Back to Journals » International Journal of Nanomedicine » Volume 16
Neuroprotective Efficiency of Prodigiosins Conjugated with Selenium Nanoparticles in Rats Exposed to Chronic Unpredictable Mild Stress is Mediated Through Antioxidative, Anti-Inflammatory, Anti-Apoptotic, and Neuromodulatory Activities
Authors Albrakati A, Alsharif KF, Al omairi NE, Alsanie WF, Almalki ASA, Abd Elmageed ZY, Elshopakey GE, Lokman MS, Bauomy AA, Abdel Moneim AE ![]() , Kassab RB
, Kassab RB ![]()
Received 4 June 2021
Accepted for publication 18 November 2021
Published 30 December 2021 Volume 2021:16 Pages 8447—8464
DOI https://doi.org/10.2147/IJN.S323436
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Farooq A. Shiekh
Ashraf Albrakati,1 Khalaf F Alsharif,2 Naif E Al omairi,3 Walaa F Alsanie,2 Abdulraheem SA Almalki,4 Zakaria Y Abd Elmageed,5 Gehad E Elshopakey,6 Maha S Lokman,7,8 Amira A Bauomy,9 Ahmed E Abdel Moneim,8 Rami B Kassab8,10
1Department of Human Anatomy, College of Medicine, Taif University, Taif, Saudi Arabia; 2Department of Clinical Laboratory Sciences, College of Applied Medical Sciences, Taif University, Taif, Saudi Arabia; 3Department of Internal Medicine, College of Medicine, Taif University, Taif, Saudi Arabia; 4Department of Chemistry, Faculty of Science, Taif University, Taif, Saudi Arabia; 5Department of Pharmacology, Edward via College of Osteopathic Medicine, University of Louisiana at Monroe, Monroe, LA, USA; 6Department of Clinical Pathology, Faculty of Veterinary Medicine, Mansoura University, Mansoura, Egypt; 7Biology Department, College of Science and Humanities, Prince Sattam bin Abdul Aziz University, Alkharj, Saudi Arabia; 8Department of Zoology and Entomology, Faculty of Science, Helwan University, Cairo, Egypt; 9Department of Science Laboratories, College of Science and Arts, Qassim University, ArRassAl-Qassim, Saudi Arabia; 10Biology Department, Faculty of Science and Arts, Al Baha University, Al Makhwah Branch, Al Baha, Saudi Arabia
Correspondence: Ahmed E Abdel Moneim
Department of Zoology and Entomology, Faculty of Science, Helwan University, Cairo, Egypt
Tel +20 1153322079
Email [email protected]
Purpose: Depression is a mood disorder accompanied by intensive molecular and neurochemical alterations. Currently, available antidepressant therapies are not fully effective and are often accompanied by several adverse impacts. Accordingly, the ultimate goal of this investigation was to clarify the possible antidepressant effects of prodigiosins (PDGs) loaded with selenium nanoparticles (PDGs-SeNPs) in chronic unpredictable mild stress (CUMS)-induced depression-like behavior in rats.
Methods: Sixty Sprague Dawley rats were randomly allocated into six groups: control, CUMS group (depression model), fluoxetine (Flu, 10 mg/kg)+CUMS, PDGs+CUMS (300 mg/kg), sodium selenite (Na2SeO3, 400 mg/kg)+CUMS, and PDGs-SeNPs+CUMS (200 mg/kg). All treatments were applied orally for 28 consecutive days.
Results: PDGs-SeNPs administration prevented oxidative insults in hippocampal tissue, as demonstrated by decreased oxidant levels (nitric oxide and malondialdehyde) and elevated innate antioxidants (glutathione, glutathione peroxidase, glutathione reductase, superoxide dismutase, and catalase), in addition to the upregulated expression of nuclear factor erythroid 2-related factor 2 and heme oxygenase-1 in rats exposed to CUMS. Additionally, PDGs-SeNPs administration suppressed neuroinflammation in hippocampal tissue, as determined by the decreased production of pro-inflammatory cytokines (tumor necrosis factor-alpha, interleukin-1β, and interleukin-6), increased anti-inflammatory cytokine interleukin-10, and decreased inflammatory mediators (prostaglandin E2, cyclooxygenase-2, and nuclear factor kappa B). Moreover, PDGs-SeNPs administration in stressed rats inhibited neuronal loss and the development of hippocampal apoptosis through enhanced levels of B cell lymphoma 2 and decreased levels of caspase 3 and Bcl-2-associated X protein. Interestingly, PDGs-SeNPs administration improved hormonal levels typically disrupted by CUMS exposure and significantly modulated hippocampal levels of monoamines, brain-derived neurotrophic factor, monoamine oxidase, and acetylcholinesterase activities, in addition to upregulating the immunoreactivity of glial fibrillary acidic protein in CUMS model rats.
Conclusion: PDGs-SeNPs may serve as a prospective antidepressant candidate due to their potent antioxidant, anti-inflammatory, and neuroprotective potential.
Keywords: chronic unpredictable mild stress, prodigiosin-loaded selenium nanoparticles, oxidative stress, neuroinflammation, apoptosis, neurochemical markers
Introduction
Depression is among the most common neuropsychiatric illnesses, affecting approximately 300 million people of all ages worldwide1 and representing a vast economic and social burden.2 Although the causative factors underlying depression are not clearly understood, sufficient evidence has suggested that neural oxidative insults, inflammation, and apoptosis, associated with neurodegeneration and reduced neurogenesis and neural plasticity, underlies the onset of major depressive disorder.3 The depletion of antioxidant molecules due to the overproduction of reactive oxygen species (ROS) has been demonstrated to play an essential role in the development of depression.4 The pathophysiology of depression is also associated with activation of monoamine oxidase activity, suppression of monoaminergic neurotransmitters, and stimulation of pro-inflammatory mediators that regulate apoptotic signaling pathways resulting in neuronal loss.5,6
Although depression is considered to be a multifactorial mood disorder, stress is believed to be a major contributor, causing neural biochemical alterations and promoting the progression of depression symptoms.7 Chronic unpredictable mild stress (CUMS) has been widely employed as a mechanism for inducing depression-like behaviors in experimental animals to scrutinize the pathophysiological mechanisms underlying depression and evaluate novel antidepressant drug efficacy.8 The CUMS model is commonly associated with increased anxiety-like behavior and impaired hypothalamic-pituitary-adrenocortical (HPA) axis function.9 The CUMS model is associated with reduced sucrose intake and reduced responsiveness, which are the primary symptoms of anhedonia, a central property of depressive disorders.10 Several medications are used for depression treatment, such as monoamine oxidase (MAO) inhibitors, tricyclic antidepressants, and selective serotonin reuptake inhibitors (SSRIs).11,12 However, the currently available depression therapies are not fully satisfactory for all patients13 and are accompanied by several adverse side effects.14 Thus, further investigations remain necessary to discover novel protocols for the development of safer and influential pharmacological therapies for depression.
Due to their availability, safety, and high efficiency, natural compounds are widely used to treat neurological disorders. Prodigiosins (PDGs), 2-methyl-3-pentyl-6-methoxyprodiginine, are atripyrrole red pigments produced by many microorganisms, including Serratia marcescens and Serratia rubidaea.15 These bacterial secondary metabolites are characterized by their varied ranges of therapeutic benefits and bioactivities, such as antibacterial, antimalarial, antifungal,16 anticancer,17 antimetastatic,18 and immunosuppressive functions.19 Intriguingly, PDGs can suppress lipopolysaccharide (LPS)-induced inflammatory responses by inhibiting the activation of c-Jun N-terminal kinase (JNK), p38 mitogen-activated kinase (MPAK), and nuclear factor kappa B (NF-κB).20 However, the neuroprotective potency of PDGs has not received sufficient attention.
Selenium (Se) is a primary trace element with enormous human health benefits and unique biological and pharmacological properties, including protective effects against neurodegenerative diseases.21 Se can interact in the dynamics of the electron transport chain, modulate neurogenesis, mediate Ca2+ transport in neurons, and maintain redox balance.22 Several Se-based nanoformulations have been developed and tested for efficacy against neurological diseases. For example, epigallocatechin-3-gallate-stabilized Se nanoparticles were found to inhibit amyloidβ (Aβ) aggregation in an Alzheimer’s disease model.23 Sialic acid and resveratrol modified with Se nanoparticles were able to effectively suppress Cu2+-mediated Aβ aggregation and promote fibril disaggregation into non-toxic oligomers.24,25 In addition, Se nanoparticles displayed powerful antioxidant potency, and selenoproteins promote endogenous antioxidant activities in brain tissue.22,26
The primary purpose of the current study was to explore the antidepressant and neuroprotective activity of PDGs-SeNPs against CUMS-induced depression-like behaviors in rats by evaluating behavioral changes, redox status, and inflammatory and apoptotic responses, in parallel with the evaluation of hormonal and neurochemical alterations in hippocampal tissue that are associated with the development of depression.
Materials and Methods
Synthesis of Prodigiosin-Loaded Selenium Nanoparticles (PDGs-SeNPs)
A volume of 10 mL of 10 mM sodium selenite (Na2SeO3) was mixed with 10 mL of PDGs (3.5 mg/mL) with magnetic stirring for 24 h. The obtained mixture (PDGs-SeNPs) was lyophilized using a vacuum freeze dryer (LabconcoFreezone 4.5 Liter Freeze Dry System, Marshall Scientific, Hampton NH, USA), and the obtained powder was used in the current investigation.
Characterization of the Prepared Nanoparticles (PDGs-SeNPs)
The average diameter, size distribution, and surface charges of PDGs-SeNPs were determined by the Zetasizer Nano ZS particle analyzer (Zetasizer Nano ZS90, MalvernPanalytical, UK). The molecular structure of PDGs-SeNPs was detected using Fourier-transform infrared spectroscopy (FTIR; PerkinElmer, USA). The crystalline size of the nanoparticles was characterized by X-ray diffraction (XRD; Malvern Panalytical, UK). Moreover, transmission electron micrographs were recorded using a high-resolution transmission electron microscope (TEM; JEOL Ltd., Japan) equipped with an electron diffraction pattern.
Animals
Sixty healthy male Sprague Dawley rats (weight 150 ± 10 g) were purchased from the animal house of King Fahd for Medical Research, King Abdulaziz University, Jeddah, Saudi Arabia. All experimental protocols were approved by the Committee of Research Ethics for Laboratory Animal Care, Taif University (approval no. HAO-02-T-105). The 8th edition of the National Institute of Health for laboratory animals uses guidelines were followed in this procedure.
Before starting the experiment, the rats were maintained for two weeks in the laboratory animal room, with an indoor temperature of 21–25°C, relative humidity of 45%–55%, and a light:dark cycle of 12:12 h. Commercial normal diet and clean drinking water ad libitum were provided to the animals.
