Back to Journals » International Journal of Nanomedicine » Volume 21
Nano-Immunotherapy Targeting TAMs: Precisely Regulating TAMs to Reverse Immunosuppressive TME
Authors Zhang JM, Dong CL ![]() , Li B
, Li B ![]()
Received 4 February 2026
Accepted for publication 21 May 2026
Published 17 June 2026 Volume 2026:21 601103
DOI https://doi.org/10.2147/IJN.S601103
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Lijie Grace Zhang
Jin-Ming Zhang,1 Chun-Ling Dong,2 Bo Li1
1Department of Oral Anatomy and Physiology, Jilin Provincial Key Laboratory of Oral Biomedical Engineering, Hospital of Stomatology, Jilin University, Changchun, People’s Republic of China; 2Department of Pulmonary and Critical Care Medicine, Second Hospital, Jilin University, Changchun, People’s Republic of China
Correspondence: Chun-Ling Dong, Department of Pulmonary and Critical Care Medicine, Second Hospital, Jilin University, Changchun, People’s Republic of China, Email [email protected] Bo Li, Department of Oral Anatomy and Physiology, Jilin Provincial Key Laboratory of Oral Biomedical Engineering, Hospital of Stomatology, Jilin University, Changchun, People’s Republic of China, Email [email protected]
Abstract: Given the crucial role of tumor-associated macrophages (TAMs) in cancer development, nano-immunotherapy targeting TAMs represents a pivotal strategy for reversing the immunosuppressive tumor microenvironment (TME) and inhibiting cancer progression. Nano-drug delivery systems have evolved into multimodal platforms that utilize multi-drug synergy and integrated physical energy interventions, showing considerable potential for enhancing drug targeting and biocompatibility. The core objective of TAM-targeted nano-immunotherapy lies in the precise regulation and functional remodeling of TAMs, as well as the synergistic reversal of the immunosuppressive TME. This review addresses the knowledge gaps and challenges in nano-immunotherapy targeting TAMs and explores recent advances in TAM-targeted immunotherapy using various strategies and nanocarriers, with particular emphasis on the relevant cell surface receptors and downstream signaling pathways. Nano-immunotherapeutic strategies targeting TAMs include reprogramming TAMs, influencing TAM polarization, regulating TAM-secreted mediators, inducing TAM exhaustion, enhancing TAM phagocytosis, modulating TAM metabolism, inhibiting TAM recruitment, and blocking PD-L1 expression on TAMs. The innovations of nanocarriers involve the entire chain intelligent design from inorganic nanocrystals, organic polymers to organic-inorganic hybrid systems, and the precise “Trojan horses” constructed using biodegradable polymers and biomimetic membrane vesicles. Intelligent nanorobots targeting TAMs, as autonomous diagnostic and therapeutic systems integrating sensing, decision-making, and execution, represent an effective approach to enhancing the precision and intelligence of anti-tumor strategies. The image shows a central cell (TAM) surrounded by various nano-drug delivery systems. In the top left, a diagram illustrates stemness with arrows pointing from a central cell to various differentiated cells. Below, a section labeled ’Metastasis’ shows tumor-associated macrophages and cytokines interacting with tumor cells, indicating movement across a membrane. The central image depicts a large cell with various nano-drug delivery systems inside. In the top right, a diagram labeled ’Drug resistance’ shows self-renewal and downregulation of MHC-I involving CD8 T cells and TCR. The bottom right section labeled ’Immunosuppressive TME’ illustrates a tumor microenvironment with various immune cells and structures. Each section highlights different aspects of tumor progression influenced by nano-immunotherapy targeting TAMs.TAM surrounded by various nano-drug delivery systems: stemness, metastasis, drug resistance and immunosuppressive TME.
Keywords: tumor-associated macrophages, immunotherapy, targeting, nanocarrier, cancer
Introduction
Tumor-associated macrophages (TAMs) are among the most abundant immune cells in the tumor microenvironment (TME) and play a core role in tumor progression (Table 1).1 Ghamangiz et al classified macrophage populations into the following phenotypes: M1, M2 (M2a–d), M3, M4, M17, Mreg, Mmox, Mhem, M(Hb), and HA-Mac.2 The functional subgroups of TAMs include IFN-TAMs (CXCL9+/ISG+), Reg-TAMs, Inflam-TAMs, LA-TAMs, Angio-TAMs, RTM-TAMs, Prolif-TAMs, FCN1+ TAMs, SPP1+ TAMs, C1Q+ TAMs, TREM2+ TAMs, and CCL18+ TAMs.3,4 TAMs are functionally plastic, exhibiting both anti-tumor (M1) and pro-tumor (M2) functions during cancer progression.5–10 M1 TAMs exhibit pro-inflammatory properties, characterized by their potent antigen presentation capability and secretion of pro-inflammatory cytokines (IL-1, IL-6, IL-12, IL-18, IL-23, and TNF-α).11 M2 TAMs are characterized by weak antigen presentation ability and anti-inflammatory effects and secrete anti-inflammatory cytokines to create an immunosuppressive environment, facilitating tumor growth and neovascularization.10,11 You et al and Jiang et al demonstrated that M1 TAMs can exhibit pro-tumor properties.12,13 Therefore, anti-tumor and pro-tumor phenotypes are currently primarily used to define functional TAM subtypes. Given the crucial role of TAMs in cancer development, nano-immunotherapy targeting TAMs represents a pivotal strategy for reversing the immunosuppressive TME and inhibiting cancer progression.
 |
Table 1 List of Abbreviations |
Nano-drug delivery systems (NDSs) are a class of advanced drug delivery platforms that encapsulate, adsorb, conjugate, or disperse therapeutic agents within nanoscale carriers, thereby enabling precise control over their in vivo fate (absorption, distribution, metabolism, and excretion). With the continuous innovation of nanotechnology, NDSs have evolved into multimodal platforms that utilize multidrug synergy and integrated physical energy interventions, showing considerable potential to enhance drug targeting and biocompatibility. Current nano-immunotherapies targeting TAMs focus mainly on the precise regulation of their recruitment, polarization, phagocytosis, exhaustion, and reprogramming. Key interventional mechanisms involve multiple signaling pathways and molecular interactions, including STAT3, TREM2, FLT3L, CD47-SIRP-α, CCL2/CCR2, the CCL5/CCR5 axis, and CSF-1/CSF-1R.14–16 Furthermore, engineered macrophages (either directly or indirectly modified as carriers), dual-targeting strategies (for both TAMs and cancer cells), and combination therapies have been developed.14–20 The core objective of TAM-targeted nano-immunotherapy lies in the precise regulation and functional remodeling of TAMs, as well as the synergistic reversal of the immunosuppressive TME.
This review addresses the knowledge gaps and challenges associated with nano-immunotherapy targeting TAMs and explores recent advances in TAM-targeted immunotherapy using various strategies and nanocarriers (Figure 1), with a particular emphasis on the relevant cell surface receptors and downstream signaling pathways. TAM heterogeneity, ECM-mediated physical barriers, and multi-agent therapeutic synergy have drawn significant attention, providing a unique perspective for nano-immunotherapy targeting TAMs.
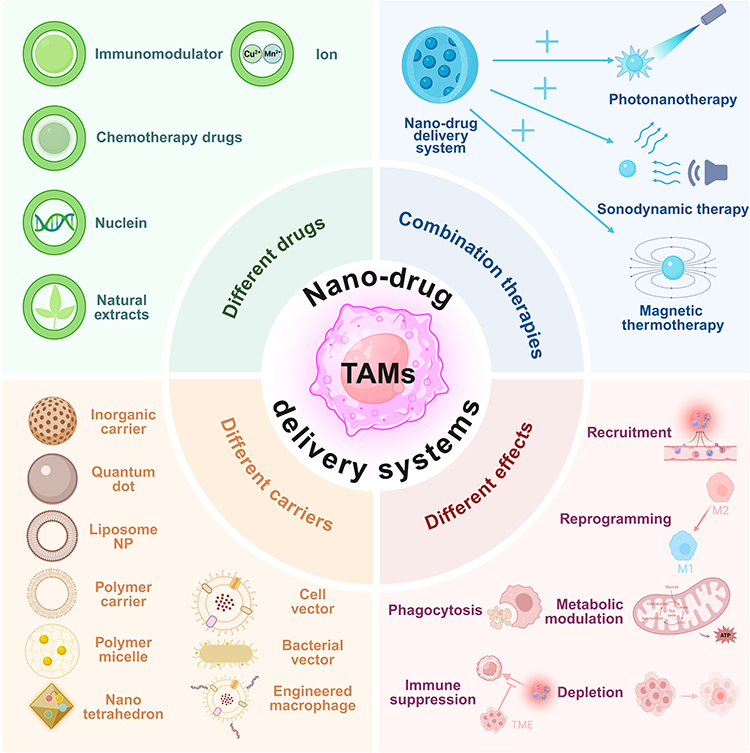 |
Figure 1 Overview of nano-drug delivery systems targeting TAMs. Nano-drug delivery systems targeting TAMs can be classified through the following approaches, different drugs, carriers, effects and combination therapies. “+” indicates combination therapy. The figure was drawn using Biorender. |
Challenges in Nano-Immunotherapy Targeting TAMs
Nano-immunotherapy targeting TAMs still faces formidable knowledge gaps and challenges, including TAM heterogeneity, the barrier effect of extracellular matrix (ECM), compensatory immunosuppression of the TME after intervention, and metabolic symbiosis between TAMs and cancer cells.
TAM Heterogeneity
TAM heterogeneity arises from cellular origin, education and reprogramming of the dynamic microenvironment, spatial distribution, cancer type specificity, and influence on tumor progression and therapeutic intervention.21,22 TAMs can be derived from adult bone marrow-derived monocytes and embryonic-derived tissue-resident macrophages. VSIG4+ TAM subpopulations derived from tissue-resident macrophages have been identified in various solid tumors such as testicular, hepatic, and lung cancer.23 In the regulation of dynamic microenvironments, metabolites, and cytokine signals in the TME precisely guide the phenotypic differentiation and spatial localization of TAMs.24 Moreover, the epigenetic suppression of GM-CSF-induced MHC-II expression leads to the spatial segregation of these functionally distinct TAM subsets within the TME.24 The functional state of TAMs is closely associated with their spatial distribution within tumors. CD68+SHP2+ immunosuppressive TAMs are enriched within tumor nests compared with the stroma in non-small cell lung cancer (NSCLC), and their density is correlated with poor patient survival.25 There are significant differences in the dominant TAM subpopulations among different cancer types and even different immune subtypes of the same cancer type. Furthermore, the composition and functional status of TAMs undergo dynamic evolution across the temporal dimensions of tumor progression and therapeutic interventions.26 To address TAM heterogeneity, strategies now emphasize subpopulation-specific identification and intervention. Single-cell RNA sequencing and spatial omics can be used to identify functional TAM subsets and their unique markers, enabling nano-targeted delivery systems that can distinguish tissue-resident TAMs (eg, VSIG4⁺) from monocyte-derived ones.27
Barrier Effect of ECM
The dense ECM within the TME acts as a critical physical barrier that impedes the penetration of both nano-drugs and immune cells, thereby limiting the efficacy of TAM-targeted nano-immunotherapy. The ECM restricts nanoparticle diffusion and compresses tumor blood vessels, thereby reducing drug delivery efficiency and creating a hypoxic microenvironment that further promotes M2-like TAM polarization.28–30 The current strategies for overcoming the ECM barrier include mainly inhibiting ECM formation, promoting ECM degradation, and remodeling the ECM, as well as altering the size of NDSs and applying surface modifications.28–32 However, the degradation of the tumor ECM may inadvertently promote the migration of cancer cells and subsequent metastasis, and the improper accumulation of these agents to overcome the ECM barrier in normal tissues can trigger adverse effects.28,29
Other Challenges
A variety of immunosuppressive cells, such as MDSCs, TANs, and Tregs, are widely present in the TME, and form a functionally redundant suppressive network with TAMs.33,34 When nano-immunotherapy eliminates or reprograms M2-type TAMs, the tumor can rapidly recruit MDSCs or TANs via chemokine signaling as a compensatory mechanism, thereby diminishing therapeutic efficacy. Nano-strategies that simultaneously modulate TAMs and MDSCs have demonstrated synergistic effects in overcoming compensatory immunosuppression in hepatocellular carcinoma models, indicating that targeting a single cell population is insufficient to fully reverse immunosuppression and underscoring the need for multi-target regulatory strategies.34
In terms of metabolic symbiosis, tumor cells abundantly secrete lactate, which, upon uptake by TAMs, drives their polarization toward an M2 phenotype through histone lactylation. Polarized M2-TAMs, in turn, increase tumor cell glycolysis, establishing a positive feedback loop that perpetuates malignancy.35 As a counterstrategy, GHB NPs disrupt the lactate metabolic coupling by blocking MCT1/MCT4 transporters, thereby reprogramming the TME.36 Moreover, the intrinsic metabolic vulnerabilities of TAMs, such as impaired glutamine metabolism, can be exploited to augment their antitumor functions.37
Biomimetic and metabolic modulation strategies are employed to resolve compensatory immunosuppression and metabolic symbiosis. Drug-free macrophage membrane-mimetic nanoparticles modulate TAM behavior without triggering compensatory immunosuppression, whereas the neutralization of acidic pH or lactate depletion alleviates macrophage dysfunction and prevents compensatory recruitment.
Nano-Immunotherapy Targeting TAMs
Nano-immunotherapeutic strategies targeting TAMs include reprogramming TAMs, influencing TAM polarization, regulating the expression of TAM-secreted mediators, inducing TAM exhaustion, enhancing TAM phagocytosis, modulating TAM metabolism, inhibiting TAM recruitment, and blocking PD-L1 expression on TAMs (Figure 2). The regulation of the pro-tumor phenotype of TAMs and the TAM-mediated immunosuppressive TME relies on the coordinated intervention of multiple pathways rather than a single target. The core design concept of NDSs lies in their ability to load drugs with different mechanisms (such as reprogramming agents, phagocytic signal regulators, immune checkpoint inhibitors, etc) onto a single platform, achieving spatiotemporal coordination. Accordingly, classification on the basis of drug quantity (single, dual, or multiple agents) directly reflects the complexity of the treatment strategy, the hierarchical integration of the underlying mechanisms, and the engineering difficulty of nanocarrier design. This logic transcends the differences in the pathways of TAMs among different cancer types and highlights synergistic immune regulation through nanotechnology.
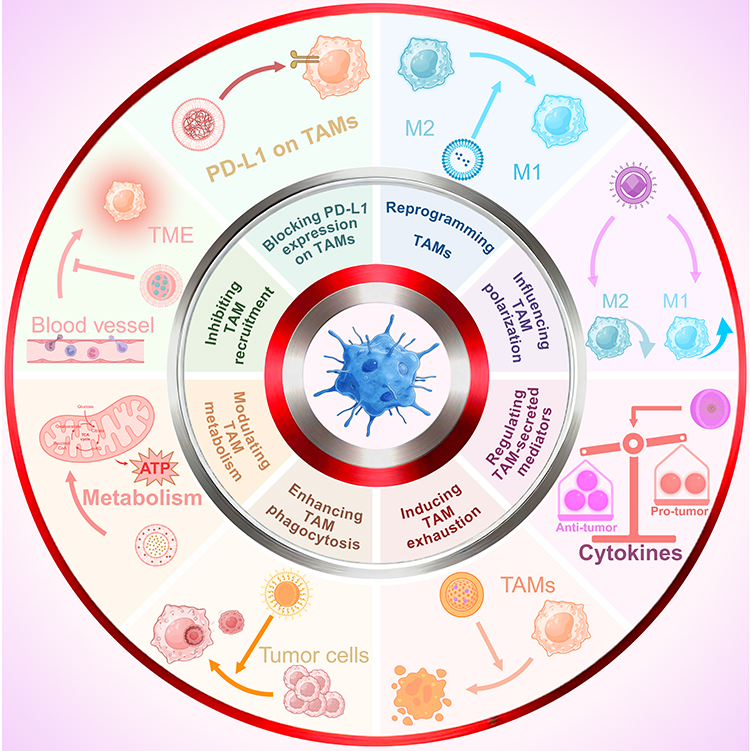 |
Figure 2 Nano-immunotherapeutic strategies targeting TAMs. Nano-immunotherapeutic strategies targeting TAMs include reprogramming TAMs, influencing TAM polarization, regulating TAM-secreted mediators, inducing TAM exhaustion, enhancing TAM phagocytosis, modulating TAM metabolism, inhibiting TAM recruitment, blocking PD-L1 expression on TAMs. “↑” indicates upregulation, while “↓” indicates downregulation. The figure was drawn using Biorender. |
Reprogramming TAMs
Single Drug
Inorganic Carriers
Inorganic carriers, such as graphene oxide and iron-based nanoscale metal‒organic frameworks, enable single-drug delivery for TAM reprogramming. The mannose-modified graphene oxide nanosystem PEG-GO-PEI-MAN@LDN193189 (PGPML) repolarizes TAMs from the M2 to M1 phenotype through ROS-mediated activation of NF-κB p65, MAPK p38, and JNK phosphorylation, thereby potentiating therapeutic efficacy in patients with HCC.38 Via the hepcidin/ferroportin-mediated reduction in iron efflux, Dic@M2pep-Fe-MOF, a nanoscale metal–organic framework, reprogrammed TAMs to the M1 phenotype and remodeled the immunosuppressive TME, generating long-term anti-tumor immune memory and promoting immunotherapy in hepatoma and breast cancer.39 The PGPML exploits a dual-targeting design to eliminate cancer stem cells and simultaneously reprogram immunosuppressive macrophages, thereby potentiating antitumor immunity in hepatocellular carcinoma. The synthetic complexity and potential immunogenicity of the graphene oxide backbone may impede scalable manufacturing and clinical translation. Functionalizing the platform with tumor-specific neoantigens could transform it into a broadly applicable personalized cancer vaccine.
Organic Carriers
DOX@FA-RHPs-SA transits TAMs from the M0 and M2 phenotypes to the M1 phenotype, promoting anti-tumor immune responses (ATIR) in breast cancer.40 MTX-PPT-micelles shift M2 to M1 TAMS via a traditional Chinese medicinal herb, astragalus polysaccharide (APS), improving tumor immunosuppression in ovarian cancer.41 CMA-R848 reprograms M2 TAMs to M1 cells via the activation of TLR7/8, reducing the secretion of IL-10 and increasing the secretion of IL-12, thereby increasing the response of gastric cancer (GC) to chemoimmunotherapy.42 5DEX-0509R (D-TAC) reprograms M2 to M1 TAMs via the TLR7 agonist DSP-0509, further inhibiting Treg recruitment, induction, and expansion and enhancing ATIR.43 BEA-C(sic)N-DOX-M reprograms M2 to M1 TAMs, promoting ATIR in HCC.44 SLNP@CpG reprograms M2 to M1 TAMs by activating the TLR9 pathway, thereby enhancing phagocytosis and antigen presentation function and promoting ATIR in T lymphoma and melanoma.45 FD9R-GW/siIRF3 reprograms M2 to M1 TAMs by downregulating the cooperative suppression of IRF3 and exosome release, promoting ATIR in breast cancer.46 Alb/LF NP albumin/lactoferrin hybrid nanoparticles carrying DDC/Cu-Fe reprogrammed M2 T cells to M1 TAMs via disulfiram, which inhibited FROUNT–chemokine receptor interactions and activated ATIR in glioma cells.47 P/PB/N/R nanoparticles reprogram M2 to M1 TAM phenotypes via the RP-182 peptide combined with the mannose receptor (MMR) CD206, promoting tumor immunotherapy of BSG.48 Pd-HA + Pd-M@R NPs, a bidirectional anisotropic palladium nanozyme, induces TAM transformation into the M1 phenotype and reprograms the immunosuppressive TME via ROS combined with R837 (agonist of TLR), promoting tumor immunotherapy in colon cancer.49 CpG-AS leads to TAM re-education via TLR9 agonists, ultimately reversing the tumor immune microenvironment in breast cancer and melanoma.50 Npgsa, which uses a PEG polymer as a carrier, repolarizes the immune-suppressive M2 subtype to anti-tumor M1 macrophages, promoting ATIR in melanoma and neuroblastoma.51 MG5-S-IMDQ converts M2 into the M1 phenotype via the use of a potent agonist of TLR7/8, imidazoquinoline (IMDQ), subsequently increasing the phagocytic activity of tumor cells, increasing the cytotoxic effects, increasing tumor antigen cross-presentation ability, and inducing macrophages to secrete TNF-α, IL-12p70, and IL-6 to combat GBM.52 The R848-GNE nanoemulsion reprograms M2 to M1 TAMs via R848, promoting ATIR of osteosarcoma.53 CSF-1Ri-loaded PMs ≈400 nm in diameter reprogram M2 to M1 TAMs via CSF-1Ri, promoting tumor immunotherapy in TNBC.54 PTX@Arg-CD-Bio/BM-HA-Man reprograms M2 to M1 TAMs, promoting ATIR in breast cancer.55 PLGA(H)-DOX@M/R837 reprograms M2 to M1 TAMs via R837 to further activate NF-κB signaling. They then secrete TNF-α and IL-6 to trigger direct tumor cytotoxicity and a CTL-mediated immune cascade, respectively, promoting tumor immunotherapy in patients with breast cancer.56
Polysaccharides, lipids, proteins, and nucleic acids have been designed to deliver single-agent therapies that repolarize TAMs from the M2 phenotype to the M1 phenotype, resulting in reversing immunosuppression and preventing tumor growth. Nevertheless, CSF 1R inhibitors induce systemic toxicity, peptide-based or natural product systems have poor stability or unclear processes, and TLR agonists run the danger of cytokine storms while having well-defined mechanisms.
Biological Carriers
Biological carriers can be classified according to the order of integrity of the organism: first, active intracellular substances such as enzymes; second, parts of cells such as exosome vesicles; and, third, the cell membrane to form the carrier plus some special ones last. MoS2@Pd-Man-mediated reprogramming of M2 to M1 TAMs and the immunosuppressive TME via catalysis of the in situ synthesis of the histone deacetylase inhibitor (HDACi) vorinostat enhances tumor growth inhibition and promotes tumor immunotherapy in patients with colon cancer.57 Repolarization of TAMs by M2Pep-Exo (pIC) via the TLR3 agonist poly(I:C) results in proliferation inhibition and apoptosis of breast cancer tumor cells.58 Lar@NP-OMV, which uses bacterial vesicles as carriers, reprograms M2 to M1 TAMs, resulting in nerve injury, and then counteracts GEM-induced neurogenesis in pancreatic cancer.59 The following two nanocarriers are characterized by their utilization of cancer cell membranes, which enables them to inherit the surface antigenic profile of host cells and enhance tumor targeting. DL@NP-M-M2pep-mediated reprogramming of M2 to M1 TAM phenotypes via modulation of the PI3K/Akt pathway promotes tumor immunotherapy in HCC.60 The PLGA-CpG@ID8-M nanovaccine leads to M1 polarization by increasing the expression of Gbp2 and promoting the recruitment of Pin1, which activates the NF-κB signaling pathway (Gbp2-Pin1-NF-κB pathway), promoting ATIR in ovarian cancer.61 HMMDN-Met@PM, which uses the macrophage membrane as a carrier, reprograms M2 to M1 TAM phenotypes via metformin, activating AMPK-NF-κB signaling, promoting tumor immunotherapy and inhibiting tumor growth and metastasis of subcutaneous breast tumors.62 Abs-CpG polarizes macrophages to the M1 type, and via CpG (a TLR9 agonist), a large amount of TNF-α is produced, which then activates cell division control protein 42 (Cdc42), enhancing the phagocytosis of adjacent macrophages and promoting tumor immunotherapy in breast cancer.63 R848@M2pep-MPs (AFPs) reprogram immunosuppressive M2 cells into the M1 TAM phenotype via the TLR7/8 agonist R848 and then present the AFP antigen to prime CD8⁺ T cells, thus providing an intratumoral niche that sustains and differentiates stem-like CD8⁺ T cells, enhancing anti-PD-1 therapy in patients with HCC and melanoma.64 PLGA NPs@PR-M reprogrammed M2 to M1 TAMs via the activation of TLR7 by R848, which regulates the secretion of cytokines, including IL-12, IFN-γ, TNF-α and IL-10, ultimately inducing ATIR in colorectal carcinoma.65 At present, it is mainly via the Gbp2-Pin1-NF-κB pathway and the PI3K/Akt/NF-κB pathway that a single drug is carried to reprogram TAMs with bioactive substances as carriers. Interestingly, Wei et al used bioorthogonal nanozymes for immunotherapy, and Zhao et al used apoptotic cancer cell bodies as carriers and cascade amplification to alleviate immunosuppression (Table 2).
