization (MR) is a powerful analytical approach that leverages genetic variations to assess potential causal relationships between exposures and outcomes. Compared to traditional randomized controlled trials (RCTs), MR is not constrained by factors such as cost, time, or ethical concerns associated with interventional studies. Additionally, it effectively minimizes confounding bias and reduces the risk of reverse causality.10 Therefore, our study aims to apply MR analysis to investigate which metabolites in CSF and plasma may have potential causal relationships with the occurrence and progression of MGMs.
Materials and Methods
Study Design
To ensure methodological rigor, this study was conducted in accordance with the Strengthening the Reporting of Observational Studies in Epidemiology using Mendelian Randomization (STROBE-MR, a guideline designed to strengthen the reporting standards for Mendelian randomization studies) guidelines.11 We strictly adhered to the three fundamental instrumental variables (IVs) assumptions in MR analysis:
- The genetic variations of the selected IVs must robustly represent the exposures of interest, which in this study are serum and CSF metabolites.
- The IVs must be independent of confounding factors.
- The IVs must influence the occurrence of meningiomas exclusively through the exposure factors (Figure 1).
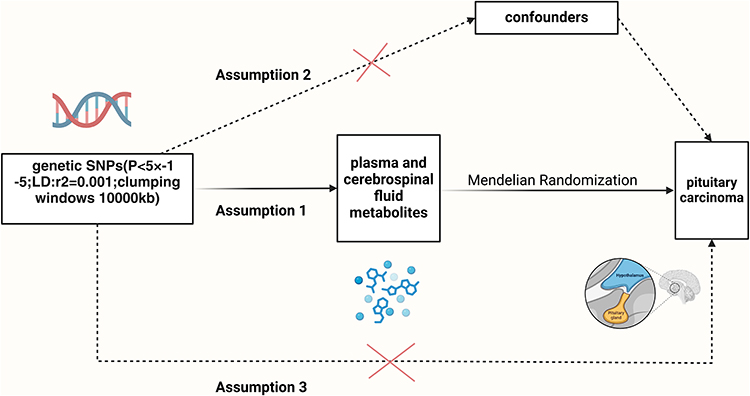 |
Figure 1 Schematic illustration of the study design. SNPs, single nucleotide polymorphisms. Created with BioRender.com. |
CSF Metabolites Data Source
The dataset for CSF metabolites was derived from the study by Panyard et al, which focused on metabolomics research. The genome-wide association study (GWAS) dataset for CSF metabolites was obtained from CSF samples of 689 participants, including 532 individuals from the Wisconsin Alzheimer’s Disease Research Center (WADRC) and 168 individuals from the Wisconsin Alzheimer’s Disease Prevention Registry (WRAP) (After excluding individual data with missing values and data with duplicate counts, the number of participants available for further analysis was 689). Of the 338 CSF metabolites analyzed, 296 have been chemically validated, while 38 remain uncharacterized. The study received approval from the Institutional Ethics Review Board of the University of Wisconsin Health Sciences.12
Serum Metabolites Data Source
The dataset for serum metabolites was obtained from the study conducted by So-Youn Shin et al, which included 7,824 adult individuals from two large European population cohorts: the German KORA F4 study (n = 1,768 participants) and the UK TwinsUK study (n = 6,056 participants). Among the 486 serum metabolites analyzed, 309 have been chemically validated, while 177 remain uncharacterized.13
MGM Data Source
The GWAS dataset for MGMs was obtained from the Round 12 version of the FinnGen biobank documentation.14 This dataset consists of 1,835 MGM patients of European ancestry and 379,509 European control participants, encompassing a total of 20,111,133 single nucleotide polymorphisms (SNPs).
Selection of Instrumental Variables
SNPs are commonly used genetic variations in MR analysis, reflecting DNA sequence diversity caused by single nucleotide changes at the genomic level. In our study, we constructed a unique set of instrumental variables for each of the CSF and plasma metabolites. This approach was adopted to ensure the accurate representation of the genetic associations specific to each metabolite. Instrumental variables were selected based on the following criteria:
- Considering the limited number of SNPs available to achieve genome-wide significance for metabolites, we relaxed the significance threshold to P < 1×10−5 to increase statistical power. However, this raised concerns about the potential influence of weak-instruments.
- To eliminate linkage disequilibrium, an r²<0.001 threshold and a genetic distance of 10,000 kb were set, ensuring independence among selected SNPs.
- The F-statistic for each SNP was calculated, and SNPs with F-values<10 were excluded to avoid weak instrument bias. Only SNPs with F-values>10 were retained.
- SNPs located within palindromic sequences were removed. To ensure the alignment of alleles across exposure and outcome datasets, we employed the harmonise function from the TwoSampleMR R package. This function systematically compares the effect and non-effect alleles for each SNP in both datasets and harmonizes them to a common reference strand. Specifically, it flips the sign of the beta coefficient when alleles are aligned in opposite directions and removes SNPs with ambiguous alignment that cannot be resolved reliably. For palindromic SNPs (ie, A/T or G/C variants), which are prone to strand ambiguity, the function excludes those with intermediate allele frequencies (typically between 0.42 and 0.58). This harmonization process is crucial for avoiding misclassification of effect directions.
- SNPs associated with potential confounding factors were excluded.
Statistical Analysis
All statistical analyses were performed using the TwoSampleMR R package (Version 0.5.8) in R-Studio (Version 4.4.1).15 The Inverse Variance Weighted (IVW) method has been reported as the most effective approach for detecting causal effects between exposures and outcomes in MR studies.16 Therefore, IVW was chosen as the primary analytical method. Additionally, MR-Egger, weighted mode, weighted median, and simple mode methods were applied as supplementary analyses to ensure robustness. However, when fewer than 10 instrumental variables were available, the results from the MR-Egger analysis exhibited a degree of unreliability. A P-value < 0.05 was set as the threshold for statistical significance. Odds ratios (ORs) were used to quantify the effect of CSF and serum metabolites on MGM risk. To effectively control the false positive rate arising from multiple testing, false discovery rate (FDR) correction was applied to enhance the accuracy and reliability of the findings. The Benjamini–Hochberg method was utilized to apply a FDR for the results.
Sensitivity Analyses
To assess the robustness of the findings, various sensitivity analyses were conducted on the significant variants. Given that weak instrumental variables were used in this study to conduct the Mendelian randomization (MR) analysis, we considered performing a sensitivity analysis using only genome-wide significant variants. Cochran’s Q test was used to evaluate heterogeneity, with a P-value <0.05 indicating significant heterogeneity.17 Potential horizontal pleiotropy was assessed using MR-Egger regression by testing for a non-zero intercept (P<0.05) and by identifying outliers using the MR-PRESSO method.18,19 Additionally, a leave-one-out analysis was performed to evaluate the potential influence of individual SNPs on the results and to mitigate bias20 (Figure 1).
Results
Mendelian Randomization Analysis of 486 Plasma Metabolites and Their Impact on Meningiomas
A total of 19 metabolites were identified to exhibit putative causal associations (IVW p < 0.05) (Table 1). Specifically, plasma levels of Glycerol 3-phosphate (G3P) (OR = 4.759, 95% CI = 1.024 − 22.116, P = 0.0466), 1-stearoylglycerophosphocholine (OR = 4.752, 95% CI = 1.314 − 17.186, P = 0.0175), Eicosapentaenoate (EPA; 20:5n3) (OR = 2.948, 95% CI = 1.119 − 7.770, P = 0.0287), and Butyrylcarnitine (OR = 1.436, 95% CI = 1.009–2.045, P = 0.0445) were positively correlated with the risk of MGMs.