Chronic Unpredictable Mild Stress (CUMS) Model
The establishment of CUMS was performed following the schedule previously described by Mao et al,27 with minor modifications. Briefly, the non-stressed control rats were maintained in standard plastic cages in another room, separate from the stressed rats. For the stress-exposed groups, each rat was separately assigned to one cage and subjected to the CUMS stress regime, which consisted of 24-h exposure to water deprivation, food deprivation, the presence of a foreign object, or moist bedding (100 g sawdust bedding in 200 mL water), combined with 6-min forced swimming (8°C), 2-h restraint in an empty water bottle, 7-h cage tilt (45°C), exposure to white noise for 1-hand light exposure overnight. The stressors were applied twice a day in a random order for four weeks while avoiding applying the same stressor on two consecutive days.
Experimental Protocol
Following the acclimatization period, the animals were randomly distributed into six groups (n= 10 animals /group) as follows: control (Control) group, unstressed rats were orally administered distilled water; CUMS group (depression model), rats were exposed to CUMS and orally administered distilled water; Flu+CUMS group, rats were exposed to CUMS and orally administered fluoxetine (Flu, 5 mg/kg/day)28 suspended in distilled water; PDGs group, rats were exposed to CUMS rats and gavaged with prodigiosin (PDGs, 300 mg/kg/day);29 Na2SeO3+CUMS group, rats were exposed to CUMS and gavaged Na2SeO3(2 mg/kg/day).28 PDGs-SeNPs+CUMS group, rats were exposed to CUMS and gavaged with PDGs-SeNPs (0.5 mg/kg/day).30 All treatments lasted for 28 consecutive days. Fluoxetine and sodium selenite were obtained from Sigma-Aldrich (St. Louis, MO, USA).
Behavioral Tests
Open-Field Test (OFT)
The open-field test (OFT) was performed as described by Dong et al.31 The open-field apparatus was a four-sided, black, square cage (100 × 100 × 40 cm), divided into 25 equal small squares using black lines. Each animal was placed alone at the center of the testing area, and their behavior was monitored freely for 5 min. The distance moved, vertical activity (rearing or wall climbing), the total number of entries to the periphery and central zone, the time spent in the center zone, and grooming activity were carefully recorded. After each test, the cage was completely cleaned with 75% alcohol.
Sucrose Preference Test (SPT)
The sucrose preference test (SPT) was conducted according to the methods previously described by Özkartal et al.32 Briefly, after 4 weeks of CUMS exposure, the animals were maintained individually in cages and provided with two bottles of sucrose solution (1%, w/v) on the first day. One of the bottles was replaced with a bottle of tap water for the second day. After the adaptation period, rats were denied nourishment and water for 24 h. Rats were then allowed free access to two bottles, one containing sucrose solution (1%, 100 mL) and the other containing tap water (100 mL), for approximately 3 h. The consumed volumes of sucrose solutions and tap water were recorded. The sucrose preference was assessed as the ratio between sucrose solution consumption and total liquid consumption [Sucrose preference (%) = sucrose consumption/(water consumption + sucrose consumption) ×100%].
Measurement of Body Weight
Each individual rat was weighed using an electronic balance every Monday (9:00 am). The overall body weight was calculated as the mean of the weekly body weights (g).
Sampling and Hippocampal Tissue Preparation
Twenty-four hours after behavioral testing, the rats were anesthetized using intraperitoneally injected ketamine (100 mg/kg) and xylazine (10 mg/kg). Individual intracardiac blood samples were collected from different groups in labeled tubes, centrifuged at 2000 x g for 10 min and stored at –80°C until use for hormonal level estimation. Additionally, the hippocampus was rapidly excised and washed twice with isotonic saline. The tissue samples were processed for tissue homogenates, gene expression analysis, and histopathological evaluation. All the biochemical markers were estimated in the hippocampal tissue homogenate (10% w/v), which was homogenized in ice-cold phosphate-buffered saline (10 mM; pH 7.4). Monoamine neurotransmitters were evaluated in hippocampal tissue homogenized in 75% methanolic high-performance liquid chromatography (HPLC) buffer (10% w/v). Both homogenates were then centrifuged at 4000 x g for 10 min, and the resulting supernatants were stored at −80 °C.
Determination of Oxidative Stress Indices in the Hippocampal Tissue
The levels of the lipid peroxidation by-product malondialdehyde (MDA) and nitric oxide (NO) in the hippocampal tissue were determined using a colorimetric assay, as reported by Ohkawa et al33 and Green et al,34 respectively. The assessment of antioxidant indices, including catalase (CAT), superoxide dismutase (SOD), glutathione (GSH), glutathione peroxidase (GPx), and glutathione reductase (GR), was performed according to the established protocols of Aebi,35 Nishikimi et al,36 Ellman,37 Paglia et al,38 and Smith et al,39 respectively. Hippocampal protein levels were determined for all measurements using the protocols described by Bradford.40
Determination of Inflammatory Mediators in the Hippocampal Tissue
Specific enzyme-linked immunosorbent assay (ELISA) kits for rats (Novus Biologicals, Centennial, CO, USA) were employed to assess the levels of inflammatory mediators in the hippocampal tissue, including tumor necrosis factor-α (TNF-α; catalog No.: NBP1-92681), interleukin-1β (IL-1β; catalog No.: NBP1-92,02), interleukin-6 (IL-6; catalog No.: NBP1-92697), interleukin-10 (IL-10; catalog No.: NBP1-92701), prostaglandin E2 (PGE2; catalog No.: NBP1-02321), and cyclooxygenase-2 (COX-2; catalog No.: NB600-971), according to the manufacturer’s guidelines.
Determination of Apoptotic Proteins in the Hippocampal Tissue
Hippocampal levels of B cell lymphoma 2 (Bcl-2) and Bcl-2-associated X protein (Bax) were quantified using ELISA kits purchased from Elabscience (Houston, TX, USA). Caspase-3 activity was estimated colorimetrically using kits obtained from Sigma-Aldrich (St. Louis, MO, USA).
Determination of Monoamine Levels in the Hippocampal Tissue
Hippocampal levels of serotonin (5-HT), dopamine (DA), and norepinephrine (NE) were estimated by HPLC using an electrochemical detector, according to the technique described by Pagel et al.41 Acetonitrile (87:13, v/v) and sodium citrate buffer (pH 4.5) were employed as the mobile phase. Frozen hippocampal tissue homogenates were filtered using nylon filters (0.22 mm) before injection into the HPLC system. Elution was performed at a 0.8 mL/min flow rate, and the working electrode potential was adjusted to 0.75 V. The standard curves were generated via the injection of standard solutions of various known concentrations into the HPLC system (Supplementary Materials; Figure S1) and were used to calculate the final concentrations of all tested neurotransmitters.
Determination of Acetylcholinesterase (AChE), Monoamine Oxidase (MAO) Activities and Brain-Derived Neurotrophic Factor (BDNF) in the Hippocampal Tissue
Hippocampal acetylcholinesterase (AChE) activity was determined using the colorimetric protocol reported by Elman et al.42 MAO activity was estimated fluorometrically at 550 nm (excitation wavelength) and 404 nm (emission wavelength), using 5-hydroxytryptamine (500 mM) as a substrate, according to the procedure described by Dar et al.43 The hippocampal level of brain-derived neurotrophic factor (BDNF) was assayed using ELISA kits obtained from Abcam (Catalog. No.: ab213899), according to the manufacturer’s instructions.
Determination of Serum Hormonal Levels
Serum cortisol levels were determined using rat-specific ELISA kits (catalog No.: K7430-100), purchased from BioVision (California, USA). Serum adrenocorticotropin hormone (ACTH) and thyroid-stimulating hormone (TSH) were assayed using the ELISA methods previously described by Arlt and Stewart44 and Nicoloff and Spencer,45 respectively.
Quantitative Real-Time PCR
The TRIzol method (Invitrogen, Carlsbad, CA, USA) was used to extract total RNA from hippocampal tissues. The corresponding cDNA was reverse transcribed from the extracted RNA using the Script cDNA synthesis kit (Bio-Rad, CA). The relative expression of each target gene was measured using SYBR Green PCR kits (Qiagen, Heidelberg, Germany) and specific primers for each gene on an Applied Biosystems 7500 Instrument. The primer sequences for nitric oxide synthase (Nos2), nuclear factor erythroid 2-related factor 2 (Nrf2, Nfe2l2), and heme oxygenase 1 (HO-1, Hmox1) are summarized in Table 1. The glyceraldehyde 3-phosphate dehydrogenase (Gapdh) gene (internal control gene) was used to normalize all target gene expression levels. PCR amplification conditions were 10 min at 95°C, followed by 40 cycles of 1 min at 94°C for denaturation, 1 min at 55°C for annealing, and 20 s at 72°C for extension. The relative fold-change levels of all target gene expression levels were calculated using the comparative 2−ΔΔCTmethod.46
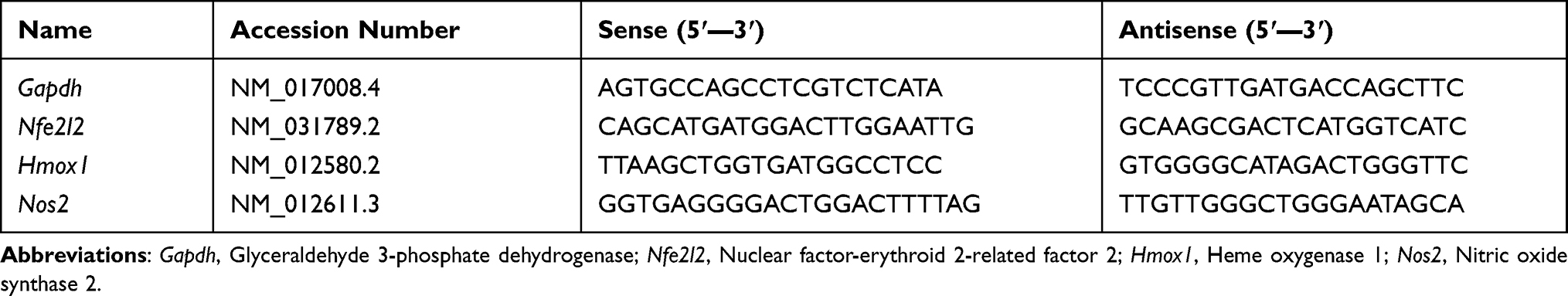 |
Table 1 Primer Sequences of Genes Analyzed in Real Time-PCR |
Light Microscopy and Immunohistochemical Examinations
For histopathological examinations, hippocampal tissue specimens were fixed in buffered neutral formalin (10%). Hippocampal specimens were dehydrated, embedded in paraffin, and sectioned (4–5 µm). Sections were stained using hematoxylin and eosin (H&E) and analyzed using a Nikon Eclipse E200-LED microscope (Tokyo, Japan, 200x and 400× magnification). The degree of hippocampal damage was characterized using a semi-quantitative scoring system, with five grades assigned according to the severity of injury, as follows: 1, minimal injury (<1%); 2, slight injury (1%–25%); 3, moderate injury (26%–50%); 4, moderate/severe injury (51%–75%); and 5, severe injury (76%–100%).47 To detect glial fibrillary acidic protein (GFAP), hippocampal tissue sections were incubated overnight with an anti-GFAP antibody (Santa Cruz, CA, USA) at 4°C, followed by incubation with biotinylated secondary antibody at room temperature for 1-h. The 3,3ʹ-diaminobenzidine (DAB) peroxidase substrate kit was used to develop a brown color. The images were observed at a magnification of 400×, using a Nikon Eclipse E200 LED (Tokyo, Japan).
