Back to Journals » Drug Design, Development and Therapy » Volume 19
Molecular Insights into Phytochemicals Mediated Epigenetic Regulation in Preclinical Models of Breast Cancer
Authors Pandey P, Alkhathami AG, Saeed M ![]() , Alshaghdali K, Kumar S, Tallei TE, Bae H, Park MN, Kumar G, Kim B, Khan F
, Alshaghdali K, Kumar S, Tallei TE, Bae H, Park MN, Kumar G, Kim B, Khan F ![]()
Received 19 July 2025
Accepted for publication 1 November 2025
Published 27 November 2025 Volume 2025:19 Pages 10589—10606
DOI https://doi.org/10.2147/DDDT.S554844
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Dr Solomon Tadesse Zeleke
Pratibha Pandey,1 Ali G Alkhathami,2 Mohd Saeed,3 Khalid Alshaghdali,4 Sandeep Kumar,5 Trina Ekawati Tallei,6,7 Hanul Bae,8 Moon Nyeo Park,8 Gaurav Kumar,5 Bonglee Kim,8 Fahad Khan9
1Centre for Research Impact and Outcome, Chitkara University, Rajpura, Punjab, India; 2Department of Clinical Laboratory Sciences, College of Applied Medical Sciences, King Khalid University, Abha, Saudi Arabia; 3Department of Biology, College of Science, University of Hail, Hail, Saudi Arabia; 4Department of Clinical Laboratory Sciences, College of Applied Medical Sciences, University of Hail, Hail, Saudi Arabia; 5Research and Innovation Cell, Bahra University, Waknaghat, HP, India; 6Department of Biology, Faculty of Mathematics and Natural Sciences, Universitas Sam Ratulangi, Manado, Indonesia; 7Department of Biology, Faculty of Medicine, Universitas Sam Ratulangi, Manado, Indonesia; 8Department of Pathology, College of Korean Medicine, Kyung Hee University, Seoul, Republic of Korea; 9Center for Global Health Research, Saveetha Medical College and Hospitals, Saveetha Institute of Medical and Technical Sciences, Chennai, TN-602105, India
Correspondence: Fahad Khan, Email [email protected] Bonglee Kim, Email [email protected]
Abstract: One of the most common malignant tumors in women worldwide is breast cancer, which affects even more than one-third of all female tumor patients. Patient outcomes and effective therapeutic strategies are frequently determined by molecular subtypes in breast cancer. However, the underlying epigenetic characteristics that could further divide breast cancer patients into groups and affect their outcomes could be the reason for the differences in therapeutic response. It is true that there have been recent findings about the role of epigenetic abnormalities in cancer, and that therapeutics targeting particular epigenetic pathways have been developed. Phytochemicals function as gene regulators in a variety of cancers and are crucial to the pathophysiology of many human cancers, including breast cancer. Preclinical studies have revealed that phytochemicals exhibit promising therapeutic efficacy against breast carcinoma by modulating several epigenetic alterations including DNA methylation, histone modifications, non-coding RNA and estrogen associated epigenetic changes. Nevertheless, despite promising in vitro and in vivo results, the clinical application of phytochemicals targeting epigenetic markers in breast cancer is limited. Further research is required to confirm their effectiveness and safety in clinical settings. Thus, this study provides a thorough summary of how epigenetic changes contribute to the development of breast cancer. This article also explores the potential benefits of phytochemicals, such as flavonoids, terpenoids, alkaloids, isothiocyanates, and quinones, in modulating these epigenetic markers in preclinical models of breast cancer.
Keywords: breast cancer, phytochemicals, epigenetic regulation, cancer therapy
Introduction
Breast cancer continues to be a tough foe of global health challenges. Complex pathophysiology and extensive range of clinical manifestations of breast cancer present significant hurdles for the development of promising treatment and prevention strategies.1,2 Breast cancer can be categorized into three principal subtypes with respect to receptor expression status including luminal A, luminal B, HER2-enriched, and triple-negative variants.3 Various breast cancer subtypes display unique biological characteristics, resulting in differences in prognosis and therapeutic efficacy.4,5
When classifying breast tumors into various molecular subgroups, the expression levels of HER2, progesterone, and estrogen play crucial roles. However, it is essential to realize that genetic variants play a substantial role in the progression of the disease, as well as the therapeutic response.6,7 Numerous studies have shown that epigenetics, apart from genomic aberrations, is a significant factor in breast cancer development. These studies focused on the initiation of molecular pathways in carcinogenesis, prediction of biomarkers for breast cancer aggressiveness and remarkable possibilities of epigenetics therapy.8,9 Recently, it has become widely recognized that abnormal gene expression, breast cancer tumorigenesis, and metastasis are linked to epigenetic dysregulation, including deregulated histone modification and DNA methylation.10,11
Phytochemicals are considered as a promising therapeutic approach that can be used to decrease side-effects related to conventional methods and increase the therapeutic efficacy for breast cancer by inhibiting disease progression. These phytochemicals have been used for decades to cure a wide range of disorders, including cancer, by modulating several cellular and molecular targets.12 In fact, a number of compounds that influence epigenetic pathways have been marketed as anticancer medications under the name “epidrugs”.13 One of the main sources of epidrugs is nature, which has a limitless supply of bioactive natural compounds that precisely target epigenetic pathways and have exceptional anticancer properties. In fact, natural compounds found in medicinal plants have demonstrated epidrug activity.14–16 Phytochemicals have been identified in a number of studies as possible regulators that can restore abnormal epigenetic changes that cause tumor growth and ultimately cancer.17,18 In this regard, many secondary metabolites that have been isolated from plants, such as alkaloids, quinones, terpenoids, and flavonoids, have demonstrated chemopreventive benefits by reversing epigenetic changes and modifying the molecular pathways linked to carcinogenesis.19–21 Phytocompounds have multiple uses and may hold great promise as a treatment for breast cancer due to their ability to target different cellular pathways that are involved in the disease’s epigenetics.22–24 Several promising phytochemicals suffer from low bioavailability and limited absorption rates. Additionally, there is a lack of reliable clinical studies validating the effects of phytochemicals on epigenetic markers in breast cancer patients. Precise mechanistic studies and comprehensive reviews of the epigenetic modulation of breast cancer by phytochemicals are limited.25 Thus, this study focused on the possible applications of specific phytochemicals in anticancer therapies, as well as their ability to combat epigenetic aberrations that support the initiation and progression of breast carcinoma in humans. As a result, a detailed description of the most studied phytochemicals that have been shown to significant modulate the epigenetic landscape in breast cancer, along with their limitations, is provided in detail.
Breast Cancer and Epigenetic Regulation
The development and advancement of cancers are significantly influenced by epigenetic changes including dysregulation of the ncRNAs, histone modifications, localized hypermethylation of CpG promoters of tumor suppressor genes, and loss of DNA methylation throughout the genome. Numerous forms of cancer, including glioblastoma, leukemia, liver, lung, prostate, breast, and gastric cancers, have been linked to abnormal DNA methylation patterns.26–32 In addition to genetic abnormalities, various epigenetic changes play a role in the development and progression of breast carcinoma. The main epigenetic alterations directly associated with breast cancer are DNA methylation, histone modifications, nucleosome rearrangement and non-coding RNAs regulation. These alterations may serve as markers for prognosis, early identification and therapy effectiveness in breast carcinoma.33–35 Additionally, a thorough understanding of underlying epigenetic regulatory markers of breast carcinoma could open up new avenues for drug development. Altogether, epigenetic abnormalities linked to the onset and spread of breast cancer significantly affect the genes that govern cell proliferation, invasion, motility, and apoptosis (Figure 1).36,37
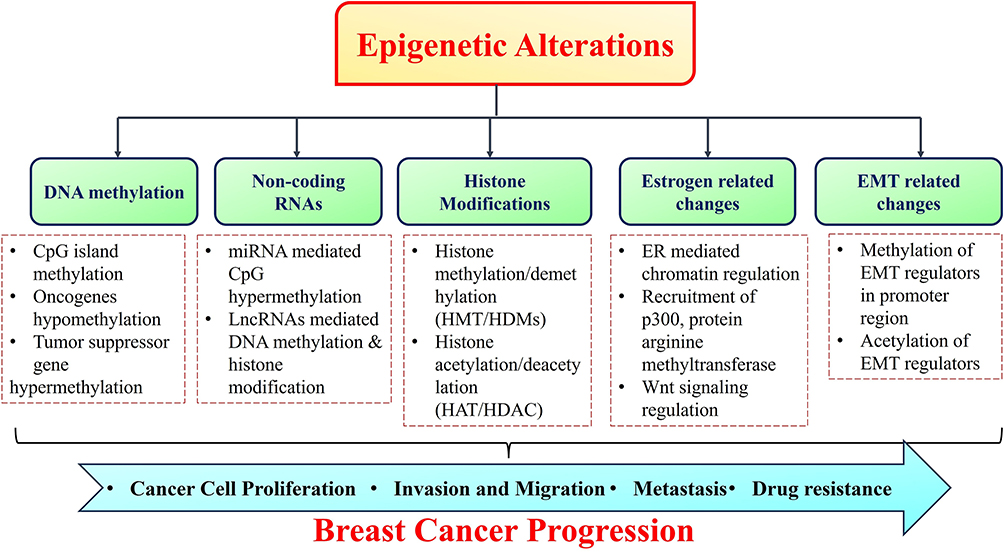 |
Figure 1 Illustration of epigenetic alterations associated with breast cancer progression, metastasis and drug resistance. |
DNA Methylation
DNA methylation, a prominent epigenetic process, involves the covalent linkage of a methyl group (CH3) to the 5′-site of cytosine preceding guanine in the DNA structure. Methylation inside CpG dinucleotides, which are densely clustered in regions known as CpG islands, affects gene expression and, consequently, influences the primary biological processes associated with cancer.38,39 Methylation results in the formation of a 5-methylcytosine (5mC) structure, which can obstruct transcription factors from retrieving DNA-binding locations or engage methyl-binding domain proteins (MBDs) alongside histone protein modifications, thereby inhibiting the expression of methylated genes. In this situation, extensive methylation of promoters of crucial tumor suppressor genes results in their suppression, whereas reduced methylation of oncogenes leads to their abnormal activation.40,41
DNA methylation is a reversible mechanism regulated by a specific class of enzymes, known as DNA methyltransferases (DNMTs). Three DNA methyltransferases are actively involved in this process: DNMT1, DNMT3a, and DNMT3b. DNA demethylation is facilitated by a family of enzymes known as ten-eleven translocation methylcytosine dioxygenases (TETs), which convert 5-methylcytosine (5mC) to 5-hydroxymethylcytosine (5-hmC) via hydroxymethylation. Three such enzymes, TET1, TET2, and TET3, are engaged in DNA demethylation, which restores genes that have been silenced as a result of DNMTs. This process collectively affects the transcriptional activation of critical genes associated with tumorigenesis and genomic stability.42–44 Numerous additional proteins exhibiting DNA demethylase activities associated with breast carcinoma such as growth arrest and DNA-damage-inducible protein (GADD45) and the cytidine deaminase family, specifically activation-induced cytidine deaminase (AID) and Apolipoprotein B mRNA editing catalytic polypeptide-like family (APOBEC). GADD45A is significantly correlated with DNA repair and epigenetic control genes.45 The relationship between GADD45 and the BRCA1 gene in breast cancer is proposed to affect the etiology of the disease, likely by activating nucleotide excision repair pathways. GADD45A exhibits aberrant methylation in breast cancer.46 AID proteins play crucial roles in active DNA demethylation, specifically the deamination of 5-mC to thymine. AID is known for its role in promoting DNA demethylation and is crucial for epithelial-mesenchymal transition (EMT) in mammary epithelial cells (non-transformed).47 Moreover, although APOBEC1 exhibits DNA demethylase activity,48–50 it has been shown that APOBEC mutagenesis affect tumor progression in ER+/HER2-positive breast carcinoma.51 Recent findings have indicated that APOBEC mutagenesis inhibits breast cancer growth by triggering immunogenic responses.52
