Back to Journals » Breast Cancer: Targets and Therapy » Volume 18
Emerging Targeted and Multimodal Therapeutic Strategies in Breast Cancer: A Comprehensive Review
Received 21 October 2025
Accepted for publication 16 December 2025
Published 7 January 2026 Volume 2026:18 575936
DOI https://doi.org/10.2147/BCTT.S575936
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Robert Clarke
Wenning Wu,1,* Yufeng He,2,3,* Chunxiang Zhang2,3
1Department of Pharmacy, School of Pharmacy, Southwest Medical University, Luzhou, Sichuan, 646000, People’s Republic of China; 2Department of Cardiology, The Affiliated Hospital of Southwest Medical University, Luzhou, Sichuan, 646000, People’s Republic of China; 3Nucleic Acid Medicine of Luzhou Key Laboratory, Southwest Medical University, Luzhou, Sichuan, 646000, People’s Republic of China
*These authors contributed equally
Correspondence: Chunxiang Zhang, Email [email protected]
Abstract: Breast cancer treatment has advanced substantially with the development of targeted and multimodal strategies, yet tumor heterogeneity and therapeutic resistance continue to limit long-term outcomes. This review provides a concise and critical overview of established and emerging targeted therapies, with emphasis on resistance mechanisms, innovations in drug delivery, and integrative therapeutic approaches. Established modalities, including endocrine therapy, HER2-directed antibody and antibody–drug conjugate therapies, and chemotherapy, are summarized alongside key subtype-specific challenges. Emerging targets such as PARP, CDK4/6, WBP2, and Trop2, as well as metabolic and tumor-microenvironment–oriented strategies, are evaluated for their potential to overcome resistance. Advances in immunotherapy, including checkpoint inhibitors, bispecific antibodies, and CAR-T cell therapies, are examined with attention to mechanistic rationale and limitations across breast cancer subtypes. Progress in nanotechnology-based delivery systems, exosome-mediated transport, and multimodal predictive frameworks, including miRNA regulatory networks, is further assessed for its relevance to personalized treatment. Overall, this review synthesizes current molecular insights and therapeutic innovations to inform the development of more effective and durable targeted treatments for breast cancer.
Plain Language Summary: Breast cancer is not a single disease. Different people develop different tumor types, which respond in different ways to treatment. Because of this diversity, many women still face treatment resistance, relapse, or limited benefit from current therapies. We wrote this review to help readers understand how new scientific advances are reshaping breast cancer treatment and what these developments may mean for future care.
In this article, we explain the major treatment approaches used today, including hormone-based therapy, HER2-targeted therapy, chemotherapy, immunotherapy, and drug–antibody combinations. We also describe important new directions, such as medicines that act on DNA repair, the cell cycle, tumor metabolism, and proteins like Trop2, WBP2, and CDK4/6. We highlight how the tumor’s surrounding environment, including immune cells, fibroblasts, blood vessels, and the extracellular matrix, can either support treatment or impede its effectiveness. We also summarize emerging tools such as nanomedicine, ultrasound-guided drug delivery, and exosome-based therapies. These technologies aim to deliver drugs more precisely and overcome resistance.
Our review shows that no single therapy is enough for all patients. Instead, combining targeted drugs, immunotherapy, metabolic treatments, or advanced delivery systems may offer stronger and longer-lasting effects. We also discuss early research using prediction models and microRNA networks, which may help doctors choose the best therapy for each person.
Together, these findings suggest that the future of breast cancer treatment will rely on personalized, multi-target strategies designed around the unique biology of each tumor and each patient.
Keywords: breast cancer, targeted therapy, drug resistance, tumor microenvironment, nanomedicine
Introduction
Breast cancer is a highly heterogeneous malignancy with marked variability in molecular subtypes, biological behavior, and therapeutic responsiveness. Although endocrine therapy, HER2 directed agents, chemotherapy, and antibody drug conjugates have all undergone continuous refinement, intrinsic and acquired resistance still limit treatment durability for many patients.1,2 These persistent challenges underscore the need to better understand breast cancer heterogeneity and to accelerate therapeutic innovation.
Previous reviews have documented advances in endocrine therapy, HER2 targeted therapeutics, or resistance mechanisms within specific subtypes, yet most discussions remain focused on isolated pathways or single categories of treatment.3–6 Existing literature has also separately addressed immunotherapy, nanomedicine, pathway inhibition, metabolic rewiring, or drug delivery technologies. However, few attempts have been made to integrate these rapidly developing fields into a broader multimodal framework that reflects the convergence of targeted therapy, tumor microenvironment modulation, immune based strategies, and advanced delivery systems.7–9 This fragmented perspective limits the ability of readers to appreciate how emerging modalities collectively reshape the therapeutic landscape.
Therefore, this review aims to provide a more unified overview of recent advances in targeted and multimodal strategies for breast cancer. By synthesizing progress in novel molecular targets, resistance related signaling networks, innovative delivery platforms such as nanoparticles and exosomes, and integrative models that incorporate immunologic and microenvironmental regulation, we aim to clarify how these approaches complement established therapies and support individualized treatment. Through this perspective, we outline evolving opportunities to overcome resistance and improve long term therapeutic outcomes across breast cancer subtypes.
Molecular Characteristics and Targetable Pathways of Breast Cancer
Breast cancer exhibits substantial molecular heterogeneity, with intrinsic subtypes defined by hormone receptor status, HER2 expression, and transcriptomic signatures.10–13 Hormone receptor positive disease depends on estrogen receptor (ER) and progesterone receptor (PR) signaling for tumor growth and therefore responds to therapies such as tamoxifen and aromatase inhibitors.14,15 HER2-positive (HER2+) tumors are driven by HER2 amplification and overexpression, and targeted agents including trastuzumab and pertuzumab significantly improve outcomes by suppressing HER2 mediated signaling.16,17
Triple-negative breast cancer (TNBC) remains the most therapeutically challenging subtype because it lacks ER, PR, and HER2 expression. Efforts to identify alternative oncogenic drivers have revealed several promising targets. Gupta et al identified CXCR4 signaling as a potential vulnerability that can be inhibited by Z-cocainone.12 Lan et al reported oncogenic circular RNAs such as circBRAF that influence DNA methylation and transcriptional regulation in TNBC.18 Additional studies have shown that proteins including CD151 regulate exosome mediated communication and contribute to metastatic progression.
Collectively, these findings broaden the spectrum of actionable biomarkers and pathways across breast cancer. Although many candidates show potential for precision treatment, most remain in preclinical or early clinical development. Continued investigation will be required to validate their therapeutic relevance and to advance them toward clinical application. Table 1 presents the current targeted treatment strategies for breast cancer subtypes.
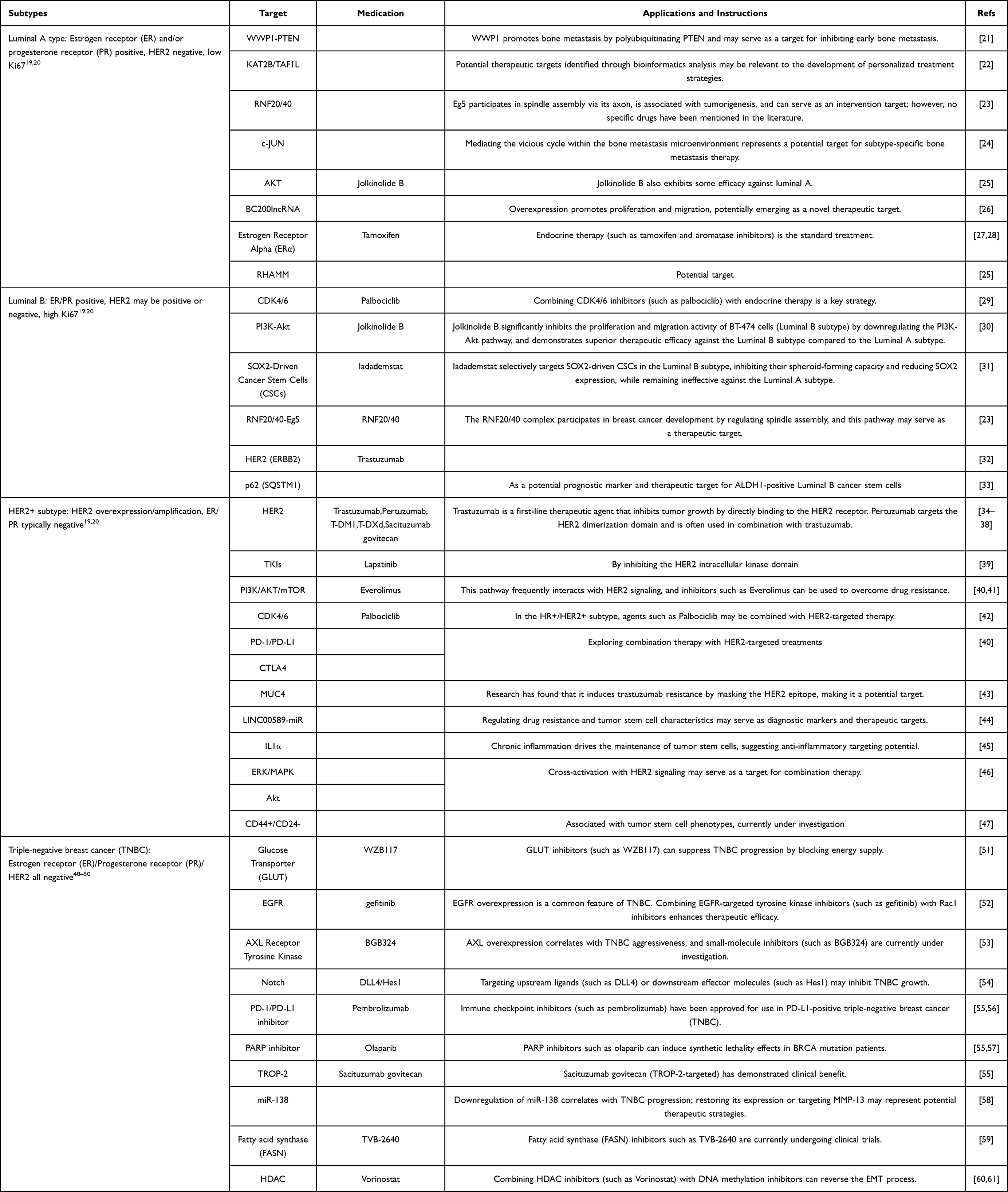 |
Table 1 Targeted Therapy Strategies for Breast Cancer Subtypes |
Current Treatment Strategies
Endocrine-Targeted Therapy for ER+ Breast Cancer
Endocrine therapy remains central to the management of ER-positive (ER+) breast cancer, yet intrinsic and acquired resistance continue to restrict its long-term effectiveness. Evidence shows that activation of the AKT and AMPK pathways drives metabolic reprogramming in endocrine resistant cells, supporting the role of altered energy metabolism as a contributor to therapeutic failure.62 Niu et al further demonstrated that high SGLT1 expression enhances glycolysis and promotes M2 macrophage polarization, thereby accelerating resistance.63
Genomic alterations also influence treatment outcomes. ESR1 mutations enable ligand independent activation of the ER pathway and are among the best-established mechanisms of aromatase inhibitor resistance.64–66 Their non-invasive detection allows earlier identification of emerging resistance.67 Additional regulatory mechanisms such as NR4A1-mediated suppression of ERK signaling and unfolded protein response related stress signaling further illustrate the complexity of endocrine resistance.68,69
New endocrine targeted agents have been developed to counter these mechanisms. The complete ER antagonist palazestrant (OP-1250) shows potent antitumor activity in preclinical studies and fully suppresses estrogen responsive transcriptional programs.70 The ER directed PROTAC vepdegestrant (ARV-471) induces proteasomal degradation of ER and has demonstrated meaningful efficacy in early clinical trials.71,72 Studies also indicate that non coding RNAs and epithelial-mesenchymal transition (EMT) related signaling shape responses to aromatase inhibitors and provide opportunities for combination strategies.73
Despite these advances, resistance remains frequent and reliable biomarkers for predicting response are still lacking. Continued research is needed to develop rational combination approaches and to identify molecular predictors that support individualized therapy for ER+ breast cancer.
Antibody Therapy for HER2+ Breast Cancer
Antibody-based therapy remains central in HER2+ breast cancer. Trastuzumab, the first humanized anti-HER2 antibody, blocks HER2 signaling and induces antibody-dependent cellular cytotoxicity (ADCC), improving outcomes in both metastatic and early-stage disease.74–76 Pertuzumab enhances this effect by inhibiting HER2–HER3 dimerization, forming the standard dual blockade.77–79 Nonetheless, resistance driven by HER3 reactivation, inadequate modulation of the tumor microenvironment (TME), and treatment-related toxicities remains common.79–81
Enhancing immune engagement has emerged as an important strategy. ADCC and antibody-dependent cellular phagocytosis (ADCP) are key components of trastuzumab activity, and CD47 blockade strengthens macrophage-mediated ADCP, improving responses in resistant models.82–87 Antibody-drug conjugates (ADCs) extend HER2 targeting by delivering cytotoxic payloads. T-DM1 (trastuzumab emtansine) provides clinical benefit in metastatic disease, although HER2 downregulation, impaired endocytosis, and compensatory pathway activation increasingly limit its efficacy.88–92
Emerging ADCs aim to overcome these limitations. ARX788, a site-specific trastuzumab–AS269 conjugate, shows robust activity in preclinical and early clinical studies, including T-DM1–resistant models.93 T-DXd (trastuzumab deruxtecan), incorporates a cleavable linker and a topoisomerase I inhibitor payload, demonstrating potent activity even in HER2-low cancers and benefiting patients with prior T-DM1 exposure.94 Disitamab vedotin, composed of an anti-HER2 monoclonal antibody and monomethyl auristatin E (MMAE), has produced encouraging results in prospective clinical studies, including in trastuzumab-resistant disease.95,96
Importantly, HER2 spatial heterogeneity profoundly influences treatment response. Tumors with heterogeneous amplification show markedly reduced pathological complete response (pCR) rates, with neoadjuvant data reporting 55% pCR in homogeneous tumors versus 0% in heterogeneous tumors treated with T-DM1 plus pertuzumab.97–101 This highlights the need for strategies that maintain efficacy despite intratumoral variability. Radionuclide-augmented ADCs such as 177Lu-labeled HER2 or TROP2 conjugates offer improved tumor control in heterogeneous and refractory models by enhancing intratumoral penetration and sustaining cytotoxicity.102
Despite their promise, ADCs carry safety concerns. Interstitial lung disease, particularly associated with T-DXd, requires close monitoring and prompt management.103–108 Toxicity profiles differ across ADC platforms, and long-term safety data remain limited. Advances in spatial proteomics and molecular profiling may refine biomarker development and patient selection. Continued investigation into resistance mechanisms, heterogeneity, and toxicity will be essential to optimize future HER2-targeted antibody and ADC therapies.
Chemotherapy and Its Auxiliary Role in Targeted Therapy
Chemotherapy remains a fundamental component of breast cancer treatment and continues to play an irreplaceable role despite advances in targeted and immune-based therapies. Cytotoxic agents exert antitumor effects by interfering with DNA replication and repair, disrupting mitotic spindle assembly, and inhibiting nucleic acid synthesis.109–112 Although targeted strategies are expanding rapidly, none can fully substitute for chemotherapy, which therefore remains widely integrated into multimodal regimens alongside surgery, radiotherapy, endocrine therapy, and targeted therapy.113,114
Chemotherapy is increasingly combined with targeted agents to enhance efficacy and delay resistance. ADCs such as T-DM1 illustrate how targeted delivery of cytotoxic payloads can strengthen antitumor activity beyond that achieved by chemotherapy alone. Improving intratumoral drug penetration has also become a priority, as abnormal vasculature contributes to chemoresistance. Strategies that promote vascular normalization, including HER2-directed antibodies and the VEGFR2 inhibitor apatinib, improve perfusion and alleviate hypoxia, thereby enhancing the distribution and activity of chemotherapeutic agents.115,116
Clinical evidence supports the auxiliary role of chemotherapy within targeted regimens. In HER2-positive patients with small node-negative tumors (≤2 cm), chemotherapy combined with HER2-targeted therapy reduces recurrence risk by roughly 30% and yields five-year survival exceeding 60%.117 In metastatic settings, dual HER2 blockade with endocrine therapy shows superior efficacy compared with chemotherapy alone, with response rates of 86–91% versus 62%–81%.118 For ER–negative (ER-) tumors, activation of the IRF7/IFN-β/IFNAR axis drives immune dormancy and chemoresistance, and inhibition of this pathway restores sensitivity and correlates with improved metastasis-free survival.119
Despite its central role, chemotherapy has important limitations. In TNBC, systemic chemotherapy remains primary, yet responses are often limited and recurrence is frequent.120–122 In hormone receptor-positive disease, chemotherapy serves mainly as an adjunct to endocrine therapy, with anti-angiogenic agents offering potential enhancement of chemosensitivity.116,123 Toxicities such as neurotoxicity, cardiotoxicity, myelosuppression, and immune suppression restrict dose intensity and may influence subsequent response to targeted or immune-based therapies. Chemotherapy also remodels the tumor microenvironment, contributing to immune suppression and altered stromal architecture, which can limit the effectiveness of later treatments.124,125 Practical and logistical barriers also influence real-world outcomes, as older adults or patients with comorbidities often tolerate standard multi-drug regimens poorly, leading to incomplete therapy and suboptimal survival benefits.
Together, these findings highlight the enduring importance of chemotherapy in breast cancer while emphasizing the need for strategies that enhance delivery, mitigate toxicity, and overcome microenvironmental and biological resistance. Figure 1 summarizes the current systemic treatment strategies.