Data Analysis
One-way analysis of variance (ANOVA), followed by Duncan's post hoc test, was used to determine significant differences between biochemical and gene expression parameters using SPSS 26.0. To visualize the data, the mean ± standard deviation (SD) was recorded. A P-value lower than 0.05 was considered significant.
Results
Physical and Chemical Characterization of PDGs-SeNPs
After adding PDGs to sodium selenite, the solution was turned from colorless to red color. Size distribution by intensity revealed that the average size of the examined SeNPs was 121.7 nm (Figure 1A) with a mean zeta potential of −23.3 mV which indicates the moderate stability of SeNPs biosynthesized with PDGs (Figure 1B). The FT-IR investigation result of synthesized PDGs-SeNPs is illustrated in Figure 1C. A broad peak observed at 3306.85 cm−1 corresponds to O–H of alcohols and phenols structure. The absorption peak at 2130.72 cm−1 corresponds to CΞCalkyne groups. The peak at 1638.39 cm−1 is due to the C=O asymmetric stretch carbon compounds. The absorption band at 1438.07 cm−1 is due to the presence of O-H bending of alcoholic group. Whereas, the band at 1140.81 cm−1 is attributed to the C-O stretching bond corresponding to aliphatic ether group. The absorption peak at 1010.68 cm−1 is attributed to the CO-O-CO stretching of the anhydride. C=C stretching in alkene causes a band at 956.98cm−1. Finally, C-Se stretching in metal compound causes is indicated by the presence band at 614.87 cm−1. These results reveal the presence of various functional groups that may be responsible for both reduction and stabilization of the PDGs-SeNPs. Furthermore, the XRD pattern records revealed a broader pattern without any definite Braggs peaks. The obtained results indicate that PDGs-SeNPs are not crystalline; rather they are more amorphous (Figure 1D). The prepared PDGs-SeNPs calculated crystalline size is < 134 nm using Scherrer’s equation. This amorphous character is in agreement with the earlier studies carried out with lycopene48 and different bacterial sp.49 Moreover, TEM is employed to analyze the shape of the PDGs-SeNPs biosynthesized with PDGs. TEM image of PDGs-SeNPs revealed spherical particles within the diameter < 120–150 nm. These particles were well distributed with mild aggregation (Figure 1E). Moreover, based on our observations, there was no sign of aggregation and the color was stable (deep red color) until 40 days.
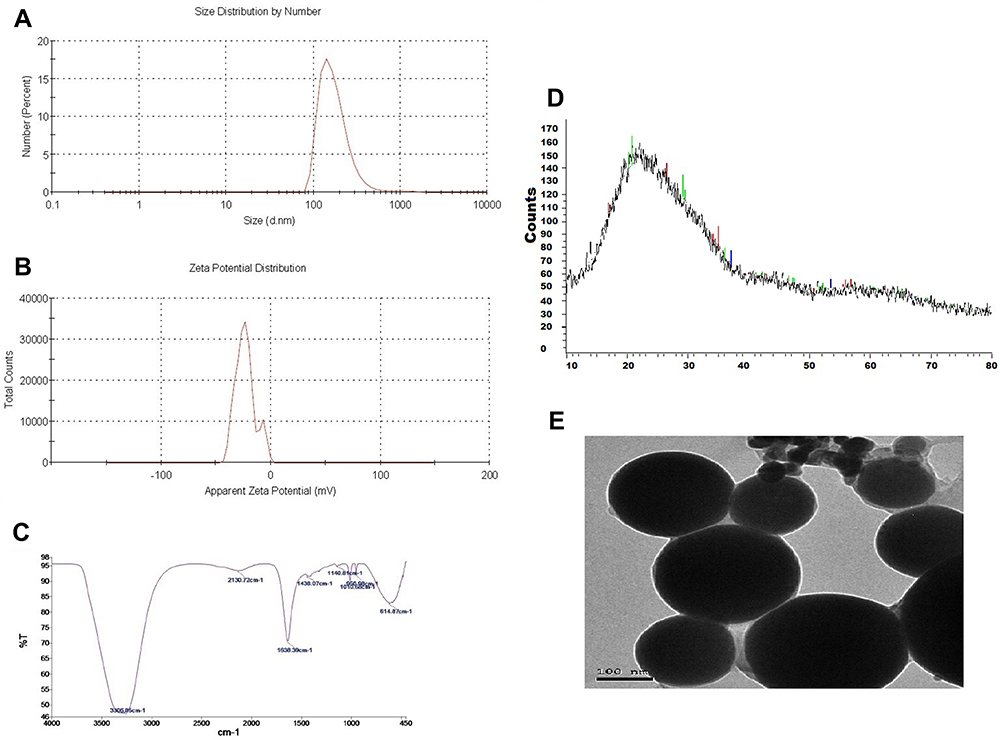 |
Figure 1 The prodigiosin-loaded selenium nanoparticles characterization, (A) Size distribution histogram as measured by a Malvern Zetasizer, (B) Surface charge histogram as measured by Zeta potential. (C) FT-IR spectra as detected using Fourier-transform infrared spectroscopy. (D) XRD spectra of PDGs-SeNPs. (E) Morphological shape of PDGs-SeNPs as observed by TEM. |
Effect of PDGs-SeNPs Treatment on Behavioral and Body Weight Changes Following CUMS Exposure
The impacts of PDGs-SeNPs treatment on CUMS-induced depression-like behaviors were measured using the OFT and SPT in the current study (Figure 2). The OFT total score was significantly reduced (p< 0.05) in the CUMS rats compared with that in control rats. Rats treated with Flu, PDGs, Na2SeO3, and PDGs-SeNPs exhibited significant improvements (p< 0.05) in OFT scores compared with the CUMS group. Additionally, sucrose consumption (%) was significantly diminished (p< 0.05) in the CUMS group compared with the control group. Although all treatments significantly (p< 0.05) restored sucrose consumption to near-normal levels, PDGs-SeNPs treatment appeared to be the most effective when compared with no treatment in CUMS model rats (Figure 2). Body weight declined significantly (p< 0.05) in the group exposed to CUMS for 28 consecutive days compared with that in the unstressed control group. Treatment with Flu, Na2SeO3, PDGs alone, or PDGs-SeNPs significantly (p< 0.05) increased the body weights of CUMS-exposed rats (Figure 2).
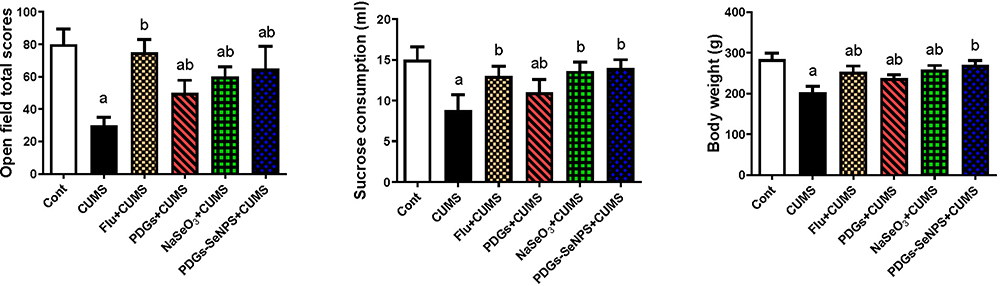 |
Figure 2 Protective impacts of prodigiosin (PDGs), sodium selenite (Na2SeO3), and prodigiosin-loaded selenium nanoparticles (PDGs-SeNPS) administered orally to CUMS-exposed rats on the open-field test, sucrose consumption percentage, and body weight changes. Data were analyzed using one-way ANOVA, followed by Duncan’s post hoc test (p< 0.05), and are expressed as the mean ± SD (n=10 rats/group). aSignificant difference relative to the control group (Non-stressed). bSignificant difference relative to the CUMS group (depression model). CUMS: chronic unpredictable mild stress. |
Effect of PDGs-SeNPs Treatment on Hippocampal Oxidative Damage Following CUMS Exposure
The hippocampal tissue in CUMS-exposed rats demonstrated a disrupted oxidative status, characterized by increased MDA and NO production and the upregulation of Nos2 mRNA expression. These changes were accompanied by a significant decrease (p< 0.05) in the levels of endogenous antioxidant proteins, including SOD, CAT, GSH, and its derived enzymes (GPx and GR), compared with those in the control group. The administration of PDGs alone or PDGs-SeNPs significantly inhibited (p< 0.05) the development of oxidative insults following exposure to CUMS by enhancing the levels of the examined antioxidant proteins and depleting the levels of pro-oxidants in hippocampal tissue. Similarly, Na2SeO3 treatment significantly prevented (p< 0.05) oxidative damage associated with CUMS (Figures 3 and 4).
