Back to Journals » Neuropsychiatric Disease and Treatment » Volume 21
Network Pharmacology and Molecular Docking Reveal Neuroprotective Potential of Ligusticum wallichii in Alzheimer’s Disease Therapy
Received 8 March 2025
Accepted for publication 20 October 2025
Published 25 November 2025 Volume 2025:21 Pages 2603—2622
DOI https://doi.org/10.2147/NDT.S523834
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Yu-Ping Ning
Chanjuan Zhou,1 Yutao Peng2
1Department of Clinical Psychology, The First Affiliated Hospital, Hengyang Medical School, University of South China, Hengyang, Hunan, 421001, People’s Republic of China; 2Department of Function, The First Affiliated Hospital, Hengyang Medical School, University of South China, Hengyang, Hunan, 421001, People’s Republic of China
Correspondence: Yutao Peng, Department of Function, The First Affiliated Hospital, Hengyang Medical School, University of South China, 69 Chuan Shan Road, Hengyang, Hunan, 421001, People’s Republic of China, Tel +86 15115465507, Email [email protected]
Purpose: In traditional Chinese medicine, Ligusticum wallichii is a prominent herb, acclaimed for its therapeutic roles, including anti-tumor, antioxidant, and anti-inflammatory benefits. Studies conducted recently suggest it may help reduce cognitive deficits linked to Alzheimer’s disease. However, the precise neuroprotective pathways through which Ligusticum wallichii exerts its effects on Alzheimer’s disease are not yet fully understood. Network pharmacology is utilized in this research to understand the mechanisms through which Ligusticum wallichii’s active ingredient might protect against Alzheimer’s disease.
Methods: The TCMSP database was utilized to extract the bioactive compounds of Ligusticum wallichii, and their related molecular targets were identified. By querying the GeneCards and OMIM databases, targets associated with Alzheimer’s disease were identified. Using Cytoscape 3.8.2, a regulatory network mapping the interactions between active compounds and their respective targets was constructed. A protein-protein interaction network was generated by analyzing the target genes influenced by Ligusticum wallichii in Alzheimer’s disease using the String database. The DAVID database was utilized to perform functional enrichment analysis, encompassing Gene Ontology (GO) and KEGG pathway analyses, to identify possible biological pathways related to these targets. Following this, molecular docking studies were carried out to confirm the interaction strength of the active compounds to the pivotal targets. Finally, in vitro experimental validation was performed to corroborate the findings.
Results: Seven bioactive compounds were identified from Ligusticum wallichii, interacting with 269 potential targets. Molecular docking revealed that Myricanone, Mandenol, and Sitosterol exhibited stable binding affinities with STAT3, HSP90AA1, and EGFR, with binding energies ranging from − 4.04 to − 5.87 kcal/mol. In vitro studies demonstrated that these compounds significantly downregulated the expression of STAT3, EGFR, and HSP90AA1 in Neuro 2A cells.
Conclusion: In conclusion, the results indicate that Ligusticum wallichii significantly downregulated STAT3, EGFR, and HSP90AA1 expression in Neuro 2A cells, providing mechanistic evidence that targeting these proteins may ameliorate neurodegenerative processes in Alzheimer’s disease and highlighting Ligusticum wallichii’s promising therapeutic potential.
Keywords: Ligusticum wallichii, molecular docking, network pharmacology, Alzheimer’s disease
Introduction
Alzheimer’s disease, which makes up 50% to 75% of dementia cases, is a widespread neurodegenerative illness with no current cure.1 Individuals affected by this disorder often experience symptoms including memory loss, cognitive deterioration, behavioral abnormalities, and reduced ability to perform daily activities.2–5 The pressing need for effective therapeutic interventions is evident. Current pharmacological strategies primarily focus on modulating cholinergic and glutamatergic systems, providing symptomatic alleviation but with debated efficacy in neuroprotection.6–8 This underscores the necessity for innovative treatment avenues. In recent years, increasing attention has also been paid to the potential of traditional Chinese medicine as a complementary strategy to conventional pharmacotherapy for Alzheimer’s disease.9–12
Ligusticum wallichii, a well-known traditional Chinese herb, has long been applied in the treatment of cerebrovascular and neurological disorders, including stroke and migraine.13,14 Pharmacological studies have revealed that its active constituent, tetramethylpyrazine, exerts neuroprotective effects by attenuating oxidative stress, suppressing inflammatory responses, and reducing neuronal apoptosis in rodent models of cerebral ischemia and oxygen-glucose deprivation experiments.15,16 Importantly, Ligusticum wallichii and its derivatives have also shown beneficial effects in Alzheimer’s disease models. For example, tetramethylpyrazine nitrone, a derivative of Ligusticum wallichii, significantly improved cognitive performance and reduced amyloid-β and hyperphosphorylated tau levels in Alzheimer’s disease mice.17 These findings suggest that Ligusticum wallichii may modulate multiple pathological processes of Alzheimer’s disease, including amyloid aggregation, tau pathology, oxidative damage, and synaptic dysfunction.
Despite accumulating evidence of its neuroprotective benefits, the precise molecular mechanisms through which Ligusticum wallichii exerts therapeutic effects on Alzheimer’s disease remain largely unexplored. Current studies have not systematically analyzed Ligusticum wallichii’s active compounds, their potential targets, and signaling pathways in Alzheimer’s disease. This knowledge gap limits the translation of Ligusticum wallichii into targeted Alzheimer’s disease therapy. Therefore, it is critical to identify the molecular targets of Ligusticum wallichii, as this would provide a mechanistic basis for its therapeutic potential and inform the development of novel multi-targeted interventions for Alzheimer’s disease.
To overcome limitations of traditional pharmacological approaches, network pharmacology offers a systems-level strategy to investigate the multi-faceted mechanisms of drug actions, integrating large-scale biological data with network visualization.18–20 This holistic approach is consistent with the principles of traditional Chinese medicine, which emphasizes systemic regulation of diseases. Moreover, molecular docking provides a powerful computational method to study how Ligusticum wallichii compounds spatially interact with their protein targets, thereby validating predicted drug–target associations.21,22 By combining these methods, we can comprehensively elucidate the active components, target proteins, and signaling pathways through which Ligusticum wallichii intervenes in Alzheimer’s disease pathology. Importantly, such computational pharmacology approaches have recently been recognized as essential tools for advancing herbal medicine research by bridging traditional pharmacology with systems biology.23–25
In this study, we integrated network pharmacology and molecular docking to systematically identify the active compounds of Ligusticum wallichii, predict their Alzheimer’s disease-related targets, and explore the pathways and mechanisms through which Ligusticum wallichii may exert neuroprotective effects. This approach provides a more rigorous and transparent foundation for understanding Ligusticum wallichii’s role in Alzheimer’s disease therapy and contributes to the development of novel multi-target therapeutic strategies.
Material and Methods
Determining the Bioactive Compounds and Targets Associated with Ligusticum Wallichii
Ligusticum wallichii’s primary components were identified via the Traditional Chinese Medicine Systems Pharmacology Database and Analysis Platform (TCMSP) (https://tcmsp-e.com/tcmsp.php), applying the standards of oral bioavailability (OB) greater than 30% and drug-likeness (DL) equal to or more than 0.18. The thresholds of OB ≥ 30% and DL ≥ 0.18 were selected because compounds with OB ≥ 30% generally ensure sufficient biological efficacy through oral administration, while compounds with DL ≥ 0.18 are structurally and chemically similar to known drugs, improving the likelihood of favorable absorption, distribution, metabolism, and excretion (ADME) properties and clinical applicability, consistent with previous studies.26,27 Using the data extracted from the TCMSP database and supplemented by a review of relevant literature, the bioactive constituents of Ligusticum wallichii were finalized. 3D Viewer was utilized to construct the 3D structures of these compounds, which were then saved in mol2 format for target prediction. Based on physicochemical properties and feature data, target information was extracted from the TCMSP database, and these bioactive constituents’ SMILES files were sourced from the PubChem database. Target prediction was conducted using these SMILES files via the SwissTargetPrediction (STP) platform (http://old.swisstargetprediction.ch/). The leading 15% of predictions in STP were true compound targets with a probability of 75%, leading to the selection of the initial set of 15 predictions. The z’-score for network proximity was determined utilizing the standard deviation and average of random distances. Compared to the “Fit score”, the Z-score utilized in analyzing PharmMapper database results provides a more robust confidence interval and statistical significance. Significance was attributed to Z-scores above 0, while negative scores were seen as non-significant. Stronger alignment between protein targets and compound molecules is reflected by higher z’-scores. The STP and TCMSP databases together highlight the primary targets of Ligusticum wallichii’s bioactive compounds.
Recognizing Targets Connected to Alzheimer’s Disease
By searching the OMIM and GeneCards databases (https://omim.org/), targets connected to Alzheimer’s disease were identified. The initial step involved querying the OMIM database’s “Gene Map” section with “Alzheimer’s disease”, and exporting the results in Excel format. Similarly, searches for “Alzheimer’s disease” are conducted in the GeneCards database, with the findings stored in Excel format. As a crucial metric, the relevance score measured the strength of a gene’s connection to a biological process or specific disease. In this study, a relevance score threshold of ≥15 in GeneCards was applied, as genes with such scores indicate a strong association with Alzheimer’s disease and reduce the risk of including weakly associated targets. The datasets of Ligusticum wallichii and Alzheimer’s disease-related targets are analyzed and compared using the online tool Venny 2.1.0 (https://bioinfogp.cnb.csic.es/tools/venny/index.html) to identify overlapping targets.
