Back to Journals » Drug Design, Development and Therapy » Volume 20
Paeoniflorin as a Promising Therapeutic Candidate in Intestinal Diseases: Experimental Evidence, Mechanistic Insights, and Translational Perspectives
Authors Chang C, Xue J, Xu Z, Hou G
Received 16 April 2026
Accepted for publication 4 June 2026
Published 9 June 2026 Volume 2026:20 617375
DOI https://doi.org/10.2147/DDDT.S617375
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Muzammal Hussain
Congcong Chang,1 Jing Xue,2 Ziyou Xu,1 Guanqun Hou1
1College of Traditional Chinese Medicine, Shandong University of Traditional Chinese Medicine, Jinan, Shandong, People’s Republic of China; 2First School of Clinical Medicine, Shandong University of Traditional Chinese Medicine, Jinan, Shandong, People’s Republic of China
Correspondence: Guanqun Hou, College of Traditional Chinese Medicine, Shandong University of Traditional Chinese Medicine, Jinan, Shandong, People’s Republic of China, Email [email protected]
Abstract: Intestinal diseases, including inflammatory bowel disease, functional bowel disorders, stress-related enteropathy, colorectal cancer, and colitis-associated colorectal cancer, share overlapping pathological processes such as epithelial barrier disruption, immune dysregulation, microbiota imbalance, oxidative stress, and inflammation-driven tissue injury. Paeoniflorin, a major monoterpene glycoside derived from Paeonia species, has attracted increasing attention because of its anti-inflammatory, antioxidant, immunomodulatory, anti-apoptotic, and barrier-protective properties. This review summarizes experimental evidence and mechanistic findings regarding the therapeutic potential of paeoniflorin in intestinal diseases. Current preclinical studies suggest that paeoniflorin can alleviate colitis, improve epithelial barrier integrity, promote mucosal repair, modulate microbiota–metabolite interactions, reduce visceral hypersensitivity, and suppress inflammation-associated colorectal tumorigenesis. Mechanistically, these effects involve coordinated regulation of inflammatory signaling, immune-cell balance, epithelial regeneration, gut–brain axis-related pathways, and oncogenic inflammatory cascades. Recent advances in colon-targeted delivery, pharmacokinetic modulation, compatibility-dependent exposure, and derivative development further support its translational potential. However, existing evidence remains largely based on in vitro and animal studies, with limited clinical data, heterogeneous models and dosing regimens, insufficient PK/PD validation, and unresolved druggability challenges. Future research should integrate mechanistic validation, optimized delivery systems, exposure–response evaluation, and clinically relevant disease stratification to support its progression from a bioactive natural compound to a viable intestinal therapeutic candidate.
Keywords: paeoniflorin, intestinal diseases, intestinal barrier, gut microbiota, translational pharmacology
Introduction
Intestinal diseases encompass a broad clinical spectrum, including inflammatory bowel disease (IBD), functional bowel disorders, stress-related intestinal injury, and colorectal cancer (CRC), and have become an increasingly important global health burden.1–3 Recent epidemiological evidence further underscores the public-health relevance of these conditions. The global incidence rate of IBD increased from 1990 to 2021, although age-standardized mortality and disability-adjusted life-year rates showed declining trends. IBS also remains highly prevalent worldwide, with pooled prevalence estimates varying according to diagnostic criteria, from approximately 3.8% under the Rome IV criteria to 9.2% under the Rome III criteria.4 CRC continues to represent a major global malignancy, with more than 1.9 million new cases and approximately 904,000 deaths estimated worldwide in 2022.5 Among these conditions, IBD, particularly Crohn’s disease (CD), often follows a chronic, relapsing course and may ultimately lead to progressive intestinal damage and functional disability.1 Disruption of the intestinal barrier is widely recognized as a shared pathological basis across multiple intestinal disorders, as it promotes the transepithelial translocation of luminal antigens and microbial products, thereby triggering mucosal immune imbalance, persistent inflammation, and tissue injury, and consequently driving disease chronicity and progression toward more complicated phenotypes.6,7 Meanwhile, irritable bowel syndrome (IBS) is now regarded as a prototypical disorder of gut–brain interaction, with core mechanisms involving visceral hypersensitivity, aberrant stress responses, dysregulated neuroimmune signaling, and impaired barrier function, rather than being considered merely a functional disturbance.8,9 In addition, prolonged chronic intestinal inflammation may facilitate the transition from inflammation to tumorigenesis through sustained activation of inflammatory signaling, barrier disruption, and abnormal epithelial renewal, thereby contributing to the initiation and progression of colitis-associated neoplasia and CRC.10,11 Therefore, it is of considerable significance to explore intervention molecules with multi-target and multi-level regulatory properties from the perspective of the continuous pathological cascade spanning barrier homeostasis, immune regulation, microbiota interaction, and inflammation-associated tumorigenic transformation.
Paeoniflorin, a representative bioactive monoterpene glycoside derived from plants of the Paeonia genus, particularly Paeonia lactiflora, is also recognized as one of the major pharmacologically active constituents of medicinal materials such as Radix Paeoniae Alba.12 In recent years, accumulating evidence has demonstrated that paeoniflorin possesses a broad spectrum of pharmacological activities, including anti-inflammatory, antioxidant, immunomodulatory, anti-apoptotic, and barrier-protective effects, thereby attracting sustained interest regarding its therapeutic potential in a variety of complex diseases.13–15 In the field of intestinal diseases, experimental studies have shown that paeoniflorin can suppress innate inflammatory signaling and abnormal immune cell infiltration in colitis models, while also alleviating colonic tissue injury.16 Further investigations have revealed that paeoniflorin not only promotes intestinal stem cell-mediated epithelial regeneration and mucosal repair, but also enhances intestinal barrier function through mechanisms related to group 3 innate lymphoid cells (ILC3s), suggesting that its effects extend beyond a purely anti-inflammatory role.17,18 In models of functional bowel disorders, paeoniflorin has likewise been shown to relieve visceral hypersensitivity, ameliorate low-grade inflammation and barrier dysfunction, and exert protective effects through pathways associated with the gut–brain axis or inflammasomes.19,20 In addition, existing studies suggest that paeoniflorin may also inhibit colitis-associated carcinogenesis and the malignant phenotype of CRC, further highlighting its potential as a candidate intervention across the continuous spectrum from inflammation and tissue injury to carcinogenesis.21,22 Nevertheless, paeoniflorin still faces practical limitations, including low oral bioavailability, limited membrane permeability, and the need for further optimization of its druggability, which is also an important reason why formulation improvement and derivative development have received increasing attention.13,14
Overall, paeoniflorin may represent a promising candidate intervention capable of acting simultaneously at multiple levels, including the intestinal barrier, mucosal immunity, microbiota-related metabolism, the gut–brain axis, and inflammation-associated tumorigenic transformation (Figure 1 and Table 1). However, the current body of evidence remains dominated by in vitro and animal studies with considerable heterogeneity, which limits the generalizability of existing conclusions and slows clinical translation. Against this background, this review systematically summarizes the experimental evidence regarding paeoniflorin in IBD, functional bowel disorders, stress-related intestinal injury and enteropathy, CRC, and CAC. It further discusses its derivatives, formulation optimization, translational prospects, current limitations, and future research directions, with the aim of providing a useful reference for subsequent mechanistic integration and preclinical development.
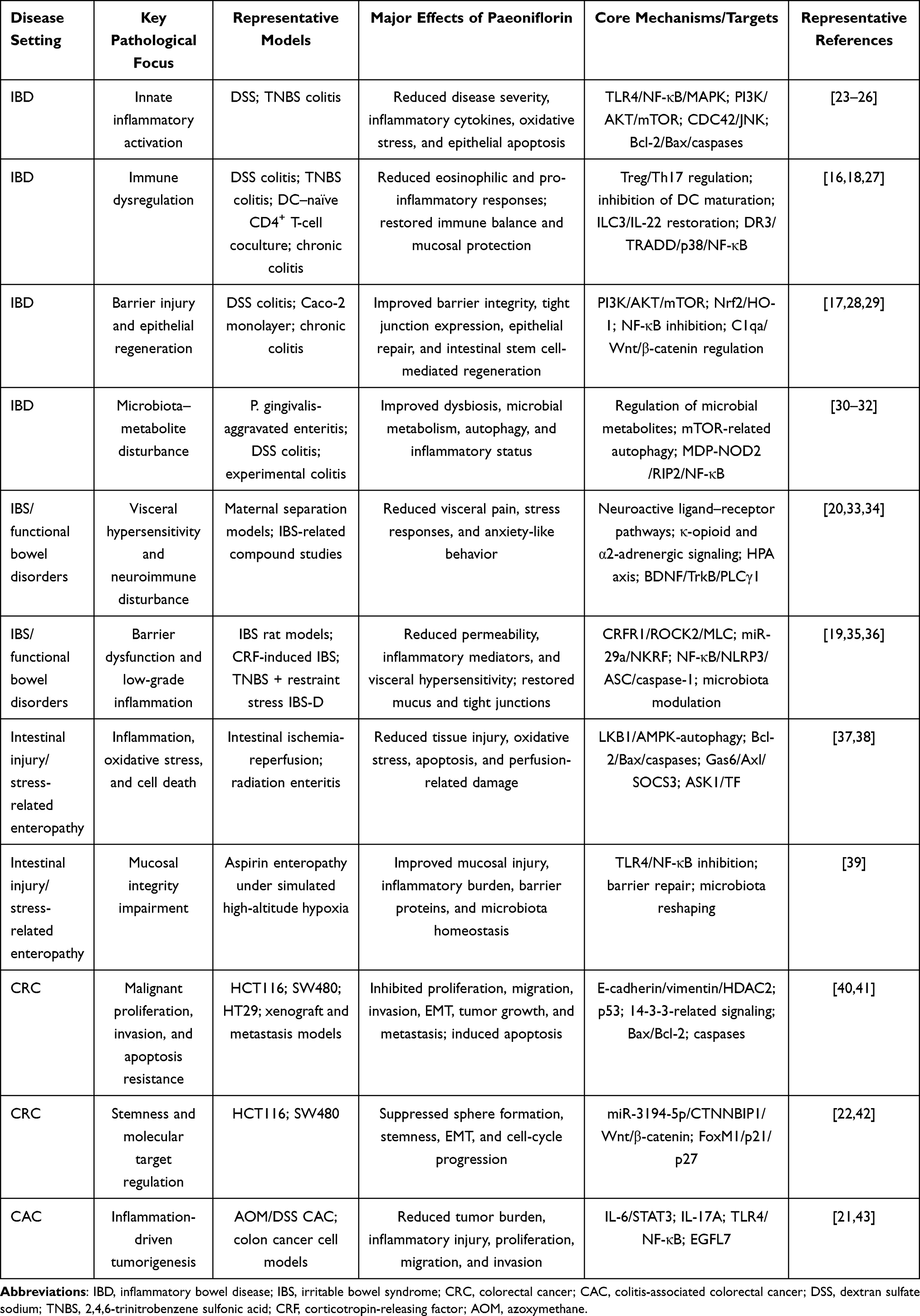 |
Table 1 Condensed Summary of the Experimental Evidence for Paeoniflorin in Intestinal Diseases |
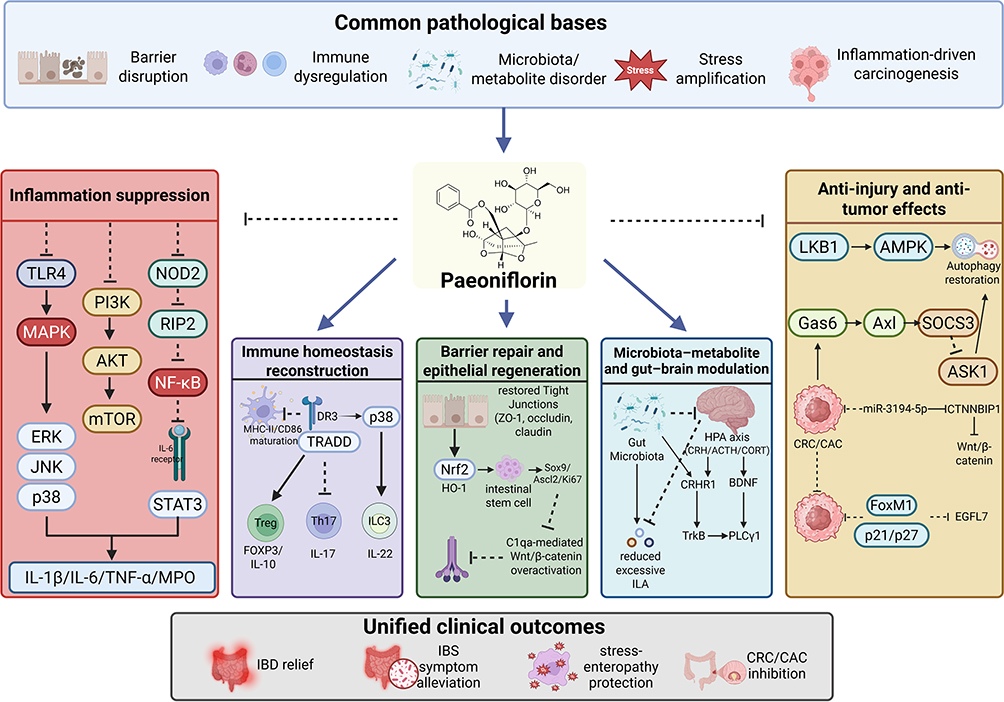 |
Figure 1 Paeoniflorin as a multi-target candidate for intestinal diseases. This schematic summarizes the shared pathological bases of intestinal diseases, including barrier disruption, immune dysregulation, microbiota–metabolite imbalance, stress amplification, and inflammation-driven carcinogenesis. Paeoniflorin may act on these interconnected axes by suppressing inflammatory signaling, restoring immune homeostasis, promoting epithelial barrier repair and regeneration, modulating microbiota–metabolite and gut–brain interactions, and attenuating intestinal injury and CRC/CAC progression. Arrows indicate activation, upregulation, or promotion; blunt-ended lines indicate inhibition or suppression; upward and downward arrows indicate increased and decreased expression or activity, respectively. Abbreviations: IBD, inflammatory bowel disease; IBS, irritable bowel syndrome; CRC, colorectal cancer; CAC, colitis-associated colorectal cancer; TLR4, Toll-like receptor 4; NF-κB, nuclear factor kappa B; MAPK, mitogen-activated protein kinase; PI3K, phosphoinositide 3-kinase; AKT, protein kinase B; mTOR, mammalian target of rapamycin; Treg, regulatory T cell; Th17, T helper 17 cell; ILC3, group 3 innate lymphoid cell; HPA, hypothalamic–pituitary–adrenal; EGFL7, epidermal growth factor-like domain-containing protein 7. |