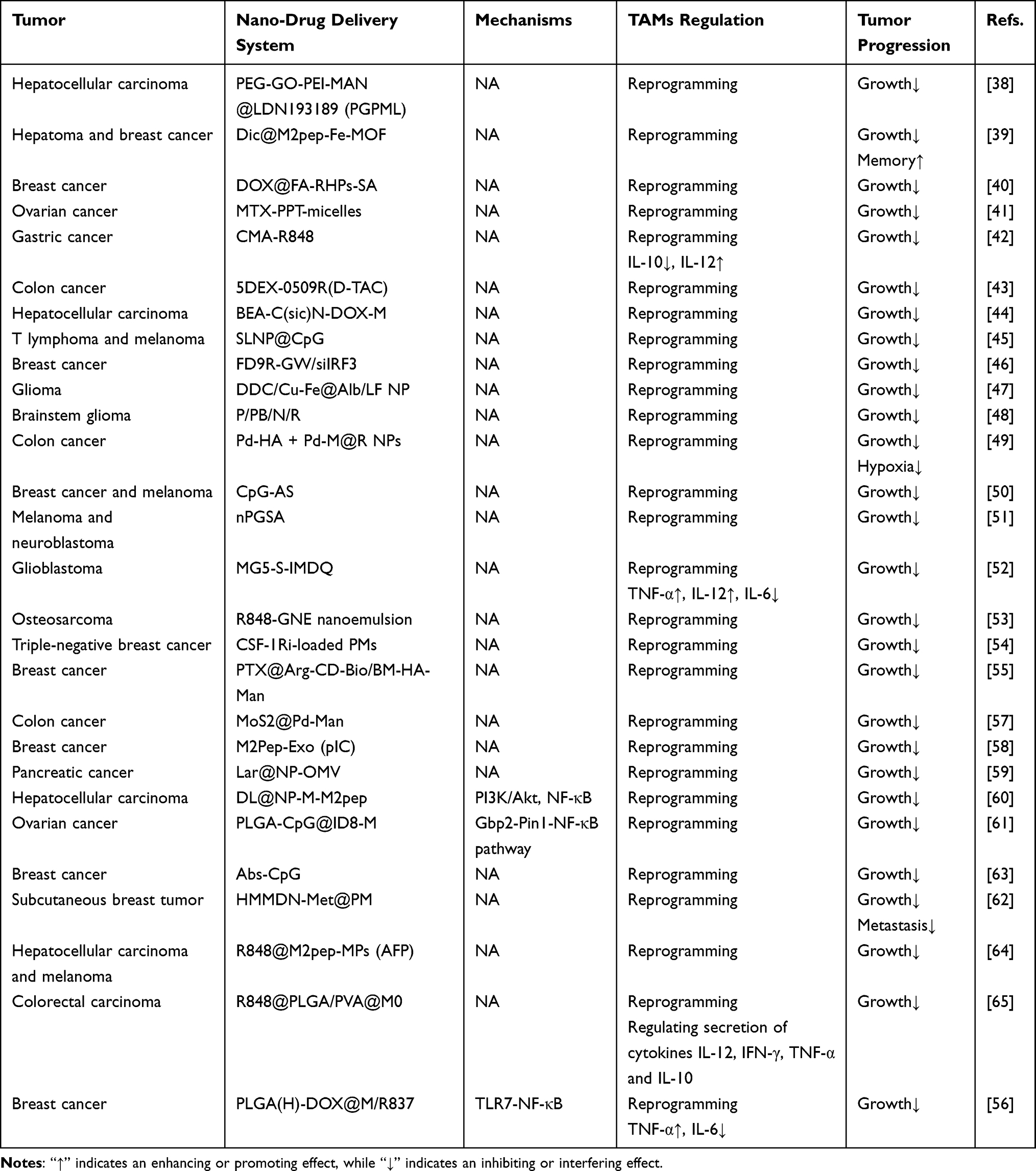 |
Table 2 Single Drug Delivery Nano-Immunotherapy Targeting TAM Reprogramming |
Exosomes, cell membrane structures, and apoptotic bodies are examples of biological carriers that provide high biocompatibility, minimal immunogenicity, and homologous targeting. TLR agonists that activate NF-κB or modify the PI3K/Akt and AMPK pathways are primarily responsible for reprogramming TAMs, through cascade reactions, a number of novel designs, such as bioorthogonal nanozymes, enhance immunological effects. The physical barrier to the ECM, heterogeneity of TAMs, and long-term in vivo safety have been completely overlooked by these systems, which also have limited scalability, high batch variability, and isolation challenges. To overcome physical obstacles and increase selectivity for TAM subsets, future approaches should integrate ECM modulation, smart responsive release, and genetic engineering of carriers.
Dual Drugs
Inorganic Carriers
An MOF-based caspase-1 nanoreporter (MCNR) (Caspase-1+R848) induces TAM reprogramming via R848-TLR7/8-NLRP3 and Fe-MAPK-NLRP3, enhancing T cell infiltration, which leads to tumor inhibition and improved survival rates in patients with breast cancer.66 This nanoplatform integrates real-time imaging feedback with immune-stimulating payload delivery, enabling visualized monitoring of the conversion of immunosuppressive macrophages into tumoricidal effectors. The inherent instability of metal-organic framework structures in physiological environments may cause premature drug leakage before reaching the intended tumor regions. Incorporating targeting ligands that respond to multiple microenvironmental cues could refine its diagnostic precision and broaden its applicability to heterogeneous solid tumors.
Organic Carriers
The coloaded liposome (CNLP) IDO-IN-7+cabozantinib reprograms M2 to M1 TAMs and the immunosuppressive TME via IDO-IN-7 and cabozantinib, promoting tumor immunotherapy in TNBC.67 PTX/DHA-FA-LNs reprogram M2 to M1 TAMs and restrict tumor growth in breast cancer.68 MAPDCs (CPT+R848) repolarize M2-GAMs to M1-GAMs via R848, increasing the expression of proinflammatory cytokines (IP-10, IL-12, and IL-6) to increase immune responses and resensitize tumors to chemotherapy, yielding a superior anti-GBM effect.69 SPADS, which combines α-tocopherol (Tpl) and toyocamycin (Tcc), reprograms M2 to M1 TAMs through the dual inhibition of the endoplasmic reticulum and oxidative stress via the synergistic inhibition of ROS and ER stress (STAT1 and STAT3) to induce the secretion of specific anti-tumor cytokines (TNF-α, IL-12, and Arg1 expression), thereby increasing macrophage phagocytic capacity and tumor-killing ability and suppressing cancer cell growth and migration.70 PIL-12+PLX@cR-PssPD promotes the repolarization of TAMs by blocking CSF-1/CSF-1R signaling and thus inhibits melanoma growth and metastasis.71 The meo-PEG-Dlinkm-PLGA copolymer (sorafenib+R848) reprograms M2 to M1 TAMs via R848-mediated repolarization, promoting tumor immunotherapy in HCC.72 GPCuD NPs (Cu+DOX) reprogram M2 to M1 TAMs and ITM and then enhance ATIR and the secretion of IL-6 and TNF-α by macrophages in breast cancer.73 PLGA-PA-DOX NPs reprogram M2 to M1 TAMs via a caspase-3-independent pathway and reduce the suppression of breast cancer cell viability and migration.74 Ultra-pH-sensitive NCPA reshapes macrophages to transform into tumoricidal M1-like cells via aCD47/aPD-L1 dual blockade, promoting ATIR in lung cancer.75 PPD/MPC (PEG-PLL-DMMA/MA-PEI-CpG) reprograms TAMs by activating TLR9 pathways, regulating the cytokine secretion of immune suppressive cells, alleviating intricate immune tolerance and remodeling the tumor immunosuppressive microenvironment, strengthening the immune response and evoking immune memory in melanoma.76 ST-NP@AS&Ab (AS+Ag9) reprograms M2 to M1 TAMs and the immunosuppressive TME via an AS-loaded nano-drug inhibiting signal transducer and activator of Transcription 6 (STAT6), promoting tumor immunotherapy in TNBC.77 PCPA&PPM@TR&PPM@TR (R848+TMZ) reprograms M2 to M1 TAMs via R848 (a TLR7/8 agonist), reducing GBM resistance to TMZ treatment.78 D&R-HM-MCA (DOX+R848) reprograms M2 to M1 TAMs by recognizing MMR on TAMs and releasing R848, leading to reprogramming of the immunosuppressive TME-enhancing ATIR of gliocytoma.79 CaNP@cAD-PEG composed of cAD loaded on CaNP enveloped with DSPE-PEG2000 could reprogram TAMs by simultaneously activating multiple inflammation-related signaling pathways (MAPK and NF-κB signaling pathways), as well as the NLRP3 inflammasome pathway, in melanoma and rectal cancer.80 R6RGD-CMβ CD@DOX NPs and M2pepCMCS@R848 NPs (RCNPDOX+MCNPR848 NPs) reprogram M2 to the M1 TAM phenotype and immunosuppressive TME by activating TLR7/8 and subsequently increasing the secretion of TNF-α to reshape the immunosuppressive tumor microenvironment, enhancing the effect of immunotherapy in breast cancer.81 R@rHDP-M2pep (OX+R837) reprograms M2 to M1 TAMs via imiquimod (R837, a hydrophobic TLR7 agonist), promoting the secretion of proinflammatory cytokines, activating CD8⁺ T cells, suppressing orthotopic tumor growth, and preventing lethal lung metastasis of TNBC.82 Au/Toy@G3 NGs, a dendrimer nanocarrier system, reprogram M2 to M1 TAMs via Au NPs, increasing the secretion of TNF-α as an effective anticancer therapy in pancreatic cancer.83 GD@PP/Wtmn (Wtmn+GEM), a raspberry-like nanocarrier system, reprograms M2 to M1 TAMs by inhibiting the PI3K pathway, synergizing with GEM to inhibit pancreatic tumor growth.84 PTX+CUR-loaded PLA/shellac-FA dimer NPs, a snowman-like nanocarrier system, reprograms M2 to M1 TAMs, promoting ATIR in pancreatic cancer.85
Dual-drug organic carriers co-load immune agonists with chemotherapeutics to achieve multidimensional TAM reprogramming and overcome resistance to single agents. Their translation is hindered by challenges in synchronizing release kinetics, optimizing drug ratios, and mitigating additive systemic toxicity, in addition to complex fabrication workflows. Future strategies should integrate sequential release schedules that match TAM polarization dynamics with ECM-degrading modules to enhance intratumoral penetration, thereby addressing functional heterogeneity and physical barriers simultaneously.
Biological Carriers
Biological carriers such as enzymes, cancer cell membranes, macrophages, exosomes, and bacteria can deliver dual drugs that precisely target TAMs as a result of their biocompatibility and subsequently reprogram them. BMCL nanozymes (pcCAT+pcLOX) reprogram M2 to M1 TAMs by inhibiting the activation of NF-κB, inducing tumor cell apoptosis and activating ATIR in colon cancer.86 CCM@HMON@CS-Pt(IV)@ANX5, which uses cancer cell membranes as carriers, reprograms M2 to M1 TAMs by initiating the TLR9/MyD88/NF-κB pathway and then suppresses efferocytosis, contributing to the activation of the innate immune response of GC.87 The following two nanocarriers are implemented using engineered macrophages as carriers, creating integrated systems that combine cellular motility with therapeutic payload delivery. Met@Man-MPs (Metformin+DOX) efficiently reprogrammed Doxil-enhanced M2 to M1 TAMs via doxil-induced activation of stimulator IFN genes (STING), facilitating macrophage maturation and antigen presentation to activate CD8⁺ T cells. It subsequently generates strong ATIR and long-term immune memory against tumor recurrence in liver cancer and breast cancer.88 Mam@NPs (IM+aCD47) effectively reprogram M2 to M1 TAMs by increasing CCL3 secretion via a combination of IM and aCD47, which further secretes inflammatory and anti-tumor cytokines, promoting ATIR in gliomas.89 The next two nanocarriers are exosome-based. CpG-EXO/TGM reprograms M2 to M1 TAMs, releasing related anti-tumor cytokines (TNF-α, IL-6, and IL-12) during this process, thereby inducing a series of ATIRs in GBM.90 SN/Mn@gHE (SN38+MnO2) reprograms M2 to M1 TAM phenotypes, promoting tumor immunotherapy in murine and murine colorectal cancer.91 OMV-CD47/GPC3 (anti-CD47 nanobodies + anti-GPC3 scFv), which uses bacterial vesicles as carriers, repolarizes TAMs to the M1 phenotype via PAMPs of OMVs, presenting TAAs to T cells and activating them, consequently initiating T-cell-specific tumor killing in HCC, thus providing a promising strategy for HCC immunotherapy.92 Among these varied approaches, the TLR9/MyD88/NF-κB signaling pathway represents one of the mechanisms through which TAM reprogramming is facilitated. Zhang, L. et al adopted the double-punch strategy, and Chen et al used a novel biomimetic nanocomposite containing copper and platinum.
Dual-drug biological carriers leverage enzymes, cell membranes, and engineered vesicles to co-deliver two agents that synergistically reprogram TAMs through multipathway engagement. Their clinical translation is constrained by the phenotypic instability of living carriers, risks of immunogenicity upon repeated dosing, and the absence of methods to independently control the release kinetics of each payload. Future approaches could integrate patient-derived avatars for prescreening to guide personalized dual-drug combinations and improve therapeutic precision.
Nanogels and Nanomicelles
D@HPMNG (metformin+DOX) reprograms TAM phenotypes via the inherited immunomodulatory effect of metformin, reshing the tumor immune microenvironment of melanoma.93 PP-RC-H reprograms M2 to M1 TAMs via the TLR7 agonist, R837, and increases the pH, promoting ATIR in head and neck squamous cell carcinoma (HNSCC).94 Gel/(REG+NG/LY) reprograms M2 to M1 TAMs, inhibiting tumor growth and liver metastases in colon cancer.95 ChiP-RS (SHP099+R848) reprograms M2 to M1 TAMs via R848, activating TLR7/8 and promoting tumor immunotherapy in mammary carcinoma.96 TfR/TATH7/PTX/R837 NMs promote M1 polarization of TAMs and reduce the immunosuppression phenomenon via R837 (an agonist of TLR7); they then regulate the secretion of cytokines (increasing TNF-α and IFN-γ and decreasing TGF-β), promoting tumor immunotherapy in normotensive glioma.97
Hydrogels have a significant benefit in that they can be administered intratumorally or peritumorally, which enhances drug retention and minimizes systemic toxicity. However, the in vivo disintegration of hydrogels is difficult to control and may cause burst or incomplete drug release, and micelles are limited by short circulation times and unresolved biosafety concerns. If the release pattern and rate are appropriately regulated, a dual-drug platform with hydrogels and micelles is promising as a successful immunotherapeutic method for locally progressed or postoperative recurring malignancies (Table 3).
 |
Table 3 Dual Drug Delivery Nano-Immunotherapy Targeting TAM Reprogramming |
Multiple Drugs
Certain NDSs have been engineered to co-deliver multiple therapeutic agents that produce synergistic effects, thereby enabling more effective TAM reprogramming through coordinated multipathway modulation. LPZ/DOX@M/R837@MPs reprogram M2 to M1 TAMs and alleviate the TME via activation of the NF-κB signaling pathway, regulating cytokine secretion (increasing IL-12 and decreasing IL-10), and promoting tumor immunotherapy in breast cancer, melanoma and colon tumors.98 Gox/CHA@MPNs reprogram M2 to M1 TAMs and the TME, promoting tumor immunotherapy and inhibiting tumor growth and metastasis in human breast carcinoma.99 Interestingly, Li et al used a multidrug, targeted local delivery system to sustainably remodel the acidic TME and enhance cancer immunotherapy.
Multi-agent co-delivery strategies counter compensatory resistance through synergistic mechanisms, offering advantages for highly heterogeneous and immunosuppressive solid tumors. Their clinical progress is limited by low manufacturing reproducibility during scale-up and a scarcity of preclinical models that recapitulate TAM diversity for rational multi-drug ratio optimization. Future development should integrate ECM-normalizing modules to improve intratumoral perfusion and employ single-cell spatial profiling to guide the design of TAM subset-discriminating delivery, and programmable sequential release systems that align with the dynamic evolution of TAM phenotypes are also essential.
Magnetothermal Therapy (MHT)
As a noninvasive physical modality, MHT reprograms TAMs through its unique capacity for spatially targeted thermal intervention, thereby altering the immunosuppressive tumor microenvironment. M-PUNCs relieve the ITM by repolarizing TAMs into the M1 phenotype via the UIONP moiety, promoting tumor immunotherapy in patients with breast cancer and melanoma.100 ZCMF@PEG-AF (ZCMF-AF) reprograms M2 to M1 TAMs via the IKKβ/IκBα/NF-κB pathway and subsequently induces lung cancer cell damage.101 FVIO reprograms M2 to M1 TAMs by interfering with the SIRPα pathway, promoting tumor immunotherapy in HCC.102 Among these varied approaches, the IKKβ/IκBα/NF-κB pathway represents one of the mechanisms through which TAM reprogramming is facilitated.
Compared with conventional drug delivery systems, MHT offers several advantages, including noninvasiveness, deep tissue penetration, remote controllability, and the avoidance of drug related toxicity. However, mechanistic investigations are still fragmented, and key issues such as thermal dosage, targeting precision, and the impact on the extracellular matrix remain poorly understood. The future potential is substantial, If combined with smart thermosensitive materials or immune agonists, MHT is expected to become a novel physical immunotherapy approach for modulating TAMs.
Photothermal Therapy
Inorganic Carriers
The photothermal–therapeutic hybrid NDS effectively reprograms TAMs through spatially and temporally controlled, unique photothermal effects. DLC-HA reprograms M2 to M1 TAMs via the ROS and cGAS-STING pathways, enhancing the efficacy of breast cancer immunotherapy.103 SnSe@ABS NS, a metal nanosheet, reprograms M2 to M1 TAMs via sustained lactate consumption and elevated intracellular oxidation levels in TAMs by SnSe NSs-mediated catalysis to amply ATIR in breast cancer.104 Fe-PHCN@DOX reprograms M2 macrophages to the M1 TAM phenotype by efficiently catalyzing H2O2 to produce O2 and relieve tumor hypoxia, promoting tumor immunotherapy in colon cancer.105 Some new NDSs, such as DLC-HA could exploit spatiotemporally controlled photothermal effects to simultaneously provoke endoplasmic reticulum stress and mitochondrial dysfunction, synergistically reprogramming the intratumoral immune landscape. The dual organelle-targeting approach risks collateral damage to peritumoral healthy cells that also internalize silica-based carriers, potentially causing unintended metabolic disruption. Future engineering efforts could harness its intrinsic photothermal property to generate in situ tumor lysates as personalized antigen reservoirs for neoantigen-based vaccination strategies.
Organic Carriers
Lipo Zol/IR NPs reprogram M2 to M1 TAMs via the immunoregulatory activity of Zol, leading to tumor death and mild immune activation in patients with breast cancer.106 AeroNP-CDNs, combined with phototherapy, reprogram M2 T cells to M1 TAMs by activating IFN gene stimulators in macrophages, mitigating the immunosuppressive TME of NSCLC107 M@SINPs transform M2 cells to the M1 TAM phenotype by generating intracellular ROS under laser irradiation, significantly inhibiting tumor growth in colon cancer.108 RPST@IMQ reprograms M2 to M1 TAMs by activating the TLR-7 pathway and enhancing phagocytosis, inflammation and antigen presentation in prostate cancer.109 PA1094T+ aCD47 polarizes TAMs to the M1 phenotype via CD47 inhibition, promoting macrophage-mediated phagocytosis of cancer cells and activating the immune response against GBM and eventually preventing tumor recurrence in GBM.110 G5-PEG-LyP-1-CuS-DMXAA NPs (GLCD NPs) repolarize M2-like macrophages to M1-like macrophages via DMXAA (a vascular disrupting agent), reversing the immunosuppressive TME and inducing ATIR in breast cancer.111 ChiP-CeR polarizes M2-type into M1-type TAMs via the TLR7/8 agonist imiquimod (R837), activating T-cell ATIR in combination with immunogenic cell death (ICD) in breast cancer.112 MCuLP nanosystems promote the repolarization of TAMs to anti-tumor M1-type macrophages via LOX, exhausting the lactate in the TME (HIF-1α) and enhancing cancer photothermal immunotherapy in patients with breast cancer.113 Ce6@PEIS-HA/Tf (CPHT) reprograms M2 to M1 TAMs to release a large amount of H2O2 via the Fenton reaction and produces hydroxyl radicals, causing tumor death in breast cancer.114 MCR reprograms M2 to M1 TAMs via the combined actions of ROS produced through photodynamic therapy (PDT) and R848 (a TLR7/8 agonist), increasing the secretion of IL-6 and TNF-α and suppressing ATIR in colon cancer.115 M.RGD@Cr-CTS-siYTHDF1 NPs reprogram M2 to M1 TAMs by depleting YTHDF1 and inhibiting STAT3 translation in a m6A-dependent manner, shifting the STAT3-STAT1 equilibrium to increase STAT1 expression, suppressing IL-10 production, and increasing the expression of IL-12 and IFN-γ, thereby remodeling TME and curbing HCC development.116 AIPH@MS-CTPP reprograms M2 to M1 TAMs via the photothermal effect of melanin and thermodynamic therapy of AIPH, enhancing systemic ATIR to combat breast cancer dissemination.117 DOX-PRO NPs and IR808/R848-PRO NPs, which integrate PDT, hypoxia-activated chemotherapy and immunotherapy, reprogram M2 to M1 TAMs via a TLR7/8 agonist, R848, to regulate cytokine secretion (TNF-α, IFN-γ, and IL-12p40 increase the decrease in IL-10 levels) and subsequently facilitate tumor cell elimination in breast cancer.118 IMD@CD/Ada-GMNPs reprogram M2 to M1 TAMs and TME through R837, leading to effective immunotherapy in breast cancer.119 R@L-MRS17 polarizes infiltrated macrophages into the M1 phenotype via the TLR7/8 agonist (R848), activating the positive ATIR and suppressing primary tumor growth in distant and metastatic tumors in breast cancer.120
Organic nanocarriers co-loaded with photosensitizers or photothermal agents and immunomodulators reprogram M2 macrophages to the M1 phenotype through light-triggered ROS or hyperthermia, which is reinforced by immunogenic cell death. Clinical application is limited by loading leakage, poor laser penetration in deep tumors, and the neglect of the phenotypic diversity of TAMs, which causes inconsistent polarization responses. Future designs should integrate ECM-lytic modules into photo-responsive carriers to improve light distribution and nanoparticle access within the desmoplastic stroma, and single-cell guided spatiotemporal light dosing that discriminates between tumor-promoting and homeostatic macrophage subsets will further enable precise reprogramming.