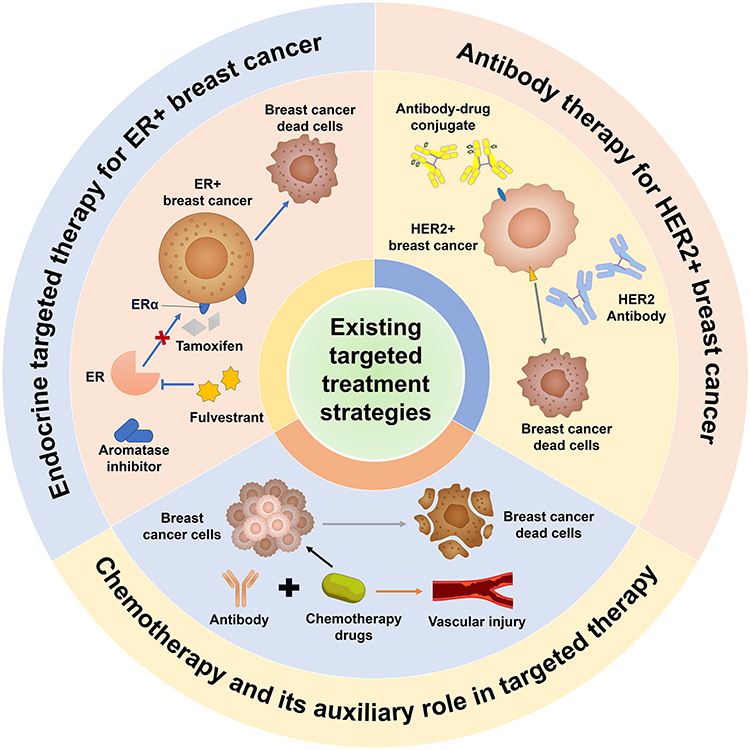 |
Figure 1 Overview of current systemic treatment strategies for breast cancer. This schematic summarizes the major systemic therapies used in breast cancer management, including endocrine therapy for ER+ tumors, HER2-targeted antibody therapy and antibody-drug conjugates for HER2+ tumors, and chemotherapy as a foundational modality across subtypes. Endocrine targeted therapy: In ER+ breast cancer, agents including tamoxifen, fulvestrant, and aromatase inhibitors block ERα signaling, leading to BCCD; Antibody therapy: In HER2+ breast cancer, HER2-targeted antibodies and antibody-drug conjugates inhibit tumor growth; Chemotherapy: Chemotherapy combined with antibody therapy induces BCDC but may also cause vascular damage. Abbreviations: BCDC, Breast cancer dead cells; ER, Estrogen Receptor; HER2, Human Epidermal growth factor Receptor 2; ERα, Estrogen Receptor alpha. |
Advanced Therapeutic Targets in Breast Cancer Management
Application of PARP Inhibitors in BRCA-Mutated Breast Cancer
Poly (ADP-ribose) polymerase (PARP) participates in multiple processes relevant to breast cancer progression, including chromatin remodeling, DNA single-strand break repair, TGFβ-driven EMT, and JAK/STAT3-mediated metabolic regulation.126–128 PARP inhibitors represent a major advance in precision oncology. By exploiting homologous recombination repair (HRR) defects caused by BRCA mutations, they induce synthetic lethality and significantly improve progression-free survival (PFS) in appropriately selected patients.129–132 Olaparib, the first approved PARP inhibitor for germline BRCA-mutated breast cancer, inhibits PARP1 and PARP2 and disrupts DNA repair. In the Phase III OlympiAD trial, olaparib prolonged PFS in gBRCA-mutated, HER2-negative metastatic breast cancer compared with physician’s-choice chemotherapy (19.3 vs 8.2 months).130,133–135 Other agents such as talazoparib have shown comparable efficacy in early- and late-phase clinical trials.133,136
Importantly, recent evidence indicates that PARP inhibitors are also effective in tumors with somatic BRCA mutations or broader HRR impairment. This phenotype, termed “BRCAness”, arises from alterations in HR-related genes beyond BRCA1/2 and confers sensitivity to PARP inhibition.137 Accordingly, potential beneficiaries extend beyond germline BRCA carriers.
PARP inhibitors are particularly relevant in TNBC with BRCA1/2 loss. PARP blockade prevents single-strand break repair, causing replication fork collapse and double-strand breaks. Tumor cells lacking BRCA-mediated HRR cannot repair such lesions, leading to apoptosis.129,138,139 However, prolonged inhibition may promote cancer stem cell–like phenotypes and activate compensatory repair pathways, attenuating long-term responsiveness.140–142
Consequently, combination strategies have therefore become a central focus. Preclinical studies show that combining PARP inhibitors with platinum agents enhances synthetic lethality in BRCA-like TNBC by simultaneously inducing double- and single-strand DNA damage.139,143 Combinations with EZH2, CDK4/6, WEE1, and c-MET inhibitors are under active investigation and exhibit promising synergy, potentially benefiting BRCA-wild-type but HRD-positive (HRD+) tumors.144–150
Despite these advances, limitations remain. Common toxicities include anemia, neutropenia, fatigue, and gastrointestinal reactions, and long-term safety data are still evolving.151 Clinical benefit is largely confined to BRCA-mutated or HRD-positive populations, and variability among HRD testing platforms complicates patient selection. Moreover, the overall survival benefit remains inconsistent across studies, and resistance mechanisms such as partial restoration of homologous recombination repair or increased drug efflux further restrict the durability of treatment responses.152,153
Future priorities include establishing reliable biomarkers for HRD, optimizing combination therapy while minimizing toxicity, and developing improved delivery systems to enhance tumor specificity. These efforts are essential to expand the clinical utility of PARP inhibitors and improve outcomes for both BRCA-mutated and HRD+ breast cancer.
Application of CDK4/6 Inhibitors in ER+ Breast Cancer
Cyclin-dependent kinase 4/6 (CDK4/6) is central to ER+ breast cancer biology through its interaction with cyclin D1, a key downstream target of ER signaling. ER activation increases cyclin D1 expression, promoting CDK4/6-mediated phosphorylation of retinoblastoma (Rb) protein and driving the G1–S transition.154–156 CDK4/6 inhibitors block this phosphorylation, induce G1 arrest, and effectively suppress tumor proliferation.157,158
Beyond classical ER+ disease, CDK4/6 inhibition may also benefit HER2-low breast cancer. Although HER2 expression is reduced, downstream signaling may still activate CDK4/6 via Rb phosphorylation. Elevated pRb levels observed in HER2+ and HER2-low tumors suggest increased cell cycle activity and potential sensitivity to CDK4/6 blockade.159
Resistance remains a major challenge. Mechanisms include RB1 alterations, activation of alternative ER pathways, and aberrant PI3K/AKT/mTOR signaling.160–163 Preclinical data indicate that combining PI3K or mTOR inhibitors with emerging CDK7 inhibitors may overcome resistance and enhance antitumor efficacy.160,164
CDK4/6 inhibitors are especially valuable in endocrine-resistant disease. Endocrine therapy suppresses estrogen-driven transcription, while CDK4/6 inhibitors block downstream cell cycle progression, generating synergistic dual inhibition. This combination is now the first-line standard for metastatic ER+ breast cancer, with trials showing that palbociclib, ribociclib, and abemaciclib combined with endocrine therapy significantly prolong PFS across diverse clinical subgroups.165–168 Ongoing studies are evaluating their role in the adjuvant setting for high-risk early breast cancer.166,169
Although CDK4/6 inhibitors have achieved clinical success by extending progression-free survival in metastatic ER+ breast cancer, several challenges remain in real-world practice. Common toxicities such as neutropenia, fatigue, and gastrointestinal discomfort often necessitate dose reduction or treatment interruption.170,171 The presence of RB1 loss, activation of downstream pathways, or altered cyclin D1 expression can markedly reduce drug sensitivity, and potential indicators remain under investigation and require prospective validation.172 Trials in the adjuvant or early-stage setting have not consistently demonstrated an improvement in overall survival, highlighting the need for careful patient selection. Future work should focus on clarifying resistance mechanisms, developing reliable biomarkers, and optimizing combination strategies, while also considering the long-term costs and follow-up requirements associated with extended combination therapy.
The Oncogenic Role and Therapeutic Potential of WBP2 in Breast Cancer
WW-binding protein 2 (WBP2) acts as an oncogenic transcriptional co-activator integrating multiple signaling cascades, including ER/PR, EGFR, PI3K, Hippo, and Wnt pathways.173 By interacting with β-catenin and YAP/TAZ complexes, WBP2 enhances tumor proliferation, invasion, and progression, and regulates key effectors such as c-Myc and cell-cycle proteins.174–178
Mechanistically, WBP2 translocates to the nucleus upon Wnt3A stimulation, binds β-catenin, and activates TCF-dependent transcription to promote invasive behavior. Its protein levels are regulated by the E3 ubiquitin ligase ITCH, which targets WBP2 for proteasomal degradation. In TNBC, impaired ITCH-mediated degradation leads to WBP2 accumulation, enhanced ERα binding, and upregulation of multidrug resistance protein 1 (MDR1), contributing to chemotherapy resistance.174,179,180 Restoration of ITCH function via the AMOTL2/c-JUN axis re-establishes WBP2 degradation and reverses chemoresistant phenotypes, highlighting a potential therapeutic strategy.
Given its functional link with YAP/TAZ, WBP2 is also modulated by upstream Hippo signaling. Activation of the Hippo/MST pathway suppresses WBP2 through Dicer-mediated miRNA processing, affecting HER2 expression and cell-cycle arrest during trastuzumab treatment.178 Conversely, elevated WBP2 enhances trastuzumab-induced HER2 internalization and cyclin D–dependent cell-cycle arrest, particularly in younger patients (<50 years old), supporting its role as both a functional effector and potential predictive biomarker for HER2-targeted therapy.176
Collectively, WBP2 serves as a central node connecting oncogenic pathways and influencing drug sensitivity, HER2-targeted therapy response, and ubiquitin-dependent degradation, making it a promising target for precision therapy. Most evidence derives from preclinical models, and clinical translation remains limited. Key challenges include incomplete understanding of WBP2 function in normal tissues, potential compensatory mechanisms, and lack of consistent biomarkers for patient selection. Future research should focus on toxicology, pharmacokinetics, early-phase clinical studies, and therapeutic strategies such as miRNA modulation or restoration of ubiquitin-mediated degradation to overcome resistance and improve breast cancer outcomes.
Trop2-Targeted Therapy for Breast Cancer
Trophoblast cell surface antigen 2 (Trop2) is a type I transmembrane glycoprotein that regulates intracellular calcium and activates PI3K/AKT signaling, promoting tumor growth, migration, and survival.181–184 In glioblastoma, Trop2 also engages JAK2/STAT3, MAPK, and Wnt/β-catenin pathways, indicating broad oncogenic involvement. Although it may act as a tumor suppressor in some contexts, these remain poorly understood. In breast cancer, particularly TNBC, Trop2 is widely overexpressed, making it an attractive therapeutic target.185
Structurally, Trop2 comprises extracellular, transmembrane, and intracellular domains. Its extracellular domain forms dimers with more extensive inter-subunit interactions than epithelial cell adhesion molecule (EpCAM), potentially influencing interactions with IGF-1, neuromodulin-1, and αβ integrins to drive tumor progression.186 These structural insights have accelerated the development of Trop2-targeted therapies, especially ADCs.
Clinical progress has been notable, particularly with sacituzumab govitecan (SG), which significantly prolongs PFS and overall survival (OS) in metastatic TNBC compared with standard chemotherapy, although direct head-to-head comparisons are limited.187 Building on these advances, new Trop2-directed ADCs continue to emerge. OBI-992, an anti-Trop2 antibody conjugated to the topoisomerase I inhibitor exatecan via an enzyme-cleavable hydrophilic spacer (DAR 4), demonstrates superior preclinical activity relative to existing agents such as Dato-DXd.188 Given Trop2’s broad expression across breast cancer subtypes, these agents may offer therapeutic utility beyond TNBC.185 Figure 2 illustrates emerging targeted therapeutic strategies in breast cancer.
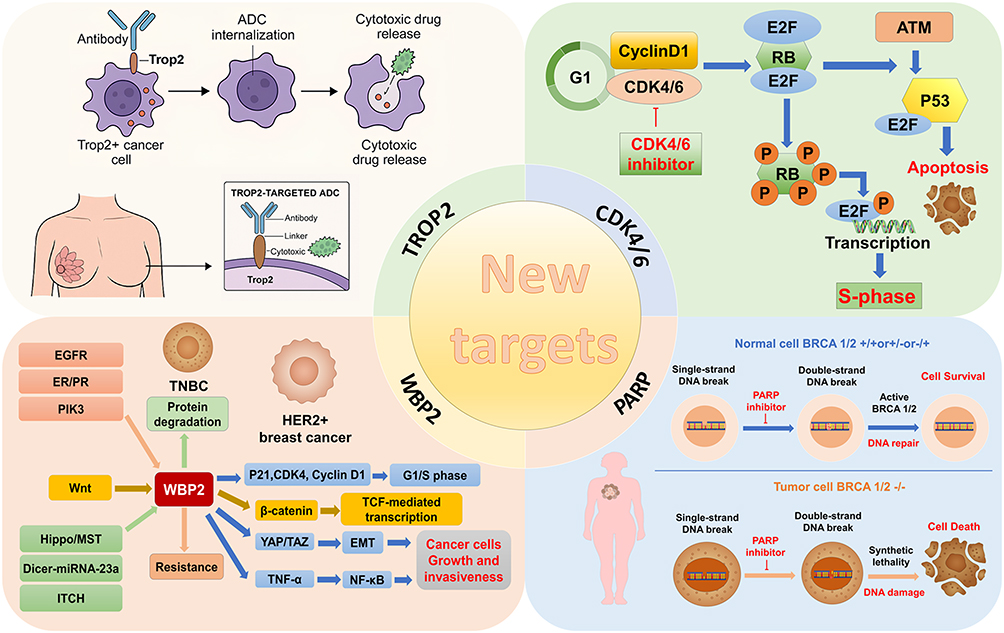 |
Figure 2 Schematic overview of novel target-directed strategies in breast cancer therapy. The figure is divided into four sections highlighting key molecular targets and their therapeutic implications. Trop2 (top left): Trop2-expressing cancer cells are selectively targeted by ADCs, which are internalized upon binding, undergo lysosomal degradation, and release cytotoxic payloads, inducing cancer cell death while sparing normal tissues. CDK4/6 (top right): CDK4/6 inhibitors block cell cycle progression by preventing RB phosphorylation and subsequent E2F activation, thereby arresting cells in G1 phase. These inhibitors also interact with DNA damage response pathways, such as ATM-p53 signaling, influencing therapeutic outcomes. WBP2 (bottom left): WBP2 integrates multiple upstream oncogenic signals, including Wnt/β-catenin, EGFR, ER/PR, PI3K, Hippo/MST, and miRNA-mediated pathways, to promote cell proliferation, EMT, and tumor progression. Modulation of WBP2 protein degradation can enhance therapeutic response and overcome resistance in TNBC. PARP (bottom right): PARP inhibitors exploit synthetic lethality in BRCA1/2-deficient tumors by preventing repair of single-strand DNA breaks, leading to accumulation of double-strand breaks and selective cancer cell death, while normal cells with functional BRCA1/2 maintain genomic integrity. This multidimensional framework illustrates how targeted interventions converge to disrupt key proliferative and survival pathways across breast cancer subtypes. Abbreviations: ADC, Antibody-drug conjugates; Trop2, Trophoblast cell surface antigen 2; CDK4/6, Cyclin-dependent kinase 4/6; RB, Retinoblastoma protein; E2F, E2 promoter-binding factor; ATM, Ataxia Telangiectasia Mutated; ER, Estrogen receptor; SLC7A11, Solute carrier family 7 member 11; WBP2, WW domain-binding protein 2; Wnt, Wingless-related integration site; EGFR, Epidermal growth factor receptor; ER/PR, Estrogen/progesterone receptor; PI3K, Phosphoinositide 3-kinase; MST, Mammalian Ste20-like kinase; TNBC, Triple-negative breast cancer; PARP, Poly-ADP-ribose polymerase; BRCA, Breast cancer susceptibility gene; EMT, Epithelial-mesenchymal transition. |
Despite clinical promise, Trop2-targeted therapies face challenges. Common toxicities include neutropenia, diarrhea, and fatigue, with some severe events requiring hospitalization.189,190 Heterogeneous Trop2 expression and functional variability contribute to differential responses, and single-target strategies may be less effective in tumors with low or heterogeneous expression.189 Nevertheless, the rapid approval of SG and ongoing ADC development underscore Trop2 as a compelling target. Future research should focus on defining biomarker thresholds, optimizing dosing, monitoring toxicity, and evaluating real-world effectiveness, particularly in aggressive, receptor-negative subtypes such as TNBC.
Advanced Targeting Strategies in Breast Cancer Management
Targeted Metabolic Therapeutic Strategies for Breast Cancer
Breast cancer stem cells (BCSCs) drive tumor initiation, progression, metastasis, and therapeutic resistance. As “seed cells,” BCSCs possess self-renewal and differentiation capacity, maintain tumor heterogeneity, and display enhanced migratory and invasive potential, contributing to metastasis to brain, lungs, liver, and bone. Resistance to chemotherapy, endocrine therapy, and radiotherapy stems from enhanced DNA repair, elevated drug efflux, and distinct metabolic states.191–193
BCSCs exhibit substantial metabolic plasticity. Mesenchymal-like (M-type) BCSCs rely on oxidative phosphorylation (OXPHOS), whereas epithelial-like (E-type) BCSCs depend on glycolysis, providing subtype-specific vulnerabilities. Lactate dehydrogenase A (LDHA) maintains stemness via lactate accumulation in the TME.194,195 Simultaneous inhibition of glycolysis and antioxidant pathways selectively eliminates dynamic BCSC populations, highlighting shared metabolic vulnerabilities.196
The TME plays a decisive role in regulating BCSC behavior. Hypoxia is a major driver of stemness maintenance and can stabilize NANOG mRNA through the N6-methyladenosine (m6A) demethylase ALKBH5, thereby promoting stemness.197 Metabolic crosstalk with mesenchymal stem cells activates P2X receptors on BCSCs, enhancing proliferation and metastasis, reversible by purinergic signaling inhibitors.198
Developmental pathways, including Notch, Hedgehog, Hippo, and Wnt, regulate BCSC self-renewal, quiescence, and differentiation. NOTCH4 maintains quiescent M-type BCSCs through SLUG and GAS1, contributing to dormancy and therapy resistance.199 Dysregulated lipid metabolism, including CD200 and CD276 expression and lipid droplet accumulation, supports immune evasion and stemness.200 Transcriptional regulators such as FoxQ1 promote stemness via Wnt/β-catenin and Snail-mediated EMT, and compounds like diallyl trisulfide (DATS) can suppress FoxQ1 and BCSC proliferation.201–203
Autophagy is essential for BCSC survival under therapeutic stress. EpCAM regulates autophagy through glycosylation-dependent PI3K/Akt/mTOR signaling, with deglycosylated EpCAM promoting autophagy and cell proliferation.204–206 Figure 3 illustrates the aforementioned process.These metabolic and signaling mechanisms that sustain BCSC stemness represent potential therapeutic targets.
 |
Figure 3 Mechanistic schematic of deglycosylated EpCAM in modulating breast cancer progression. Deglycosylated EpCAM exerts multifaceted effects on breast cancer cells. It promotes autophagy either directly or via inhibition of the Akt/mTOR signaling axis, and concurrently enhances apoptosis, resulting in a marked reduction in proliferative capacity. Additionally, deglycosylated EpCAM suppresses invasion and metastatic potential by downregulating MAPK and PI3K/Akt pathways, which leads to decreased expression of EMT-associated proteins, including N-cadherin and vimentin. This integrated modulation of survival, growth, and metastatic signaling highlights the critical role of deglycosylated EpCAM in restraining breast cancer progression. Abbreviations: EpCAM, Epithelial Cell Adhesion Molecule; mTOR, mammalian target of rapamycin; AKT, Protein Kinase B; PI3K, Phosphatidylinositol 3-kinase; MAPK, Mitogen-activated protein kinase; EMT, Epithelial-mesenchymal transition. |
However, translating metabolic targeting into clinical practice remains challenging. Many metabolic enzymes and pathways are active in normal tissues, raising concerns about selectivity and systemic toxicity. Evidence supporting metabolic interventions is largely preclinical, and early-phase clinical trials have yielded modest or inconsistent responses. Tumor metabolism is highly adaptive, capable of switching to compensatory pathways, and pronounced metabolic heterogeneity within and across tumors further complicates patient selection. Future progress will require reliable biomarkers, rational combination strategies to prevent metabolic escape, and rigorous clinical studies to define therapeutic windows and long-term safety.