Experimental Evidence of Paeoniflorin in Inflammatory Intestinal Diseases
Suppression of Innate Inflammatory Signaling
The pathogenesis and progression of IBD are closely associated with excessive activation of innate immunity. Aberrant innate inflammatory signaling, by continuously amplifying inflammation, oxidative stress, and epithelial injury, has become a major driving force that promotes mucosal damage and impairs repair.44,45
Available experimental evidence indicates that paeoniflorin exerts relatively consistent anti-inflammatory effects across multiple colitis models. A common finding is its significant amelioration of inflammatory phenotypes, including body weight loss, increased disease activity index, colon shortening, and histopathological injury, accompanied by concomitant reductions in the levels of inflammatory markers such as myeloperoxidase (MPO), interleukin-1β (IL-1β), interleukin-6 (IL-6), and tumor necrosis factor-α (TNF-α).23–26 At the molecular level, the inhibitory effects of paeoniflorin on innate inflammatory signaling are not restricted to a single pathway. On the one hand, it suppresses the Toll-like receptor 4 (TLR4)-dependent nuclear factor kappa B (NF-κB)/mitogen-activated protein kinase (MAPK) cascade, thereby reducing the activation of p65, extracellular signal-regulated kinase (ERK), JNK, and p38 and ultimately attenuating inflammatory cytokine transcription and downstream amplification.24 On the other hand, paeoniflorin also inhibits phosphoinositide 3-kinase (PI3K)/protein kinase B (AKT)/mammalian target of rapamycin (mTOR)-related inflammatory signaling, suggesting that it may act at a more upstream level within the regulatory networks governing inflammatory responses, cell survival, and metabolism.25 In addition, in 2,4,6-trinitrobenzene sulfonic acid (TNBS)- or DSS-induced colitis models, paeoniflorin has been shown to alleviate inflammation-associated epithelial apoptosis by suppressing NF-κB-mediated inflammatory responses and modulating apoptosis-related molecules such as B-cell lymphoma 2 (Bcl-2), Bcl-2-associated X protein (Bax), caspase-3, and caspase-9.26 Moreover, by inhibiting cell division cycle 42 (CDC42)/JNK signaling and correcting oxidative stress as well as certain metabolic disturbances, paeoniflorin further demonstrates a multilayered protective profile characterized by coordinated anti-inflammatory, antioxidant, and anti-apoptotic effects.23
Regulation of Immune Cell Balance
IBD is, in essence, also a disorder of mucosal homeostasis caused by dysregulation of the intestinal immune cell network. Accordingly, restoring the functional balance among dendritic cells, the regulatory T cell (Treg)/T helper 17 (Th17) axis, and innate lymphoid cells is critical for shifting the therapeutic paradigm from simple suppression of inflammation to reconstruction of mucosal homeostasis.46–48
Available studies indicate that paeoniflorin can reshape intestinal immune balance at multiple levels. First, in DSS-induced colitis models, paeoniflorin not only reduces pro-inflammatory cytokines such as IL-1β, IL-6, and TNF-α, but also decreases the expression of eosinophil-related chemotactic and marker molecules, including chemokine (C-C motif) ligand 11 (CCL11), chemokine (C-C motif) ligand 24 (CCL24), C-C chemokine receptor 3 (CCR3), and sialic acid-binding immunoglobulin-like lectin F (Siglec-F), thereby suppressing eosinophil infiltration. This effect is accompanied by restoration of the Treg proportion, suggesting that paeoniflorin exerts dual actions in attenuating aberrant inflammatory cell recruitment and correcting immune skewing.16 Second, in TNBS-induced ulcerative colitis models and dendritic cell–naïve CD4⁺ T-cell coculture systems, paeoniflorin inhibits dendritic cell maturation, downregulates major histocompatibility complex class II (MHC-II), CD86, and IL-12, and consequently corrects the Th17/Treg imbalance by reducing interleukin-17 (IL-17) levels while increasing forkhead box P3 (Foxp3) and IL-10 expression. These findings suggest that paeoniflorin can re-establish immune tolerance through modulation of antigen-presenting cell function.27 Furthermore, in chronic colitis models, paeoniflorin also acts on the ILC3-related immune network by restoring the proportions of ILC3s, natural cytotoxicity receptor (NCR)⁺ ILC3s, and IL-22⁺ ILC3s, while reducing NCR− ILC3s and pro-inflammatory effector molecules such as IL-17A and granulocyte-macrophage colony-stimulating factor (GM-CSF). At the same time, it suppresses activation of the death receptor 3 (DR3)/TNFR1-associated death domain protein (TRADD)/p38/NF-κB signaling pathway, thereby promoting IL-22-dominant mucosal protective effects.18 Collectively, paeoniflorin facilitates the restoration of intestinal immunity from a pro-inflammatory dysregulated state toward one characterized by mucosal tolerance and repair through coordinated regulation of multiple immune cell populations and their functional networks.16,18,27
Barrier Repair and Epithelial Regeneration
Disruption of the intestinal mucosal barrier constitutes an important basis for the persistence and recurrence of IBD. Therefore, beyond suppressing inflammation, promoting epithelial regeneration and mucosal healing has become a key therapeutic dimension for achieving genuine disease remission.49–51
In this regard, paeoniflorin has demonstrated particularly notable capacities to repair barrier function and promote epithelial regeneration. Studies have shown that paeoniflorin not only ameliorates inflammatory phenotypes in DSS-induced colitis in mice, but also upregulates the expression of intestinal stem cell-related molecules such as Lgr5, Sox9, and Ascl2, as well as Ki67, restores differentiation markers including ChgA, Muc2, Lyz, and Villin, and enhances organoid growth together with intestinal epithelial cell proliferation, migration, and repair. These effects are closely associated with activation of the PI3K-AKT-mTOR signaling pathway and the promotion of intestinal stem cell renewal and differentiation.17 In an in vitro Caco-2 monolayer model, paeoniflorin increases transepithelial electrical resistance (TEER), decreases fluorescein isothiocyanate (FITC)-dextran permeability, and restores the expression of tight junction proteins such as zonula occludens-1 (ZO-1), occludin, and claudin-5. At the same time, by activating nuclear factor erythroid 2-related factor 2 (Nrf2)/heme oxygenase-1 (HO-1) signaling and inhibiting NF-κB signaling, it exerts coordinated protective effects that integrate anti-inflammatory activity with barrier repair.28 Moreover, in chronic colitis models, paeoniflorin has been shown to directly target C1qa, reduce the release of soluble C1q from macrophages, enhance membrane-anchored C1q-associated phagocytic clearance, and suppress C1q-mediated overactivation of Wnt/β-catenin signaling, thereby correcting aberrant intestinal stem cell proliferation and differentiation imbalance and promoting mucosal structural reconstruction and restoration of homeostasis.29 Taken together, paeoniflorin not only repairs the intestinal epithelial barrier and restores permeability homeostasis, but also promotes intestinal stem cell-mediated mucosal regeneration, thereby exhibiting dual potential in IBD as both a mucosal protective agent and a facilitator of tissue repair.17,28,29
Microbiota-Metabolite Interactions
In recent years, the gut microbiota and its metabolites have been increasingly recognized as a central link among mucosal inflammation, immune dysregulation, and barrier damage. Accordingly, examining the mechanisms of paeoniflorin from an integrated microbiota–metabolite–host response perspective may offer deeper insight into the basis of its intestinal protective effects.52,53
Relevant studies have suggested that paeoniflorin can alleviate colitis by modulating microbial composition and microbial metabolism. First, in a model of enteritis aggravated by Porphyromonas gingivalis, paeoniflorin was shown to improve gut microbial dysbiosis and correct abnormalities in pathways such as arginine biosynthesis, phenylalanine metabolism, galactose metabolism, and arachidonic acid metabolism, accompanied by reduced inflammatory cytokine levels and improved immune imbalance. These findings indicate that paeoniflorin can ameliorate inflammation through coordinated regulation of both the intestinal microecology and metabolic networks.30 Second, in DSS-induced colitis, paeoniflorin was found to reduce the level of the microbiota-derived tryptophan metabolite indole-3-lactic acid (ILA), whereas exogenous ILA supplementation attenuated its protective effect. Further mechanistic studies showed that ILA aggravates inflammatory injury by suppressing epithelial autophagy and activating mTOR, suggesting that paeoniflorin may exert its protective action through correction of microbiota-derived metabolic disturbances and restoration of epithelial autophagy.31 Third, paeoniflorin can also reduce the abundance and mucosal infiltration of Gram-positive bacteria, inhibit activation of the MDP-triggered NOD2/RIP2/NF-κB pathway, and downregulate multiple pro-inflammatory mediators, thereby underscoring its potential to intervene in the pathogenic cascade linking microbial compositional alterations, innate immune recognition, and inflammatory amplification32 (Figure 2).