Biological Carriers
DFMO + N-aP@ICG synergistic light therapy repolarizes TAMs by restraining the expression of aconitate decarboxylase 1 (ACOD1) and releasing aPD-L1, which binds to PD-L1 on TAMs, reversing immune suppression at the tumor site and inhibiting the proliferation of glioma.121 ICG/CMSN@GsE repolarizes M2-like macrophages to the M1 phenotype by entering the nucleus, mediated by a variety of membrane receptors, such as MyD88, TLR2/4, TNFR 1, IL-17R, CD40, and RAGE, thereby increasing their transcription, upregulating the expression of upstream mediators (MyD88 and NF-κB), and culminating the production of proinflammatory cytokines (TNF-α and IL-12), enhancing cancer immunotherapy efficacy in breast cancer.122 RB-OL@M reprograms M2 to M1 TAMs by activating TLR-7 in the NF-κB signaling pathway, ultimately cooperating with phototherapy-driven ICD to increase therapeutic efficacy against breast cancer.123 CW-camouflaged nanospores repolarize immunosuppressive TAMs via the activation of STING and can effectively suppress the Rubicon–LAP–efferocytosis axis, thereby increasing the expression of IFN-β and the proliferation and promotion of ATIR in melanoma and breast cancer.124
The biocarriers described above, including microglial vesicles, ginseng exosomes, germs, and fungi, are loaded with photosensitizers or photothermal agents in combination with drugs to reprogram TAMs with improved biocompatibility and targeting capability. However, this approach still faces certain issues, such as biosafety concerns. In summary, in combination with the photothermal effect, the aforementioned carriers achieved TAM reprogramming through various mechanisms, including the induction of the (TLR2/4, TNFR 1, IL-17R, CD40, and RAGE)-MyD88-NF-κB, STING, and TLR-7-NF-κB signaling pathways.
NDSs subsequently employ nanomotors or various cell membranes, including erythrocyte membranes, platelet membranes, tumor cell membranes, and macrophage membranes, in combination with photothermal therapy to achieve TAM reprogramming. HPA/AS/CQ@Man-EM reprograms M2 to M1 TAMs through p38 and NF-κB in response to artemisinin derivatives, effectively suppressing tumor growth in colorectal cancer.125 MOAP reprograms M2 to M1 TAMs via MnOx (a hydrogen peroxide nanozyme), alleviating hypoxia and suppressing postresection relapse and dissemination in patients with breast cancer.126 Au@Fe-PMs reprogram M2 to M1 TAMs via P38- and STAT1-mediated pathways, promoting ATIR mediated by NIR-II PTT in breast cancer.127 RNP@PNM-IR reprograms M2 macrophages into M1 macrophages via R848, thereby enhancing the phagocytic function of macrophages, which effectively inhibits postsurgical tumor recurrence and metastasis in patients with breast cancer.128 Rg3-Exos/ATO@Ce6 reprograms M2 to M1 TAMs and the immunosuppressive TME via the immunomodulatory effects of Rg3, promoting tumor immunotherapy in GBM.129 SEx@Fc-NPs convert M2 macrophages via generation of ROS by the type-I photosensitizer and blocking the immune checkpoint protein Siglec-10, further enhancing the ATIR of M1 macrophages in breast cancer.130 CS-I/J@CM NPs repolarize M2 macrophages into the M1 phenotype by generating high levels of ROS and oxygen through a Fenton-like reaction to relieve the suppression of the TME in GBM.131 P-I@M1E/AAL reprograms M2 to M1 TAMs by alleviating cancer hypoxia, releasing proinflammatory cytokines reprograming the TME, and augmenting PDT immunotherapy in TNBC.132 SPI@hEL-RS17 reduces TAM phenotypes, exerting potent anti-tumor efficacy against both breast tumors and melanoma.133 IC@MSA JNM reprograms M2 to M1 TAMs via ibrutinib (IBR), the secretion of proinflammatory cytokines, and the mobilization of T cells for tumor clearance, collectively suppressing breast cancer progression.134 Pd@ZIF-8/R848@M JNMs reprogram M2 to M1 TAMs via R848, thereby converting the TME to activate ATIR and achieve immunotherapy in HCC.135 Sheng et al hybridized platelet membranes and neutrophil membranes to obtain a new membrane system to better target and reprogram TAMs. Cell membrane-camouflaged nanomotors combine prolonged circulation, homologous targeting, and autonomous motility to reprogram TAMs via photothermal effects and the Siglec-1-BTK pathway. Their progress is limited by incomplete in vivo degradation of motor components, potential thermal injury to peritumoral normal tissue, and the inability to distinguish TAM subsets with opposing functions. Future strategies could harness the autonomous movement to generate microfluidic perturbations that loosen the dense ECM and improve photosensitizer penetration into deep tumor regions, while the development of motor materials undergoing disassembly triggered by endogenous enzymes or low pH would facilitate gradual clearance of residual components and reduce long-term retention risk (Table 4).
 |
Table 4 Photodynamic Therapy Nano-Immunotherapy Targeting TAM Reprogramming |
Sonodynamic Therapy
The combination of sonodynamic therapy (SDT) with NDSs enables superior tissue penetration and precise spatiotemporal control, thereby enhancing targeted reprogramming of TAMs. AIE/Biotin/Mannose-M, combined with SDT, reprograms M2 to M1 TAMs via low-dose SDT, generating ROS in macrophages and promoting ATIR in breast cancer.136 G5-CHC-R, combined with SDT, repolarizes TAMs and downregulates MDSCs via R848 (a potent TLR7/8 agonist), ultimately promoting local and systemic cytotoxic T-cell function in pancreatic cancer.137 ICG@C3F8-R848 NBs, combined with SDT, reprogram M2 to M1 TAMs via R848 (a TLR agonist), enhancing ATIR activity in HCC.138 GNP@EXO-AP, combined with SDT, reprograms TAMs in the ultrasound environment to an anti-tumor phenotype via US-induced TAM reprogramming, exhibiting a tumor growth inhibition effect on oral squamous cell carcinoma.139 PATCM, combined with SDT, reprograms M2 to M1 TAMs via TMP195-induced immune responses in TNBC.140 Gal/Ce6@NCG reprograms M2 to M1 TAMs by blocking phosphorylation, activating immunosuppressive Smad2/3 signaling, curbing MDSC differentiation, and promoting tumor immunotherapy in colorectal cancer and liver metastasis.141 Interestingly, Peng et al used dual-targeting aggregation-induced emission polymer micelles to simultaneously target cancer cells and TAMs; Chen et al used nanobubbles to reprogram TAMs; and Tang et al used macrophage exosomes as carriers. Gong et al fabricated a hybrid membrane consisting of a cancer cell membrane and a macrophage membrane to form a nanocarrier system.
Sonodynamic therapy achieves considerably deeper tissue penetration than photodynamic therapy, reaching several centimeters, without ionizing radiation, and enables noninvasive treatment of deep-seated or large-volume tumors. However, the tumor-targeting efficiency of sonosensitizers remains limited, and the long-term safety and in vivo metabolic behavior of sonosensitizers are not yet well understood. The future potential is substantial, the development of smart responsive sonosensitizers combined with spatially precise regulation could establish SDT as a novel physical immunotherapy for modulating TAMs in deep tumors.
Others
Galactan-enriched nanocomposites (Gal-NCs) reprogram M2 to M1 TAM phenotypes by enabling selective and multivalent pattern recognition (TLR4) on macrophage surfaces by activating NF-κB, inhibiting tumor growth, increasing the intratumoral population of cytotoxic T cells, and promoting tumor immunotherapy in colon adenocarcinoma.142 N-M2T-gp100 HBc NVs reprogram M2 cells to M1 TAMs, promoting the activation of Th and CD8+ T cells and reshaping the TME, thus curtailing the growth of both local and distant tumors and triggering long-lasting immune memory protection in melanoma.143 TDN@siCSF-1R, a DNA tetrahedron inspired by ancient mortise and tenon architectural principles, reprograms M2 to M1 TAMs via sustained inhibition of the CSF-1R signaling axis, promoting ATIR in breast cancer.144 FA-D27-tEV-miR-138, which simultaneously targets tumor cells and TAMs, reprograms M2 to M1 TAMs, promoting ATIR in GBM.145 IFN-γ in the BBV reprograms M2 macrophages to M1 macrophages via NF-κB and JAK-STAT signaling, releasing the nitric elements IL-12 and iNOS and promoting ATIR in colon cancer, breast cancer, and HPV.146 FlhDC VNP reprograms M2 to M1 TAMs via the Arts–Heme complex, enabling sustained ROS generation, eliciting mitochondrial stress, and consequently NLRP3 activation, leading to inflammatory immune cell homing and reprogramming of the TME in patients with breast cancer and melanoma.147 CPMV-CD206s+CPMV-CD206 converts M2 macrophages to M1 macrophages via CPMV, a triple TLR agonist (TLRs 2, 4, and 7), inducing ATIR in melanoma.148 Exosomes derived from M1-type macrophages (M1-Exos) reprogram M2 to M1 TAMs via the RNA contained within them, including miR-27a, miR-125b, miR-155, and miR-199a, which exerts significant anti-tumor effects in breast cancer.149
These specifically designed NDSs primarily achieve precise regulation of TAM-mediated phenotype reprogramming by activating ROS-NLRP3 inflammatory bodies, modulating NF-κB signal transduction, and interfering with key molecular mechanisms such as the JAK-STAT pathway. The system developed by Wang et al is based on virus-like particles and the hepatitis B antigen, whereas Zhang et al were inspired by ancient mortise and tenon technology to develop DNA tetrahedra without encapsulation. In addition, Kim et al used a macrophage vesicle in situ to reprogram TAMs, and Xu et al developed nanoparticles that induced local tumor hemorrhage due to A, leading to TAM reprogramming of artesunate and heme to form a complex. Cumulatively, these pioneering studies provide valuable insights and a forward-looking roadmap for the future development of targeted TAM interventions (Table 5).
 |
Table 5 Other Nano-Immunotherapy Targeting TAM Reprogramming |
In summary, reprogramming strategies utilize nanocarriers to deliver therapeutic agents that convert M2 type TAMs into the M1 phenotype, serving as a core cornerstone approach for reversing immunosuppression. The advantage lies in fundamentally restoring the anti-tumor function of TAMs and potentially inducing long term immunological memory. However, current challenges include suboptimal therapeutic efficacy due to TAM heterogeneity, as well as issues related to the stability of the delivery system and its ability to penetrate the ECM.
Influencing TAM Polarization
Promoting M1 and Inhibiting M2 TAMs
NDSs can mediate a dual regulatory effect by potentiating the pro-inflammatory activity of M1 macrophages while concurrently suppressing M2 macrophage function. ZNPs/I@CML reduces M2 TAMs and promotes M1 TAMs by restraining the 3-kinase (PI3K) pathway and subsequently enhancing the immune response in prostate cancer.150 FucSSeDHA (FSSeD)-CFZs reduce M2 TAMs and promote M1 TAMs polarization by downregulating HIF-1α expression and subsequently increasing the mature tumor antigen presentation of DCs to enhance immunogenic cell death-induced immunoreaction, preventing metastasis formation in a comprehensive manner in breast cancer r.151 ACEA@CL(XGMA)-Fe(III) MSs, in combination with photodynamic therapy, increase the number of M1-like TAMs and suppress M2 macrophage expression via activation of ACEA and MSs, leading to remodeling of the TME and suppression of metastatic dissemination in colon cancer.152 R-CM@MSN@BC, with both photodynamic and glutamine-metabolic therapies, and synergistic light therapy reduce the number of immunosuppressive macrophages, particularly M2 and CX3CR1 macrophages, and promote M1 macrophage infiltration via inhibition of glutamine metabolism (the GLS1 inhibitor BPTES), reversing the immunosuppressive cholangiocarcinoma (CCA) TME.153 Interestingly, Deng et al and Zheng et al developed nanocarrier systems integrated with phototherapy, and the system engineered by Zheng et al incorporated glutamine metabolism and demonstrated efficacy in inhibiting immunosuppressive CX3CR1-expressing macrophages However, current studies have largely focused on the glutamine metabolic pathway, while systematic investigations into other key metabolic routes, such as glucose metabolism and amino acid processing, are lacking. Future potential lies in expanding combined interventions that target multiple metabolic pathways, including glycolysis and fatty acid oxidation, thereby constructing a more precise platform for metabolic immunotherapy of TAMs.
Inhibiting M2 TAMs
At present, we mainly inhibit M2 subtype TAMs through SOCS3-STAT6-PPAR-γ signaling to reverse immunosuppression. Man@pSiNP-erastin inhibits M2-type polarization and cause macrophage phenotypic shifting via SOCS3-STAT6-PPAR-γ signaling, increase the expression of PD-L1 in TAMs, and increases the efficacy of anti-PD-L1 therapy against HCC.154 PCL/PTX@DSPE/ET can modulate fatty acid metabolism, blocking M2-like macrophage polarization in the TME, and alleviating immune suppression via etomoxir, blocking the generation of IL-4-mediated M2 TAMs by disrupting intracellular CoA homeostasis, promoting tumor immunotherapy, and suppressing lymphatic metastasis in breast cancer.155 Compared with strategies that both inhibit M2 and promote M1, the advantage of this approach lies in its well-defined mechanism and focused targets. However, this strategy has notable limitations, simply inhibiting M2 macrophages without promoting M1 macrophages may not be sufficient to fully activate anti-tumor immunity. Moreover, long-term inhibition of M2 may interfere with normal tissue repair functions.
Promoting M1 TAMs
Inorganic Carriers
Currently, iron and its derivatives, such as magnetite and magnetically loaded carbon nanoparticles, are primarily used to deliver drugs that promote m1-type macrophage activation. FH-MPLA activates macrophages in an antitumorigenic phenotype at clinically relevant concentrations via monophosphoryl lipid A (MPLA) against TLR4, promoting ATIR in breast cancer.156 Magnetic Sia-IMNPFe triggers the polarization of TAMs to the M1 phenotype via cytokine secretion and ROS generation elicited by its silica–iron matrix, thus promoting ATIR of TAMs in breast cancer.157 CM NPs, which enable photoacoustic (PA)/magnetic resonance (MR) dual-mode imaging, polarize macrophages toward the M1 phenotype prompted by the tetrapeptide tuftsin, eliciting anti-tumor cytokine secretion (eg, TNF-α) and potent anti-Rb immune effects.158 Notably, Zou et al combined photoinduced magnetoresistance imaging and successfully observed and inhibited tumor growth However, nanocarrier systems that use other inorganic materials, such as copper and silicon, as carriers to promote M1 TAMs are lacking. FH-MPLA delivers a translatable TLR4 agonist at clinically feasible doses, effectively polarizing suppressive macrophages toward a tumoricidal state and sensitizing melanoma to monoclonal antibody immunotherapy. Its therapeutic impact as a standalone treatment remains modest, with pronounced tumor regression occurring only when combined with checkpoint blockade, revealing a dependence on complementary immune interventions. Engineering the platform to achieve active tumor targeting and to co-incorporate neoepitopes could convert it into a personalized in situ vaccine capable of eliciting broad-spectrum antitumor immunity.
Organic Carriers
A range of organic carriers, including lipids, nucleotides, carbohydrates, and other biomolecules, can effectively encapsulate and deliver drugs to the M1 phenotype. Bosutinib-HDL increases the population of pro-inflammatory TAMs via a combination of bosutinib-HDL NPs and radiation-promoted tumor immunotherapy in the ATCC.159 GM NPs promote M1 macrophage polarization, thus yielding a synergistic therapeutic response against both primary tumors and metastatic lesions in breast cancer.160 A UIRN molecular targeting drug, a regorafenib-encapsulating nanoparticle constructed with an amphiphilic inulin-ursodeoxycholic acid conjugate, induces the polarization of M1-type TAMs via gut microbiota modulation and the function of regorafenib (REG), a multi-kinase inhibitor, to conquer immunosuppression and enhance ATIR in colorectal cancer (CRC).161 The delivery of LCL-161, R84 and ruxolitinib by carbohydrate nanoparticles (CANDI) induces a distinctive M1-like TAM phenotype with high IL-12 production through the activation of the TLR7/8 signaling pathway and inhibition of the JAK/STAT and NF-κB pathways, after which IL-12 mediates an effective anti-tumor T-cell response, promoting melanoma and colon cancer cell death.162 It is worth noting that Fredrich et al identified a novel TAM subset that exhibits a unique transcriptional profile distinct from that of conventional M1/M2 polarization states and is characterized by exceptionally high IL-12 production The primary advantage of organic carriers lies in their excellent modifiability and relative stability. Despite the inherent advantages of proteins as drug carriers, their systematic development and application as core components in nanocarrier systems remain underexplored.
Carbon Dots
RCD@DOX/BA-O induces the polarization of TAMs to M1 macrophages, further enhancing ATIR in liver and breast cancer.163 This NDS orchestrates a hierarchical delivery cascade of chemotherapeutic and immune-adjuvant payloads, converting tumor-supportive macrophages into antitumor effectors and achieving amplified immune-mediated tumor elimination in liver and breast cancers. The long-term biosafety and metabolic clearance pathways of the carbon-based carrier remain poorly characterized, raising concerns about chronic tissue accumulation. Its theranostic architecture holds clear promise for integrating real-time imaging modules to monitor macrophage repolarization kinetics and adapt dosing schedules for patient-specific immunochemotherapy.
Biological Carriers
Telratolimod@adipocytes promote the polarization of tumor-suppressive M1-phenotype TAMs via TLR7/8 stimulation of type-1 IFN-mediated immune responses, thereby suppressing tumor growth in patients with melanoma.164 This living adipocyte carrier system leverages innate macrophage-recruiting capacity to drive sustained proinflammatory reprogramming via TLR7/8-mediated type I interferon signaling, generating potent antitumor immunity against melanoma. The requirement for autologous adipocyte isolation introduces substantial donor-dependent variability and a complex manufacturing workflow that complicates standardization and scale-up. Engineering these cellular vehicles to co-deliver complementary stimulatory signals could yield a modular personalized immunotherapy platform effective across a wider range of solid tumors.
Photothermal Therapy (PTT)
When combined with photothermal therapy (PTT), NDSs can more effectively promote M1 polarization of TAMs by inducing localized ATIR, thereby reshaping the immunosuppressive TME. M@P@HA polarizes TAMs into anti-tumor M1-type macrophages by activating cGAS STING signaling to further activate innate immunity in breast cancer.165 DPTT-Mn Lipo NPs promoted M1 polarization of TAMs via PTT-mediated ICD and transactivation of the cGAS-STING pathway, leading to efficient tumor suppression, induction of tumor clearance, and establishment of durable immunological memory in breast cancer.166 M1mDDTF reprograms M2 to M1 TAMs, subsequently stimulating ICD and enhancing immunotherapy in bone tumors.167 Among these varied approaches, the cGAS-STING signaling pathway represents one of the mechanisms through which the M1 TAM phenotype is facilitated. Interestingly, the drug delivery system developed by Wang et al uses M1 macrophage membrane camouflage to improve targeting and treatment. This synergistic strategy provides a remotely and precisely triggered photothermal effect that overcomes resistance to conventional immunotherapy by coupling physical hyperthermia with innate immune activation. Insufficient light penetration depth, however, restricts its efficacy against deep-seated tumors. Future advances in more sensitive photothermal materials and integrated ECM penetration strategies could enable precise and controllable synergistic photothermal immune therapy for deep tumors.
Engineered Macrophages
Engineered macrophages can rapidly and precisely target TAMs by exploiting their native chemotactic pathways and surface-displayed targeting ligands. MINS@M Φ (MNC-ICG-NIG@SiO2 with MINS-loaded macrophages) promotes M1 polarization of macrophages via the release of Fe2+ and CpG caused by self-pyroptosis of engineered macrophages, resulting in the release of anti-tumor cytokines (IL-2, IL-6, and IFN-γ), enhancing its efficacy against bladder cancer.168 FUSION (ZGSO@MSN@R848 encapsulated by dipalmitoylphosphatidylcholine [DPPC]) achieves closed-loop macrophage engineering via a TLR7/8 agonist (R848), promoting ATIR in breast cancer.169 Interestingly, the NDS studied by Xu et al focused on the ultrasound response and combined on-demand activation with feedback to form a closed loop. Engineered macrophages exploit innate tumor-homing chemotactic pathways to achieve precise delivery of immunostimulatory payloads, with controlled self-pyroptosis enabling on-demand release of metal ions and nucleic acid adjuvants that reshape the intratumoral cytokine milieu against bladder cancer. Premature or asynchronous pyroptotic death during circulation risks off-target payload release and systemic inflammatory complications before these cellular carriers reach the tumor bed. Incorporating external trigger-responsive genetic circuits into these living delivery platforms could enable spatiotemporally tunable pyroptosis, substantially elevating their safety margin and therapeutic precision across multiple cancer types.
Special Phenotypes
CANDI400 polarizes TAMs toward CXCL9 (an anti-tumor phenotype) via RBN2397-tank binding kinase 1-IRF3, STING-I IFN, and TLR7/8-NFκB, promoting ATIR in colon cancer170 (Table 6). The novel TAMs identified in his study provide a new target for TAM-targeted nano immunotherapy and offer researchers a fresh perspective. In the future, overcoming immunosuppressive resistance in solid tumors and advancing personalized precision immunotherapy may become important breakthroughs.
 |
Table 6 Nano-Immunotherapy of Promoting M1 and Inhibiting M2 TAMs |
Polarization modulation strategies intervene in microenvironmental signals to guide TAM differentiation toward the M1 phenotype, thereby preventing M2 polarization at the source. These strategies are suitable for early-stage tumors or postoperative adjuvant therapy and can synergize with immune checkpoint blockade. Compared with reprogramming approaches, polarization modulation has a slower onset of action but fewer side effects and requires sustained release to maintain the direction of polarization. In addition, the ability to penetrate the ECM remains a challenge.
Regulating TAM-Secreted Mediators
Inorganic Carriers
By acting through macrophages, SPION-CCPMs elicit the production of RNS and cytokines (eg, IL-1β and TNFα), which initiate a multifaceted mechanism against lung cancer cells, leading to functional impairment and cell death, which is suppressed by the inhibitors QVD and 3MA.171 MTCZC+cGAMP-CPS@Gel activates the STING pathway (STING-TBK1-IRF3) and promotes the release of proinflammatory cytokines (IFN-I), triggering an IFN-I-driven inflammatory program that reverses the TME and enhances ATIR in TNBC.172 The advantage of this strategy lies in directly harnessing the natural secretory function of TAMs, thereby avoiding the complexity of drug loading, while inorganic carriers can integrate imaging or magnetothermal functions. However, the risk of excessive cytokine activation and the long-term toxicity of inorganic materials remain challenges. Future potential lies in designing tumor microenvironment-responsive inorganic carriers that enable spatiotemporally controlled release of cytokines, thereby converting secretory modulation into a precise local immune activation strategy.