Building a Network of Active Ingredient-Target Interactions
To find overlapping targets between Ligusticum wallichii and disease pathways, the online Venn diagram tool at https://bioinfogp.cnb.csic.es/tools/venny/ was utilized. This is achieved by cross-referencing Alzheimer’s disease-associated targets with those of active compounds, subsequently visualized through Venn diagram construction. The overlapping genes suggested potential therapeutic targets for Ligusticum wallichii in Alzheimer’s disease treatment. Gene nomenclature was used to computationally predict and standardize both the active compound and Alzheimer’s disease-associated targets. The curated Alzheimer’s disease targets were incorporated into Cytoscape 3.8.2 to build a detailed compound-target interaction network.
Constructing a Network for Protein-Protein Interactions and Identifying Main Targets
The STRING database was employed to assess the identified overlapping target proteins, with “Homo sapiens” designated as the target species for extracting protein-protein interaction (PPI) data. The data, organized as a Tab Separated Values (TSV) file, was then brought into the Cytoscape 3.8.2 for visualizing the network. Network topology analysis was performed utilizing the CytoHubba plug-in, leveraging eleven distinct algorithms based on shortest path principles, such as maximum clique centrality (MCC), maximum neighborhood component density, edge percolation component, and degree, as well as six centrality measures (stress, closeness, radiality, betweenness, degree, and bottleneck). Notably, in terms of accuracy, the MCC algorithm has been established as superior in identifying critical targets. Applying the MCC method in this study enabled the identification of core therapeutic targets, representing key nodes in Ligusticum wallichii’s mechanism of action combating Alzheimer’s disease. Additionally, critical sub-networks of Ligusticum wallichii’s active compounds were mapped within the PPI network, highlighting their functional significance.
Functional Enrichment Analysis
The clusterProfilerGO.R package in R software, available at https://www.r-project.org/, along with Perl scripting, was utilized to conduct functional annotation and pathway enrichment analyses. Enrichment scores were calculated based on standard hypergeometric testing, where P-values represent the likelihood of observing the number of genes associated with a GO term in our target set compared to the background genome. These P-values were then transformed to –log10(P-value) for visualization, with higher scores indicating stronger enrichment. The analysis applied a significance threshold of a modified p-value under 0.05, with the statistically enriched outcomes visualized through the ggplot2 package. The target genes of interest were initially analyzed through the DAVID online platform and subsequently processed in Omicsbean software, with Gene Symbol designated as the identifier type and “Homo sapiens” selected as the target species. Comprehensive functional characterization was achieved through Gene Ontology (GO) annotation and Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway mapping of the target proteins. The analytical outputs are systematically organized in Excel for functional and pathway filtering, maintaining a statistically meaningful level of P-value under 0.05. For clarity and focus on the most relevant functions, we selected the top 10 enriched terms in each GO category (BP, CC, MF) based on enrichment scores. For KEGG analysis, we focused on the top 20 pathways with nominal p-values well below 0.05. Although no formal multiple testing correction was applied, the consistently low values support the biological relevance of these pathways. This enrichment analysis was intended to be exploratory, highlighting meaningful pathways for further discussion and experimental validation rather than providing strict statistical inference. The results were presented in a bar graph format for enhanced interpretability.
Confirmation of the Drug-Component-Target-Disease Network
The creation of an integrated drug-component-target-disease network involved merging overlapping genes from both drug datasets and disease with the 20 most significant pathways derived from KEGG enrichment analysis.
Molecular Docking
This computational approach, known as molecular docking, predicts the binding orientation of small molecules at the active sites of macromolecules, enabling the prediction of binding affinity. This research employed AutoDock4 to examine the interactions between potential bioactive compounds from Ligusticum wallichii and their target proteins. The PubChem database was utilized to obtain the chemical structures of these active compounds in SDF format, then processed using ChemOffice software for building 3D structures, minimizing energy, and converting to mol2 format. Structures of target proteins are acquired from the PDB database (http://www.rcsb.org/) and prepared by removing water molecules with PyMOL 2.3, while hydrogen atoms were added with AutoDock4. The pdbqt format was used to save both target and ligand files for analysis with the Vina docking program. Finally, LigPlot 2.23 was employed to generate 2D interaction diagrams, while PyMOL 2.3 was used to visualize the 3D docking complexes. Docking was performed using a grid box centered on the binding pocket of each target protein. The grid box was set to cover the protein’s active sites with the following search space in Å: EGFR-Mandenol (Center XYZ = 6.082, 9.592, and 58.764), EGFR-Myricanone (Center XYZ = 10.69, 9.408, and 58.597), EGFR-Sitosterol (Center XYZ = 8.555, 10.187, and 59.186), HSP90AA1-Mandenol (Center XYZ = 0.433, 14.681, and 20.381), HSP90AA1-Myricanone (Center XYZ = 2.656, 14.681, and 19.707), HSP90AA1-Sitosterol (Center XYZ = 0.181, 14.681, and 19.707), STAT3-Mandenol (Center XYZ = −2.599, 19.619, and 27.129), STAT3-Myricanone (Center XYZ = −2.264, 19.582, and 27.129), and STAT3-Sitosterol (Center XYZ = 8.555, 16.255, and 26.782). Validation was performed through re-docking of co-crystallized ligands, yielding RMSD values below 2.0 Å, confirming docking reliability. AutoDock4’s Lamarckian Genetic Algorithm (GA) scoring function was used, with parameters including 10 GA runs and a population size of 150.
Cell Culture
Neuro 2A cells, provided by Procell Life Science & Technology Co., Ltd. in Wuhan, China, were kept in Dulbecco’s Modified Eagle’s Medium (DMEM) with 10% fetal bovine serum, plus 100 U/mL streptomycin and 100 U/mL penicillin. In a humidified atmosphere with 5% CO2, the cells were incubated at 37°C. Neuro 2A cells were selected because they are widely used in Alzheimer’s disease research for studying neuronal differentiation, signaling pathways, and Aβ metabolism, providing a relevant and established in vitro model for evaluating neuroprotective mechanisms. Subsequently, they were exposed to 80 μM concentrations of Myricanone, Mandenol, and Sitosterol over the course of 12 hours.
Cell Activity Detection
Myricanone, Mandenol, and Sitosterol, each with 98% purity and respective CAS numbers 32492–74-3, 544–35-4, and 83–46-5, were sourced from Baoji Chenguang Biotechnology Co., Ltd. in China. The CCK-8 assay was employed to evaluate cell viability. Briefly, in 96-well culture plates, Neuro 2A cells were placed and incubated at 37°C for a duration of 12 hours. They were then treated with varying concentrations (0–160 μM) of Myricanone, Mandenol, and Sitosterol for 24 hours. After adding 10 µL of CCK-8 solution to each well, the plates were incubated at 37°C for 1 hour. A 96-well microplate reader from Bio-Tek Instruments Inc. in Winooski, VT, USA, was utilized to assess absorbance at 450 nm. To ensure reliability, all experiments were executed a minimum of three times.
Real-Time Quantitative PCR
Cells were used to extract total RNA with TRIzol reagent from Invitrogen, USA. The process of synthesizing complementary DNA (cDNA) involved a high-capacity reverse transcription kit (Takara, Kyoto, Japan). The ABI7900HT Fast Real-Time PCR System from Applied Biosystems in Foster City, CA, USA, was used to perform quantitative real-time PCR (qRT-PCR) for 40 cycles with the following settings: 95°C for 3 minutes, then 90°C for 15 seconds, and 60°C for 1 minute. The sequences of primers utilized for qRT-PCR were: β-actin-forward primer, 5′-TGAGGCCCAGAGCAAGAGA-3′; reverse primer, 5′-TCGTCCCAGTTGGTGACGAT-3′. HSP90AA1-forward primer, 5′-GACGCTCTGGATAAAATCCGTT-3′; reverse primer, 5′-TGGGAATGAGATTGATGTGCAG-3′; EGFR-forward primer, 5′-TCCCTCAGCCACCCATATGTAC-3′; reverse primer, 5′-GTCTCGGGCCATTTTGGAGAATCC-3′; STAT3-forward primer, 5′-TGCAGTTTGGAAATAATGGTGA-3′; reverse primer, 5′-CATGTCAAAGGTGAGGGACTC-3′. For mRNA normalization, β-actin acted as the internal reference. To ensure the specificity of PCR products, melting curve analysis was employed. To determine relative gene expression levels, the 2−ΔΔCt method was applied.