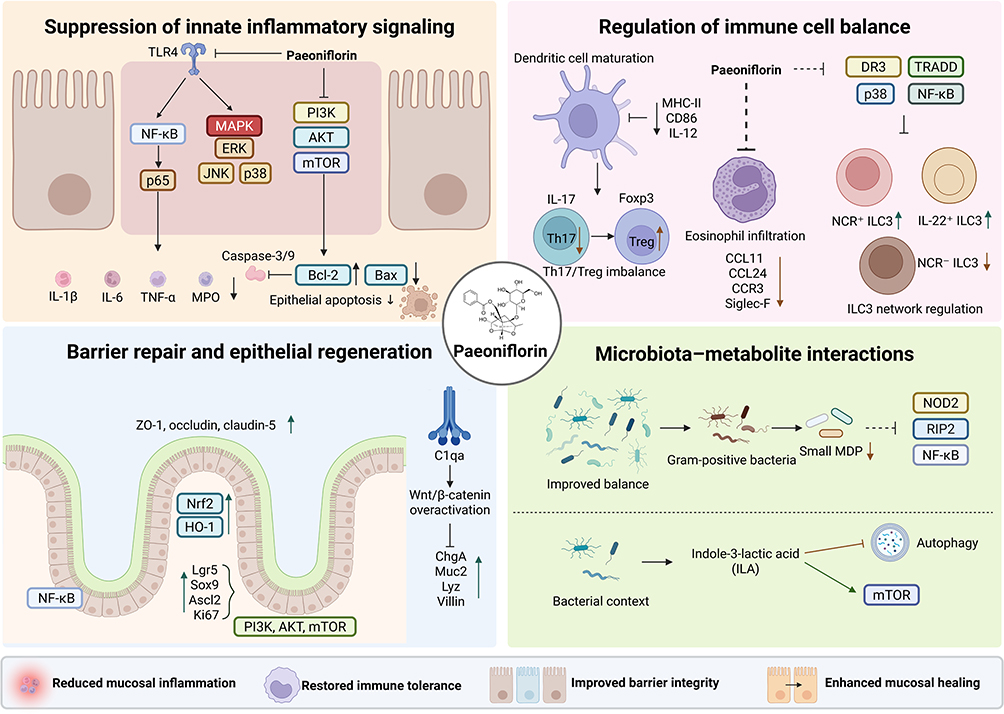 |
Figure 2 Paeoniflorin alleviates inflammatory intestinal diseases through anti-inflammatory, immunomodulatory, barrier-repairing, and microbiota-regulating effects. Paeoniflorin suppresses innate inflammatory signaling mainly through TLR4/NF-κB/MAPK, PI3K/AKT/mTOR, and NOD2/RIP2/NF-κB pathways; restores immune balance by regulating dendritic cell maturation, Th17/Treg balance, eosinophil infiltration, and ILC3 responses; promotes epithelial barrier repair and regeneration by enhancing tight junctions, Nrf2/HO-1 signaling, and intestinal stem cell-related repair; and modulates microbiota–metabolite interactions, including Gram-positive bacteria-related MDP signaling and ILA-associated autophagy. Arrows indicate activation, upregulation, or promotion; blunt-ended lines indicate inhibition or suppression; upward and downward arrows indicate increased and decreased expression or activity, respectively. Abbreviations: IL, interleukin; TNF-α, tumor necrosis factor-α; MPO, myeloperoxidase; Bcl-2, B-cell lymphoma 2; Bax, Bcl-2-associated X protein; MHC-II, major histocompatibility complex class II; Foxp3, forkhead box P3; CCL, C-C motif chemokine ligand; CCR3, C-C chemokine receptor 3; Siglec-F, sialic acid-binding immunoglobulin-like lectin F; DR3, death receptor 3; TRADD, TNFR1-associated death domain protein; NCR, natural cytotoxicity receptor; ZO-1, zonula occludens-1; Nrf2, nuclear factor erythroid 2-related factor 2; HO-1, heme oxygenase-1; MDP, muramyl dipeptide; ILA, indole-3-lactic acid. |
Paeoniflorin in Functional Intestinal Disorders
Visceral Hypersensitivity and Neuroimmune Regulation
One of the core pathological bases of functional bowel disorders, particularly IBS, lies in visceral hypersensitivity and abnormalities in gut–brain axis–neuroimmune regulation. Accordingly, alleviating pain sensitization, modulating stress-related signaling, and intervening in neuroinflammatory networks have become important therapeutic strategies.54,55
Existing studies have shown that paeoniflorin can markedly attenuate visceral hypersensitivity and abdominal pain-related manifestations in multiple IBS-like models. In a study investigating the synergistic effects of active components in Sini San, paeoniflorin was identified as one of the key constituents involved in IBS intervention. In combination with synephrine and naringin, it increased the visceral pain threshold, alleviated pain sensitization, and acted on pathways related to neuroactive ligand–receptor interaction, transient receptor potential (TRP) channel-mediated inflammatory mediator regulation, and serotonergic synapses, suggesting its multi-target regulatory potential within the neuroinflammatory and neuroendocrine network.33 In a neonatal maternal separation-induced rat model of visceral hypersensitivity, paeoniflorin also dose-dependently reduced colorectal distension-induced pain responses, with a relatively rapid onset of analgesic action. Further evidence from central administration, brain region distribution, and receptor blockade experiments suggested that its effects may primarily depend on the central nervous system and may be associated with descending inhibitory pain pathways mediated by κ-opioid receptors and α2-adrenergic receptors.34
Moreover, in maternal separation-induced IBS-like rats, paeoniflorin not only reduced the abdominal withdrawal reflex (AWR) score and electromyographic (EMG) responses, thereby relieving visceral hypersensitivity, but also improved anxiety-like behavior; decreased adrenocorticotropic hormone (ACTH), corticosterone (CORT), and corticotropin-releasing hormone (CRH) levels; restored hippocampal glucocorticoid receptor (GR) expression; downregulated corticotropin-releasing hormone receptor 1 (CRHR1) in both the hippocampus and colon; and suppressed aberrant activation of the colonic brain-derived neurotrophic factor (BDNF)/tropomyosin receptor kinase B (TrkB)/phospholipase Cγ1 (PLCγ1) signaling pathway. These findings suggest that paeoniflorin can ameliorate stress-related pain amplification in functional bowel disorders by modulating the hypothalamic–pituitary–adrenal (HPA) axis, the gut–brain axis, and neuroplasticity-related pathways.20
Intestinal Barrier Dysfunction in IBS
Both clinical and experimental studies suggest that a subset of patients with IBS exhibit increased intestinal permeability, downregulation of tight junction proteins, disruption of the mucus layer, and persistent activation of low-grade inflammation. These alterations allow luminal antigens, bacterial metabolites, and inflammatory mediators to more readily access the mucosal immune system, thereby triggering or exacerbating abdominal pain, diarrhea, and stress-related symptoms.56–58 Therefore, restoring barrier integrity, reducing intestinal hyperpermeability, and attenuating secondary inflammatory activation have become important therapeutic goals.
Available evidence has relatively consistently shown that paeoniflorin can ameliorate IBS-associated barrier injury and increased permeability. In rat models of IBS, paeoniflorin treatment alleviated colonic mucosal structural disruption and inflammatory cell infiltration, reduced fecal water content and related indicators such as D-lactate, TNF-α, and IL-1β, and upregulated ZO-1 and occludin expression. Meanwhile, 16S ribosomal RNA (16S rRNA) analysis showed that paeoniflorin also increased beneficial bacteria, including Lactobacillus, Akkermansia, Alistipes, and Bacteroides, while decreasing potentially detrimental taxa such as Desulfovibrio, Parasutterella, and Enterococcus, suggesting a coordinated effect involving microbiota modulation, inflammation alleviation, and barrier repair.35 In CRF-induced IBS models, paeoniflorin not only improved anxiety-like behavior and visceral hypersensitivity, but also reduced FITC-dextran permeability, increased goblet cell mucus secretion, restored tight junction ultrastructure, and upregulated occludin and claudin-1. Mechanistically, these effects were associated with direct targeting of corticotropin-releasing factor receptor 1 (CRFR1) and inhibition of downstream Rho-associated coiled-coil containing protein kinase 2 (ROCK2)/myosin light chain (MLC) pathway activation, indicating that paeoniflorin may interrupt barrier disruption by interfering with stress receptor signaling.36 In addition, in a mouse model of IBS with diarrhea (IBS-D) induced by TNBS enema combined with restraint stress, paeoniflorin reduced the levels of diamine oxidase (DAO), D-lactic acid (D-LA), MPO, IL-1β, interleukin-18 (IL-18), and TNF-α; ameliorated diarrhea, defecation frequency, and visceral hypersensitivity; and restored claudin-1 and ZO-1 expression. Mechanistically, paeoniflorin downregulated microRNA-29a (miR-29a), upregulated its target molecule nuclear factor-κB repressing factor (NKRF), and inhibited aberrant activation of the NF-κB/NLR family pyrin domain-containing 3 (NLRP3)/apoptosis-associated speck-like protein containing a CARD (ASC)/caspase-1 inflammasome pathway, thereby alleviating low-grade inflammation and re-establishing epithelial barrier homeostasis19 (Figure 3).
 |
Figure 3 Paeoniflorin improves functional intestinal disorders by regulating visceral hypersensitivity, gut–brain stress signaling, and epithelial barrier dysfunction. In IBS-like models, paeoniflorin may reduce visceral hypersensitivity by modulating HPA-axis activity, restoring glucocorticoid receptor signaling, inhibiting BDNF/TrkB/PLCγ1 activation, and engaging possible central descending inhibitory pathways. It also improves barrier dysfunction by targeting CRHR1 and inhibiting ROCK2/MLC signaling, restoring tight junction proteins and mucus secretion, modulating gut microbiota, and suppressing the miR-29a/NKRF/NF-κB/NLRP3 inflammasome axis. Arrows indicate activation, upregulation, or promotion; blunt-ended lines indicate inhibition or suppression; upward and downward arrows indicate increased and decreased expression or activity, respectively. Abbreviations: IBS, irritable bowel syndrome; CNS, central nervous system; CRH, corticotropin-releasing hormone; ACTH, adrenocorticotropic hormone; CORT, corticosterone; GR, glucocorticoid receptor; BDNF, brain-derived neurotrophic factor; TrkB, tropomyosin receptor kinase B; PLCγ1, phospholipase Cγ1; CRHR1, corticotropin-releasing hormone receptor 1; ROCK2, Rho-associated coiled-coil containing protein kinase 2; MLC, myosin light chain; NKRF, nuclear factor-κB repressing factor; NLRP3, NLR family pyrin domain-containing 3; ASC, apoptosis-associated speck-like protein containing a CARD. |
Paeoniflorin in Intestinal Injury and Stress-Related Enteropathy
Anti-Inflammatory and Anti-Oxidative Protection
The development and progression of intestinal injury-related disorders and stress-associated intestinal diseases generally involve the combined actions of multiple injurious mechanisms, including inflammation, ischemia-hypoxia, oxidative stress, microcirculatory disturbance, and cell death, which together continuously disrupt mucosal architecture and amplify local tissue injury.59,60
Available studies indicate that paeoniflorin exerts clear protective effects across different models of stress-induced intestinal injury. In intestinal ischemia-reperfusion models, paeoniflorin markedly alleviates histopathological intestinal damage, reduces inflammatory indicators such as IL-6, IL-1β, TNF-α, and MPO, and simultaneously decreases malondialdehyde (MDA) while increasing superoxide dismutase (SOD) and glutathione (GSH) levels, suggesting that it is capable of concurrently suppressing inflammatory responses and attenuating oxidative stress.37 Further studies have shown that paeoniflorin upregulates Bcl-2 while downregulating Bax and cleaved caspase-3/caspase-9, thereby attenuating epithelial cell apoptosis. It also restores autophagic flux, enhances Beclin-1 and LC3-II expression, promotes p62 degradation, and activates LKB1/AMPK signaling. Notably, knockdown of LKB1 markedly weakens its pro-autophagic, anti-inflammatory, and anti-apoptotic effects, indicating that the LKB1/AMPK-mediated restoration of autophagy is a key mechanism underlying its protective effect against intestinal ischemic injury.37
In models of radiation enteritis, paeoniflorin likewise reduces mortality, ameliorates body weight loss, mucosal damage, and inflammatory cell infiltration, and restores local intestinal blood flow, suggesting that its effects extend beyond anti-inflammation to the alleviation of ischemic alterations following radiation injury.38 Mechanistically, paeoniflorin upregulates the growth arrest-specific 6 (Gas6)/AXL receptor tyrosine kinase (Axl)/suppressor of cytokine signaling 3 (SOCS3) axis, thereby inhibiting the apoptosis signal-regulating kinase 1 (ASK1)/tissue factor (TF) signaling pathway, reducing TF-related microthrombosis and microcirculatory dysfunction, and ultimately attenuating radiation-induced intestinal injury through multiple coordinated processes, including suppression of inflammation, improvement of perfusion, and reduction of secondary tissue damage.38
Preservation of Mucosal Integrity
In stress-related intestinal disorders, impaired mucosal integrity can further aggravate inflammation and hinder repair through barrier disruption, increased permeability, and microecological imbalance. Therefore, protecting mucosal architecture, preserving barrier function, and reconstructing the local microecological environment are important therapeutic objectives.6
Studies have shown that, in a model of aspirin-induced small intestinal enteropathy combined with simulated high-altitude hypoxia, hypoxia exacerbates small intestinal mucosal injury, whereas paeoniflorin dose-dependently ameliorates body weight loss and histopathological damage. In addition, paeoniflorin reduces inflammatory and oxidative stress-related indicators, including IL-1β, TNF-α, and MPO, while increasing interleukin-10 (IL-10), SOD, and the expression of ZO-1 and occludin, suggesting that it exerts both anti-inflammatory and barrier-repairing effects.39 Mechanistically, paeoniflorin suppresses TLR4/NF-κB signaling and reshapes the gut microbial community structure, thereby producing an integrated protective effect characterized by inflammation inhibition, barrier repair, and restoration of microbiota homeostasis (Figure 4).