Targeting the Tumor Immune System
Application of Immune Checkpoint Inhibitors
Immune checkpoint inhibitors (ICIs) reactivate antitumor T-cell responses by blocking inhibitory pathways such as PD-1/PD-L1 and CTLA-4. Their efficacy in breast cancer varies across subtypes, highlighting the need to understand subtype-specific immunogenic features and optimize combination strategies.
In HR+/HER2− breast cancer, low tumor-infiltrating lymphocyte (TIL) density and limited PD-L1 expression result in weak immunogenicity and poor ICI responsiveness.207 Combination strategies aim to sensitize these immunologically “cold” tumors. CDK4/6 inhibition with palbociclib enhances antigen presentation and T-cell priming, and when combined with pembrolizumab and the aromatase inhibitor letrozole, produces encouraging antitumor activity in HR+/HER2− metastatic disease.208 These findings emphasize the integration of endocrine therapy and cell cycle inhibition to improve ICI efficacy.
HER2+ tumors are more immunogenic, with higher TIL infiltration and PD-L1 expression, which enhances ICI responsiveness.209,210 HER2-targeted antibodies such as trastuzumab activate natural killer cells via ADCC, facilitating adaptive T-cell responses.211 Moreover, radiotherapy further amplifies HER2-specific cytotoxic T-cell infiltration, strengthening ICI activity in preclinical models.212,213
TNBC exhibits the highest immunogenicity, with abundant TILs and frequent PD-L1 upregulation, making it the most responsive subtype to ICIs.209 Chemotherapy enhances antigen release and presentation while reducing immunosuppressive cells, synergistically improving ICI responses.214–217 Mechanistically, PD-L1 overexpression allows immune evasion; PD-L1 blockade with atezolizumab combined with albumin-bound paclitaxel is approved for PD-L1–positive metastatic TNBC.218 However, therapeutic benefit remains limited to PD-L1–positive populations, and primary or acquired resistance continues to pose clinical challenges.
Emerging strategies explore ICIs in combination with anti-angiogenic agents, PARP inhibitors, or TME modulators to enhance immunogenicity and overcome resistance across subtypes.219,220 These combinatorial approaches aim to enhance immunogenicity, remodel suppressive microenvironments, and overcome intrinsic resistance mechanisms across breast cancer subtypes. Continued refinement of these strategies is essential for maximizing the therapeutic potential of immune checkpoint inhibitors in breast cancer.
Despite these advances, clinical benefit remains largely confined to PD-L1–positive TNBC, with limited efficacy in HR+ disease. Immune-related adverse events, including thyroid dysfunction, colitis, hepatitis, and pneumonitis, can be severe and require immunosuppressive therapy.221,222 Low tumor mutational burden, poor antigenicity, and immunosuppressive TME further limit response, while cost and accessibility constrain broader application. These challenges highlight the need for predictive biomarkers, optimized combination regimens, and rigorous safety management to expand the therapeutic impact of ICIs in breast cancer.
Novel Immunotherapy
Bispecific antibodies (BsAbs) are engineered molecules capable of simultaneously binding tumor-associated antigens on breast cancer cells and immune effectors such as CD3 on T cells, thereby recruiting T cells to tumor cells and inducing antigen-specific cytotoxicity.223–225 Over 200 BsAb constructs, including BsAb drug conjugates, are under preclinical or early clinical investigation across breast cancer subtypes.226,227 Mechanistic studies have expanded applications beyond direct cytotoxicity; for example, IL-6/IL-8 dual-receptor–targeting BsAbs suppress migration and metastasis, while DB-1419 (B7-H3×PD-L1) BsAbs combine immune checkpoint blockade with direct tumor targeting.228 Despite these promising directions, BsAb development faces several challenges. Complex structural formats, such as asymmetric IgG-like molecules, complicate large-scale manufacturing, whereas smaller constructs, such as Fab fragments, often display reduced half-lives and suboptimal pharmacokinetic properties.229
BsAb research laid the conceptual foundation for chimeric antigen receptor T-cell (CAR-T) therapy, which genetically modifies patient T cells to express tumor antigen–binding receptors fused to activation domains such as CD3ζ, enabling continuous T-cell activation independent of MHC presentation.224,230 CAR-T therapy has achieved clinical success in hematologic malignancies through standardized workflows such as viral transduction and ex vivo expansion. Solid tumors, including breast cancer, pose additional challenges due to heterogeneous antigen expression, variable target distribution, and an immunosuppressive microenvironment.231,232 Antigen loss through epigenetic reprogramming or clonal selection, including CD19-negative relapse in hematologic malignancies and antigen-negative outgrowth in glioblastoma, further limits CAR-T efficacy.233–237
Dual-targeting strategies, exemplified by CD79a/b in B-cell malignancies, demonstrate the potential to mitigate antigen escape, yet dynamic antigen plasticity in solid tumors remains a major challenge. Emerging approaches combine CAR-T and BsAb strategies. For instance, dual-target CAR-T cells recognizing EpCAM and intercellular adhesion molecule-1 (ICAM-1) enhance antigen coverage and tumor suppression, while p95HER2-CAR-T combined with HER2/CD3 BsAbs achieved complete tumor regression in HER2-positive breast cancer models.238 Fine-tuning BsAb affinity can further guide CAR-T cells toward high-antigen tumors while minimizing off-tumor toxicity.
Despite progress, solid tumor microenvironments limit engineered immunotherapies. TGF-β and dense stroma restrict T-cell infiltration and persistence, contributing to T-cell exhaustion and antigen escape.239 Toxicities such as cytokine release syndrome and neurotoxicity further constrain treatment. BsAbs and cancer vaccines face similar obstacles, including short half-lives, on-target off-tumor activity, and limited antigenicity. Most evidence remains preclinical or early-phase, and uncertainties persist regarding feasibility, manufacturing, scalability, and long-term safety. Future research should focus on integrating BsAbs, CAR-T engineering, and microenvironment-modulating strategies to enhance tumor penetration, expand antigen coverage, and broaden the therapeutic window, thereby improving clinical efficacy and durability in breast cancer.
Targeting the Tumor Microenvironment
The breast cancer TME comprises heterogeneous cellular and non-cellular components, including suppressor myeloid cells, tumor-associated macrophages (TAMs), cancer-associated fibroblasts (CAFs), extracellular matrix (ECM), and inflammatory mediators, collectively driving tumor progression, immune suppression, and therapeutic resistance.240–244 Therapeutic strategies targeting the TME aim to reverse immunosuppression, enhance intratumoral drug delivery, and overcome therapy resistance.
Reversing immune suppression is central to improving therapeutic efficacy. Tumor-derived granulocyte–macrophage colony-stimulating factor (GM-CSF) upregulates ARG1 expression in myeloid cells via STAT3, p38 MAPK, and cAMP signaling under acidic conditions, promoting local immunosuppression. Blocking GM-CSF reduces ARG1 expression, restoring antitumor immunity and enhancing T-cell–based therapies and immune checkpoint inhibitor responses.242
Structural and biophysical barriers within the TME also limit drug delivery. Dense ECM, elevated interstitial pressure, and poor perfusion impede penetration of conventional therapies, while tumor acidity can be exploited for pH-responsive drug release. Nanotechnology-based strategies, including hypoxia- or esterase-responsive nanomicelles, functionalized liposomes, and sonodynamic therapy (SDT)-assisted delivery, enhance spatiotemporal drug release, prolong intratumoral retention, and improve therapeutic efficacy.240,245–252
Given the high heterogeneity of the TME, precision oncology increasingly relies on high-resolution profiling. Eftekhari et al demonstrated that dynamic remodeling of CD4+ T-cell subsets (Treg, Th1, Th2, Th17), inflammatory cytokines (TNF-α, IL-6), and immune mediators (FASL, IDO, SOCS1, VEGF, CCR7) correlates with breast cancer progression.253 Single-cell RNA sequencing by Yang et al identified regulatory T cells, M2 macrophages, angiogenesis-associated mast cells, DNA-repair-deficient stem-like cells, angiogenic endothelial cells, collagen-producing fibroblasts, and proliferative epithelial cells as key drivers of a pro-tumorigenic interface zone.254 These findings highlight the cellular complexity that shapes tumor behavior and therapeutic response.
Despite advances in understanding the tumor microenvironment, TME-targeted therapies face substantial translational challenges. Conventional two-dimensional models inadequately recapitulate physiological conditions, emphasizing the need for advanced three-dimensional models, while integration of multi-omics data, including genomics, transcriptomics, proteomics, and metabolomics, is essential to map dynamic cellular interactions and identify therapeutic vulnerabilities.255–259 Clinically, approaches such as ECM remodeling, CAF depletion, or vascular normalization have shown only variable or transient benefits, sometimes disrupting stromal homeostasis or producing short-lived effects. Lack of predictive biomarkers and off-tumor toxicity further limit efficacy, highlighting the need for combination strategies rather than single-target interventions.
Overall, integrating TME characterization with precision medicine offers a promising path for breast cancer therapy. Approaches that combine microenvironmental modulation, personalized therapeutic selection, and multi-omics profiling may enhance treatment efficacy and improve clinical outcomes.
Novel Drug Delivery Systems
Nanotechnology Innovation
Nanotechnology has gained substantial traction in breast cancer management through diverse platforms, including liposomes, polymeric nanoparticles, and inorganic nanostructures, which function as drug carriers to overcome key limitations of conventional chemotherapy. These systems improve drug stability, enhance tumor-specific targeting, enable controlled or stimuli-responsive release, facilitate delivery of hydrophobic agents, and reduce toxicity to normal tissues.260–264 Increasingly, nanocarriers are being explored for combination therapies, gene silencing, and diagnostic applications.
Kahlous et al engineered gold nanoparticle–based carriers, including chitosan–citrate–tamoxifen nanocomposites and layered gold nanoparticle systems, demonstrating enhanced antiproliferative activity against MCF-7 cells, supporting potential synergy with endocrine therapy.7 Wang et al developed a biomimetic dual-targeted protein nanocage with three-dimensional drug-loading capacity, illustrating how rational nanoplatform design can improve delivery efficiency, enhance therapeutic synergy, and mitigate resistance.265
Nanocarriers have also advanced gene-silencing strategies. Yang et al designed an siRNA system targeting BCSG1, achieving effective molecular-level gene knockdown.266 Similarly, Alipoor et al developed DNAi peptide nanohybrids against BCL-2, inducing apoptosis in breast cancer cells.267 To overcome siRNA instability and poor bioavailability, hybrid systems combining liposomes with mesoporous silica nanoparticles co-delivering siRNA and doxorubicin have been proposed, enhancing intracellular accumulation and overcoming multidrug resistance.268
In diagnostics, nanotechnology enables sensitive detection of early disease biomarkers. Cheng et al employed surface-enhanced Raman spectroscopy (SERS) with silver nanoparticle substrates to detect serum biomarkers with high sensitivity.269 In addition, Heikal et al developed atorvastatin-loaded pumpkin seed oil nanovesicles delivered via microneedles, providing localized therapy while minimizing systemic toxicity.
Despite these advances, clinical translation of nanocarriers faces challenges, including reproducibility, long-term biosafety, scalable manufacturing, regulatory hurdles, and variable intratumoral distribution in heterogeneous microenvironments. Combining nanotechnology with targeted therapies or immunotherapy represents a promising direction to overcome resistance and improve precision in breast cancer treatment.
Ultrasound Delivery Technology
Ultrasound-mediated drug delivery has emerged as a promising targeted therapeutic strategy in breast cancer by leveraging acoustic energy, mechanical forces, and cavitation to enhance intratumoral drug penetration while minimizing systemic toxicity. Among these, ultrasound-targeted microbubble cavitation (UTMC) transiently disrupts tumor vasculature: upon ultrasound activation, microbubbles (MBs) or nanobubbles (NBs) undergo inertial cavitation, widening endothelial gaps, increasing vascular permeability, and promoting deeper drug distribution.270,271 Using microbubble/microdroplet clusters (PS101), J. Yu et al demonstrated enhanced drug accumulation and improved therapeutic outcomes in breast cancer models.252
Ultrasound also facilitates drug loading and targeted release. Italiya et al developed a microbubble-based system delivering virus-sensitized therapeutics, where ultrasonic cavitation enhanced intratumoral deposition of oncolytic agents in 4T1 breast cancer.272 Beyond mechanical effects, ultrasound-generated thermal energy and transient membrane permeabilization improve drug internalization, with kinetic models confirming controllable release profiles under acoustic stimulation.273,274
Recent innovations extend ultrasound applications beyond drug delivery. Wang et al engineered a system activating plant chlorophyll–based sonosensitizers (C34), generating reactive oxygen species, triggering immunogenic cell death, suppressing primary tumor growth, and reducing metastatic spread.273 Additionally, responsive nanocarriers such as ultrasound-triggered liposomes and polymeric nanoparticles enable site-specific release, reducing off-target effects. Li et al evaluated IR-780 iodide as a sonodynamic therapy agent, where ultrasound exposure induced necrotic and apoptotic cell death in 4T1 cells via reactive oxygen species.275 Acoustic-responsive nanobubbles further integrate diagnostic imaging and therapeutic enhancement, while approaches like acoustic droplet vaporization and phase-change nanomaterials improve spatial precision and efficacy.276–279
Despite these advantages, including non-invasive activation, real-time imaging, and controllable release, clinical translation remains limited due to technical variability, incomplete long-term safety data, and lack of standardized protocols. Future directions include optimizing multimodal combinations with immunotherapy, developing intelligent acoustically responsive carriers, and establishing robust clinical validation frameworks.
Exosome Delivery
Exosomes have emerged as versatile platforms for breast cancer diagnosis and therapy due to their unique biochemical composition, intrinsic stability, and natural capacity for intercellular communication. Carrying proteins, nucleic acids, lipids, and metabolites, exosomes reflect the molecular profile of tumor cells and enable minimally invasive biomarker detection. MUC1, a membrane-bound glycoprotein linked to breast cancer progression, is enriched on tumor-derived exosomes and correlates with metastatic potential.280 Rui et al developed an electrochemiluminescent aptamer sensor to quantify exosomal MUC1 across breast cancer subtypes, offering a sensitive tool for early detection and molecular monitoring.281
Beyond diagnosis, exosomes serve as therapeutic carriers. Their lipid bilayer confers high biocompatibility, low immunogenicity, and favorable biodistribution, allowing efficient delivery of chemotherapeutics, proteins, and nucleic acids such as siRNA, circRNA, and mRNA.282,283 Compared with synthetic nanocarriers, exosomes exhibit superior stability and reduced off-target accumulation. Zheng et al highlighted their dual role in regulating breast cancer initiation, progression, metastasis, and treatment resistance, emphasizing their potential as both vehicles and therapeutic targets.284
Exosomes also modulate the tumor microenvironment (TME). Hypoxia-induced exosomes promote angiogenesis via SNHG1, while exosome-loaded siRNA against SNHG1 can reverse this effect. They enhance intracellular retention of chemotherapeutics and reduce drug efflux in resistant cells. Engineered exosomes are being explored for immunomodulation; Niu et al proposed exosomes carrying chimeric antigen receptors as a cell-free alternative to CAR-T therapy, potentially reducing cytokine toxicity and improving tumor infiltration.285
Despite these advantages, challenges remain, such as large-scale isolation, consistent purification, efficient cargo loading, and predictable pharmacokinetics. Continued refinement of exosome characterization and manufacturing technologies is essential for clinical translation. Their multifunctional roles as biomarkers and drug carriers position exosomes as promising components of next-generation breast cancer therapeutics. Figure 4 illustrates emerging exosome-based targeted strategies.