Data Analysis
For network pharmacology analysis, data processing was conducted extensively using R software. GraphPad Prism 8.0.1, provided by GraphPad Software Inc. in San Diego, CA, USA, was adopted for the purpose of statistical analysis of the in vitro experimental data. Pairwise comparisons were conducted utilizing a Student’s t-test, and multiple group analyses were performed with one-way ANOVA followed through Tukey’s post hoc test. Sample sizes (n = 5 independent biological replicates for each assay, with each condition measured in five technical replicates) were explicitly reported. Prior to parametric analysis, normality was assessed using the Shapiro–Wilk test. P-values were adjusted using Tukey’s post hoc correction for multiple comparisons, and exact p-values are provided where appropriate. A p-value smaller than 0.05 was the threshold for statistical significance.
Results
Computational Prediction of Ligusticum wallichii’s Active Ingredients and Their Potential Connection to Alzheimer’s Disease Targets
The TCMSP database was utilized to identify the bioactive components of Ligusticum wallichii, yielding seven compounds in accordance with the criteria of OB being no less than 30% and DL being no less than 0.18 (Table 1). Corresponding target proteins for these bioactive compounds were also retrieved from the TCMSP database, resulting in 269 unique targets after compilation and removal of duplicates. Additionally, the GeneCards and OMIM databases were utilized to extract targets linked to Alzheimer’s disease with the indexed term “Alzheimer’s disease.” The datasets from both sources were merged, and redundant entries were eliminated, ultimately identifying a total of 2,345 unique targets (Figure 1). Among these, 124 targets overlapped with Ligusticum wallichii–related targets, and the remaining 2,221 are non-overlapping Alzheimer’s disease targets.
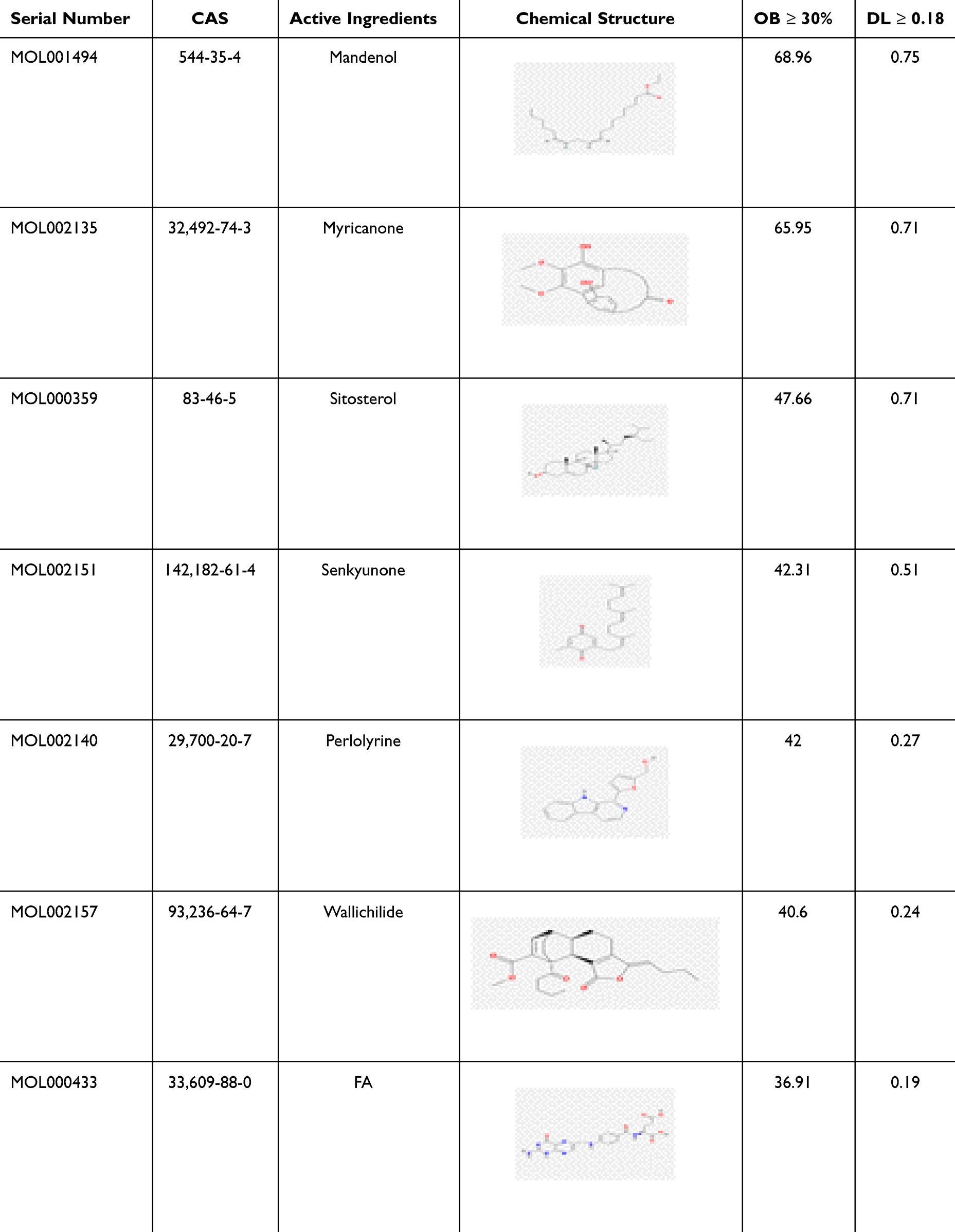 |
Table 1 Active Ingredients in Ligusticum wallichii |
 |
Figure 1 Overlap of target genes between Ligusticum wallichii’s active compounds and those associated with Alzheimer’s disease. Bioactive compounds of Ligusticum wallichii were identified from the TCMSP database, yielding seven compounds and 269 unique targets. Alzheimer’s disease-related targets were obtained from GeneCards and OMIM, resulting in 2,345 unique genes. After integration, 124 overlapping targets were identified, leaving 2,221 Alzheimer’s disease-specific and 145 Ligusticum wallichii-specific targets. |
Ligusticum wallichii’s potential targets were matched with targets linked to Alzheimer’s disease, identifying 124 overlapping targets through the Venn platform (Figure 1). These shared targets, along with the active compounds, were then entered into Cytoscape 3.8.2 for visualization, generating a “compound-target network” (Figure 2). The network analysis revealed that while some targets are influenced by multiple active compounds, others, such as NLRP3, PRKCA, HTR2A, and ESR1, are modulated by only one. Notably, targets like HSP90AA1, EGFR, STAT3, CYP27A1, and PRKCG were found to be regulated by all identified compounds.
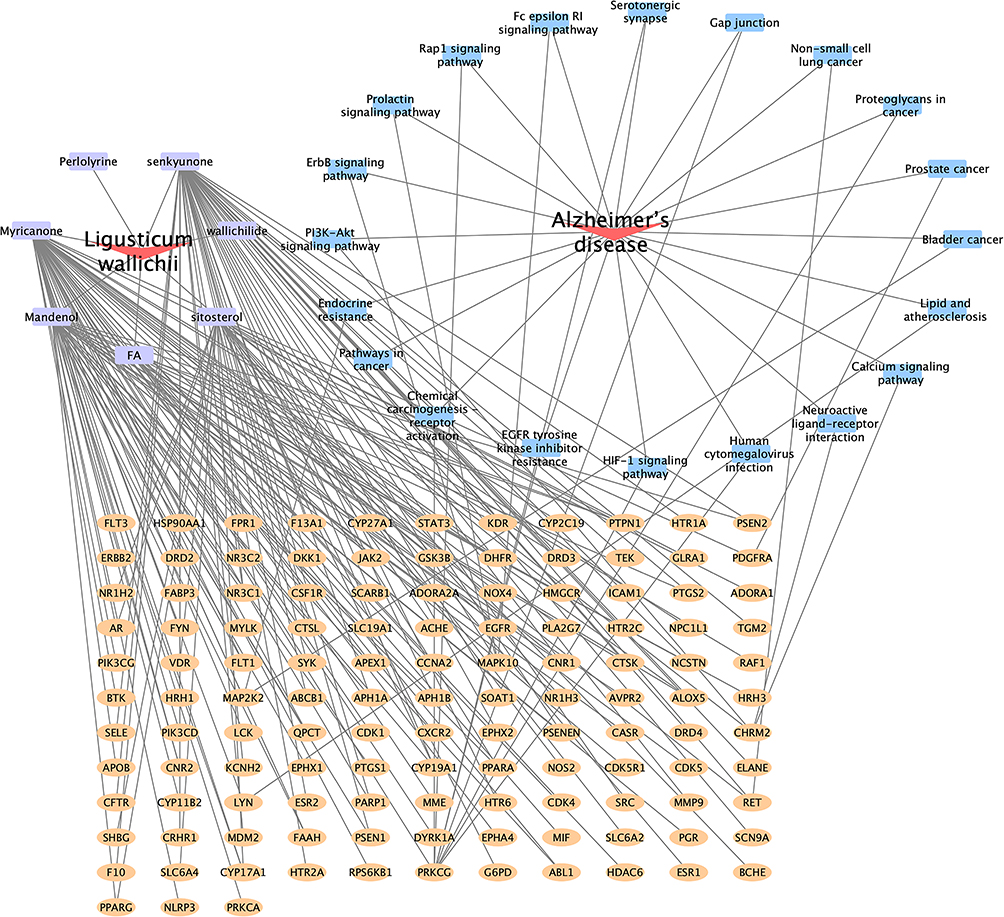 |
Figure 2 Network construction for active ingredients and targets. The purple square denotes the bioactive compound of Ligusticum wallichii, while the yellow square signifies the shared targets between this compound and Alzheimer’s disease. The blue square indicates the signaling pathways associated with Alzheimer’s disease-related targets. |
Computational Prediction of the Protein–Protein Interaction (PPI) Network
The initial step in investigating Ligusticum wallichii’s protective mechanism against Alzheimer’s disease involved analyzing intersecting targets with the STRING database, with “Homo sapiens” selected as the species while keeping default settings. The network of drug-disease interactions, featuring 124 nodes and 343 edges (Figure 3). Enrichment analysis revealed that the PPI sub-network contained far more interactions than a random protein set of similar size from the genome (P < 1.0e-16). The network’s characteristics included a local clustering coefficient of 0.516 and an average node degree of 5.53. Specifically, the average node degree of 5.53 reflects that each target is connected to more than five other proteins on average, while the local clustering coefficient of 0.516 indicates a high likelihood of neighboring proteins also being interconnected, consistent with functional convergence of Ligusticum wallichii-related targets in Alzheimer’s disease. This means that these proteins form a biologically interconnected cluster. Following this, within Cytoscape 3.8.2, the network data were imported, and the NetworkAnalyzer plug-in was used to establish a PPI network (Figure 4A). Nodes in the diagram varied in size based on degree values, with larger ones representing more connected genes. The ten highest-ranked proteins by degree, betweenness, and closeness centrality were identified in Figure 4B. Among these, we selected HSP90AA1, EGFR, and STAT3 as key targets not only because they consistently ranked at the top across all three centrality measures (Figure 5A and Table 2), but also due to their well-documented roles in Alzheimer’s disease pathology, including regulation of neuroinflammation, amyloid-β aggregation, and neuronal survival.28–30 Additionally, the active compounds Myricanone, Mandenol, and Sitosterol exhibited strong links to these targets (Figure 5B). This dual consideration of network centrality and disease relevance provides a more rigorous justification for focusing on these three proteins. In the PPI network, these fundamental proteins are crucial, highlighting their significance in the therapeutic benefits of Ligusticum wallichii against Alzheimer’s disease.