Numerous genes associated with breast cancer demonstrate CpG island hypermethylation, and in several cases, aberrant activity of DNA methyltransferases results in the hypermethylation and silencing of tumor suppressor genes like HOXA5, RASSF1A, TMS1, p16, and BRCA1.53,54 Moreover, genes repressed by promoter hypermethylation include E-cadherin, GSTP1, TMS1, and p16.55–57 Some of the important biological functions of these genes include estrogen signaling, pro-apoptosis (HOXA5 and TMS1), cell cycle progression (RASSF1A and p16), and DNA repair pathways (BRCA1). BRCA1, a prominent breast cancer susceptibility gene often suppressed in sporadic breast cancers, has been related to CpG hypermethylation linked to DNMT3b upregulation.58 The initial phases of sporadic breast cancer demonstrate the loss of the cell cycle checkpoint gene p16INK4a due to abnormal promoter methylation of CpG, and approximately 80% of breast tumors show downregulation of the CDK inhibitor p21, due to its hypermethylation.59,60
Non-Coding RNAs
Several epigenetic alterations are associated with breast cancer. Non-coding RNAs, especially microRNAs (miRNAs), play a role in the post-transcriptional control of breast cancer development, progression, and metastasis.61 miRNAs, non-coding RNA of 17–25 nucleotides, can modulate gene expression in both healthy and cancerous cells. Polymerase II facilitates transcription of primary miRNAs. Primary miRNAs are converted into pre-miRNAs by the Drosha–Dicer complex, which enables mature versions of miRNAs to control post-transcriptional levels of gene expression. In cancer, miRNAs improperly control genes due to CpG hypermethylation in miRNA genes or deregulation of miRNA manufacturing mechanisms.62 Aberrant miRNA mechanisms have been implicated in all phases of cancer, from carcinogenesis to metastasis in breast carcinoma. For instance, miRNA profiling has revealed that tumor suppressor miRNAs, such as miR-4458 linked to SOC1 signaling, are downregulated, while the oncogenic miRNA miR-214, involved in PI3K/Akt/mTOR signaling, is elevated in breast tumors.63 Dysregulated expression of these miRNAs is responsible for the emergence of cancer characteristics such as cell proliferation, hypoxia, metastasis, apoptosis, and angiogenesis.64 Breast tumor is represented by the presence of both miRNAs and long noncoding RNAs. LncRNAs, which are non-coding RNAs measuring between 200 nucleotides and 100 kilobases, can inhibit target genes by facilitating DNA methylation or histone modification. The development of tumors can result from dysregulation of these mechanisms.65,66 The lncRNA GAS5 functions as a tumor suppressor by modulating several tumor suppressor proteins, including PTEN, PDCD4, FOXO1, OKK2, and SUFU, and is significantly decreased in breast cancer. Moreover, GAS5 expression is inhibited by promoter methylation in triple-negative breast cancer, indicating that lncRNAs are pivotal in the pathogenesis of breast cancer.67
Histone Modifications
Histones function as crucial proteins for DNA packaging, thus preserving the chromatin architecture. Histone proteins (H2A, H2B, H3, and H4) facilitate the formation of nucleosomes by encasing octamers of approximately 147 bp of DNA. Post-translational modifications of histones induce alterations in the chromatin structure, thereby influencing gene expression regulation.68 Multiple modifications were carried out on particular residues located at the amino and carboxy ends of the histone tails. These modifications include methylation, acetylation, ubiquitination, phosphorylation, SUMOylation, and glycosylation. These changes are facilitated by various enzymes, including histone acetyltransferases (HATs), histone methyltransferases (HMTs), histone deacetylases (HDACs), and histone demethylases (HDMs). The development and progression of cancer are also linked to changes in the expression of these enzymes. Multiple changes have been observed in breast carcinomas, including overexpression of HDAC1, HDAC2, HDAC3, and HDAC6.69
Breast cancer cell proliferation and metastasis have been associated with histone methyltransferases, such as lysine methyl transferase 2 (KMT2). KMT2 promotes the expression of oncogenes and metastatic genes through the methylation of H3K4 at enhancer and promoter sites.70 Abnormal expression of the essential histone methyltransferase enhancer of zeste homolog 2 (EZH2) is prevalent in breast tumors. EZH2 promotes transcriptional silencing of various genes by inducing H3K27 methylation, thereby altering the EMT and metastasis in breast carcinoma.71,72 In addition, another important HMT known as Disruptor silencing 1 like (DOT1L), is also recognized to be able to enhance metastatic in breast cancer cells.73 The development of breast cancer is also linked to a number of histone demethylases, such as lysine-specific demethylase 4A (KDM4A), Lysine Specific Demethylase 4B (KDM4B), and lysine-specific demethylase 4C (KDM4C). Cells of the ERα-positive subtype demonstrate overexpression of KDM4A and KDM4B, whereas triple-negative breast cancer is characterized by elevated levels of KDM4C.74 KDM4A activates a Notch1-dependent signaling cascade that facilitates breast cancer proliferation and metastasis.75 KDM4B modulates estrogen signaling and its downregulation restricts breast cancer proliferation.76 KDM4C functions as a coactivator of the HIF-1α/VEGF pathway, thereby facilitating breast cancer tumorigenesis.77
Estrogen Associated Epigenetic Alterations
Breast cancer subtypes can be distinguished based on their epigenetic processes. ER-linked epigenetic modifications in breast tumors are governed by transcription factors and co-regulators. Estrogen governs mitotic and epigenetic processes involved in mammary gland development. Estrogen is categorized into five distinct forms: estrogen, estrone, 17-estradiol, estriol, and estrone sulfate.78 Estradiol (E2) serves as a catalyst for breast cancer development. E2 treatment promotes breast cancer carcinogenesis in vitro through anchorage-independent growth, loss of ductulogenesis in collagen, and increased invasiveness.79 E2 activation is facilitated by the ER and involves the recruitment of numerous co-regulators to chromatin, including the p160 family, p300, protein arginine methyltransferases, and certain mediator complexes.80,81 Polycomb proteins serve as a connection between the ER and Wnt signaling networks. One significant component that can directly interact with ER and beta-catenin is the polycomb group protein enhancer of zeste homolog 2 (EZH2), which connects the estrogen and Wnt pathways, indicating a strong correlation between Wnt signaling and carcinogenesis as well as metastasis in ER-positive breast carcinoma.82 In breast cancer, DNA methylation-mediated epigenetic silencing controls Wnt antagonist genes including SERP and DKK. Methylation of Wnt antagonistic genes constantly stimulates β-catenin, which in turn triggers Wnt-associated genes. This increases the rate of stem cell growth and regeneration, leading to poor prognosis and cancer recurrence.83 For instance, DKK3 promoter methylation was observed in 78% of individuals with primary breast cancer, and these individuals had a worse prognosis and elevated metastatic rates than those without DKK3 methylation.84 Furthermore, epigenetic silencing of DKK3 led to lymph node metastases and positive ERα status.85 Targeting these epigenetic mechanisms typically regulates oncogenic signals such as Wnt signaling.
Epigenetic Alterations and EMT
The EMT plays a pivotal role in cancer metastasis. In various malignancies, including breast cancer, the interplay between EMT transcription factors and epigenetic regulators is significant, resulting in EMT through the modulation of genes associated with this process.86,87 In breast cancer, DOT1L interacts with c-Myc and p300 to facilitate the methylation and acetylation of H3K79 in the promoter regions of EMT transcription regulators, resulting in enhanced levels of associated genes. Thus, this mechanism enhances the hallmarks of cancer stemness related to epithelial-mesenchymal transition (EMT). Moreover, invasive breast cancer cells exhibit abnormally elevated CDH1 methylation, which is associated with downregulated E-cadherin production.88 E-cadherin is a tumor suppressor that promotes cell-to-cell contact between adjacent cells.89 The interaction between G9a and Snail in breast cancer inhibits transcription at E-cadherin promoter sites. By reducing Slug expression, the histone deacetylase inhibitor Trichostatin A (TSA) can reverse epithelial-mesenchymal transition (EMT) in breast carcinoma.90,91 Furthermore, blocking bromodomain protein 4 (BRD4) inhibits Gli1, which is essential for transcriptional promotion of Snail proteins. This indicates that BRD4 modulates the tumorigenicity of breast cancer cells via regulation of Snail expression and post-translational processes.92 Consequently, recognizing the interaction between EMT and epigenetics presents novel opportunities for cancer therapy. As a result, epigenetic pathways present numerous avenues for breast cancer treatment. Given that epigenetic modifications caused by DNMTs and HDACs are temporary and reversible, numerous investigations are presently underway to determine the optimal dosage and treatment regimens for various epigenetic agents associated with breast cancer.
Phytochemicals as Epigenetic Modulators in Breast Cancer
Numerous studies have elucidated the potential of phytochemicals in cancer therapy. These plant-derived therapeutic substances are mainly extracted from fruits, vegetables, cereals, spices, and medicinal herbs.93 There has been data from preclinical, clinical, and epidemiological studies that a high intake of plant-based diets high in phytochemicals is associated with a lower incidence of certain types of cancer.94 The mechanisms that collectively strengthen the chemopreventive and therapeutic effects of these phytochemicals include antioxidant, antiproliferative, apoptosis-inducing, anti-inflammatory properties, alteration of oncogenic signaling, anti-angiogenesis, modulation of the tumor microenvironment, and immune checkpoints.95 Additionally, several phytochemicals demonstrate significant potential as epigenetic regulators in cancer therapy.96 For example, a phytochemical sulforaphane exposure resulted in DNMT downregulation and promoter demethylation in prostate and colon cancer cells.97,98 In a similar manner, various other phytochemicals including quercetin, rosmarinic acid, ursolic acid, and resveratrol also modulated the crucial targets linked to epigenetic dysregulation in cancer.99–102 Numerous preclinical studies have shown that phytochemicals can regulate epigenetic alterations for chemotherapy, but clinical trials are limited. A phase II clinical investigation on the effect of sulforaphane in males with recurrent cancer revealed an enhancement in histone acetylation subsequent to sulforaphane administration103. A phase I clinical trial examined the influence of quercetin on EGCG absorption in prostate cancer, while simultaneously evaluating the inhibitory efficacy of quercetin on enzymatic activity and expression of catechol-O-methyltransferase (COMT) and DNMT1 (NCT01912820).
Recent studies have indicated that phytochemicals may demonstrate health-promoting benefits through epigenetic pathways. These phytochemicals contribute to the regulation of gene expression related to cancer and age-associated illnesses linked to oxidative stress and inflammation.104 Various plant-derived phytochemicals, including flavonoids, isothiocyanates, quinones, and alkaloids, have been investigated as effective agents for modulating epigenetic pathways. In this regard, phytochemicals could offer an alternative therapeutic approach for the management of cancers, including breast cancer, by targeting epigenetic pathways (Figure 2).105 Phytochemicals that target epigenetic regulatory mechanisms in breast cancer are listed in Table 1.