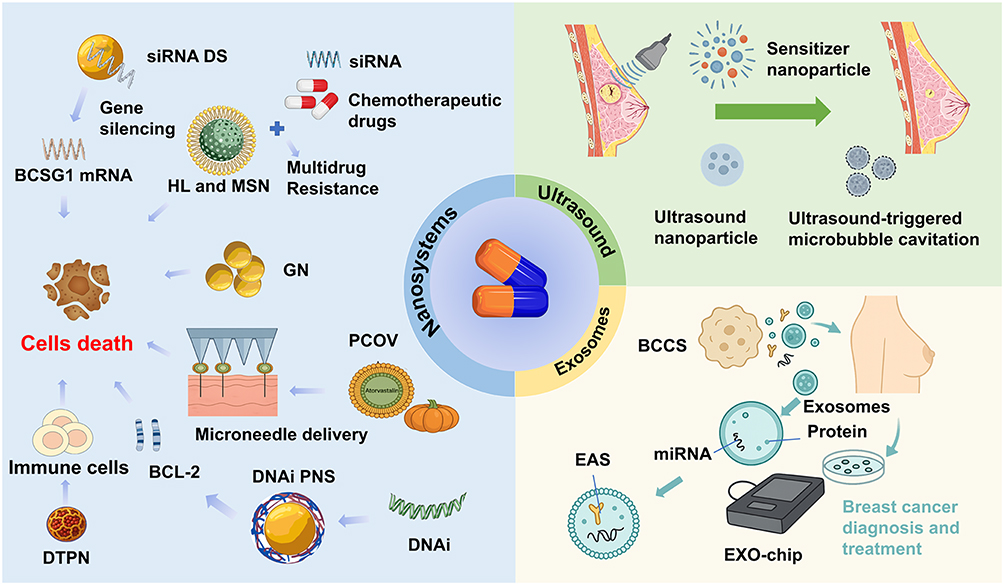 |
Figure 4 Schematic of emerging drug delivery platforms for breast cancer therapy. This schematic illustrates innovative strategies that enhance the precision, efficacy, and selectivity of anticancer interventions. Nanosystems: These include multifunctional nanoparticles that co-deliver siRNA and chemotherapeutics to overcome multidrug resistance, targeted nanoparticles such as HL, MSN, GN, and PCOV to induce selective cancer cell death, microneedle arrays enabling localized delivery of BCL-2 inhibitors and DNA interference agents, and DTPN that both elicit tumor cell apoptosis and activate antitumor immunity. Ultrasound-assisted systems: Techniques leveraging ultrasound-mediated microbubble cavitation enhance drug permeability and therapeutic penetration, while ultrasound-responsive nanoparticles facilitate spatiotemporal control of drug and gene delivery. Exosome-based approaches: Breast cancer cells secrete exosomes containing proteins and miRNAs that can serve as diagnostic biomarkers; these vesicles can be captured and analyzed via EAS or EXO chips, and engineered exosomes offer promising platforms for targeted therapeutic delivery. Collectively, these systems exemplify how advanced nanotechnology, acoustic modulation, and extracellular vesicle engineering converge to overcome biological barriers, maximize tumor-specific effects, and enable personalized breast cancer treatment. Abbreviations: BCCS, Breast cancer cells; DTPN, Dual-targeted protein nanoparticles; EAS, Electrochemiluminescent aptamer sensor; HL, Hybrid liposomes; MSN, Mesoporous silica nanoparticles; PCOV, Pumpkin seed oil vesicles; siRNA DS, siRNA delivery systems; GN, Gold nanoparticles; DNAi PNS, DNAi peptide nanocomposites; BCL-2, B-cell lymphoma 2; DNAi, DNA interference. |
Combined Treatment Strategies
Synergistic Effects of Autophagy Regulation and Targeted Therapy
Autophagy plays a context-dependent role in breast cancer, acting both as a tumor suppressor by removing damaged cellular components and as a survival mechanism that enables cancer cells to withstand metabolic and therapeutic stress.286,287 This duality positions autophagy as a pivotal regulator of treatment efficacy, and small-molecule modulators have shown promise. Liu et al identified LZ02/01, which suppresses breast cancer cell proliferation by impairing autophagic flux and promoting apoptosis, while You et al reported that St. John’s Wort induces pro-death autophagy and apoptosis in TNBC, demonstrating dual-pathway potential.288,289
Modulating autophagy can synergize with targeted therapies to overcome resistance. Paclitaxel resistance in TNBC is promoted by neddylation-activated TRIM25 driving TFEB-mediated autophagy, and inhibition of this pathway restores drug sensitivity.290 Autophagy also intersects with tumor immunoregulation: inhibition of glutamine metabolism elevates reactive oxygen species (ROS), inducing autophagy and enhancing antitumor immune responses, suggesting integration with immune checkpoint blockade may improve efficacy.291 Nanotechnology offers additional avenues, as NIR-activated nanogenerators can trigger autophagy-related signaling while inducing immunogenic cell death, suppressing tumor growth and metastasis.292 Epigenetic modulation further regulates autophagy-related pathways: Chidamide, a histone deacetylase inhibitor (HDACi), modulates BTG1 and FOXO1 and, in combination with androgen receptor (AR) antagonists, destabilizes AR signaling, enhancing therapeutic response in luminal androgen receptor (LAR)–type TNBC.293
Despite these advances, clinical translation faces challenges. Autophagy is highly context-dependent, and indiscriminate modulation may have pro-survival effects in tumors or normal tissues. Many findings derive from preclinical models that may not capture human breast cancer complexity, including metabolic plasticity and immune interactions. Agents often lack specificity and can affect parallel pathways, complicating toxicity and dosing, while predictive biomarkers remain limited.
Collectively, these findings underscore that strategic modulation of autophagy, either through inhibition or induction, can enhance the effectiveness of chemotherapy, immunotherapy, nanomedicine, and epigenetic therapies, providing a versatile framework for overcoming resistance and improving breast cancer treatment outcomes.
JAM-A/LAT1 Dual-Target Combination Therapy for Breast Cancer
Junctional adhesion molecule-A (JAM-A) is a core tight-junction protein whose expression and stability are closely linked to HER2 signaling in breast cancer.294 LAT1 (L-type amino acid transporter 1) mediates essential amino acid uptake, activates mTORC1, and promotes proliferation, invasion, and metastasis.295 Clinically, high LAT1 expression correlates with advanced stage, lymph node involvement, and poor response to trastuzumab in HER2+ metastatic patients, highlighting its role in resistance.296
Mounting evidence indicates a synergistic oncogenic relationship between JAM-A and LAT1. Co-expression associates with poor prognosis, especially in subtype B and HER2+ disease.296–298 Mechanistic studies show that JAM-A regulates mTORC1 signaling by controlling LAT1 expression, affecting protein synthesis and proliferation, particularly in estrogen-deprivation-resistant (EDR) breast cancer models.299 Proteomics analyses reveal that JAM-A forms a membrane complex with LAT1, enhancing amino acid uptake and tumor growth.298
Preclinical studies demonstrate therapeutic potential for dual targeting. JAM-A extracellular blockade reduces proliferation by >60%, and combining JAM-A inhibition with the LAT1 inhibitor JPH203 markedly enhances efficacy. In 4T1–BALB/c models, dual blockade reduced tumor volume by 72% compared with monotherapies.298,300,301 At the resistance level, JAM-A also stabilizes HER2/HER3, contributing to therapy resistance; combining JAM-A/LAT1 inhibition with HER2 downregulation reverses aromatase inhibitor resistance.
Dual-target intervention additionally modulates the tumor immune microenvironment. LAT1 inhibition reshapes amino-acid–dependent metabolic pathways in immune cells, enhancing PD-1 responsiveness, while JAM-A deletion reduces regulatory T-cell infiltration. In TNBC, combining the JAM-A antagonist peptide P4D with LAT1 silencing reduced lung metastasis by 89%, demonstrating strong synergy against metastatic spread.301–303
Overall, JAM-A/LAT1 dual targeting holds promise for AI-resistant or trastuzumab-resistant subtype B/HER2+ breast cancer and high-tumor-mutation-burden TNBC, where junctional signaling and metabolic rewiring drive progression and immune evasion. However, translation challenges remain. Current evidence is largely preclinical, and co-dependence of JAM-A and LAT1 across heterogeneous human tumors is uncertain. Both proteins are physiologically essential, raising safety concerns. LAT1 expression varies across subtypes, and predictive biomarkers are lacking. Integrating dual targeting with endocrine therapy, HER2 blockade, or immunotherapy introduces further complexity in toxicity, sequencing, and cost. Early-phase clinical studies are needed to define therapeutic windows, long-term safety, and feasibility.
Cutting-Edge Research Directions
Multi-Target Prediction Model
Multi-target prediction models in breast cancer are designed to simultaneously forecast multiple biomarkers or clinical outcomes, integrating conventional clinical factors with advanced machine learning and multi-omics data. Compared with single-target models, these frameworks leverage diverse inputs, including pathological slides and multi-modal imaging, for a more comprehensive disease assessment. Models based on multi-modal imaging have been shown to outperform unimodal approaches.304 For instance, Song et al developed a multiparametric ultrasound radiomics model capable of predicting postoperative recurrence risk and molecular subtypes, providing non-invasive guidance for diagnosis and therapeutic planning.305
In surgical risk prediction, the Breast Cancer Surgery Risk Calculator (BCSRc) uses patient-specific factors from the NSQIP database to estimate postoperative complication probabilities, aiding preoperative risk stratification and clinical decision-making.306 At the molecular level, multi-target frameworks can integrate cellular and pathway-level mechanisms to identify therapeutic targets and combination strategies. Qayoom et al analyzed molecular interactions affected by Bruginin using TCGA, STRING, and AutoDock Vina, establishing a basis for predictive modeling that links molecular networks with drug-target relationships.307
From a technical perspective, machine learning algorithms such as random forests and naive Bayes, along with deep learning approaches like convolutional neural networks (CNNs), are commonly employed. DeepProg, proposed by Poirion et al, combines deep learning with machine learning to integrate multi-omics data and achieve robust prognostic modeling in breast cancer datasets.308 Additionally, Jain et al applied multivariable logistic regression and predictive margin models to identify sociodemographic predictors of treatment delays exceeding 90 days in young female patients, emphasizing the need to incorporate social determinants into predictive frameworks.309
Despite their promise, multi-target models face challenges for clinical translation. Most rely on retrospective datasets with limited annotation, introducing selection bias and reducing generalizability. Multi-omics integration adds complexity due to batch effects, variable data resolution, and inconsistent preprocessing. Deep learning–based models offer strong predictive performance but limited interpretability, which impedes clinical adoption and regulatory approval. Data privacy concerns, lack of cross-institutional standardization, and resource-intensive prospective validation further restrict implementation. Future advances will require harmonized data-sharing, rigorous external validation, and interpretable AI architectures that can be integrated seamlessly into clinical workflows.
miRNA Regulatory Network
In breast cancer, miRNA regulatory networks critically influence tumor progression, invasion, metastasis, and drug resistance. The EGFR-driven cell cycle pathway is central to proliferation and pathogenesis. Uhlmann et al integrated large-scale miRNA screening with proteomics to identify three novel tumor-suppressor miRNAs, miR-124, miR-147, and miR-193a-3p, which collectively target proteins in the EGFR-driven cell cycle network and demonstrate the broad regulatory impact of miRNAs on proliferation.310
Regarding chemotherapy resistance, MDR refers to the phenomenon whereby cancer cells develop cross-resistance to multiple chemotherapeutic agents with distinct structures and mechanisms. Zhang’s team showed that β-limonene reverses resistance to doxorubicin and docetaxel by regulating MDR-associated miRNAs, affecting targets such as PTEN and P-glycoprotein.311 In diagnosis and subtype classification, deep sequencing has revealed subtype-specific miRNA dysregulation. Chang et al identified 25 miRNA biomarkers distinguishing TNBC from normal tissue, while Xu et al constructed miRNA–TF–mRNA regulatory networks to classify subtypes and guide personalized therapy.312,313
In prognosis, Lirussi et al reported a Let-7–SMUG1 network, with specific miRNA–mRNA interactions correlating with favorable outcomes in ER+ breast cancer.314 Zhang et al built a lncRNA–miRNA–mRNA interaction network identifying miR-510 as a central node, and Sarkar et al applied feature selection with survival analysis to pinpoint core miRNAs associated with subtypes and prognosis.315,316
Despite these advances, clinical translation is limited by context-dependent miRNA expression, intra-tumor heterogeneity, reliance on computational predictions, and challenges in therapeutic delivery due to off-target effects and pleiotropy.317 Furthermore, the lack of standardized pipelines for miRNA detection, normalization, and network construction complicates reproducibility across studies. These limitations underscore the need for integrative multi-layer validation strategies and improved delivery platforms before miRNA-directed therapies can reach clinical applicability.
Overall, miRNAs exert multi-layered control over breast cancer initiation, progression, drug resistance, and prognosis. Integrating bioinformatics and machine learning enhances mechanistic insights and supports the development of novel miRNA-directed therapeutic strategies.
Conclusion and Prospect
Breast cancer therapeutics are converging toward a treatment paradigm in which established targeted strategies remain foundational, yet their long-term effectiveness increasingly depends on overcoming the biological constraints that limit durable responses. Endocrine therapy, HER2-directed agents, and inhibitors of CDK4/6 and PARP have produced substantial survival gains, but their efficacy is frequently compromised by tumor heterogeneity, metabolic and immune suppression within the microenvironment, and the emergence of adaptive resistance. As this review highlights, these challenges underscore an urgent need for therapeutic concepts that extend beyond single-pathway inhibition toward integrated approaches capable of modifying the tumor ecosystem as a whole.
Recent advances outline several promising directions. Metabolic targeting, regulation of cancer stem cells, and modulation of autophagy offer new routes to address refractory disease, while emerging molecular axes such as WBP2, Trop2 and the JAM A–LAT1 pathway provide additional leverage to disrupt resistance networks. Progress in drug delivery science, including nanocarriers, ultrasound-responsive formulations and engineered exosomes, further suggests that enhancing intratumoral access may be as crucial as improving intrinsic drug potency. Ultrasound-assisted nanobubbles exemplify this shift by enabling spatial and temporal control of drug release, although their translation will require rigorous validation of long-term safety, scalable production and biomarker-driven patient selection.
Taken together, the evidence reviewed in this work points toward a future in which effective breast cancer treatment relies on the coordinated use of multi omic profiling, imaging-based monitoring and artificial intelligence to align therapeutic decisions with the evolving biology of each tumor. Such convergence is expected to yield treatment strategies that are more precise, more resilient to resistance and ultimately more compatible with the complex architecture of breast cancer.
Funding
This work was supported by National Natural Science Foundation of China (Grant Nos. U23A20398; Grant Nos. 82030007), Noncommunicable Chronic Diseases-National Science and Technology Major Project (Grant Nos. 2024ZD0537707),Open Research Projects of the Ministry of Education Innovation Center for Basic Medical Research on Metabolic Cardiovascular Diseases (Grant Nos. xnykdxcxzx-2024-13).
Disclosure
The authors declare that they have no competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
References
1. Lau KH, Tan AM, Shi Y. New and emerging targeted therapies for advanced breast cancer. Int J Mol Sci. 2022;23(4):2288. doi:10.3390/ijms23042288
2. Okumura S, Hirano Y, Komatsu Y. Inhibition of breast cancer cell proliferation with anti-microRNA oligonucleotides flanked by interstrand cross-linked duplexes. Nucleosides Nucleotides Nucleic Acids. 2020;39(1–3):225–28. doi:10.1080/15257770.2019.1671595
3. Gautam P, Karhinen L, Szwajda A, et al. Identification of selective cytotoxic and synthetic lethal drug responses in triple negative breast cancer cells. Mol Cancer. 2016;15(1):34. doi:10.1186/s12943-016-0517-3
4. Mistry T, Nath A, Pal R, et al. Emerging futuristic targeted therapeutics: a comprising study towards a new era for the management of TNBC. Am J Clin Oncol. 2024;47(3):132–148. doi:10.1097/COC.0000000000001071
5. Liu Y, He E, Zhang Y, et al. WW domain binding protein 2 (WBP2) as an oncogene in breast cancer: mechanisms and therapeutic prospects-a narrative review. Gland Surg. 2022;11(12):1984–2002. doi:10.21037/gs-22-716
6. Chu D, Lu J. Novel therapies in breast cancer: what is new from ASCO 2008. J Hematol Oncol. 2008;1:16. doi:10.1186/1756-8722-1-16
7. Kahlous Y, PALANIRAJAN VK, STARLIN M, et al. Preparation and characterization of chitosan and inclusive compound-layered gold nanocarrier to improve the antiproliferation effect of tamoxifen citrate in colorectal adenocarcinoma (Caco-2) and breast cancer (MCF-7) cells. Turk J Pharm Sci. 2022;19(4):391–399. doi:10.4274/tjps.galenos.2021.48961
8. Zheng J, Hao H. The importance of cancer-associated fibroblasts in targeted therapies and drug resistance in breast cancer. Front Oncol. 2023;13:1333839. doi:10.3389/fonc.2023.1333839
9. Jacobs AT, Martinez Castaneda-Cruz D, Rose MM, et al. Targeted therapy for breast cancer: an overview of drug classes and outcomes. Biochem Pharmacol. 2022;204:115209. doi:10.1016/j.bcp.2022.115209
10. Denkert C, Seither F, Schneeweiss A, et al. Clinical and molecular characteristics of HER2-low-positive breast cancer: pooled analysis of individual patient data from four prospective, neoadjuvant clinical trials. Lancet Oncol. 2021;22(8):1151–1161. doi:10.1016/S1470-2045(21)00301-6
11. El-Sahli S, Hua K, Sulaiman A, et al. A triple-drug nanotherapy to target breast cancer cells, cancer stem cells, and tumor vasculature. Cell Death Dis. 2021;12(1):8. doi:10.1038/s41419-020-03308-w
12. Gupta N, Mohan CD, Shanmugam MK, et al. CXCR4 expression is elevated in TNBC patient derived samples and Z-guggulsterone abrogates tumor progression by targeting CXCL12/CXCR4 signaling axis in preclinical breast cancer model. Environ Res. 2023;232:116335. doi:10.1016/j.envres.2023.116335
13. Zhu Y, Zhu X, Wei X, et al. HER2-targeted therapies in gastric cancer. Biochim Biophys Acta Rev Cancer. 2021;1876(1):188549. doi:10.1016/j.bbcan.2021.188549
14. Cong Y, Zhang S-Y, Li H-M, et al. A truncated DNA aptamer with high selectivity for estrogen receptor-positive breast cancer cells. Int J Biol Macromol. 2023;252:126450. doi:10.1016/j.ijbiomac.2023.126450
15. Xu Z, Yang Y, Li B, et al. Checkpoint suppressor 1 suppresses transcriptional activity of ERalpha and breast cancer cell proliferation via deacetylase SIRT1. Cell Death Dis. 2018;9(5):559. doi:10.1038/s41419-018-0629-3
16. Medford A, Spring LM, Moy B, et al. Antibody drug conjugates for patients with breast cancer. Curr Probl Cancer. 2021;45(5):100795. doi:10.1016/j.currproblcancer.2021.100795
17. Vargas C, Aguirre-Ducler A, Cereceda K, et al. CD44 marks dormant tumor cells after HER2 inhibition in breast cancer cells. Int J Mol Sci. 2025;26(10):4907. doi:10.3390/ijms26104907
18. Lan J, Wang L, Cao J, Wan Y, Zhou Y. circBRAF promotes the progression of triple-negative breast cancer through modulating methylation by recruiting KDM4B to histone H3K9me3 and IGF2BP3 to mRNA. Am J Cancer Res. 2024;14(5):2020–2036. doi:10.62347/OOLG5765
19. Carvalho E, Canberk S, Schmitt F, et al. Molecular subtypes and mechanisms of breast cancer: precision medicine approaches for targeted therapies. Cancers. 2025;17(7):1102. doi:10.3390/cancers17071102
20. Ehab M, Elbaz M. Profile of palbociclib in the treatment of metastatic breast cancer. Breast Cancer. 2016;8:83–91. doi:10.2147/BCTT.S83146
21. Jiang H, Li Z, Xu W, et al. WWP1 targeting PTEN for polyubiquitination to promote bone metastasis of luminal breast cancer. Sci Rep. 2024;14(1):29950. doi:10.1038/s41598-024-81541-5
22. Zhu M, Lin J, Liu H, et al. Construction and validation of acetylation-related gene signatures for immune landscape analysis and prognostication risk prediction in luminal breast cancer. Cancer Cell Int. 2025;25(1):287. doi:10.1186/s12935-025-03920-w
23. Duan Y, Huo D, Gao J, et al. Ubiquitin ligase RNF20/40 facilitates spindle assembly and promotes breast carcinogenesis through stabilizing motor protein Eg5. Nat Commun. 2016;7(1):12648. doi:10.1038/ncomms12648
24. Han Y, Katayama S, Futakuchi M, et al. Targeting c-jun is a potential therapy for luminal breast cancer bone metastasis. Mol Cancer Res. 2023;21(9):908–921. doi:10.1158/1541-7786.MCR-22-0695
25. Wang J, Li D, Shen W, et al. RHAMM inhibits cell migration via the AKT/GSK3beta/Snail axis in luminal A subtype breast cancer. Anat Rec. 2020;303(9):2344–2356. doi:10.1002/ar.24321
26. Barton M, Santucci-Pereira J, Vaccaro OG, et al. BC200 overexpression contributes to luminal and triple negative breast cancer pathogenesis. BMC Cancer. 2019;19(1):994. doi:10.1186/s12885-019-6179-y
27. Cano-Sanchez J, Murillo-González FE, de Jesús-Aguilar J, et al. The aryl hydrocarbon receptor ligand 6-formylindolo(3,2-b)carbazole promotes estrogen receptor alpha and c-fos protein degradation and inhibits MCF-7 cell proliferation and migration. Pharmacology. 2023;108(2):157–165. doi:10.1159/000527993
28. Telang NT, Li G, Katdare M, et al. The nutritional herb epimedium grandiflorum inhibits the growth in a model for the luminal A molecular subtype of breast cancer. Oncol Lett. 2017;13(4):2477–2482. doi:10.3892/ol.2017.5720
29. Cocco S, Leone A, Piezzo M, et al. Targeting autophagy in breast cancer. Int J Mol Sci. 2020;21(21):7836. doi:10.3390/ijms21217836
30. Wang Y, Shen S-Y, Liu L, et al. Jolkinolide B inhibits proliferation or migration and promotes apoptosis of MCF-7 or BT-474 breast cancer cells by downregulating the PI3K-Akt pathway. J Ethnopharmacol. 2022;282:114581. doi:10.1016/j.jep.2021.114581
31. Cuyas E, Gumuzio J, Verdura S, et al. The LSD1 inhibitor iadademstat (ORY-1001) targets SOX2-driven breast cancer stem cells: a potential epigenetic therapy in luminal-B and HER2-positive breast cancer subtypes. Aging. 2020;12(6):4794–4814. doi:10.18632/aging.102887
32. Orea-Soufi A, Castillo-Lluva S, Salvador-Tormo N, et al. The pseudokinase TRIB3 negatively regulates the HER2 receptor pathway and is a biomarker of good prognosis in luminal breast cancer. Cancers. 2021;13(21):5307. doi:10.3390/cancers13215307
33. Ozaki A, Motomura H, Tamori S, et al. High expression of p62 and ALDH1A3 is associated with poor prognosis in luminal B breast cancer. Anticancer Res. 2022;42(7):3299–3312. doi:10.21873/anticanres.15818
34. Cavallaro PA, De Santo M, Belsito EL, et al. Peptides targeting HER2-positive breast cancer cells and applications in tumor imaging and delivery of chemotherapeutics. Nanomaterials. 2023;13(17):2476. doi:10.3390/nano13172476
35. Murase Y, Nanjo S, Ueda T, et al. Mechanisms of resistance to antibody-drug conjugates in cancers. Respir Investig. 2025;63(4):693–698. doi:10.1016/j.resinv.2025.05.012
36. Randall M, Akers R, Rao R. A review of current and future antibody drug conjugates in breast cancer. Curr Treat Options Oncol. 2024;25(12):1506–1516. doi:10.1007/s11864-024-01276-3