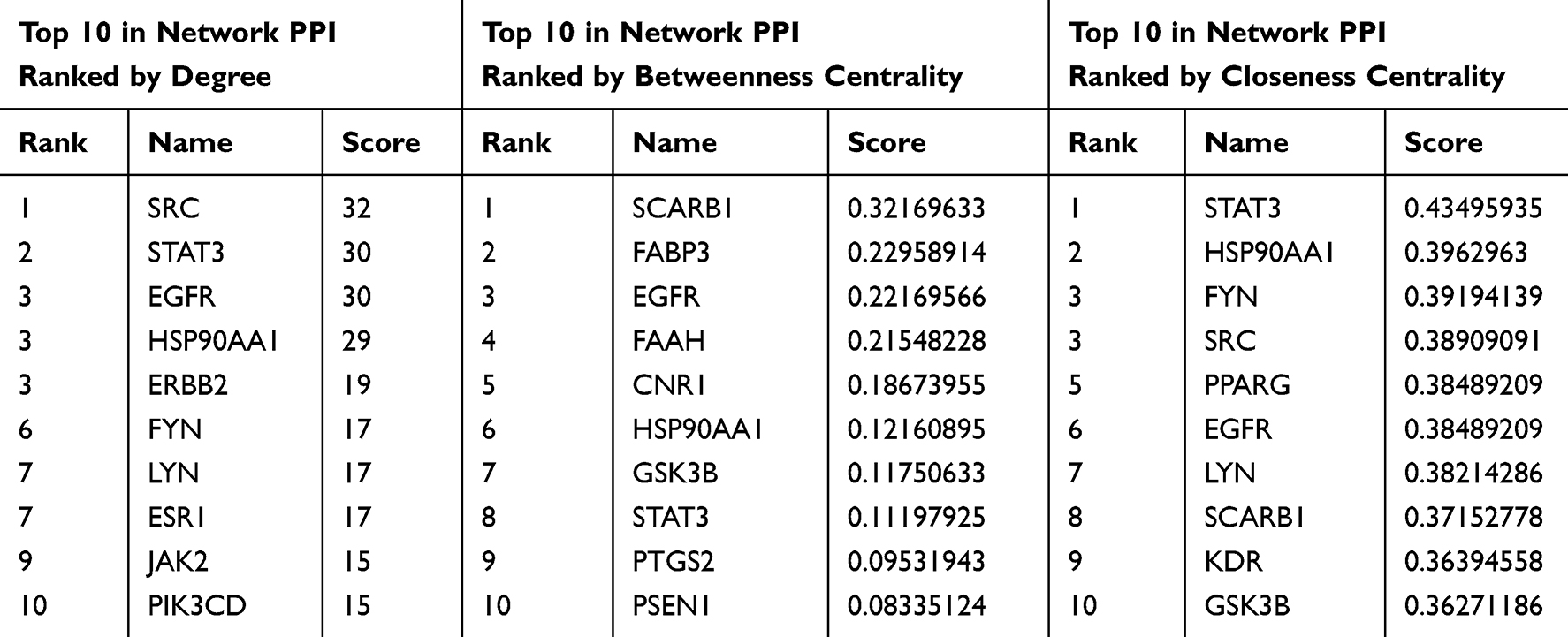 |
Table 2 Details of the 10 Most Critical Targets of Ligusticum wallichii in Alzheimer’s Disease Therapy |
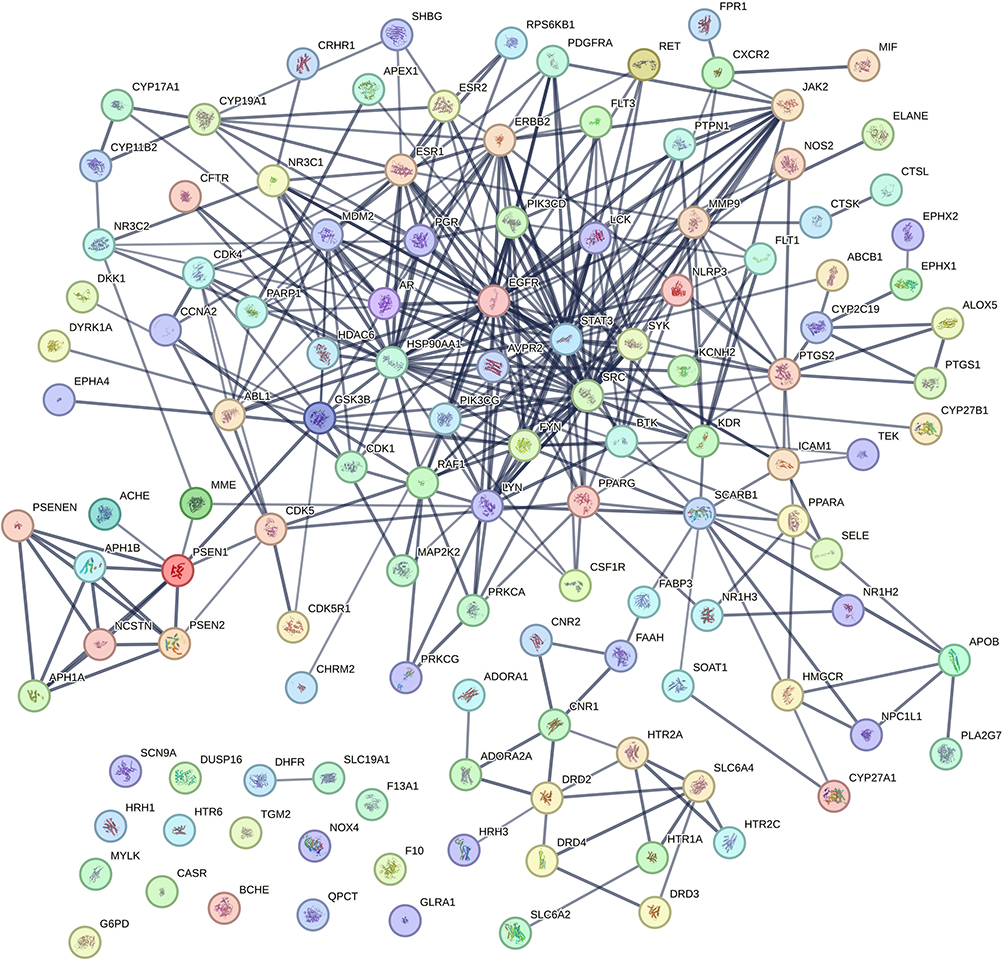 |
Figure 3 The network of protein-protein interactions for Ligusticum wallichii was created through STRING. |
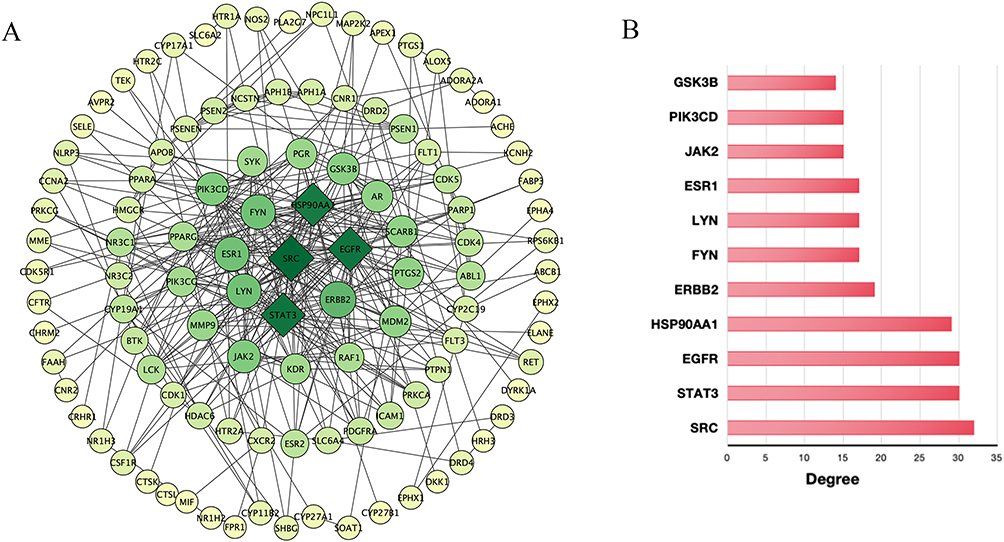 |
Figure 4 The PPI network of Ligusticum wallichii. (A) Visualization of the PPI network constructed using Cytoscape 3.8.2. Nodes with larger size and darker color denote proteins exhibiting higher connectivity. (B) A histogram illustrating the top 10 genes ranked by degree values. |
 |
Figure 5 Identification of pivotal targets. (A) The shared targets identified by degree centrality parameters (Degree, Betweenness, and Closeness). (B) The heat map represents the binding affinities of active components to crucial targets as determined by molecular docking. Myricanone-HSP90AA1 (−5.12 kcal/mol), Mandenol-HSP90AA1 (−4.49 kcal/mol), Sitosterol-HSP90AA1 (−5.27 kcal/mol), Myricanone-EGFR (−4.34 kcal/mol), Mandenol-EGFR (−5.46 kcal/mol), Sitosterol-EGFR (−4.93 kcal/mol), Myricanone-STAT3 (−4.63 kcal/mol), Mandenol-STAT3 (−5.87 kcal/mol), and Sitosterol-STAT3 (−4.04 kcal/mol). |
Computational Prediction of Pathway Enrichment Results
Subsequently, biological functions and metabolic pathways related to the intersecting targets were explored using GO and KEGG enrichment analyses. The possible impact of Ligusticum wallichii on Alzheimer’s disease is highlighted in the KEGG analysis bubble plot (Figure 6). Within the 130 pathways identified, the top 20 were selected, encompassing key pathophysiological processes such as PI3K-Akt, insulin resistance, EGFR tyrosine kinase inhibitor resistance, and Rap1 signaling pathway. Although several of these pathways are annotated as “cancer-related” in KEGG, it is increasingly recognized that these signaling pathways also play crucial roles in neuronal survival, apoptosis, inflammation, and synaptic plasticity, which are highly relevant to AD pathophysiology.28,31,32 Therefore, their enrichment in our study likely reflects shared molecular mechanisms rather than direct oncogenic processes. Cytoscape software was utilized to produce a “target-pathway” network diagram, which further illustrates the connections between important pathways and their targets (Figure 7).
 |
Figure 6 KEGG-based analysis of biological pathway enrichment. The x-axis (gene ratio) represents the proportion of Ligusticum wallichii target genes relative to the total number of genes in each KEGG pathway. The observed range (0.05–0.2) reflects the typical overlap pattern in network pharmacology studies, where pathways contain many genes but only a subset is mapped to the identified targets. The count legend represents the number of overlapping genes between Ligusticum wallichii targets and each KEGG pathway. |
 |
Figure 7 Visual representation of the target-pathway network. Nodes represent target genes and KEGG pathways, with edges indicating gene–pathway associations. Elliptical nodes represent pathways, while rectangular nodes represent target genes. A color scale from light yellow to orange reflects increasing pathway relevance. |