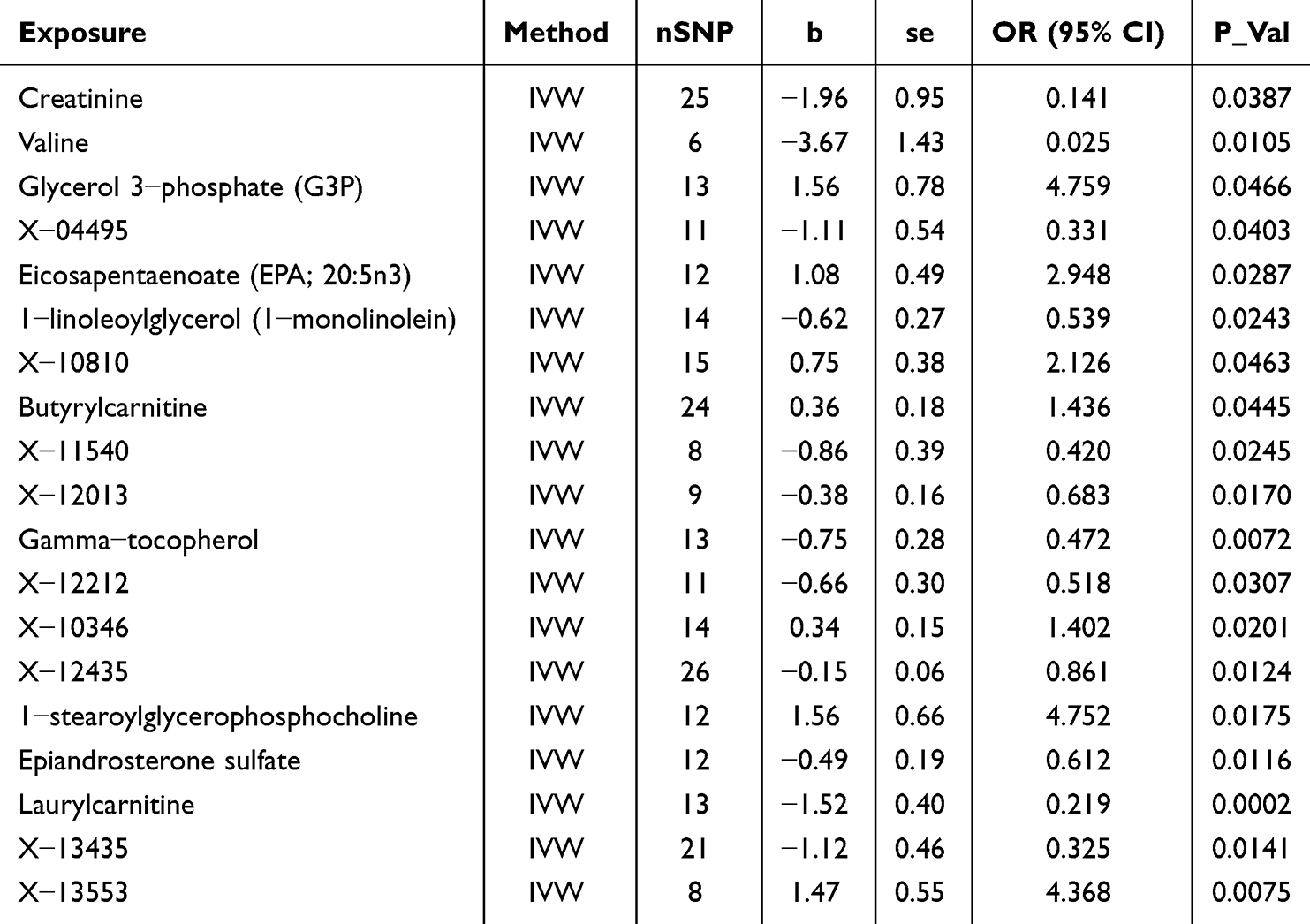 |
Table 1 The Significant IVW Results of Plasma Metabolites and MGMs |
Conversely, several plasma metabolites exhibited negative correlations with MGM risk, including Epiandrosterone sulfate (OR = 0.612, 95% CI = 0.418–0.896, P = 0.0116), 1-linoleoylglycerol (1-monolinolein) (OR = 0.539, 95% CI = 0.315–0.923, P = 0.0243), Gamma-tocopherol (OR = 0.472, 95% CI = 0.273–0.816, P = 0.0072), Laurylcarnitine (OR = 0.219, 95% CI = 0.099–0.482, P = 0.0002), Creatinine (OR = 0.141, 95% CI = 0.022–0.903, P = 0.0387), and Valine (OR = 0.025, 95% CI = 0.002–0.423, P = 0.0105).
Additionally, nine unidentified metabolites exhibited a causal relationship with MGM occurrence (Figures 2–4). While, after FDR correction, no associations between metabolites and meningioma remained statistically significant (q < 0.05). Scatter plots were generated to visualize these causal associations (Figure 5). Moreover, reverse MR analysis confirmed that no causal relationship exists from MGMs to these metabolites (P < 0.05).
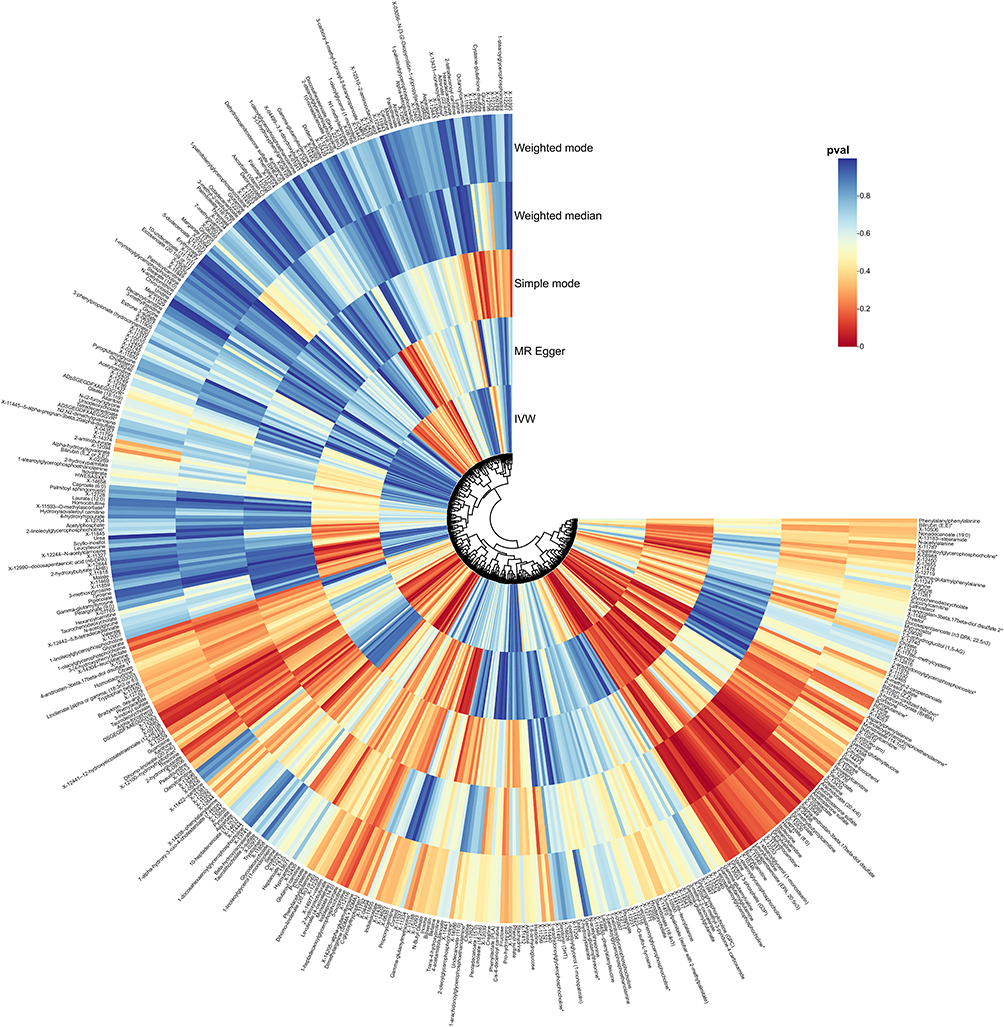 |
Figure 2 Summary of causal estimates regarding the impact of all plasma metabolites on MGMs in MR analysis. From outside to inside, the corresponding P-values of Weighted mode, Weighted median, Simple mode, MR Egger, IVW are represented, respectively. Abbreviations: MR, Mendelian randomization; IVW, inverse-variance weighted. |