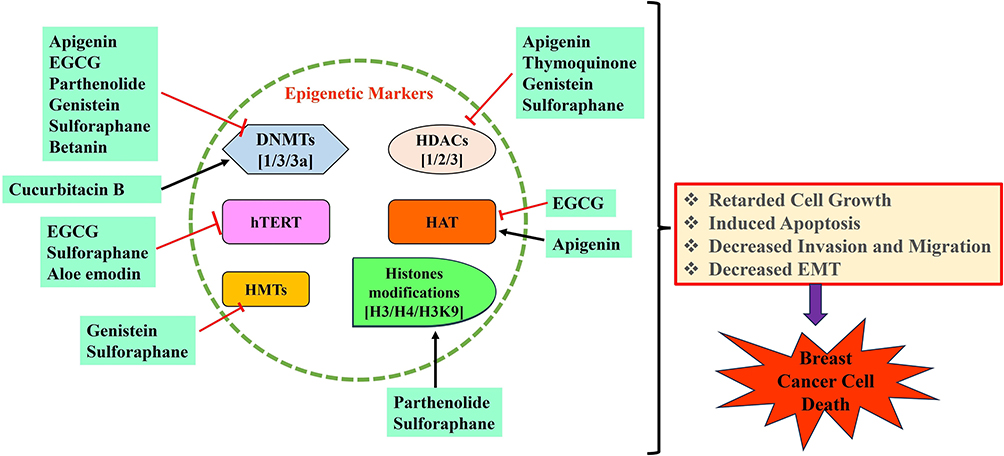 |
Figure 2 An overview of mechanistic effects of natural phytochemicals in modulation of epigenetic pathways in breast cancer cells. |
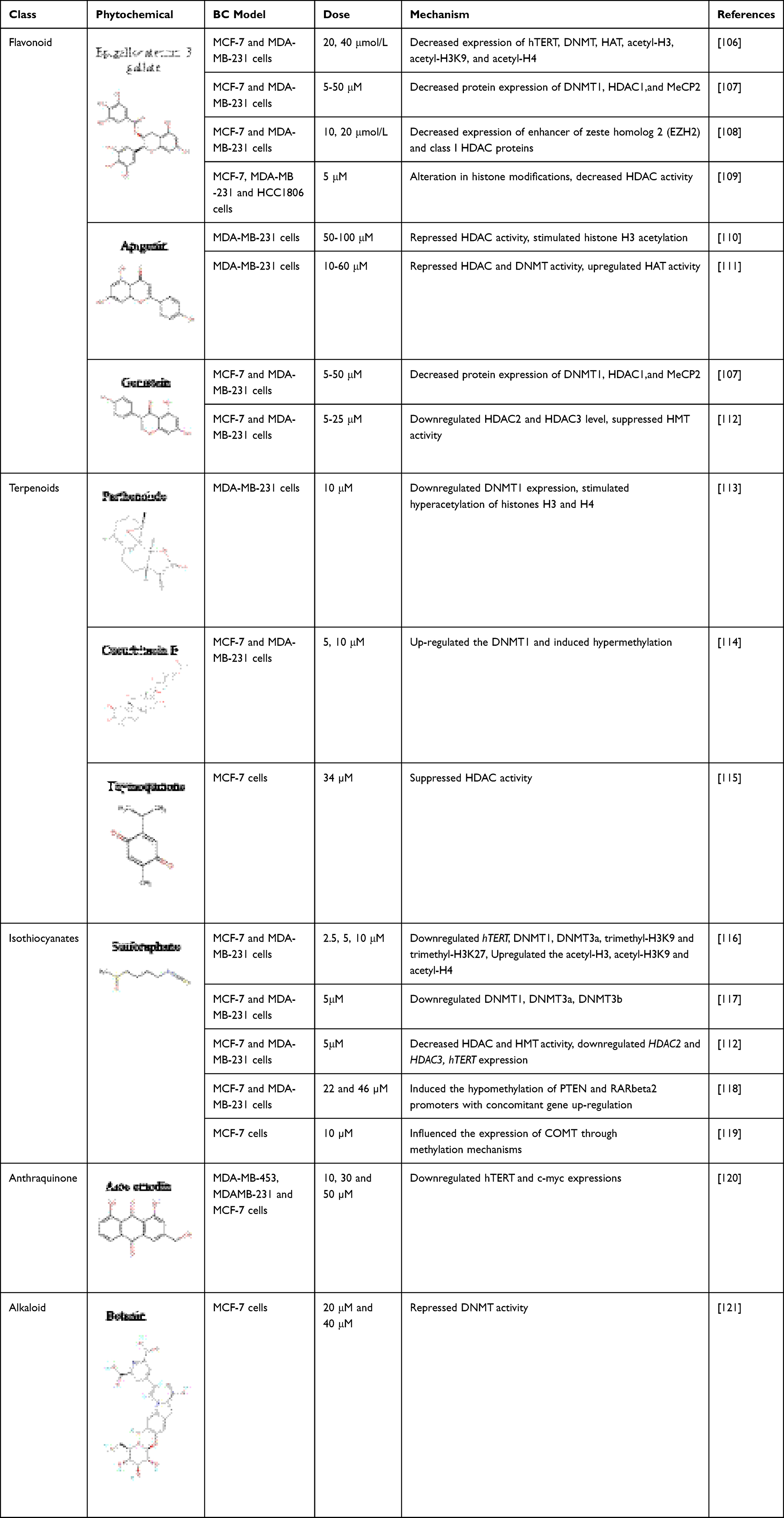 |
Table 1 List of Phytochemicals Implicated in Epigenetic Alterations in Breast Cancer |
Flavonoids
Among the several possibilities for the treatment of breast cancer, flavonoids, which belong to the polyphenol subgroup, show great therapeutic potential. Recent data indicate that beyond their antioxidant capabilities, flavonoids can directly engage with proteins, rendering them suitable small molecules for the control of enzymes, transcription elements, and cell surface receptors.122 A number of flavonoids, including apigenin, genkwanin, silymarin, kaempferol, icariin, silibinin, and luteolin, have been found to repress cell proliferation and trigger apoptosis, cell cycle arrest and autophagy related cell death in breast cancer cells.123–127 The potential of flavonoids to alter the epigenetic regulatory mechanisms in breast cancer is particularly intriguing. Nonetheless, the clinical use of flavonoids in cancer trials has been limited.128 Phytochemicals, a family of pleiotropic compounds, have shown considerable promise in modifying several cancer processes through epigenetic mechanisms. The green tea polyphenol, epigallocatechin-3-gallate (EGCG), is one of the most widely studied flavonoids. The majority of studies indicate that the inhibitory mechanisms of EGCG include alterations in cell cycle progression, proliferation, and apoptotic cell death through the modulation of various signaling pathways. Besides these, another mechanism elucidating the diverse effects of EGCG in tumors is the epigenetic alteration through multiple processes.129
Meeran et al106 found that EGCG and pro-EGCG blocked the transcription of hTERT (human telomerase reverse transcriptase) via epigenetic pathways in MCF-7 and MDA-MB-231 cells. At least largely due to DNA methyltransferase and histone acetyltransferase suppression, hTERT promoter hypomethylation and histone deacetylation downregulate hTERT expression. Furthermore, EGCG and pEGCG can alter the chromatin architecture of the hTERT promoter by reducing the levels of acetyl-H3, acetyl-H3K9, and acetyl-H4. EGCG and pEGCG prompted chromatin modifications that enhanced the association of numerous hTERT repressors such as E2F-1 and MAD1.106
Mirza et al107 elucidated the efficacy of many phytochemicals, including EGCG, in reversing epigenetic alterations in breast cancer. This study demonstrated that EGCG administration significantly reduced the transcript levels of all the examined DNMTs. The levels of HDAC1, DNMT1, and MeCP2 proteins were significantly reduced as a result of these phytochemicals.107
A separate study demonstrated the capacity of EGCG to epigenetically induce the expression of tissue inhibitor of matrix metalloproteinase-3 (TIMP-3) in breast cancer cells. The molecular mechanism demonstrated that TIMP-3 suppression in breast cancer cells is facilitated by epigenetic regulatory mechanisms involving the enhanced activity of EZH2 and class I HDACs, irrespective of promoter DNA hypermethylation. Administration of GTP and EGCG to these cancer cells markedly decreased expression of EZH2 and class I HDAC proteins. Moreover, the transcriptional activation of TIMP-3 was correlated with reduced EZH2 localization and a higher level of H3K27 trimethylation at the TIMP-3 promoter, alongside a rise in histone H3K9/18 acetylation.108 Steed et al109 further revealed that EGCG reduced the levels of cIAP2, while augmenting the level of the pro-apoptotic marker caspase-7. Histone modifications also exhibited alterations, indicating the involvement of epigenetic factors in the variations in cIAP2 expression. A substantial reduction in AcH3 enrichment within the selected promoter region was observed in the two breast cancer cell lines after the combinatorial administration of SAHA and EGCG. A notable reduction in HDAC activity was observed in all three breast cancer cell types, as well as in the MCF-7 cell line, when treated with SAHA and EGCG in combination. Notably, the combinatorial action of SAHA and EGCG enhanced the transcription of HMTs in MCF-7 cells.109
An example of a plant flavone is apigenin, which can be isolated from a variety of fruits and vegetables, such as parsley and celery. Apigenin is recognized as a powerful anticancer compound that interacts with oncogenes and obstructs DNA replication, induces apoptosis via caspase activation and ROS generation in diverse cancer types including colon, breast, skin, leukemia, prostate, and thyroid, thus inhibiting cancer progression.130 In this context, preclinical findings have revealed that apigenin significantly influences cancer management by altering epigenetic processes and tumorigenesis. Apigenin treatment reduced cell growth and cell cycle arrest (G2/M) in MDA-MB-231 breast cancer cells. Immunoblot analysis revealed that apigenin modulated the expression of cell cycle markers, including cyclin A, cyclin B, CDK1, and p21WAF1/CIP1, and augmented its association with proliferating cell nuclear antigen (PCNA), thereby inhibiting cell cycle progression. Moreover, apigenin markedly suppresses HDAC activity and promotes histone H3 acetylation. Apigenin enhanced the acetylation of histone H3 in the p21WAF1/CIP1 promoter, leading to increased p21WAF1/CIP1 expression. In a tumor xenograft model, apigenin significantly inhibited tumor proliferation. As a result of apigenin treatment, cyclin A and cyclin B levels decreased in these xenograft model, but p21WAF1/CIP1 and acetylated histone H3 levels increased.110 A separate study examined the anti-breast cancer efficacy of apigenin and its combination with Vorinostat against MDA-MB-231 cells. The findings indicated that apigenin decreased the expression of class I HDACs at both transcriptional and proteomic levels. Apigenin suppresses the enzymatic function of HDAC/DNMT and enhances HAT activity. Apigenin has been shown to affect miRNA expression by stimulating the tumor suppressor miR-200b and downregulating oncomiR-21. The combinatorial treatment repressed the growth of breast carcinoma cells by regulating the levels of epigenetic and apoptotic markers. The in vitro investigations were supported by in-silico findings, which investigated the mechanism of catalytic suppression by HDAC1 and HDAC3.111
Genistein is a bioactive isoflavone present in soybeans and in several soy-derived products. They are referred to as phytoestrogens or estrogenic substances with anti-cancer effects. Genistein exhibits dose-dependent epigenetic effects on DNMT suppression, gene transcription, and histone acetylation.131 Mirza et al107 also reported the ability of genistein to reverse epigenetic alterations in breast cancer. This study demonstrated that genistein therapy significantly reduced the expression levels of all the examined DNMTs. Significantly, these natural substances reduced the expression of HDAC1, DNMT1, and MeCP2 in breast cancer.107
Paul et al112 revealed the synergistic benefits of genistein and sulforaphane in inhibiting breast tumors through epigenetic regulation. These findings demonstrated that the synergistic effect of genistein and sulforaphane significantly surpassed their individual dosages in enhancing apoptotic rates and reducing the colony-forming capacity of breast cancer cells. These phytochemicals augmented cell cycle arrest at G2-phase in MDA-MB-231 cells and at G1-phase in MCF-7 breast cancer cells. Moreover, the results indicated that the combination functioned effectively as an HDAC and HMT repressor. This combination also reduced the expression of HDAC2 and HDAC3 at both the gene and protein level. Likewise, the combination of genistein and sulforaphane is efficacious in diminishing hTERT levels, which are known to be activated upon KLF4 binding to its promoter region.112
Terpenoids
Terpenoids, the predominant class of chemicals synthesized by plants, are formed from mevalonic acid and are distinguished by a molecular structure composed of isoprene units.132 Terpenoids have considerable significance in biology, chemistry, and pharmacology. In addition to many other biological features, its anticancer effects are particularly remarkable. These effects include anti-proliferative, apoptosis-inducing, anti-migratory, and anti-metastatic properties.133 Terpenoids are extensively found in nature and have also shown anti-breast cancer effects through the targeting of epigenetic markers. Parthenolide is a sesquiterpene lactone extracted from the medicinal plant, Tanacetum parthenium. Because of its multi-targeted modes of action, parthenolide has emerged as a potentially useful therapeutic compound. These mechanisms of action interfere with important signaling pathways that are associated with the proliferation and survival of cancer cells.134 In recent years, the emergence of epigenetic therapy has unveiled a growing number of parthenolide’s epigenetic modifications in breast carcinoma. Carlisi et al113 demonstrated that treatment with parthenolide alone activated the Akt/mTOR survival pathway and subsequently facilitated the nuclear transport of Nrf2, whereas administration with SAHA alone prompted autophagic effect. Nevertheless, when cancer cells were exposed to SAHA and parthenolide in combination, the impact of parthenolide on Akt/mTOR/Nrf2 signaling was mitigated by SAHA, and the autophagic action of SAHA was diminished by parthenolide. The combinatorial treatment with these two compounds resulted in GSH depletion, a decrease in MMP, cytochrome c release, and caspase-mediated apoptosis. Ultimately, they exhibited that the combination treatment preserved both the hyperacetylation of histones H3 and H4 caused by SAHA and the decreased expression of DNMT1 caused by parthenolide.113
Cucurbitacin B, another terpenoid, has similarly demonstrated an antiproliferative action against a variety of human breast cancer cells. Dittharot et al114 investigated the effect of cucurbitacin B derived from Trichosanthes cucumerina on the promoter methylation levels of cyclin D1, c-Myc, and survivin in breast cancer. These findings demonstrate that cucurbitacin B can impede cellular proliferation in breast carcinoma cells. Oncogene promoters are typically hypomethylated in neoplastic cells. Following treatment with cucurbitacin B, there was an overexpression of DNMT1 and significant methylation in the promoters of target genes, resulting in the downregulation of these oncogenes.114
Thymoquinone is the active compound in black cumin seeds and has been utilized in various conventional healthcare systems for thousands of years. Thymoquinone is a natural phytochemical predominantly extracted from Nigella sativa (black cumin or black seed), utilized in various ancient traditional medicines, particularly within Ayurvedic and Unani practices across Arabian, South Asian, Mediterranean, and African countries. Numerous studies have demonstrated that thymoquinone has significant effects on the management of multiple illnesses, including cancers, by regulating diverse biochemical, molecular, and physiological processes.135 Thymoquinone has been shown to suppress DNA replication and survival in cancerous cells by disrupting the DNA structure. Thymoquinone is a prospective anticancer agent as it can target several cell signaling pathways implicated in carcinogenesis, including the modification of epigenetic characteristics of cancer cells, including DNA methylation or demethylation and histone acetylation or deacetylation.136 Parbin et al used an in vitro cell culture investigation to examine the effects of these interactions in addition to an in-silico technique to shed light on the basic interactions of thymoquinone with HDAC. Combined analysis of docking and molecular dynamics modeling revealed the fundamental engagement and stability of thymoquinone with HDACs. Thymoquinone demonstrated HDAC repression at an IC50 of 34 mM in a time-dependent manner. MCF-7 cells subjected to thymoquinone treatment for 24 h demonstrated a substantial reduction in HDAC activity compared with the control. A significant alteration in HDAC activity was noted following thymoquinone exposure for 48 and 72 h.115