The 124 identified targets were analyzed for GO functional enrichment utilizing the Metascape platform, revealing the leading 10 most significant terms. The analysis identified 122 molecular function pathways, such as protein tyrosine kinase activity, transmembrane receptor protein tyrosine kinase activity, protein kinase activity, enzyme binding, ATP binding, identical protein binding, and nuclear receptor activity. For biological processes (BP), enrichment analysis identified 478 pathways, suggesting that Ligusticum wallichii may exert therapeutic effects on Alzheimer’s disease through mechanisms like positive regulation of the ERK1/2 cascade, kinase activity modulation, peptidyl-tyrosine phosphorylation, protein autophosphorylation, response to xenobiotic stimuli, and protein phosphorylation. Additionally, 81 cellular component (CC) terms were identified, encompassing structures such as dendrites, axons, neuronal cell bodies, cytoplasm, membrane rafts, presynaptic membranes, and the plasma membrane. A visual representation of these components is provided in Figure 8.
 |
Figure 8 Analysis of GO enrichment. The enrichment score is defined as the proportion of input genes annotated to a given GO term (Gene Ratio = Count / total number of input genes × 100%). Higher enrichment scores indicate stronger association of target genes with the respective GO terms. |
Computational Prediction of Molecular Docking Interactions
Molecular docking was conducted with AutoDock and PyMOL to investigate how the active compounds of Ligusticum wallichii interact with their target genes. Docking simulations were carried out for three key targets (HSP90AA1, EGFR, and STAT3) and three major bioactive components selected based on highest OB and DL values (Myricanone, Mandenol, and Sitosterol) (Figure 5B). The main bioactive components showed high binding energies with these targets: HSP90AA1 (−5.12 kcal/mol, −4.49 kcal/mol, −5.27 kcal/mol), EGFR (−4.34 kcal/mol, −5.46 kcal/mol, −4.93 kcal/mol), and STAT3 (−4.63 kcal/mol, −5.87 kcal/mol, −4.04 kcal/mol), indicating robust binding interactions between these components and their targets. The 2D structures of these computationally analyzed compounds are illustrated in Figure 9. Molecular docking analysis (Figures 10–12) revealed that EGFR formed hydrogen bonds with Mandenol via residues ARG-807, TYR-803, VAL-745, and SER-744, while HSP90AA1 interacted through residue LEU-107, ILE-96, and LYS-58. STAT3 formed hydrogen bonds with Mandenol through LEU-260, GLU-324, CYS-328, and PRO-330. Similarly, Myricanone engaged EGFR via VAL-745, LEU-754, and ASP-746, HSP90AA1 through THR-184, ASP-54, and GLY-135, and STAT3 through LYS-658, ILE-659, TYR-657, PRO-639, and GLU-638. Sitosterol formed interactions with EGFR at LEU-680, ARG-681, LEU-754, and GLY-755, HSP90AA1 via LEU-220, LYS-204, ILE-218, and PRO-217, and STAT3 through PHE-172, GLN-202, LEU-207, and LYS-294. These findings further substantiate that the bioactive components of Ligusticum wallichii have a robust binding interaction with their specific targets.