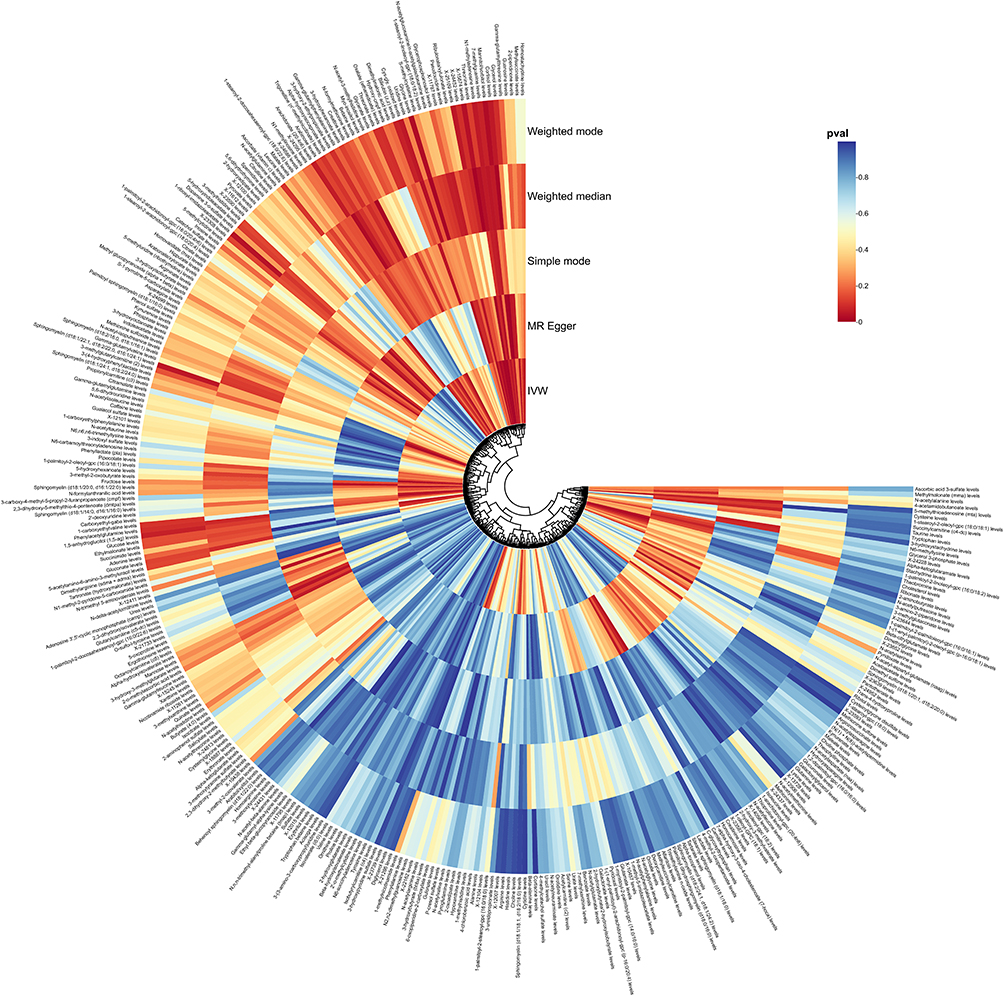 |
Figure 3 Summary of causal estimates regarding the impact of all CSF metabolites on MGMs in MR analysis. From outside to inside, the corresponding P-values of Weighted mode, Weighted median, Simple mode, MR Egger, IVW are represented, respectively. Abbreviations: MR, Mendelian randomization; IVW, inverse-variance weighted. |
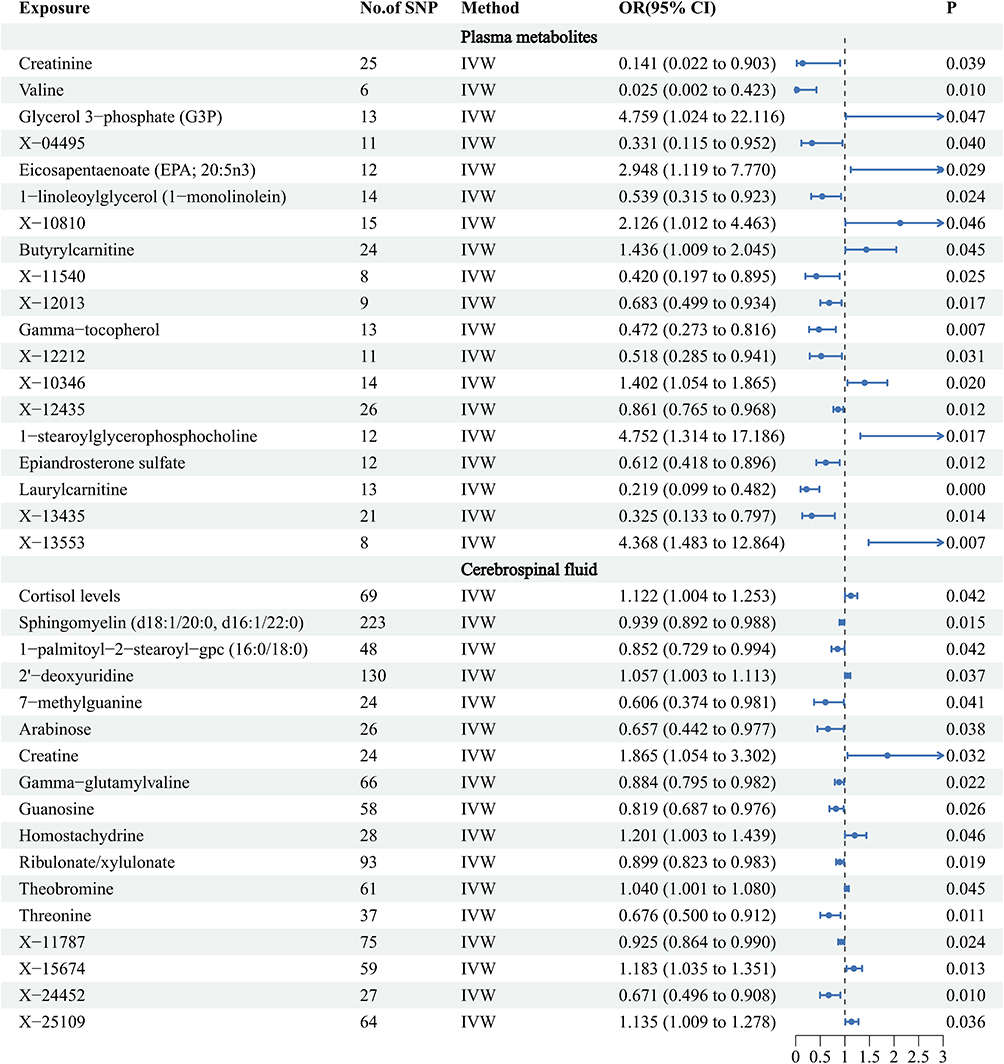 |
Figure 4 Forest plots illustrating the significant causal estimates of plasma and CSF metabolites on MGMs are presented. Abbreviations: SNPs, single nucleotide polymorphisms; OR, odds ratio; IVW, inverse-variance weighted. |
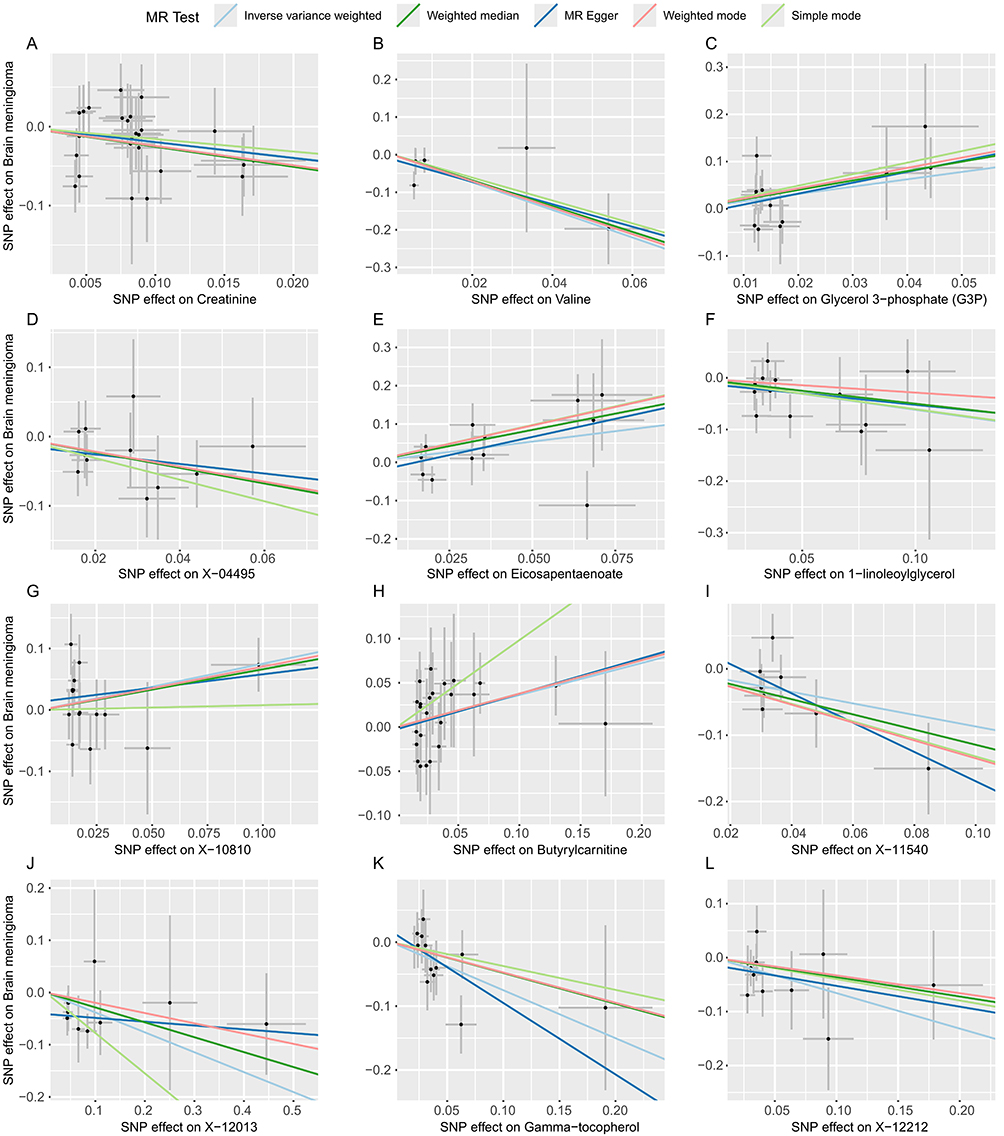 |
Figure 5 Continued. |
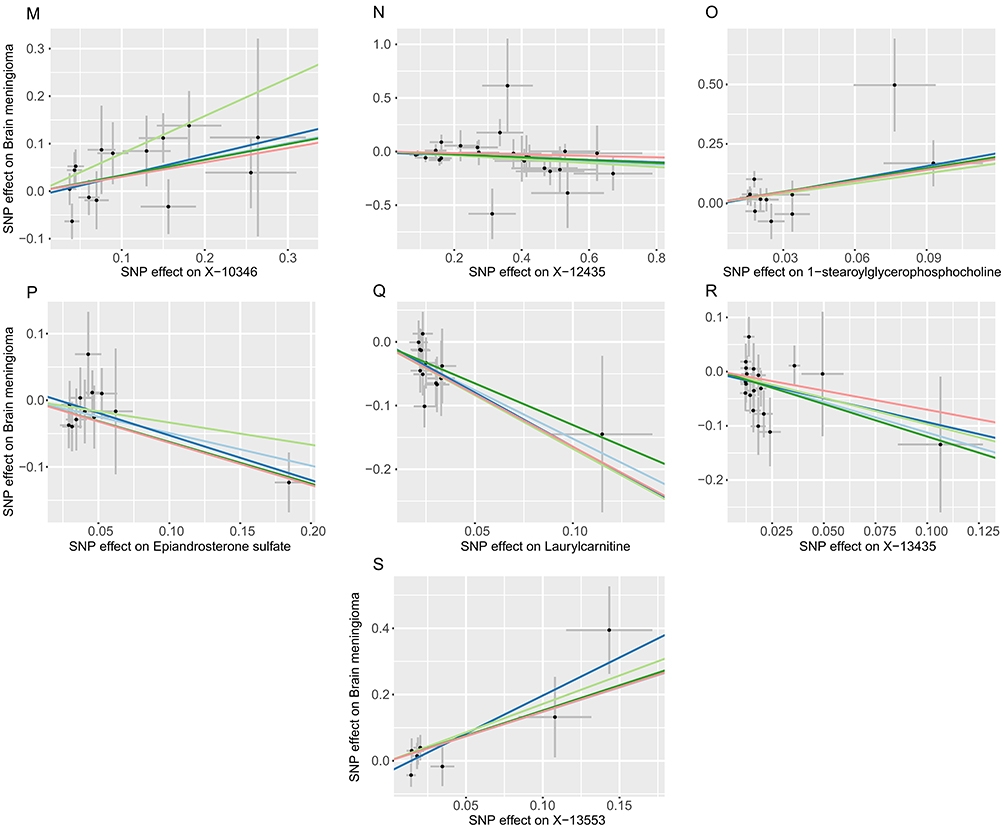 |
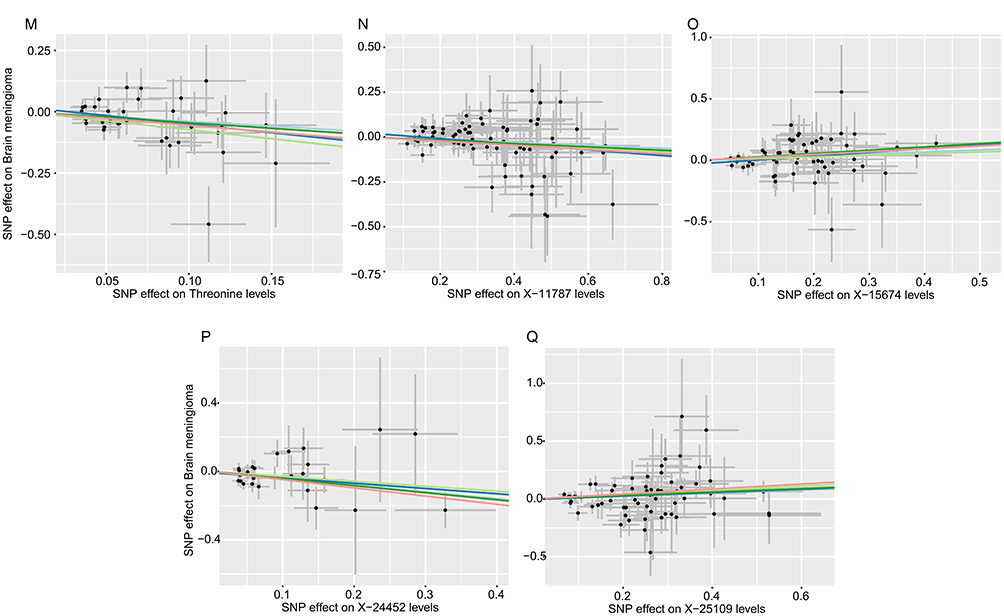
Figure 5 Scatter plots for the causal association between plasma metabolites and MGMs.(A) Creatinine (B) Valine (C) Glycerol 3−phosphate (G3P) (D) X−04495 (E) Eicosapentaenoate (F) 1−linoleoylglycerol (G) X−10810 (H) Butyrylcarnitine (I) X−11540 (J) X−12013 (K) Gamma−tocopherol (L) X−12212 (M) X−10346 (N) X−12435 (O) 1−stearoylglycerophosphocholine (P) Epiandrosterone sulfate (Q) Laurylcarnitine (R) X−13435 (S) X−13553. |