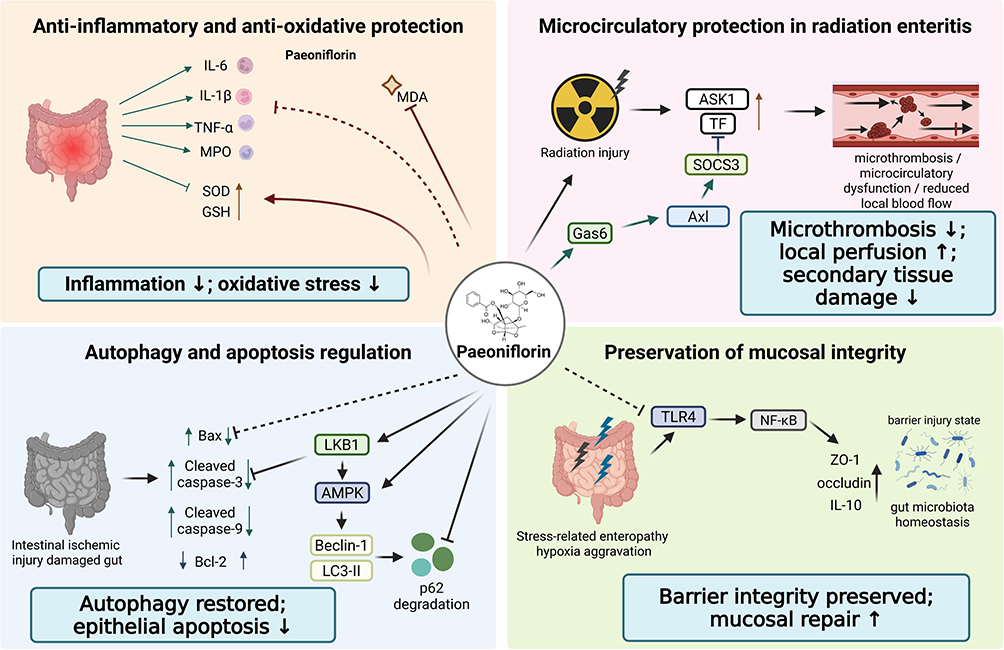 |
Figure 4 Paeoniflorin protects against intestinal injury and stress-related enteropathy through anti-inflammatory, pro-autophagic, microcirculatory, and barrier-preserving effects. Paeoniflorin attenuates intestinal ischemia-reperfusion injury, radiation enteritis, and stress-related enteropathy by reducing inflammatory mediators and oxidative stress, activating LKB1/AMPK-mediated autophagy, inhibiting epithelial apoptosis, improving Gas6/Axl/SOCS3-mediated microcirculatory protection, suppressing ASK1/TF-related microthrombosis, and preserving mucosal integrity through TLR4/NF-κB inhibition, tight junction restoration, and gut microbiota regulation. Arrows indicate activation, upregulation, or promotion; blunt-ended lines indicate inhibition or suppression; upward and downward arrows indicate increased and decreased expression or activity, respectively. Abbreviations: MDA, malondialdehyde; SOD, superoxide dismutase; GSH, glutathione; LKB1, liver kinase B1; AMPK, AMP-activated protein kinase; LC3-II, microtubule-associated protein 1 light chain 3-II; p62, sequestosome 1; Gas6, growth arrest-specific 6; Axl, AXL receptor tyrosine kinase; SOCS3, suppressor of cytokine signaling 3; ASK1, apoptosis signal-regulating kinase 1; TF, tissue factor; ZO-1, zonula occludens-1. |
Paeoniflorin in Colorectal Cancer and Colitis-Associated Tumorigenesis
Direct Anti-Tumor Effects
CRC is one of the most common malignant tumors of the digestive tract,61 and its initiation and progression involve multiple key processes, including aberrant tumor cell proliferation, evasion of apoptosis, enhanced invasion and metastasis, and EMT.62,63
Available studies indicate that paeoniflorin exerts relatively clear direct antitumor activity against CRC. On the one hand, in HCT116 and SW480 cells, paeoniflorin dose-dependently inhibits cell proliferation, migration, and invasion, and reverses EMT by upregulating E-cadherin while downregulating vimentin and histone deacetylase 2 (HDAC2), thereby reducing the invasive and metastatic potential of tumor cells. In an in vivo nude mouse model of liver metastasis, paeoniflorin likewise decreases both the number and volume of hepatic metastatic lesions, providing further in vivo support for its antimetastatic effects.40 On the other hand, in HT29 cells and nude mouse xenograft models, paeoniflorin suppresses tumor cell growth and induces cell-cycle arrest and apoptosis. These effects are associated with upregulation of tumor protein p53 (p53), modulation of 14-3-3-related proteins, activation of caspase-3/caspase-9, and an increased Bax/Bcl-2 ratio. In vivo, paeoniflorin also reduces the positivity rate of proliferating cell nuclear antigen (PCNA) and increases the number of terminal deoxynucleotidyl transferase dUTP nick end labeling (TUNEL)-positive cells, without showing obvious hematological toxicity.41
The direct antitumor effects of paeoniflorin in CRC are mainly manifested as a continuous process of inhibiting proliferation, inducing apoptosis, suppressing migration and invasion, and restraining metastatic progression. Current evidence suggests that paeoniflorin does not merely confer indirect benefit by improving the intestinal inflammatory milieu, but can also directly interfere with the malignant phenotype of tumor cells. Therefore, it has a sound experimental basis as a potential candidate for adjuvant therapy in CRC.40,41
Molecular Targets in Colorectal Cancer
With advances in mechanistic studies of CRC, the maintenance of tumor stemness, aberrant activation of Wnt/β-catenin signaling, dysregulation of the cell cycle, and disruption of transcriptional regulatory networks have been recognized as major drivers of persistent tumor progression and therapeutic resistance.64,65 Compared with nonspecific cytotoxic effects, identifying the key molecular targets of paeoniflorin in CRC may help clarify its deeper antitumor mechanisms and better support subsequent mechanistic integration and translational research.
Studies have shown that paeoniflorin suppresses sphere formation, EMT, and stemness-related phenotypes in HCT116 and SW480 cells, while downregulating OCT4, Nanog, Sox2, p-β-catenin, Axin2, and C-myc. Further mechanistic investigations demonstrated that paeoniflorin downregulates abnormally elevated microRNA-3194-5p (miR-3194-5p) and upregulates its target gene catenin beta interacting protein 1 (CTNNBIP1), thereby inhibiting Wnt/β-catenin signaling and weakening tumor cell stemness. Overexpression of miR-3194-5p or knockdown of CTNNBIP1 partially reverses these effects, suggesting that the miR-3194-5p/CTNNBIP1/Wnt/β-catenin axis is an important pathway underlying the action of paeoniflorin.22 In addition, in HCT116 cells, paeoniflorin markedly decreases the mRNA and protein expression of forkhead box M1 (FoxM1), while increasing the expression of its downstream cell-cycle inhibitory molecules p21 and p27, and inducing G0/G1-phase arrest, apoptosis, and reduced migratory and invasive capacities. Moreover, FoxM1 overexpression partially reverses the inhibitory effects of paeoniflorin, whereas FoxM1 knockdown further enhances its antitumor activity, indicating that FoxM1 is another key molecular target through which paeoniflorin suppresses CRC progression.42
Mechanistic research on paeoniflorin in CRC has progressed beyond simple phenotypic inhibition to the level of target-specific intervention. Current evidence indicates that paeoniflorin can both suppress tumor stemness by modulating the miRNA–target gene network and interfere with cell-cycle progression, invasion, and metastasis through downregulation of FoxM1, highlighting its distinct multi-target properties. Nevertheless, the available evidence in this area is still derived mainly from in vitro cell-based studies, and additional animal and preclinical investigations are needed to verify the stability and translational relevance of these molecular targets in vivo.
Chemopreventive Potential in CAC
Colitis-associated colorectal cancer (CAC) is a representative form of tumorigenic transformation that arises after prolonged chronic intestinal inflammation. Its pathological nature is characterized not by inflammation alone or tumorigenesis alone, but rather by the reciprocal promotion and stepwise evolution of both processes within the same disease continuum.11,66,67 Persistent inflammatory stimulation can drive abnormal epithelial proliferation, dysplasia, and ultimately malignant transformation through sustained activation of pro-inflammatory cytokines, aberrant immune responses, barrier disruption, and local carcinogenic signaling.68 Therefore, molecules capable of simultaneously targeting the inflammatory microenvironment and tumor-promoting pathways are often of particular value for the chemoprevention and early intervention of CAC.
Available studies indicate that paeoniflorin exhibits relatively clear chemopreventive potential in CAC models. In an azoxymethane (AOM)/DSS-induced mouse model of CAC, network pharmacology and molecular docking analyses suggested that IL-6 may represent a key target of paeoniflorin. Animal experiments further confirmed that paeoniflorin improves survival, reduces the number and volume of colonic tumors, alleviates colonic inflammation and histopathological injury, and decreases the expression of proliferation markers such as Ki-67 and PCNA. Mechanistically, paeoniflorin downregulates IL-6 levels in both serum and colonic tissue, inhibits signal transducer and activator of transcription 3 (STAT3) phosphorylation, and reduces local IL-17A expression, suggesting that it primarily delays CAC progression by suppressing the IL-6/STAT3 inflammation-driven oncogenic signaling axis.21 Another study, using both an AOM/DSS-induced CAC model and colon cancer cell experiments, similarly found that paeoniflorin significantly reduces tumor burden, attenuates inflammatory cell accumulation and intestinal mucosal barrier injury, and downregulates inflammatory cytokines including TNF-α, IL-1β, IL-6, and IL-13, while also inhibiting TLR4/NF-κB signaling and epidermal growth factor-like domain-containing protein 7 (EGFL7) expression. In vitro, paeoniflorin also suppresses cancer cell proliferation, migration, invasion, and colony formation while promoting apoptosis, indicating that it not only improves the inflammatory microenvironment but also directly blocks inflammation-driven tumor deterioration43 (Figure 5).
 |
Figure 5 Paeoniflorin suppresses CRC and CAC through cell-autonomous anti-tumor effects and inhibition of inflammation-driven tumorigenesis. In CRC, paeoniflorin inhibits EMT, migration, invasion, proliferation, tumor growth, and stemness by regulating E-cadherin/vimentin/HDAC2, p53/caspase signaling, miR-3194-5p/CTNNBIP1/Wnt/β-catenin signaling, and FoxM1/p21/p27-mediated cell-cycle control. In CAC, paeoniflorin may interrupt the inflammation-to-cancer continuum by suppressing IL-6/STAT3, IL-17A, TLR4/NF-κB, and EGFL7-related signaling, thereby reducing inflammatory injury, barrier damage, tumor burden, and CAC progression. Arrows indicate activation, upregulation, or promotion; blunt-ended lines indicate inhibition or suppression; upward and downward arrows indicate increased and decreased expression or activity, respectively. Abbreviations: CRC, colorectal cancer; CAC, colitis-associated colorectal cancer; EMT, epithelial–mesenchymal transition; HDAC2, histone deacetylase 2; PCNA, proliferating cell nuclear antigen; CTNNBIP1, catenin beta interacting protein 1; FoxM1, forkhead box M1; STAT3, signal transducer and activator of transcription 3; EGFL7, epidermal growth factor-like domain-containing protein 7. |
The significance of paeoniflorin in CAC is better understood as that of an intervention molecule targeting the inflammation-to-carcinogenesis continuum. It can both alleviate chronic inflammation and the amplification of tumor-promoting signals, and inhibit tumor cell proliferation and metastasis-related behaviors, thereby showing considerable theoretical promise for chemoprevention and early intervention. Although current studies remain largely limited to animal and cell-based models, they already suggest that paeoniflorin may be relevant not only to isolated IBD or CRC, but may be particularly well suited for intervention research in inflammation-driven tumorigenesis.