 |
Figure 9 The two-dimensional arrangements of the in silico active components. (A) Mandenol (molecular weight: 308.50 g/mol, logP: 8.17); (B) Myricanone (molecular weight: 356.41 g/mol, logP: 3.79); (C) Sitosterol (molecular weight: 414.71 g/mol, logP: 10.73). |
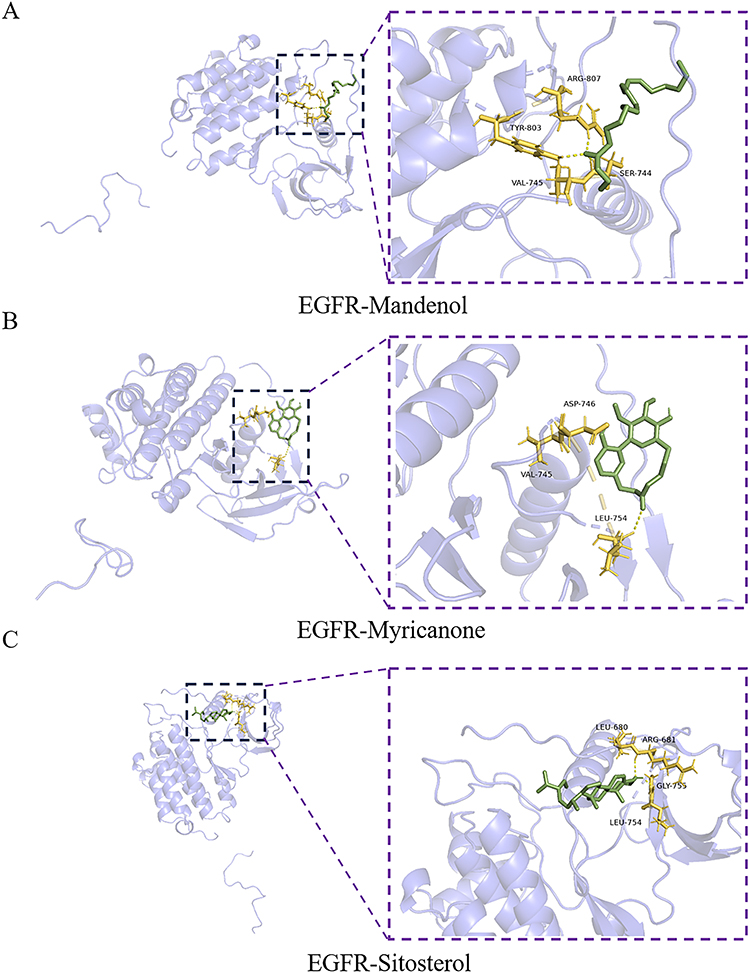 |
Figure 10 Molecular docking model of EGFR-active ingredients. (A) EGFR-Mandenol; (B) EGFR-Myricanone; (C) EGFR-Sitosterol. |
 |
Figure 11 Molecular docking model of HSP90AA1-active ingredients. (A) HSP90AA1-Mandenol; (B) HSP90AA1-Myricanone; (C) HSP90AA1-Sitosterol. |
 |
Figure 12 Molecular docking model of STAT3-active ingredients. (A) STAT3-Mandenol; (B) STAT3-Myricanone; (C) STAT3-Sitosterol. |
Experimental Validation of’s Active Ingredients in Modulating Target Gene Expression
The development of various cell lines, including Neuro 2A cells, SH-SY5Y, adipose-derived mesenchymal stem cells, and rat pheochromocytoma (PC12), has provided essential in vitro strategies for investigating Alzheimer’s disease mechanisms and assessing the neuroprotective properties of bioactive ingredients.33 Among these, Neuro 2A cell lines are extensively employed to examine neuronal differentiation, signal transduction pathways, and Aβ metabolism, making them a crucial tool for exploring the pathology of Alzheimer’s disease and assessing potential therapies. To assess the regulatory effects of Ligusticum wallichii’s active components on target gene expression, we executed CCK-8 and qRT-PCR analyses. The CCK-8 results confirmed that intervention with 80 μM Myricanone, Mandenol, and Sitosterol in Neuro 2A cells did not exhibit cytotoxicity (Figure 13A), prompting the use of this concentration in subsequent analyses. However, at 160 μM, a sharp drop in viability was observed, which is likely attributable to dose-dependent cytotoxicity. At high concentrations, lipophilic compounds such as Myricanone, Mandenol, and Sitosterol may destabilize cellular membranes and perturb homeostatic pathways, leading to non-linear toxic responses. This biphasic pattern, protective at moderate doses but harmful at excessive levels, is consistent with previous reports on natural product bioactivity. As depicted in Figures 13B–D, these compounds significantly suppressed the expression of HSP90AA1, EGFR, and STAT3. These findings at 80 μM offer preliminary but reliable evidence for the modulatory effects of Ligusticum wallichii’s bioactive ingredients, supporting insights gained from molecular docking and network pharmacology studies.
 |
Figure 13 Ligusticum wallichii’s active ingredients suppress the expression of crucial target genes. The measurement of cell viability was conducted with the CCK-8 assay. (A) Neuro 2A cells were treated with Mandenol, Myricanone, and Sitosterol at 0 μM, 10 μM, 20 μM, 40 μM, 80 μM, and 160 μM (n = 5). Neuro 2A cells were treated with Mandenol, Myricanone, and Sitosterol at 80 μM. Real-time qRT-PCR was used to examine the mRNA expression of EGFR (B), HSP90AA1 (C), and STAT3 (D) in Neuro 2A cells exposed to active ingredients (n = 5). Error bars indicate the standard deviation (SD) of five independent biological replicates; each measured in five technical replicates. |
Discussion
As a widely used traditional tonic herb, Ligusticum wallichii is known for its neuroprotective, anti-tumor, and anti-inflammatory effects.34,35 In this study, its potential therapeutic mechanisms against Alzheimer’s disease were explored through molecular docking and network pharmacology approaches. Traditional herbal medicine follows a holistic treatment philosophy, and network pharmacology aligns with this by employing a systems biology framework that emphasizes the interactions of multiple pathways, targets, and components in disease intervention. Meanwhile, molecular docking enables computational simulations to predict drug-target binding, aiding in the rational design of targeted therapeutics. By integrating these two methodologies, we systematically analyzed Ligusticum wallichii’s therapeutic mechanisms for complex diseases. Protein binding emerged as the most significantly enriched term in the GO analysis of 124 core targets, according to KEGG and GO database analysis. This suggests that Ligusticum wallichii could influence Alzheimer’s disease through its modulation of PPI. Additionally, within the 130 pathways enriched in KEGG, those associated with Alzheimer’s disease were highly represented, indicating that Ligusticum wallichii demonstrate neuroprotective properties by modulating key signaling pathways implicated in Alzheimer’s pathology.
Interestingly, several enriched cancer-associated pathways such as PI3K-Akt, EGFR tyrosine kinase inhibitor, and Rap1 signaling were also identified in our KEGG analysis. Although often categorized as cancer-related pathways, these signaling cascades are increasingly implicated in Alzheimer’s disease pathogenesis. PI3K-Akt signaling regulates neuronal survival, synaptic plasticity, and oxidative stress responses, while its dysregulation contributes to impaired neurogenesis and memory loss. EGFR signaling, when aberrantly activated, promotes neuroinflammation and amyloid pathology;36 recent evidence suggests that repurposing EGFR inhibitors can mitigate Aβ-induced toxicity and reactive astrocytosis in Alzheimer’s disease models.37 Rap1 signaling, traditionally associated with cell adhesion and migration, has also been linked to calcium homeostasis and neuronal hyperexcitability in Alzheimer’s disease. Pharmacological inhibition of Rap1-Pde6δ interactions was shown to normalize abnormal Ca²+ influx and restore synaptic function in Alzheimer’s disease models.31 These findings suggest that Ligusticum wallichii may exert pleiotropic neuroprotective effects by acting on signaling pathways that overlap with oncogenic networks but are also critical in neurodegeneration.
The neuroprotective effects of Ligusticum wallichii’s active ingredients on neurodegenerative diseases have been previously reported.35,38,39 Our study further demonstrated that Myricanone, Mandenol, and Sitosterol exert significant therapeutic potential against Alzheimer’s disease. Molecular docking analysis confirmed that these compounds effectively bind to HSP90AA1, EGFR, and STAT3, highlighting their key role in modulating disease-related pathways. These targets are not only critical for the pharmacological effects of Ligusticum wallichii but also have significant roles in the onset and evolution of Alzheimer’s disease. For example, studies have shown that dysregulated expression of HSP90AA1 significantly impacts synaptic stability and Aβ pathology in Alzheimer’s disease by influencing astroglial and microglial cell function.40 HSP90AA1 modulation also affects tau protein homeostasis, as it interacts with molecular chaperones and tau kinases, potentially reducing tau hyperphosphorylation and aggregation in neurons.41 Suppressing HSP90AA1 has been shown to reduce neuropathic pain and mitigate inflammation in chronic constrictive injury rats by regulating microglia-astrocyte interactions.42 EGFR inhibition has been reported to reduce reactive astrocyte activation, promote autophagy, mitigate Aβ-induced neurotoxicity and neuroinflammation, and facilitate axonal repair.37 Mechanistically, EGFR modulation may prevent Aβ accumulation and associated tau hyperphosphorylation by enhancing neuronal autophagic clearance and dampening pro-inflammatory signaling.43 Additionally, STAT3 is a multifunctional transcription factor that mediates neuroinflammation through the JAK2-STAT3 pathway in microglia, astrocytes, and hippocampal neurons. Inhibition of STAT3 suppresses microglial and astrocytic activation, reduces pro-inflammatory cytokine release, and thereby limits Aβ deposition and downstream tau phosphorylation.44,45 Together, these discoveries imply that Ligusticum wallichii’s bioactive ingredients offer protection against Alzheimer’s disease in vitro via targeting these key molecular regulators.
From a network pharmacology perspective, the mechanisms of Ligusticum wallichii show both similarities and differences when compared with other natural neuroprotective compounds such as ginsenosides and curcumin. Ginsenosides Rg1 and Rb1, identified through network pharmacology, exert neuroprotection against Alzheimer’s diseases mainly through anti-inflammatory, antioxidant, and anti-apoptotic effects mediated by PI3K/Akt, NF-κB, Nrf2, MAPKs, and Wnt/β-catenin pathways.46 Curcumin, likewise, has been predicted to target key Alzheimer’s diseases-related genes including APP, STAT3, MAPK1, and PPARγ and involves PI3K/Akt, BDNF/CREB, and autophagy-related pathways.47 Similar to ginsenosides and curcumin, Ligusticum wallichii also regulates the PI3K-Akt, EGFR tyrosine kinase inhibitor resistance, and Rap1 signaling pathways linked to Alzheimer’s diseases; however, its distinctive regulation of HSP90AA1 and EGFR provides unique therapeutic potential that complements these well-studied agents. This comparison highlights how network pharmacology can systematically reveal convergent and divergent pathways among different herbal medicines, thereby supporting rational multi-target strategies for Alzheimer’s diseases. In addition, our findings are consistent with previous experimental studies of Ligusticum wallichii in Alzheimer’s disease models. For instance, tetramethylpyrazine nitrone, a derivative of Ligusticum wallichii, significantly improved cognition and reduced amyloid-β and tau pathology in Alzheimer’s disease mice.48 These results reinforce the plausibility of our computational predictions.