Mendelian Randomization Analysis of 338 CSF Metabolites and Their Impact on Meningiomas
A total of 17 metabolites were identified to exhibit putative causal associations (IVW p < 0.05) (Table 2). Notably, Creatine (OR = 1.865, 95% CI = 1.054–3.302, P = 0.0324), Homostachydrine (OR = 1.201, 95% CI = 1.003–1.439, P = 0.0465), Cortisol (OR = 1.122, 95% CI = 1.004–1.253, P = 0.0422), 2’-deoxyuridine (OR = 1.057, 95% CI = 1.003–1.113, P = 0.0369), and Theobromine (OR = 1.040, 95% CI = 1.001–1.080, P = 0.0453) were positively correlated with MGM risk.
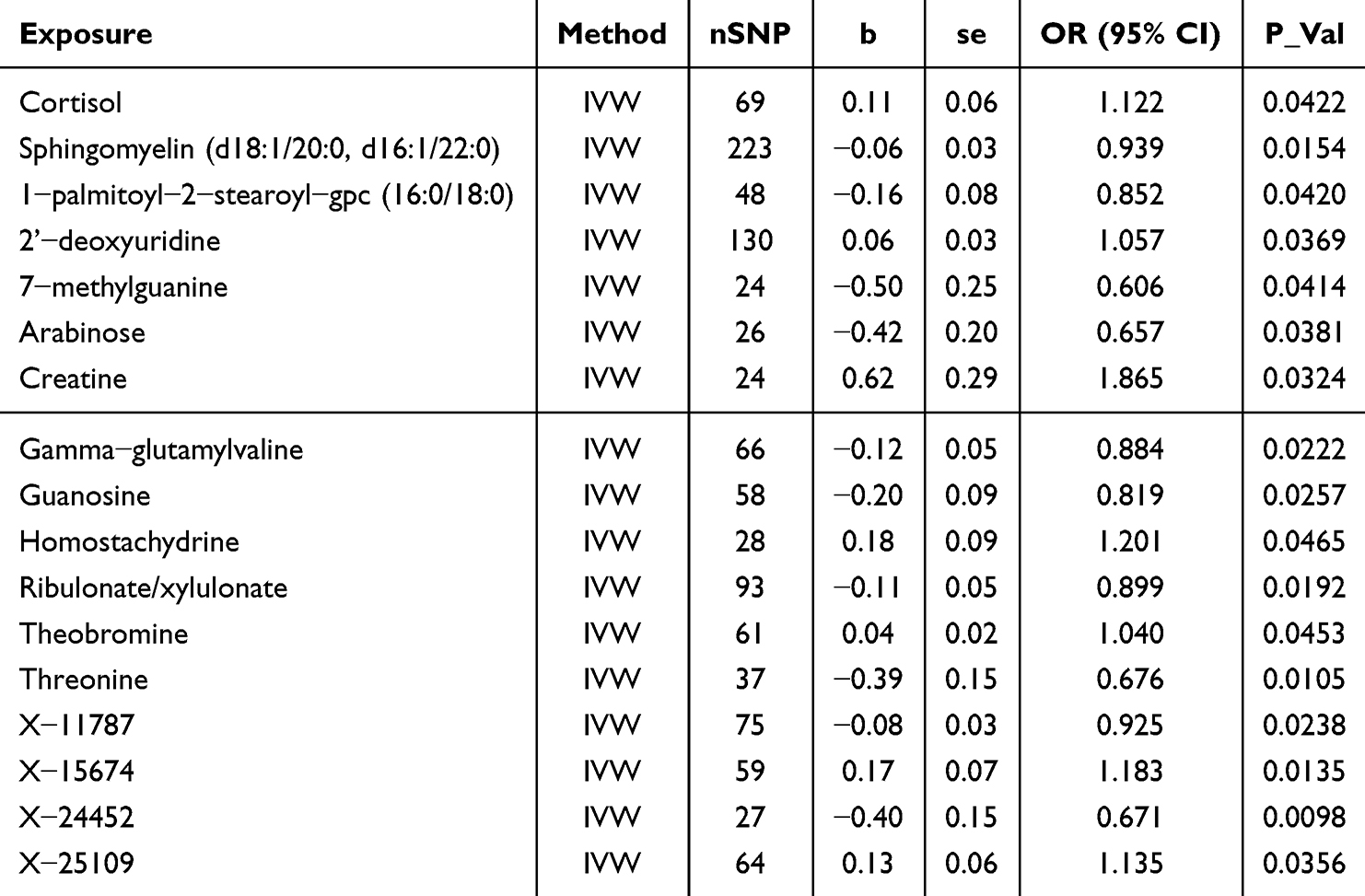 |
Table 2 The Significant IVW Results of CSF Metabolites and MGMs |
Conversely, several CSF metabolites exhibited negative correlations with MGM risk, including Sphingomyelin (d18:1/20:0, d16:1/22:0) (OR = 0.939, 95% CI = 0.892–0.988, P = 0.0154), Ribulonate/xylulonate (OR = 0.899, 95% CI = 0.823–0.983, P = 0.0192), Gamma-glutamylvaline (OR = 0.884, 95% CI = 0.795–0.982, P = 0.0222), 1-palmitoyl-2-stearoyl-gpc (16:0/18:0) (OR = 0.852, 95% CI = 0.729–0.994, P = 0.0420), Guanosine (OR = 0.819, 95% CI = 0.687–0.976, P = 0.0257), Threonine (OR = 0.676, 95% CI = 0.500–0.912, P = 0.0105), Arabinose (OR = 0.657, 95% CI = 0.442–0.977, P = 0.0381), and 7-methylguanine (OR = 0.606, 95% CI = 0.374–0.981, P = 0.0414).