Organic Carriers
Organic carriers are readily internalized by TAMs through membrane fusion or receptor-mediated endocytosis, thereby enabling targeted regulation of cytokine secretion profiles. LPS-PEC (DOX@LPS-PEC:Sirtinol 1:5 w/w) decreases TAM-secreted IL6 levels by targeting the TLR4/SIRT2 axis and suppressing the metastatic capability of TNBC.173 ZnCDA increases systemic levels of IFN-β, TNF-α, IFN-γ, IL-6 and CCL-2 through STING activation through the phosphorylation of IRF3, potentiating the efficacy of radiotherapy and immune checkpoint blockade in immunosuppressive pancreatic cancer and glioma.174 IL-13-LCL-SIM targets the proangiogenic functions of TAMs via IL-13, suppressing key mediators (VEGF, leptin, FasL) and increasing the susceptibility of the melanoma TME to PEG-EV-DOX, ultimately triggering Bax-dependent apoptosis and oxidative stress in a potent anti-tumor response.175 MAPDCs (CPT+R848) reprogram GAMs from M2 to M1 via R848, increasing the secretion of proinflammatory cytokines (IP-10, IL-12 and IL-6), amplifying ATIR and restoring chemosensitivity, and yielding superior anti-GBM outcomes.69 SPADS, which combines α-tocopherol (Tpl) and toyocamycin (Tcc), results in the distinct cytokine profiles of TAMs (increases in TNF-α, IL-12, and Arg1 levels and decreases in TGF-β levels) via coinhibition of ER activity and oxidative stress (STAT1/STAT3), potentiating their phagocytic function and tumoricidal activity, which collectively curb breast oncogenesis.70 GPCuD NPs (Cu+DOX) increase the secretion of IL-6 and TNF-α by macrophages, promoting ATIR in breast cancer.73 CaNP@cAD-PEG composed of cAD loaded on CaNP enveloped with DSPE-PEG2000 promoted IL-1β and IFN-γ secretion through the simultaneous activation of multiple inflammation-related signaling pathways (MAPK signaling pathway and NF-κB signaling pathway), as well as the NLRP3 inflammasome pathway, in melanoma and rectal cancer.80 M.RGD@Cr-CTS-siYTHDF1 NPs suppressed immunosuppressive IL-10 production, increased the expression of immunostimulatory factors (IL-12 and IFN-γ), improved the TME and inhibited tumor progression in patients with HCC.116 MG5-S-IMDQ converts M2 TAMs into M1 TAM phonotypes via a potent agonist of TLR7/8, imidazoquinoline (IMDQ), subsequently increasing their phagocytic activity toward tumor cells, enhancing their cytotoxic effects, improving tumor antigen cross-presentation ability and inducing macrophages to secrete TNF-α, IL-12p70 and IL-6 to combat GBM.52 Interestingly, by employing an innovative “double-punch” approach, Mahmoud et al designed a combinatorial nanoformulation that concurrently targeted both TAMs and tumor cells, effectively overcoming compensatory resistance mechanisms while achieving comprehensive tumor suppression through coordinated immune activation and direct cytotoxicity. However, nanocarrier drug delivery systems with other organic substances, such as sugars or other natural substances, are still lacking.
Biological Carriers
Biological carriers, such as enzymes, cancer cell membranes, macrophages, exosomes, and bacteria, can deliver drugs that precisely target TAMs owing to their biocompatibility and subsequently regulate TAM secretion. Abs-CpG polarizes M1-type macrophages and, via CpG (TLR9 agonist), produces a large amount of TNF-α, which then activates cell division control protein 42 (Cdc42), enhancing the phagocytosis of adjacent macrophages and promoting tumor immunotherapy in breast cancer.63 CpG-EXO/TGM induce TAMs, releasing anti-tumor cytokines (TNF-α, IL-6, and IL-12) in the process, thereby promoting ATIR in GBM.90 CW-camouflaged nanospores increase the expression of IFN-β, promoting ATIR in melanoma and breast cancer.124 PR-M can regulate the secretion of cytokines (IL-12, IFN-γ, TNF-α and IL-10), increasing ATIR levels in colorectal carcinoma.65 Mam@NPs could increase CCL3 secretion through the combination of IM and aCD47, which further secreted inflammatory and anti-tumor cytokines, promoting ATIR in gliomas.89 MINS@M Φ (MNC-ICG-NIG@SiO2 with MINS-loaded macrophages) promoted the release of anti-tumor cytokines, including IL-2, IL-6 and IFN-γ, enhancing its efficacy against bladder cancer.168 Some researchers use cancer cells as carriers, and some use fungi, which can better target and phagocytose TAMs. Interestingly, Zhang et al, Guo et al, and Long Zhang used engineered macrophages to regulate mediators of TAM secretion
Compared with organic or inorganic carriers, biological carriers can mimic natural intercellular communication, evade immune clearance, and actively bind to tumor sites However, this strategy still faces several challenges, including complex preparation processes, off-target binding, and potential tumorigenic risks In the future, combining gene editing technologies to modify these carriers, for example, by knocking out genes associated with immune rejection, could transform secretion modulation into a precise and safe local immunotherapy strategy.
Others
Some carriers that combine magnetic thermal therapy, deliver multiple drugs, or use a motor that can regulate TAM secretion in a unique way. M-PUNCs promote intratumoral IFN-β secretion by activating STING signaling in M1 TAMs and then recruiting tumor-infiltrating CD8α/CD103 DCs to prime CTLs, promoting tumor immunotherapy in breast cancer and melanoma.100 IC@MSA JNM facilitates the reprogramming of M2 to M1 macrophages via ibrutinib (IBR), the production of pro-inflammatory cytokines and the recruitment of more T cells to kill tumor cells, ultimately inhibiting breast cancer growth.134 LPZ/DOX@M/R837@MPs could regulate cytokine secretion (increase in IL-12 and decrease in IL-10 levels) and promote tumor immunotherapy in breast cancer, melanoma, and colon tumors.98
Strategies that modulate TAM-secreted mediators employ nanocarriers to deliver agents such as antibodies, antisense oligonucleotides, or small molecules, thereby influencing cytokine release from TAMs. Their importance lies in the relatively well-defined targets and the potential for synergy with anti-PD-1 therapy. Compared with comprehensive reprogramming, modulating secreted mediators is more precise and results in lower systemic toxicity. However, the complex cytokine network means that blocking a single mediator may easily induce compensatory upregulation, necessitating multi-target co-delivery systems.
Inducing TAM Exhaustion
Inorganic Carriers
siRNAaRGODox, combined with phototherapy, causes M2 TAM obliteration via site-specific shuttling of M2 TAMs via SR-BI receptor recognition and apolipoprotein A-I crown binding with overexpressed scavenger receptors, blocking the TGF-β-Snail pathway and reversing the expression of EMT-related proteins to rebuild the anti-metastatic niche in breast cancer.176 Man@pSiNPs-erastin causes the activation of intracellular ferroptosis via GPX4/RRM2 signaling regulation, promoting ATIR in liver, lung, colon, and breast cancers.154 Although metal-based carriers exhibit considerable potential in biomedical applications, the current research landscape remains limited. The long-term in vivo accumulation toxicity of metal-based materials remains unclear, and their biodegradability is poor. Future potential lies in developing biodegradable metal organic frameworks or hybrid materials combined with tumor microenvironment responsive release, thereby achieving a precise and safe strategy for TAM depletion.
Organic Carriers
A CSP-cationic folate nanoconjugate delivery system (glucose-based carbon nanospheres) targets and depletes SR-expressing TAMs via the cationic lipid-conjugated neuropsychotic drug haloperidol (H8), promoting ATIR in gliomas and GBM.177 PTX-SAL depletes TAMs via endocytosis mediated by SA receptors such as Siglec-1, promoting ATIR in sarcoma.178 SA-CH modified liposomal ANB (ANB-SL) targets and eradicates TAMs via Siglec-1, effectively terminating TAM-driven angiogenesis and tumor progression to promote vascular normalization, activate ATIR, and reverse the immunosuppressive TME in sarcoma.179 CML, a click chemistry-based liposome nanoplatform, causes M2 macrophage exhaustion via clodronate-mediated apoptosis, promoting ATIR in breast cancer.180 M-H@lip-ZA, in combination with SDT, depletes M2 TAMs via a combination of HMME-mediated SDT and ZA, thereby restraining tumor growth, inducing vascular normalization to improve perfusion, and alleviating hypoxia to enhance ATIR in breast cancer.181 Man-LP@ZOL/DOX depletes M2 TAMs via ZOL-promoting tumor immunotherapy in TNBC.182 THP-PSA NPs effectively eliminate TAMs via a scavenger receptor-A (SR-A) mediated endocytosis pathway, and lipid-raft/caveolae and clathrin-mediated endocytosis pathways, promoting tumor immunotherapy in breast cancer.183 A-PER-p(TMZ)29/CLo causes TAM exhaustion via clodronate (Clo), which inhibits tumor growth and markedly extends the survival of brain astroblastoma and glioma.184 IMNP (nCS(ALN + Bin)-Man) synergistically depletes TAMs via interactions between mannose and MMR on the surface of TAMs and ALN released intracellularly, leading to the apoptosis of TAMs, and promoting tumor immunotherapy in melanoma and breast cancer.185 Compared with metal-based or biological carriers, organic carriers offer better biocompatibility and ease of surface modification; however, this strategy is limited by the poor subtype selectivity of drugs such as clodronate, which may also deplete normal tissue macrophages, and by the potential immunodeficiency or infection risks from long-term repeated administration. In addition, the in vivo stability and burst drug release of organic carriers have not been fully resolved. Future potential lies in developing prodrug strategies (as exemplified by Zhang et al), combining physical therapies, and designing smart responsive release systems, thereby reducing systemic toxicity and enhancing anti-tumor immune efficacy
Biological Carriers
PROTAC (CRV-LLC membrane/DS-PLGA/dBET6) depletes M2 TAMs via BRD4, promoting tumor immunotherapy in lung cancer186 DCC@M2 eliminates M2-type TAMs and disrupts their crosstalk with malignant cells by amplifying ROS-induced immunogenic cell death and blocking CSF-1, thereby increasing ATIR in breast cancer.187 Achieving selective depletion without harming M1-type or homeostatic macrophages remains challenging due to shared surface marker expression across myeloid lineages. Incorporating logic-gated dual-recognition modules could refine target cell discrimination and extend this degradation-based strategy to other malignancies driven by tumor-supportive myeloid compartments.
Photothermal Therapy
AuNH-2-Ab combined with phototherapy attenuates TAMs by blocking the immune checkpoint PD-L1 through the heat-induced release of anti-PD-L1, eliminating local and metastatic HCC tumors.188 DS-Ce6, combined with phototherapy, induces apoptosis of M2-like macrophages via uptake by specific binding to SR-A and generating cytotoxic O2 under laser irradiation, inducing effective tumor cell death and regression of breast cancer.189 CBE combined with phototherapy targets M2 TAMs and eliminates them via photodynamics and PTT in the presence of laser irradiation, promoting ATIR in LLC cells.190 Upon light exposure, the mUNO-EB-ICG-Fc@Alb formulation mediated the full removal of M2 TAMs. Light-triggered production of reactive oxygen species directly targets these cells. Simultaneously, an Fc-catalyzed chemical reaction is initiated, which not only generates highly destructive hydroxyl radicals but also alleviates low-oxygen conditions by producing oxygen gas, thereby increasing the effectiveness of phototherapeutic interventions for treating breast cancer.191
Nanocarrier systems using biological carriers, or in combination with phototherapy, effectively deplete TAMs by exploiting the favorable biocompatibility of biological carriers or by combining them with photothermal therapy to achieve improved targeting and therapeutic efficacy. However, challenges in targeting specificity remain, as these agents are still partially distributed to the liver and spleen after systemic administration, posing a risk of immunodeficiency, and their ability to penetrate the ECM must be improved to achieve high therapeutic efficacy. Compared with reprogramming strategies, depletion is more thorough but irreversible. Future efforts should aim for selective depletion that avoids the elimination of normal tissue macrophages (Table 7).
 |
Table 7 Nano-Immunotherapy Targeting of TAM Exhaustion |
Enhancing TAM Phagocytosis
Organic Carriers
Organic carriers, including lipids, nucleotides, carbohydrates, and other biomaterials, can effectively deliver agents that specifically target the phagocytic function of TAMs by leveraging their inherent biocompatibility and biomimetic properties to facilitate recognition and internalization. M@SINPs, combined with photothermal effects, restore M1 macrophage phagocytic activity by inhibiting SHP2 expression, blocking the CD47-SIRPa pathway and significantly inhibiting tumor growth in colon cancer.108 cGAMP@CaP@Abs, dual-target liposomes, enhances macrophage phagocytosis and promotes cross presentation of tumor antigen via blocking the CD47-SIRPα axis and activating the STING pathway, respectively, resulting in tumor growth inhibition in breast cancer and melanoma.192 cGAMP@CaP@Abs reinstate phagocytic clearance of tumor cells by disrupting CD47-SIRPα interactions while simultaneously triggering STING-dependent antigen cross-presentation, driving potent antitumor immunity in both breast cancer and melanoma. The dependence on intratumoral injection and the risk of systemic STING-mediated inflammation constrain its translational safety and practicality for tumors that are difficult to access. Derivatizing the platform for systemic administration with tumor-restricted STING activation could extend its therapeutic benefit to metastatic lesions and a wide range of solid malignancies.
Biological Carriers
Biological carriers can deliver drugs that precisely target TAMs owing to their biocompatibility and subsequently promote the phagocytic function of these cells. ICG/CMSN@GsE enhances the phagocytic capacity of TAMs by entering the nucleus through a variety of membrane receptors, such as MyD88, TLR2/4, TNFR 1, IL-17R, CD40, and RAGE, thereby upregulating the expression of upstream mediators (MyD88 and NF-κB) and enhancing cancer immunotherapy efficacy in breast cancer.122 The subsequent four nanocarrier systems uniformly employ engineered bacterial OMVs as their foundation platform, capitalizing on the inherent immunostimulatory properties and biocompatible nanostructure of the vesicles to regulate the phagocytosis of TAMs. OMV-SIRP α@CaP/GM-CSF induces Fc-SIRPα-mediated phagocytosis to present a tumor antigen to activate T cells, promoting ATIR in adenocarcinoma of the colon and melanoma.193 OMV-CD47/GPC3 promotes the phagocytosis of tumor cells by macrophages by blocking CD47/SIRPα, thus providing a promising strategy for HCC immunotherapy.92 CR/si@ab-OMV/PTX NPs increase macrophage phagocytosis of tumors via TAMs repolarizing to the M1 type by delivering siRNAs to silence Arg-1 and blocking the “do not eat me” signal with antibodies, subsequently activating ATIR in TNBC.194 PEG/Se@OMV-CD47nb reprogrammed the immunosuppressive TME, driving macrophages toward a proinflammatory state and neutralizing the “do not eat me” signal, thus inducing T-cell-mediated ATIR to promote tumor immunotherapy in colon cancer and melanocytic tumors.195 The following two NDSs are based on engineered macrophages as carriers, leveraging their innate tumor-homing capabilities and phagocytic functions to achieve precise spatial delivery of therapeutic agents and facilitate profound remodeling of the TME. SIRP α-M@nanoPB restores macrophage phagocytosis and M1 polarization by blocking the CD47–SIRPα checkpoint, triggering potent immune responses in osteosarcoma.196 PMMNPs increase macrophage-mediated cancer cell phagocytosis and antigen presentation by blocking CD47 and PD-L1 expression, promoting anti-tumor synergy between T cells and macrophages in colon cancer.197 Interestingly, Gu et al used ginseng exosomes as carriers, whereas Li et al used multimodule nanocomplexes to reconcile the competitive cooperative patterns between malignant and immune cells and activate ATIR in TNBC models Ji et al used engineered macrophages to enhance the phagocytic function of TAMs, and the system developed by Moon et al promoted anti-tumor synergy between T cells and TAMs.
The biological carriers described above, including plant-derived exosomes, bacterial outer membrane vesicles, and engineered macrophages, exploit their natural biocompatibility and immunomodulatory properties to increase the ability of TAMs to phagocytose tumor cells. Compared with organic or inorganic carriers, biological carriers offer the advantage of mimicking pathogen-associated molecular patterns to activate innate immunity while also utilizing homologous targeting and tumor-homing capabilities for precise delivery. However, the preparation processes for outer membrane vesicles and engineered macrophages are complex, and the release of large amounts of inflammatory mediators following enhanced phagocytosis carries the risk of triggering a cytokine storm, and most studies have not adequately evaluated long-term safety.
Others
ChiP-RS (SHP099+R848) enhances macrophage phagocytosis via R848 by activating TLR7/8 and promoting tumor immunotherapy in mammary carcinoma.96 By sensitizing tumors to αCD47 checkpoint inhibition, a supramolecular hydrogel (PF) facilitates macrophage-mediated phagocytosis, thereby triggering a potent anti-GBM immune response.198 FVIO, incorporated with MHT, synergizes to increase macrophage phagocytosis by interfering with the CD47-SIRPα checkpoint, resulting in a robust ATIR response against HCC.102
Enhanced phagocytosis strategies directly mobilize innate immunity and synergize with antibody-dependent cellular phagocytosis, resulting in few side effects. However, challenges include off-target effects and insufficient penetration through the tumor extracellular matrix. Future efforts should focus on site-specific penetration of the tumor ECM to achieve precise and efficient therapeutic outcomes.
Modulating TAM Metabolism
We targeted TAM ferroptosis, NO metabolism, glucose metabolism, gene expression regulation, and antigen cross-presentation restoration to regulate TAM metabolism. CQ@MIL-GOX@PB perturbs the metabolism of neoplastic cells and TAMs via the release of a CQ autophagy inhibitor (chloroquine), which can drive the NO anabolism of TAMs to further sensitize them to CDT in breast cancer.199 M-s/W-NP blocks the glycolysis of TAMs by suppressing GLUT1 through inhibition of PI3K expression to increase the intratumor glucose level for sustained effector functions of anti-tumor immune cells in pancreatic tumors.200 M/LM-Lipo causes mannose to suppress glycolysis in TAMs by recruiting lysosomes to degrade M6P isomerase enzyme (MPI), thus achieving a prominent abscopal effect in metastatic lesions by potentiating ATIR in breast cancer.201 H-M@siS15/Cis-Asp8 reverses tumor immunosuppression and inhibits osteoclast differentiation via downregulation of Siglec15 expression in TAMs among lung adenocarcinoma cells.202 Cy5-D-PEG4-SPDP has a gene knockdown effect on TAMs and achieves a gene silencing effect on GFP-expressing tumors in glioma.203 Man@pSiNP-erastin activates intracellular ferroptosis via GPX4/RRM2 signaling regulation, promoting ATIR in liver, lung, colon and breast cancers.154 ME@C combined with anti–PD-L1 antibodies reshapes antigen cross-presentation by M2 TAMs via CD302 receptor-mediated endocytosis targeting, releasing E64 to inhibit the activity of cysteine proteases and then priming the clonal expansion of tumor-antigen-specific CD8⁺ T cells in breast cancer and melanoma.204 The aforementioned carriers achieve TAM metabolic modulation through various mechanisms, including the induction of the GPX4/RRM2 signaling pathway. Interestingly, the NDS studied by Gao et al targets a variety of metabolites, and the study by Xiao et al not only regulates glucose metabolism but also regulates the extracellular matrix. Currently, research on nanomaterial systems targeting other metabolic modes, such as lipid and protein metabolism, is limited, and much room for development remains.
This metabolic modulation strategy precisely targets TAM-specific metabolic vulnerabilities while sparing normal macrophages and offers greater sustainability compared with depletion or reprogramming approaches. Tumor metabolic plasticity frequently activates compensatory pathways that demand co-delivery of multiple inhibitors, and clinical translation is further constrained by the lack of long-term safety data and the limited ECM penetration of delivery vehicles. Future advances in ECM-penetrating and safe delivery systems combined with multi-inhibitor co-delivery could unlock the sustainable and selective metabolic reprogramming of TAMs.
Inhibiting TAM Recruitment
Nanocarrier systems can effectively inhibit tumor progression by precisely regulating TAM recruitment, which subsequently disrupts the establishment of an immunosuppressive TME. Telratolimod@adipocytes increased the recruitment of TAMs and suppressed tumor growth in a melanoma model.164 GENs induce the recruitment of M1 TAMs to the TME via GENs, promoting ATIR in glioma.205 NP-BUP suppresses norepinephrine (NE) release from adrenergic cells, thereby curtailing NE-driven macrophage infiltration via neuroimmune signaling and enhancing ATIR in TNBC.206 Interestingly, Kim et al used ginseng-derived exosome-like nanoparticles to inhibit tumor progression by precisely regulating the recruitment of TAMs Wu et al suppressed noradrenergic nerves in the TME to diminish the infiltration of TAMs targeting TAMs via neuroimmune signaling, which represents a novel therapeutic avenue.
The strategy of inhibiting TAM recruitment holds significant value for postoperative recurrence prevention and early intervention. Compared with reprogramming or depletion approaches, inhibition of recruitment results in fewer side effects. However, it is ineffective against TAMs already present within the tumor and therefore often needs to be combined with depletion or polarization modulation strategies.
Blocking PD-L1 Expression on TAMs
Nanocarriers can effectively deliver specialized therapeutic agents to precisely regulate PD-L1 expression on TAMs, thereby reversing PD-L1-mediated immunosuppression. Man@pSiNPs-erastin increased PD-L1 expression in macrophages and increased the ATIR in anti-PD-L1 therapy.154 AuNH-2-Ab alleviated PD-L1-expressing TAMs by blocking the immune checkpoint PD-L1 through the heat-induced release of anti-PD-L1, eliminating local and metastatic HCC tumors.188 DFMO+N-aP@ICG, synergistic photothermal therapy, repolarizes TAMs by inhibiting the expression of aconitate decarboxylase 1 (ACOD1) and releasing aPD-L1, which binds to PD-L1 on TAMs, reverses immune suppression at the tumor site, and inhibits the proliferation of glioma cells.121 Interestingly, Tang et al used porous silicon nanoparticles modified with mannose, whereas Zhang et al used microglial cell vesicles as carriers (Table 8).
 |
Table 8 Nano-Immunotherapy of Regulating PD-L1 Expression on TAMs |
The strategy of blocking PD-L1 expression on TAMs is critical because PD-L1 directly targets immune checkpoints. However, the ability to penetrate the ECM and the stability of the delivery system remain significant challenges. Compared with reprogramming, PD-L1 blockade is more specifically focused on adaptive immunosuppression. Nevertheless, TAMs also evade immunity through other molecules, such as CD47; therefore, using this strategy alone has limited efficacy, necessitating combination with phagocytosis enhancement or reprogramming approaches to achieve comprehensive remodeling.