Derivatives, Formulations, and Translational Extension
Formulation Optimization and Colon-Targeted Delivery
For natural bioactive compounds such as paeoniflorin, their clinical translational value depends not only on whether they possess anti-inflammatory or barrier-protective activities per se, but also to a great extent on their delivery efficiency within the intestine, local retention capacity, and effective exposure at the lesion site. This is particularly relevant in diseases such as ulcerative colitis, which are characterized by localized colonic inflammation and barrier disruption. In such settings, improving site-specific drug release and local bioavailability in the colon is a critical step in translating natural products from pharmacologically active compounds into clinically applicable therapeutic agents.69–71
Available studies suggest that formulation optimization can further enhance the intestinal protective effects of paeoniflorin. For example, an oral colon-targeted bigel co-loaded with patchouli oil and paeoniflorin has been shown to meet the dual delivery requirements of both hydrophilic and hydrophobic active components, while exhibiting more pronounced overall protective effects than administration of the individual constituents alone at both the cellular and animal levels. Specifically, this bigel formulation alleviates DSS-induced body weight loss, elevated DAI, and colon shortening, improves mucosal structural damage and goblet cell depletion, and reduces the levels of inflammatory cytokines such as IL-1β, IL-6, and TNF-α. At the same time, it restores the expression of barrier-related molecules including ZO-1, occludin, claudin-4, and mucin 2 (MUC-2), and promotes an increase in M2 macrophages while reducing neutrophil and CD8⁺ T-cell infiltration, suggesting that the optimized delivery system can simultaneously potentiate the anti-inflammatory, barrier-repairing, and immunomodulatory effects of paeoniflorin.72 Beyond improving colonic targeting, the manner in which paeoniflorin is incorporated into a delivery matrix may directly influence its release kinetics and local pharmacodynamic effects. For hydrogel-, bigel-, nanoparticle-, or polymer-based systems, material swelling, erosion, enzymatic degradation, pH responsiveness, and mucoadhesive retention can affect the timing, rate, and spatial distribution of paeoniflorin release in the inflamed intestine. These parameters are translationally important because sustained local exposure may be required not only for suppressing inflammatory cytokine production, but also for maintaining effective concentrations that support epithelial barrier restoration, intestinal stem cell-mediated regeneration, and functional mucosal recovery. Future formulation studies should therefore report not only therapeutic endpoints, but also drug loading efficiency, encapsulation efficiency, in vitro and ex vivo release profiles, degradation behavior, local colonic concentration, and exposure–response relationships.69–72
Pharmacokinetic Modulation and Compatibility-Dependent Exposure
Another key challenge frequently encountered in the clinical translation of natural products is insufficient in vivo exposure, an incomplete understanding of pharmacokinetic behavior under pathological conditions, and the difficulty of predicting how multi-component formulations may affect absorption and distribution.73,74 For paeoniflorin, efficacy evaluation should not remain limited to the question of whether it is effective; it is also necessary to determine whether its systemic exposure and local bioavailability vary across different disease states, dosing frequencies, and compatibility contexts, and whether such changes may in turn influence therapeutic efficacy.75
In this regard, several studies have provided valuable supplementary evidence from a pharmacokinetic perspective. First, in a dinitrobenzene sulfonic acid (DNBS)-induced colitis model, repeated high-dose administration of Moutan Cortex extract increased systemic exposure to paeoniflorin, accompanied by improvements in body weight loss, disease activity index, and colonic injury. These changes were also associated with downregulation of matrix metalloproteinase-9 (MMP-9), transforming growth factor-β1 (TGF-β1), and AMPK, together with upregulation of occludin, suggesting a potential link between enhanced in vivo exposure of paeoniflorin and its barrier-protective as well as anti-inflammatory benefits.76 Second, in IBS model rats, a polysaccharide-containing multi-herb formulation significantly increased the maximum plasma concentration (Cmax) and area under the concentration-time curve (AUC) of multiple active constituents, including paeoniflorin. After depletion of the gut microbiota, however, this exposure-enhancing effect was markedly attenuated, indicating that the absorption and bioavailability of paeoniflorin are influenced not only by co-administered components but also closely related to the participation of the gut microbiota.77 These findings suggest that, in the context of intestinal diseases, the in vivo disposition of paeoniflorin may not be fixed, but rather jointly regulated by disease status, formulation compatibility, and the intestinal microecological environment.
Taken together, pharmacokinetic and compatibility studies have advanced our understanding of paeoniflorin beyond that of a compound with demonstrated efficacy, positioning it instead as a candidate drug with considerable potential for further optimization. Although the current evidence remains insufficient to establish a complete model for clinical pharmacokinetic translation, it already suggests two important points: first, repeated dosing and dose design may substantially affect the in vivo exposure of paeoniflorin; second, formulation context and the gut microbiota may reshape its absorption process. Accordingly, future development of paeoniflorin should not focus solely on pharmacodynamic endpoints, but should also systematically incorporate factors such as dosing regimen, compatibility conditions, and microecological background in order to enhance its true translational value.
Structural Modification and Derivative Development
Although natural paeoniflorin itself has already demonstrated certain anti-inflammatory, barrier-protective, and immunomodulatory potential, the parent molecule may still have limitations in terms of pharmacological potency, target specificity, stability, and druggability. Therefore, structural modification of paeoniflorin to develop derivatives with greater activity or novel mechanistic features represents an important direction for advancing this class of compounds from basic research toward candidate drug development.
At present, two representative studies support this concept. First, the paeoniflorin derivative CP-25 has shown clear protective effects in both a TNF-α-induced Caco-2 cell model of barrier injury and a DSS-induced mouse model of colitis. CP-25 increases TEER, reduces fluorescent permeability, restores the expression of ZO-1 and occludin, decreases epithelial cell apoptosis, and suppresses the release of inflammatory cytokines such as IL-1β, IL-6, and interleukin-8 (IL-8). Mechanistically, CP-25 primarily acts by inhibiting activation of ERK1/2-NF-κB signaling, reducing the membrane translocation of G protein-coupled receptor kinase 2 (GRK2) and β-arrestin 2, and restoring their cytoplasmic inhibitory effects on ERK1/2, thereby maintaining intestinal epithelial barrier homeostasis.78 Second, another structurally modified product, benzoylpaeoniflorin (BP), has also been shown to markedly attenuate body weight loss, increased DAI, colonic injury, and barrier disruption in both DSS-induced ulcerative colitis mice and DSS-stimulated NCM460 intestinal epithelial cells, while upregulating the expression of tight junction proteins such as ZO-1 and occludin. More importantly, BP does not act merely through conventional anti-inflammatory mechanisms, but instead exerts anti-inflammatory and mucosal protective effects by targeting phosphogluconate dehydrogenase (PGD), restoring pentose phosphate pathway activity, suppressing ferroptosis in intestinal epithelial cells, and activating the Nrf2/solute carrier family 7 member 11 (SLC7A11)/glutathione peroxidase 4 (GPX4) axis, suggesting that paeoniflorin derivatives may acquire mechanistic features more distinctive than those of the parent compound.79
Overall, studies on structural modification indicate that paeoniflorin is not merely a natural product suitable for direct application, but also a lead scaffold with substantial room for further optimization. Whether reflected in the enhanced regulation of barrier-related signaling by CP-25 or the novel mechanisms of BP in anti-ferroptosis and metabolic reprogramming, current evidence suggests that the development of paeoniflorin derivatives may represent a key direction for expanding its therapeutic indications in intestinal diseases and improving its druggability.
Translational Extension: From Active Natural Compound to Drug Development Candidate
Overall, research on paeoniflorin in intestinal diseases has moved beyond the preliminary question of whether the isolated compound is effective, and is gradually extending toward issues more directly relevant to drug development, including formulation optimization, pharmacokinetic modulation, mechanisms of multi-herb compatibility, and structural modification (Table 2). This research trajectory in fact reflects a broader trend in the translation of natural products: only by integrating pharmacological activity, delivery, exposure, and druggability can the preclinical value of a candidate molecule be truly maximized.
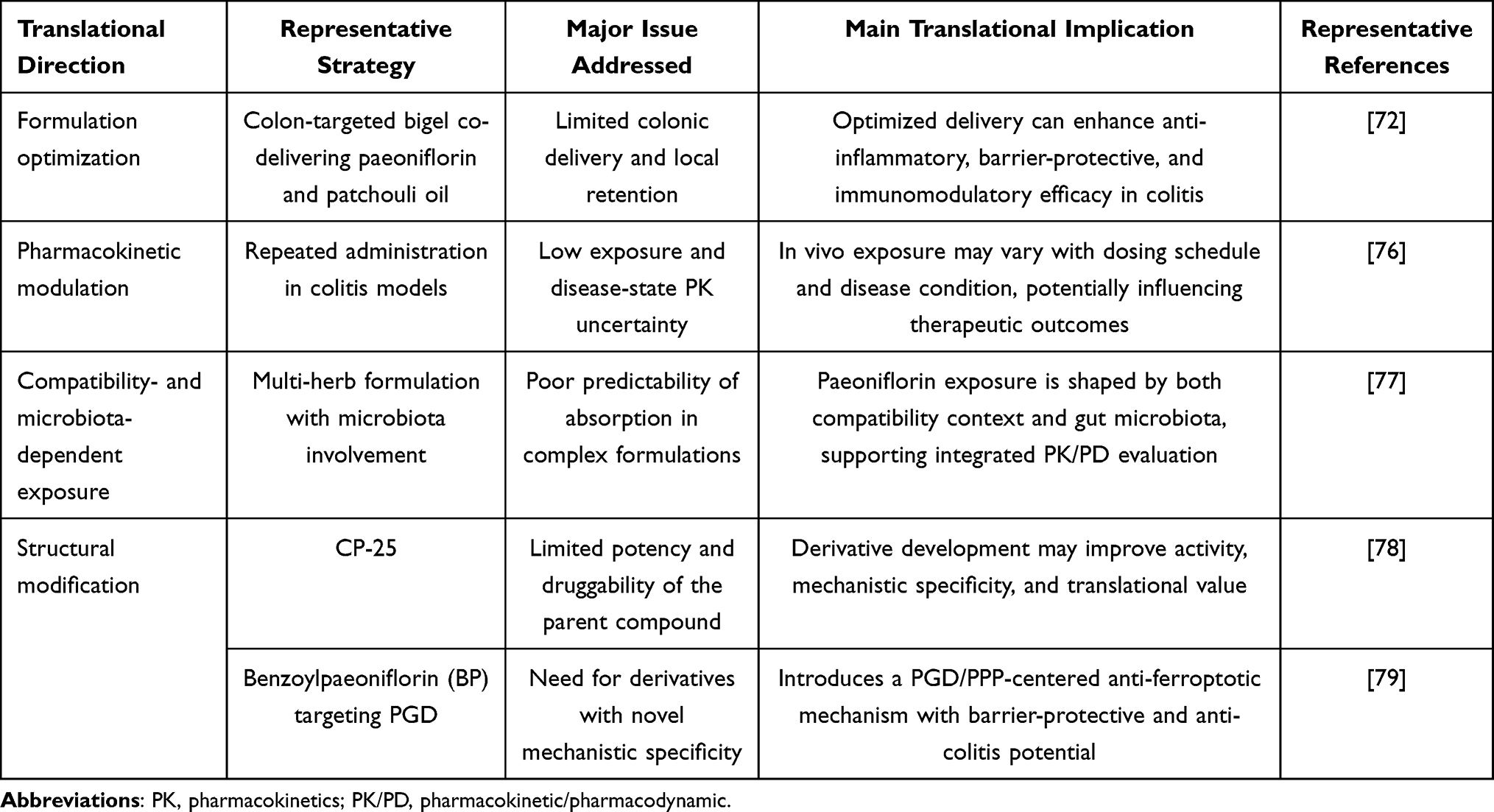 |
Table 2 Condensed Summary of the Translational Extension of Paeoniflorin |
Based on the current evidence, the translational extension of paeoniflorin is mainly reflected in three complementary pathways. First, targeted formulations may improve colonic delivery efficiency and enhance local therapeutic efficacy.72 Second, PK and multi-herb compatibility studies may help clarify the tunability of its in vivo exposure and its dependence on the intestinal microecological environment.76,77 Third, structural modification may yield derivatives with greater activity, novel mechanisms, or improved druggability.78,79 For paeoniflorin-based interventions, clinical viability should not be judged solely by pharmacological activity in experimental models. A clinically viable strategy would need to satisfy several interconnected criteria: sufficient systemic or local bioavailability at the intestinal lesion site; reproducible and controllable release behavior; an acceptable safety margin after repeated administration; formulation stability and scalable manufacturing; compatibility with standard quality-control requirements; and a feasible regulatory pathway. For colon-targeted oral formulations, development may be relatively more straightforward if the product can be regulated as a conventional drug product with well-characterized composition, release specifications, and pharmacokinetic behavior. By contrast, delivery platforms that combine paeoniflorin with device-like components, biologics, or complex multifunctional carriers may raise additional regulatory considerations, including product classification, quality documentation, review-strategy issues, and potential combination-product concerns.