 |
Figure 3 Protective impacts of prodigiosin (PDGs), sodium selenite (Na2SeO3), and prodigiosin-loaded selenium nanoparticles (PDGs-SeNPS) administered orally to CUMS-exposed rats on hippocampal mRNA and levels of oxidative stress indicators. Data were analyzed using one-way ANOVA, followed by Duncan’s post hoc test (p< 0.05), and are expressed as the mean ± SD (n=10 rats/group). aSignificant difference relative to the control group (Non-stressed). bSignificant difference relative to the CUMS group (depression model). Abbreviations: MDA, malondialdehyde; NO, nitric oxide; Nos2, inducible nitric oxide synthase; CUMS, chronic unpredictable mild stress. |
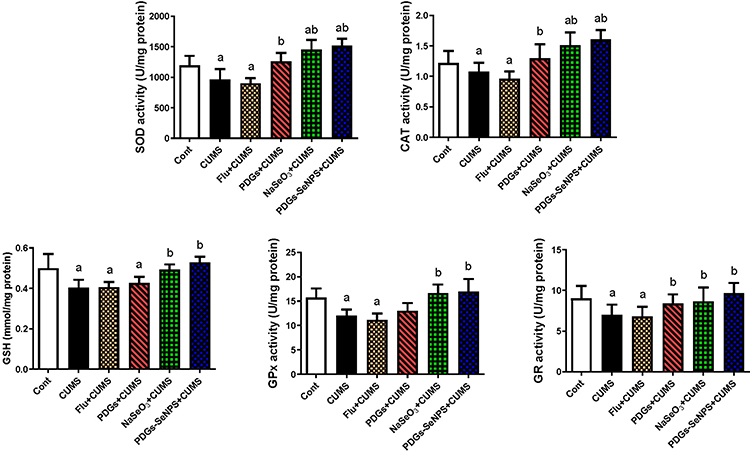 |
Figure 4 Protective impacts of prodigiosin (PDGs), sodium selenite (Na2SeO3), and prodigiosin-loaded selenium nanoparticles (PDGs-SeNPS) administered orally to CUMS-exposed rats on hippocampal antioxidant enzyme activities. Data were analyzed using one-way ANOVA, followed by Duncan’s post hoc test (p< 0.05), and are expressed as the mean ± SD (n=10 rats/group). aSignificant difference relative to the control group (Non-stressed). bSignificant difference relative to the CUMS group (depression model). Abbreviations: CAT, catalase; SOD, superoxide dismutase; GSH, glutathione; GPx, glutathione peroxidase; GR, glutathione reductase; CUMS, chronic unpredictable mild stress. |
To elucidate the molecular mechanism underlying the antioxidant properties of PDGs-SeNPs treatment following CUMS exposure, Nfe2l2 and Hmox1 mRNA expression levels were investigated. Rats exposed to CUMS exhibited the downregulation of Nrf2 and HO-1 compared with the expression levels observed in the control group. By contrast, both antioxidant promoters were upregulated in the hippocampal tissue following the administration of PDGs alone or PDGs-SeNPs relative to the levels observed in the untreated CUMS group (Figure 5), demonstrating the potent antioxidant capacities of PDGs and PDGs-SeNPs against hippocampal oxidative damage induced by CUMS exposure (Figure 5).
 |
Figure 5 Protective impacts of prodigiosin (PDGs), sodium selenite (Na2SeO3), and prodigiosin-loaded selenium nanoparticles (PDGs-SeNPS) administered orally to CUMS-exposed rats on the mRNA expression levels of Nfe2l2 and Hmox1. Data were analyzed using one-way ANOVA, followed by Duncan’s post hoc test (p< 0.05), and are expressed as the mean ± SD (n=10 rats/group). aSignificant difference relative to the control group (Non-stressed). bSignificant difference relative to the CUMS group (depression model). Abbreviations: Nfe2l2, Nuclear factor erythroid 2-related factor 2; Hmox1, heme oxygenase-1; CUMS, chronic unpredictable mild stress. |
Effects of PDGs-SeNPs Treatment on Neuroinflammation Following CUMS Exposure
CUMS-induced depression caused neuronal inflammation, identified by significant elevations (p< 0.05) in the hippocampal levels of pro-inflammatory cytokines (TNF-α, IL-1β, and IL-6) and other inflammatory mediators (PGE2 and COX-2), the upregulation of NF-κB, and reduced IL-10 levels compared with the levels observed in the control group. However, these hippocampal inflammatory responses were significantly suppressed in all treated groups, especially in PDGs and PDGs-SeNPs treated rats, compared with the CUMS group, indicating the potent anti-inflammatory activity of these treatments in the CUMS-induced depression model (Figure 6).
 |
Figure 6 Protective impacts of prodigiosin (PDGs), sodium selenite (Na2SeO3), and prodigiosin-loaded selenium nanoparticles (PDGs-SeNPS) administered orally to CUMS-exposed rats on hippocampal levels of neuroinflammatory markers. Data were analyzed using one-way ANOVA, followed by Duncan’s post hoc test (p< 0.05), and are expressed as the mean ± SD (n=10 rats/group). aSignificant difference relative to the control group (Non-stressed). bSignificant difference relative to the CUMS group (depression model). Abbreviations: TNF-α, tumor necrosis factor-alpha; IL-1β, interleukin-1β; IL-6, interleukin-6; IL-10, interleukin-10; PGE2, prostaglandin E2; COX-2, cyclooxygenase-2; NF-κB, nuclear factor kappa-B transcription complex; CUMS, chronic unpredictable mild stress. |
Effects of PDGs-SeNPs Treatment on Apoptotic Proteins Following CUMS Exposure
To explore neuronal apoptotic events in the CUMS-induced depression model rats and the potential anti-apoptotic role of PDGs-SeNPs treatment, the levels of Bcl-2 and Bax and caspase-3 activity were examined in hippocampal tissue. Compared with the control group, rats exposed to CUMS exhibited significant elevations (p< 0.05) in the levels of apoptogenic proteins (Bax and caspase-3), whereas a significant reduction (p< 0.05) in the Bcl-2 level (anti-apoptotic protein) was observed. However, PDGs-SeNPs administration prevented the apoptotic cascade and reversed the CUMS-exposure-induced changes in apoptotic proteins compared with the untreated CUMS levels, indicating the protective role played by PDGs-SeNPs against neuronal loss following CUMS exposure. Flu and NaSeO3 treatments also restored alterations in the apoptotic proteins near the normal levels when compared with the control group (Figure 7).
 |
Figure 7 Protective impacts of prodigiosin (PDGs), sodium selenite (Na2SeO3), and prodigiosin-loaded selenium nanoparticles (PDGs-SeNPS) administered orally to CUMS-exposed rats on hippocampal levels of apoptosis markers. Data were analyzed using one-way ANOVA, followed by Duncan’s post hoc test (p< 0.05), and are expressed as the mean ± SD (n=10 rats/group). aSignificant difference relative to the control group (Non-stressed). bSignificant difference relative to the CUMS group (depression model). Abbreviations: Bcl2, B cell lymphoma 2; Bax, Bcl-2-associated X protein; CUMS, chronic unpredictable mild stress. |
Effects of PDGs-SeNPs Treatment on Hormonal Levels Following CUMS Exposure
Serum cortisol and ACTH levels were significantly increased (p< 0.05), whereas TSH levels were decreased in CUMS-exposed rats compared with those in control animals. The levels of the tested hormones returned to near-normal values, and the maximum effect was observed in the PDGs-SeNPs treated group (Figure 8).
 |
Figure 8 Protective impacts of prodigiosin (PDGs), sodium selenite (Na2SeO3), and prodigiosin-loaded selenium nanoparticles (PDGs-SeNPS) administered orally to CUMS-exposed rats on serum hormonal levels. Data were analyzed using one-way ANOVA, followed by Duncan’s post hoc test (p< 0.05), and are expressed as the mean ± SD (n=10 rats/group). aSignificant difference relative to the control group (Non-stressed). bSignificant difference relative to the CUMS group (depression model). Abbreviations: ACTH, Adrenocorticotropic hormone; TSH, Thyroid-stimulating hormone; CUMS, chronic unpredictable mild stress. |
Effects of PDGs-SeNPs Treatment on the Levels of Monoamines, AChE, MAO, and BDNF Following CUMS Exposure
A disturbance in cholinergic and monoaminergic neurotransmission was observed in CUMS-exposed rats, as indicated by decreased levels of 5-HT, DA, and NE and reduced AChE activity in hippocampal tissue. Additionally, the CUMS-exposed rats revealed a significant increase in MAO activity compared with that in the control group. Interestingly, PDGs administration (either alone or as PDGs-SeNPs) significantly restored (p< 0.05) the levels of these neurotransmitters, and AChE and MAO activities were restored to near-normal values, demonstrating the potent neuromodulatory role played by PDGs-SeNPs against CUMS-mediated depression-like symptoms. Flu and Na2SeO3administration were also found to normalize the changes in the examined neurotransmitters to levels similar to those observed in the unstressed control group (Table 2).
 |
Table 2 Protective Impacts of Prodigiosin (PDGs), Sodium Selenite (NaSeO3), and Prodigiosin-Loaded Selenium Nanoparticles (PDGs-SeNPs) Administered Orally to CUMS-Exposed Rats on Hippocampal Monoamine Levels and Acetylcholinesterase and Monoamine Oxidase Activities |
CUMS-exposed rats exhibited a marked decrease (p< 0.05) in BDNF levels compared with that in the control rats. However, all treated groups showed a significant increase (p< 0.05) in BDNF levels (which returned to the control level) when compared with theCUMS group (Table 2).
Effects of PDGs-SeNPs Treatment on Histological Alteration in the Hippocampal Tissue Following CUMS Exposure
As presented in Figure 9, the hippocampal tissue sections obtained from the CUMS group exhibited severe neural cell degeneration, indicated by the presence of apoptotic neurons, pyknotic nuclei, vacuolation in the intraneuronal space, and inflammatory cell infiltration. By contrast, the groups that received PDGs and PDGs-SeNPs administration markedly improved hippocampal structures, and the development of neuronal impairments associated with CUMS was prevented (Supplementary Materials: Figure S2).
 |
Figure 9 Protective impacts of prodigiosin (PDGs), sodium selenite (Na2SeO3), and prodigiosin-loaded selenium nanoparticles (PDGs-SeNPS) administered orally to CUMS-exposed rats on hippocampal histological alterations and glial fibrillary acidic protein (GFAP) expression. Black arrow: blood vessels congestion; red arrow: apoptotic neurons; blue arrow: neuron edema and degeneration. |
Effects of PDGs-SeNPs Treatment on Glial Fibrillary Acidic Protein (GFAP) Expression in Hippocampal Tissue Following CUMS Exposure
Immunohistochemical examination revealed the downregulation in GFAP immunoreactivity in CUMS-treated rats compared with that in the control group, indicating astrocytic deactivation. However, PDGs-SeNPs administration effectively upregulated the GFAP immunoreactivity in hippocampal tissue following CUMS exposure (Figure 9).
Discussion
Depression is a multifactorial disease that involves both hereditary and environmental factors, and identifying the underlying pathways associated with the development of depression is urgently necessary to gain new insights regarding how to prevent and treat depression using newly developed pharmaceutical strategies.50 The currently available classical antidepressant drugs are typically associated with reduced efficacy in some populations and the occurrence of adverse effects.51 The use of metal-based nanoformulations has become a promising trend in the pharmaceutical industries to the enhanced bioavailability, delivery progression, and drug inflow to the target tissues provided by these treatment formulations compared with typical drug formulations.52 Accumulating evidence has indicated the potential role of Se-based nanoparticles for the treatment of neural diseases.30 However, the role and mechanisms through which Se might regulate depressive disorders have not yet been fully determined. To the author’s knowledge, this study is the first to investigate the neuroprotective effects of PDGs, either alone or in the form of stabilized-selenium nanoparticles (PDGs-SeNPs), in a CUMS-induced depression-like animal model. Nanoparticles’ size has a significant effect on their biological activity. Generally, nanoparticles with a smaller size are more active than those with a bigger size.53 In the case of elemental Se, certain properties are certainly size-dependent. It is physiologically inactive above the micrometre size range.54 SeNPs have been shown to have a size effect on redox reactivity,55 and SeNPs with a size range of 5–200 nm have a size-dependent effect on directly scavenging several free radicals in vitro, including 1,1-diphenyl-2-picrylhydrazyl and superoxide anion.56 According to Pelyhe and Mézes57 and Wang et al,58 selenium at the nanoscale exhibits a reduced hazardous effect. Additionally, these researchers demonstrated that preparing selenium in the form of nanoselenium increases selenium bioavailability in the body.