Isothiocyanates
Cruciferous vegetables contain abundant bioactive compounds, particularly isothiocyanates, which offer various health benefits. Isothiocyanates are the hydrolytic intermediates of glucosinolates. Some examples of isothiocyanates include sulforaphane, allyl isothiocyanate, benzyl isothiocyanate, and phenethyl isothiocyanate.137 Despite exhibiting numerous features, the most significant attribute of sulforaphane is its anticancer potential. Sulforaphane prevents tumor progression by suppressing the growth of cancer cells, halting the cell cycle, and promoting apoptosis.138–140 Sulforaphane also offers cancer prevention through the modification of many epigenetic and non-epigenetic processes, as evidenced in multiple cancer types. Sulforaphane inhibits HDAC function in malignant cells. Suppression of histone deacetylase is crucial for cancer prevention because it amplifies various mechanisms, including apoptosis and cell cycle arrest.141–143 Moreover, sulforaphane inhibits histone phosphorylation by augmenting the activity of phosphatases, particularly PP1β and PP2α.144 The epigenetic regulation of sulforaphane in breast cancer was evaluated in multiple research.145
In MCF-7 and MDA-MB-231 cells, sulforaphane exposure suppressed hTERT activity in a dose- and time-responsive manner. DNMTs, particularly DNMT1 and DNMT3a, were reduced in sulforaphane-treated cells, indicating that sulforaphane may suppress hTERT expression by influencing epigenetic mechanisms. Decreased DNMTs expression in response to sulforaphane induces site-dependent CpG demethylation, predominantly inside the first exon of the hTERT gene, thereby enhancing CTCF binding linked to hTERT repression. Immunoprecipitation study of the hTERT promoter shown that sulforaphane elevated the levels of acetyl-H3, acetyl-H3K9, and acetyl-H4, while concurrently reducing the inactive chromatin markers trimethyl-H3K9 and trimethyl-H3K27. Sulforaphane-mediated hyperacetylation enhances the association of numerous hTERT repressor proteins, including MAD1 and CTCF, with the hTERT regulatory area. CTCF depletion with siRNA diminished the SFN-mediated reduction in hTERT transcription in these breast carcinoma cells. Moreover, decreased hTERT expression promotes the apoptotic stimulation in breast carcinoma cells.116
A separate study conducted by Royston et al examined the combinatorial effect of withaferin and sulforaphane on breast cancer cell growth and HDAC1 and DNMTs activity. This combination reduced the enzymatic function of DNMTs in both the breast cancer cell lines. The combination therapy of withaferin and sulforaphane in MCF-7 cells demonstrated superior efficacy in inhibiting DNMT activity compared to sulforaphane alone; however, withaferin alone did not exhibit similar effectiveness. In MDA-MB-231 cell line, the combinatorial treatment effect was markedly significant, surpassing that of withaferin alone. Sulforaphane, in conjunction with withaferin, downregulated the expression levels of DNMT1, DNMT3A, and DNMT3B in both the cancer cells. Furthermore, combination therapy markedly reduced HDAC1 levels in MCF-7 and MDA-MB-231 breast cancer cell lines.117
A different combination of genistein and sulforaphane demonstrated breast tumor suppression through an epigenetic regulatory mechanism. This combination reduced the mRNA and protein levels of HDAC2 and HDAC3. The conjunction of genistein and sulforaphane effectively downregulates hTERT levels, which are activated by KLF4 binding to its promoter region.112
Lubecka-Pietruszewska et al118 demonstrated that sulforaphane induces hypomethylation of PTEN and RARbeta2 promoters, resulting in enhanced gene regulation in breast cancer MCF-7 and MDA-MB-231 cells. The amalgamation of sulforaphane and ClF amplifies these effects, leading to augmentation of cell growth inhibition and apoptotic effects in breast cancer cells.118 Cao et al119 examined the metabolic alterations in ER-positive breast cancer MCF-7 cells exposed to estradiol (E2) and sulforaphane to determine critical metabolite profiles that could elucidate the mechanisms behind the anticancer effects of sulforaphane. These findings indicate that the capacity of sulforaphane to epigenetically regulate COMT expression subsequently affects E2 metabolism.119
Alkaloids and Quinones
Quinones are chemical compounds that are distinguished by their cyclic diketone structure. Anthraquinones originating from the rigid, planar tricyclic aromatic structure of anthracene (3), possess two carbonyl groups located at the 9-, 10-positions and the 1-, 4-positions, respectively.146
Anthraquinones are a class of natural substances that are distinguished by their extensive structural diversity, significant biological activity, and minimal toxicity. Aloe emodin (anthraquinone derivative) is a phytocompound present in the roots and rhizomes of several plants. This phytochemical has demonstrated growth-inhibitory, antiangiogenic, and antiproliferative properties, along with its capacity to reverse multidrug resistance in cancerous cells. Aloe emodin is a broad-spectrum suppressive agent of cancer cells that has been shown to have anticancer properties in a variety of biological pathways, particularly epigenetic control.147,148 A study by Wang et al120 shown that 48 hours of aloe emodin administration leads to telomere shortening and telomerase suppression in breast cancer cells. The expression of hTERT was inhibited by the induction of E2F1 and deactivation of c-Myc proteins. Marked demethylation of CpG islands in the hTERT promoter was found in both breast cancer cells. Aloe emodin contended with dNTP for binding to the enzyme’s active site. Aloe emodin functioned as a stabilizer of telomeric G-quadruplex structures, as demonstrated by titration assays and FRET investigations.120
Alkaloids are a category of natural chemicals that have garnered significant interest owing to their possible therapeutic applications. Alkaloids frequently influence various critical cellular systems associated with cancer growth including epigenetic regulation. Multitargeting capabilities can provide substantial advantages to alkaloids in surmounting resistance mechanisms. Alkaloids have consistently affected cancer pharmacotherapy and scientific research. Several alkaloids are currently used as chemotherapeutic drugs for cancer treatment.149,150 A phase 2 study revealed that the combined administration of vinflunine and trastuzumab showed significant efficacy in treating EGFR-2 positive metastatic breast cancer. Furthermore, vinflunine demonstrated efficacy in a phase 3 trial, although its anticancer effectiveness was not superior to that of other alkylating agents.151 Berberine is notably efficacious against colorectal adenomas and acts as a chemopreventive agent, reducing the recurrence of colorectal adenomas and polypoid lesions when ingested in dietary form (0.3 g twice daily).152,153
Betanin, a water-soluble nitrogenous substance derived from beetroot, possesses several advantageous biological properties, including antioxidant, anti-inflammatory, and anti-tumor effects, attributed to its aromatic amino molecular constituents. Furthermore, it affects the mechanisms of altered cells by slowing their development, promoting apoptosis through the upregulation of specific apoptotic proteins, and inducing alterations in membrane integrity, including apoptotic pathways.154,155 Paluszczak et al121 assessed the impact of various dietary phytochemicals, including betanin, on the expression of DNMTs in human breast cancer MCF7 cells as well as their influence on DNA and histone H3 methylation. The results indicated that betanin reduced DNA methyltransferase activity in breast cancer MCF-7 cells at concentrations of 20 μM and 40 μM.122
Limitations and Challenges
This review focused on the anti-breast cancer potential of phytochemicals by targeting on epigenetic regulatory markers; nevertheless, it is important to note that there are several limitations. The low solubility, high metabolism, and rapid elimination rate of many phytochemicals result in poor bioavailability.156 In order to improve solubility and stability, liposomal and nanoformulations based delivery methods have been employed. These drug delivery methods enhance pharmacokinetics, improved absorption and prevent drug degradation.157 To confirm the effectiveness of phytochemicals in breast cancer treatment, more extensive and rigorous clinical trials are needed. Limited sample size and inadequate standardization are common problems in contemporary research. To address these problems, greater industry-researcher cooperation should be encouraged, and phytochemicals should be incorporated into traditional treatments to enable more thorough studies and improved clinical trials.158 Dose optimization is crucial for phytochemicals because, despite their natural origin, they can have adverse effects if used in high dose range. Determining the ideal dosage that optimizes effectiveness while limiting toxicity is crucial for safe and long-term use.159 The quick metabolism and excretion of phytochemicals from the body limit their potential as medicinal drug or formulation. Many of these phytochemicals including polyphenols, flavonoids, terpenoids and alkaloids, undergo substantial metabolism by cytochrome P450 and phase II conjugation enzymes in the liver and gastrointestinal tract. Drug delivery methods have been revolutionized by nanotechnology, significantly improving the stability, solubility, and bioavailability of phytochemicals employed in breast cancer treatment.160 Targeting overexpressed proteins and enhancing therapeutic results in breast cancer can be accomplished in various ways via combination therapy, which combines phytochemicals with traditional cancer treatments. Altogether, to advance the use of phytochemicals as viable drug candidate options for breast cancer therapy, it is imperative to address these limitations through rigorous clinical studies, enhanced drug delivery systems, and standardized extraction techniques.161
Conclusion and Future Directions
This study emphasizes the potential of phytochemicals including flavonoids, terpenoids, isothiocyanates, quinones, and alkaloids in mitigating the epigenetic instabilities associated with breast cancer, along with the prospective therapeutic advantages that may arise from further exploration in this domain. Various studies have underscored the potential of phytochemicals to inhibit breast cancer carcinogenesis by regulating epigenetic modifications through the targeting of DNMTs, HDACs, HMTs, and other indicators. The investigated phytochemicals appeared to regulate epigenetic alterations in vitro and in certain in vivo cancer models, thereby limiting or reducing cancer cell survival, metastasis, and angiogenesis.