Although this study clearly demonstrates that Ligusticum wallichii’s bioactive ingredients are capable of downregulating key Alzheimer’s disease-related target genes, there have been several limitations to be considered. Firstly, our in vitro studies mainly assessed gene expression suppression without directly evaluating functional consequences such as amyloid-beta deposition, tau phosphorylation, or neuroinflammatory responses. Secondly, positive controls using established Alzheimer’s disease drugs or known inhibitors were not included, and protein-level confirmation of target modulation, in vivo validation, as well as long-term effects was not performed. Thirdly, molecular docking provides predictive insights into ligand-target interactions but does not directly measure binding affinities, which would require techniques such as surface plasmon resonance or isothermal titration calorimetry. Moreover, we acknowledge that the TCMSP and SwissTargetPrediction databases, while useful for network construction and target prediction, have inherent algorithmic limitations and incomplete datasets that may introduce biases or omit relevant targets. Future studies should therefore focus on (i) in vivo validation of Ligusticum wallichii compounds in Alzheimer’s disease animal models, (ii) protein-level confirmation of predicted target engagement, (iii) pharmacokinetic and blood–brain barrier penetration studies, and (iv) investigation of potential synergistic effects when combined with other natural neuroprotective compounds such as ginsenosides or curcumin. These approaches will be crucial for translating our computational predictions into clinically meaningful therapeutic strategies.
Conclusion
Using an integrated network pharmacology and molecular docking strategy, this study identified Myricanone, Mandenol, and Sitosterol as the major bioactive compounds of Ligusticum wallichii that interact with critical Alzheimer’s disease-related targets, namely HSP90AA1, EGFR, and STAT3. In vitro assays further confirmed that these compounds significantly reduce the expression of these targets, suggesting a potential role in modulating amyloid aggregation, tau pathology, and neuroinflammation. By revealing these multitarget interactions, our findings advance the mechanistic understanding of how Ligusticum wallichii may exert neuroprotective effects in Alzheimer’s disease. Moreover, the identification of these compounds provides a promising basis for the development of novel multitarget therapeutic candidates derived from or inspired by traditional herbal medicine. Future in vivo validation and pharmacokinetic studies will be essential to translate these discoveries into clinically applicable strategies.
Data Sharing Statement
The datasets utilized or examined in this research could be obtained from the corresponding author if requested reasonably through suitable communication methods.
Disclosure
This article does not have any conflicts of interest.
References
1. Jia J, Ning Y, Chen M, et al. Biomarker changes during 20 years preceding alzheimer’s disease. N Engl J Med. 2024;390(8):712–722. doi:10.1056/NEJMoa2310168
2. Liu Y, Tan Y, Zhang Z, Yi M, Zhu L, Peng W. The interaction between ageing and Alzheimer’s disease: insights from the hallmarks of ageing. Transl Neurodegener. 2024;13(1):7. doi:10.1186/s40035-024-00397-x
3. Liu E, Zhang Y, Wang JZ. Updates in Alzheimer’s disease: from basic research to diagnosis and therapies. Transl Neurodegener. 2024;13(1):45. doi:10.1186/s40035-024-00432-x
4. Menkü BE, Akın S, Tamdemir SE, Genis B, Altıparmak T, Cosar B. Diagnostic transitions from primary psychiatric disorders to underlying medical conditions: a 5-year retrospective survey from a university hospital sample. Alpha Psychiatry. 2024;25(2):226–232. doi:10.5152/alphapsychiatry.2024.231274
5. Kim HG, Kim Y, Edmonds EC, Bondi MW. Comparison of cognitive impairment diagnosis criteria in clinical settings: conventional vs. Neuropsychological Alpha Psychiatry. 2024;25(2):212–219. doi:10.5152/alphapsychiatry.2024.231448
6. Bleibel M, El Cheikh A, Sadier NS, Abou-Abbas L. The effect of music therapy on cognitive functions in patients with Alzheimer’s disease: a systematic review of randomized controlled trials. Alzheimers Res Ther. 2023;15(1):65. doi:10.1186/s13195-023-01214-9
7. Buccellato FR, D’Anca M, Tartaglia GM, Del Fabbro M, Scarpini E, Galimberti D. Treatment of alzheimer’s disease: beyond symptomatic therapies. Int J Mol Sci. 2023;24:18. doi:10.3390/ijms241813900
8. Yuan L, Ye J, Wang W, et al. Research of arithmetic and drawing writing in improving communication and cognitive function in patients with mild-to-moderate dementia: a cluster randomized controlled trial. Alpha Psychiatry. 2024;25(2):262–268. doi:10.5152/alphapsychiatry.2023.1470
9. Ma L, Jiang X, Huang Q, et al. Traditional Chinese medicine for the treatment of Alzheimer’s disease: a focus on the microbiota-gut-brain axis. Biomed Pharmacother. 2023;165:115244. doi:10.1016/j.biopha.2023.115244
10. Fang Z, Tang Y, Ying J, Tang C, Wang Q. Traditional Chinese medicine for anti-Alzheimer’s disease: berberine and evodiamine from Evodia rutaecarpa. Chin Med. 2020;15:82. doi:10.1186/s13020-020-00359-1
11. Tan W, Qi L, Hu X, Tan Z. Research progress in traditional Chinese medicine in the treatment of Alzheimer’s disease and related dementias. Front Pharmacol. 2022;13:921794. doi:10.3389/fphar.2022.921794
12. Chen Z, Wang X, Du S, et al. A review on traditional Chinese medicine natural products and acupuncture intervention for Alzheimer’s disease based on the neuroinflammatory. Chin Med. 2024;19(1):35. doi:10.1186/s13020-024-00900-6
13. Pang HQ, Guo JX, Yang Y, et al. Elucidating the chemical interaction effects of herb pair Danshen-Chuanxiong and its anti-ischemic stroke activities evaluation. J Ethnopharmacol. 2024;318(Pt B):117058. doi:10.1016/j.jep.2023.117058
14. Xing Z, Peng F, Chen Y, Wan F, Peng C, Li D. Metabolomic profiling integrated with molecular exploring delineates the action of Ligusticum chuanxiong hort. on migraine. Phytomedicine. 2024;134:155977. doi:10.1016/j.phymed.2024.155977
15. Chang CY, Wu CC, Pan PH, et al. Tetramethylpyrazine alleviates mitochondrial abnormality in models of cerebral ischemia and oxygen/glucose deprivation Reoxygenation. Exp Neurol. 2023;367:114468. doi:10.1016/j.expneurol.2023.114468
16. Zhong KX, Zeng Q, Tang H, Tang B, Wang H. Tetramethylpyrazine attenuates cerebral ischemia-reperfusion injury by inhibiting ferroptosis via the AMPK / Nrf2 pathways. J Stroke Cerebrovasc Dis. 2025;34(2):108196. doi:10.1016/j.jstrokecerebrovasdis.2024.108196
17. Zhou X, Zhu Z, Kuang S, et al. Tetramethylpyrazine Nitrone (TBN) reduces amyloid β deposition in alzheimer’s disease models by modulating app expression, bace1 activity, and autophagy pathways. Pharmaceuticals. 2024;17(8):1005. doi:10.3390/ph17081005
18. Zhang P, Zhang D, Zhou W, et al. Network pharmacology: towards the artificial intelligence-based precision traditional Chinese medicine. Brief Bioinform. 2023;25:1. doi:10.1093/bib/bbad518
19. Zhao L, Zhang H, Li N, et al. Network pharmacology, a promising approach to reveal the pharmacology mechanism of Chinese medicine formula. J Ethnopharmacol. 2023;309:116306. doi:10.1016/j.jep.2023.116306
20. Li X, Liu Z, Liao J, Chen Q, Lu X, Fan X. Network pharmacology approaches for research of Traditional Chinese Medicines. Chin J Nat Med. 2023;21(5):323–332. doi:10.1016/S1875-5364(23)60429-7
21. Pinzi L, Rastelli G. Molecular Docking: shifting Paradigms in Drug Discovery. Int J Mol Sci. 2019;20:18. doi:10.3390/ijms20184331
22. Sahu D, Rathor LS, Dwivedi SD, et al. A review on molecular docking as an interpretative tool for molecular targets in disease management. Assay Drug Dev Technol. 2024;22(1):40–50. doi:10.1089/adt.2023.060