Additionally, four unidentified metabolites demonstrated causal associations with MGMs (Figures 3 and 4). Scatter plots were generated to illustrate these relationships (Figure 6). While, after FDR correction, no associations between metabolites and meningioma remained statistically significant (q < 0.05). Reverse MR analysis confirmed that no causal relationship exists from MGMs to these metabolites (P < 0.05). Figure 6 Continued. Figure 6 Scatter plots for the causal association between cerebrospinal fluid metabolites and MGMs.(A) Cortisol (B) Sphingomyelin (C) 1−palmitoyl−2−stearoyl−gpc (D) 2ʹ−deoxyuridine (E) 2ʹ−deoxyuridine (F) Arabinose (G) Creatine (H) Gamma−glutamylvaline (I) Guanosine (J) Homostachydrine (K) Ribulonate/xylulonate (L) Theobromine (M) Threonine (N) X−11787 (O) X−15674 (P) X−24452 (Q) X−25109 and IVW results of plasma metabolites and MGMs.
Sensitivity Analyses
To ensure the robustness of our findings, several sensitivity analyses were conducted. The MR-Egger intercept test was performed to assess the presence of pleiotropy, while Cochran’s Q test was used to evaluate heterogeneity.
For plasma metabolites and MGMs, no evidence of heterogeneity or pleiotropy was detected among the 19 plasma metabolites.
For CSF metabolites and MGMs, with the exception of Sphingomyelin (d18:1/20:0, d16:1/22:0), no significant heterogeneity was observed in the other 16 CSF metabolites. Additionally, except for Cortisol, no significant pleiotropy was detected among the remaining CSF metabolites.
Furthermore, leave-one-out analysis identified no potential outliers, and after outlier correction, MR-PRESSO analysis did not yield any additional abnormal instrumental variables. These results collectively reinforce the robustness and reliability of our findings (Tables 3 and 4), while illustrations of leave-one-out analysis and funnel plots can be discovered in Supplementary Figure S1–S6.
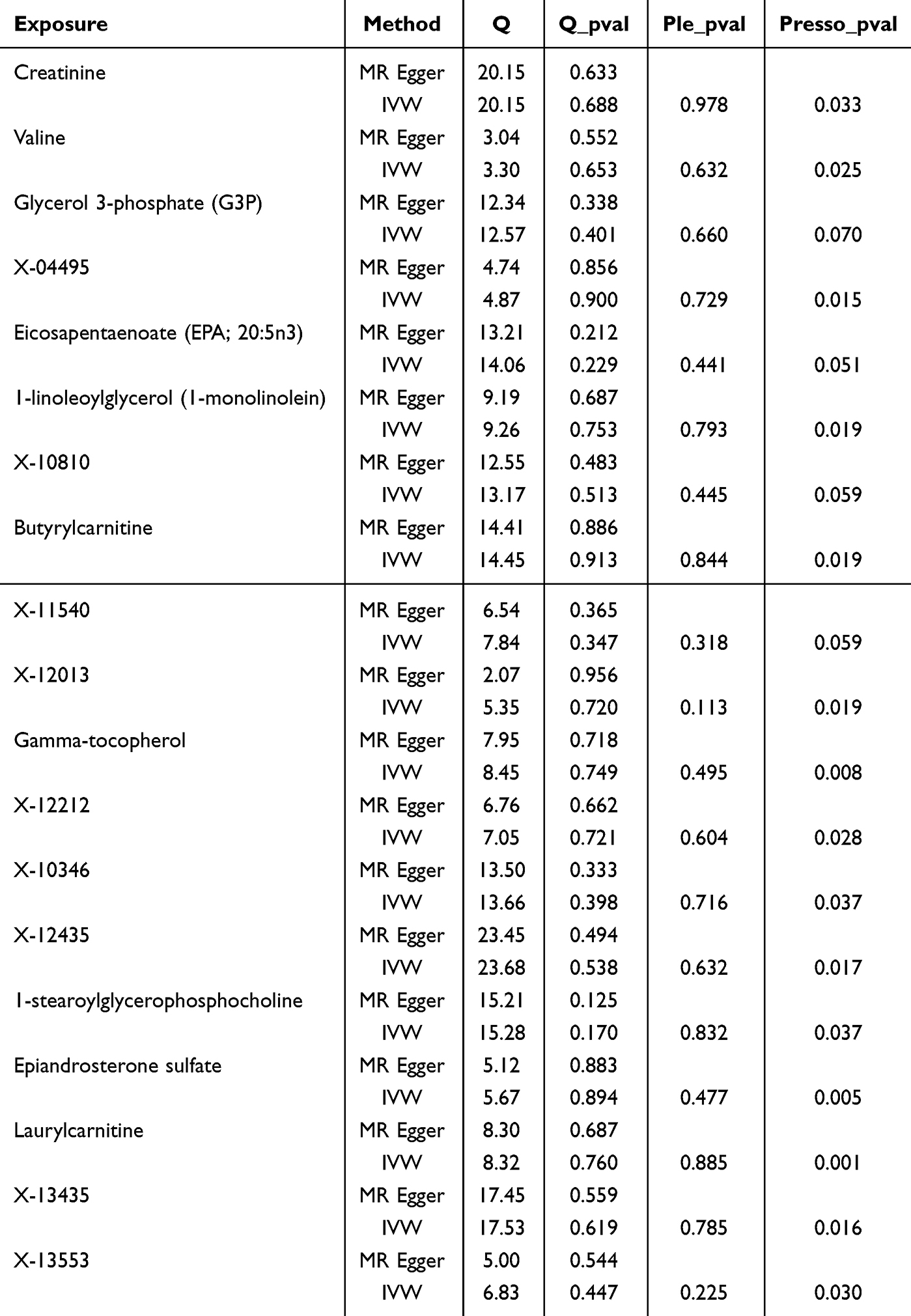 |
Table 3 The Sensitivity Analysis Results of Plasma Metabolites and MGMs |
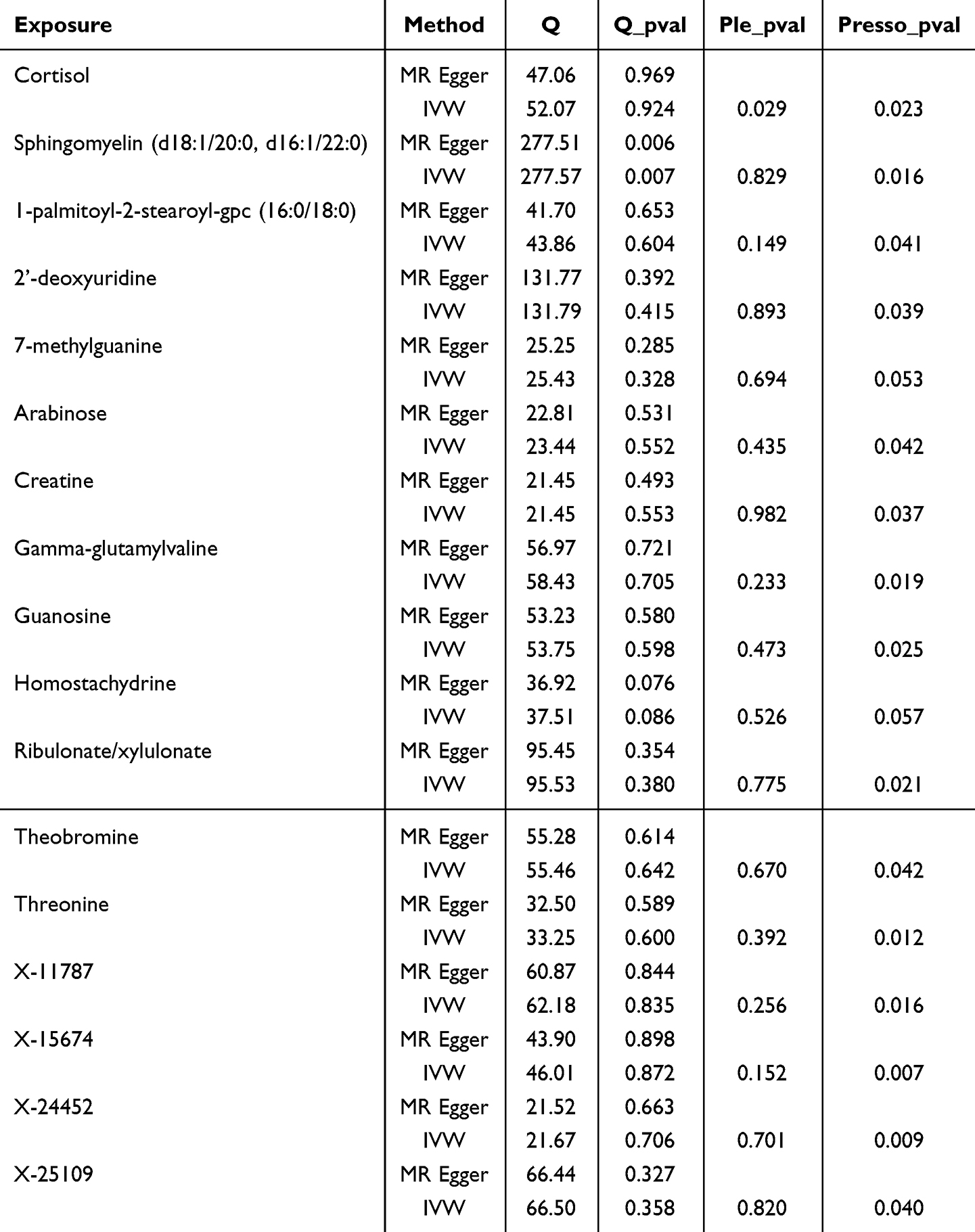 |
Table 4 The Sensitivity Analysis Results of CSF Metabolites and MGM |
Discussion
To elucidate the potential causal relationship between various metabolites and the occurrence of MGMs, we conducted a MR analysis utilizing serum and CSF metabolite datasets alongside MGM genetic data. Our study identified 17 CSF metabolites and 19 plasma metabolites associated with MGM occurrence. Among these, 14 metabolites exhibited a positive correlation, while 22 metabolites showed a negative correlation with MGMs. Among these metabolites, 13 remained unidentified (“X-”), a phenomenon attributed to the presence of features labeled as “ unidentified metabolites” in the metabolomic dataset. In the metabolomics datasets, certain features were labeled as “unidentified metabolites”, denoting signals detected via mass spectrometry that lacked confirmed structural annotation at the time of data release. These entries were retained in the analysis to preserve potentially informative biological signals. These findings provide valuable insights into potential novel biomarkers for MGM diagnosis and prognosis.
The IV method is a statistical approach designed to address endogeneity issues, particularly in regression analyses, by leveraging external variables—termed IVs—to infer causal relationships.21 The IVW method, a widely adopted statistical approach in IV analysis, is commonly employed for the combined analysis of multiple instrumental variables to enhance the reliability of causal inference.
The MR framework is based on Mendel’s second law, which posits that alleles are randomly distributed during meiosis. This method exploits genetic variations, such as SNPs, as IVs for exposure factors that are indezatiopendent of outcome risk, unaffected by confounding factors, and not influenced by acquired characteristics. This enables robust causal inference between exposure variables and health outcomes.22,23