Relevant Receptors and Pathways
Reprogramming TAMs
Related receptors include CSF-1R, TLR3, TLR4/9, CD47, Siglec, TLR2/4, TNFR 1, IL-17R, CD40, and RAGE. Receptors on the membranes of intracellular vesicles include TLR7/8. In HCC, DL@NP-M-M2pep reprograms TAMs from the M2 to the M1 phenotype via the PI3K/Akt and NF-κB pathways. In ovarian cancer, PLGA-CpG@ID8-M reprograms TAMs from the M2 to the M1 phenotype via the Gbp2-Pin1-NF-κB pathway. In breast cancer, PLGA(H)-DOX@M/R837 and RB-OL@M reprogram TAMs from M2 to the M1 phenotype via the TLR7-NF-κB pathway. In breast cancer, MNCRs reprogram TAMs from the M2 to the M1 phenotype via the TLR7/8-NLRP3 and MAPK-NLRP3 pathways. In melanoma and rectal cancer, CaNP@cAD-PEG reprograms M2 TAMs to M1 TAMs via the NLRP3-inflammasome pathway and the AMPK, and NF-κB pathways. In GC, CCM@HMON@CS-Pt(IV)@ANX5 reprograms TAMs from the M2 to the M1 phenotype via the TLR9/MyD88/NF-κB signaling pathway. In breast cancer, ICG/CMSN@GsE reprograms M2 TAMs to the M1 phenotype via the (TLR2/4, TNFR 1, IL-17R, CD40, RAGE)-MyD88-NF-κB pathway. In melanoma and breast cancer, PC-CW reprograms M2 TAMs to M1 TAMs via the STING pathway. In breast cancer, IC@MSA JNM reprograms M2 TAMs to M1 TAMs via the Siglec-1-BTK pathway. In colon cancer, breast cancer, and HPV, IFN-γ BBV reprograms M2 TAMs to M1 TAMs via the NF-κB and JAK-STAT signaling pathways. In breast cancer and melanoma, FlhDC VNP reprograms M2 TAMs to M1 TAMs via the ROS-NLRP3 pathway. In lung cancer, ZCMF@PEG-AF reprograms M2 TAMs to M1 TAMs via the IKKβ/IκBα/NF-κB pathway. In HCC, M.RGD@Cr-CTS-siYTHDF1 NPs reprogram M2 TAMs to M1 TAMs via the siYTHDF1-M6A-STAT3 pathway (Figure 3).
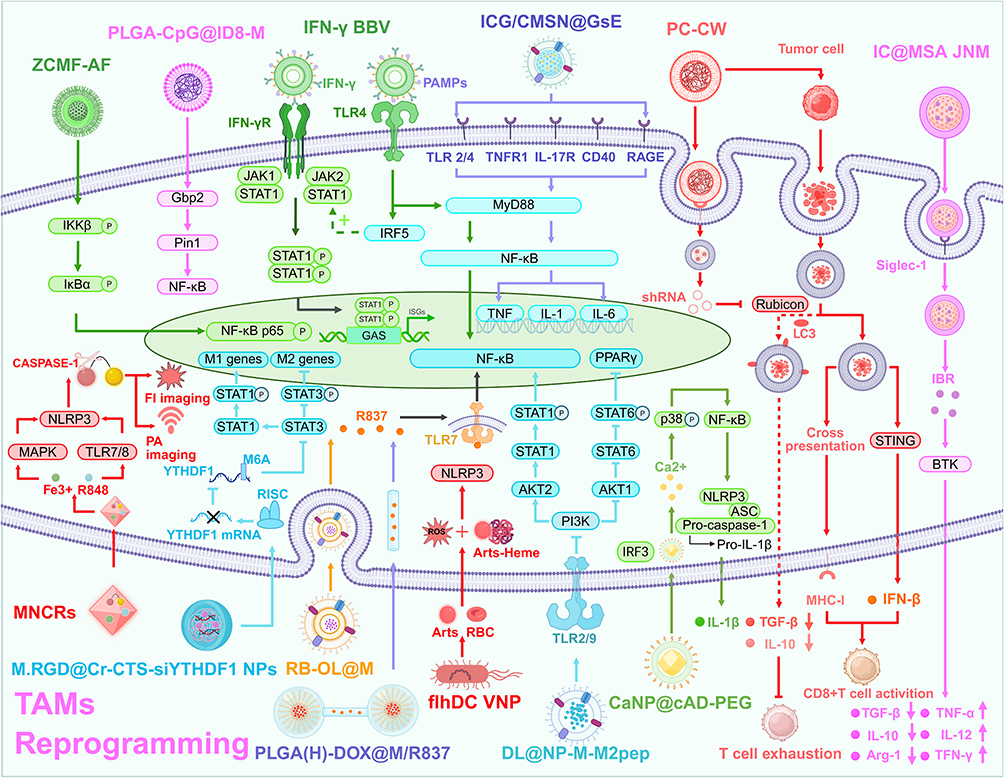 |
Figure 3 Nano-immunotherapy targeting reprogramming TAMs. TAMs can be reprogrammed from M2 towards M1 phenotype via PI3K/Akt, NF-κB, Gbp2-Pin1-NF-κB, TLR7-NF-κB, TLR7/8-NLRP3, MAPK-NLRP3, NLRP3-inflammasome pathway, AMPK NF-κB, TLR9/MyD88/NF-κB, (TLR2/4, TNFR 1, IL-17R, CD40, RAGE)-MyD88-NF-κB pathway, STING pathway, Siglec-1-BTK, NF-κB JAK-STAT, ROS-NLRP3, IKKβ/IκBα/NF-κB and siYTHDF1-M6A-STAT3 pathway. “↑” indicates upregulation, while “↓” indicates downregulation. “+” indicates positive feedback regulation. The figure was drawn using Biorender. |
Influencing TAM Polarization
The relevant receptors include TLR7/8, TLR4, EGFR, and CB1. Receptors on the membranes of intracellular vesicles include TLR7/8. We mainly promote the M1 TAM phenotype and inhibit the M2 TAM phenotype through the TLR7/8-TAK1-p38-NF-ΚB, NF-κB-CD40-clAP/TRAF-NIK-Nc-NF-κB, IL-10-JAK1/Tyk2-STAT3, cGAS-STING, SOCS3-STAT6-PPAR-γ signaling and RelA/NF-κB/CSF-1 pathway (Figure 4).
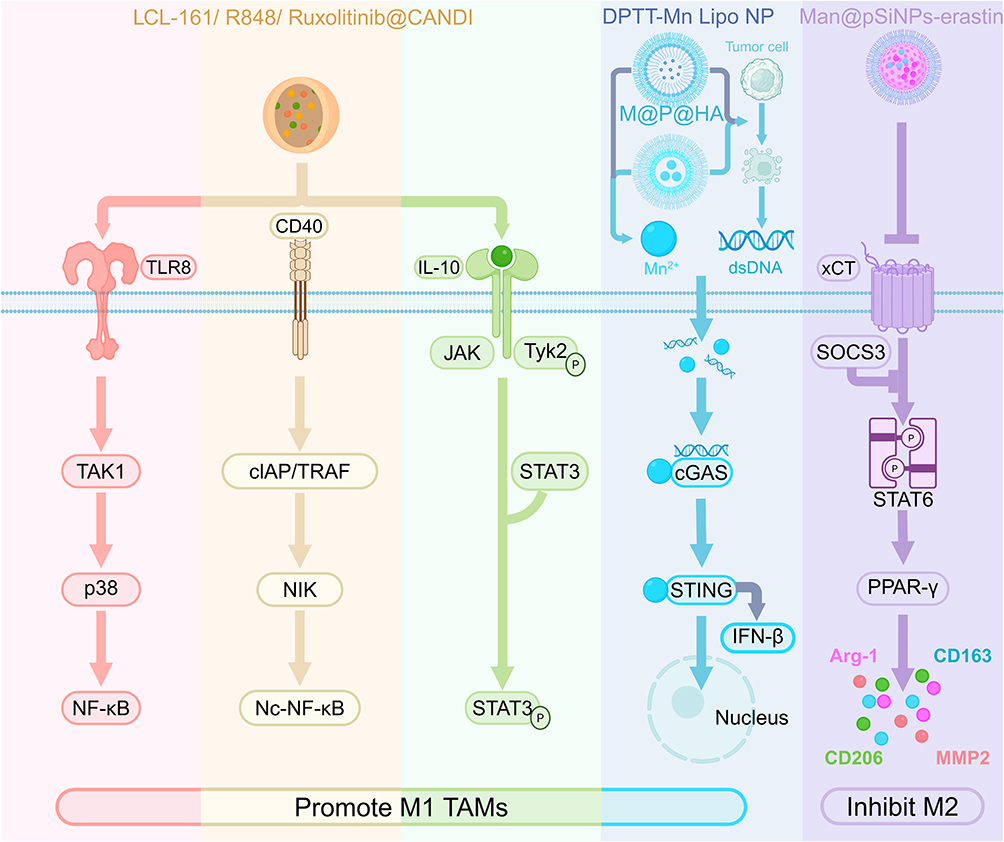 |
Figure 4 Nano-immunotherapy targeting promoting M1 and inhibiting M2 phenotype. Promoting the M1 phenotype of TAMs and inhibiting the M2 phenotype of TAMs could through TLR7/8-TAK1-p38-NF-κB, NF-κB-CD40-clAP/TRAF-NIK-Nc-NF-κB, IL-10-JAK1/Tyk2-STAT3, cGAS-STING, SOCS3-STAT6-PPAR-γ signaling and RelA/NF-κB/CSF-1 pathway. The figure was drawn using Biorender. |
Regulating TAM-Secreted Mediators
The relevant receptors include TLR4, TLR9, CSF-1R, SR-A, and IL-13R. Receptors on the membranes of intracellular vesicles include TLR7/8. In TNBC, MTCZC + cGAMP-CPS@Gel regulated the expression of TAM-secreted mediators via the STING-TBK1-IRF3 pathway. In TNBC, LPS-PEC (DOX@LPS-PEC:Sirtinol 1:5 w/w) regulates TAM-secreted mediators via the TLR4/SIRT2 axis. In melanoma and rectal cancer, CaNP@cAD-PEG regulates TAM-secreted mediators via the MAPK, NF-κB, and NLRP3 inflammasome pathways. In melanoma and breast cancer, CW-camouflaged nanopores regulate TAM-secreted mediators by blocking the Rubicon-LAP efferocytosis axis. In breast cancer and melanoma, m-PUNCs regulate TAM-secreted mediators via the STING pathway. In breast cancer, melanoma, and colonic tumors, LPZ/DOX@M/R837@MPs regulated the secretion of mediators by TAMs via the intracellular NF-κB signaling pathway. Additionally, TAMs primarily secrete the following mediators: Arg-1, CCL2, CCL3, CXCL9, IFN-β, IFN-γ, IFN-I, TNF-α, TGF-β, IL-6, IL-10, IL-12, IL-1β, VEGF, leptin, FasL, and RNS (Figure 5).
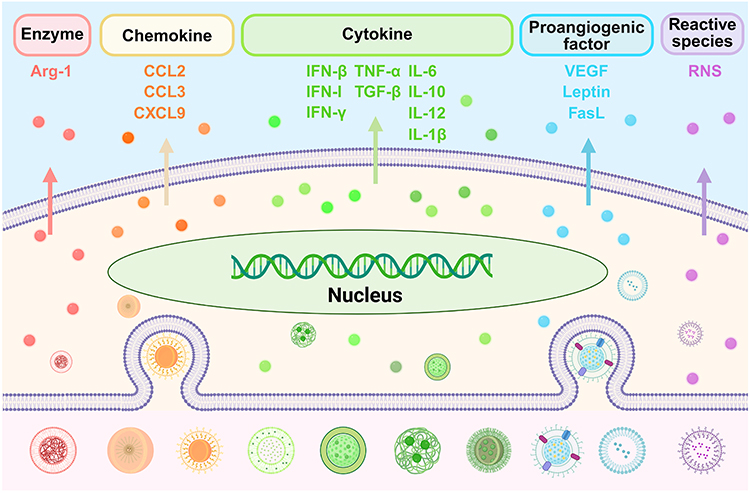 |
Figure 5 Mediator molecules of TAMs. TAMs primarily secrete the following mediators: Arg-1, CCL2, CCL3, CXCL9, IFN-β, IFN-γ, IFN-I, TNF-α, TGF-β, IL-6, IL-10, IL-12, IL-1β, VEGF, leptin, FasL, RNS. The figure was drawn using Biorender. |
Inducing TAM Exhaustion
The relevant receptors include Siglec-1, SR-A, CSF-1, PD-L1, MMR, and scavengers. In breast cancer, siRNAaRGODox promotes exhaustion via the TGF-β-Snail pathway. In liver, lung, colon, and breast cancers, Man@pSiNPs-erastin promotes TAM exhaustion via the GPX4/RRM2 signaling pathway.
Enhancing TAM Phagocytosis
The relevant receptors included TLR2/4, TNFR 1, IL-17R, CD40, and RAGE. In breast cancer and melanoma, cGAMP@CaP@Abs promote macrophage phagocytosis via the STING pathway. In breast cancer and melanoma, ICG/CMSN@GsE promotes macrophage phagocytosis via the (TLR2/4, TNFR 1, IL-17R, CD40, RAGE)-MyD88-NF-κB pathway.
Nano-Immunotherapy of TAMs Through Tumor Cells
NDS can regulate the production and release of mediators (cytokines and metabolites) derived from tumor cells, and these released mediators can target TAMs for intervention. More specifically, by internalizing the drug-loaded system into tumor cells, these systems induce the secretion of CSF-1, ROS, mitochondrial DNA, DAMPs, and HIF-1α or inhibit the LOX pathway to achieve therapeutic effects. FG-CD@Cu reprograms M2 to M1 TAMs by generating O-2, effectively improving the hypoxic environment of the cells and leading to a reduction in HIF-1α expression, promoting tumor immunotherapy, and restraining tumor growth in colon cells.207 MT-NPs (siATP6), which use peptides and PEG polymers as carriers, indirectly reprogram M2 to M1 TAMs via activation of the TLR9 pathway (which involves localizing siRNA within mitochondria to deplete proteins such as ATP6/CYB, increasing ROS levels, inducing organelle damage, and culminating in the release of mtDNA into the tumor stroma), promoting tumor immunotherapy in BCa.208 N-PB, when combined with photodynamic therapy, decreases M2 TAMs by downregulating CSF-1 expression by antagonizing the RelA/NF-κB axis to elicit a systemic ATIR that suppresses metastatic breast cancer.209 In combination with photodynamic therapy, PLNR840 reprograms M2 to M1 TAMs via the LOX pathway, reprogramming ATIR metabolism in HCC and breast cancer.210 M@Au-D/B NCs, in combination with photodynamic therapies, reprogram M2 to M1 TAMs via ROS generation and PTT, promoting ATIR in breast cancer.211 Combining photodynamic therapies with LASNEO triggers effective PDT and augments M1 macrophage polarization via ROS, resulting in excellent anti-tumor effects on colon cancer and HCC.212 MNP-RGD-TPP, combined with magnetic hyperthermia treatment, reprograms M2 macrophages to M1 TAMs through surface exposure to calreticulin and the emission of immunogenic DAMPs (ATP/HSP70) from dying cancer cells, which triggers macrophage activation, promoting tumor immunotherapy in HCC.213 Interestingly, multiple research groups are actively pursuing photothermal combination therapy targeting TAMs through tumor cells. Specifically, biological carriers have emerged as good choices for targeting TAMs because of their intrinsic targeting capabilities, which are mediated by naturally occurring membrane proteins and surface markers that enable precise recognition of and interaction with specific receptors overexpressed on TAMs. Wei et al developed gold nanocages that mimic cancer cell membranes as carriers, whereas Zhang et al utilized exosomes from natural killer (NK) cells. Additionally, Han et al integrated magnetic hyperthermia, not only by leveraging its physical ablation effects but also by efficiently activating the body’s own ATIR by targeting and regulating TAMs.
Alternative Strategies for Nano-Immunotherapy Targeting TAMs
TME Modulation
NDSs indirectly affect TAMs mainly by reversing acidity in the TME. AL@O2-FMRC suppresses lactic acid production and accumulation while neutralizing the acidic microenvironment shaped by lactic acid, thereby abrogating lactic acid-driven immunosuppression.214 DMGTF NCs block MCT4-mediated lactate efflux and impair mitochondrial lactate oxidation upon microwave triggering, converting metabolic fuel into inert waste to eliminate lactate-imposed immunosuppressive pressure on TAMs.215
ECM Targeting
The ECM acts as a physical barrier that restricts the penetration and distribution of nano-drug delivery systems into TAM-enriched regions. Moreover, its composition and density can regulate the polarization state of TAMs. Therefore, overcoming the ECM barrier is critical for improving the therapeutic efficacy of TAM-targeted nano immunotherapy. NF#PTX@Soluplus/DG-pp-FA, a needleless jet injection system, employs a dual strategy combining “physical breakthrough” and “biological modification” to overcome the barrier of the ECM, achieving efficient drug penetration and retention.216 STAR preferentially releases the small molecule losartan to actively loosen the tumor matrix, reducing the density of the fiber network and thereby creating permeable interstitial pathways for subsequent nanoparticles to carry TAM-modulating payloads.217 Biomimetic oxidized mRNA lipid nanoparticles directly deliver collagenase-encoding mRNA into macrophages, enabling them to secrete matrix-degrading enzymes while expressing chimeric antigen receptors, thus transforming engineered TAMs into mobile ECM-degrading units that clear barrier matrices as they migrate.218
Drug Penetration
Drug penetration is now achieved mainly through charge-driven translocation, in situ synthesis, particle size modulation, and surface modification. Cou/SN38 NPs rely on site-specific ultrasound to cleave coumarin bonds, switching the surface charge from negative to positive and then driving iterative transcellular transport through Golgi-dependent transcytosis to achieve deep penetration and deliver chemotherapeutics to TAMs in the core region.219 PL@mBiME exploits the acidic tumor microenvironment to trigger charge reversal, efficiently crosses the blood-brain barrier, and expresses the bispecific macrophage engager BiME in situ within the glioblastoma parenchyma, thereby establishing physical connections between deep M2 macrophages and tumor cells through in situ protein synthesis.220 LNP-panCAR achieves efficient transvascular extravasation and intratumoral diffusion by modulating the particle size and hydrophilic–hydrophobic balance and delivering multiple chimeric antigen receptor-encoding mRNAs deep into macrophages and other immune cells to accomplish in situ engineering of TAMs directly in the tumor core.221
Clinical Relevance
Nano-immunotherapeutic strategies targeting TAMs exhibit varying degrees of clinical maturity. TAM depletion using CSF-1R inhibitors such as pexidartinib has received FDA approval for tenosynovial giant cell tumor.222 Phagocytosis enhancement with anti-CD47 antibodies has advanced to Phase II/III trials in both solid and hematologic tumors.223 Inhibition of TAM recruitment via CCR2 antagonists and reprogramming using TLR agonists has completed phase I/II exploration in early stage solid tumors.224 In contrast, strategies including polarization modulation, regulation of secreted mediators, blockade of PD-L1 expression on TAMs, and metabolic regulation remain predominantly at the preclinical or early Phase I stage. Collectively, these strategies have shown the potential for clinical application by reversing the immunosuppressive TME, overcoming drug resistance, and inhibiting metastasis. However, they face challenges such as systemic toxicity, including cytokine storm, anemia, and immunodeficiency, as well as tumor heterogeneity and poor delivery efficiency caused by the extracellular matrix barrier. The rapid update and iteration of NDSs aim to enhance safety and efficacy through active targeting, intelligent response release, and local administration methods. Future efforts should focus on combination therapies and multicenter randomized controlled trials to further validate their clinical value. Overall, the clinical translation of nano-immunotherapy targeting tumor-associated macrophages still has a long way to go.
Conclusion
Nano-immunotherapy targeting TAMs adopts multi-modal collaborative intervention, aiming to break through the ECM barrier, enhance drug penetration, precisely regulate TAMs, and reverse the immunosuppressive TME. The innovations of nanocarriers involve the entire chain intelligent design from inorganic nanocrystals, organic polymers to organic-inorganic hybrid systems, and the precise “Trojan horses” constructed using biodegradable polymers and biomimetic membrane vesicles. These carriers not only enable precise antigen presentation through surface functionalization but also achieve spatiotemporally controlled drug release by responding to the unique pH levels of TME, enzymatic activity, and ROS. Therapeutic strategies have ushered in the era of multi-modal therapy characterized by “multi-drug synergy” and “combined physical energy interventions”. Nanocarriers co-deliver reprogrammed drugs and immune checkpoint inhibitors to reverse the M2 phenotype to the anti-tumor M1 phenotype. Physical energy is transformed into a “trigger” that activates ATIR by integrating photothermal, sonodynamic, or magnetothermal therapies. This approach not only induces ICD and in situ tumor vaccine generation but also enables the intelligently controlled release of TAMs to regulate drug delivery. This temporal-spatial coordination of chemical-immunological-physical therapies generates potent distal and memory immune effects.20,225
Current strategies to extend circulation time and enhance active targeting of nanomedicines rely primarily on biomimetic cell membrane coating and ligand modification. Further conjugation of TAM-specific targeting peptides or antibodies enables active receptor recognition.226 Surface modification with TAM-specific ligands (eg, mannose, TLR4 agonists, or CRV peptides) drives receptor-mediated endocytosis, fundamentally shifting away from passive uptake.226 Moreover, engineered bacteria that colonize hypoxic tumor cores can continuously secrete reprogramming factors and consume immunosuppressive metabolites, offering a solution to the limited durability of conventional drug delivery and the difficulty of penetrating the ECM barrier.
NDSs can not only achieve targeting through pathological chemotaxis, but also synthesize and release reprogrammed drugs in situ when recognizing M2-specific surface markers, enabling integrated “diagnostic-treatment” precision strikes.227,228 Recent studies have revealed that tumor cells can remotely reshape myeloid generation in bone marrow by inducing NRF2-mediated oxidative stress responses in myeloid progenitor cells, while continuously amplifying the immunosuppressive program of the monocyte–macrophage lineage within the TME.229 These discoveries have brought about a comprehensive revolution in future therapeutic strategies, spanning from treatment concepts to target design. Intelligent nanorobots targeting TAMs, as autonomous diagnostic and therapeutic systems integrating sensing, decision-making, and execution, represent an effective approach to enhancing the precision and intelligence of anti-tumor strategies.