Current Clinical Evidence and Evidence Gap
At present, published direct clinical evidence supporting paeoniflorin as a treatment for intestinal diseases remains limited. Although paeoniflorin-containing preparations have been investigated in human pharmacokinetic or non-intestinal clinical studies, these data cannot be directly extrapolated to IBD, IBS, CRC, or CAC.80,81 Therefore, current translational claims for intestinal diseases should remain grounded in preclinical evidence. Future clinical development should begin with standardized paeoniflorin preparations or clearly defined formulations, followed by Phase I safety and pharmacokinetic studies, biomarker-based proof-of-mechanism studies, and carefully selected early-phase trials in disease settings where paeoniflorin’s mechanisms are most biologically plausible, such as barrier repair in ulcerative colitis, pain-predominant or diarrhea-predominant IBS, and chemoprevention in high-risk inflammation-associated populations.17,19,21,28,43,82
Discussion
Taken together, the available evidence indicates that the effects of paeoniflorin in intestinal diseases are not confined to a single pathological node, but rather exhibit multi-target regulatory properties across disease spectra and biological hierarchies. In IBD, functional bowel disorders, stress-related intestinal injury, CRC, and CAC, its overall protective pattern is relatively consistent, namely, suppression of aberrant inflammatory activation, maintenance or reconstruction of the epithelial barrier, modulation of host immunity and microecological interactions, and, in specific contexts, further regulation of the gut–brain axis or inflammation-driven tumorigenic transformation.
Nevertheless, several important limitations in the current body of evidence should be acknowledged. First, most available studies remain at the cell and animal levels, and substantial heterogeneity exists among studies in terms of model selection, dose regimens, treatment windows, endpoint measurements, and the depth of mechanistic validation, making cross-study comparisons difficult.75 Second, paeoniflorin itself faces druggability-related barriers, including low bioavailability, limited membrane permeability, and insufficient in vivo exposure, whereas the relationship among effective dose, local exposure, and pharmacodynamic outcome remains insufficiently characterized in most current studies.75,83,84 Third, although some studies have begun to address the influence of multi-herb formulations, the gut microbiota, and disease status on the pharmacokinetic behavior of paeoniflorin, systematic pharmacokinetic/pharmacodynamic (PK/PD) evidence is still clearly lacking, and the extrapolation of preclinical findings to human dosing, exposure, and efficacy remains insufficiently developed.74,85 Finally, many proposed key targets are still inferred from correlative omics, docking, or pathway-intervention data, whereas direct target engagement and causal validation remain limited in most current studies. Therefore, a more cautious conclusion at this stage is that paeoniflorin possesses considerable preclinical development potential, but a sufficiently robust and clinically translatable evidence chain has not yet been established. To further strengthen the translational framework of paeoniflorin research, future studies should incorporate preclinical-to-clinical extrapolation strategies. Network pharmacology may be useful for integrating the multi-target and multi-pathway characteristics of paeoniflorin, particularly in the context of traditional Chinese medicine-derived natural products, and for prioritizing candidate targets, signaling networks, and pharmacodynamic biomarkers for subsequent experimental validation.86 However, network-based predictions should be regarded as hypothesis-generating rather than confirmatory and should be combined with molecular docking, omics-based validation, direct target engagement assays, and disease-relevant functional readouts. In parallel, allometric scaling may provide an initial quantitative approach for extrapolating animal pharmacokinetic parameters or effective exposure ranges to humans, thereby supporting preliminary dose estimation and study design.87 Nevertheless, such extrapolation should be interpreted cautiously because paeoniflorin has low oral bioavailability and its intestinal absorption may be influenced by gut microbiota, disease status, and formulation context. Physiologically based pharmacokinetic (PBPK) modeling may offer a more mechanistic framework by integrating physicochemical properties, intestinal permeability, metabolism, transporter involvement, gastrointestinal physiology, disease-related changes, and formulation-dependent release profiles.88 In this regard, combining network pharmacology, allometric scaling, PBPK modeling, and experimental PK/PD validation may help bridge the gap between current preclinical evidence and rational clinical development of paeoniflorin-based interventions.
Based on the above analysis, future research should proceed in three major directions. First, stronger emphasis should be placed on closed-loop mechanistic validation, particularly with respect to the microbiota–metabolite–host response axis, the immune cell–intestinal stem cell interaction axis, and the gut–brain axis–barrier–symptom phenotype framework. By integrating organoid models, coculture systems, single-cell transcriptomics, spatial omics, and target validation strategies, future studies should distinguish the core primary effects of paeoniflorin from secondary downstream changes.89–92 Second, translational optimization should be further promoted, including colon-targeted delivery, nanomedicine design, enhanced local retention, derivative development, and pharmacokinetic evaluation under disease conditions, in order to address the practical bottleneck of compounds that are effective but fail to adequately reach the lesion site.93–95 Third, research should be designed around questions that are more clinically relevant; for example, focusing on barrier healing and relapse prevention in IBD, on pain-predominant or IBS-D subtypes in IBS,55,96,97 and on early chemoprevention in high-risk inflammatory populations in CAC.98–101 Only through the progressive integration of mechanistic validation, delivery optimization, and disease stratification can paeoniflorin truly evolve from a bioactive natural product into a candidate intestinal therapeutic molecule with meaningful developmental potential.
Overall, research on paeoniflorin in intestinal diseases has gradually shifted from the early stage of simply determining whether it has pharmacological activity to a more advanced stage focused on clarifying its multi-level mechanisms and optimizing its translational pathway. Its greatest appeal lies not merely in the fact that it has shown activity across multiple intestinal disease models, but in that these diseases share common pathological axes, including barrier disruption, immune dysregulation, microecological imbalance, stress amplification, and inflammation-driven carcinogenesis, all of which appear to be modulated by paeoniflorin to some extent in a coordinated manner. Therefore, paeoniflorin may be more appropriately regarded as a candidate molecule with systemic interventional potential. With further advances in mechanistic validation, pharmacokinetic optimization, and precision disease stratification, its prospects in the field of intestinal diseases remain worthy of continued attention.
Conclusion and Translational Perspective
In conclusion, paeoniflorin represents a promising multi-target natural compound for intestinal disease intervention, with preclinical evidence supporting its roles in suppressing inflammation, restoring epithelial barrier integrity, modulating immune and microbiota-related networks, reducing visceral hypersensitivity, and interfering with inflammation-driven colorectal tumorigenesis. However, its current developmental status should be viewed as preclinical rather than clinically established. The next stage of research should move beyond descriptive efficacy studies and focus on building a translational evidence chain that includes standardized preparations, optimized colon-targeted delivery, defined release and exposure profiles, PK/PD modeling, direct target validation, repeated-dose safety assessment, and disease-specific patient stratification. From this perspective, paeoniflorin is unlikely to progress simply as a conventional natural product; instead, its clinical potential will depend on whether formulation science, pharmacokinetic optimization, and mechanism-based clinical positioning can be integrated into a coherent development strategy.
Acknowledgments
Figures were created with BioRender.com.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
National Administration of Traditional Chinese Medicine High-level Key Discipline Construction Project of Traditional Chinese Medicine (zyyzdxk-2023118).
Disclosure
Dr Guanqun Hou reports Support for the manuscript from National Administration of Traditional Chinese Medicine High-level Key Discipline Construction Project of Traditional Chinese Medicine (zyyzdxk-2023118), during the conduct of the study, outside the submitted work. The authors declare that they have no competing interests.