37. Shin I. HER2 signaling in breast cancer. Adv Exp Med Biol. 2021;1187:53–79.
38. Stanowicka-Grada M, Senkus E. Anti-HER2 drugs for the treatment of advanced HER2 positive breast cancer. Curr Treat Options Oncol. 2023;24(11):1633–1650. doi:10.1007/s11864-023-01137-5
39. Singla H, Munshi A, Banipal RPS, et al. Recent updates on the therapeutic potential of HER2 tyrosine kinase inhibitors for the treatment of breast cancer. Curr Cancer Drug Targets. 2018;18(4):306–327. doi:10.2174/1568009617666170623122213
40. Agostinetto E, Curigliano G, Piccart M. Emerging treatments in HER2-positive advanced breast cancer: keep raising the bar. Cell Rep Med. 2024;5(6):101575. doi:10.1016/j.xcrm.2024.101575
41. Pan L, Li J, Xu Q, et al. HER2/PI3K/AKT pathway in HER2-positive breast cancer: a review. Medicine. 2024;103(24):e38508. doi:10.1097/MD.0000000000038508
42. Di Cosimo S, Porcu L, Cardoso F. CDK 4/6 inhibitors mired in uncertainty in HR positive and HER2 negative early breast cancer. Breast. 2021;55:75–78. doi:10.1016/j.breast.2020.12.006
43. Bruni S, Mauro FL, Proietti CJ, et al. Blocking soluble TNFalpha sensitizes HER2-positive breast cancer to trastuzumab through MUC4 downregulation and subverts immunosuppression. J Immunother Cancer. 2023;11(3):e005325. doi:10.1136/jitc-2022-005325
44. Bai W, Peng H, Zhang J, et al. LINC00589-dominated ceRNA networks regulate multiple chemoresistance and cancer stem cell-like properties in HER2(+) breast cancer. NPJ Breast Cancer. 2022;8(1):115. doi:10.1038/s41523-022-00484-0
45. Liu S, Lee JS, Jie C, et al. HER2 overexpression triggers an IL1alpha proinflammatory circuit to drive tumorigenesis and promote chemotherapy resistance. Cancer Res. 2018;78(8):2040–2051. doi:10.1158/0008-5472.CAN-17-2761
46. Peippo M, Gardberg M, Kronqvist P, et al. Characterization of expression and function of the formins FHOD1, INF2, and DAAM1 in HER2-positive breast cancer. J Breast Cancer. 2023;26(6):525–543. doi:10.4048/jbc.2023.26.e47
47. Feng L, Huang S, An G, et al. Identification of new cancer stem cell markers and signaling pathways in HER‑2‑positive breast cancer by transcriptome sequencing. Int J Oncol. 2019;55(5):1003–1018. doi:10.3892/ijo.2019.4876
48. Ahn SG, Kim SJ, Kim C, et al. Molecular classification of triple-negative breast cancer. J Breast Cancer. 2016;19(3):223–230. doi:10.4048/jbc.2016.19.3.223
49. Matossian MD, Burks HE, Bowles AC, et al. A novel patient-derived xenograft model for claudin-low triple-negative breast cancer. Breast Cancer Res Treat. 2018;169(2):381–390. doi:10.1007/s10549-018-4685-2
50. Wang X, Wang L, Liu Y. Current status of immune checkpoint inhibitors and treatment responsive biomarkers for triple-negative breast cancer. Thorac Cancer. 2025;16(9):e70072. doi:10.1111/1759-7714.70072
51. Li YL, Weng H-C, Hsu J-L, et al. The combination of MK-2206 and WZB117 exerts a synergistic cytotoxic effect against breast cancer cells. Front Pharmacol. 2019;10:1311. doi:10.3389/fphar.2019.01311
52. Kang J, Zhong Y, Tian W, et al. A novel anthraquinone‑quinazoline hybrid 7B blocks breast cancer metastasis and EMT via targeting EGFR and Rac1. Int J Oncol. 2021;58(5). doi:10.3892/ijo.2021.5199
53. Li P, Niu Y, Li S, et al. Identification of an AXL kinase inhibitor in triple-negative breast cancer by structure-based virtual screening and bioactivity test. Chem Biol Drug Des. 2022;99(2):222–232. doi:10.1111/cbdd.13977
54. Pardo I, Fagundes PB, Oliveira RSD, et al. A molecular approach to triple-negative breast cancer: targeting the notch signaling pathway. Einstein. 2024;22:eRW0552. doi:10.31744/einstein_journal/2024RW0552
55. Peddi PF. Triple negative breast cancer: any closer to cracking the code? Curr Opin Obstet Gynecol. 2022;34(1):52–55. doi:10.1097/GCO.0000000000000769
56. Young JA, Tan AR. Targeted treatment of triple-negative breast cancer. Cancer J. 2021;27(1):50–58. doi:10.1097/PPO.0000000000000495
57. Kagihara JA, Shagisultanova E, Afghahi A, et al. Moving towards targeted therapies for triple-negative breast cancer. Curr Breast Cancer Rep. 2021;13(3):216–226. doi:10.1007/s12609-021-00416-0
58. Elmasry H, Khadrawy SM, Kamel MM, et al. Evaluation of MMP-13 and micro RNA-138 as prognostic biomarkers for breast cancer in Egyptian women patients. Pathol Res Pract. 2024;253:155045. doi:10.1016/j.prp.2023.155045
59. Dadhich R, Kapoor S. Lipidomic and membrane mechanical signatures in triple-negative breast cancer: scope for membrane-based theranostics. Mol Cell Biochem. 2022;477(11):2507–2528. doi:10.1007/s11010-022-04459-4
60. Huang Y, Zeng A, Song L. Facts and prospects of peptide in targeted therapy and immune regulation against triple-negative breast cancer. Front Immunol. 2023;14:1255820. doi:10.3389/fimmu.2023.1255820
61. Nair A, Singh R, Gautam N, et al. Multifaceted role of phytoconstituents based nano drug delivery systems in combating TNBC: a paradigm shift from chemical to natural. Naunyn Schmiedebergs Arch Pharmacol. 2024;397(12):9207–9226. doi:10.1007/s00210-024-03234-0
62. Duan L, Calhoun S, Shim D, et al. Fatty acid oxidation and autophagy promote endoxifen resistance and counter the effect of AKT inhibition in ER-positive breast cancer cells. J Mol Cell Biol. 2021;13(6):433–444. doi:10.1093/jmcb/mjab018
63. Niu X, Ma J, Li J, et al. Sodium/glucose cotransporter 1-dependent metabolic alterations induce tamoxifen resistance in breast cancer by promoting macrophage M2 polarization. Cell Death Dis. 2021;12(6):509. doi:10.1038/s41419-021-03781-x
64. Gaillard SL, Andreano KJ, Gay LM, et al. Constitutively active ESR1 mutations in gynecologic malignancies and clinical response to estrogen-receptor directed therapies. Gynecol Oncol. 2019;154(1):199–206. doi:10.1016/j.ygyno.2019.04.010
65. Kinslow CJ, Tang A, Chaudhary KR, et al. Prevalence of Estrogen Receptor Alpha (ESR1) somatic mutations in breast cancer. JNCI Cancer Spectr. 2022;6(5). doi:10.1093/jncics/pkac060
66. Zhang J, Wang Q, Wang Q, et al. Mechanisms of resistance to estrogen receptor modulators in ER+/HER2- advanced breast cancer. Cell Mol Life Sci. 2020;77(4):559–572. doi:10.1007/s00018-019-03281-4
67. Sundaresan TK, Dubash TD, Zheng Z, et al. Evaluation of endocrine resistance using ESR1 genotyping of circulating tumor cells and plasma DNA. Breast Cancer Res Treat. 2021;188(1):43–52. doi:10.1007/s10549-021-06270-z
68. Kim YC, Kim CY, Oh JH, et al. NR4A1 regulates tamoxifen resistance by suppressing ERK signaling in ER-positive breast cancer. Cells. 2021;10(7):1633. doi:10.3390/cells10071633
69. Fan P, Jordan VC. Estrogen receptor and the unfolded protein response: double-edged swords in therapy for estrogen receptor-positive breast cancer. Target Oncol. 2022;17(2):111–124. doi:10.1007/s11523-022-00870-5
70. Parisian AD, Barratt SA, Hodges-Gallagher L, et al. Palazestrant (OP-1250), A complete estrogen receptor antagonist, inhibits wild-type and mutant ER-positive breast cancer models as monotherapy and in combination. Mol Cancer Ther. 2024;23(3):285–300. doi:10.1158/1535-7163.MCT-23-0351
71. Gough SM, Flanagan JJ, Teh J, et al. Oral estrogen receptor PROTAC vepdegestrant (ARV-471) is highly efficacious as monotherapy and in combination with CDK4/6 or PI3K/mTOR pathway inhibitors in preclinical ER+ breast cancer models. Clin Cancer Res. 2024;30(16):3549–3563. doi:10.1158/1078-0432.CCR-23-3465
72. Hu B, Hu J. Complete elimination of estrogen receptor α by PROTAC estrogen receptor α degrader ERD-148 in breast cancer cells. Breast Cancer Res Treat. 2024;203(2):383–396. doi:10.1007/s10549-023-07136-2
73. Xiang Y, Liu H, Hu H, et al. LINC00094/miR-19a-3p/CYP19A1 axis affects the sensitivity of ER positive breast cancer cells to Letrozole through EMT pathway. Aging. 2022;14(11):4755–4768. doi:10.18632/aging.204110
74. Li F, Liu S. Focusing on NK cells and ADCC: a promising immunotherapy approach in targeted therapy for HER2-positive breast cancer. Front Immunol. 2022;13:1083462. doi:10.3389/fimmu.2022.1083462
75. Truffi M, Colombo M, Sorrentino L, et al. Multivalent exposure of trastuzumab on iron oxide nanoparticles improves antitumor potential and reduces resistance in HER2-positive breast cancer cells. Sci Rep. 2018;8(1):6563. doi:10.1038/s41598-018-24968-x
76. Veeraraghavan J, De Angelis C, Gutierrez C, et al. HER2-positive breast cancer treatment and resistance. Adv Exp Med Biol. 2025;1464:495–525.
77. Avelino ARM, Pulipati S, Jamouss K, et al. Updates in treatment of HER2-positive metastatic breast cancer. Curr Treat Options Oncol. 2024;25(12):1471–1481. doi:10.1007/s11864-024-01277-2
78. Watanabe S, Yonesaka K, Tanizaki J, et al. Targeting of the HER2/HER3 signaling axis overcomes ligand-mediated resistance to trastuzumab in HER2-positive breast cancer. Cancer Med. 2019;8(3):1258–1268. doi:10.1002/cam4.1995
79. Kuchukulla RR, Hwang I, Park SW, et al. Novel 2,6,9-trisubstituted purines as potent CDK inhibitors alleviating trastuzumab-resistance of HER2-positive breast cancers. Pharmaceuticals. 2022;15(9):1041. doi:10.3390/ph15091041
80. Exman P, Tolaney SM. HER2-positive metastatic breast cancer: a comprehensive review. Clin Adv Hematol Oncol. 2021;19(1):40–50.
81. Qian Y, Peng Y, Zhou H, et al. Trastuzumab plus pertuzumab in combination with chemotherapy in metastatic HER2-positive breast cancer: a retrospective single-armed cohort study in China. Ann Transl Med. 2022;10(16):877. doi:10.21037/atm-22-3592
82. Kamen L, Myneni S, Langsdorf C, et al. A novel method for determining antibody-dependent cellular phagocytosis. J Immunol Methods. 2019;468:55–60. doi:10.1016/j.jim.2019.03.001
83. Shi Y, Sun Y, Seki A, et al. A real-time antibody-dependent cellular phagocytosis assay by live cell imaging. J Immunol Methods. 2024;531:113715. doi:10.1016/j.jim.2024.113715
84. Stanganello E, Brkic M, Zenner S, et al. Protocol for the murine antibody-dependent cellular phagocytosis assay. Methods Cell Biol. 2023;173:109–120.
85. Upton R, Banuelos A, Feng D, et al. Combining CD47 blockade with trastuzumab eliminates HER2-positive breast cancer cells and overcomes trastuzumab tolerance. Proc Natl Acad Sci U S A. 2021;118(29). doi:10.1073/pnas.2026849118
86. Sakamoto M, Murata Y, Tanaka D, et al. Anticancer efficacy of monotherapy with antibodies to SIRPalpha/SIRPbeta1 mediated by induction of antitumorigenic macrophages. Proc Natl Acad Sci U S A. 2022;119(1). doi:10.1073/pnas.2109923118
87. Tsao LC, Crosby EJ, Trotter TN, et al. CD47 blockade augmentation of trastuzumab antitumor efficacy dependent on antibody-dependent cellular phagocytosis. JCI Insight. 2019;4(24). doi:10.1172/jci.insight.131882
88. Ferraro E, Drago JZ, Modi S. Implementing antibody-drug conjugates (ADCs) in HER2-positive breast cancer: state of the art and future directions. Breast Cancer Res. 2021;23(1):84. doi:10.1186/s13058-021-01459-y
89. Barok M, Le Joncour V, Martins A, et al. ARX788, a novel anti-HER2 antibody-drug conjugate, shows anti-tumor effects in preclinical models of trastuzumab emtansine-resistant HER2-positive breast cancer and gastric cancer. Cancer Lett. 2020;473:156–163. doi:10.1016/j.canlet.2019.12.037
90. Sauveur J, Conilh L, Beaumel S, et al. Characterization of T-DM1-resistant breast cancer cells. Pharmacol Res Perspect. 2020;8(4):e00617. doi:10.1002/prp2.617
91. Skeie M, Nikolaysen F, Chitano Y, et al. Hsp90 inhibition and co-incubation with pertuzumab induce internalization and degradation of trastuzumab: implications for use of T-DM1. J Cell Mol Med. 2020;24(17):10258–10262. doi:10.1111/jcmm.15643
92. Takegawa N, Nonagase Y, Yonesaka K, et al. DS-8201a, a new HER2-targeting antibody-drug conjugate incorporating a novel DNA topoisomerase I inhibitor, overcomes HER2-positive gastric cancer T-DM1 resistance. Int J Cancer. 2017;141(8):1682–1689. doi:10.1002/ijc.30870
93. Skidmore L, Sakamuri S, Knudsen NA, et al. ARX788, a site-specific Anti-HER2 antibody-drug conjugate, demonstrates potent and selective activity in HER2-low and T-DM1-resistant breast and gastric cancers. Mol Cancer Ther. 2020;19(9):1833–1843. doi:10.1158/1535-7163.MCT-19-1004
94. Yin O, Iwata H, Lin -C-C, et al. Exposure-response relationships in patients with HER2-positive metastatic breast cancer and other solid tumors treated with trastuzumab deruxtecan. Clin Pharmacol Ther. 2021;110(4):986–996. doi:10.1002/cpt.2291
95. Wang K, Xu T, Wu J, et al. Real-world application of disitamab vedotin (RC48-ADC) in patients with breast cancer with different HER2 expression levels: efficacy and safety analysis. Oncologist. 2025;30(8). doi:10.1093/oncolo/oyae304
96. Mercogliano MF, Bruni S, Mauro FL, Schillaci R. Emerging targeted therapies for HER2-positive breast cancer. Cancers. 2023;15(7).