Abbreviations
NDSs, Nano-drug delivery systems; PI3K, Phosphoinositide 3-kinase; ECM, Extracellular matrix; TME, Tumor microenvironment; Akt, Protein kinase B; PAMP, Pathogen-associated molecular pattern; TAMs, Tumor-associated macrophages; Gbp2, Guanylate-binding protein 2; TAA, Tumor-associated antigen; ATIR, Anti-tumor immune responses; Pin1, Peptidyl-prolyl cis-trans isomerase NIMA-interacting 1; PLAG, Poly (lactic acid-co-glycolic acid); ROS, Reactive oxygen species; AMPK, AMP-activated protein kinase; HNSCC, Head and neck squamous cell carcinoma; OSCC, Oral squamous cell carcinoma; AFP, Alpha-fetoprotein; REG, Regorafenib; iNOS, Inducible nitric oxide synthase; PD-1, Programmed cell death protein 1; SHP099, SHP2 inhibitor; MMP, Matrix metalloproteinase; CTLs, Cytotoxic T lymphocytes; TfR, Transferrin receptor; LA-NSCLC, Locally advanced non-small cell lung cancer; STING, Stimulator of interferon genes; NMs, Nanomicelles; CSF-1, Colony-stimulating factor 1; cGAS, Cyclic GMP-AMP synthase; IFN-γ, Interferon gamma; CSF-1R, Colony-stimulating factor 1 receptor; NLRP3, NLR family pyrin domain containing 3; GOx, Glucose oxidase; CCL2, C-C motif chemokine ligand 2; MAPK, Mitogen-activated protein kinase; CHA, Chlorogenic acid; CXCR2, C-X-C motif chemokine receptor 2; JNK, c-Jun N-terminal kinase; MPNs, Metal-phenolic networks; CCR5, C-C motif chemokine receptor 5; NF-κB, Nuclear factor kappa B; UIONPs, Ultra-small iron oxide nanoparticles; CCL5, C-C motif chemokine ligand 5; STAT6, Signal transducer and activator of transcription 6; IKKβ, Inhibitor of nuclear factor kappa-B kinase beta; CD47, Cluster of differentiation 47; HIF-1α, Hypoxia-inducible factor 1-alpha; IκBα, Nuclear factor of kappa light polypeptide gene enhancer in B-cells inhibitor alpha; SIRP-α, Signal regulatory protein alpha; VEGF, Vascular endothelial growth factor; FVIO, Ferrimagnetic vortex-domain iron oxide; TREM2, Triggering receptor expressed on myeloid cells 2; FasL, Fas ligand; DLC, Material candidate; FLT3L, Fms-like tyrosine kinase 3 ligand; Bax, BCL2-associated X protein; SnSe, Tin selenide; STAT3, Signal transducer and activator of transcription 3; SHP2, Src homology region 2-containing protein tyrosine phosphatase 2; NSs, Nanosheets; IFN, Interferon; SIRPα, Signal regulatory protein alpha; scFv, Single-chain variable fragment; TNF, Tumor necrosis factor; DC, Dendritic cells; Lipo, Liposome; TLR, Toll-like receptor; MDSCs, Myeloid-derived suppressor cells; Zol, Zoledronic acid; IL-4, Interleukin 4; Treg, Regulatory T cells; AeroNP, Aerosolized nanoparticle; IL-13, Interleukin 13; ICD, Immunogenic cell death; CDN, Cyclic dinucleotide; IL-1, Interleukin 1; PDT, Photodynamic therapy; SINPs, Silicon nanoparticles; IL-6, Interleukin 6; PTT, Photothermal therapy; RPST, Polymer system; IL-12, Interleukin 12; SDT, Sonodynamic therapy; IMQ, Imiquimod; IL-18, Interleukin 18; MHT, Magnetothermal therapy; LyP-1, Targeting peptide; IL-23, Interleukin 23; CSF-1Ri, CSF-1 receptor inhibitor; CuS, Copper sulfide; TNF-α, Tumor necrosis factor alpha; IDO, Indoleamine 2,3-dioxygenase; DMXAA, 5,6-dimethylxanthenone-4-acetic acid; M2pep, M2-targeting peptide; DHA, Docosahexaenoic acid; LOX, Lysyl oxidase; CCL3, C-C motif chemokine ligand 3; CPT, Camptothecin; Ce6, Chlorin e6; scRNA-seq, Single-cell RNA sequencing; Tpl, α-Tocopherol; Tf, Transferrin; PEG, Polyethylene glycol; Tcc, Toyocamycin; IL-10, Interleukin 10; GO, Graphene oxide; ER, Endoplasmic reticulum; IL-12, Interleukin 12; PEI, Polyethylenimine; IFN-β, Interferon beta; OMV, Outer membrane vesicles; MAN, Mannose; PGE2, Prostaglandin E2; Cr, Chromium; MOF, Metal-organic framework; GM-CSF, Granulocyte-macrophage colony stimulating factor; CTS, Chitosan; DOX, Doxorubicin; Arg1, Arginase 1; siYTHDF1, Small interfering RNA against YTHDF1; FA, Folic acid; TGF-β, Transforming growth factor beta; m6A, N6-methyladenosine; RHPs, Rhodiola rosea polysaccharides; LAP, LC3-associated phagocytosis; STAT1, Signal transducer and activator of transcription 1; SA, Sodium alginate; PA, Palmitic acid; MS, Mesoporous silica; MTX, Methotrexate; caspase-3, Cysteinyl aspartate specific proteinase 3; CTPP, Targeting peptide; PPT, Podophyllotoxin; aCD47, Anti-CD47 antibody; IMD, Imidazoquinoline; APS, Astragalus polysaccharide; aPD-L1, Anti-PD-L1 antibody; CD, Cyclodextrin; CMA, Carboxymethylated alginate; PLL, Poly-L-lysine; Ada, Adamantane; R848, Resiquimod; DMMA, Dimethylmaleic anhydride; GMNPs, Gold nanoparticles; GC, Gastric cancer; MA, Maleic anhydride; DFMO, Difluoromethylornithine; DEX, Dexamethasone; Ab, Antibody; ICG, Indocyanine green; TLR7, Toll-like receptor 7; TMZ, Temozolomide; aPD-L1, Anti-PD-L1 antibody; HCC, Hepatocellular carcinoma; MMR, Mannose receptor; ACOD1, Aconitate decarboxylase 1; CpG, Cytosine-phosphate-guanine; CaNP, Calcium nanoparticle; CMSN, Mesoporous silica nanoparticles; TLR9, Toll-like receptor 9; NGs, Nanogels; GsE, Ginseng exosomes; IRF3, Interferon regulatory factor 3; PTX, Paclitaxel; TLR2, Toll-like receptor 2; DDC, Diethyldithiocarbamate; CUR, Curcumin; TLR4, Toll-like receptor 4; GPC3, Glypican-3; PLA, Polylactic acid; TNFR1, Tumor necrosis factor receptor 1; BSG, Brainstem glioma; GEM, Gemcitabine; IL-17R, Interleukin 17 receptor; Pd, Palladium; CS, Chitosan; CD40, Cluster of differentiation 40; HA, Hyaluronic acid; ANX5, Annexin V; RAGE, Receptor for advanced glycation end products; R837, Imiquimod; MyD88, Myeloid differentiation 88; CW, Cell wall; GBM, Glioblastoma; Met, Metformin; PSS, Poly (sodium 4-styrenesulfonate); TNBC, Triple-negative breast cancer; MPs, Microparticles; PEI, Polyethylenimine; PLGA, Poly (lactic-co-glycolic acid); IM, Imipramine; shRNA, Short hairpin RNA.
Acknowledgments
We extend our appreciation to the authors, whose figures and work have been referenced in this review, contributing significantly to the depth of our discussion. We also acknowledge the use of BioRender for assisting with professional preparation of our figures.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
The research was supported by the International Science and Technology Cooperation Project, Jilin Provincial Department of Science and Technology (20250205010GH).
Disclosure
The authors declare no competing interests in this work.
References
1. Sun XQ, Park MD, Merad M, Brown BD. Macrophages: targets for next-generation cancer immunotherapy. Cancer Cell. 2026;44(3):498–42. doi:10.1016/j.ccell.2026.01.020
2. Ghamangiz S, Jafari A, Maleki-Kakelar H, Azimi H, Mazloomi E. Reprogram to heal: macrophage phenotypes as living therapeutics. Life Sci. 2025;371:123601. doi:10.1016/j.lfs.2025.123601
3. Ma R-Y, Black A, Qian B-Z. Macrophage diversity in cancer revisited in the era of single-cell omics. Review. Trends Immunol. 2022;43(7):546–563. doi:10.1016/j.it.2022.04.008
4. Wang J, Zhu N, Su X, Gao Y, Yang R. Novel tumor-associated macrophage populations and subpopulations by single cell RNA sequencing. Review. Front Immunol. 2024;14:1264774. doi:10.3389/fimmu.2023.1264774
5. Shao N, Qiu H, Liu J, et al. Targeting lipid metabolism of macrophages: a new strategy for tumor therapy Article. J Adv Res. 2025;68:99–114. doi:10.1016/j.jare.2024.02.009
6. Jin R, Neufeld L, McGaha TL. Linking macrophage metabolism to function in the tumor microenvironment. Review. Nat Cancer. 2025;6(2):239–252. doi:10.1038/s43018-025-00909-2
7. Bied M, Ho WW, Ginhoux F, Bleriot C. Roles of macrophages in tumor development: a spatiotemporal perspective. Review. Cell Mol Immunol. 2023;20(9):983–992. doi:10.1038/s41423-023-01061-6
8. Basak U, Sarkar T, Mukherjee S, et al. Tumor-associated macrophages: an effective player of the tumor microenvironment Review. Front Immunol. 2023;14:1295257. doi:10.3389/fimmu.2023.1295257
9. Mercedes SVA, Bocci F, Levine H, Onuchic JN, Jolly MK, Wong PK. Decoding leader cells in collective cancer invasion. Nat Rev Cancer. 2021;21(9):592–604. doi:10.1038/s41568-021-00376-8
10. Wang S, Wang J, Chen Z, et al. Targeting M2-like tumor-associated macrophages is a potential therapeutic approach to overcome antitumor drug resistance. Review. Npj Precision Oncol. 2024;8(1):31. doi:10.1038/s41698-024-00522-z
11. Jumaniyazova E, Lokhonina A, Dzhalilova D, Miroshnichenko E, Kosyreva A, Fatkhudinov T. The role of macrophages in various types of tumors and the possibility of their use as targets for antitumor therapy. review. Cancers. 2025;17(3):342. doi:10.3390/cancers17030342
12. You YH, Tian ZW, Du Z, et al. M1-like tumor-associated macrophages cascade a mesenchymal/stem-like phenotype of oral squamous cell carcinoma via the IL6/Stat3/THBS1 feedback loop. J Exp Clin Cancer Res. 2022;41(1):10. doi:10.1186/s13046-021-02222-z
13. Jiang MJ, Liu LY, Huang W, Qi Y, Li YF, Li B. HMGB1-activated tumor-associated macrophages promote migration and invasion via NF-xB/IL-6 signaling in oral squamous cell carcinoma. Int Immunopharmacol. 2024;126:111200. doi:10.1016/j.intimp.2023.111200
14. Wang Y, Johnson KCC, Gatti-Mays ME, Li Z. Emerging strategies in targeting tumor-resident myeloid cells for cancer immunotherapy. Review. J Hematol Oncol. 2022;15(1):118. doi:10.1186/s13045-022-01335-y
15. Li MJ, He LY, Zhu J, Zhang P, Liang SF. Targeting tumor-associated macrophages for cancer treatment. Cell Biosci. 2022;12(1):85. doi:10.1186/s13578-022-00823-5
16. Kloosterman DJ, Akkari L. Macrophages at the interface of the co-evolving cancer ecosystem. Review. Cell. 2023;186(8):1627–1651. doi:10.1016/j.cell.2023.02.020
17. Yang L, Zhang Y. Tumor-associated macrophages: from basic research to clinical application. Review. J Hematol Oncol. 2017;10:58. doi:10.1186/s13045-017-0430-2
18. Li Q, Xu J, Hua R, Xu H, Wu Y, Cheng X. Nano-strategies for targeting tumor-associated macrophages in cancer immunotherapy. Review. J Cancer. 2025;16(7):2261–2274. doi:10.7150/jca.108194
19. Xia Y, Rao L, Yao H, Wang Z, Ning P, Chen X. Engineering macrophages for cancer immunotherapy and drug delivery. Article. Adv Mater. 2020;32(40):2002054. doi:10.1002/adma.202002054
20. Yang Q, Guo N, Zhou Y, Chen J, Wei Q, Han M. The role of tumor-associated macrophages (TAMs) in tumor progression and relevant advance in targeted therapy. Review. Acta Pharmaceutica Sinica B. 2020;10(11):2156–2170. doi:10.1016/j.apsb.2020.04.004
21. Chu XY, Tian Y, Lv C. Decoding the spatiotemporal heterogeneity of tumor-associated macrophages. Mol Cancer. 2024;23(1):150. doi:10.1186/s12943-024-02064-1
22. Nabet BY, Hamidi H, Lee MC, et al. Immune heterogeneity in small-cell lung cancer and vulnerability to immune checkpoint blockade. Cancer Cell. 2024;42(3):429–443. doi:10.1016/j.ccell.2024.01.010
23. Ma ZK, Wang YZ, Wang WK, et al. Targeting VSIG4+ tissue-resident macrophages enhances T cell cytotoxicity and immunotherapy efficacy in cancer. Dev Cell. 2025;60(19):2594–2610. doi:10.1016/j.devcel.2025.04.011
24. Lu Y, Luo C, Huang L, et al. Functional genetic screens reveal key pathways instructing the molecular phenotypes of tumor-associated macrophages. Cancer Immunol Res. 2025. doi:10.1158/2326-6066.Cir-25-0488
25. Liu X, Zhang ZF, Yuan JP, Yu JM, Chen DW. Spatial interaction and functional status of CD68+SHP2+ macrophages in tumor microenvironment correlate with overall survival of NSCLC. Front Immunol. 2024;15:1396719. doi:10.3389/fimmu.2024.1396719
26. Lyu A, Fan ZH, Clark M, et al. Evolution of myeloid-mediated immunotherapy resistance in prostate cancer. Nature. 2025;637(8048):1207–1217. doi:10.1038/s41586-024-08290-3
27. Sazinsky S, Zafari M, Klebanov B, et al. Antibodies targeting human or mouse VSIG4 repolarize tumor-associated macrophages providing the potential of potent and specific clinical anti-tumor response induced across multiple cancer types. Int J Mol Sci. 2024;25(11):6160. doi:10.3390/ijms25116160
28. Li M, Zhang YJ, Zhang Q, Li JC. Tumor extracellular matrix modulating strategies for enhanced antitumor therapy of nanomedicines. Mater Today Bio. 2022;16100364. doi:10.1016/j.mtbio.2022.100364
29. Hu J, Yuan XW, Wang F, Gao HL, Liu XL, Zhang W. The progress and perspective of strategies to improve tumor penetration of nanomedicines. Chin Chem Lett. 2021;32(4):1341–1347. doi:10.1016/j.cclet.2020.11.006
30. Zhang YR, Lin R, Li HJ, He WL, Du JZ, Wang J. Strategies to improve tumor penetration of nanomedicines through nanoparticle design. Wiley Interdiscip Rev. 2019;11(1):e1519. doi:10.1002/wnan.1519
31. Xu XX, Wu Y, Qian XD, et al. Nanomedicine strategies to circumvent intratumor extracellular matrix barriers for cancer therapy. Adv Healthcare Mater. 2022;11(1):2101428. doi:10.1002/adhm.202101428
32. Choi I-K, Strauss R, Richter M, Yun C-O, Lieber A. Strategies to increase drug penetration in solid tumors. Front Oncol. 2013;3:193. doi:10.3389/fonc.2013.00193
33. Yan WL, Li Y, Zou YT, et al. Breaking tumor immunosuppressive network by regulating multiple nodes with triadic drug delivery nanoparticles. Acs Nano. 2023;17(18):17826–17844. doi:10.1021/acsnano.3c03387
34. Liu SJ, Tian BL, Wang N, et al. MARCO expression on myeloid-derived suppressor cells is essential for their differentiation and immunosuppression. Cell Death Discovery. 2025;11(1):337. doi:10.1038/s41420-025-02627-1
35. Cai JL, Zhang PL, Cai YF, et al. Lactylation-driven NUPR1 promotes immunosuppression of tumor-infiltrating macrophages in hepatocellular carcinoma. Adv Sci. 2025;12(20):2413095. doi:10.1002/advs.202413095
36. Liu R, Guo L, Shi DD, et al. Multilayer cascade-response nanoplatforms as metabolic symbiotic disruptors to reprogram the immunosuppressive microenvironment. J Control Release. 2025:383113797. 10.1016/j.jconrel.2025.113797.
37. Liu MZ, Chen QX, Zhang LL, et al. Metabolic engineering of SLC38A2 reprograms glutamine utilization and enhances CAR-macrophage antitumor function in solid tumors. Cancer Biol Med. 2026;23(3):392–417. doi:10.20892/j.issn.2095-3941.2025.0775
38. Wang J, Liang Z, Wang Y, et al. Mannose modified graphene oxide drug-delivery system targets cancer stem cells and tumor-associated macrophages to promote immunotherapeutic efficacy. Article. Colloids Surf B Biointerfaces. 2025;253:114710. doi:10.1016/j.colsurfb.2025.114710
39. Wei Z, Zhang X, Zhang Z, et al. Engineered iron-based nanoplatform amplifies repolarization of M2-like tumor-associated macrophages for enhanced cancer immunotherapy. Article. Chem Eng J. 2022;433:133847. doi:10.1016/j.cej.2021.133847
40. Xiong Y-X, Li N, Han -M-M, et al. Rhodiola rosea polysaccharides-based nanoparticles loaded with DOX boosts chemo-immunotherapy for triple-negative breast cancer by re-educating Tumor-associated macrophages. Article. Int J Biol Macromol. 2023;239:124110. doi:10.1016/j.ijbiomac.2023.124110
41. Liu M, Zhang Z-X, Wang J-H, et al. Immunomodulatory and anti-ovarian cancer effects of novel astragalus polysaccharide micelles loaded with podophyllotoxin. Article. Int J Biol Macromol. 2025;290:138960. doi:10.1016/j.ijbiomac.2024.138960
42. Chen J, Liu X, Zhao S, et al. Carboxymethylated alginate-resiquimod micelles reverse the immunosuppressive tumor microenvironment and synergistically enhance the chemotherapy and immunotherapy for gastric cancer. Article. ACS Appl Mater Interfaces. 2023;15(30):35999–36012. doi:10.1021/acsami.3c06828
43. Ota Y, Inagaki R, Takanashi Y, et al. Targeting tumor-associated macrophages with the immune-activating nanomedicine for achieving strong antitumor activity with rapid clearance from the body. Article. Acs Nano. 2024;18(34):23757–23772. doi:10.1021/acsnano.4c08811
44. Huang C, Xie T, Liu Y, et al. A sodium alginate-based multifunctional nanoplatform for synergistic chemo-immunotherapy of hepatocellular carcinoma. Article. Adv Mater. 2023;35(33):2301352. doi:10.1002/adma.202301352
45. Kim Y, Lee S, Jon S. Liposomal delivery of an immunostimulatory CpG induces robust antitumor immunity and long-term immune memory by reprogramming tumor-associated macrophages. Article. Adv Healthcare Mater. 2024;13(6):2300549. doi:10.1002/adhm.202300549
46. Kim J, Kim M, Han H, Kim S, Lahiji SF, Kim Y-H. Dual-delivery of exosome inhibitor and immune-activating gene via lipid nano-assemblies for tumor immune evasion inhibition. J Control Release. 2025;381:113569. doi:10.1016/j.jconrel.2025.02.065
47. Wang R, Song W, Zhu J, et al. Biomimetic nano-chelate diethyldithiocarbamate Cu/Fe for enhanced metalloimmunity and ferroptosis activation in glioma therapy. Article. J Control Release. 2024;368:84–96. doi:10.1016/j.jconrel.2024.02.004
48. Gao L, Shi C, Yang Z, et al. Convection-enhanced delivery of nanoencapsulated gene locoregionally yielding ErbB2/Her2-specific CAR-macrophages for brainstem glioma immunotherapy. Article. J Nanobiotechnol. 2023;21(1):56. doi:10.1186/s12951-023-01810-9
49. Chen X, Jia Z, Wen Y, et al. Bidirectional anisotropic palladium nanozymes reprogram macrophages to enhance collaborative chemodynamic therapy of colorectal cancer. Article. Acta Biomater. 2022;151:537–548. doi:10.1016/j.actbio.2022.08.020
50. Lu D, Di Z, Li L, Zhao J, Zheng L. An aptamer-driven DNA nanodevice for improved delivery of synthetic immunostimulants. Article; Early Access. Nano Res. 2024;17(10):9078–9083. doi:10.1007/s12274-024-6632-3
51. Lu X, Xia H, Gao W, et al. A pH-Responsive and Guanidinium-Rich Nanoadjuvant Efficiently Delivers STING Agonist for Cancer Immunotherapy. Article. Acs Nano. 2025;19(7):6758–6770. doi:10.1021/acsnano.4c10202
52. Wang Y, Li G, Su J, et al. Tumor-associated macrophages nano-reprogrammers induce “gear effect” to empower glioblastoma immunotherapy. Article. Small. 2025;21(6):2406839. doi:10.1002/smll.202406839
53. Goswami R, Nabawy A, Jiang M, et al. All-natural gelatin-based nanoemulsion loaded with TLR 7/8 agonist for efficient modulation of macrophage polarization for immunotherapy. Article. Nanomaterials. 2024;14(19):1556. doi:10.3390/nano14191556
54. Rodriguez-Perdigon M, Jimaja S, Haeni L, Bruns N, Rothen-Rutishauser B, Rueegg C. Polymersomes-mediated delivery of CSF1R inhibitor to tumor associated macrophages promotes m2 to m1-like macrophage repolarization. Article. Macromol biosci. 2022;22(8):2200168. doi:10.1002/mabi.202200168
55. Fu J, Zhao W, Liang N, Sun S. Functionalized hydroxypropyl-β-cyclodextrin inclusion complex for combined tumor therapy through intelligent delivery of paclitaxel and polarization of M2-like tumor associated macrophages. Article. Colloids Surf B. 2025;252:114654. doi:10.1016/j.colsurfb.2025.114654
56. Li X, Pan J, Li Y, et al. Development of a localized drug delivery system with a step-by-step cell internalization capacity for cancer immunotherapy. Article. Acs Nano. 2022;16(4):5778–5794. doi:10.1021/acsnano.1c10892
57. Wei Y, Wu S, Liu Z, et al. Tumor associated macrophages reprogrammed by targeted bifunctional bioorthogonal nanozymes for enhanced tumor immunotherapy. Article. Mater Today. 2022;56:16–28. doi:10.1016/j.mattod.2022.01.024
58. Yang M, Pu L, Yang S, et al. Engineered antler stem cells derived exosomes potentiate anti-tumor efficacy of immune checkpoint inhibitor by reprogramming immunosuppressive tumor microenvironment. Article. Chem Eng J. 2024;479:147421. doi:10.1016/j.cej.2023.147421
59. Qin J, Liu J, Wei Z, et al. Targeted intervention in nerve-cancer crosstalk enhances pancreatic cancer chemotherapy. Article. Nature Nanotechnol. 2025;20(2):311–324. doi:10.1038/s41565-024-01803-1
60. Han S, Bao X, Zou Y, et al. d-lactate modulates M2 tumor-associated macrophages and remodels immunosuppressive tumor microenvironment for hepatocellular carcinoma. Article. Sci Adv. 2023;9(29):EADG2697. doi:10.1126/sciadv.adg2697
61. Xiong J, Huang J, Xu H, et al. CpG-based nanovaccines enhance ovarian cancer immune response by Gbp2-mediated remodeling of tumor-associated macrophages. Article. Adv Sci. 2025;12(15):2412881. doi:10.1002/advs.202412881
62. Chong L, Jiang Y-W, Wang D, Chang P, Xu K, Li J. Targeting and repolarizing M2-like tumor-associated macrophage-mediated MR imaging and tumor immunotherapy by biomimetic nanoparticles. Article. J Nanobiotechnol. 2023;21(1):401. doi:10.1186/s12951-023-02122-8
63. Zhao G, Liu H, Wang Z, et al. Exosome transportation-me diate d immunosuppression relief through cascade amplification for enhanced apoptotic body vaccination. Article. Acta Biomater. 2022;153:529–539. doi:10.1016/j.actbio.2022.09.014
64. Zhang X, Wei Z, Yong T, et al. Cell microparticles loaded with tumor antigen and resiquimod reprogram tumor associated macrophages and promote stem like CD8+ T cells to boost anti-PD-1 therapy. Article. Nat Commun. 2023;14(1):5653. doi:10.1038/s41467-023-41438-9