Although several treatment modalities exist for CNS tumors, they are often accompanied by substantial limitations. Identifying biomarkers for early diagnosis and prognosis assessment is therefore of paramount importance. Advances in metabolomics research within neurosurgery have provided novel insights and methodologies for biomarker discovery.24 However, metabolomics-based research on MGMs remains in its infancy. Nonetheless, accumulating evidence underscores the promising potential of metabolomics in elucidating the mechanisms underlying MGMs, as well as in their clinical staging and prognostic evaluation.4
Metabolomics and the Pathophysiology of Meningiomas
Recent metabolomics researches had yielded crucial insights into the interplay between metabolic pathways and the biological behavior of MGMs. The anaplerotic role of glutamine facilitates the replenishment of tricarboxylic acid (TCA) cycle intermediates, supporting amino acid synthesis and thereby influencing tumor cell growth and survival. This pathway appears particularly critical in high-grade MGMs.25
Advanced analytical approaches, such as liquid chromatography-high resolution mass spectrometry (LC-HRMS) and machine learning algorithms, have enabled the effective differentiation between aggressive and low-grade tumors based on their metabolic and lipidomic profiles.26 Additionally, High-Resolution Magic Angle Spinning Nuclear Magnetic Resonance (HR-MAS NMR) has been instrumental in distinguishing metabolic alterations across different meningioma grades, with metabolites such as glutamate and alanine emerging as key determinants of tumor invasiveness and therapeutic response.27
Collectively, these findings indicate that metabolic reprogramming plays a pivotal role in MGM progression, with metabolic profiles serving as biomarkers for tumor aggressiveness, aiding in prognostic prediction and therapeutic decision-making. Notably, as tumor grade increases, the clinical utility of metabolic alterations becomes increasingly pronounced.28
Glycerol-3-Phosphate (G3P) as Potential Biomarkers
In our MR analysis results, G3P exhibited the highest OR (4.759) among all plasma and CSF metabolites associated with MGM occurrence, warranting further investigation. G3P, a central metabolic intermediate, participates in glycolysis and gluconeogenesis, two essential pathways for cellular energy homeostasis. Dysregulated glucose metabolism is a hallmark of meningiomas, often manifesting as significantly enhanced glycolysis to support the rapid proliferation of tumor cells.28
Although most studies on G3P have focused on renal metabolism, emerging evidence suggests that G3P is involved in multiple metabolic pathways, including NAD+ regeneration, oxidative stress regulation, and fatty acid metabolism. In chronic kidney disease (CKD), G3P enhances gluconeogenesis, mitigates oxidative stress-induced cell damage, and aids in restoring cellular energy balance.29,30 Additionally, G3P modulates phosphate metabolism by regulating FGF23 synthesis, which in turn affects vascular calcification and phosphate reabsorption.31,32
Intriguingly, the cross-regulation between glycolysis and phosphate metabolism is particularly relevant in MGMs. Lactate, a key glycolytic product, is known to regulate renal phosphate transporters, influencing phosphate homeostasis.33 Similarly, during aerobic glycolysis (Warburg effect), lactate accumulation may promote malignant transformation in MGMs, allowing tumors to adapt to hypoxic conditions.27 Recent metabolomic analyses have linked hyperphosphatemia to MGM progression and calcification, highlighting the importance of phosphate metabolism in tumor biology.25
Amino Acid Metabolism and Meningiomas
The reprogramming of amino acid metabolism has emerged as a major focus in cancer research. Amino acid dysregulation plays a critical role in tumor initiation and progression, with diagnostic, prognostic, and therapeutic implications.