97. Giugliano F, Carnevale Schianca A, Corti C, et al. Unlocking the resistance to Anti-HER2 treatments in breast cancer: the issue of HER2 spatial distribution. Cancers. 2023;15(5):1385. doi:10.3390/cancers15051385
98. Kim JJ, Kim JY, Suh HB, et al. Characterization of breast cancer subtypes based on quantitative assessment of intratumoral heterogeneity using dynamic contrast-enhanced and diffusion-weighted magnetic resonance imaging. Eur Radiol. 2022;32(2):822–833. doi:10.1007/s00330-021-08166-4
99. Lei H, Ling Y, Yuan P, et al. Assessment of the expression pattern of HER2 and its correlation with HER2-targeting antibody-drug conjugate therapy in urothelial cancer. J Natl Cancer Cent. 2023;3(2):121–128. doi:10.1016/j.jncc.2023.02.003
100. Menezes B, Linderman JJ, Thurber GM. Simulating the selection of resistant cells with bystander killing and antibody coadministration in heterogeneous human epidermal growth factor receptor 2-positive tumors. Drug Metab Dispos. 2022;50(1):8–16. doi:10.1124/dmd.121.000503
101. Ocana A, Amir E, Pandiella A. HER2 heterogeneity and resistance to anti-HER2 antibody-drug conjugates. Breast Cancer Res. 2020;22(1):15. doi:10.1186/s13058-020-1252-7
102. Yamaguchi A, Yamazaki CM, Anami Y, et al. 177Lu-labeled antibody-drug conjugate: a dual-mechanistic treatment modality in solid tumors. Mol Cancer Ther. 2025;24(6):907–919. doi:10.1158/1535-7163.MCT-24-0254
103. Hackshaw MD, Danysh HE, Singh J, et al. Incidence of pneumonitis/interstitial lung disease induced by HER2-targeting therapy for HER2-positive metastatic breast cancer. Breast Cancer Res Treat. 2020;183(1):23–39. doi:10.1007/s10549-020-05754-8
104. Lin W, Xu J, Liao Y, et al. Assessing safety concerns of interstitial lung disease associated with antibody-drug conjugates: a real-world pharmacovigilance evaluation of the FDA adverse event reporting system. Int J Clin Pharm. 2024;46(3):614–622. doi:10.1007/s11096-023-01673-y
105. Ma Z, Zhang Y, Zhu M, et al. Interstitial lung disease associated with anti-HER2 anti-body drug conjugates: results from clinical trials and the WHO’s pharmacovigilance database. Expert Rev Clin Pharmacol. 2022;15(11):1351–1361. doi:10.1080/17512433.2022.2121705
106. Maurier L, Chéné A-L, Hulo P, et al. Diffuse interstitial lung disease induced by antibody-drug conjugates. Rev Mal Respir. 2025;42(5):274–285. doi:10.1016/j.rmr.2025.03.003
107. Riudavets M, Azarine A, Smaali S, et al. Unexpected cardiotoxicity in patients with HER2-mutant NSCLC treated with trastuzumab deruxtecan: a case report. JTO Clin Res Rep. 2022;3(12):100432. doi:10.1016/j.jtocrr.2022.100432
108. Xu C, Chen Z, Xia Y, et al. Clinical best practices in interdisciplinary management of human epidermal growth factor receptor 2 antibody-drug conjugates-induced interstitial lung disease/pneumonitis: an expert consensus in China. Cancer. 2024;130(S17):3054–3066. doi:10.1002/cncr.35475
109. Al Khafaji AT, Barakat AM, Shayyal AJ, et al. Managing doxorubicin cardiotoxicity: insights into molecular mechanisms and protective strategies. J Biochem Mol Toxicol. 2025;39(2):e70155. doi:10.1002/jbt.70155
110. Bandari SK, Tripathi K, Rangarajan S, et al. Therapy-induced chemoexosomes: sinister small extracellular vesicles that support tumor survival and progression. Cancer Lett. 2020;493:113–119. doi:10.1016/j.canlet.2020.08.022
111. Reginelli A, Sangiovanni A, Vacca G, et al. Chemotherapy-induced bowel ischemia: diagnostic imaging overview. Abdom Radiol. 2022;47(5):1556–1564. doi:10.1007/s00261-021-03024-9
112. Zeien J, Qiu W, Triay M, et al. Clinical implications of chemotherapeutic agent organ toxicity on perioperative care. Biomed Pharmacother. 2022;146:112503. doi:10.1016/j.biopha.2021.112503
113. Oshiro-Junior JA, Sato MR, Boni FI, et al. Phthalocyanine-loaded nanostructured lipid carriers functionalized with folic acid for photodynamic therapy. Mater Sci Eng C Mater Biol Appl. 2020;108:110462. doi:10.1016/j.msec.2019.110462
114. Zhang J, Pan S, Jian C, et al. Immunostimulatory properties of chemotherapy in breast cancer: from immunogenic modulation mechanisms to clinical practice. Front Immunol. 2021;12:819405. doi:10.3389/fimmu.2021.819405
115. Harbeck N. Neoadjuvant and adjuvant treatment of patients with HER2-positive early breast cancer. Breast. 2022;62(Suppl 1):S12–s16. doi:10.1016/j.breast.2022.01.006
116. Waks AG, Winer EP. Breast cancer treatment: a review. JAMA. 2019;321(3):288–300. doi:10.1001/jama.2018.19323
117. An SJ, Duchesneau ED, Strassle PD, et al. Pathologic complete response and survival after neoadjuvant chemotherapy in cT1-T2/N0 HER2+ breast cancer. NPJ Breast Cancer. 2022;8(1):65. doi:10.1038/s41523-022-00433-x
118. Wang Y, Xu H, Han Y, et al. Identifying the optimal therapeutics for patients with hormone receptor-positive, HER2-positive advanced breast cancer: a systematic review and network meta-analysis. ESMO Open. 2023;8(3):101216. doi:10.1016/j.esmoop.2023.101216
119. Lan Q, Peyvandi S, Duffey N, et al. Type I interferon/IRF7 axis instigates chemotherapy-induced immunological dormancy in breast cancer. Oncogene. 2019;38(15):2814–2829. doi:10.1038/s41388-018-0624-2
120. Lan KW, Huang W-Y, Chiu Y-L, et al. In vivo investigation of boron-rich nanodrugs for treating triple-negative breast cancers via boron neutron capture therapy. Biomater Adv. 2023;155:213699. doi:10.1016/j.bioadv.2023.213699
121. Yang T, Dai L, Liu J, et al. Metal-phenolic-network-coated gold nanoclusters for enhanced photothermal/chemodynamic/immunogenic cancer therapy. Bioact Mater. 2025;44:447–460. doi:10.1016/j.bioactmat.2024.10.021
122. Azari M, Bahreini F, Uversky VN, et al. Current therapeutic approaches and promising perspectives of using bioengineered peptides in fighting chemoresistance in triple-negative breast cancer. Biochem Pharmacol. 2023;210:115459. doi:10.1016/j.bcp.2023.115459
123. Mou J, Li C, Zheng Q, et al. Research progress in tumor angiogenesis and drug resistance in breast cancer. Cancer Biol Med. 2024;21(7):571–585. doi:10.20892/j.issn.2095-3941.2023.0515
124. Koelsch N, Manjili MH. Moving beyond cytotoxicity in cancer immunotherapy: embracing tumor microenvironment remodeling for durable control. Br J Cancer. 2025;133(9):1233–1240. doi:10.1038/s41416-025-03133-y
125. Zhang M, Liu C, Tu J, et al. Advances in cancer immunotherapy: historical perspectives, current developments, and future directions. Mol Cancer. 2025;24(1):136. doi:10.1186/s12943-025-02305-x
126. Beck C, Rodriguez-Vargas JM, Boehler C, et al. PARP3, a new therapeutic target to alter Rictor/mTORC2 signaling and tumor progression in BRCA1-associated cancers. Cell Death Differ. 2019;26(9):1615–1630. doi:10.1038/s41418-018-0233-1
127. Das C, Dash SR, Sinha S, et al. Talazoparib enhances the quinacrine-mediated apoptosis in patient-derived oral mucosa CSCs by inhibiting BER pathway through the modulation of GCN5 and P300. Med Oncol. 2023;40(12):351. doi:10.1007/s12032-023-02222-3
128. Wang T, Fahrmann JF, Lee H, et al. JAK/STAT3-regulated fatty acid beta-oxidation is critical for breast cancer stem cell self-renewal and chemoresistance. Cell Metab. 2018;27(1):136–150.e5. doi:10.1016/j.cmet.2017.11.001
129. Oshi M, Gandhi S, Wu R, et al. Development of a novel BRCAness score that predicts response to PARP inhibitors. Biomark Res. 2022;10(1):80. doi:10.1186/s40364-022-00427-8
130. Turk AA, Wisinski KB. PARP inhibitors in breast cancer: bringing synthetic lethality to the bedside. Cancer. 2018;124(12):2498–2506. doi:10.1002/cncr.31307
131. Tutuncuoglu B, Krogan NJ. Mapping genetic interactions in cancer: a road to rational combination therapies. Genome Med. 2019;11(1):62. doi:10.1186/s13073-019-0680-4
132. Yuan P, Ma N, Xu B. Poly (adenosine diphosphate-ribose) polymerase inhibitors in the treatment of triple-negative breast cancer with homologous repair deficiency. Med Res Rev. 2024;44(6):2774–2792. doi:10.1002/med.22058
133. Jerez Y, Márquez-Rodas I, Aparicio I, et al. Poly (ADP-ribose) polymerase inhibition in patients with breast cancer and BRCA 1 and 2 mutations. Drugs. 2020;80(2):131–146. doi:10.1007/s40265-019-01235-5
134. Tung NM, Garber JE. BRCA1/2 testing: therapeutic implications for breast cancer management. Br J Cancer. 2018;119(2):141–152. doi:10.1038/s41416-018-0127-5
135. Wang X, Shi Y, Huang D, et al. Emerging therapeutic modalities of PARP inhibitors in breast cancer. Cancer Treat Rev. 2018;68:62–68. doi:10.1016/j.ctrv.2018.05.014
136. Zhang D, Baldwin P, Leal AS, et al. A nano-liposome formulation of the PARP inhibitor Talazoparib enhances treatment efficacy and modulates immune cell populations in mammary tumors of BRCA-deficient mice. Theranostics. 2019;9(21):6224–6238. doi:10.7150/thno.36281
137. Murai J, Pommier Y. BRCAness, homologous recombination deficiencies, and synthetic lethality. Cancer Res. 2023;83(8):1173–1174. doi:10.1158/0008-5472.CAN-23-0628
138. Morganti S, Marra A, De Angelis C, et al. PARP inhibitors for breast cancer treatment: a review. JAMA Oncol. 2024;10(5):658–670. doi:10.1001/jamaoncol.2023.7322
139. Rodler E, Sharma P, Barlow WE, et al. Cisplatin with veliparib or placebo in metastatic triple-negative breast cancer and BRCA mutation-associated breast cancer (S1416): a randomised, double-blind, placebo-controlled, phase 2 trial. Lancet Oncol. 2023;24(2):162–174. doi:10.1016/S1470-2045(22)00739-2
140. Dibitetto D, Widmer CA, Rottenberg S. PARPi, BRCA, and gaps: controversies and future research. Trends Cancer. 2024;10(9):857–869. doi:10.1016/j.trecan.2024.06.008
141. Loehr A, Hussain A, Patnaik A, et al. Emergence of BRCA reversion mutations in patients with metastatic castration-resistant prostate cancer after treatment with rucaparib. Eur Urol. 2023;83(3):200–209. doi:10.1016/j.eururo.2022.09.010
142. Liu Y, Burness ML, Martin-Trevino R, et al. RAD51 mediates resistance of cancer stem cells to PARP inhibition in triple-negative breast cancer. Clin Cancer Res. 2017;23(2):514–522. doi:10.1158/1078-0432.CCR-15-1348
143. Dieras V, Han HS, Kaufman B, et al. Veliparib with carboplatin and paclitaxel in BRCA-mutated advanced breast cancer (BROCADE3): a randomised, double-blind, placebo-controlled, phase 3 trial. Lancet Oncol. 2020;21(10):1269–1282. doi:10.1016/S1470-2045(20)30447-2
144. Li M, Gong Y, Pang Y, et al. A novel colorectal cancer screening framework with feature interpretability to identify high-risk populations for colonoscopy. J Gastroenterol Hepatol. 2024;39(9):1827–1836. doi:10.1111/jgh.16600
145. Wang C, Qu L, Li S, et al. Discovery of first-in-class dual PARP and EZH2 inhibitors for triple-negative breast cancer with wild-type BRCA. J Med Chem. 2021;64(17):12630–12650. doi:10.1021/acs.jmedchem.1c00567
146. Yamaguchi H, Du Y, Nakai K, et al. EZH2 contributes to the response to PARP inhibitors through its PARP-mediated poly-ADP ribosylation in breast cancer. Oncogene. 2018;37(2):208–217. doi:10.1038/onc.2017.311
147. Huang YZ, Sang M-Y, Xi P-W, et al. FANCI inhibition induces PARP1 redistribution to enhance the efficacy of PARP inhibitors in breast cancer. Cancer Res. 2024;84(20):3447–3463. doi:10.1158/0008-5472.CAN-23-2738
148. Zhang Y, Yin F, Luo Z, et al. Design and synthesis of PARP/CDK6 dual-target inhibitors modulating of wnt/beta-catenin signaling pathway for the treatment of BRCA wild-type TNBC. J Med Chem. 2025;68(12):13030–13056. doi:10.1021/acs.jmedchem.5c01058
149. Wang W, Xiong Y, Hu X, et al. Codelivery of adavosertib and olaparib by tumor-targeting nanoparticles for augmented efficacy and reduced toxicity. Acta Biomater. 2023;157:428–441. doi:10.1016/j.actbio.2022.12.021
150. Gao Y, Chen M-K, Chu -Y-Y, et al. Nuclear translocation of the receptor tyrosine kinase c-MET reduces the treatment efficacies of olaparib and gemcitabine in pancreatic ductal adenocarcinoma cells. Am J Cancer Res. 2021;11(1):236–250.
151. Yamaoka K, Fujiwara M, Uchida M, et al. Comprehensive analysis of adverse events induced by PARP inhibitors using JADER and time to onset. Life. 2022;12(9):1355. doi:10.3390/life12091355
152. Zhou T, Zhang J. Therapeutic advances and application of PARP inhibitors in breast cancer. Transl Oncol. 2025;57:102410. doi:10.1016/j.tranon.2025.102410
153. Fu X, Li P, Zhou Q, et al. Mechanism of PARP inhibitor resistance and potential overcoming strategies. Genes Dis. 2024;11(1):306–320. doi:10.1016/j.gendis.2023.02.014
154. Maskey RS, Wang F, Lehman E, et al. Sustained mTORC1 activity during palbociclib-induced growth arrest triggers senescence in ER+ breast cancer cells. Cell Cycle. 2021;20(1):65–80. doi:10.1080/15384101.2020.1859195
155. Vigano L, Locatelli A, Ulisse A, et al. Modulation of the Estrogen/erbB2 receptors cross-talk by CDK4/6 inhibition triggers sustained senescence in estrogen receptor- and ErbB2-positive breast cancer. Clin Cancer Res. 2022;28(10):2167–2179. doi:10.1158/1078-0432.CCR-21-3185
156. Zhou FH, Downton T, Freelander A, et al. CDK4/6 inhibitor resistance in estrogen receptor positive breast cancer, a 2023 perspective. Front Cell Dev Biol. 2023;11:1148792. doi:10.3389/fcell.2023.1148792
157. Ji W, Shi Y, Wang X, et al. Combined Androgen receptor blockade overcomes the resistance of breast cancer cells to palbociclib. Int J Biol Sci. 2019;15(3):522–532. doi:10.7150/ijbs.30572
158. Wagner V, Gil J. Senescence as a therapeutically relevant response to CDK4/6 inhibitors. Oncogene. 2020;39(29):5165–5176. doi:10.1038/s41388-020-1354-9
159. Sinclair WD, Cui X. The effects of HER2 on CDK4/6 activity in breast cancer. Clin Breast Cancer. 2022;22(3):e278–e285. doi:10.1016/j.clbc.2021.08.007
160. Cai Z, Wang J, Li Y, et al. Overexpressed cyclin D1 and CDK4 proteins are responsible for the resistance to CDK4/6 inhibitor in breast cancer that can be reversed by PI3K/mTOR inhibitors. Sci China Life Sci. 2023;66(1):94–109. doi:10.1007/s11427-021-2140-8
161. De Angelis C, Fu X, Cataldo ML, et al. Activation of the IFN signaling pathway is associated with resistance to CDK4/6 inhibitors and immune checkpoint activation in ER-positive breast cancer. Clin Cancer Res. 2021;27(17):4870–4882. doi:10.1158/1078-0432.CCR-19-4191
162. Li Z, Razavi P, Li Q, et al. Loss of the FAT1 tumor suppressor promotes resistance to CDK4/6 inhibitors via the hippo pathway. Cancer Cell. 2018;34(6):893–905.e8. doi:10.1016/j.ccell.2018.11.006
163. Palafox M, Monserrat L, Bellet M, et al. High p16 expression and heterozygous RB1 loss are biomarkers for CDK4/6 inhibitor resistance in ER(+) breast cancer. Nat Commun. 2022;13(1):5258. doi:10.1038/s41467-022-32828-6
164. Pascual J, Lim JSJ, Macpherson IR, et al. Triplet therapy with palbociclib, taselisib, and fulvestrant in PIK3CA-mutant breast cancer and doublet palbociclib and taselisib in pathway-mutant solid cancers. Cancer Discov. 2021;11(1):92–107. doi:10.1158/2159-8290.CD-20-0553
165. Chong QY, Kok Z-H, Bui N-L-C, et al. A unique CDK4/6 inhibitor: current and future therapeutic strategies of abemaciclib. Pharmacol Res. 2020;156:104686. doi:10.1016/j.phrs.2020.104686
166. Cunningham NC, Turner NC. Understanding divergent trial results of adjuvant CDK4/6 inhibitors for early stage breast cancer. Cancer Cell. 2021;39(3):307–309. doi:10.1016/j.ccell.2021.02.011
167. Haslam A, Ranganathan S, Prasad V, et al. CDK4/6 inhibitors as adjuvant therapy in early breast cancer? Uncertain benefits, guaranteed harms. Eur J Cancer. 2024;207:114192. doi:10.1016/j.ejca.2024.114192
168. Loibl S, Furlanetto J. Integrating CDK4/6 inhibitors in the treatment of patients with early breast cancer. Breast. 2022;62(Suppl 1):S70–s79. doi:10.1016/j.breast.2021.12.008
169. Gil-Gil M, Alba E, Gavilá J, et al. The role of CDK4/6 inhibitors in early breast cancer. Breast. 2021;58:160–169. doi:10.1016/j.breast.2021.05.008
170. Klocker EV, Egle D, Bartsch R, et al. Efficacy and safety of CDK4/6 inhibitors: a focus on HR+/HER2- early breast cancer. Drugs. 2025;85(2):149–169. doi:10.1007/s40265-024-02144-y
171. Thill M, Schmidt M. Management of adverse events during cyclin-dependent kinase 4/6 (CDK4/6) inhibitor-based treatment in breast cancer. Ther Adv Med Oncol. 2018;10:1758835918793326. doi:10.1177/1758835918793326
172. Shi C, Ju H, Wu Y, et al. Potential role of cyclin-dependent kinase 4/6 inhibitors in the treatment of mucosal melanoma. Holistic Integrative Oncology. 2024;3(1):24. doi:10.1007/s44178-024-00090-z
173. Tabatabaeian H, Rao A, Ramos A, et al. The emerging roles of WBP2 oncogene in human cancers. Oncogene. 2020;39(24):4621–4635. doi:10.1038/s41388-020-1318-0
174. Lim SK, Lu SY, Kang S-A, et al. Wnt signaling promotes breast cancer by blocking ITCH-mediated degradation of YAP/TAZ transcriptional coactivator WBP2. Cancer Res. 2016;76(21):6278–6289. doi:10.1158/0008-5472.CAN-15-3537
175. Chen S, Zhang Y, Wang H, et al. WW domain-binding protein 2 acts as an oncogene by modulating the activity of the glycolytic enzyme ENO1 in glioma. Cell Death Dis. 2018;9(3):347. doi:10.1038/s41419-018-0376-5
176. Kang SA, Guan JS, Tan HJ, et al. Elevated WBP2 expression in HER2-positive breast cancers correlates with sensitivity to trastuzumab-based neoadjuvant therapy: a retrospective and multicentric study. Clin Cancer Res. 2019;25(8):2588–2600. doi:10.1158/1078-0432.CCR-18-3228
177. Lim YX, Lin H, Seah SH, et al. Reciprocal regulation of hippo and WBP2 signalling-implications in cancer therapy. Cells. 2021;10(11):3130. doi:10.3390/cells10113130
178. Lim SK, Tabatabaeian H, Lu SY, et al. Hippo/MST blocks breast cancer by downregulating WBP2 oncogene expression via miRNA processor dicer. Cell Death Dis. 2020;11(8):669. doi:10.1038/s41419-020-02901-3
179. Chen S, Wang H, Li Z, et al. Interaction of WBP2 with ERalpha increases doxorubicin resistance of breast cancer cells by modulating MDR1 transcription. Br J Cancer. 2018;119(2):182–192. doi:10.1038/s41416-018-0119-5
180. Zhu M, Zhong W, Wong S, et al. E3 ubiquitin ligase ITCH-mediated proteasomal degradation of WBP2 sensitizes breast cancer cells to chemotherapy through restraining AMOTL2/c-JUN axis. Biochem Pharmacol. 2025;232:116720. doi:10.1016/j.bcp.2024.116720
181. Aslan M, Hsu E-C, Garcia-Marques FJ, et al. Oncogene-mediated metabolic gene signature predicts breast cancer outcome. NPJ Breast Cancer. 2021;7(1):141. doi:10.1038/s41523-021-00341-6
182. Hou J, Lv A, Deng Q, et al. TROP2 promotes the proliferation and metastasis of glioblastoma cells by activating the JAK2/STAT3 signaling pathway. Oncol Rep. 2019;41(2):753–764. doi:10.3892/or.2018.6859
183. Li N, Xu J, Yan X, et al. TROP2 promotes the proliferation of triple-negative breast cancer cells via calcium ion-dependent ER stress signaling pathway. Cell Biochem Biophys. 2024;82(3):2205–2216. doi:10.1007/s12013-024-01327-4
184. Zhao W, Kuai X, Zhou X, et al. Trop2 is a potential biomarker for the promotion of EMT in human breast cancer. Oncol Rep. 2018;40(2):759–766. doi:10.3892/or.2018.6496
185. Hu Y, Zhu Y, Qi D, et al. Trop2-targeted therapy in breast cancer. Biomark Res. 2024;12(1):82. doi:10.1186/s40364-024-00633-6
186. Pavšič M. Trop2 forms a stable dimer with significant structural differences within the membrane-distal region as compared to EpCAM. Int J Mol Sci. 2021;22(19):10640. doi:10.3390/ijms221910640
187. Jabbarzadeh Kaboli P, Shabani S, Sharma S, et al. Shedding light on triple-negative breast cancer with Trop2-targeted antibody-drug conjugates. Am J Cancer Res. 2022;12(4):1671–1685.