Nonetheless, limitations persist in utilizing these phytochemicals as chemopreventive agents, especially concerning the effective concentration range of individual natural product compounds in preliminary in vitro models, which complicates the identification of optimal and biocompatible concentrations in in vivo models. Moreover, in many instances, the in vivo outcomes are not as favorable or encouraging as those observed in in vitro investigations; hence, they are not suitable for clinical trials. Furthermore, a detailed examination of the proper validation and administration routes for natural products is necessary because of their reactivity and ambiguous characteristics. Future research should concentrate on clinical studies, the optimization of drug delivery technologies, including nanoparticles, micelles, and liposomes, and synergistic effects of phytochemicals in conjunction with conventional therapies to enhance efficacy. Phytochemical therapy combined with genetic profiling and biomarker identification may allow for therapy customization, optimize outcomes, reduce toxicity and provide a cost-effective approach for low-income populations. To enhance breast cancer management, research into phytochemical mode of action, dosage strategies, and dietary inclusion is prioritized.
Data Availability Statement
Not applicable as this is a review article.
Acknowledgment
The authors express their gratitude to the Deanship of Scientific Research at King Khalid University for funding this work through the Large Research Group Project under grant number RGP.02/725/46.
Author Contributions
All authors made a significant contribution to the work reported, whether in the conception, study design, execution, acquisition of data, analysis, and interpretation, or in all these areas, took part in drafting, revising, or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This work was supported by the Basic Science Research Program through the National Research Foundation of Korea (NRF) funded by the Ministry of Education (Grant No. NRF-2020R1I1A2066868); the National Research Foundation of Korea (NRF) grant funded by the Korea government (MSIT, Grant No. RS-2020-NR049559); a grant of the Korea Health Technology R&D Project through the Korea Health Industry Development Institute (KHIDI), funded by the Ministry of Health & Welfare, Republic of Korea (Grant No. RS-2020-KH087790); the Starting Growth Technological R&D Program (TIPS Program, Grant No. RS-2024-00507224) funded by the Ministry of SMEs and Startups (MSS, Korea) in 2024; and the National Research Foundation of Korea (NRF) grant funded by the Korea government (MSIT, Grant No. RS-2024-00350362). The funders had no role in the study design, data collection and analysis, decision to publish, or preparation of the manuscript.
Disclosure
The authors declare no conflicts of interest.
References
1. Giaquinto AN, Sung H, Miller KD, et al. Breast Cancer Statistics, 2022. CA Cancer J Clin. 2022;72(6):524–541. doi:10.3322/caac.21754
2. Bray F, Laversanne M, Sung H, et al. Global cancer statistics 2022: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J Clin. 2024;74(3):229–263. doi:10.3322/caac.21834
3. Dai X, Li T, Bai Z, et al. Breast cancer intrinsic subtype classification, clinical use and future trends. Am J Cancer Res. 2015;5(10):2929–2943.
4. Yap KM, Sekar M, Fuloria S, et al. Drug delivery of natural products through nanocarriers for effective breast cancer therapy: a comprehensive review of literature. Int J Nanomed. 2021;16:7891–7941. doi:10.2147/IJN.S328135
5. Xiong X, Zheng LW, Ding Y, et al. Breast cancer: pathogenesis and treatments. Signal Transduct Target Ther. 2025;10(1):49. doi:10.1038/s41392-024-02108-4
6. Ng CKY, Bidard FC, Piscuoglio S, et al. Genetic heterogeneity in therapy-naïve synchronous primary breast cancers and their metastases. Clin Cancer Res. 2017;23(15):4402–4415. doi:10.1158/1078-0432.CCR-16-3115
7. Schrijver WAME, Selenica P, Lee JY, et al. Mutation profiling of key cancer genes in primary breast cancers and their distant metastases. Cancer Res. 2018;78(12):3112–3121. doi:10.1158/0008-5472.CAN-17-2310
8. Dagdemir A, Durif J, Ngollo M, Bignon YJ, Bernard-Gallon D. Histone lysine trimethylation or acetylation can be modulated by phytoestrogen, estrogen or anti-HDAC in breast cancer cell lines. Epigenomics. 2013;5(1):51–63. doi:10.2217/epi.12.74
9. Suzuki J, Chen YY, Scott GK, et al. Protein acetylation and histone deacetylase expression associated with malignant breast cancer progression. Clin Cancer Res. 2009;15(9):3163–3171. doi:10.1158/1078-0432.CCR-08-2319
10. Duan YT, Sangani CB, Liu W, Soni KV, Yao Y. New promises to cure cancer and other genetic diseases/disorders: epi-drugs through epigenetics. Curr Top Med Chem. 2019;19(12):972–994. doi:10.2174/1568026619666190603094439
11. Thakur C, Qiu Y, Fu Y, et al. Epigenetics and environment in breast cancer: new paradigms for anti-cancer therapies. Front Oncol. 2022;12:971288. doi:10.3389/fonc.2022.971288
12. Israel BB, Tilghman SL, Parker-Lemieux K, Payton-Stewart F. Phytochemicals: current strategies for treating breast cancer. Oncol Lett. 2018;15(5):7471–7478. doi:10.3892/ol.2018.8304
13. El Omari N, Bakrim S, Bakha M, et al. Natural bioactive compounds targeting epigenetic pathways in cancer: a review on alkaloids, terpenoids, quinones, and isothiocyanates. Nutrients. 2021;13(11):3714. doi:10.3390/nu13113714
14. Collins HM, Abdelghany MK, Messmer M, et al. Differential effects of garcinol and curcumin on histone and p53 modifications in tumour cells. BMC Cancer. 2013;13:37. doi:10.1186/1471-2407-13-37
15. Bouyahya A, Mechchate H, Oumeslakht L, et al. The role of epigenetic modifications in human cancers and the use of natural compounds as epidrugs: mechanistic pathways and pharmacodynamic actions. Biomolecules. 2022;12(3):367. doi:10.3390/biom12030367
16. Santarelli R, Currà P, Di Crosta M, Gonnella R, Gilardini Montani MS, Cirone M. Changes in lysine methylation contribute to the cytotoxicity of curcumin in colon cancer cells. Molecules. 2025;30(2):335. doi:10.3390/molecules30020335
17. Li S, Kuo HD, Yin R, et al. Epigenetics/epigenomics of triterpenoids in cancer prevention and in health. Biochem Pharmacol. 2020;175:113890. doi:10.1016/j.bcp.2020.113890
18. Khan A, Khan A, Khan MA, et al. Phytocompounds targeting epigenetic modulations: an assessment in cancer. Front Pharmacol. 2024;14:1273993. doi:10.3389/fphar.2023.1273993
19. Gilbert ER, Liu D. Flavonoids influence epigenetic-modifying enzyme activity: structure - function relationships and the therapeutic potential for cancer. Curr Med Chem. 2010;17(17):1756–1768. doi:10.2174/092986710791111161
20. Anantharaju PG, Reddy DB, Padukudru MA, Chitturi CMK, Vimalambike MG, Madhunapantula SV. Induction of colon and cervical cancer cell death by cinnamic acid derivatives is mediated through the inhibition of Histone Deacetylases (HDAC). PLoS One. 2017;12(11):e0186208. doi:10.1371/journal.pone.0186208
21. Khan H, Belwal T, Efferth T, et al. Targeting epigenetics in cancer: therapeutic potential of flavonoids. Crit Rev Food Sci Nutr. 2021;61(10):1616–1639. doi:10.1080/10408398.2020.1763910
22. Gu HF, Mao XY, Du M. Prevention of breast cancer by dietary polyphenols-role of cancer stem cells. Crit Rev Food Sci Nutr. 2020;60(5):810–825. doi:10.1080/10408398.2018.1551778
23. Ros M, Riesco-Llach G, Polonio-Alcalá E, et al. Inhibition of cancer stem-like cells by curcumin and other polyphenol derivatives in MDA-MB-231 TNBC cells. Int J Mol Sci. 2024;25(13):7446. doi:10.3390/ijms25137446
24. Li H, Li J, Zhang Y, et al. The therapeutic effect of traditional Chinese medicine on breast cancer through modulation of the Wnt/β-catenin signaling pathway. Front Pharmacol. 2024;15:1401979. doi:10.3389/fphar.2024.1401979
25. Goleij P, Sanaye PM, Alam W, et al. Unlocking daidzein’s healing power: present applications and future possibilities in phytomedicine. Phytomedicine. 2024;134:155949. doi:10.1016/j.phymed.2024.155949
26. Kim HJ, Kim N, Kim HW, Park JH, Shin CM, Lee DH. Promising aberrant DNA methylation marker to predict gastric cancer development in individuals with family history and long-term effects of H. pylori eradication on DNA methylation. Gastric Cancer. 2021;24(2):302–313. doi:10.1007/s10120-020-01117-w
27. Yang J, Wang Q, Zhang ZY, et al. DNA methylation-based epigenetic signatures predict somatic genomic alterations in gliomas. Nat Commun. 2022;13(1):4410. doi:10.1038/s41467-022-31827-x
28. Božić T, Kuo CC, Hapala J, et al. Investigation of measurable residual disease in acute myeloid leukemia by DNA methylation patterns. Leukemia. 2022;36(1):80–89. doi:10.1038/s41375-021-01316-z
29. Nepal C, Andersen JB. Alternative promoters in CpG depleted regions are prevalently associated with epigenetic misregulation of liver cancer transcriptomes. Nat Commun. 2023;14(1):2712. doi:10.1038/s41467-023-38272-4
30. Tran TO, Lam LHT, Le NQK. Hyper-methylation of ABCG1 as an epigenetics biomarker in non-small cell lung cancer. Funct Integr Genomics. 2023;23(3):256. doi:10.1007/s10142-023-01185-y
31. Wang C, Leavenworth J, Zhang C, et al. Epigenetic regulation of EIF4A1 through DNA methylation and an oncogenic role of eIF4A1 through BRD2 signaling in prostate cancer. Oncogene. 2022;41(19):2778–2785. doi:10.1038/s41388-022-02272-3
32. Turvey GL, López de Alba E, Stewart E, et al. Epigenetic deprogramming by disruption of CIZ1-RNA nuclear assemblies in early-stage breast cancers. J Cell Biol. 2025;224(5):e202409123. doi:10.1083/jcb.202409123
33. Sarvari P, Sarvari P, Ramírez-Díaz I, Mahjoubi F, Rubio K. Advances of epigenetic biomarkers and epigenome editing for early diagnosis in breast cancer. Int J Mol Sci. 2022;23(17):9521. doi:10.3390/ijms23179521
34. Alalhareth IS, Alyami SM, Alshareef AH, et al. Cellular epigenetic targets and epidrugs in breast cancer therapy: mechanisms, challenges, and future perspectives. Pharmaceuticals. 2025;18(2):207. doi:10.3390/ph18020207
35. Cortellesi E, Savini I, Veneziano M, et al. Decoding the epigenome of breast cancer. Int J Mol Sci. 2025;26(6):2605. doi:10.3390/ijms26062605
36. Mangrulkar SV, Kulkarni SS, Nanepag PV, et al. A comprehensive review on pleiotropic effects and therapeutic potential of soy lecithin. Adv Tradit Med. 2025;25(1):145–164.