One of the most commonly applied tests for evaluating anxiety-like behavior is the OFT, which can also be used to assess spontaneous activity in rodents.59 The SPT is commonly used to evaluate notable depression-like symptoms in rodents, anhedonia, or the lack of pleasure.32 Hu et al9 reported reduced horizontal and vertical movement, locomotion latencies, and prominent reduction of number of central zone entries in CUMS rats. A substantial reduction in sucrose solution consumption has also been reported in CUMS-exposed animals.7 Homogeneity in the reported responses of CUMS-treated animals in both the OFT and SPT has been identified across multiple studies.10,31,60 The observed changes in OFT performance in CUMS-exposed animals relative to unstressed animals indicate that CUMS exposure has a significant influence on anxiety, exercise, and exploration levels.61 The observed reduction in sucrose consumption in the SPT reflects the suppression of the brain reward system associated with CUMS exposure, which serves as a proxy for depression in rodents.62 CUMS has also been associated with significant changes in body weight, which aligns with previous studies and likely reflects combined effects of stress, food deprivation, and sucrose consumption paradigms used during the CUMS protocol.31,60,63 Comparably, Zhang et al64 reported that animals exposed to the CUMS paradigm showed reduced body weight gains relative to non-stressed animals. Following our discriminate analysis, we observed diminished OFT scores, reduced sucrose intake, and lower body weights in rats exposed to CUMS for four continuous weeks compared with those measures in control unstressed rats. However, our study showed that these changes could be ameliorated by PDGs and PDGs-SeNPs administration. This finding agrees with the findings of Ebokaiwe et al,65 Ali et al,66 and Yuan et al,30 who reported that SeNP administration attenuated neurobehavioral alterations following brain injury. Our study is considered the first investigation to assess the modification of depression-related behaviors through the administration of PDGs, either alone as PDGs-SeNPs in CUMS-exposed rats.
Oxidative stress plays a fundamental role in the pathogenesis and progression of psychiatric diseases, including depression or bipolar disorder.67 One of the pathophysiologies of depression is the repeated or unpredictable exposure to stress, which results in the upregulation of ROS generation and secretion, the depletion of innate antioxidant proteins, and the induction of neurodegeneration and necrosis.68 The current study examined CUMS-induced hippocampal oxidative injury and identified the elevation of MDA and NO levels and Nos2 mRNA levels, in addition to the suppression of the antioxidant system, indicated by reduced antioxidant molecule activities, including GSH, GPx, GR, SOD, and GR, and the downregulation of the Nrf2/HO-1 pathway. PDGs-SeNPs administration protected hippocampal tissues by enhancing endogenous antioxidants and inhibiting pro-oxidant formation.
These results align with those reported by previous studies, in which a link between CUMS and the initiation of oxidative insults in brain tissues was demonstrated by increased levels of neuronal lipoperoxidation and NO production, which was associated with a reduction in antioxidant markers and enzymes.6,69,70 This imbalance between oxidants and antioxidants has been attributed to ROS overproduction. In support of previous studies, our findings indicated the significant downregulation of Nfe2l2 and Hmox1 mRNA expression levels in the hippocampal tissue of CUMS-exposed rats. The deactivation of the Nrf2/HO-1 signaling pathway plays an essential role in the development of oxidative injury following CUMS exposure.71 Nrf2 is a transcriptional mediator with cytoprotective impacts mediated by controlling the expression of an array of antioxidant proteins, such as HO-1.72 The antioxidant activity of Se has been correlated with the incorporation of Se in selenoproteins and selenoenzyme structures, which can dampen ROS and reduce oxidative damage.73 In accordance with our findings, the treatment of epileptic mice with Se nanoparticles restored neuronal oxidative damage through the upregulation of GR, SOD, GPx, and CAT mRNA expression, increased GSH levels, and reduced MDA and NO generation,30 which were attributed to the enhancement of antioxidant regulators (Nrf2/HO-1). Se nanoparticles have been reported to have potent ROS scavenging activity and boosted several endogenous antioxidant proteins due to the small size of Se, which allows for more Se molecules to accumulate and scavenge free radicals.74 Due to their small size and large surface area, SeNPs had a greater number of atoms exposed to free radicals for the electron exchanger, implying a greater capacity for scavenging multiple free radicals than larger sized antioxidant enzymes, particularly SOD and GPx. Thus, SeNPs were able to significantly suppress oxidative stress markers and restore non-enzymatic and enzymatic antioxidants.58 Previous investigations examining Se or SeNPs have indicated their abilities to improve the activities and expression of endogenous antioxidant proteins and suppress ROS production following neuronal damage.66,75,76 Chitosan-stabilized Se nanoparticles also displayed more antioxidant power toward acrylamide-induced neural damage, as demonstrated by reduced cerebral MDA and NO levels and increased GSH levels.77 The antioxidant potential of PDGs may also be associated with free radical-scavenging properties and the ability to interfere with NOS activity to suppress NO production.20 PDGs have also been demonstrated to effectively inhibit ROS production associated with hypoxia-mediated brain injury by minimizing nitrotyrosine formation and hindering blood–brain barrier leakage, which suppresses CD11b leukocyte infiltration.78 Similarly, PDGs administration prevented the imbalance between gastric oxidants and antioxidants induced by HCl/ethanol through reduced lipid peroxide and NO levels and elevated antioxidant enzyme and GSH levels.29
Neural inflammation and apoptosis are prevalent etiologic factors associated with neurodegenerative and depressive disorders.79 Following CUMS exposure, our results revealed the overproduction of pro-inflammatory cytokines (TNF-α, IL-1β, and IL-6), COX-2, PGE2, and NF-κB in the hippocampal tissue. Earlier studies have reported increased levels of pro-inflammatory cytokines in the prefrontal cortex of animal models of depression.4,59,80 The inflammatory response that follows CUMS exposure has been suggested to be associated with enhanced COX-2/PGE2-driven inflammatory pathways and the increased activation of NF-κB, leading to increased pro-inflammatory cytokines production.81,82 By contrast, the administration of Se nanoformulations exhibited neuroprotective activity via the suppression of the inflammatory cascade in epileptic mice and diabetic rats.30,65 PDGs-SeNPs administration also successfully prevented hippocampal inflammatory events associated with CUMS exposure. Similarly, treatment with SeNPs was shown to inhibit the release of pro-inflammatory cytokines and reduce microglial inflammatory responses through the suppression of NF-κB.66 The neural anti-inflammatory properties of PDGs were previously illustrated in oxygen-glucose deprivation (OGD)-activated microglial cells and middle cerebral artery occlusion/reperfusion (MCAO/R)-injured mouse brains, through the regulation of NF-κB activation and nuclear translocation. Additionally, PDGs can suppress the activation of the COX-2/PGE2 pathway and myeloperoxidase activity, which are considered to serve as vital mediators of tissue oxidative stress and inflammation.29
A major underlying component associated with neurodegenerative disorders, including depression, is neuronal cell death. Apoptosis is the primary form of cell death and is tightly linked to oxidative insults, either directly or through the activation of inflammatory pathways.83 Our results revealed a significant acceleration in the apoptosis rate in neuronal cells of CUMS-exposed rats, as demonstrated by a decline in the Bcl-2 level and an increase in Bax and caspase-3 levels. Previous studies have also reported disruptions in the Bcl-2/Bax regulatory pathways during depression.6,31,84 Emerging evidence suggests that neuronal cell death in depressed patients is coupled with excessive ROS levels and the activation of stress kinases, in addition to the activation of NF-κB.3 However, treatment with PDGs-SeNPs prevented CUMS-induced neuronal cell loss, as indicated by the decreased expression of pro-apoptotic proteins (Bax and caspase-3) the increased expression of the anti-apoptotic protein Bcl-2. Earlier reports have demonstrated the anti-apoptotic activity of Se and Se-based nanoformulations.29,30,85 The anti-apoptotic effects of SeNPs have been suggested to be associated with the ability to modulate Ca2+ homeostasis and decrease intracellular Ca2+ concentrations via the regulation of parvalbumin protein expression because increased Ca2+ levels induce apoptosis and necrotic cell death.65 Additionally, PDGs administration has been reported to stimulate anti-apoptotic protein (Bcl-2) expression and downregulate Bax and caspase-3 activation.86
The HPA axis plays a vital role in the stress response.60 The unusual release of cortisol has been associated with exposure to chronic stress.50,59,63 Here, we recorded a significant increase in cortisol and ACTH levels, accompanied by a decrease in the TSH level. In line with our study, CUMS exposure induced the elevation of serum ACTH and cortisol levels through the enhancement of HPA axis activity.4,6 The elevation of pro-inflammatory cytokine release following stress could activate the HPA, subsequently increasing cortisol secretion and inducing neuronal damage in brain tissue.87 Treatment with PDGs, either alone or in the form of PDGs-SeNPs, normalized serum levels of ACTH, TSH, and cortisol in CUMS-exposed rats, suggesting that these treatments were able to ameliorate the overactivity of the HPA axis mediated by stress. Se is necessary for peripheral corticosterone and ACTH metabolism and the activity of the selenoenzyme deiodinase, which catalyzes the deiodination of T4 into T3,88 which might explain the restoration of hormonal balance following SeNPs administration.