188. Li WF, Chiang M-F, Weng H-C, et al. OBI-992, a novel TROP2-targeted antibody-drug conjugate, demonstrates antitumor activity in multiple cancer models. Mol Cancer Ther. 2025;24(2):163–175. doi:10.1158/1535-7163.MCT-24-0588
189. Liu X, Deng J, Yuan Y, et al. Advances in Trop2-targeted therapy: novel agents and opportunities beyond breast cancer. Pharmacol Ther. 2022;239:108296. doi:10.1016/j.pharmthera.2022.108296
190. Bardia A, Messersmith WA, Kio EA, et al. Sacituzumab govitecan, a Trop-2-directed antibody-drug conjugate, for patients with epithelial cancer: final safety and efficacy results from the phase I/II IMMU-132-01 basket trial. Ann Oncol. 2021;32(6):746–756. doi:10.1016/j.annonc.2021.03.005
191. Lv L, Shi Y, Wu J, et al. Nanosized drug delivery systems for breast cancer stem cell targeting. Int J Nanomed. 2021;16:1487–1508. doi:10.2147/IJN.S282110
192. Pavlopoulou A, Oktay Y, Vougas K, et al. Determinants of resistance to chemotherapy and ionizing radiation in breast cancer stem cells. Cancer Lett. 2016;380(2):485–493. doi:10.1016/j.canlet.2016.07.018
193. Song K, Farzaneh M. Signaling pathways governing breast cancer stem cells behavior. Stem Cell Res Ther. 2021;12(1):245. doi:10.1186/s13287-021-02321-w
194. Luo M, Shang L, Brooks MD, et al. Targeting breast cancer stem cell state equilibrium through modulation of redox signaling. Cell Metab. 2018;28(1):69–86.e6. doi:10.1016/j.cmet.2018.06.006
195. Wang S, Ma L, Wang Z, et al. Lactate dehydrogenase-A (LDH-A) preserves cancer stemness and recruitment of tumor-associated macrophages to promote breast cancer progression. Front Oncol. 2021;11:654452. doi:10.3389/fonc.2021.654452
196. Luo M, Davis A, McDermott S, et al. Abstract 2311: targeting EMT and MET breast cancer stem cell states through simultaneous inhibition of glycolytic and antioxidant pathways. Cancer Res. 2015;75(15_Supplement):2311. doi:10.1158/1538-7445.AM2015-2311
197. Zhang C, Samanta D, Lu H, et al. Hypoxia induces the breast cancer stem cell phenotype by HIF-dependent and ALKBH5-mediated m(6)A-demethylation of NANOG mRNA. Proc Natl Acad Sci U S A. 2016;113(14):E2047–56. doi:10.1073/pnas.1602883113
198. Maffey A, Storini C, Diceglie C, et al. Mesenchymal stem cells from tumor microenvironment favour breast cancer stem cell proliferation, cancerogenic and metastatic potential, via ionotropic purinergic signalling. Sci Rep. 2017;7(1):13162. doi:10.1038/s41598-017-13460-7
199. Zhou L, Wang D, Sheng D, et al. NOTCH4 maintains quiescent mesenchymal-like breast cancer stem cells via transcriptionally activating SLUG and GAS1 in triple-negative breast cancer. Theranostics. 2020;10(5):2405–2421. doi:10.7150/thno.38875
200. Hershey BJ, Vazzana R, Joppi DL, et al. Lipid droplets define a sub-population of breast cancer stem cells. J Clin Med. 2019;9(1):87. doi:10.3390/jcm9010087
201. Kim SH, Hahm E-R, Singh KB, et al. Novel mechanistic targets of forkhead box Q1 transcription factor in human breast cancer cells. Mol, Carcinog. 2020;59(10):1116–1128. doi:10.1002/mc.23241
202. Mitchell AV, Wu J, Meng F, et al. DDR2 coordinates EMT and metabolic reprogramming as a shared effector of FOXQ1 and SNAI1. Cancer Res Commun. 2022;2(11):1388–1403. doi:10.1158/2767-9764.CRC-22-0013
203. Kim SH, Kaschula CH, Priedigkeit N, et al. Forkhead box Q1 is a novel target of breast cancer stem cell inhibition by diallyl trisulfide. J Biol Chem. 2016;291(26):13495–13508. doi:10.1074/jbc.M116.715219
204. Raza S, Siddiqui JA, Srivastava A, et al. Autophagy as a therapeutic target in breast tumors: the cancer stem cell perspective. Autophagy Rep. 2024;3(1). doi:10.1080/27694127.2024.2358648
205. Liu X, Yang L, Zhang D, et al. Deglycosylation of epithelial cell adhesion molecule affects epithelial to mesenchymal transition in breast cancer cells. J Cell Physiol. 2019;234(4):4504–4514. doi:10.1002/jcp.27256
206. Zhang HW, Hu -J-J, Fu R-Q, et al. Flavonoids inhibit cell proliferation and induce apoptosis and autophagy through downregulation of PI3Kgamma mediated PI3K/AKT/mTOR/p70S6K/ULK signaling pathway in human breast cancer cells. Sci Rep. 2018;8(1):11255. doi:10.1038/s41598-018-29308-7
207. Waks AG, Stover DG, Guerriero JL, et al. The immune microenvironment in hormone receptor-positive breast cancer before and after preoperative chemotherapy. Clin Cancer Res. 2019;25(15):4644–4655. doi:10.1158/1078-0432.CCR-19-0173
208. Yuan Y, Lee JS, Yost SE, et al. Phase I/II trial of palbociclib, pembrolizumab and letrozole in patients with hormone receptor-positive metastatic breast cancer. Eur J Cancer. 2021;154:11–20. doi:10.1016/j.ejca.2021.05.035
209. Dieci MV, Miglietta F, Guarneri V. Immune infiltrates in breast cancer: recent updates and clinical implications. Cells. 2021;10(2):223. doi:10.3390/cells10020223
210. Pan JW, Zabidi MMA, Ng P-S, et al. The molecular landscape of Asian breast cancers reveals clinically relevant population-specific differences. Nat Commun. 2020;11(1):6433. doi:10.1038/s41467-020-20173-5
211. Griguolo G, Pascual T, Dieci MV, et al. Interaction of host immunity with HER2-targeted treatment and tumor heterogeneity in HER2-positive breast cancer. J Immunother Cancer. 2019;7(1):90. doi:10.1186/s40425-019-0548-6
212. Goto T. Radiation as an in situ auto-vaccination: current perspectives and challenges. Vaccines. 2019;7(3).
213. Verma S, Young S, Boldt G, et al. Immunotherapy and radiation therapy sequencing in breast cancer: a systematic review. Int J Radiat Oncol Biol Phys. 2024;118(5):1422–1434. doi:10.1016/j.ijrobp.2024.01.001
214. Kleinendorst SC, Oosterwijk E, Bussink J, et al. Combining targeted radionuclide therapy and immune checkpoint inhibition for cancer treatment. Clin Cancer Res. 2022;28(17):3652–3657. doi:10.1158/1078-0432.CCR-21-4332
215. Tian C, Wang X, Zhang S. CTLA-4 and its inhibitors in esophageal cancer: efficacy of therapy and potential mechanisms of adverse events. Am J Cancer Res. 2023;13(7):3140–3156.
216. Galluzzi L, Humeau J, Buqué A, et al. Immunostimulation with chemotherapy in the era of immune checkpoint inhibitors. Nat Rev Clin Oncol. 2020;17(12):725–741. doi:10.1038/s41571-020-0413-z
217. Page DB, Bear H, Prabhakaran S, et al. Two may be better than one: PD-1/PD-L1 blockade combination approaches in metastatic breast cancer. NPJ Breast Cancer. 2019;5(1):34. doi:10.1038/s41523-019-0130-x
218. Benthami H, Zohair B, Rezouki I, et al. Elevated Siglec-7 expression correlates with adverse clinicopathological, immunological, and therapeutic response signatures in breast cancer patients. Front Immunol. 2025;16:1573365. doi:10.3389/fimmu.2025.1573365
219. Li Q, Wang Y, Jia W, et al. Low-dose anti-angiogenic therapy sensitizes breast cancer to PD-1 blockade. Clin Cancer Res. 2020;26(7):1712–1724. doi:10.1158/1078-0432.CCR-19-2179
220. Nishida N, Sakai K, Morita M, et al. Association between genetic and immunological background of hepatocellular carcinoma and expression of programmed cell death-1. Liver Cancer. 2020;9(4):426–439. doi:10.1159/000506352
221. Abraham PE, Johnson DB. Long-term toxicities of immune checkpoint inhibitors. Drugs. 2025;85(12):1535–1549. doi:10.1007/s40265-025-02243-4
222. Yin Q, Wu L, Han L, et al. Immune-related adverse events of immune checkpoint inhibitors: a review. Front Immunol. 2023;14:1167975. doi:10.3389/fimmu.2023.1167975
223. Al Hadidi S, Heslop HE, Brenner MK, et al. Bispecific antibodies and autologous chimeric antigen receptor T cell therapies for treatment of hematological malignancies. Mol Ther. 2024;32(8):2444–2460. doi:10.1016/j.ymthe.2024.05.039
224. Aldoss I, Bargou RC, Nagorsen D, et al. Redirecting T cells to eradicate B-cell acute lymphoblastic leukemia: bispecific T-cell engagers and chimeric antigen receptors. Leukemia. 2017;31(4):777–787. doi:10.1038/leu.2016.391
225. Viardot A, Bargou R. Bispecific antibodies in haematological malignancies. Cancer Treat Rev. 2018;65:87–95. doi:10.1016/j.ctrv.2018.04.002
226. Goebeler ME, Stuhler G, Bargou R. Bispecific and multispecific antibodies in oncology: opportunities and challenges. Nat Rev Clin Oncol. 2024;21(7):539–560. doi:10.1038/s41571-024-00905-y
227. Ran R, Chen X, Yang J, et al. Immunotherapy in breast cancer: current landscape and emerging trends. Exp Hematol Oncol. 2025;14(1):77. doi:10.1186/s40164-025-00667-y
228. Yang H, Karl MN, Wang W, et al. Engineered bispecific antibodies targeting the interleukin-6 and −8 receptors potently inhibit cancer cell migration and tumor metastasis. Mol Ther. 2022;30(11):3430–3449. doi:10.1016/j.ymthe.2022.07.008
229. Mortezaee K. B7-H3 immunoregulatory roles in cancer. Biomed Pharmacother. 2023;163:114890. doi:10.1016/j.biopha.2023.114890
230. Besliu C, Tanase AD, Rotaru I, et al. The evolving landscape in multiple myeloma: from risk stratification to T cell-directed advanced therapies. Cancers. 2025;17(3):525. doi:10.3390/cancers17030525
231. Hoseini SS, Dobrenkov K, Pankov D, et al. Bispecific antibody does not induce T-cell death mediated by chimeric antigen receptor against disialoganglioside GD2. Oncoimmunology. 2017;6(6):e1320625. doi:10.1080/2162402X.2017.1320625
232. Zhou K, Liu Y, Tang C, et al. Pancreatic cancer: pathogenesis and clinical studies. MedComm. 2025;6(4):e70162. doi:10.1002/mco2.70162
233. Hupperetz C, Lah S, Kim H, et al. CAR T cell immunotherapy beyond haematological malignancy. Immune Netw. 2022;22(1):e6. doi:10.4110/in.2022.22.e6
234. Lu Y, Zhao F. Strategies to overcome tumour relapse caused by antigen escape after CAR T therapy. Mol Cancer. 2025;24(1):126. doi:10.1186/s12943-025-02334-6
235. Salas-Benito D, Birocchi F, Park S, et al. Tandem CAR-T cells targeting mesothelin and MUC16 overcome tumor heterogeneity by targeting one antigen at a time. J Immunother Cancer. 2025;13(9):e012822. doi:10.1136/jitc-2025-012822
236. Hegde M, Mukherjee M, Grada Z, et al. Tandem CAR T cells targeting HER2 and IL13Ralpha2 mitigate tumor antigen escape. J Clin Invest. 2016;126(8):3036–3052. doi:10.1172/JCI83416
237. Kourtesakis A, Bailey E, Chow HNH, et al. Utilization of universal-targeting mSA2 CAR-T cells for the treatment of glioblastoma. Oncoimmunology. 2025;14(1):2518631. doi:10.1080/2162402X.2025.2518631
238. Yang Y, McCloskey JE, Yang H, et al. Bispecific CAR T cells against EpCAM and Inducible ICAM-1 overcome antigen heterogeneity and generate superior antitumor responses. Cancer Immunol Res. 2021;9(10):1158–1174. doi:10.1158/2326-6066.CIR-21-0062
239. Roman Alonso M, Grinyó-Escuer A, Duro-Sánchez S, et al. Generation of chimeric antigen receptor T cells targeting p95HER2 in solid tumors. Nat Commun. 2024;15(1):9589. doi:10.1038/s41467-024-53265-7
240. Kolosnjaj-Tabi J, Marangon I, Nicolas-Boluda A, et al. Nanoparticle-based hyperthermia, a local treatment modulating the tumor extracellular matrix. Pharmacol Res. 2017;126:123–137. doi:10.1016/j.phrs.2017.07.010
241. Rahman SM, Campbell JM, Coates RN, et al. Evaluation of intercellular communication between breast cancer cells and adipose-derived stem cells via passive diffusion in a two-layer microfluidic device. Lab Chip. 2020;20(11):2009–2019. doi:10.1039/D0LC00142B
242. Su X, Xu Y, Fox GC, et al. Breast cancer-derived GM-CSF regulates arginase 1 in myeloid cells to promote an immunosuppressive microenvironment. J Clin Invest. 2021;131(20). doi:10.1172/JCI145296
243. Yu T, Di G. Role of tumor microenvironment in triple-negative breast cancer and its prognostic significance. Chin J Cancer Res. 2017;29(3):237–252. doi:10.21147/j.issn.1000-9604.2017.03.10
244. Zhang H, Wang M, Xu Y. Understanding the mechanisms underlying obesity in remodeling the breast tumor immune microenvironment: from the perspective of inflammation. Cancer Biol Med. 2023;20(4):268–286. doi:10.20892/j.issn.2095-3941.2022.0547
245. Chu S, Shi X, Tian Y, et al. pH-responsive polymer nanomaterials for tumor therapy. Front Oncol. 2022;12:855019. doi:10.3389/fonc.2022.855019
246. Feng Y, Li Z, Song L, et al. A ZIF-8-based dual-modal smart responsive nanoplatform for overcoming radiotherapy resistance in advanced tumors. Nanoscale. 2025;17(19):12134–12148. doi:10.1039/D5NR01093D
247. Li D, Ren T, Ge Y, et al. A multi-functional hypoxia/esterase dual stimulus responsive and hyaluronic acid-based nanomicelle for targeting delivery of chloroethylnitrosouea. J Nanobiotechnology. 2023;21(1):291. doi:10.1186/s12951-023-02062-3
248. Liu X, Sun M, Dai L, et al. Functional nucleic acid-powered hybrid nanocarriers for synchronized targeted delivery of dual-solubility therapeutics. J Biotechnol. 2025;406:127–135. doi:10.1016/j.jbiotec.2025.07.002
249. Ryu K, Park J, Kim TI. Effect of pH-responsive charge-conversional polymer coating to cationic reduced graphene oxide nanostructures for tumor microenvironment-targeted drug delivery systems. Nanomaterials. 2019;9(9):1289. doi:10.3390/nano9091289
250. Timaner M, Letko-Khait N, Kotsofruk R, et al. Therapy-educated mesenchymal stem cells enrich for tumor-initiating cells. Cancer Res. 2018;78(5):1253–1265. doi:10.1158/0008-5472.CAN-17-1547
251. Nel J, Elkhoury K, Velot É, et al. Functionalized liposomes for targeted breast cancer drug delivery. Bioact Mater. 2023;24:401–437. doi:10.1016/j.bioactmat.2022.12.027
252. Yu J, Hu J-R, Tian Y, et al. Nanosensitizer-assisted sonodynamic therapy for breast cancer. J Nanobiotechnology. 2025;23(1):281. doi:10.1186/s12951-025-03311-3
253. Eftekhari R, Esmaeili R, Mirzaei R, et al. Study of the tumor microenvironment during breast cancer progression. Cancer Cell Int. 2017;17(1):123. doi:10.1186/s12935-017-0492-9
254. Yang W, Xu M, Xu S, et al. Single-cell RNA reveals a tumorigenic microenvironment in the interface zone of human breast tumors. Breast Cancer Res. 2023;25(1):100. doi:10.1186/s13058-023-01703-7
255. Read RD, Tapp ZM, Rajappa P, et al. Glioblastoma microenvironment-from biology to therapy. Genes Dev. 2024;38(9–10):360–379. doi:10.1101/gad.351427.123
256. Xiao Y, Yu D. Tumor microenvironment as a therapeutic target in cancer. Pharmacol Ther. 2021;221:107753. doi:10.1016/j.pharmthera.2020.107753