37. Kim A, Mo K, Kwon H, et al. Epigenetic regulation in breast cancer: insights on epidrugs. Epigenomes. 2023;7(1):6. doi:10.3390/epigenomes7010006
38. Esteller M. Relevance of DNA methylation in the management of cancer. Lancet Oncol. 2003;4(6):351–358. doi:10.1016/s1470-2045(03)01115-x
39. Mo S, Ye L, Wang D, et al. Early detection of molecular residual disease and risk stratification for stage I to III colorectal cancer via circulating tumor DNA methylation. JAMA Oncol. 2023;9(6):770–778. doi:10.1001/jamaoncol.2023.0425
40. Feinberg AP. Phenotypic plasticity and the epigenetics of human disease. Nature. 2007;447(7143):433–440. doi:10.1038/nature05919
41. Papanicolau-Sengos A, Aldape K. DNA methylation profiling: an emerging paradigm for cancer diagnosis. Annu Rev Pathol. 2022;17:295–321. doi:10.1146/annurev-pathol-042220-022304
42. Kareta MS, Botello ZM, Ennis JJ, Chou C, Chédin F. Reconstitution and mechanism of the stimulation of de novo methylation by human DNMT3L. J Biol Chem. 2006;281(36):25893–25902. doi:10.1074/jbc.M603140200
43. Scourzic L, Mouly E, Bernard OA. TET proteins and the control of cytosine demethylation in cancer. Genome Med. 2015;7(1):9. doi:10.1186/s13073-015-0134-6
44. Yagi M, Kabata M, Tanaka A, et al. Identification of distinct loci for de novo DNA methylation by DNMT3A and DNMT3B during mammalian development. Nat Commun. 2020;11(1):3199. doi:10.1038/s41467-020-16989-w
45. Niehrs C, Schäfer A. Active DNA demethylation by Gadd45 and DNA repair. Trends Cell Biol. 2012;22(4):220–227. doi:10.1016/j.tcb.2012.01.002
46. Pietrasik S, Zajac G, Morawiec J, Soszynski M, Fila M, Blasiak J. Interplay between BRCA1 and GADD45A and its potential for nucleotide excision repair in breast cancer pathogenesis. Int J Mol Sci. 2020;21(3):870. doi:10.3390/ijms21030870
47. Muñoz DP, Lee EL, Takayama S, et al. Activation-induced cytidine deaminase (AID) is necessary for the epithelial-mesenchymal transition in mammary epithelial cells. Proc Natl Acad Sci U S A. 2013;110(32):E2977–E2986. doi:10.1073/pnas.1301021110
48. Morgan HD, Dean W, Coker HA, Reik W, Petersen-Mahrt SK. Activation-induced cytidine deaminase deaminates 5-methylcytosine in DNA and is expressed in pluripotent tissues: implications for epigenetic reprogramming. J Biol Chem. 2004;279(50):52353–52360. doi:10.1074/jbc.M407695200
49. Popp C, Dean W, Feng S, et al. Genome-wide erasure of DNA methylation in mouse primordial germ cells is affected by AID deficiency. Nature. 2010;463(7284):1101–1105. doi:10.1038/nature08829
50. Guo JU, Su Y, Zhong C, Ming GL, Song H. Hydroxylation of 5-methylcytosine by TET1 promotes active DNA demethylation in the adult brain. Cell. 2011;145(3):423–434. doi:10.1016/j.cell.2011.03.022
51. Bos MK, Smid M, Sleijfer S, Martens JWM. Apolipoprotein B mRNA-editing catalytic polypeptide-like-induced protein changes in estrogen receptor-positive, human epidermal growth factor receptor 2-negative breast cancer throughout disease progression. JCO Precis Oncol. 2022;
52. DiMarco AV, Qin X, McKinney BJ, et al. APOBEC mutagenesis inhibits breast cancer growth through induction of T cell-mediated antitumor immune responses. Cancer Immunol Res. 2022;10(1):70–86. doi:10.1158/2326-6066.CIR-21-0146
53. Fujikane T, Nishikawa N, Toyota M, et al. Genomic screening for genes upregulated by demethylation revealed novel targets of epigenetic silencing in breast cancer. Breast Cancer Res Treat. 2010;122(3):699–710. doi:10.1007/s10549-009-0600-1
54. Ciriello G, Sinha R, Hoadley KA, et al. The molecular diversity of Luminal A breast tumors. Breast Cancer Res Treat. 2013;141(3):409–420. doi:10.1007/s10549-013-2699-3
55. Shargh SA, Sakizli M, Khalaj V, et al. Downregulation of E-cadherin expression in breast cancer by promoter hypermethylation and its relation with progression and prognosis of tumor. Med Oncol. 2014;31(11):250. doi:10.1007/s12032-014-0250-y
56. Mirza S, Sharma G, Prasad CP, et al. Promoter hypermethylation of TMS1, BRCA1, ERalpha and PRB in serum and tumor DNA of invasive ductal breast carcinoma patients. Life Sci. 2007;81(4):280–287. doi:10.1016/j.lfs.2007.05.012
57. Ma L, Li C, Yin H, et al. The mechanism of DNA Methylation and miRNA in breast cancer. Int J Mol Sci. 2023;24(11):9360. doi:10.3390/ijms24119360
58. Butcher DT, Rodenhiser DI. Epigenetic inactivation of BRCA1 is associated with aberrant expression of CTCF and DNA methyltransferase (DNMT3B) in some sporadic breast tumours. Eur J Cancer. 2007;43(1):210–219. doi:10.1016/j.ejca.2006.09.002
59. Vallian S, Sedaghat M, Nassiri I, Frazmand A. Methylation status of p16 INK4A tumor suppressor gene in Iranian patients with sporadic breast cancer. J Cancer Res Clin Oncol. 2009;135(8):991–996. doi:10.1007/s00432-008-0534-8
60. Askari M, Sobti RC, Nikbakht M, Sharma SC. Aberrant promoter hypermethylation of p21 (WAF1/CIP1) gene and its impact on expression and role of polymorphism in the risk of breast cancer. Mol Cell Biochem. 2013;382(1–2):19–26. doi:10.1007/s11010-013-1696-5
61. Parfenyev S, Singh A, Fedorova O, Daks A, Kulshreshtha R, Barlev NA. Interplay between p53 and non-coding RNAs in the regulation of EMT in breast cancer. Cell Death Dis. 2021;12(1):17. doi:10.1038/s41419-020-03327-7
62. Jorge AL, Pereira ER, Oliveira CS, et al. MicroRNAs: understanding their role in gene expression and cancer. Einstein. 2021;19:eRB5996. doi:10.31744/einstein_journal/2021RB5996
63. Rahman MM, Brane AC, Tollefsbol TO. MicroRNAs and epigenetics strategies to reverse breast cancer. Cells. 2019;8(10):1214. doi:10.3390/cells8101214
64. Baylie T, Kasaw M, Getinet M, et al. The role of miRNAs as biomarkers in breast cancer. Front Oncol. 2024;14:1374821. doi:10.3389/fonc.2024.1374821
65. Mondal P, Meeran SM. Long non-coding RNAs in breast cancer metastasis. Noncoding RNA Res. 2020;5(4):208–218. doi:10.1016/j.ncrna.2020.11.004
66. Yan S, Teng L, Du J, et al. Long non-coding RNA DANCR aggravates breast cancer through the miR-34c/E2F1 feedback loop. Mol Med Rep. 2024;29(6):93. doi:10.3892/mmr.2024.13217
67. Filippova EA, Fridman MV, Burdennyy AM, et al. Long Noncoding RNA GAS5 in Breast Cancer: epigenetic Mechanisms and Biological Functions. Int J Mol Sci. 2021;22(13):6810. doi:10.3390/ijms22136810
68. Khan SA, Reddy D, Gupta S. Global histone post-translational modifications and cancer: biomarkers for diagnosis, prognosis and treatment? World J Biol Chem. 2015;6(4):333–345. doi:10.4331/wjbc.v6.i4.333
69. Zaib S, Rana N, Khan I. Histone modifications and their role in epigenetics of cancer. Curr Med Chem. 2022;29(14):2399–2411. doi:10.2174/0929867328666211108105214
70. Ansari KI, Kasiri S, Mandal SS. Histone methylase MLL1 has critical roles in tumor growth and angiogenesis and its knockdown suppresses tumor growth in vivo. Oncogene. 2013;32(28):3359–3370. doi:10.1038/onc.2012.352
71. Yang X, Karuturi RK, Sun F, et al. CDKN1C (p57) is a direct target of EZH2 and suppressed by multiple epigenetic mechanisms in breast cancer cells. PLoS One. 2009;4(4):e5011. doi:10.1371/journal.pone.0005011
72. Xie Y, Shi Z, Qian Y, et al. HDAC2- and EZH2-mediated histone modifications induce PDK1 expression through miR-148a downregulation in breast cancer progression and adriamycin resistance. Cancers. 2022;14(15):3600. doi:10.3390/cancers14153600
73. Byun WS, Kim WK, Han HJ, et al. Targeting histone methyltransferase DOT1L by a Novel Psammaplin A analog inhibits growth and metastasis of triple-negative breast cancer. Mol Ther Oncolytics. 2019;15:140–152. doi:10.1016/j.omto.2019.09.005
74. Yang G, Li C, Tao F, et al. The emerging roles of lysine-specific demethylase 4A in cancer: implications in tumorigenesis and therapeutic opportunities. Genes Dis. 2023;11(2):645–663. doi:10.1016/j.gendis.2022.12.020
75. Pei J, Zhang S, Yang X, et al. Epigenetic regulator KDM4A activates Notch1-NICD-dependent signaling to drive tumorigenesis and metastasis in breast cancer. Transl Oncol. 2023;28:101615. doi:10.1016/j.tranon.2022.101615
76. Gaughan L, Stockley J, Coffey K, et al. KDM4B is a master regulator of the estrogen receptor signalling cascade. Nucleic Acids Res. 2013;41(14):6892–6904. doi:10.1093/nar/gkt469
77. Li Z, Peluffo G, Stevens LE, et al. KDM4C inhibition blocks tumor growth in basal breast cancer by promoting cathepsin L-mediated histone H3 cleavage. Nat Genet. 2025;57(6):1463–1477. doi:10.1038/s41588-025-02197-z
78. Thomas MP, Potter BV. The structural biology of oestrogen metabolism. J Steroid Biochem Mol Biol. 2013;137:27–49. doi:10.1016/j.jsbmb.2012.12.014
79. Russo J, Fernandez SV, Russo PA, et al. 17-Beta-estradiol induces transformation and tumorigenesis in human breast epithelial cells. FASEB J. 2006;20(10):1622–1634. doi:10.1096/fj.05-5399com
80. Garcia-Martinez L, Zhang Y, Nakata Y, Chan HL, Morey L. Epigenetic mechanisms in breast cancer therapy and resistance. Nat Commun. 2021;12(1):1786. doi:10.1038/s41467-021-22024-3
81. Lakshmanan MD, Shaheer K. Endocrine disrupting chemicals may deregulate DNA repair through estrogen receptor mediated seizing of CBP/p300 acetylase. J Endocrinol Invest. 2020;43(9):1189–1196. doi:10.1007/s40618-020-01241-5
82. Shi B, Liang J, Yang X, et al. Integration of estrogen and Wnt signaling circuits by the polycomb group protein EZH2 in breast cancer cells. Mol Cell Biol. 2007;27(14):5105–5119. doi:10.1128/MCB.00162-07
83. Sharma A, Mir R, Galande S. Epigenetic regulation of the Wnt/β-catenin signaling pathway in cancer. Front Genet. 2021;12:681053. doi:10.3389/fgene.2021.681053