Although the specific pathophysiological mechanisms underlying depression are not yet completely clear, animal and human studies have illustrated that dysfunctional intracerebral neurotransmission, such as 5-HT, NE, and DA, is positively associated with the development of depression.80 In the current study, CUMS exposure disrupted cholinergic and monoaminergic transmission, as demonstrated by reduced hippocampal levels of AChE, 5-HT, DA and NE, which was associated with increased MAO activity. A previous report showed that the hyperactivity of the HPA axis is correlated with a reduction in 5-HT and NE levels in brain tissue.89 Additionally, a connection between the overproduction of inflammatory cytokines during stress and monoamine neurotransmitter metabolism has also been reported.90 Previous findings have demonstrated diminished levels of cholinergic and monoamine neurotransmitters in depressed animals.80,87 Liu91 also described reduced AChE activity in CUMS-exposed animals, resulting in acetylcholine accumulation and subsequent neurological consequences. The decrease in 5-HT content in depression has been correlated with reduced synaptosomal uptake and the suppression of tryptophan hydroxylase activity.92 In addition, increased cytokine expression can promote NO synthesis and minimize tetrahydropterpine, resulting in a decline in tyrosine hydroxylase activity, preventing the conversion of tyrosine into DA.93 NE depletion in neural disorders, including depression, is likely due to the downregulation of α1 receptor density or the suppression of the dopamine-β-hydroxylase enzyme that is responsible for NE synthesis.30,94 Another theory that might explain decreased monoamine levels in depression is increased MAO activity, which is involved in the monoamine degradation process95 and was observed in our results. Our study is the first to explore the potential neuroprotective effect of PDGs, either alone or in the form of PDGs-SeNPs, highlighting the ability of PDGs to modulate neurotransmission in hippocampal tissues, particularly monoamines. In agreement with our results, SeNPs administration elevated AChE activity and DA, NE, and 5-HT contents in an epileptic model induced by neuronal intoxication with cadmium or acrylamide.30,96,97 The authors attributed this outcome to the ability of Se to inhibit MAO activity, in addition to the ROS scavenging activity and anti-inflammatory potency of Se.97 The administration of Se was also shown to suppress neurodegeneration and reduce the loss of dopaminergic neurons in a Parkinson’s disease model.98
BDNF is a neurotrophic factor that contributes to the maintenance and survival of neurons and is involved in synaptic plasticity. Accumulating data suggest that BDNF plays an important role in the development of depression and has been considered as a potential therapeutic target for antidepressant drugs.99 Our findings showed a significant decline in BDNF levels in the hippocampal tissue following CUMS exposure. These results are in line with those of previous studies.100,101 Reduced BDNF levels correlate with ROS production and the development of oxidative damage following exposure to CUMS.6 PDGs and SeNPs co-treatment was shown to significantly counteract the prominent changes in BDNF levels, which could be attributed to their potent antioxidant capacities. Previous reports showed that Se and its nanoformulations could restore BDNF levels following neuronal damage.102,103
GFAP is the primary intermediate filament component of astrocytes and is widely used as a marker for astrocytic activation.104 In agreement with previous studies, our findings revealed a decrease in the immunoreactivity of hippocampal GFAP in CUMS-exposed rats,105,106 which was attributed to apoptosis induced by chronic stress, reduced neurogenesis, and increased neuronal atrophy and cell death.107 Treatment with both PDGs and PDGs-SeNPs was able to efficiently restore GFAP immunoreactivity, indicating the activation of astrocytes. Although no previous reports have examined the effects of PDGs, either alone or combined with SeNPs, on GFAP expression in brain tissue, Se and SeNPs were found to dampen apoptotic cell death processes in the hippocampus, increase GFAP expression, reduce reactive astrogliosis, increase neuronal survival, and mitigate overall neuronal damage.85,108
Conclusion
Based on our findings, SeNPs with a size range of 120–150 loaded with PDGs (PDGs-SeNPs) exerted neuroprotective effects against CUMS-induced depression-like behaviors, as demonstrated by ameliorated behavioral responses, the restoration of normal body weight, and the prevention of hippocampal oxidative insults associated with neuroinflammation and neuronal cells loss. Additionally, PDGs-SeNPs displayed a neuroprotective role through the modulation of disrupted monoaminergic and cholinergic transmission, increased BDNF levels, and the activation of GFAP in hippocampal tissue.
Funding
The authors extend their appreciation to the Deputyship for Research and Innovation, Ministry of Education in Saudi Arabia for funding this research work through the project number [135-1441-1].
Disclosure
The authors report no conflicts of interest for this work.
References
1. Kim YH, Im A, Park B-K, et al. Antidepressant-like and neuroprotective effects of ethanol extract from the root bark of hibiscus syriacus L. Biomed Res Int. 2018;2018:1–13. doi:10.1155/2018/7383869
2. Ferrari AJ, Charlson FJ, Norman RE, et al. The epidemiological modelling of major depressive disorder: application for the Global Burden of Disease Study 2010. PLoS One. 2013;8(7):e69637. doi:10.1371/journal.pone.0069637
3. Bakunina N, Pariante CM, Zunszain PA. Immune mechanisms linked to depression via oxidative stress and neuroprogression. Immunology. 2015;144(3):365–373. doi:10.1111/imm.12443
4. Liao D, Lv C, Cao L, et al. Curcumin Attenuates Chronic Unpredictable Mild Stress-Induced Depressive-Like Behaviors via Restoring Changes in Oxidative Stress and the Activation of Nrf2 Signaling Pathway in Rats. Oxid Med Cell Longev. 2020;2020:1–11. doi:10.1155/2020/9268083
5. Lopresti AL, Maker GL, Hood SD, Drummond PD. A review of peripheral biomarkers in major depression: the potential of inflammatory and oxidative stress biomarkers. Prog Neuropsychopharmacol Biol Psychiatry. 2014;48:102–111. doi:10.1016/j.pnpbp.2013.09.017
6. Li X, Wu T, Yu Z, et al. Apocynum venetum leaf extract reverses depressive-like behaviors in chronically stressed rats by inhibiting oxidative stress and apoptosis. Biomed Pharmacother. 2018;100:394–406. doi:10.1016/j.biopha.2018.01.137
7. Rai A, Gill M, Kinra M, et al. Catechin ameliorates depressive symptoms in Sprague Dawley rats subjected to chronic unpredictable mild stress by decreasing oxidative stress. Biomed Rep. 2019;11(2):79–84. doi:10.3892/br.2019.1226
8. Willner P. Chronic mild stress (CMS) revisited: consistency and behavioural-neurobiological concordance in the effects of CMS. Neuropsychobiology. 2005;52(2):90–110. doi:10.1159/000087097
9. Hu C, Luo Y, Wang H, et al. Re-evaluation of the interrelationships among the behavioral tests in rats exposed to chronic unpredictable mild stress. PLoS One. 2017;12(9):e0185129. doi:10.1371/journal.pone.0185129
10. Yu H, Fan C, Yang L, et al. Ginsenoside Rg1 prevents chronic stress-induced depression-like behaviors and neuronal structural plasticity in rats. Cell Physiol Biochem. 2018;48(6):2470–2482. doi:10.1159/000492684
11. Matreja PS, Badyal DK, Deswal RS, Sharma A. Efficacy and safety of add on low-dose mirtazapine in depression. Indian J Pharmacol. 2012;44(2):173. doi:10.4103/0253-7613.93843
12. Holtzheimer III PE, Nemeroff CB. Advances in the treatment of depression. NeuroRx. 2006;3(1):42–56. doi:10.1016/j.nurx.2005.12.007
13. Mojtabai R, Olfson M, Sampson NA, et al. Barriers to mental health treatment: results from the National Comorbidity Survey Replication (NCS-R). Psychol Med. 2011;41(8):1751. doi:10.1017/S0033291710002291
14. Licinio J, Wong M-L. Depression, antidepressants and suicidality: a critical appraisal. Nat Rev Drug Discov. 2005;4(2):165–171. doi:10.1038/nrd1634
15. Chang -C-C, Chen W-C, Ho T-F, Wu H-S, Wei Y-H. Development of natural anti-tumor drugs by microorganisms. J Biosci Bioeng. 2011;111(5):501–511. doi:10.1016/j.jbiosc.2010.12.026
16. Darshan N, Manonmani H. Prodigiosin and its potential applications. J Food Sci Technol. 2015;52(9):5393–5407. doi:10.1007/s13197-015-1740-4
17. Pérez-Tomás R, Vinas M. New insights on the antitumoral properties of prodiginines. Curr Med Chem. 2010;17(21):2222–2231. doi:10.2174/092986710791331103
18. Williamson NR, Fineran PC, Leeper FJ, Salmond GP. The biosynthesis and regulation of bacterial prodiginines. Nat Rev Microbiol. 2006;4(12):887–899. doi:10.1038/nrmicro1531
19. Han S-B, Lee CW, Yoon YD, et al. Effective prevention of lethal acute graft-versus-host disease by combined immunosuppressive therapy with prodigiosin and cyclosporine A. Biochem Pharmacol. 2005;70(10):1518–1526. doi:10.1016/j.bcp.2005.08.017
20. Huh J-E, Yim J-H, Lee H-K, Moon E-Y, Rhee D-K, Pyo S. Prodigiosin isolated from Hahella chejuensis suppresses lipopolysaccharide-induced NO production by inhibiting p38 MAPK, JNK and NF-κB activation in murine peritoneal macrophages. Int Immunopharmacol. 2007;7(13):1825–1833. doi:10.1016/j.intimp.2007.09.002
21. Yang L, Sun J, Xie W, Liu Y, Liu J. Dual-functional selenium nanoparticles bind to and inhibit amyloid β fiber formation in Alzheimer’s disease. J Mater Chem B. 2017;5(30):5954–5967. doi:10.1039/C6TB02952C
22. Cardoso BR, Roberts BR, Bush AI, Hare DJ. Selenium, selenoproteins and neurodegenerative diseases. Metallomics. 2015;7(8):1213–1228. doi:10.1039/C5MT00075K
23. Zhang J, Zhou X, Yu Q, et al. Epigallocatechin-3-gallate (EGCG)-stabilized selenium nanoparticles coated with Tet-1 peptide to reduce amyloid-β aggregation and cytotoxicity. ACS Appl Mater Interfaces. 2014;6(11):8475–8487. doi:10.1021/am501341u
24. Yin T, Yang L, Liu Y, Zhou X, Sun J, Liu J. Sialic acid (SA)-modified selenium nanoparticles coated with a high blood–brain barrier permeability peptide-B6 peptide for potential use in Alzheimer’s disease. Acta biomaterialia. 2015;25:172–183. doi:10.1016/j.actbio.2015.06.035
25. Yang L, Wang W, Chen J, Wang N, Zheng G. A comparative study of resveratrol and resveratrol‐functional selenium nanoparticles: inhibiting amyloid β aggregation and reactive oxygen species formation properties. J Biomed Mater Res A. 2018;106(12):3034–3041. doi:10.1002/jbm.a.36493
26. Wadhwani SA, Shedbalkar UU, Singh R, Chopade BA. Biogenic selenium nanoparticles: current status and future prospects. Appl Microbiol Biotechnol. 2016;100(6):2555–2566. doi:10.1007/s00253-016-7300-7
27. Mao QQ, Ip S-P, Ko K-M, Tsai S-H, Che C-T. Peony glycosides produce antidepressant-like action in mice exposed to chronic unpredictable mild stress: effects on hypothalamic-pituitary-adrenal function and brain-derived neurotrophic factor. Prog Neuropsychopharmacol Biol Psychiatry. 2009;33(7):1211–1216. doi:10.1016/j.pnpbp.2009.07.002
28. Kędzierska E, Dąbkowska L, Obierzyński P, et al. Synergistic Action of Sodium Selenite with some Antidepressants and Diazepam in Mice. Pharmaceutics. 2018;10(4):270. doi:10.3390/pharmaceutics10040270
29. Abdelfattah MS, Elmallah MI, Ebrahim HY, Almeer RS, Eltanany RM, Abdel Moneim AE. Prodigiosins from a marine sponge-associated actinomycete attenuate HCl/ethanol-induced gastric lesion via antioxidant and anti-inflammatory mechanisms. PLoS One. 2019;14(6):e0216737. doi:10.1371/journal.pone.0216737
30. Yuan X, Fu Z, Ji P, et al. Selenium nanoparticles pre-treatment reverse behavioral, oxidative damage, neuronal loss and neurochemical alterations in pentylenetetrazole-induced epileptic seizures in mice. Int J Nanomedicine. 2020;15:6339. doi:10.2147/IJN.S259134
31. Dong H, Cong W, Guo X, et al. β‑asarone relieves chronic unpredictable mild stress induced depression by regulating the extracellular signal‑regulated kinase signaling pathway. Exp Ther Med. 2019;18(5):3767–3774. doi:10.3892/etm.2019.8018
32. Sahin Ozkartal C, Aricioglu F, Tuzun E, Kucukali Cİ. Chronic mild stress-induced anhedonia in rats is coupled with the upregulation of inflammasome sensors: a possible involvement of NLRP1. Psychiatry Clin Psychopharmacol. 2018;28(3):236–244. doi:10.1080/24750573.2018.1426694
33. Ohkawa H, Ohishi N, Yagi K. Assay for lipid peroxides in animal tissues by thiobarbituric acid reaction. Anal Biochem. 1979;95(2):351–358. doi:10.1016/0003-2697(79)90738-3
34. Green LC, Wagner DA, Glogowski J, Skipper PL, Wishnok JS, Tannenbaum SR. Analysis of nitrate, nitrite, and [15N]nitrate in biological fluids. Research Support, U.S. Gov’t, P.H.S. Anal Biochem. 1982;126(1):131–138. doi:10.1016/0003-2697(82)90118-X