23. Cheung S, Zhong Y, Wu L, et al. Mechanism interpretation of Guhan Yangshengjing for protection against Alzheimer’s disease by network pharmacology and molecular docking. J Ethnopharmacol. 2024;328:117976. doi:10.1016/j.jep.2024.117976
24. Selvaraj M, Sadasivam K, Jothimani M, Muthusamy K. A comprehensive computational perspective in drug discovery for alzheimer’s disease. Comb Chem High Throughput Screen. 2023;26(12):2113–2123. doi:10.2174/1386207325666220606142910
25. Wang T, Zhang X, Liu W, et al. Identification of diagnostic molecules and potential traditional Chinese medicine components for Alzheimer’s disease by single cell RNA sequencing combined with a systematic framework for network pharmacology. Front Med Lausanne. 2023;10:1335512. doi:10.3389/fmed.2023.1335512
26. Zhang JC, Zhang HL, Xin XY, et al. Mechanisms of Bushen Tiaoxue Granules against controlled ovarian hyperstimulation-induced abnormal morphology of endometrium based on network pharmacology. J Ovarian Res. 2024;17(1):25. doi:10.1186/s13048-023-01339-3
27. Shi T, Hou C, Duan Y, et al. Mechanism of Smilax China L. in the treatment of intrauterine adhesions based on network pharmacology, molecular docking and experimental validation. BMC Complement Med Ther. 2024;24(1):150. doi:10.1186/s12906-024-04414-4
28. Choi HJ, Jeong YJ, Kim J, Hoe HS. EGFR is a potential dual molecular target for cancer and Alzheimer’s disease. Front Pharmacol. 2023;14:1238639. doi:10.3389/fphar.2023.1238639
29. Liu M, Pan J, Li X, et al. Interleukin-6 deficiency reduces neuroinflammation by inhibiting the STAT3-cGAS-STING pathway in Alzheimer’s disease mice. J Neuroinflammation. 2024;21(1):282. doi:10.1186/s12974-024-03277-3
30. Huang Q, Zhang Y, Huang L, Guo Z, Wu D. Research on the protective effect of Rhizoma of Anemarrhena asphodeloides on TMT induced AD mice model based on network pharmacology combined with in vitro and in vivo experimental validation. J Pharmacol Sci. 2025;157(2):75–87. doi:10.1016/j.jphs.2024.12.005
31. Dumbacher M, Van Dooren T, Princen K, et al. Modifying Rap1-signalling by targeting Pde6δ is neuroprotective in models of Alzheimer’s disease. Mol Neurodegener. 2018;13(1):50. doi:10.1186/s13024-018-0283-3
32. Huang J, Xu Z, Yu C, et al. The volatile oil of Acorus tatarinowii Schott ameliorates Alzheimer’s disease through improving insulin resistance via activating the PI3K/AKT pathway. Phytomedicine. 2024;135:156168. doi:10.1016/j.phymed.2024.156168
33. Vicente-Zurdo D, Rosales-Conrado N, León-González ME. Unravelling the in vitro and in vivo potential of selenium nanoparticles in Alzheimer’s disease: a bioanalytical review. Talanta. 2024;269:125519. doi:10.1016/j.talanta.2023.125519
34. Wei Z, Dong C, Guan L, Wang Y, Huang J, Wen X. A metabolic exploration of the protective effect of Ligusticum wallichii on IL-1β-injured mouse chondrocytes. Chin Med. 2020;15:12. doi:10.1186/s13020-020-0295-0
35. He P, Wang Z, Yang J, et al. Mechanism of ligusticum wallichii-borneol in the treatment of cerebral ischemic stroke in rats based on network pharmacology, molecular docking, and experimental verification. Chem Biodivers. 2024;e202401893.
36. Chen Y, Zhang Y, Chen Q, et al. Inhibition of mGluR5/PI3K-AKT Pathway Alleviates Alzheimer’s disease-like pathology through the activation of autophagy in 5XFAD mice. J Alzheimers Dis. 2023;91(3):1197–1214. doi:10.3233/JAD-221058
37. Mansour HM, Fawzy HM, El-Khatib AS, Khattab MM. Potential repositioning of anti-cancer EGFR inhibitors in alzheimer’s disease: current perspectives and challenging prospects. Neuroscience. 2021;469:191–196. doi:10.1016/j.neuroscience.2021.06.013
38. Lin X, Xu B, Zhang Z, et al. Proteomic analysis reveals the potential neuroprotective effects of tetramethylpyrazine dimer in neuro2a/APPswe cells. RSC Adv. 2019;9(33):18776–18784. doi:10.1039/C9RA03054A
39. Tan F, Fu W, Cheng N, Meng DI, Gu Y. Ligustrazine reduces blood-brain barrier permeability in a rat model of focal cerebral ischemia and reperfusion. Exp Ther Med. 2015;9(5):1757–1762. doi:10.3892/etm.2015.2365
40. Astillero-Lopez V, Villar-Conde S, Gonzalez-Rodriguez M, et al. Proteomic analysis identifies HSP90AA1, PTK2B, and ANXA2 in the human entorhinal cortex in Alzheimer’s disease: potential role in synaptic homeostasis and Aβ pathology through microglial and astroglial cells. Brain Pathol. 2024;34(4):e13235. doi:10.1111/bpa.13235
41. Sordu P, Alaylıoğlu M, Samancı B, et al. Cerebrospinal fluid HSP90AA1, HSPA4, and STUB1/CHIP levels in Alzheimer’s disease, mild cognitive impairment, and frontotemporal dementia. J Alzheimers Dis. 2025;105(2):481–493. doi:10.1177/13872877251329540
42. Luo F, Zhang J, Miao Y, Wu D, Shen H, Lu M. Paeoniflorin regulates microglia-astrocyte crosstalk, inhibits inflammatory response, and alleviates neuropathic pain through HSP90AA1/HMGB1 signaling pathway. Int J Biochem Cell Biol. 2024;176:106675. doi:10.1016/j.biocel.2024.106675
43. Lee HJ, Hwang JW, Kim J, et al. Erlotinib regulates short-term memory, tau/Aβ pathology, and astrogliosis in mouse models of AD. Front Immunol. 2024;15:1421455. doi:10.3389/fimmu.2024.1421455
44. Reichenbach N, Delekate A, Plescher M, et al. Inhibition of Stat3-mediated astrogliosis ameliorates pathology in an Alzheimer’s disease model. EMBO Mol Med. 2019;11(2). doi:10.15252/emmm.201809665
45. Choi M, Kim H, Yang EJ, Kim H. Inhibition of STAT3 phosphorylation attenuates impairments in learning and memory in 5XFAD mice, an animal model of Alzheimer’s disease. J Pharmacol Sci. 2020;143(4):290–299. doi:10.1016/j.jphs.2020.05.009
46. Jiang M, Chi J, Qiao Y, et al. Rb1 and rare ginsenosides: promising candidate agents for Parkinson’s disease and Alzheimer’s disease and network pharmacology analysis. Pharmacol Res. 2025;212:107578. doi:10.1016/j.phrs.2025.107578
47. Lou S, Gong D, Yang M, Qiu Q, Luo J, Chen T. Curcumin improves neurogenesis in alzheimer’s disease mice via the upregulation of Wnt/β-catenin and BDNF. Int J Mol Sci. 2024;25:10. doi:10.3390/ijms25105123
48. Zhou X, Huang K, Wang Y, et al. Evaluation of therapeutic effects of tetramethylpyrazine nitrone in Alzheimer’s disease mouse model and proteomics analysis. Front Pharmacol. 2023;14:1082602. doi:10.3389/fphar.2023.1082602
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Anti-Inflammatory Effects and Molecular Mechanisms of Shenmai Injection in Treating Acute Pancreatitis: Network Pharmacology Analysis and Experimental Verification
He Y, Hu C, Liu S, Xu M, Liang G, Du D, Liu T, Cai F, Chen Z, Tan Q, Deng L, Xia Q
Drug Design, Development and Therapy 2022, 16:2479-2495
Published Date: 2 August 2022
A Novel Approach Based on Gut Microbiota Analysis and Network Pharmacology to Explain the Mechanisms of Action of Cichorium intybus L. Formula in the Improvement of Hyperuricemic Nephropathy in Rats

Amatjan M, Li N, He P, Zhang B, Mai X, Jiang Q, Xie H, Shao X
Drug Design, Development and Therapy 2023, 17:107-128
Published Date: 20 January 2023
Network Pharmacology and Experimental Validation to Explore That Celastrol Targeting PTEN is the Potential Mechanism of Tripterygium wilfordii (Lév.) Hutch Against IgA Nephropathy

Zhao J, Liu H, Xia M, Chen Q, Wan L, Leng B, Tang C, Chen G, Liu Y, Zhang L, Liu H
Drug Design, Development and Therapy 2023, 17:887-900
Published Date: 23 March 2023
Investigating the Mechanism of Action of Schisandra chinensis Combined with Coenzyme Q10 in the Treatment of Heart Failure Based on PI3K-AKT Pathway

Wen S, Yang K, Bai Y, Wu Y, Liu D, Wu X, Zhang X, Sun J
Drug Design, Development and Therapy 2023, 17:939-957
Published Date: 27 March 2023
Network Pharmacology Analysis of the Mechanisms Underlying the Therapeutic Effects of Yangjing Zhongyu Tang on Thin Endometrium
Zhang L, Li H, Zhang L, Zu Z, Xu D, Zhang J
Drug Design, Development and Therapy 2023, 17:1805-1818
Published Date: 17 June 2023