84. Khan Z, Arafah M, Shaik JP, Mahale A, Alanazi MS. High-frequency deregulated expression of Wnt signaling pathway members in breast carcinomas. Onco Targets Ther. 2018;11:323–335. doi:10.2147/OTT.S154395
85. Shailesh H, Siveen KS, Sif S. Protein arginine methyltransferase 5 (PRMT5) activates WNT/β-catenin signalling in breast cancer cells via epigenetic silencing of DKK1 and DKK3. J Cell Mol Med. 2021;25(3):1583–1600. doi:10.1111/jcmm.16260
86. Su Y, Hopfinger NR, Nguyen TD, Pogash TJ, Santucci-Pereira J, Russo J. Epigenetic reprogramming of epithelial mesenchymal transition in triple negative breast cancer cells with DNA methyltransferase and histone deacetylase inhibitors. J Exp Clin Cancer Res. 2018;37(1):314. doi:10.1186/s13046-018-0988-8
87. Khaled N, Bidet Y. New insights into the implication of epigenetic alterations in the EMT of triple negative breast cancer. Cancers. 2019;11(4):559. doi:10.3390/cancers11040559
88. Cho MH, Park JH, Choi HJ, et al. DOT1L cooperates with the c-Myc-p300 complex to epigenetically derepress CDH1 transcription factors in breast cancer progression. Nat Commun. 2015;6:7821. doi:10.1038/ncomms8821
89. Pal M, Bhattacharya S, Kalyan G, Hazra S. Cadherin profiling for therapeutic interventions in epithelial mesenchymal transition (EMT) and tumorigenesis. Exp Cell Res. 2018;368(2):137–146. doi:10.1016/j.yexcr.2018.04.014
90. Dong C, Wu Y, Yao J, et al. G9a interacts with Snail and is critical for Snail-mediated E-cadherin repression in human breast cancer. J Clin Invest. 2012;122(4):1469–1486. doi:10.1172/JCI57349
91. Wang X, Chen S, Shen T, et al. Trichostatin A reverses epithelial-mesenchymal transition and attenuates invasion and migration in MCF-7 breast cancer cells. Exp Ther Med. 2020;19(3):1687–1694. doi:10.3892/etm.2020.8422
92. Yang GJ, Song YQ, Wang W, Han QB, Ma DL, Leung CH. An optimized BRD4 inhibitor effectively eliminates NF-κB-driven triple-negative breast cancer cells. Bioorg Chem. 2021;114:105158. doi:10.1016/j.bioorg.2021.105158
93. Chauhan P, Pandey P, Ramniwas S, et al. Unveiling the anticancer potential of phytocompounds: pioneering future cancer therapies. Curr Top Med Chem. 2025. doi:10.2174/0115680266354680250121154720
94. Kubczak M, Szustka A, Rogalińska M. Molecular targets of natural compounds with anti-cancer properties. Int J Mol Sci. 2021;22(24):13659. doi:10.3390/ijms222413659
95. Utpal BK, Bouenni H, Zehravi M, et al. Exploring natural products as apoptosis modulators in cancers: insights into natural product-based therapeutic strategies. Naunyn Schmiedebergs Arch Pharmacol. 2025. doi:10.1007/s00210-025-03876-8
96. Dorna D, Grabowska A, Paluszczak J. Natural products modulating epigenetic mechanisms by affecting histone methylation/demethylation: targeting cancer cells. Br J Pharmacol. 2025;182(10):2137–2158. doi:10.1111/bph.16237
97. Wong CP, Hsu A, Buchanan A, et al. Effects of sulforaphane and 3,3’-diindolylmethane on genome-wide promoter methylation in normal prostate epithelial cells and prostate cancer cells. PLoS One. 2014;9(1):e86787.
98. Zhou JW, Wang M, Sun NX, et al. Sulforaphane-induced epigenetic regulation of Nrf2 expression by DNA methyltransferase in human Caco-2 cells. Oncol Lett. 2019;18(3):2639–2647.
99. Wang J, Li Y, Wang X, Jiang C. Ursolic acid inhibits proliferation and induces apoptosis in human glioblastoma cell lines U251 by suppressing TGF-β1/miR-21/PDCD4 pathway. Basic Clin Pharmacol Toxicol. 2012;111(2):106–112. doi:10.1111/j.1742-7843.2012.00870.x
100. Kedhari Sundaram M, Hussain A, Haque S, Raina R, Afroze N. Quercetin modifies 5’CpG promoter methylation and reactivates various tumor suppressor genes by modulating epigenetic marks in human cervical cancer cells. J Cell Biochem. 2019;120(10):18357–18369. doi:10.1002/jcb.29147
101. Han Y, Ma L, Zhao L, Feng W, Zheng X. Rosmarinic inhibits cell proliferation, invasion and migration via up-regulating miR-506 and suppressing MMP2/16 expression in pancreatic cancer. Biomed Pharmacother. 2019;115:108878. doi:10.1016/j.biopha.2019.108878
102. Zhao J, Fang Z, Zha Z, et al. Quercetin inhibits cell viability, migration and invasion by regulating miR-16/HOXA10 axis in oral cancer. Eur J Pharmacol. 2019;847:11–18. doi:10.1016/j.ejphar.2019.01.006
103. Alumkal JJ, Slottke R, Schwartzman J, et al. A phase II study of sulforaphane-rich broccoli sprout extracts in men with recurrent prostate cancer. Invest New Drugs. 2015;33(2):480–489. doi:10.1007/s10637-014-0189-z
104. Bouyahya A, El Menyiy N, Oumeslakht L, et al. Preclinical and clinical antioxidant effects of natural compounds against oxidative stress-induced epigenetic instability in tumor cells. Antioxidants. 2021;10(10):1553. doi:10.3390/antiox10101553
105. Shah D, Gandhi M, Kumar A, Cruz-Martins N, Sharma R, Nair S. Current insights into epigenetics, noncoding RNA interactome and clinical pharmacokinetics of dietary polyphenols in cancer chemoprevention. Crit Rev Food Sci Nutr. 2023;63(12):1755–1791. doi:10.1080/10408398.2021.1968786
106. Meeran SM, Patel SN, Chan TH, Tollefsbol TO. A novel prodrug of epigallocatechin-3-gallate: differential epigenetic hTERT repression in human breast cancer cells. Cancer Prev Res. 2011;4(8):1243–1254. doi:10.1158/1940-6207.CAPR-11-0009
107. Mirza S, Sharma G, Parshad R, Gupta SD, Pandya P, Ralhan R. Expression of DNA methyltransferases in breast cancer patients and to analyze the effect of natural compounds on DNA methyltransferases and associated proteins. J Breast Cancer. 2013;16(1):23–31. doi:10.4048/jbc.2013.16.1.23
108. Deb G, Thakur VS, Limaye AM, Gupta S. Epigenetic induction of tissue inhibitor of matrix metalloproteinase-3 by green tea polyphenols in breast cancer cells. Mol Carcinog. 2015;54(6):485–499. doi:10.1002/mc.22121
109. Steed KL, Jordan HR, Tollefsbol TO. SAHA and EGCG promote apoptosis in triple-negative breast cancer cells, possibly through the modulation of cIAP2. Anticancer Res. 2020;40(1):9–26. doi:10.21873/anticanres.13922
110. Tseng TH, Chien MH, Lin WL, et al. Inhibition of MDA-MB-231 breast cancer cell proliferation and tumor growth by apigenin through induction of G2/M arrest and histone H3 acetylation-mediated p21WAF1/CIP1 expression. Environ Toxicol. 2017;32(2):434–444. doi:10.1002/tox.22247
111. Nimal S, Kumbhar N, Saruchi, et al. Apigenin and its combination with Vorinostat induces apoptotic-mediated cell death in TNBC by modulating the epigenetic and apoptotic regulators and related miRNAs. Sci Rep. 2024;14(1):9540. doi:10.1038/s41598-024-60395-x
112. Paul B, Li Y, Tollefsbol TO. The effects of combinatorial genistein and sulforaphane in breast tumor inhibition: role in epigenetic regulation. Int J Mol Sci. 2018;19(6):1754. doi:10.3390/ijms19061754
113. Carlisi D, Lauricella M, D’Anneo A, et al. The synergistic effect of SAHA and parthenolide in MDA-MB231 breast cancer cells. J Cell Physiol. 2015;230(6):1276–1289. doi:10.1002/jcp.24863
114. Dittharot K, Dakeng S, Suebsakwong P, Suksamrarn A, Patmasiriwat P, Promkan M. Cucurbitacin b induces hypermethylation of oncogenes in breast cancer cells. Planta Med. 2019;85(5):370–378. doi:10.1055/a-0791-1591
115. Parbin S, Shilpi A, Kar S, et al. Insights into the molecular interactions of thymoquinone with histone deacetylase: evaluation of the therapeutic intervention potential against breast cancer. Mol Biosyst. 2016;12(1):48–58. doi:10.1039/c5mb00412h
116. Meeran SM, Patel SN, Tollefsbol TO. Sulforaphane causes epigenetic repression of hTERT expression in human breast cancer cell lines. PLoS One. 2010;5(7):e11457. doi:10.1371/journal.pone.0011457
117. Royston KJ, Udayakumar N, Lewis K, Tollefsbol TO. A novel combination of Withaferin A and sulforaphane inhibits epigenetic machinery, cellular viability and induces apoptosis of breast cancer cells. Int J Mol Sci. 2017;18(5):1092. doi:10.3390/ijms18051092
118. Lubecka-Pietruszewska K, Kaufman-Szymczyk A, Stefanska B, Cebula-Obrzut B, Smolewski P, Fabianowska-Majewska K. Sulforaphane alone and in combination with clofarabine epigenetically regulates the expression of DNA methylation-silenced tumour suppressor genes in human breast cancer cells. J Nutrigenet Nutrigenomics. 2015;8(2):91–101. doi:10.1159/000439111
119. Cao S, Wang L, Zhang Z, Chen F, Wu Q, Li L. Sulforaphane-induced metabolomic responses with epigenetic changes in estrogen receptor positive breast cancer cells. FEBS Open Bio. 2018;8(12):2022–2034. doi:10.1002/2211-5463.12543
120. Wang S, Yan WW, He M, Wei D, Long ZJ, Tao YM. Aloe emodin inhibits telomerase activity in breast cancer cells: transcriptional and enzymological mechanism. Pharmacol Rep. 2020;72(5):1383–1396. doi:10.1007/s43440-020-00062-w
121. Paluszczak J, Krajka-Kuźniak V, Baer-Dubowska W. The effect of dietary polyphenols on the epigenetic regulation of gene expression in MCF7 breast cancer cells. Toxicol Lett. 2010;192(2):119–125. doi:10.1016/j.toxlet.2009.10.010
122. Sahoo S, Mohapatra P, Sahoo SK. Flavonoids for the Treatment of breast cancer, present status and future prospective. Anticancer Agents Med Chem. 2023;23(6):658–675. doi:10.2174/1871520623666221024114521
123. Zhang HW, Hu JJ, Fu RQ, et al. Flavonoids inhibit cell proliferation and induce apoptosis and autophagy through downregulation of PI3K$\gamma$ mediated PI3K/AKT/mTOR/p70S6K/ULK signaling pathway in human breast cancer cells. Sci Rep. 2018;8(1):11255. doi:10.1038/s41598-018-29308-7