References
1. Dolinger M, Torres J, Vermeire S. Crohn’s disease. Lancet. 2024;403(10432):1177–21. doi:10.1016/S0140-6736(23)02586-2
2. Whelan K, Ford AC, Burton-Murray H, et al. Dietary management of irritable bowel syndrome: considerations, challenges, and solutions. Lancet Gastroenterol Hepatol. 2024;9(12):1147–1161. doi:10.1016/S2468-1253(24)00238-3
3. Lin D, Jin Y, Shao X, et al. Global, regional, and national burden of inflammatory bowel disease, 1990-2021: insights from the global burden of disease 2021. Int J Colorectal Dis. 2024;39(1):139. doi:10.1007/s00384-024-04711-x
4. Oka P, Parr H, Barberio B, et al. Global prevalence of irritable bowel syndrome according to Rome III or IV criteria: a systematic review and meta-analysis. Lancet Gastroenterol Hepatol. 2020;5(10):908–917. doi:10.1016/S2468-1253(20)30217-X
5. Bray F, Laversanne M, Sung H, et al. Global cancer statistics 2022: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J Clin. 2024;74(3):229–263. doi:10.3322/caac.21834
6. Neurath MF, Artis D, Becker C. The intestinal barrier: a pivotal role in health, inflammation, and cancer. Lancet Gastroenterol Hepatol. 2025;10(6):573–592. doi:10.1016/S2468-1253(24)00390-X
7. Dunleavy KA, Raffals LE, Camilleri M. Intestinal barrier dysfunction in inflammatory bowel disease: underpinning pathogenesis and therapeutics. Dig Dis Sci. 2023;68(12):4306–4320. doi:10.1007/s10620-023-08122-w
8. Mayer EA, Ryu HJ, Bhatt RR. The neurobiology of irritable bowel syndrome. Mol Psychiatry. 2023;28(4):1451–1465. doi:10.1038/s41380-023-01972-w
9. Zhang X, Shi L, Chen C, et al. The role of miRNA in IBS pathogenesis, diagnosis and therapy: the latest thought. Dig Liver Dis. 2024;56(9):1433–1441. doi:10.1016/j.dld.2024.01.209
10. Hisamatsu T, Miyoshi J, Oguri N, et al. Inflammation-associated carcinogenesis in inflammatory bowel disease: clinical features and molecular mechanisms. Cells. 2025;
11. Shahgoli VK, Noorolyai S, Ahmadpour Youshanlui M, et al. Inflammatory bowel disease, colitis, and cancer: unmasking the chronic inflammation link. Int J Colorectal Dis. 2024;39(1):173. doi:10.1007/s00384-024-04748-y
12. Ma W, Ren H, Meng X, et al. A review of the ethnopharmacology, phytochemistry, pharmacology, pharmacokinetics and quality control of paeonia lactiflora pall. J Ethnopharmacol. 2024;335:118616. doi:10.1016/j.jep.2024.118616
13. Ou X, Yu Z, Pan C, et al. Paeoniflorin: a review of its pharmacology, pharmacokinetics and toxicity in diabetes. Front Pharmacol. 2025;16:1551368. doi:10.3389/fphar.2025.1551368
14. Hong H, Lu X, Wu C, et al. A review for the pharmacological effects of paeoniflorin in the nervous system. Front Pharmacol. 2022;13:898955. doi:10.3389/fphar.2022.898955
15. Ma X, Zhang W, Jiang Y, et al. Paeoniflorin, a natural product with multiple targets in liver diseases-a mini review. Front Pharmacol. 2020;11:531. doi:10.3389/fphar.2020.00531
16. Li J, Ren S, Li M, et al. Paeoniflorin protects against dextran sulfate sodium (DSS)-induced colitis in mice through inhibition of inflammation and eosinophil infiltration. Int Immunopharmacol. 2021;97:107667. doi:10.1016/j.intimp.2021.107667
17. Ma Y, Lang X, Yang Q, et al. Paeoniflorin promotes intestinal stem cell-mediated epithelial regeneration and repair via PI3K-AKT-mTOR signalling in ulcerative colitis. Int Immunopharmacol. 2023;119:110247. doi:10.1016/j.intimp.2023.110247
18. Huang S, Xie X, Xu B, et al. Paeoniflorin ameliorates chronic colitis via the DR3 signaling pathway in group 3 innate lymphoid cells. J Pharm Anal. 2024;14(6):100940. doi:10.1016/j.jpha.2024.01.008
19. Ke W, Wang Y, Huang S, et al. Paeoniflorin alleviates inflammatory response in IBS-D mouse model via downregulation of the NLRP3 inflammasome pathway with involvement of miR-29a. Heliyon. 2022;8(12):e12312. doi:10.1016/j.heliyon.2022.e12312
20. Liang R, Ge W, Song X, et al. Paeoniflorin alleviates anxiety and visceral hypersensitivity via HPA axis and BDNF/TrkB/PLCγ1 pathway in maternal separation-induced IBS-like rats. Curr Mol Pharmacol. 2024;17:e18761429280572. doi:10.2174/0118761429280572240311060851
21. Si XL, Wang Y, Song B-N, et al. Potential chemoprevention of paeoniflorin in colitis-associated colorectal cancer by network pharmacology, molecular docking, and in vivo experiment. Chem Biodivers. 2022;19(8):e202200295. doi:10.1002/cbdv.202200295
22. Su Z, Hu B, Li J, et al. Paeoniflorin inhibits colorectal cancer cell stemness through the miR −3194-5p/catenin beta-interacting protein 1 axis. Kaohsiung J Med Sci. 2023;39(10):1011–1021. doi:10.1002/kjm2.12736
23. Hu Q, Xie J, Jiang T, et al. Paeoniflorin alleviates DSS-induced ulcerative colitis by suppressing inflammation, oxidative stress, and apoptosis via regulating serum metabolites and inhibiting CDC42/JNK signaling pathway. Int Immunopharmacol. 2024;142(Pt A):113039. doi:10.1016/j.intimp.2024.113039
24. Zhang J, Dou W, Zhang E, et al. Paeoniflorin abrogates DSS-induced colitis via a TLR4-dependent pathway. Am J Physiol Gastrointest Liver Physiol. 2014;306(1):G27–36. doi:10.1152/ajpgi.00465.2012
25. Li Q, Zheng S, Niu K, et al. Paeoniflorin improves ulcerative colitis via regulation of PI3K‑AKT based on network pharmacology analysis. Exp Ther Med. 2024;27(4):125. doi:10.3892/etm.2024.12414
26. Gu P, Zhu L, Liu Y, et al. Protective effects of paeoniflorin on TNBS-induced ulcerative colitis through inhibiting NF-kappaB pathway and apoptosis in mice. Int Immunopharmacol. 2017;50:152–160. doi:10.1016/j.intimp.2017.06.022
27. Zheng K, Jia J, Yan S, et al. Paeoniflorin ameliorates ulcerative colitis by modulating the dendritic cell-mediated T(H)17/T(reg) balance. Inflammopharmacology. 2020;28(6):1705–1716. doi:10.1007/s10787-020-00722-6
28. Wu XX, Huang X-L, Chen -R-R, et al. Paeoniflorin prevents intestinal barrier disruption and inhibits lipopolysaccharide (LPS)-induced inflammation in Caco-2 cell monolayers. Inflammation. 2019;42(6):2215–2225. doi:10.1007/s10753-019-01085-z
29. Wang Y, You K, You Y, et al. Paeoniflorin prevents aberrant proliferation and differentiation of intestinal stem cells by controlling C1q release from macrophages in chronic colitis. Pharmacol Res. 2022;182:106309. doi:10.1016/j.phrs.2022.106309
30. Qu R, Li M, Li P, et al. Mitigation of P. gingivalis -exacerbated intestinal inflammation by paeoniflorin through alteration of the gut microbiota. Microbiol Spectr. 2025;13(8):e0000224. doi:10.1128/spectrum.00002-24
31. Fan Q, Guan X, Hou Y, et al. Paeoniflorin modulates gut microbial production of indole-3-lactate and epithelial autophagy to alleviate colitis in mice. Phytomedicine. 2020;79:153345. doi:10.1016/j.phymed.2020.153345
32. Luo X, Wang X, Huang S, et al. Paeoniflorin ameliorates experimental colitis by inhibiting gram-positive bacteria-dependent MDP-NOD2 pathway. Int Immunopharmacol. 2021;90:107224. doi:10.1016/j.intimp.2020.107224
33. Cai T, Wang X, Li B, et al. Deciphering the synergistic network regulation of active components from SiNiSan against irritable bowel syndrome via a comprehensive strategy: combined effects of synephrine, paeoniflorin and naringin. Phytomedicine. 2021;86:153527. doi:10.1016/j.phymed.2021.153527
34. Zhang XJ, Li Z, Leung W-M, et al. The analgesic effect of paeoniflorin on neonatal maternal separation-induced visceral hyperalgesia in rats. J Pain. 2008;9(6):497–505. doi:10.1016/j.jpain.2007.12.009
35. Wang L, Lei J, Zhao Z, et al. Therapeutic effects of paeoniflorin on irritable bowel syndrome in rats. J Vet Sci. 2023;24(3):e23. doi:10.4142/jvs.22083
36. Jin R, Zhou Z, Lou Y, et al. Peoniflorin alleviates corticotropin-releasing factor-induced irritable bowel syndrome by improving the intestinal barrier function through the inhibition of the CRFR1/ROCK2/MLC signaling pathway. J Agric Food Chem. 2026;74(3):2560–2573. doi:10.1021/acs.jafc.5c10339
37. Wen J, Xu B, Sun Y, et al. Paeoniflorin protects against intestinal ischemia/reperfusion by activating LKB1/AMPK and promoting autophagy. Pharmacol Res. 2019;146:104308. doi:10.1016/j.phrs.2019.104308
38. Sheng L, Hu F, Yu H, et al. Paeoniflorin inhibits ASK1-TF axis by up-regulating SOCS3 to alleviate radiation enteritis. Front Pharmacol. 2022;13:743708. doi:10.3389/fphar.2022.743708
39. Tuerxuntayi A, Shi W, Shi T, et al. Paeoniflorin attenuates ASA-induced enteropathy in simulated high-altitude hypoxia in rats by regulating intestinal inflammation, intestinal barrier, and intestinal flora. Toxicol Appl Pharmacol. 2026;506:117641. doi:10.1016/j.taap.2025.117641
40. Zhang JW, Li L-X, Wu W-Z, et al. Anti-tumor effects of paeoniflorin on epithelial-to-mesenchymal transition in human colorectal cancer cells. Med Sci Monit. 2018;24:6405–6413. doi:10.12659/MSM.912227
41. Wang H, Zhou H, Wang C-X, et al. Paeoniflorin inhibits growth of human colorectal carcinoma HT 29 cells in vitro and in vivo. Food Chem Toxicol. 2012;50(5):1560–1567. doi:10.1016/j.fct.2012.01.035
42. Yue M, Li S, Yan G, et al. Paeoniflorin inhibits cell growth and induces cell cycle arrest through inhibition of FoxM1 in colorectal cancer cells. Cell Cycle. 2018;17(2):240–249. doi:10.1080/15384101.2017.1407892
43. Wang Y, Zhou Y, Lin H, et al. Paeoniflorin inhibits the proliferation and metastasis of ulcerative colitis-associated colon cancer by targeting EGFL7. J Oncol. 2022;2022:7498771. doi:10.1155/2022/7498771
44. Wan T, Wang Y, He K, et al. Microbial sensing in the intestine. Protein Cell. 2023;14(11):824–860. doi:10.1093/procel/pwad028
45. Tratenšek A, Locatelli I, Grabnar I, et al. Oxidative stress-related biomarkers as promising indicators of inflammatory bowel disease activity: a systematic review and meta-analysis. Redox Biol. 2024;77:103380. doi:10.1016/j.redox.2024.103380
46. Zhang S, Zhong R, Tang S, et al. Metabolic regulation of the Th17/Treg balance in inflammatory bowel disease. Pharmacol Res. 2024;203:107184. doi:10.1016/j.phrs.2024.107184
47. Horn V, Sonnenberg GF. Group 3 innate lymphoid cells in intestinal health and disease. Nat Rev Gastroenterol Hepatol. 2024;21(6):428–443. doi:10.1038/s41575-024-00906-3
48. Bai X, Chen S, Chi X, et al. Reciprocal regulation of T follicular helper cells and dendritic cells drives colitis development. Nat Immunol. 2024;25(8):1383–1394. doi:10.1038/s41590-024-01882-1
49. Rath T, Atreya R, Bodenschatz J, et al. Intestinal barrier healing is superior to endoscopic and histologic remission for predicting major adverse outcomes in inflammatory bowel disease: the prospective ERIca trial. Gastroenterology. 2023;164(2):241–255. doi:10.1053/j.gastro.2022.10.014
50. Mansouri P, Mansouri P, Najafipour S, et al. Comprehensive computational strategies for multi-target drug discovery in inflammatory bowel disease utilizing bioactive compounds. Scientific Reports. 2025;144:113544.
51. Villablanca EJ, Selin K, Hedin CRH. Mechanisms of mucosal healing: treating inflammatory bowel disease without immunosuppression? Nat Rev Gastroenterol Hepatol. 2022;19(8):493–507. doi:10.1038/s41575-022-00604-y
52. Arifuzzaman M, Collins N, Guo C-J, et al. Nutritional regulation of microbiota-derived metabolites: implications for immunity and inflammation. Immunity. 2024;57(1):14–27. doi:10.1016/j.immuni.2023.12.009
53. Ghosh S, Whitley CS, Haribabu B, et al. Regulation of intestinal barrier function by microbial metabolites. Cell Mol Gastroenterol Hepatol. 2021;11(5):1463–1482. doi:10.1016/j.jcmgh.2021.02.007
54. Vanuytsel T, Bercik P, Boeckxstaens G. Understanding neuroimmune interactions in disorders of gut–brain interaction: from functional to immune-mediated disorders. Gut. 2023;72(4):787–798. doi:10.1136/gutjnl-2020-320633
55. Black CJ, Ford AC. Personalisation of therapy in irritable bowel syndrome: a hypothesis. Lancet Gastroenterol Hepatol. 2024;9(12):1162–1176. doi:10.1016/S2468-1253(24)00245-0
56. Yuan Y, Wang X, Huang S, et al. Low-level inflammation, immunity, and brain-gut axis in IBS: unraveling the complex relationships. Gut Microbes. 2023;15(2):2263209. doi:10.1080/19490976.2023.2263209
57. Grover M, Vanuytsel T, Chang L. Intestinal permeability in disorders of gut–brain interaction: from bench to bedside. Gastroenterology. 2025;168(3):480–495. doi:10.1053/j.gastro.2024.08.033
58. Hanning N, Edwinson AL, Ceuleers H, et al. Intestinal barrier dysfunction in irritable bowel syndrome: a systematic review. Ther Adv Gastroenterol. 2021;14:1756284821993586. doi:10.1177/1756284821993586
59. Wang F, Huang H, Wei X, et al. Targeting cell death pathways in intestinal ischemia-reperfusion injury: a comprehensive review. Cell Death Discov. 2024;10(1):112. doi:10.1038/s41420-024-01891-x
60. Dvornikova KA, Platonova ON, Bystrova EY. Hypoxia and intestinal inflammation: common molecular mechanisms and signaling pathways. Int J Mol Sci. 2023;24(3).