Our MR analysis revealed that valine exhibited the lowest OR (0.025) among both plasma and CSF metabolites, indicating a negative association with MGM occurrence. In contrast, creatine demonstrated the highest OR (1.865) among CSF metabolites, suggesting a positive correlation with MGM risk. Retrospective analysis of 62 MGM samples using HRMAS spectroscopy revealed that high-grade MGMs (WHO grade II and III) are associated with lower valine concentrations, which correlate with poorer histopathological prognostic markers.27
Similarly, creatine, a key metabolite in energy metabolism, has been identified as a potential CSF biomarker for MGMs. Creatine levels are significantly associated with MGM presence, with plasma creatine and creatinine levels demonstrating strong discriminatory power in distinguishing MGM patients from healthy individuals.34 Postoperative metabolic profiling reveals elevated glutamine-to-creatinine ratios following extensive tumor resections,35 while CSF ribulonate/xylulonate and arabinose levels demonstrate negative correlations with MGM development - patterns mirroring hepatocellular carcinoma metabolomes.36
Lipid Metabolism and Meningiomas
Lipid metabolic dysregulation is implicated in the pathogenesis of multiple cancers, including MGMs.37 Our MR analysis identified four lipid metabolites with causal relationships to MGM occurrence. Plasma 1-stearoylglycerophosphocholine and eicosapentaenoate (EPA; 20:5n3) were positively correlated, whereas plasma 1-linoleoylglycerol (1-monolinolein) and CSF 1-palmitoyl-2-stearoyl-glycerophosphocholine (16:0/18:0) were negatively correlated with MGM occurrence.
Interestingly, EPA exhibits neuroprotective and anti-tumor properties,38,39 but its susceptibility to oxidative stress may compromise these effects. Vitamin E, a potent antioxidant, may help preserve EPA’s biological activity, thereby modulating its role in tumor biology. Our MR analysis identified plasma gamma-tocopherol levels as inversely associated with MGM occurrence, further supporting this hypothesis.
Nucleotide Metabolism and Meningiomas
Nucleotide metabolism dysregulation emerges through CSF biomarkers: 2’-deoxyuridine shows positive correlation, contrasting with negative associations for guanosine, threonine, and 7-methylguanine. The proliferative marker 5-ethynyl-2’-deoxyuridine (EdU) derivatives and guanosine catabolites like neopterin warrant further investigation.40 Threonine-mediated tRNA modification mechanisms observed in glioblastoma suggest parallel metabolic reprogramming in meningiomas.41
Peptide Metabolism and Meningiomas
Peptide metabolism alterations manifest through plasma epiandrosterone sulfate (inverse correlation) and CSF homostachydrine/cortisol (positive correlations). The sexual dimorphism in meningioma incidence and cortisol’s modulation of steroid hormone receptors suggest endocrine-metabolic crosstalk in tumorigenesis.42 Acylcarnitine profiling reveals contrasting associations for short-chain (butyrylcarnitine, positive) versus long-chain (laurylcarnitine, negative) species, implicating fatty acid metabolism in MGM pathophysiology.
Study Limitations
While our study provides novel insights into metabolite-associated MGM pathogenesis, several limitations must be acknowledged.
- Population Specific Bias – Our MR analysis was conducted using GWAS data exclusively from European cohorts, potentially limiting generalizability across ethnic groups.
- Lack of Experimental Validation – The causal relationships identified require further validation through experimental and clinical studies.
- Statistical Adjustments – As an exploratory study, multiple testing adjustments were limited, although we employed multiple MR methodologies to ensure robustness.
An important limitation of the two-sample MR analysis conducted herein lies in the pronounced sample size disparity between the exposure and outcome datasets (689/7,824: 379,509). As a result, we applied a more relaxed threshold and used weaker instruments for the analysis (P<1×10−5). This imbalance may introduce multiple methodological concerns.
Firstly, the difference in sample sizes may inflate the FDR, as the large outcome sample size amplifies the significance of instrument bias, thereby significantly increasing the probability of type I errors and potentially reducing the likelihood of type II errors.
Secondly, the relatively small sample size of the exposure dataset may lead to the “winner’s curse” phenomenon—due to sampling variability, the effect sizes of genetic variants associated with the exposure may be overestimated. This issue is particularly evident when the exposure GWAS lacks sufficient statistical power to robustly detect true associations. The winner’s curse can further inflate the instrument-exposure association, thereby exacerbating the bias in MR causal effect estimates, especially when the outcome sample size is large.
Thirdly, despite implementing sensitivity analyses such as MR-Egger regression and MR-PRESSO, the imbalance in sample sizes may exacerbate residual pleiotropy, where genetic instruments influence the outcome through non-target exposure pathways, leading to confounding effects. Although these biases were assessed and mitigated through sensitivity analyses, the risk of overestimated causal effects and potential residual pleiotropy due to unequal sample sizes cannot be entirely ruled out.
Therefore, due to the presence of these potential sources of bias, the Mendelian randomization approach may not completely rule out the possibility of vertical pleiotropy or violations of the exclusion restriction assumption. Consequently, interpretations of the results should be approached with caution, particularly for metabolites exhibiting notably strong OR, as further in-depth experimental validation is warranted to clarify their true biological relevance and research potential.
Conclusions
In this study, we investigated the potential causal relationships between metabolites in cerebrospinal fluid and plasma and the development of meningioma. Provides evidence consistent with a causal role for the identified metabolites. These findings aim to further advance the application of metabolomics in elucidating meningioma the pathogenic mechanisms of meningioma and underscore the potential utility of plasma and CSF metabolites as valuable diagnostic and prognostic indicators.
This study provides novel insights into the metabolic landscape of meningiomas, providing indicative value for early detection, tumor grading, and prognosis assessment. Future investigations, including large-scale validation studies and mechanistic explorations, are warranted to confirm these findings and facilitate their translation into clinical applications.
Abbreviations
MGM, Meningioma; CNS, Central nervous system; WHO, World Health Organization; CSF, Cerebrospinal fluid; MR, Mendelian randomization; RCT, Randomized controlled trial; IV, Instrumental variable; IVW, Inverse Variance Weighted, GWAS, Genome-wide association study; SNP, Single nucleotide polymorphism; ORs, Odds ratios; G3P, Glycerol-3-phosphate; TCA, Tricarboxylic acid; LC-HRMS, Liquid chromatography-high resolution mass spectrometry; HR-MAS NMR, High-Resolution Magic Angle Spinning Nuclear Magnetic Resonance; CKD, Chronic kidney disease; EdU, 5-ethynyl-2’-deoxyuridine; STROBE-MR, Strengthening the Reporting of Observational Studies in Epidemiology using Mendelian Randomization.
Data Sharing Statement
The original contributions presented in the study are included in the article/Supplementary Material. Further inquiries can be directed to the corresponding author.
Ethics Approval and Consent to Participate
The study protocol was reviewed and approved by the Medical Ethics Committee of the Affiliated Hospital of Qingdao University (Approval No. QYFYwzll30212).
Acknowledgments
We sincerely thank the investigators and participants of the original GWAS datasets used in this study. We are grateful for all GWAS sharing summary data used in this study.
Funding
This research was funded by the China Postdoctoral Science Foundation funded project (grant no. 2021M691696).
Disclosure
The authors declare no competing interests.
References
1. Ostrom QT, Gittleman H, Xu J, et al. CBTRUS statistical report: primary brain and other central nervous system tumors diagnosed in the United States in 2009–2013. Neuro-Oncology. 2016;18(suppl_5):v1–v75. doi:10.1093/neuonc/now207
2. Nyalundja AD, Mugisha F, Karekezi C. The natural history and treatment of meningiomas: an update. Seminars Neurol. 2024;44(1):1–15. doi:10.1055/s-0043-1777352
3. Gritsch S, Batchelor TT, Gonzalez Castro LN. Diagnostic, therapeutic, and prognostic implications of the 2021 world health organization classification of tumors of the central nervous system. Cancer. 2021;128(1):47–58. doi:10.1002/cncr.33918
4. Halabi R, Dakroub F, Haider MZ, et al. Unveiling a biomarker signature of meningioma: the need for a panel of genomic, epigenetic, proteomic, and RNA biomarkers to advance diagnosis and prognosis. Cancers. 2023;15(22):5339. doi:10.3390/cancers15225339
5. Kurokawa GA, Hamamoto Filho PT, Delafiori J, et al. Differential plasma metabolites between high- and low-grade meningioma cases. Int J Mol Sci. 2022;24(1):394. doi:10.3390/ijms24010394
6. Negroni C, Hilton DA, Ercolano E, et al. GATA-4, a potential novel therapeutic target for high-grade meningioma, regulates miR-497, a potential novel circulating biomarker for high-grade meningioma. eBioMedicine. 2020;59.