35. Aebi H. Catalase in vitro. Methods Enzymol. 1984;105:121–126.
36. Nishikimi M, Appaji N, Yagi K. The occurrence of superoxide anion in the reaction of reduced phenazine methosulfate and molecular oxygen. Biochem Biophys Res Commun. 1972;46(2):849–854. doi:10.1016/S0006-291X(72)80218-3
37. Ellman GL. Tissue sulfhydryl groups. Arch Biochem Biophys. 1959;82(1):70–77. doi:10.1016/0003-9861(59)90090-6
38. Paglia DE, Valentine WN. Studies on the quantitative and qualitative characterization of erythrocyte glutathione peroxidase. J Lab Clin Med. 1967;70(1):158–169.
39. Smith IK, Vierheller TL, Thorne CA. Assay of glutathione reductase in crude tissue homogenates using 5, 5′-dithiobis (2-nitrobenzoic acid). Anal Biochem. 1988;175(2):408–413. doi:10.1016/0003-2697(88)90564-7
40. Bradford MM. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem. 1976;72(1):248–254. doi:10.1016/0003-2697(76)90527-3
41. Pagel P, Blome J, Wolf HU. High-performance liquid chromatographic separation and measurement of various biogenic compounds possibly involved in the pathomechanism of Parkinson’s disease. J Chromatogr B Biomed Sci Appl. 2000;746(2):297–304. doi:10.1016/S0378-4347(00)00348-0
42. Ellman GL, Courtney KD, Andres JV, Featherstone RM. A new and rapid colorimetric determination of acetylcholinesterase activity. Biochem Pharmacol. 1961;7(2):88–95. doi:10.1016/0006-2952(61)90145-9
43. Dar A, Khan KM, Ateeq HS, et al. Inhibition of monoamine oxidase–A activity in rat brain by synthetic hydrazines: structure-activity relationship (SAR). J Enzyme Inhib Med Chem. 2005;20(3):269–274. doi:10.1080/14756360400026212
44. Arlt W, Stewart PM. Adrenal corticosteroid biosynthesis, metabolism, and action. Endocrinol Metab Clin. 2005;34(2):293–313. doi:10.1016/j.ecl.2005.01.002
45. Nicoloff JT, Spencer CA. The use and misuse of the sensitive thyrotropin assays. J Clin Endocrinol Metab. 1990;71(3):553–558. doi:10.1210/jcem-71-3-553
46. Livak KJ, Schmittgen TD. Analysis of relative gene expression data using real-time quantitative PCR and the 2− ΔΔCT method. methods. 2001;25(4):402–408. doi:10.1006/meth.2001.1262
47. Nakaji M, Hayashi Y, Ninomiya T, et al. Histological grading and staging in chronic hepatitis: its practical correlation. Pathol Int. 2002;52(11):683–690. doi:10.1046/j.1440-1827.2002.01410.x
48. Al-Brakati A, Alsharif KF, Alzahrani KJ, et al. Using Green Biosynthesized Lycopene-Coated Selenium Nanoparticles to Rescue Renal Damage in Glycerol-Induced Acute Kidney Injury in Rats. Int J Nanomedicine. 2021;16:4335–4349. doi:10.2147/IJN.S306186
49. Rajkumar K, Mvs S, Koganti S, Burgula S. Selenium Nanoparticles Synthesized Using Pseudomonas stutzeri (MH191156) Show Antiproliferative and Anti-angiogenic Activity Against Cervical Cancer Cells. Int J Nanomedicine. 2020;15:4523–4540. doi:10.2147/IJN.S247426
50. Fatima M, Srivastav S, Ahmad MH, Mondal AC. Effects of chronic unpredictable mild stress induced prenatal stress on neurodevelopment of neonates: role of GSK-3β. Sci Rep. 2019;9(1):1–13. doi:10.1038/s41598-018-38085-2
51. Foyet HS, Deffo ST, Yewo PK, et al. Ficus sycomorus extract reversed behavioral impairment and brain oxidative stress induced by unpredictable chronic mild stress in rats. BMC Complement Altern Med. 2017;17(1):1–15. doi:10.1186/s12906-016-1505-2
52. Hasan A, Morshed M, Memic A, Hassan S, Webster TJ, Marei HE. Nanoparticles in tissue engineering: applications, challenges and prospects. Int J Nanomedicine. 2018;13:5637. doi:10.2147/IJN.S153758
53. Peng D, Zhang J, Liu Q, Taylor EW. Size effect of elemental selenium nanoparticles (Nano-Se) at supranutritional levels on selenium accumulation and glutathione S-transferase activity. J Inorg Biochem. 2007;101(10):1457–1463. doi:10.1016/j.jinorgbio.2007.06.021
54. Zhang J-S, Gao X-Y, Zhang L-D, Bao Y-P. Biological effects of a nano red elemental selenium. Biofactors. 2001;15(1):27–38. doi:10.1002/biof.5520150103
55. Mishra B, Hassan P, Priyadarsini K, Mohan H. Reactions of biological oxidants with selenourea: formation of redox active nanoselenium. J Phys Chem B. 2005;109(26):12718–12723. doi:10.1021/jp051328n
56. Huang B, Zhang J, Hou J, Chen C. Free radical scavenging efficiency of Nano-Se in vitro. Free Radic Biol Med. 2003;35(7):805–813. doi:10.1016/S0891-5849(03)00428-3
57. Pelyhe C, Mézes M. Myths and facts about the effects of nano selenium in farm animals–mini-review. Eur Chem Bull. 2013;2(12):1049–1052.
58. Wang Y, Yan X, Fu L. Effect of selenium nanoparticles with different sizes in primary cultured intestinal epithelial cells of crucian carp, Carassius auratus gibelio. Int J Nanomedicine. 2013;8:4007. doi:10.2147/IJN.S43691
59. Yang X-H, Song S-Q, Xu Y. Resveratrol ameliorates chronic unpredictable mild stress-induced depression-like behavior: involvement of the HPA axis, inflammatory markers, BDNF, and Wnt/β-catenin pathway in rats. Neuropsychiatr Dis Treat. 2017;13:2727. doi:10.2147/NDT.S150028
60. Gáll Z, Farkas S, Albert A, et al. Effects of chronic cannabidiol treatment in the rat chronic unpredictable mild stress model of depression. Biomolecules. 2020;10(5):801. doi:10.3390/biom10050801
61. Wang L, Li -X-X, Chen X, Qin X-Y, Kardami E, Cheng Y. Antidepressant-like effects of low-and high-molecular weight FGF-2 on chronic unpredictable mild stress mice. Front Mol Neurosci. 2018;11:377. doi:10.3389/fnmol.2018.00377
62. Strekalova T, Spanagel R, Bartsch D, Henn FA, Gass P. Stress-induced anhedonia in mice is associated with deficits in forced swimming and exploration. Neuropsychopharmacology. 2004;29(11):2007–2017. doi:10.1038/sj.npp.1300532
63. He LW, Zeng L, Tian N, et al. Optimization of food deprivation and sucrose preference test in SD rat model undergoing chronic unpredictable mild stress. Animal Models Exp Med. 2020;3(1):69–78. doi:10.1002/ame2.12107
64. Zhang L, Luo J, Zhang M, Yao W, Ma X, Yu SY. Effects of curcumin on chronic, unpredictable, mild, stress-induced depressive-like behaviour and structural plasticity in the lateral amygdala of rats. Int J Neuropsychopharmacol. 2014;17(5):793–806. doi:10.1017/S1461145713001661
65. Ebokaiwe AP, Okori S, Nwankwo JO, Ejike CE, Osawe SO. Selenium nanoparticles and metformin ameliorate streptozotocin-instigated brain oxidative-inflammatory stress and neurobehavioral alterations in rats. Naunyn schmiedeberg’s Arch Pharmacol. 2020;1:1–12.