257. Semenza GL. Hypoxia-inducible factors: coupling glucose metabolism and redox regulation with induction of the breast cancer stem cell phenotype. EMBO j. 2017;36(3):252–259. doi:10.15252/embj.201695204
258. Singh M, Venkata Krishnan H, Ranganathan S, et al. Controlled three-dimensional tumor microenvironments recapitulate phenotypic features and differential drug response in early vs advanced stage breast cancer. ACS Biomater Sci Eng. 2018;4(2):421–431. doi:10.1021/acsbiomaterials.7b00081
259. Trzeciak ER, Zimmer N, Gehringer I, et al. Oxidative stress differentially influences the survival and metabolism of cells in the melanoma microenvironment. Cells. 2022;11(6):930. doi:10.3390/cells11060930
260. Alven S, Gandidzanwa S, Ngalo B, et al. Platinum group metals nanoparticles in breast cancer therapy. Pharmaceutics. 2024;16(9):1162. doi:10.3390/pharmaceutics16091162
261. He L, Gu J, Lim LY, et al. Nanomedicine-mediated therapies to target breast cancer stem cells. Front Pharmacol. 2016;7:313. doi:10.3389/fphar.2016.00313
262. Mu Q, Wang H, Zhang M. Nanoparticles for imaging and treatment of metastatic breast cancer. Expert Opin Drug Deliv. 2017;14(1):123–136. doi:10.1080/17425247.2016.1208650
263. Mukherjee D, Raikwar S. Recent update on Nanocarrier(s) as the targeted therapy for breast cancer. AAPS Pharm Sci Tech. 2024;25(6):153. doi:10.1208/s12249-024-02867-x
264. Zeng M, Hu C, Chen T, et al. Advancements in cell membrane-derived biomimetic nanotherapeutics for breast cancer. Int J Nanomed. 2025;20:6059–6083. doi:10.2147/IJN.S502144
265. Wang Z, Zhang S, Zhang R, et al. Bioengineered dual-targeting protein nanocage for stereoscopical loading of synergistic hydrophilic/hydrophobic drugs to enhance anticancer efficacy. Adv Funct Mater. 2021;31(29):2102004. doi:10.1002/adfm.202102004
266. Yang C, Cheng X, Shen P. Silencing of BCSG1 with specific siRNA via nanocarriers for breast cancer treatment. Bull Cancer. 2021;108(3):323–332. doi:10.1016/j.bulcan.2020.10.022
267. Alipour M, Sheikhnejad R, Fouani MH, et al. DNAi-peptide nanohybrid smart particles target BCL-2 oncogene and induce apoptosis in breast cancer cells. Biomed Pharmacother. 2023;166:115299. doi:10.1016/j.biopha.2023.115299
268. Yang Y, Yang S, Zhang B, et al. Hybrid liposome-MSN system with co-delivering potential effective against multidrug-resistant tumor targets in mice model. Int J Nanomed. 2024;19:8949–8970. doi:10.2147/IJN.S472276
269. Cheng Z, Li H, Chen C, et al. Application of serum SERS technology based on thermally annealed silver nanoparticle composite substrate in breast cancer. Photodiagnosis Photodyn Ther. 2023;41:103284. doi:10.1016/j.pdpdt.2023.103284
270. Liu B, Du F, Feng Z, et al. Ultrasound-augmented cancer immunotherapy. J Mater Chem B. 2024;12(15):3636–3658. doi:10.1039/D3TB02705H
271. Sabuncu S, Yildirim A. Gas-stabilizing nanoparticles for ultrasound imaging and therapy of cancer. Nano Converg. 2021;8(1):39. doi:10.1186/s40580-021-00287-2
272. Italiya KS, Mullins-Dansereau V, Geoffroy K, et al. Ultrasound and microbubble mediated delivery of virus-sensitizing drugs improves in vitro oncolytic virotherapy against breast cancer cells. Ultrasound Med Biol. 2025;51(7):1124–1133. doi:10.1016/j.ultrasmedbio.2025.03.010
273. Wang L, Cao L, Shao K, et al. Phytochlorin-based sonosensitizers combined with free-field ultrasound for immune-sonodynamic cancer therapy. Adv Mater. 2025;37(8):e2410559. doi:10.1002/adma.202410559
274. Zhang S, Wu Y, Lyu C, et al. Magnetothermal and ultrasound-activated nanoplatform for the inhalable therapy of bacterial lung infections. Acta Biomater. 2025;199:428–442. doi:10.1016/j.actbio.2025.04.041
275. Li Y, Zhou Q, Deng Z, et al. IR-780 dye as a sonosensitizer for sonodynamic therapy of breast tumor. Sci Rep. 2016;6(1):25968. doi:10.1038/srep25968
276. Baghbani F, Moztarzadeh F, Mohandesi JA, et al. Novel alginate-stabilized doxorubicin-loaded nanodroplets for ultrasounic theranosis of breast cancer. Int J Biol Macromol. 2016;93(Pt A):512–519. doi:10.1016/j.ijbiomac.2016.09.008
277. Ho YJ, Yeh CK. Theranostic performance of acoustic nanodroplet vaporization-generated bubbles in tumor intertissue. Theranostics. 2017;7(6):1477–1488. doi:10.7150/thno.19099
278. Ho YJ, Yeh CK. Concurrent anti-vascular therapy and chemotherapy in solid tumors using drug-loaded acoustic nanodroplet vaporization. Acta Biomater. 2017;49:472–485. doi:10.1016/j.actbio.2016.11.018
279. Liu WW, Liu S-W, Liou Y-R, et al. Nanodroplet-vaporization-assisted sonoporation for highly effective delivery of photothermal treatment. Sci Rep. 2016;6(1):24753. doi:10.1038/srep24753
280. Sun L, Yuan Y, Chen J, et al. Brahma related gene 1 (BRG1) regulates breast cancer cell migration and invasion by activating MUC1 transcription. Biochem Biophys Res Commun. 2019;511(3):536–543. doi:10.1016/j.bbrc.2019.02.088
281. Rui L, An Y, Jin T, et al. Detection of MUC1 protein on tumor cells and their derived exosomes for breast cancer surveillance with an electrochemiluminescence aptasensor. J Electroanal Chem. 2021;882:115011. doi:10.1016/j.jelechem.2021.115011
282. Hesari A, Golrokh Moghadam SA, Siasi A, et al. Tumor-derived exosomes: potential biomarker or therapeutic target in breast cancer? J Cell Biochem. 2018;119(6):4236–4240. doi:10.1002/jcb.26364
283. Thakur P, Dahiya H, Kaushal A, et al. Exosomal miRNAs as next-generation therapy vehicles in breast cancer. Curr Gene Ther. 2023;23(5):330–342. doi:10.2174/1566523223666230215103524
284. Zheng Y, Li M, Weng B, et al. Exosome-based delivery nanoplatforms: next-generation theranostic platforms for breast cancer. Biomater Sci. 2022;10(7):1607–1625. doi:10.1039/D2BM00062H
285. Niu Z, Wu J, Zhao Q, et al. CAR-based immunotherapy for breast cancer: peculiarities, ongoing investigations, and future strategies. Front Immunol. 2024;15:1385571. doi:10.3389/fimmu.2024.1385571
286. Beilankouhi EAV, Valilo M, Dastmalchi N, et al. The function of autophagy in the initiation, and development of breast cancer. Curr Med Chem. 2024;31(20):2974–2990. doi:10.2174/0929867330666230503145319
287. Yu T, Rui L, Jiumei Z, et al. Advances in the study of autophagy in breast cancer. Breast Cancer. 2024;31(2):195–204. doi:10.1007/s12282-023-01541-7
288. Liu L, Tian Z, Zhang Y, et al. Ligand-based discovery of small molecules suppressing cancer cell proliferation via autophagic flux inhibition. J Mol Med. 2020;98(11):1573–1589. doi:10.1007/s00109-020-01971-2
289. You M, Lee Y-H, Kim H-J, et al. St. john’s wort suppresses growth in triple-negative breast cancer cell line MDA-MB-231 by inducing prodeath autophagy and apoptosis. Nutrients. 2020;12(10):3175. doi:10.3390/nu12103175
290. Zheng B, Qian F, Wang X, et al. Neddylation activated TRIM25 desensitizes triple-negative breast cancer to paclitaxel via TFEB-mediated autophagy. J Exp Clin Cancer Res. 2024;43(1):177. doi:10.1186/s13046-024-03085-w
291. Li Q, Zhong X, Yao W, et al. Inhibitor of glutamine metabolism V9302 promotes ROS-induced autophagic degradation of B7H3 to enhance antitumor immunity. J Biol Chem. 2022;298(4):101753. doi:10.1016/j.jbc.2022.101753
292. Shaojun P, Wang H, Xin Y, et al. Second near-infrared photoactivatable hydrogen selenide nanogenerators for metastasis-inhibited cancer therapy. Nano Today. 2021;40:101240. doi:10.1016/j.nantod.2021.101240
293. Zhao YX, Wang H, Zhang S-W, et al. Enhancing therapeutic efficacy in luminal androgen receptor triple-negative breast cancer: exploring chidamide and enzalutamide as a promising combination strategy. Cancer Cell Int. 2024;24(1):131. doi:10.1186/s12935-024-03313-5
294. Bonilha CS, Benson RA, Brewer JM, et al. Targeting opposing immunological roles of the junctional adhesion Molecule-A in autoimmunity and cancer. Front Immunol. 2020;11:602094. doi:10.3389/fimmu.2020.602094
295. Zhou Z, Zhu T, Zheng W, et al. LAT1 transporter as a target for breast cancer diagnosis and therapy. Eur J Med Chem. 2025;283:117064. doi:10.1016/j.ejmech.2024.117064
296. Kurozumi S, Kaira K, Matsumoto H, et al. Association of L-type amino acid transporter 1 (LAT1) with the immune system and prognosis in invasive breast cancer. Sci Rep. 2022;12(1):2742. doi:10.1038/s41598-022-06615-8
297. Ichinoe M, Mikami T, Yanagisawa N, et al. Prognostic values of L-type amino acid transporter 1 and CD98hc expression in breast cancer. J Clin Pathol. 2021;74(9):589–595. doi:10.1136/jclinpath-2020-206457
298. Magara K, Takasawa A, Takasawa K, et al. Multilayered proteomics reveals that JAM -A promotes breast cancer progression via regulation of amino acid transporter LAT1. Cancer Sci. 2024;115(9):3153–3168. doi:10.1111/cas.16259
299. Shindo H, Harada-Shoji N, Ebata A, et al. Targeting amino acid metabolic reprogramming via L-type amino acid transporter 1 (LAT1) for endocrine-resistant breast cancer. Cancers. 2021;13(17):4375. doi:10.3390/cancers13174375
300. Fedoroff MY, Zhao L, Wang S, et al. Amino acid transporter LAT1 (SLC7A5) promotes metabolic rewiring in TNBC progression through the L-Trp/QPRT/NAD(+) pathway. J Exp Clin Cancer Res. 2025;44(1):190. doi:10.1186/s13046-025-03446-z
301. Zhao Y, Pu C, Liu K, et al. Targeting LAT1 with JPH203 to reduce TNBC proliferation and reshape suppressive immune microenvironment by blocking essential amino acid uptake. Amino Acids. 2025;57(1):27. doi:10.1007/s00726-025-03456-3
302. Burugu S, Gao D, Leung S, et al. LAG-3+ tumor infiltrating lymphocytes in breast cancer: clinical correlates and association with PD-1/PD-L1+ tumors. Ann Oncol. 2017;28(12):2977–2984. doi:10.1093/annonc/mdx557
303. Ren T, Zheng Y, Liu F, et al. Identification and validation of JAM-A as a novel prognostic and immune factor in human tumors. Biomedicines. 2024;12(7):1423. doi:10.3390/biomedicines12071423
304. Chen J, Pan T, Zhu Z, et al. A deep learning-based multimodal medical imaging model for breast cancer screening. Sci Rep. 2025;15(1):14696. doi:10.1038/s41598-025-99535-2
305. Song X, Xu H, Wang X, et al. Use of ultrasound imaging omics in predicting molecular typing and assessing the risk of postoperative recurrence in breast cancer. BMC Womens Health. 2024;24(1):380. doi:10.1186/s12905-024-03231-8
306. Jonczyk MM, Fisher CS, Babbitt R, et al. Surgical predictive model for breast cancer patients assessing acute postoperative complications: the breast cancer surgery risk calculator. Ann Surg Oncol. 2021;28(9):5121–5131. doi:10.1245/s10434-021-09710-8
307. Qayoom H, Alkhanani M, Almilaibary A, et al. A network pharmacology-based investigation of brugine reveals its multi-target molecular mechanism against breast cancer. Med Oncol. 2023;40(7):202. doi:10.1007/s12032-023-02067-w
308. Poirion OB, Jing Z, Chaudhary K, et al. DeepProg: an ensemble of deep-learning and machine-learning models for prognosis prediction using multi-omics data. Genome Med. 2021;13(1):112. doi:10.1186/s13073-021-00930-x
309. Jain U, Jain B, Fayanju OM, et al. Disparities in timely treatment among young women with breast cancer. Am J Surg. 2022;224(2):811–815. doi:10.1016/j.amjsurg.2022.01.019
310. Uhlmann S, Mannsperger H, Zhang JD, et al. Global microRNA level regulation of EGFR-driven cell-cycle protein network in breast cancer. Mol Syst Biol. 2012;8(1):570. doi:10.1038/msb.2011.100
311. Zhang J, Zhang HD, Chen L, et al. β-elemene reverses chemoresistance of breast cancer via regulating MDR-related microRNA expression. Cell Physiol Biochem. 2014;34(6):2027–2037. doi:10.1159/000366398
312. Chang YY, Kuo W-H, Hung J-H, et al. Deregulated microRNAs in triple-negative breast cancer revealed by deep sequencing. Mol Cancer. 2015;14(1):36. doi:10.1186/s12943-015-0301-9
313. Xu T, Le TD, Liu L, et al. Identifying cancer subtypes from miRNA-TF-mRNA regulatory networks and expression data. PLoS One. 2016;11(4):e0152792. doi:10.1371/journal.pone.0152792
314. Lirussi L, Ayyildiz D, Liu Y, et al. A regulatory network comprising let-7 miRNA and SMUG1 is associated with good prognosis in ER+ breast tumours. Nucleic Acids Res. 2022;50(18):10449–10468. doi:10.1093/nar/gkac807
315. Zhang D, An X, Yu H, et al. The regulatory effect of 6-TG on lncRNA-miRNA-mRNA ceRNA network in triple-negative breast cancer cell line. Biosci Rep. 2021;41(2). doi:10.1042/BSR20203890
316. Sarkar JP, Saha I, Sarkar A, et al. Machine learning integrated ensemble of feature selection methods followed by survival analysis for predicting breast cancer subtype specific miRNA biomarkers. Comput Biol Med. 2021;131:104244. doi:10.1016/j.compbiomed.2021.104244
317. Das N, Bhat S, Fugo P, et al. miRNAs as neuro-oncologic therapeutics: a narrative review. Mol Ther. 2025;33(6):2740–2752. doi:10.1016/j.ymthe.2024.12.045
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Breast Cancer: An Overview of Current Therapeutic Strategies, Challenge, and Perspectives
Wang J, Wu SG
Breast Cancer: Targets and Therapy 2023, 15:721-730
Published Date: 20 October 2023
Dysregulation of Fatty Acid Metabolism in Breast Cancer and Its Targeted Therapy
Chang Y, Du R, Xia F, Xu X, Wang H, Chen X
Breast Cancer: Targets and Therapy 2024, 16:825-844
Published Date: 29 November 2024
From Pioneering Discoveries to Innovative Therapies: A Journey Through the History and Advancements of Nanoparticles in Breast Cancer Treatment
Basingab FS, Alshahrani OA, Alansari IH, Almarghalani NA, Alshelali NH, Alsaiary AH, Alharbi N, Zaher KA
Breast Cancer: Targets and Therapy 2025, 17:27-51
Published Date: 21 January 2025
Advances in Targeted Therapy for Brain Metastases in Human Epidermal Growth Factor Receptor 2 (HER2)-Positive Breast Cancer: A Focus on ADCs and TKIs
Rao B, Huo P, Lu J, Huang W
Breast Cancer: Targets and Therapy 2025, 17:423-432
Published Date: 19 May 2025
Molecular Insights into Phytochemicals Mediated Epigenetic Regulation in Preclinical Models of Breast Cancer
Pandey P, Alkhathami AG, Saeed M, Alshaghdali K, Kumar S, Tallei TE, Bae H, Park MN, Kumar G, Kim B, Khan F
Drug Design, Development and Therapy 2025, 19:10589-10606
Published Date: 27 November 2025