65. Zhang X, Yue L, Cao L, et al. Tumor microenvironment-responsive macrophage-me diate d immunotherapeutic drug delivery. Article. Acta Biomater. 2024;186:369–382. doi:10.1016/j.actbio.2024.07.042
66. Fan Z, Jiang X, Sun T, et al. In vivo visualization of tumor-associated macrophages re-education by photoacoustic/fluorescence dual-modal imaging with a metal-organic frames-based caspase-1 nanoreporter. Article. J Colloid Interface Sci. 2024;659:48–59. doi:10.1016/j.jcis.2023.12.123
67. Gu Z, Da silva CG, Ma S, et al. Dual-targeting nanoliposome improves proinflammatory immunomodulation of the tumor microenvironment. Article. Adv Healthcare Mater. 2023;12(31):2302046. doi:10.1002/adhm.202302046
68. Li B, Tan T, Chu W, et al. Co-delivery of paclitaxel (PTX) and docosahexaenoic acid (DHA) by targeting lipid nanoemulsions for cancer therapy. Article. Drug Delivery. 2022;29(1):75–88. doi:10.1080/10717544.2021.2018523
69. Li Z, Jiang S, Wang J, et al. Peptide-drug conjugates repolarize glioblastoma-associated macrophages to resensitize chemo-immunotherapy of glioblastoma. Article. Sci Adv. 2025;11(3):EADR8841. doi:10.1126/sciadv.adr8841
70. Xiao Q, Huang J, Wang X, et al. Supramolecular peptide amphiphile nanospheres reprogram tumor-associated macrophage to reshape the immune microenvironment for enhanced breast cancer immunotherapy. Article. Small. 2024;20(21):2307390. doi:10.1002/smll.202307390
71. Hu Y, Nie W, Lyu L, et al. Tumor-microenvironment-activatable nanoparticle mediating immunogene therapy and M2 macrophage-targeted inhibitor for synergistic cancer immunotherapy. Article. Acs Nano. 2024;18(4):3295–3312. doi:10.1021/acsnano.3c10037
72. Huang L, Xu R, Li W, et al. Repolarization of macrophages to improve sorafenib sensitivity for combination cancer therapy. Article. Acta Biomater. 2023;162:98–109. doi:10.1016/j.actbio.2023.03.014
73. Zhang H, Chen F, Cheng W, et al. Dynamic polyphenol nanoparticles boost cuproptosis-driven metalloimmunotherapy of breast cancer. Article. Nano Today. 2024;58:102442. doi:10.1016/j.nantod.2024.102442
74. He Y, de Araujo RF, Cavalcante RS, et al. Effective breast cancer therapy based on palmitic acid-loaded PLGA nanoparticles. Article. Biomat Adv. 2023;145:213270. doi:10.1016/j.bioadv.2022.213270
75. Su Z, Dong S, Chen Y, et al. Microfluidics-enabled nanovesicle delivers CD47/PD-L1 antibodies to enhance antitumor immunity and reduce immunotoxicity in lung adenocarcinoma. Article. Adv Sci. 2023;10(20):2206213. doi:10.1002/advs.202206213
76. Meng S, Du H, Li X, et al. An adjuvant micelle-based multifunctional nanosystem for tumor immunotherapy by remodeling three types of immunosuppressive cells. Article. Acs Nano. 2024;18(4):3134–3150. doi:10.1021/acsnano.3c08792
77. Huang T, Zhang Q, Yi J, et al. PEG-sheddable nanodrug remodels tumor microenvironment to promote effector t cell infiltration and revise their exhaustion for breast cancer immunotherapy. Article. Small. 2023;19(38):2301749. doi:10.1002/smll.202301749
78. Li J, Yang J, Jiang S, et al. Targeted reprogramming of tumor-associated macrophages for overcoming glioblastoma resistance to chemotherapy and immunotherapy. Article. Biomaterials. 2024;311:122708. doi:10.1016/j.biomaterials.2024.122708
79. Jiang S, Li W, Yang J, et al. Cathepsin B-responsive programmed brain targeted delivery system for chemo-immunotherapy combination therapy of glioblastoma. Article. Acs Nano. 2024;18(8):6445–6462. doi:10.1021/acsnano.3c11958
80. An J, Liu M, Zhao L, et al. Boosting tumor immunotherapy by bioactive nanoparticles via ca2+ interference mediated TME reprogramming and specific PD-L1 depletion. Article. Adv Funct Mater. 2022;32(24):2201275. doi:10.1002/adfm.202201275
81. Chen Z, Wen T, Wang X, et al. Co-delivery of immunochemotherapeutic by classified targeting based on chitosan and cyclodextrin derivatives. Article. Int J Biol Macromol. 2023;226:1396–1410. doi:10.1016/j.ijbiomac.2022.11.253
82. Tang L, Yin Y, Zhang Z, et al. Size-switchable and dual-targeting nanomedicine for cancer chemoimmunotherapy by potentiating deep tumor penetration and antitumor immunity. Article. Chem Eng J. 2024;493:152590. doi:10.1016/j.cej.2024.152590
83. Zhang G, Zhan M, Zhang C, et al. Redox-responsive dendrimer nanogels enable ultrasound-enhanced chemoimmunotherapy of pancreatic cancer via endoplasmic reticulum stress amplification and macrophage polarization. Article. Adv Sci. 2023;10(24):2301759. doi:10.1002/advs.202301759
84. Li T, Chen D, Liu H, et al. Spatially targeting and regulating tumor-associated macrophages using a raspberry-like micellar system sensitizes pancreatic cancer chemoimmunotherapy. Article. Nanoscale. 2022;14(36):13098–13112. doi:10.1039/d2nr03053e
85. Ye C, Yan X, Dai X, et al. Biocompatible snowman-like tumor-targeting dimer nanoparticles for improved delivery efficiency and enhanced anti-tumor therapy. Article. Chem Eng J. 2023;474:145766. doi:10.1016/j.cej.2023.145766
86. Sun L, Gao W, Liu J, et al. O2-supplying nanozymes alleviate hypoxia and deplete lactate to eliminate tumors and activate antitumor immunity. Article. ACS Appl Mater Interfaces. 2022;14(51):56644–56657. doi:10.1021/acsami.2c18960
87. Chen Z, Li Z, Huang H, et al. Cancer immunotherapy based on cell membrane-coated nanocomposites augmenting cGAS/STING activation by efferocytosis blockade. Article. Small. 2023;19(43):2302758. doi:10.1002/smll.202302758
88. Zhang X, Wei Z, Ding Z, et al. Boosting doxil-based chemoimmunotherapy via reprogramming tumor-associated macrophages. Article. Chem Eng J. 2023;451:138971. doi:10.1016/j.cej.2022.138971
89. Zhang L, Zhang Y, Qi J, et al. Double-punch strategy: macrophage membrane-enveloped imipramine and aCD47 reprograms tumor microenvironment to enhance glioblastoma immunotherapy. Article. Chem Eng J. 2025;515:163715. doi:10.1016/j.cej.2025.163715
90. Cui J, Wang X, Li J, et al. Immune exosomes loading self-assembled nanomicelles traverse the blood-brain barrier for chemo-immunotherapy against glioblastoma. Article. Acs Nano. 2023;17(2):1464–1484. doi:10.1021/acsnano.2c10219
91. Cheng L, Zhang P, Liu Y, et al. Multifunctional hybrid exosomes enhanced cancer chemo-immunotherapy by activating the STING pathway. Article. Biomaterials. 2023;301:122259. doi:10.1016/j.biomaterials.2023.122259
92. Li Y, Zhu T, Chen J, et al. Dual-targeted engineered bacterial outer membrane vesicles for hepatocellular carcinoma immunotherapy. Article. Adv Funct Mater. 2024;34(41):2401355. doi:10.1002/adfm.202401355
93. Tang L, Fu C, Liu H, et al. Chemoimmunotherapeutic nanogel for pre- and postsurgical treatment of malignant melanoma by reprogramming tumor-associated macrophages. Article. Nano Lett. 2024;24(5):1717–1728. doi:10.1021/acs.nanolett.3c04563
94. Wu Y, Chang X, Yang G, et al. A physiologically responsive nanocomposite hydrogel for treatment of head and neck squamous cell carcinoma via proteolysis-targeting chimeras enhanced immunotherapy. Article. Adv Mater. 2023;35(12):2210787. doi:10.1002/adma.202210787
95. Li Z, Xu W, Yang J, et al. A tumor microenvironments-adapted polypeptide hydrogel/nanogel composite boosts antitumor molecularly targeted inhibition and immunoactivation. Article. Adv Mater. 2022;34(21):2200449. doi:10.1002/adma.202200449
96. Chen X-Y, Yan M-Y, Liu Q, Yu B-X, Cen Y, Li S-Y. Chimeric peptide engineered bioregulator for metastatic tumor immunotherapy through macrophage polarization and phagocytosis restoration. Article. Acs Nano. 2023;17(16):16056–16068. doi:10.1021/acsnano.3c04778
97. Sun P, Wu Z, Xiao Y, et al. TfR-T12 short peptide and pH sensitive cell transmembrane peptide modified nano-composite micelles for glioma treatment via remodeling tumor microenvironment. Article. Nanomed Nanotechnol Biol Med. 2022;41:102516. doi:10.1016/j.nano.2022.102516
98. Li Y, Yuan R, Luo Y, et al. A hierarchical structured fiber device remodeling the acidic tumor microenvironment for enhanced cancer immunotherapy. Article. Adv Mater. 2023;35(21):2300216. doi:10.1002/adma.202300216
99. Fu Z, Zhang Y, Chen X, et al. A versatile nanoplatform based on metal-phenolic networks inhibiting tumor growth and metastasis by combined starvation/chemodynamic/immunotherapy. Article. Adv Funct Mater. 2023;33(7):2211869. doi:10.1002/adfm.202211869
100. Li T, Song R, Sun F, et al. Bioinspired magnetic nanocomplexes amplifying STING activation of tumor-associated macrophages to potentiate cancer immunotherapy. Article. Nano Today. 2022;43:101400. doi:10.1016/j.nantod.2022.101400
101. Min T, Yang C, Zhang M, Hu P, Shi J. Mild magnetothermal immunotherapy for malignant pleural effusion. Article; Early Access. Small. 2024;2407734. doi:10.1002/smll.202407734
102. Wang S, Jiao W, Yan B, et al. Intracellular magnetic hyperthermia enables concurrent down-regulation of CD47 and SIRPα to potentiate antitumor immunity. Article. Nano Lett. 2024;24(9):2894–2903. doi:10.1021/acs.nanolett.4c00003
103. Ni J, Tian H, Qin S, et al. Disulfide bond-rich nano-silica for high-efficiency cancer immunotherapy using synergistic endoplasmic reticulum stress cooperated mitochondrial dysfunction. Article. Chem Eng J. 2024;498:154877. doi:10.1016/j.cej.2024.154877
104. Ling J, Chang Y, Yuan Z, Chen Q, He L, Chen T. Designing lactate dehydrogenase-mimicking SnSe nanosheets to reprogram tumor-associated macrophages for potentiation of photothermal immunotherapy. Article. ACS Appl Mater Interfaces. 2022;14(24):27651–27665. doi:10.1021/acsami.2c05533
105. Zhu L, Li J, Guo Z, Kwok HF, Zhao Q. Synergistic combination of targeted nano-nuclear-reactors and anti-PD-L1 nanobodies evokes persistent T cell immune activation for cancer immunotherapy. Article. J Nanobiotechnol. 2022;20(1):521. doi:10.1186/s12951-022-01736-8
106. Jian H, Wang X, Song P, et al. Tumor microcalcification-mediate d relay drug delivery for photodynamic immunotherapy of breast cancer. Article. Acta Biomater. 2022;140:518–529. doi:10.1016/j.actbio.2021.12.014
107. Liu Y, Crowe WN, Wang L, Petty WJ, Habib AA, Zhao D. Aerosolized immunotherapeutic nanoparticle inhalation potentiates PD-L1 blockade for locally advanced lung cancer. Article. Nano Res. 2023;16(4):5300–5310. doi:10.1007/s12274-022-5205-6
108. Gong Y, Gao W, Zhang J, Dong X, Zhu D, Ma G. Engineering nanoparticles-enabled tumor-associated macrophages repolarization and phagocytosis restoration for enhanced cancer immunotherapy. Article. J Nanobiotechnol. 2024;22(1):341. doi:10.1186/s12951-024-02622-1
109. Zhang C, Yin X, Hao L, et al. Integrin-targeted, activatable nanophototherapeutics for immune modulation: enhancing photoimmunotherapy efficacy in prostate cancer through macrophage reprogramming. Article; Early Access. Aggregate. 2025;6(5):E70001. doi:10.1002/agt2.70001
110. Li S, Zeng F, Zhou Q, et al. NIR-II photoacoustic imaging-guided chemo-photothermal therapy using PA1094T combined with Anti-CD47 antibody: activating pyroptosis against orthotopic glioblastoma. Article; Early Access. Adv Healthcare Mater. 2024:2403108. 10.1002/adhm.202403108.
111. Zhang Y, Ouyang Z, Zhan M, et al. An intelligent vascular disrupting dendritic nanodevice incorporating copper sulfide nanoparticles for immune modulation-mediated combination tumor therapy. Article. Small. 2023;19(39):2301914. doi:10.1002/smll.202301914
112. Liu Y-B, Chen X-Y, Yu B-X, et al. Chimeric peptide-engineered self-delivery nanomedicine for photodynamic-triggered breast cancer immunotherapy by macrophage polarization. Article. Small. 2024;20(22):2309994. doi:10.1002/smll.202309994
113. Zheng X, Liu Y, Liu Y, et al. Dual closed-loop of catalyzed lactate depletion and immune response to potentiate photothermal immunotherapy. Article. ACS Appl Mater Interfaces. 2022;14(20):23260–23276. doi:10.1021/acsami.2c07254
114. Zhang L, Cui S, Ding N, et al. Tumor-associated macrophages regulating a polymer nanoplatform for synergistic treatment of breast tumors. Article. ACS Appl Mater Interfaces. 2023;15(29):34527–34539. doi:10.1021/acsami.3c05497
115. Uthaman S, Pillarisetti S, Lim Y-M, et al. Light and immunostimulant mediated in situ re-education of tumor-associated macrophages using photosensitizer conjugated mannan nanoparticles for boosting immuno-photodynamic anti-metastasis therapy. Article. Biomater Sci. 2022;11(1):298–306. doi:10.1039/d2bm01508k
116. Chen S, He Y, Huang X, et al. Photosensitive and dual-targeted chromium nanoparticle delivering small interfering RNA YTHDF1 for molecular-targeted immunotherapy in liver cancer. Article. J Nanobiotechnol. 2024;22(1):348. doi:10.1186/s12951-024-02612-3
117. Zhang Y, Wang Q, Ji Y, et al. Mitochondrial targeted melanin@mSiO2 yolk-shell nanostructures for NIR-II-driven photo-thermal-dynamic/immunotherapy. Article. Chem Eng J. 2022;435:134869. doi:10.1016/j.cej.2022.134869
118. Xu C, Ding Y, Qi J, et al. Cascade-responsive nanoplatform for synergistic cancer therapy via ROS amplification and hypoxia targeting. Article. Chem Eng J. 2025;516:164112. doi:10.1016/j.cej.2025.164112
119. Li J, Ding Y-F, Wang Z, et al. TAM-preferential nanoparticles intracellularly self-assembled for enhanced macrophage repolarization and cancer immunotherapy. Article. Nano Today. 2024;54:102104. doi:10.1016/j.nantod.2023.102104
120. Qiu Z-W, Zhong Y-T, Lu Z-M, et al. Pyroptosis-inducing self-adaptor to potentiate immune checkpoint blockade therapy for breast cancer by reeducating the treatment-recruited macrophages. Article. Adv Funct Mater. 2025;35(7):2414847. doi:10.1002/adfm.202414847
121. Zhang X, Zhou W, Yu J, et al. Reprogramming tumor-associated macrophage by ornithine decarboxylase inhibitor and immune checkpoint for orthotopic glioblastoma photothermal immunotherapy. Article. ACS Appl Mater Interfaces. 2025;17(24):35155–35167. doi:10.1021/acsami.5c04588
122. Gu W, Guo W, Ren Z, et al. A bioactive nanocomposite integrated specific TAMs target and synergistic TAMs repolarization for effective cancer immunotherapy. Article. Bioact Mater. 2024;38:472–485. doi:10.1016/j.bioactmat.2024.04.029
123. Fu C, Tang L, Cao Y, et al. Bacterial outer membrane vesicles-clothed nanoparticles delivered by live macrophages for potentiating antitumor photoimmunotherapy. Article. Chem Eng J. 2024;499:156420. doi:10.1016/j.cej.2024.156420
124. Shang Y, Lu H, Liao L, Li S, Xiong H, Yao J. Bioengineered nanospores selectively blocking lc3-associated phagocytosis in tumor-associated macrophages potentiate antitumor immunity. Article. Acs Nano. 2023;17(11):10872–10887. doi:10.1021/acsnano.3c02657
125. Peng J, Zhou J, Sun R, et al. Dual-targeting of artesunate and chloroquine to tumor cells and tumor-associated macrophages by a biomimetic PLGA nanoparticle for colorectal cancer treatment. Article. Int J Biol Macromol. 2023;244:125163. doi:10.1016/j.ijbiomac.2023.125163
126. Zhang K, Long Y, Ma Z, Li S, Zhao Y, Han H. Artificial nanoplatelet regulation of tumor immune microenvironment to inhibit post- surgical tumor recurrence and lung g metastasis. Article. Mater Today. 2023;67:68–83. doi:10.1016/j.mattod.2023.05.024
127. Du Y, Qian X, Lin F, et al. A biomimetic nanoplatform for precise reprogramming of tumor-associated macrophages and NIR-II me diate d antitumor immune activation. Article. Acta Biomater. 2023;162:85–97. doi:10.1016/j.actbio.2023.03.021
128. Sheng S, Jin L, Zhang Y, et al. A twindrive precise delivery system of platelet-neutrophil hybrid membrane regulates macrophage combined with CD47 blocking for postoperative immunotherapy. Article. Acs Nano. 2024;18(6):4981–4992. doi:10.1021/acsnano.3c10862
129. Li J, Zhang Y, Zhu A, Zeng W, Di L, Wang R. Ginsenoside Rg3-engineered exosomes as effective delivery platform for potentiated chemotherapy and photoimmunotherapy of glioblastoma. Article. Chem Eng J. 2023;471:144692. doi:10.1016/j.cej.2023.144692
130. Sun Z, Sun Z, Liu J, et al. Engineered extracellular vesicles expressing siglec-10 camouflaged AIE photosensitizer to reprogram macrophages to active M1 phenotype and present tumor-associated antigens for photodynamic immunotherapy. Article. Small. 2024;20(12):2307147. doi:10.1002/smll.202307147
131. Wang T, Zhang H, Qiu W, Han Y, Liu H, Li Z. Biomimetic nanoparticles directly remodel immunosuppressive microenvironment for boosting glioblastoma immunotherapy. Article. Bioact Mater. 2022;16:418–432. doi:10.1016/j.bioactmat.2021.12.029
132. Zhen X, Li Y, Yuan W, et al. Biointerface-engineered hybrid nanovesicles for targeted reprogramming of tumor microenvironment. Article. Adv Mater. 2024;36(41):2401495. doi:10.1002/adma.202401495
133. Tang L, Yin Y, Cao Y, et al. Extracellular vesicles-derived hybrid nanoplatforms for amplified CD47 blockade-based cancer immunotherapy. Article. Adv Mater. 2023;35(35):2303835. doi:10.1002/adma.202303835
134. Zhou X, Ma E, Zhang Y, et al. NIR-actuated targeted janus nanomotors remodel immunosuppressive tumor microenvironment for augmented cancer immunotherapy. Article. Adv Healthcare Mater. 2024;13(2):2302272. doi:10.1002/adhm.202302272
135. Chen Y, Xu W, Tian H, et al. NIR-II light-actuated nanomotors for enhanced photoimmunotherapy toward hepatocellular carcinoma. Article. ACS Appl Mater Interfaces. 2024;16(30):39051–39063. doi:10.1021/acsami.4c06994
136. Peng H, Wang D, Huang S, Yu A. Dual-targeting aggregation-induced emission polymer micelles mediate immunogenic sonodynamic therapy for Tumor cell growth inhibition and macrophage reprogramming. Acta Biomater. 2025;195:321–337. doi:10.1016/j.actbio.2025.01.065
137. Wang Y, Li G, Su J, et al. Spatiotemporal controllable sono-nanovaccines driven by free-field based whole-body ultrasound for personalized cancer therapy. Article. Adv Sci. 2024;11(14):2307920. doi:10.1002/advs.202307920
138. Chen Y, Wu B, Shang H, et al. Sono-immunotherapy mediated controllable composite nano fluorescent probes reprogram the immune microenvironment of hepatocellular carcinoma. Article. Int J Nanomed. 2023;18:6059–6073. doi:10.2147/ijn.S426297
139. Tang Q, Zhang F, Luo L, et al. Ultrasound-induced gold nanoparticle united with acoustic reprogramming of macrophages for enhanced cancer therapy. Article. ACS Appl Mater Interfaces. 2023;15(44):50926–50939. doi:10.1021/acsami.3c12599
140. Gong J, Cheng D, Liu C, et al. Hybrid cell membrane-coated nanoparticles for synergizing sonodynamic therapy and immunotherapy against triple-negative breast cancer. Article. Adv Healthcare Mater. 2025;14(3):2404184. doi:10.1002/adhm.202404184
141. Huang S, Ding D, Lan T, et al. Multifunctional nanodrug performs sonodynamic therapy and inhibits TGF-β to boost immune response against colorectal cancer and liver metastasis. Article. Acta Biomater. 2023;164:538–552. doi:10.1016/j.actbio.2023.04.001
142. Hyun GH, Jeong D-H, Yang YY, et al. Multivalent carbohydrate nanocomposites for tumor microenvironment remodeling to enhance antitumor immunity. Article. Acs Nano. 2023;17(12):11567–11582. doi:10.1021/acsnano.3c01645
143. Wang C, Zhao J, Duan Y, et al. Tumor-associated myeloid cells selective delivery of a therapeutic tumor nano-vaccine for overcoming immune barriers for effective and long-term cancer immunotherapy. Article. Adv Healthcare Mater. 2024;13(26):2401416. doi:10.1002/adhm.202401416
144. Zhang N, Qian L, Xu C, et al. Innovative DNA tetrahedron inspired by ancient mortise-and-tenon technique offers new immunotherapy strategy for metastatic breast cancer. Article. Biomaterials. 2025;322:123390. doi:10.1016/j.biomaterials.2025.123390
145. Nguyen GH, Noh M, Kang JM, et al. Simultaneous targeting of tumor cells and tumor-associated macrophages to reprogram glioblastoma using trypsinized extracellular vesicles carrying tumor suppressive MicroRNA. Article. Nano Lett. 2025;25(20):8414–8422. doi:10.1021/acs.nanolett.5c01897
146. Zheng P, He J, Fu Y, et al. Engineered bacterial biomimetic vesicles reprogram tumor-associated macrophages and remodel tumor microenvironment to promote innate and adaptive antitumor immune responses. Article. Acs Nano. 2024;18(9):6863–6886. doi:10.1021/acsnano.3c06987
147. Xu H, Xiong S, Chen Y, et al. Flagella of tumor-targeting bacteria trigger local hemorrhage to reprogram tumor-associated macrophages for improved antitumor therapy. Article. Adv Mater. 2023;35(38):2303357. doi:10.1002/adma.202303357