66. Ali HF, El-Sayed NM, Ahmed AA, Hanna PA, Moustafa YM. Nano selenium ameliorates oxidative stress and inflammatory response associated with cypermethrin-induced neurotoxicity in rats. Ecotoxicol Environ Saf. 2020;195:110479. doi:10.1016/j.ecoenv.2020.110479
67. Lucca G, Comim CM, Valvassori SS, et al. Increased oxidative stress in submitochondrial particles into the brain of rats submitted to the chronic mild stress paradigm. J Psychiatr Res. 2009;43(9):864–869. doi:10.1016/j.jpsychires.2008.11.002
68. Lindqvist D, Dhabhar FS, James SJ, et al. Oxidative stress, inflammation and treatment response in major depression. Psychoneuroendocrinology. 2017;76:197–205. doi:10.1016/j.psyneuen.2016.11.031
69. Hu M, Zou W, Wang C-Y, et al. Hydrogen sulfide protects against chronic unpredictable mild stress-induced oxidative stress in hippocampus by upregulation of BDNF-TrkB pathway. Oxid Med Cell Longev. 2016;2016:1–10. doi:10.1155/2016/2153745
70. Li Y, Wang L, Wang P, et al. Ginsenoside-Rg1 Rescues Stress-Induced Depression-Like Behaviors via Suppression of Oxidative Stress and Neural Inflammation in Rats. Oxid Med Cell Longev. 2020;2020:2325391. doi:10.1155/2020/2325391
71. Liao D, Lv C, Cao L, et al. Curcumin Attenuates Chronic Unpredictable Mild Stress-Induced Depressive-Like Behaviors via Restoring Changes in Oxidative Stress and the Activation of Nrf2 Signaling Pathway in Rats. Oxid Med Cell Longev. 2020;2020:9268083. doi:10.1155/2020/9268083
72. Kassab RB, Lokman MS, Daabo HM, et al. Ferulic acid influences Nrf2 activation to restore testicular tissue from cadmium‐induced oxidative challenge, inflammation, and apoptosis in rats. J Food Biochem. 2020;44(12):e13505. doi:10.1111/jfbc.13505
73. Ahmed HH, Abd El-Maksoud MD, Moneim AEA, Aglan HA. Pre-clinical study for the antidiabetic potential of selenium nanoparticles. Biol Trace Elem Res. 2017;177(2):267–280. doi:10.1007/s12011-016-0876-z
74. Shalby AB, Abd El-Maksoud MD, Moneim AEA, Ahmed HH. Antifibrotic candidates of Selenium nanoparticles and selenium in the experimental model. J Appl Pharm Sci. 2017;7(09):191–198.
75. Cao C, Li X, Qin L, et al. High Selenium Yeast mitigates aluminum-induced cerebral inflammation by increasing oxidative stress and blocking NO production. Biometals. 2018;31(5):835–843. doi:10.1007/s10534-018-0128-0
76. Nazıroğlu M, Muhamad S, Pecze L. Nanoparticles as potential clinical therapeutic agents in Alzheimer’s disease: focus on selenium nanoparticles. Expert Rev Clin Pharmacol. 2017;10(7):773–782. doi:10.1080/17512433.2017.1324781
77. Khiralla G, Elhariry H, Selim SM. Chitosan‐stabilized selenium nanoparticles attenuate acrylamide‐induced brain injury in rats. J Food Biochem. 2020;44(10):e13413. doi:10.1111/jfbc.13413
78. Chang -C-C, Wang Y-H, Chern C-M, et al. Prodigiosin inhibits gp91phox and iNOS expression to protect mice against the oxidative/nitrosative brain injury induced by hypoxia–ischemia. Toxicol Appl Pharmacol. 2011;257(1):137–147. doi:10.1016/j.taap.2011.08.027
79. Cigliano L, Spagnuolo MS, Boscaino F, et al. Dietary Supplementation with fish oil or conjugated linoleic acid relieves depression markers in mice by modulation of the Nrf2 pathway. Mol Nutr Food Res. 2019;63(21):1900243. doi:10.1002/mnfr.201900243
80. Zhou Y, Cong Y, Liu H. Folic acid ameliorates depression-like behaviour in a rat model of chronic unpredictable mild stress. BMC Neurosci. 2020;21(1):1–8. doi:10.1186/s12868-020-0551-3
81. Chen Q, Luo Y, Kuang S, et al. Cyclooxygenase-2 signalling pathway in the cortex is involved in the pathophysiological mechanisms in the rat model of depression. Sci Rep. 2017;7(1):1–12. doi:10.1038/s41598-016-0028-x
82. Song Q, Fan C, Wang P, Li Y, Yang M, Yu SY. Hippocampal CA1 βCaMKII mediates neuroinflammatory responses via COX-2/PGE2 signaling pathways in depression. J Neuroinflammation. 2018;15(1):1–17. doi:10.1186/s12974-018-1377-0
83. You -M-M, Chen Y-F, Pan Y-M, et al. Royal Jelly Attenuates LPS-Induced Inflammation in BV-2 Microglial Cells through Modulating NF-κB and p38/JNK Signaling Pathways. Mediators Inflamm. 2018;2018:11. doi:10.1155/2018/7834381
84. Wang Z, Liu S, Pan W, Guo Y, Shen Z. Bafilomycin A1 alleviates depression‑like symptoms in chronic unpredictable mild stress rats. Mol Med Rep. 2018;18(5):4587–4594. doi:10.3892/mmr.2018.9431
85. Yeo JE, Kang SK. Selenium effectively inhibits ROS-mediated apoptotic neural precursor cell death in vitro and in vivo in traumatic brain injury. Biochimica et Biophysica Acta. 2007;1772(11–12):1199–1210. doi:10.1016/j.bbadis.2007.09.004
86. Hassankhani R, Sam MR, Esmaeilou M, Ahangar P. Prodigiosin isolated from cell wall of Serratia marcescens alters expression of apoptosis-related genes and increases apoptosis in colorectal cancer cells. Med Oncol. 2015;32(1):366. doi:10.1007/s12032-014-0366-0
87. Wu G-F, Ren S, Tang R-Y, et al. Antidepressant effect of taurine in chronic unpredictable mild stress-induced depressive rats. Sci Rep. 2017;7(1):1–14.
88. Chanoine J-P, Wong AC, Lavoie J-C. Selenium deficiency impairs corticosterone and leptin responses to adrenocorticotropin in the rat. Biofactors. 2004;20(2):109–118. doi:10.1002/biof.5520200205
89. Peng Z, Zhang C, Yan L, et al. EPA is more effective than DHA to improve depression-like behavior, glia cell dysfunction and hippcampal apoptosis signaling in a chronic stress-induced rat model of depression. Int J Mol Sci. 2020;21(5):1769. doi:10.3390/ijms21051769
90. Perona MT, Waters S, Hall FS, et al. Animal models of depression in dopamine, serotonin and norepinephrine transporter knockout mice: prominent effects of dopamine transporter deletions. Behav Pharmacol. 2008;19(5–6):566. doi:10.1097/FBP.0b013e32830cd80f
91. Liu C, Wang J, Xu S, et al. Paecilomyces tenuipes extract prevents depression-like behaviors in chronic unpredictable mild stress-induced rat model via modulation of neurotransmitters. Mol Med Rep. 2017;16(2):2172–2178. doi:10.3892/mmr.2017.6807
92. Stockmeier CA. Involvement of serotonin in depression: evidence from postmortem and imaging studies of serotonin receptors and the serotonin transporter. J Psychiatr Res. 2003;37(5):357–373. doi:10.1016/S0022-3956(03)00050-5
93. Huang D, Zhang L, Yang JQ, et al. Evaluation on monoamine neurotransmitters changes in depression rats given with sertraline, meloxicam or/and caffeic acid. Genes Dis. 2019;6(2):167–175. doi:10.1016/j.gendis.2018.05.005
94. Fernandes J, Gupta GL. N-acetylcysteine attenuates neuroinflammation associated depressive behavior induced by chronic unpredictable mild stress in rat. Behav Brain Res. 2019;364:356–365. doi:10.1016/j.bbr.2019.02.025
95. Lim DW, Park J, Jung J, et al. Dicaffeoylquinic acids alleviate memory loss via reduction of oxidative stress in stress-hormone-induced depressive mice. Pharmacol Res. 2020;161:105252. doi:10.1016/j.phrs.2020.105252
96. Al Kahtani M. Effect of both selenium and biosynthesized nanoselenium particles on cadmium-induced neurotoxicity in albino rats. Hum Exp Toxicol. 2020;39(2):159–172. doi:10.1177/0960327119880589
97. Abou Zaid OA. Effect of Acrylamide on neurotransmitters and acetyl-cholinestrase activity in the brain of rats: therapeutic effect of ferulic acid and selenium nanoparticles. Ann Brit Med Sci. 2017;3(1):18–25.
98. Zafar KS, Siddiqui A, Sayeed I, Ahmad M, Salim S, Islam F. Dose‐dependent protective effect of selenium in rat model of Parkinson’s disease: neurobehavioral and neurochemical evidences. J Neurochem. 2003;84(3):438–446. doi:10.1046/j.1471-4159.2003.01531.x
99. Henderson CE. Role of neurotrophic factors in neuronal development. Curr Opin Neurobiol. 1996;6(1):64–70. doi:10.1016/S0959-4388(96)80010-9
100. Li K, Yan L, Zhang Y, et al. Seahorse treatment improves depression-like behavior in mice exposed to CUMS through reducing inflammation/oxidants and restoring neurotransmitter and neurotrophin function. J Ethnopharmacol. 2020;250:112487. doi:10.1016/j.jep.2019.112487
101. Zhang -S-S, Tian Y-H, Jin S-J, et al. Isoflurane produces antidepressant effects inducing BDNF-TrkB signaling in CUMS mice. Psychopharmacology. 2019;236(11):3301–3315. doi:10.1007/s00213-019-05287-z
102. Abou Zaid OAR, Sm E-S, Barakat W. Ameliorative effect of selenium nanoparticles and ferulic acid on acrylamide-induced neurotoxicity in rats. Ann Med Biomed Sci. 2017;3(2):35–45.
103. Abdelfattah MS, Badr SEA, Lotfy SA, et al. Rutin and Selenium Co-administration Reverse 3-Nitropropionic Acid-Induced Neurochemical and Molecular Impairments in a Mouse Model of Huntington’s Disease. Neurotox Res. 2020;37(1):77–92. doi:10.1007/s12640-019-00086-y
104. Anděrová M, Kubinová S, Mazel T, et al. Effect of elevated K+, hypotonic stress, and cortical spreading depression on astrocyte swelling in GFAP‐deficient mice. Glia. 2001;35(3):189–203. doi:10.1002/glia.1084
105. Ali S, Abd El Wahab M, Ayuob N, Suliaman M. The antidepressant-like effect of Ocimum basilicum in an animal model of depression. Biotechnic Histochem. 2017;92(6):390–401. doi:10.1080/10520295.2017.1323276
106. Li M, Fu Q, Li Y, Li S, Xue J, Ma S. Emodin opposes chronic unpredictable mild stress induced depressive-like behavior in mice by upregulating the levels of hippocampal glucocorticoid receptor and brain-derived neurotrophic factor. Fitoterapia. 2014;98:1–10. doi:10.1016/j.fitote.2014.06.007
107. Yao Z, Zhang Z, Zhang J, et al. Electroacupuncture alleviated the depression-like behavior by regulating FGF2 and astrocytes in the hippocampus of rats with chronic unpredictable mild stress. Brain Res Bull. 2021;169:43–50. doi:10.1016/j.brainresbull.2021.01.005
108. Ibrahim H, Zommara M, Elnaggar M. Ameliorating effect of selenium nanoparticles on cyclophosphamide induced hippocampal neurotoxicity in male rats: light, electron microscopic and immunohistochemical study. Folia Morphologica. 2020. doi:10.5603/FM.a2020.0117
 © 2021 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2021 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.