61. Eng C, Yoshino T, Ruíz-García E, et al. Colorectal cancer. Lancet. 2024;404(10449):294–310. doi:10.1016/S0140-6736(24)00360-X
62. Li Q, Geng S, Luo H, et al. Signaling pathways involved in colorectal cancer: pathogenesis and targeted therapy. Signal Transduct Target Ther. 2024;9(1):266. doi:10.1038/s41392-024-01953-7
63. Shin AE, Giancotti FG, Rustgi AK. Metastatic colorectal cancer: mechanisms and emerging therapeutics. Trends Pharmacol Sci. 2023;44(4):222–236. doi:10.1016/j.tips.2023.01.003
64. Sun L, Xing J, Zhou X, et al. Wnt/β-catenin signalling, epithelial-mesenchymal transition and crosslink signalling in colorectal cancer cells. Biomed Pharmacother. 2024;175:116685. doi:10.1016/j.biopha.2024.116685
65. Merhi M, Ahmad F, Taib N, et al. The complex network of transcription factors, immune checkpoint inhibitors and stemness features in colorectal cancer: a recent update. Semin Cancer Biol. 2023;89:1–17. doi:10.1016/j.semcancer.2023.01.001
66. Zhou RW, Harpaz N, Itzkowitz SH, et al. Molecular mechanisms in colitis-associated colorectal cancer. Oncogenesis. 2023;12(1):48. doi:10.1038/s41389-023-00492-0
67. Nardone OM, Zammarchi I, Santacroce G, et al. Inflammation-driven colorectal cancer associated with colitis: from pathogenesis to changing therapy. Cancers. 2023;15(8).
68. Dan WY, Zhou G-Z, Peng L-H, et al. Update and latest advances in mechanisms and management of colitis-associated colorectal cancer. World J Gastrointest Oncol. 2023;15(8):1317–1331. doi:10.4251/wjgo.v15.i8.1317
69. Zheng B, Wang L, Yi Y, et al. Design strategies, advances and future perspectives of colon-targeted delivery systems for the treatment of inflammatory bowel disease. Asian J Pharm Sci. 2024;19(4):100943. doi:10.1016/j.ajps.2024.100943
70. Gvozdeva Y, Staynova R. pH-dependent drug delivery systems for ulcerative colitis treatment. Pharmaceutics. 2025;17(2).
71. Kotla NG, Singh R, Baby BV, et al. Inflammation-specific targeted carriers for local drug delivery to inflammatory bowel disease. Biomaterials. 2022;281:121364. doi:10.1016/j.biomaterials.2022.121364
72. Wu Y, Jiang Y, He Y, et al. A bigel co-delivering highly hydrophilic and hydrophobic natural compounds for enhanced ulcerative colitis therapy. Int J Pharm. 2025;678:125706. doi:10.1016/j.ijpharm.2025.125706
73. Huang L, Huang X-H, Yang X, et al. Novel nano-drug delivery system for natural products and their application. Pharmacol Res. 2024;201:107100. doi:10.1016/j.phrs.2024.107100
74. Li M, Wang Y, Chen Y, et al. A comprehensive review on pharmacokinetic mechanism of herb-herb/drug interactions in Chinese herbal formula. Pharmacol Ther. 2024;264:108728. doi:10.1016/j.pharmthera.2024.108728
75. Zhou YX, Gong X-H, Zhang H, et al. A review on the pharmacokinetics of paeoniflorin and its anti-inflammatory and immunomodulatory effects. Biomed Pharmacother. 2020;130:110505. doi:10.1016/j.biopha.2020.110505
76. Kim JH, Jeong J-S, Kim J-W, et al. Pharmacokinetic profiles of Moutan Cortex after single and repeated administration in a dinitrobenzene sulfonic acid-induced colitis model. PLoS One. 2025;20(12):e0337713. doi:10.1371/journal.pone.0337713
77. Zhong P, Zhou J, Fan Y-T, et al. Co-existing polysaccharides affect the systemic exposure of major bioactive ingredients in Chang-Kang-Fang, a multi-herb prescription for treatment of irritable bowel syndrome. J Ethnopharmacol. 2022;298:115601. doi:10.1016/j.jep.2022.115601
78. Jiang M, Tang C, Yang M, et al. Paeoniflorin-6′-O-benzene sulfonate protected the intestinal epithelial barrier by restoring the inhibitory effect of GRK2 and β-arrestin 2 on ERK1/2-NF-κB. Phytother Res. 2023;37(2):743–758. doi:10.1002/ptr.7657
79. Tang Y, Wang Z, Zhou F, et al. Benzoylpaeoniflorin alleviates ulcerative colitis by inhibiting ferroptosis through targeting phosphogluconic dehydrogenase. Phytomedicine. 2025;147:157111. doi:10.1016/j.phymed.2025.157111
80. Li X, Shi F, Zhang R, et al. Pharmacokinetics, safety, and tolerability of amygdalin and paeoniflorin after single and multiple intravenous infusions of huoxue-tongluo lyophilized powder for injection in healthy chinese volunteers. Clin Ther. 2016;38(2):327–337. doi:10.1016/j.clinthera.2015.12.005
81. Zhou L, Cao T, Wang Y, et al. Clinical observation on the treatment of oral lichen planus with total glucosides of paeony capsule combined with corticosteroids. Int Immunopharmacol. 2016;36:106–110. doi:10.1016/j.intimp.2016.03.035
82. Jiang H, Li J, Wang L, et al. Total glucosides of paeony: a review of its phytochemistry, role in autoimmune diseases, and mechanisms of action. J Ethnopharmacol. 2020;258:112913. doi:10.1016/j.jep.2020.112913
83. Bhalani DV, Nutan B, Kumar A, et al. Bioavailability enhancement techniques for poorly aqueous soluble drugs and therapeutics. Biomedicines. 2022;10(9).
84. Lv Y, Li W, Liao W, et al. Nano-drug delivery systems based on natural products. Int J Nanomedicine. 2024;19:541–569. doi:10.2147/IJN.S443692
85. Chen F, Wen Q, Jiang J, et al. Could the gut microbiota reconcile the oral bioavailability conundrum of traditional herbs? J Ethnopharmacol. 2016;179:253–264. doi:10.1016/j.jep.2015.12.031
86. Zhai Y, Liu L, Zhang F, et al. Network pharmacology: a crucial approach in traditional Chinese medicine research. Chin Med. 2025;20(1):8. doi:10.1186/s13020-024-01056-z
87. Choi GW, Lee YB, Cho HY. Interpretation of Non-Clinical Data for Prediction of Human Pharmacokinetic Parameters: In Vitro-In Vivo Extrapolation and Allometric Scaling. Pharmaceutics; 2019:11(4).
88. Chou P, Shannar A, Pan Y, et al. Application of Physiologically-Based Pharmacokinetic (PBPK) model in drug development and in dietary phytochemicals. Curr Pharmacol Rep. 2025;11(1):45. doi:10.1007/s40495-025-00427-w
89. Chelakkot C, Ghim J, Ryu SH. Mechanisms regulating intestinal barrier integrity and its pathological implications. Exp Mol Med. 2018;50(8):1–9. doi:10.1038/s12276-018-0126-x
90. Huang R, Wu F, Zhou Q, et al. Lactobacillus and intestinal diseases: mechanisms of action and clinical applications. Microbiol Res. 2022;260:127019. doi:10.1016/j.micres.2022.127019
91. Meng X, Zhang G, Cao H, et al. Gut dysbacteriosis and intestinal disease: mechanism and treatment. J Appl Microbiol. 2020;129(4):787–805. doi:10.1111/jam.14661
92. Özkan A, LoGrande NT, Feitor JF, et al. Intestinal organ chips for disease modelling and personalized medicine. Nat Rev Gastroenterol Hepatol. 2024;21(11):751–773. doi:10.1038/s41575-024-00968-3
93. Hu S, Zhao R, Xu Y, et al. Orally-administered nanomedicine systems targeting colon inflammation for the treatment of inflammatory bowel disease: latest advances. J Mater Chem B. 2023;12(1):13–38. doi:10.1039/D3TB02302H
94. Hua S, Marks E, Schneider JJ, et al. Advances in oral nano-delivery systems for colon targeted drug delivery in inflammatory bowel disease: selective targeting to diseased versus healthy tissue. Nanomedicine. 2015;11(5):1117–1132. doi:10.1016/j.nano.2015.02.018
95. Zhang S, Langer R, Traverso G. Nanoparticulate drug delivery systems targeting inflammation for treatment of inflammatory bowel disease. Nano Today. 2017;16:82–96. doi:10.1016/j.nantod.2017.08.006
96. Black CJ, Yiannakou Y, Guthrie EA, et al. A novel method to classify and subgroup patients with ibs based on gastrointestinal symptoms and psychological profiles. Am J Gastroenterol. 2021;116(2):372–381. doi:10.14309/ajg.0000000000000975
97. Heitkemper M, Cain KC, Shulman R, et al. Subtypes of irritable bowel syndrome based on abdominal pain/discomfort severity and bowel pattern. Dig Dis Sci. 2011;56(7):2050–2058. doi:10.1007/s10620-011-1567-4
98. Dyson JK. Colorectal cancer in inflammatory bowel disease: what is the real magnitude of the risk? World J Gastroenterol. 2012;18(29):3839–3848. doi:10.3748/wjg.v18.i29.3839
99. Hsiao S-W, Yen -H-H, Chen -Y-Y. Chemoprevention of colitis-associated dysplasia or cancer in inflammatory bowel disease. Gut Liver. 2022;16(6):840–848. doi:10.5009/gnl210479
100. Triantafillidis JK, Nasioulas G, Kosmidis PA. Colorectal cancer and inflammatory bowel disease: epidemiology, risk factors, mechanisms of carcinogenesis and prevention strategies. Anticancer Res. 2009;29(7):2727–2737.
101. Zhang H, Shi Y, Lin C, et al. Overcoming cancer risk in inflammatory bowel disease: new insights into preventive strategies and pathogenesis mechanisms including interactions of immune cells, cancer signaling pathways, and gut microbiota. Front Immunol. 2023;14:1338918. doi:10.3389/fimmu.2023.1338918
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Taurine Regulates the Expression of Interleukin −17/10 and Intestinal Flora and Protects the Liver and Intestinal Mucosa in a Nonalcoholic Fatty Liver Disease Rat Model
Zhu FL, Huang T, Lv ZL, Liang G, Yao Z, Lan LC, Qadir A, Chen XQ, Shan QW
Diabetes, Metabolic Syndrome and Obesity 2024, 17:675-689
Published Date: 9 February 2024
Portulaca Oleracea L. as a Potential Therapeutic Drug Intervention in Ulcerative Colitis: Mechanisms of Action and Clinical Studies
Liu XY, Liu ZX, Tan WW, Zhang WB, Zhang YL, Zheng L, Que RY, Wen HZ, Dai YC
Drug Design, Development and Therapy 2024, 18:5931-5946
Published Date: 11 December 2024
Metagenomic Sequencing Combined with Metabolomics to Explore Gut Microbiota and Metabolic Changes in Mice with Acute Myocardial Infarction and the Potential Mechanism of Allicin

Gao Y, Qin G, Liang S, Yin J, Wang B, Jiang H, Liu M, Luo F, Li X
Drug Design, Development and Therapy 2025, 19:771-791
Published Date: 3 February 2025
Coix Seed Oil Alleviates DSS-Induced Ulcerative Colitis via Intestinal Barrier Repair and Ferroptosis Regulation
Zeng YX, Li NR, Deng BY, Gu YF, Lu SF, Liu Y
Journal of Inflammation Research 2025, 18:2557-2581
Published Date: 20 February 2025
Si-Ni Decoction as a Potential Treatment for Ulcerative Colitis: Modulation of Gut Microbiota and AKT1 Inhibition Through Network Pharmacology and in vivo Validation
Shi L, Chen L, Jin G, Yang Y, Zhu F, Zhou G
Journal of Inflammation Research 2025, 18:6263-6280
Published Date: 14 May 2025