124. Song L, Chen X, Mi L, et al. Icariin-induced inhibition of SIRT6/NF-κB triggers redox mediated apoptosis and enhances anti-tumor immunity in triple-negative breast cancer. Cancer Sci. 2020;111(11):4242–4256. doi:10.1111/cas.14648
125. Jiang K, Wang W, Jin X, Wang Z, Ji Z, Meng G. Silibinin, a natural flavonoid, induces autophagy via ROS-dependent mitochondrial dysfunction and loss of ATP involving BNIP3 in human MCF7 breast cancer cells. Oncol Rep. 2015;33(6):2711–2718. doi:10.3892/or.2015.3915
126. Kim SH, Choo GS, Yoo ES, et al. Silymarin inhibits proliferation of human breast cancer cells via regulation of the MAPK signaling pathway and induction of apoptosis. Oncol Lett. 2021;21(6):492. doi:10.3892/ol.2021.12753
127. Pham TH, Page YL, Percevault F, Ferrière F, Flouriot G, Pakdel F. Apigenin, a partial antagonist of the estrogen receptor (ER), Inhibits ER-positive breast cancer cell proliferation through Akt/FOXM1 signaling. Int J Mol Sci. 2021;22(1):470. doi:10.3390/ijms22010470
128. Sudhakaran M, Sardesai S, Doseff AI. Flavonoids: New Frontier for immuno-regulation and breast cancer control. Antioxidants. 2019;8(4):103. doi:10.3390/antiox8040103
129. Yang L, Zhang W, Chopra S, et al. The epigenetic modification of epigallocatechin gallate (EGCG) on cancer. Curr Drug Targets. 2020;21(11):1099–1104. doi:10.2174/1389450121666200504080112
130. Pandey P, Khan F, Upadhyay TK. Deciphering the modulatory role of apigenin targeting oncogenic pathways in human cancers. Chem Biol Drug Des. 2023;101(6):1446–1458. doi:10.1111/cbdd.14206
131. Xiang Z, Ma B, Pei X, Wang W, Gong W. Mechanism of action of genistein on breast cancer and differential effects of different age stages. Pharm Biol. 2025;63(1):141–155. doi:10.1080/13880209.2025.2469607
132. Nagegowda DA, Gupta P. Advances in biosynthesis, regulation, and metabolic engineering of plant specialized terpenoids. Plant Sci. 2020;294:110457. doi:10.1016/j.plantsci.2020.110457
133. Guo J, Huang M, Hou S, et al. Therapeutic potential of terpenoids in cancer treatment: targeting mitochondrial pathways. Cancer Rep. 2024;7(9):e70006. doi:10.1002/cnr2.70006
134. Carlisi D, Lauricella M, D’Anneo A, et al. Parthenolide and its soluble analogues: multitasking compounds with antitumor properties. Biomedicines. 2022;10(2):514. doi:10.3390/biomedicines10020514
135. Rahmani AH, Alzohairy MA, Khan MA, Aly SM. Therapeutic implications of Black Seed and its constituent thymoquinone in the prevention of cancer through inactivation and activation of molecular pathways. Evid Based Complement Alternat Med. 2014;2014:724658. doi:10.1155/2014/724658
136. Khan MA, Tania M, Fu J. Epigenetic role of thymoquinone: impact on cellular mechanism and cancer therapeutics. Drug Discov Today. 2019;24(12):2315–2322. doi:10.1016/j.drudis.2019.09.007
137. Fofaria NM, Ranjan A, Kim SH, Srivastava SK. Mechanisms of the anticancer effects of isothiocyanates. Enzymes. 2015;37:111–137. doi:10.1016/bs.enz.2015.06.001
138. Myzak MC, Hardin K, Wang R, Dashwood RH, Ho E. Sulforaphane inhibits histone deacetylase activity in BPH-1, LnCaP and PC-3 prostate epithelial cells. Carcinogenesis. 2006;27(4):811–819. doi:10.1093/carcin/bgi265
139. Clarke JD, Hsu A, Yu Z, Dashwood RH, Ho E. Differential effects of sulforaphane on histone deacetylases, cell cycle arrest and apoptosis in normal prostate cells versus hyperplastic and cancerous prostate cells. Mol Nutr Food Res. 2011;55(7):999–1009. doi:10.1002/mnfr.201000547
140. Su X, Jiang X, Meng L, et al. Anticancer activity of sulforaphane: the epigenetic mechanisms and the Nrf2 signaling pathway. Oxid Med Cell Longev. 2018;2018:5438179. doi:10.1155/2018/5438179
141. Atwell LL, Beaver LM, Shannon J, Williams DE, Dashwood RH, Ho E. Epigenetic regulation by sulforaphane: opportunities for breast and prostate cancer chemoprevention. Curr Pharmacol Rep. 2015;1(2):102–111. doi:10.1007/s40495-014-0002-x
142. Jiang LL, Zhou SJ, Zhang XM, Chen HQ, Liu W. Sulforaphane suppresses in vitro and in vivo lung tumorigenesis through downregulation of HDAC activity. Biomed Pharmacother. 2016;78:74–80. doi:10.1016/j.biopha.2015.11.007
143. Dos Santos PWDS, Machado ART, De Grandis RA, et al. Transcriptome and DNA methylation changes modulated by sulforaphane induce cell cycle arrest, apoptosis, DNA damage, and suppression of proliferation in human liver cancer cells. Food Chem Toxicol. 2020;136:111047. doi:10.1016/j.fct.2019.111047
144. Abbaoui B, Telu KH, Lucas CR, et al. The impact of cruciferous vegetable isothiocyanates on histone acetylation and histone phosphorylation in bladder cancer. J Proteomics. 2017;156:94–103. doi:10.1016/j.jprot.2017.01.013
145. Asif Ali M, Khan N, Kaleem N, et al. Anticancer properties of sulforaphane: current insights at the molecular level. Front Oncol. 2023;13:1168321. doi:10.3389/fonc.2023.1168321
146. Malik MS, Alsantali RI, Jassas RS, et al. Journey of anthraquinones as anticancer agents - a systematic review of recent literature. RSC Adv. 2021;11(57):35806–35827. doi:10.1039/d1ra05686g
147. Karatoprak G Ş, Küpeli Akkol E, Ç Y, Acıkara Ö B, Sobarzo-Sánchez E. Advances in understanding the role of aloe emodin and targeted drug delivery systems in cancer. Oxid Med Cell Longev. 2022;2022:7928200. doi:10.1155/2022/7928200
148. Zhang H, Chen L, Bu HQ, et al. Effects of emodin on the demethylation of tumor-suppressor genes in pancreatic cancer PANC-1 cells. Oncol Rep. 2015;33(6):3015–3023. doi:10.3892/or.2015.3914
149. Koochaki R, Amini E, Zarehossini S, et al. Alkaloids in Cancer therapy: targeting the tumor microenvironment and metastasis signaling pathways. Fitoterapia. 2024;179:106222. doi:10.1016/j.fitote.2024.106222
150. Ahmad I, Fakhri S, Khan H, Jeandet P, Aschner M, Yu ZL. Targeting cell cycle by β-carboline alkaloids in vitro: novel therapeutic prospects for the treatment of cancer. Chem Biol Interact. 2020;330:109229. doi:10.1016/j.cbi.2020.109229
151. Yardley DA, McCleod M, Schreiber F, et al. A phase II trial of vinflunine as monotherapy or in combination with trastuzumab as first-line treatment of metastatic breast cancer. Cancer Invest. 2010;28(9):925–931. doi:10.3109/07357907.2010.496755
152. Chen YX, Gao QY, Zou TH, et al. Berberine versus placebo for the prevention of recurrence of colorectal adenoma: a multicentre, double-blinded, randomised controlled study. Lancet Gastroenterol Hepatol. 2020;5(3):267–275. doi:10.1016/S2468-1253(19)30409-1
153. Rampogu S, Balasubramaniyam T, Lee JH. Phytotherapeutic applications of alkaloids in treating breast cancer. Biomed Pharmacother. 2022;155:113760. doi:10.1016/j.biopha.2022.113760
154. Vieira Teixeira da Silva D, Dos santos baião D, de Oliveira Silva F, et al. Betanin, a natural food additive: stability, bioavailability, antioxidant and preservative ability assessments. Molecules. 2019;24(3):458. doi:10.3390/molecules24030458
155. Hirad AH, Alarfaj AA, Ravindran B, Narasimhamoorthi SP. Betanin inspired zinc oxide nanoparticles: the potential antioxidant and anticancer activity against human lung cancer cell line (A549). Biochem Biophys Res Commun. 2025;742:151019. doi:10.1016/j.bbrc.2024.151019
156. Aqil F, Munagala R, Jeyabalan J, Vadhanam MV. Bioavailability of phytochemicals and its enhancement by drug delivery systems. Cancer Lett. 2013;334(1):133–141. doi:10.1016/j.canlet.2013.02.032
157. Bertoncini-Silva C, Vlad A, Ricciarelli R, et al. Enhancing the bioavailability and bioactivity of curcumin for disease prevention and treatment. Antioxidants. 2024;13(3):331. doi:10.3390/antiox13030331
158. Choudhari AS, Mandave PC, Deshpande M, Ranjekar P, Prakash O. Phytochemicals in cancer treatment: from preclinical studies to clinical practice. Front Pharmacol. 2020;10:1614. Erratum in: Front Pharmacol. 2020;11:175. doi:10.3389/fphar.2019.01614
159. Domingo-Fernández D, Gadiya Y, Preto AJ, et al. Natural products have increased rates of clinical trial success throughout the drug development process. J Nat Prod. 2024;87(7):1844–1851. doi:10.1021/acs.jnatprod.4c00581
160. Chavda VP, Nalla LV, Balar P, et al. Advanced phytochemical-based nanocarrier systems for the treatment of breast cancer. Cancers. 2023;15(4):1023. doi:10.3390/cancers15041023
161. Atanasov AG, Zotchev SB, Dirsch VM, International Natural Product Sciences Taskforce, Supuran CT, et al. Natural products in drug discovery: advances and opportunities. Nat Rev Drug Discov. 2021;20(3):200–216. doi:10.1038/s41573-020-00114-z
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Immunotherapy: Constructive Approach for Breast Cancer Treatment
Anayyat U, Ahad F, Muluh TA, Zaidi SAA, Usmani F, Yang H, Li M, Hassan HA, Wang X
Breast Cancer: Targets and Therapy 2023, 15:925-951
Published Date: 15 December 2023
Antibody-Drug Conjugates in Breast Cancer: A Comprehensive Review of How to Selectively Deliver Payloads
Monteiro MR, Nunes NCC, Junior AADS, Fêde ABDS, Bretas GDO, Souza CDP, Mano M, da Silva JL
Breast Cancer: Targets and Therapy 2024, 16:51-70
Published Date: 28 February 2024
GCDH Promotes Breast Cancer Glutaminolysis Reprogramming by Inducing GLS1 Expression Through Histone Crotonylation at Its Promoter Region
Zhang J, Zou M
Cancer Management and Research 2025, 17:3185-3196
Published Date: 17 December 2025
Emerging Targeted and Multimodal Therapeutic Strategies in Breast Cancer: A Comprehensive Review
Wu W, He Y, Zhang C
Breast Cancer: Targets and Therapy 2026, 18:575936
Published Date: 7 January 2026