148. Zhao Z, Chung YH, Steinmetz NF. Melanoma immunotherapy enabled by M2 macrophage targeted immunomodulatory cowpea mosaic virus. Article. Mater Adv. 2024;5(4):1473–1479. doi:10.1039/d3ma00820g
149. Kim H, Park H-J, Chang HW, et al. Exosome-guided direct reprogramming of tumor-associated macrophages from protumorigenic to antitumorigenic to fight cancer. Article. Bioact Mater. 2023;25:527–540. doi:10.1016/j.bioactmat.2022.07.021
150. Zhang Z, Hou L, Yu Z, et al. Biomimetic Small-Molecule Self-Assembly of PI3K inhibitor integrated with immunomodulator to amplify anticancer efficacy. Article. Chem Eng J. 2022;433:133747. doi:10.1016/j.cej.2021.133747
151. Jiang T, Wang B, Wang T, Zhang L, Chen X, Zhao X. TAM-hijacked immunoreaction rescued by hypoxia-pathway-intervened strategy for enhanced metastatic cancer immunotherapy. Article. Small. 2024;20(5):2305728. doi:10.1002/smll.202305728
152. Deng Y, Li J, Tao R, et al. Molecular engineering of electrosprayed hydrogel microspheres to achieve synergistic anti-tumor chemo-immunotherapy with ACEA cargo. Article. Adv Sci. 2024;11(17):2308051. doi:10.1002/advs.202308051
153. Zheng Q, Zou T, Wang W, et al. Necroptosis-mediated synergistic photodynamic and glutamine-metabolic therapy enabled by a biomimetic targeting nanosystem for cholangiocarcinoma. Article. Adv Sci. 2024;11(29):2309203. doi:10.1002/advs.202309203
154. Tang B, Zhu J, Wang Y, et al. Targeted xCT-mediated ferroptosis and protumoral polarization of macrophages is effective against HCC and enhances the efficacy of the Anti-PD-1/L1 response. Article. Adv Sci. 2023;10(2):2203973. doi:10.1002/advs.202203973
155. He X, Deng T, Li J, et al. A core-satellite micellar system against primary tumors and their lymphatic metastasis through modulation of fatty acid metabolism blockade and tumor-associated macrophages. Article. Nanoscale. 2023;15(18):8320–8336. doi:10.1039/d2nr04693h
156. Stater EP, Morcos G, Isaac E, et al. Translatable drug-loaded iron oxide nanophore sensitizes murine melanoma tumors to monoclonal antibody immunotherapy. Article. Acs Nano. 2023;17(7):6178–6192. doi:10.1021/acsnano.2c05800
157. Xu S, Gu Z, Lu H, Guan P, Liu Z. Leveraging macrophage-mediated cancer immunotherapy via a cascading effect induced by a molecularly imprinted nanocoordinator. Article. ACS Appl Mater Interfaces. 2023;15(23):27658–27669. doi:10.1021/acsami.3c03950
158. Zou H, Li M, Li X, et al. Multimodal imaging and photothermal synergistic immunotherapy of retinoblastoma with tuftsin-loaded carbonized MOF nanoparticles. Article. Drug Delivery. 2022;29(1):1785–1799. doi:10.1080/10717544.2022.2081379
159. Dehghankelishadi P, Badiee P, Maritz MF, Dmochowska N, Thierry B. Bosutinib high density lipoprotein nanoformulation has potent tumour radiosensitisation effects. Article. J Nanobiotechnol. 2023;21(1):102. doi:10.1186/s12951-023-01848-9
160. Wang Y, Yang L, Yan C, et al. Supramolecular artificial Nano-AUTACs enable tumor-specific metabolism protein degradation for synergistic immunotherapy. Article. Sci Adv. 2024;10(25):EADN8079. doi:10.1126/sciadv.adn8079
161. Zhu R, Yuan W, Xia A, et al. Inulin-based nanoparticle modulates gut microbiota and immune microenvironment for improving colorectal cancer therapy. Article. Adv Funct Mater. 2024;34(46):2407685. doi:10.1002/adfm.202407685
162. Fredrich IF, Halabi EA, Kohler RH, Ge X, Garris CS, Weissleder R. Highly active myeloid therapy for cancer. Article. Acs Nano. 2023;17(20):20666–20679. doi:10.1021/acsnano.3c08034
163. Chen C, Yu Q, Wang L, et al. Boric acid functionalized carbon nanotheranostic platform for the hierarchical delivery of immune adjuvants and chemotherapy agents. Article. J Drug Delivery Sci Technol. 2025;109:106993. doi:10.1016/j.jddst.2025.106993
164. Wen D, Liang T, Chen G, et al. Adipocytes encapsulating telratolimod recruit and polarize tumor-associated macrophages for cancer immunotherapy. Article. Adv Sci. 2023;10(5):2206001. doi:10.1002/advs.202206001
165. Song W, Song S-J, Kuang J, et al. Activating innate immunity by a sting signal amplifier for local and systemic immunotherapy. Article. Acs Nano. 2022;16(10):15977–15993. doi:10.1021/acsnano.2c03509
166. Yuan H, Qiu C, Wang X, et al. Engineering semiconducting polymeric nanoagonists potentiate cGAS-STING pathway activation and elicit long term memory against recurrence in breast cancer. Article. Adv Mater. 2025;37(4):2406662. doi:10.1002/adma.202406662
167. Wang Y, Xu H, Wang D, et al. Synergistic reinforcement of immunogenic cell death and transformation of tumor-associated macrophages via an M1-type macrophage membrane-camouflaged ferrous-supply-regeneration nanoplatform. Article. Acta Biomater. 2024;174:358–371. doi:10.1016/j.actbio.2023.11.041
168. Guo P, Dai P, Yang S, et al. Engineered macrophages tune intratumoral cytokines through precisely controlled self-pyroptosis to enhance bladder cancer immunotherapy. Article. Small. 2024;20(13):2306699. doi:10.1002/smll.202306699
169. Xu S, Meng L, Hu Q, et al. Closed-loop control of macrophage engineering enabled by focused-ultrasound responsive mechanoluminescence nanoplatform for precise cancer immunotherapy. Article. Small. 2024;20(46):2401398. doi:10.1002/smll.202401398
170. Enbergs N, Halabi EA, Goubet A-G, et al. Pharmacological polarization of tumor-associated macrophages toward a CXCL9 antitumor phenotype. Article. Adv Sci. 2024;11(15):2309026. doi:10.1002/advs.202309026
171. Horvat NK, Chocarro S, Marques O, et al. Superparamagnetic iron oxide nanoparticles reprogram the tumor microenvironment and reduce lung cancer regrowth after crizotinib treatment. Article. Acs Nano. 2024;18(17):11025–11041. doi:10.1021/acsnano.3c08335
172. Zhang T, Zhang L, Huang X, et al. Dual nanoparticle immunostimulant hydrogel for synergistic combating “Cold” tumor. Article. Chem Eng J. 2024;490:151663. doi:10.1016/j.cej.2024.151663
173. Mahmoud YS, Hassanin IA, Sabra SA, et al. Lipopolysaccharide nanomedicine-based reversion of chemotherapy-induced metastatic potential of breast cancer via hampering tumoral TLR4/SIRT2 axis and IL6 secretion from tumor-associated macrophages. Article. ISPRS J Photogram Remote Sens. 2025;222:141396. doi:10.1016/j.ijbiomac.2025.141396
174. Yang K, Han W, Jiang X, et al. Zinc cyclic di-AMP nanoparticles target and suppress tumours via endothelial STING activation and tumour-associated macrophage reinvigoration. Article. Nature Nanotechnol. 2022;17(12):1322. doi:10.1038/s41565-022-01225-x
175. Negrea G, Rauca V-F, Meszaros MS, et al. Active tumor-targeting nano-formulations containing simvastatin and doxorubicin inhibit melanoma growth and angiogenesis. Article. Front Pharmacol. 2022;13:870347. doi:10.3389/fphar.2022.870347
176. Wang B, Cheng H, Ji Z, et al. Synergistic target-attacking tumor cells and M2 macrophages via a triple-responsive nanoassembly for complete metastasis blocking. Article; Early Access. Adv Healthcare Mater. 2024:2304096. 10.1002/adhm.202304096.
177. Jaggarapu MMCS, Ansari A, Jinka S, et al. Surface modified glucose-derived, blood-brain barrier-crossing nanospheres dually targets macrophage and cancer cells for effective in situ anti-glioma effect. Article. Adv Ther. 2024;7(9):2400100. doi:10.1002/adtp.202400100
178. Sun W, Li Y, Sui D, et al. A potential platform of combining sialic acid derivative-modified paclitaxel cationic liposomes with antibody-drug conjugates inspires robust tumor-specific immunological memory in solid tumors. Article. Biomater Sci. 2023;11(8):2787–2808. doi:10.1039/d2bm01769e
179. Qi Z, Yang Q, Wang Y, Song Y, Deng Y. Vicious-cycle-breaking antiangiogenic nano-delivery systems potentiate and simplify the tumor vascular normalization strategy. Article. Int J Pharm. 2025;681:125788. doi:10.1016/j.ijpharm.2025.125788
180. Choi TH, Yoo RJ, Park JY, et al. Development of finely tuned liposome nanoplatform for macrophage depletion. Article. J Nanobiotechnol. 2024;22(1):83. doi:10.1186/s12951-024-02325-7
181. Cao Y, Qiao B, Chen Q, et al. Tumor microenvironment remodeling via targeted depletion of M2-like tumor-associated macrophages for cancer immunotherapy. Article. Acta Biomater. 2023;160:239–251. doi:10.1016/j.actbio.2023.02.006
182. Yao W, Xu H, Chen Y, et al. Mannose modified co-loaded zoledronic liposomes deplete M2-tumor-associated macrophages to enhance anti-tumor effect of doxorubicin on TNBC. Article. J Drug Delivery Sci Technol. 2022;74:103551. doi:10.1016/j.jddst.2022.103551
183. Feng J, Xiang L, Fang C, et al. Dual-targeting of tumor cells and tumor-associated macrophages by palmitic acid modified albumin nanoparticles for antitumor and antimetastasis therapy. Article. ACS Appl Mater Interfaces. 2022;14(13):14887–14902. doi:10.1021/acsami.1c23274
184. Zhang X, Guo Q, Zhao Z, Cheng P, Wu A, Liu H. Engineering prodrug nanoparticles for targeted therapy in heterogeneous glioblastoma. Article. Chem Eng J. 2023;474:145557. doi:10.1016/j.cej.2023.145557
185. Zheng C, Zhao X, Wang Y, et al. Immune modulating nanoparticles depleting tumor-associated macrophages to enhance immune checkpoint blockade therapy. Article. Chem Eng J. 2022;435:134779. doi:10.1016/j.cej.2022.134779
186. Zhang H-T, Peng R, Chen S, et al. Versatile Nano-PROTAC-induced epigenetic reader degradation for efficient lung cancer therapy. Article. Adv Sci. 2022;9(29):2202039. doi:10.1002/advs.202202039
187. Zhang J, Sun X, Xu M, et al. A self-amplifying ROS-sensitive prodrug-based nanodecoy for circumventing immune resistance in chemotherapy-sensitized immunotherapy. Article. Acta Biomater. 2022;149:307–320. doi:10.1016/j.actbio.2022.06.035
188. Cheng H-W, Lee W, Hsu F-T, et al. Manipulating the crosstalk between cancer and immunosuppressive cells with phototherapeutic gold-nanohut for reprogramming tumor microenvironment. Article. Adv Sci. 2024;11(32):2404347. doi:10.1002/advs.202404347
189. Park K, Ahn JW, Kim JH, Kim JW. Tumor-associated macrophage-targeted photodynamic cancer therapy using a dextran sulfate-based nano-photosensitizer. Article. Int J Biol Macromol. 2022;218:384–393. doi:10.1016/j.ijbiomac.2022.07.159
190. Lin Y, Yi M, Guan X, et al. “Two birds with one stone” strategy for the lung cancer therapy with bioinspired AIE aggregates. Article. J Nanobiotechnol. 2023;21(1):49. doi:10.1186/s12951-023-01799-1
191. Yu C, Hu L, Yu Q, et al. In vivo self-assembled albumin nanoparticle elicit antitumor immunity of PD-1 inhibitor by imaging and clearing tumor-associated macrophages. Article. Front Chem. 2024;12:1469568. doi:10.3389/fchem.2024.1469568
192. Nie C, Ma T, Ye J, et al. A STING agonist-loaded bispecific nanobioconjugate modulates macrophage immune responses to enhance antitumor immunotherapy. Article. Chem Eng J. 2024;485:149901. doi:10.1016/j.cej.2024.149901
193. Liang J, Zhu F, Cheng K, et al. Outer membrane vesicle-based nanohybrids target tumor-associated macrophages to enhance trained immunity-related vaccine-generated antitumor activity. Article. Adv Mater. 2023;35(46):2306158. doi:10.1002/adma.202306158
194. Li X, Guo Q, Chen Q, et al. Reconciling the cooperative-competitive patterns among tumor and immune cells for triple-negative breast cancer treatment using multimodule nanocomplexes. Article. Adv Mater. 2024;36(26):2312219. doi:10.1002/adma.202312219
195. Feng Q, Ma X, Cheng K, et al. Engineered bacterial outer membrane vesicles as controllable two-way adaptors to activate macrophage phagocytosis for improved tumor immunotherapy. Article. Adv Mater. 2022;34(40):2206200. doi:10.1002/adma.202206200
196. Ji X, Qian X, Luo G, et al. Engineered macrophage nanoparticles enhance microwave ablation efficacy in osteosarcoma via targeting the CD47-SIRPα Axis: a novel Biomimetic immunotherapeutic approach. Article. Bioact Mater. 2025;47:248–265. doi:10.1016/j.bioactmat.2025.01.012
197. Moon S, Jung M, Go S, et al. Engineered nanoparticles for enhanced antitumoral synergy between macrophages and T cells in the tumor microenvironment. Article. Adv Mater. 2024;36(44):2410340. doi:10.1002/adma.202410340
198. Wang F, Huang Q, Su H, et al. Self-assembling paclitaxel-mediated stimulation of tumor-associated macrophages for postoperative treatment of glioblastoma. Article. Proc Natl Acad Sci USA. 2023;120(18):e2204621120. doi:10.1073/pnas.2204621120
199. Gao F, Dong J-H, Xue C, et al. Tumor-targeting multiple metabolic regulations for bursting antitumor efficacy of chemodynamic therapy. Article. Small. 2024;20(26):2310248. doi:10.1002/smll.202310248
200. Xiao Z, Cai Y, Wang X, et al. Nanodrug simultaneously regulates stromal extracellular matrix and glucose metabolism for effective immunotherapy against orthotopic pancreatic cancer. Article. Nano Today. 2022;44:101490. doi:10.1016/j.nantod.2022.101490
201. Shen W, Liu T, Pei P, et al. Metabolic homeostasis-regulated nanoparticles for antibody-independent cancer radio-immunotherapy. Article. Adv Mater. 2022;34(51):2207343. doi:10.1002/adma.202207343
202. Liang H, Zhou L, Hu Z, et al. Siglec15 checkpoint blockade for simultaneous immunochemotherapy and osteolysis inhibition in lung adenocarcinoma spinal metastasis via a hollow nanoplatform. Article. Small. 2022;18(29):2107787. doi:10.1002/smll.202107787
203. Liyanage W, Wu T, Kannan S, Kannan RM. Dendrimer-siRNA conjugates for targeted intracellular delivery in glioblastoma animal models. Article; Early Access. ACS Appl Mater Interfaces. 2022;46290–46303. doi:10.1021/acsami.2c13129
204. Qiao G, Li S, Pan X, et al. Surgical tumor-derived nanoplatform targets tumor-associated macrophage for personalized postsurgical cancer immunotherapy. Article. Sci Adv. 2024;10(13):EADK7955. doi:10.1126/sciadv.adk7955
205. Kim J, Zhu Y, Chen S, et al. Anti-glioma effect of ginseng-derived exosomes-like nanoparticles by active blood-brain-barrier penetration and tumor microenvironment modulation. Article. J Nanobiotechnol. 2023;21(1):253. doi:10.1186/s12951-023-02006-x
206. Wu H, Huang X, Xu H, et al. Bupivacaine nanoparticles inhibit triple-negative breast tumor growth by suppressing the noradrenergic nerves in tumor microenvironment. Article. Int J Nanomed. 2025;20:6023–6041. doi:10.2147/ijn.S515895
207. Bao Y, Li G, Li S, et al. Multifunctional tumor-targeting carbon dots for tumor microenvironment activated ferroptosis and immunotherapy in cancer treatment. Article. ACS Appl Mater Interfaces. 2023;15(49):56834–56845. doi:10.1021/acsami.3c13867
208. Xu R, Huang L, Liu J, et al. Remodeling of mitochondrial metabolism by a mitochondria-targeted RNAi nanoplatform for effective cancer therapy. Article. Small. 2024;20(10):2305923. doi:10.1002/smll.202305923
209. Cen Y, Chen Y, Li X, et al. Optical controlled and nuclear targeted CECR2 competitor to downregulate CSF-1 for metastatic breast cancer immunotherapy. Article. Biomaterials. 2024:308122568. 10.1016/j.biomaterials.2024.122568.
210. Lu Y, Wang Y, Liu W, et al. Photothermal ?nano-dot? reactivate ?immune-hot? for tumor treatment via reprogramming cancer cells metabolism. Article. Biomaterials. 2023:296122089. 10.1016/j.biomaterials.2023.122089.
211. Wei Y, Wang Z, Yang J, et al. Reactive oxygen species/photothermal therapy dual-triggered biomimetic gold nanocages nanoplatform for combination cancer therapy via ferroptosis and tumor-associated macrophage repolarization mechanism. Article. J Colloid Interface Sci. 2022;606:1950–1965. doi:10.1016/j.jcis.2021.09.160
212. Zhang M, Shao W, Yang T, et al. Conscription of immune cells by light-activatable silencing NK-derived exosome (LASNEO) for synergetic tumor eradication. Article. Adv Sci. 2022;9(22):2201135. doi:10.1002/advs.202201135
213. Jiang H, Fu H, Guo Y, Hu P, Shi J. Evoking tumor associated macrophages by mitochondria-targeted magnetothermal immunogenic cell death for cancer immunotherapy. Article. Biomaterials. 2022;289121799. doi:10.1016/j.biomaterials.2022.121799
214. Fang C, Peng QX, Li XY, et al. Multichannel immune nanoregulators suppress lactic acid metabolism and lactic acid-shaped acidic microenvironment to uproot anti-tumor immunosuppression. Adv Mater. 2026;38(3). doi:10.1002/adma.202512230
215. Li Y, Wei Y, Ma S, Li F, Meng X, Yu S. A dual-transformable MgGa-MOF nanoplatform for HCC therapy via lactate metabolism blockade and immune reactivation. J Nanobiotechnol. 2026. doi:10.1186/s12951-026-04356-8
216. Song KC, Hao YM, Lei P, et al. In situ needle-free injection of multiretention micelles for melanoma therapy with multiomics insights into tumor targeting and immune modulation. Acs Nano. 2026;20(11):9539–9559. doi:10.1021/acsnano.6c01481
217. Feng CY, Yang ZY, Lan YF, et al. STAR Therapy: spatiotemporal antitumor regimen via a size-gated DNA hydrogel drug delivery system. Adv Mater. 2026;38(14). doi:10.1002/adma.202517487
218. Mukalel AJ, Tylek T, O’Brien E, et al. Bioinspired oxidized mRNA lipid nanoparticles for ex vivo engineering of chimeric antigen receptor macrophages targeting solid tumors. Bioeng Transl Med. 2026;2026. doi:10.1002/btm2.70138
219. Kong CY, Luo CW, Xue FX, et al. Ultrasound-triggered charge-reversal nanoparticles via golgi-dependent iterative transcytosis for enhanced deep tumor penetration. Acs Nano. 2026;20(6):4741–4757. doi:10.1021/acsnano.5c14557
220. Zhang H, Miao J, Gao L, et al. Co-delivering macrophage engager mRNA and PD-L1 antibody via tumor-responsive nanoparticles for glioblastoma immunotherapy. Nat Commun. 2026. doi:10.1038/s41467-026-71646-y
221. Yin QM, Liang X, Zhang CC, et al. Lipid nanoparticle-mediated in vivo generation of panCAR immune cells for solid tumor immunotherapy. Proc Natl Acad Sci USA. 2026;123(8):2509698123. doi:10.1073/pnas.2509698123
222. Wagner AJ, Tap WD, Bauer S, et al. Long-term efficacy and safety of pexidartinib in patients with tenosynovial giant cell tumor: final results of the ENLIVEN study. Oncologist. 2025;30(7):oyae345. doi:10.1093/oncolo/oyae345
223. Xiao AN, Mei MT. Selection of a salvage regimen for relapsed or refractory classic hodgkin lymphoma in the era of novel therapies. Hematol Oncol Clin North Am. 2026;40(2):275–288. doi:10.1016/j.hoc.2025.12.004
224. Matsuda K, Ota Y, Uemachi H, et al. Examination of the potential clinical application of 5DEX-0509R, the tumor macrophage-targeting nanomedicine. Cytokine. 2025;186:156842. doi:10.1016/j.cyto.2024.156842
225. Wang MN, Zhao JZ, Xiong HJ, Lu HB, Jiang H, Wang XM. Advance of nano anticancer therapies targeted on tumor-associated macrophages. Coord Chem Rev. 2021;446:214126. doi:10.1016/j.ccr.2021.214126
226. Huang H, Tong QS, Zhang JY, et al. Phagocytosis-activating nanocomplex orchestrates macrophage-mediated cancer immunotherapy. Adv Mater. 2025;37(28). doi:10.1002/adma.202500982
227. Cheng YF, Zhu KN, Yang L, et al. A self-homing liposomal nanobot for active therapy of glioblastoma. Angew Chem Int Ed. 2025;2025:e202512948. doi:10.1002/anie.202512948
228. Luo TY, Zhang MY, Xu J, et al. Jahn-Teller distortion-engineered self-propelled nanorobots for mitochondrial targeting and bioenergetic disruption in tumor therapy. Bioact Mater. 2025;54:273–290. doi:10.1016/j.bioactmat.2025.08.021
229. Hegde S, Giotti B, Soong BY, et al. Myeloid progenitor dysregulation fuels immunosuppressive macrophages in tumours. Nature. 2025;646(8087):1214–1222. doi:10.1038/s41586-025-09493-y
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Combined Photodynamic and Photothermal Therapy and Immunotherapy for Cancer Treatment: A Review
Kong C, Chen X
International Journal of Nanomedicine 2022, 17:6427-6446
Published Date: 16 December 2022
Ability of Blood Cell Parameters to Predict Clinical Outcomes of Nivolumab Monotherapy in Advanced Esophageal Squamous Cell Carcinoma
Hamai Y, Emi M, Ibuki Y, Kurokawa T, Yoshikawa T, Ohsawa M, Hirohata R, Kitasaki N, Okada M
OncoTargets and Therapy 2023, 16:263-273
Published Date: 10 April 2023
Cancer Immunotherapy and Medical Imaging Research Trends from 2003 to 2023: A Bibliometric Analysis

Tang S, Fan T, Wang X, Yu C, Zhang C, Zhou Y
Journal of Multidisciplinary Healthcare 2024, 17:2105-2120
Published Date: 6 May 2024
Impact of Caspase3/GSDME-Mediated Pyroptosis on Tumor Immune Microenvironment and Clinical Prognosis Across Multiple Cancers
Huang Y, Liu J, Lin C, Zhu Q, Wu L
Cancer Management and Research 2024, 16:1663-1683
Published Date: 26 November 2024
Artificial Intelligence–Enabled Multi-Omics for Predicting Immune Checkpoint Inhibitor Response and Resistance
Wang X, He J, Ding G, Tang Y, Wang Q
Journal of Multidisciplinary Healthcare 2026, 19:572089
Published Date: 26 February 2026