7. Herrgott GA, Snyder JM, She R, et al. Detection of diagnostic and prognostic methylation-based signatures in liquid biopsy specimens from patients with meningiomas. Nat Commun. 2023;14(1). doi:10.1038/s41467-023-41434-z
8. Ricklefs FL, Maire CL, Wollmann K, et al. Diagnostic potential of extracellular vesicles in meningioma patients. Neuro-Oncology. 2022;24(12):2078–2090. doi:10.1093/neuonc/noac127
9. Rudà R, Pellerino A, Soffietti R. Blood and cerebrospinal fluid biomarkers in neuro-oncology. Curr Opinion Neurol. 2024;37(6):693–701. doi:10.1097/WCO.0000000000001317
10. Connor A, Davk E, Kathiresan S. Mendelian randomization. JAMA. 2017;318(19):1925–1926. doi:10.1001/jama.2017.17219
11. Skrivankova VW, Richmond RC, Woolf BAR, et al. Strengthening the reporting of observational studies in epidemiology using Mendelian randomization. JAMA. 2021;326(16):1614. doi:10.1001/jama.2021.18236
12. Panyard DJ, Kim KM, Darst BF, et al. Cerebrospinal fluid metabolomics identifies 19 brain-related phenotype associations. Commun Biol. 2021;4(1). doi:10.1038/s42003-020-01583-z
13. Shin S-Y, Fauman EB, Petersen A-K, et al. An atlas of genetic influences on human blood metabolites. Nature Genet. 2014;46(6):543–550. doi:10.1038/ng.2982
14. Kurki MI, Karjalainen J, Palta P, et al. FinnGen provides genetic insights from a well-phenotyped isolated population. Nature. 2023;613(7944):508–518.
15. Bao H, Chen Y, Meng Z, Chu Z. The causal relationship between CSF metabolites and GBM: a two-sample Mendelian randomization analysis. BMC Cancer. 2024;24(1). doi:10.1186/s12885-024-12901-7
16. Hartwig FP, Davey Smith G, Bowden J. Robust inference in summary data Mendelian randomization via the zero modal pleiotropy assumption. Int J Epidemiol. 2017;46(6):1985–1998. doi:10.1093/ije/dyx102
17. Papadimitriou N, Dimou N, Tsilidis KK, et al. Physical activity and risks of breast and colorectal cancer: a Mendelian randomisation analysis. Nat Commun. 2020;11(1). doi:10.1038/s41467-020-14389-8
18. Burgess S, Thompson SG. Interpreting findings from Mendelian randomization using the MR-Egger method. Eur J Epidemiol. 2017;32(5):377–389. doi:10.1007/s10654-017-0255-x
19. Verbanck M, Chen C-Y, Neale B, Do R. Detection of widespread horizontal pleiotropy in causal relationships inferred from Mendelian randomization between complex traits and diseases. Nature Genet. 2018;50(5):693–698. doi:10.1038/s41588-018-0099-7
20. Hemani G, Bowden J, Davey Smith G. Evaluating the potential role of pleiotropy in Mendelian randomization studies. Human Molecular Genetics. 2018;27(R2):R195–R208. doi:10.1093/hmg/ddy163
21. Millard LAC, Davey Smith G, Tilling K. Using the global randomization test as a Mendelian randomization falsification test for the exclusion restriction assumption. Eur J Epidemiol. 2024;39(8):843–855. doi:10.1007/s10654-024-01097-6
22. Davey Smith G, Ebrahim S. ‘Mendelian randomization’: can genetic epidemiology contribute to understanding environmental determinants of disease?*. Int J Epidemiol. 2003;32(1):1–22. doi:10.1093/ije/dyg070
23. Davies NM, Holmes MV, Davey Smith G. Reading Mendelian randomisation studies: a guide, glossary, and checklist for clinicians. BMJ. 2018;k601. doi:10.1136/bmj.k601
24. Kumari S, Gupta R, Ambasta RK, Kumar P. Emerging trends in post-translational modification: shedding light on Glioblastoma multiforme. BBA-Rev Cancer. 2023;1878(6).
25. Ijare OB, Hambarde S, Brasil da Costa FH, et al. Glutamine anaplerosis is required for amino acid biosynthesis in human meningiomas. Neuro-Oncology. 2022;24(4):556–568. doi:10.1093/neuonc/noab219
26. Safari Yazd H, Bazargani SF, Fitzpatrick G, Yost RA, Kresak J, Garrett TJ. Metabolomic and lipidomic characterization of meningioma grades using LC–HRMS and machine learning. J Am Soc Mass Spectrom. 2023;34(10):2187–2198. doi:10.1021/jasms.3c00158
27. Bender L, Somme F, Ruhland E, Cicek AE, Bund C, Namer IJ. Metabolomic profile of aggressive meningiomas by using high-resolution magic angle spinning nuclear magnetic resonance. J Proteome Res. 2019;19(1):292–299. doi:10.1021/acs.jproteome.9b00521
28. Masalha W, Daka K, Woerner J, et al. Metabolic alterations in meningioma reflect the clinical course. BMC Cancer. 2021;21(1). doi:10.1186/s12885-021-07887-5
29. Li Y, Gu W, Hepokoski M, et al. Energy metabolism dysregulation in chronic kidney disease. Kidney360. 2023;4(8):1080–1094. doi:10.34067/KID.0000000000000153
30. Ferkingstad E, Sulem P, Atlason BA, et al. Large-scale integration of the plasma proteome with genetics and disease. Nature Genet. 2021;53(12):1712–1721. doi:10.1038/s41588-021-00978-w
31. Simic P, Kim W, Zhou W, et al. Glycerol-3-phosphate is an FGF23 regulator derived from the injured kidney. J Clin Investig. 2020;130(3):1513–1526. doi:10.1172/JCI131190
32. Shinobu miyazaki-anzai ALK, Blaine J, Miyazaki M, Miyazaki M. Targeted disruption of a proximal tubule specific TMEM174 gene in mice causes hyperphosphatemia and vascular calcification. J Am Soc Nephrol. 2022;33(8):1477–1486. doi:10.1681/ASN.2021121578
33. Zhou W, Simic P, Zhou IY, et al. Kidney glycolysis serves as a mammalian phosphate sensor that maintains phosphate homeostasis. J Clin Investig. 2023;133(8). doi:10.1172/JCI164610
34. Baranovičová E, Galanda T, Galanda M, et al. Metabolomic profiling of blood plasma in patients with primary brain tumours: basal plasma metabolites correlated with tumour grade and plasma biomarker analysis predicts feasibility of the successful statistical discrimination from healthy subjects – a preliminary study. IUBMB Life. 2019;71(12):1994–2002. doi:10.1002/iub.2149
35. Tomomi Kuninaka KK, Kobayashi S, Nagamine H, Hokama Y. Metabolic profiling of atypical meningioma and recurrent meningioma: a comparative analysis with proton magnetic resonance spectroscopy. J Neurosurg. 2024;142(1):214–223. doi:10.3171/2024.4.JNS24187
36. Osman D, Ali O, Obada M, El‐Mezayen H, El‐Said H. Chromatographic determination of some biomarkers of liver cirrhosis and hepatocellular carcinoma in Egyptian patients. Biomed Chromatogr. 2016;31(6). doi:10.1002/bmc.3893
37. Yoon H, Shaw JL, Haigis MC, Greka A. Lipid metabolism in sickness and in health: emerging regulators of lipotoxicity. Molecular Cell. 2021;81(18):3708–3730. doi:10.1016/j.molcel.2021.08.027
38. Kou W, Luchtman D, Song C. Eicosapentaenoic acid (EPA) increases cell viability and expression of neurotrophin receptors in retinoic acid and brain-derived neurotrophic factor differentiated SH-SY5Y cells. Eur J Nutr. 2008;47(2):104–113. doi:10.1007/s00394-008-0703-1
39. Newell M, Mazurak V, Postovit LM, Field CJ. N-3 long-chain polyunsaturated fatty acids, eicosapentaenoic and docosahexaenoic acid, and the role of supplementation during cancer treatment: a scoping review of current clinical evidence. Cancers. 2021;13(6):1206. doi:10.3390/cancers13061206
40. Shi Y, Tang Y, Sun Z, et al. Scutellarein inhibits osteosarcoma growth by targeting the TLR4/TRAF6/NF-κB pathway. Drug Des Devel Ther. 2025;19:51–64. doi:10.2147/DDDT.S489092
41. Wu X, Yuan H, Wu Q, et al. Threonine fuels glioblastoma through YRDC-mediated codon-biased translational reprogramming. Nat Cancer. 2024;5(7):1024–1044. doi:10.1038/s43018-024-00748-7
42. Miyagishima DF, Moliterno J, Claus E, Günel M. Hormone therapies in meningioma-where are we? J Neuro-Oncol. 2022;161(2):297–308. doi:10.1007/s11060-022-04187-1
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Identification of 7-HOCA as a Potential Biomarker in Glioblastoma: Evidence from Genome-Wide Association Study and Clinical Validation
Zhao Z, Xing N, Sun G
International Journal of General Medicine 2024, 17:6185-6197
Published Date: 13 December 2024
The Causal Association Analysis between Depression and Cerebrospinal Fluid: From the Perspective of Mendelian Randomization
Zhang Y, Wu P, Liu Z
Psychology Research and Behavior Management 2025, 18:1085-1097
Published Date: 5 May 2025
Causal Association Between Genetically Predicted Circulating Metabolites and Pressure Ulcers: A Two-Sample Mendelian Randomization Study
Hu X, Zhang Y, Wu Y, Peng M
Journal of Inflammation Research 2025, 18:6907-6926
Published Date: 28 May 2025