Back to Journals » International Journal of Nanomedicine » Volume 20
Mesenchymal Stem Cell-Derived Exosomes in Anti-NET Therapy: Mechanisms, Challenges, and Future Perspectives
Authors Ye Y, Ye Y, Tian M, Zhao Y, Guo Z, Jin C, Duan S, Zheng Y ![]()
Received 26 March 2025
Accepted for publication 14 October 2025
Published 4 December 2025 Volume 2025:20 Pages 14481—14497
DOI https://doi.org/10.2147/IJN.S528771
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Dr Krishna Nune
Yihao Ye,1,2,* Yushan Ye,3,* Mei Tian,3– 5 Yitao Zhao,3 Ziwei Guo,2,6 Chenghong Jin,3 Shiwei Duan,3 Yueliang Zheng2
1The Second School of Clinical Medicine, Hangzhou Normal University, Hangzhou, Zhejiang, 311121, People’s Republic of China; 2Emergency and Critical Care Center, Department of Emergency Medicine, Zhejiang Provincial People’s Hospital (Affiliated People’s Hospital), Hangzhou Medical College, Hangzhou, Zhejiang, 314408, People’s Republic of China; 3Key Laboratory of Novel Targets and Drug Study for Neural Repair of Zhejiang Province, School of Medicine, Hangzhou City University, Hangzhou, Zhejiang, 310015, People’s Republic of China; 4Geriatric Medicine Center, Department of Endocrinology, Zhejiang Provincial People’s Hospital (Affiliated People’s Hospital), Hangzhou Medical College, Hangzhou, Zhejiang, 314408, People’s Republic of China; 5College of Pharmacy, Zhejiang University of Technology, Hangzhou, Zhejiang, 310014, People’s Republic of China; 6School of Basic Medicine and Forensic Medicine Hangzhou Medical College, Hangzhou, Zhejiang, 310063, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Shiwei Duan, Key Laboratory of Novel Targets and Drug Study for Neural Repair of Zhejiang Province, School of Medicine, Hangzhou City University, Hangzhou, Zhejiang, 310015, People’s Republic of China, Email [email protected] Yueliang Zheng, Emergency and Critical Care Center, Department of Emergency Medicine, Zhejiang Provincial People’s Hospital (Affiliated People’s Hospital), Hangzhou Medical College, Hangzhou, Zhejiang, 314408, People’s Republic of China, Email [email protected]
Abstract: Neutrophils, a key component of the innate immune system, play a crucial role in immune responses. In 2004, Brinkmann et al identified neutrophil extracellular traps (NETs) as a novel antibacterial mechanism. However, NETs have since been implicated in the pathogenesis of various diseases, including autoimmune disorders, sepsis, and cancer. Consequently, targeting NETs has emerged as a promising therapeutic approach. Mesenchymal stem cells (MSCs) have demonstrated efficacy in modulating NET formation, but MSC-derived exosomes offer distinct advantages over whole MSCs due to their lower immunogenicity, higher biological stability, and ability to deliver bioactive molecules like miRNAs and CD59. These exosomes can block critical signaling pathways involved in NET formation and protect neutrophil mitochondria, inhibiting NET release. Despite challenges such as low yield and targeting efficiency, ongoing research has made significant strides in addressing these issues. This article reviews the current progress in MSC-derived exosome-based anti-NET therapies and discusses potential strategies to enhance their therapeutic application.
Keywords: neutrophils, neutrophil extracellular traps, exosomes, MSC, MSC-exo
Introduction
Neutrophils are a crucial component of the innate immune system and serve as a vital defense against microbial pathogens. Upon infection or inflammation, neutrophils are attracted to the site of infection through chemotaxis, where they perform phagocytosis, bind pathogens, and release various antimicrobial substances to eliminate microbes.1 In 2004, Brinkmann, Reichard, Goosmann, Fauler, Uhlemann, Weiss, Weinrauch and Zychlinsky2 made a groundbreaking discovery that neutrophils possess an additional mechanism for killing bacteria outside the cell: neutrophil extracellular traps (NETs). NETs are a fibrous network structure composed of DNA, nuclear proteins, histones, granular proteins, and cytoplasmic proteins. When neutrophils are stimulated by interleukin-8 (IL-8), phorbol myristate (PMA), or lipopolysaccharide (LPS), they release NETs to capture and kill bacteria extracellularly.3
While NETs have extracellular antibacterial and pro-inflammatory effects, excessive neutrophil activation can lead to uncontrollable NET release, resulting in persistent inflammation, vascular occlusion, tissue damage, and an exaggerated immune response.4 Increasing evidence suggests that NETs are closely linked to a range of diseases, including autoimmune disorders, cancer, atherosclerosis, sepsis, and acute lung injury.5 As a result, anti-NET therapy has emerged as a potential therapeutic target.
Traditional anti-NET drugs aim to either inhibit NET formation or degrade existing NETs. For instance, statins,6 cyclosporine A,7 and metformin8 can suppress NET formation by reducing reactive oxygen species (ROS) levels, while DNase I, which is clinically approved, degrades released NETs.9 However, these traditional anti-NET strategies lack specificity and often inhibit NET release by neutrophils throughout the body. This broad inhibition can compromise normal immune function, making the body more susceptible to infections.10 Consequently, these therapies are challenging to apply in clinical settings, highlighting the need for more targeted and effective drugs to regulate NETs in a controlled manner.11
Mesenchymal stem cell exosomes (MSC-Exos) are considered a promising alternative to traditional drugs for targeted therapy.12 MSCs have anti-inflammatory and immunomodulatory properties, and their ability to inhibit NET formation has been well-documented.13 MSCs primarily exert their biological effects via paracrine signaling through exosomes—nano-sized vesicles encapsulated by a lipid bilayer that are rich in proteins, lipids, miRNAs, mRNAs, and other molecules, playing a key role in intercellular communication in both normal and pathological conditions.14 Moreover, MSC-derived exosomes can serve as drug delivery systems, enabling the targeted delivery of therapeutic agents and showing considerable potential for various medical applications.15 Recent studies have demonstrated the therapeutic potential of MSC-Exos in inhibiting NET formation, further supporting their use in anti-NET therapy. This review aims to summarize the progress of MSC-Exos in anti-NET therapy and explore possible future therapeutic strategies and prospects.
Mechanism and Pathophysiology of Neutrophil Extracellular Trap Formation
NETs are extracellular fibrous structures released by activated neutrophils in response to specific stimuli. These structures primarily consist of DNA fibers, histones, and antimicrobial proteins that capture and neutralize pathogens such as bacteria, fungi, protozoa, and viruses outside the cell. The process of NET formation, known as “NETosis”, represents a unique form of cell death distinct from necrosis and apoptosis.4
NETosis typically occurs via the NADPH oxidase (NOX)-dependent pathway, which usually takes 3–4 hours. This programmed cell death process is characterized by the breakdown of the nuclear membrane, chromatin decondensation, and the binding of decondensed chromatin to cytoplasmic proteins.16 Initially, neutrophils can be stimulated by various factors, including pathogens, cytokines, LPS, and PMA, leading to the release of calcium from the endoplasmic reticulum and the opening of calcium channels in the plasma membrane.17 This activation triggers NOX through protein kinase C (PKC) and Raf-MEK (MAPK/ERK kinase)-ERK signaling pathways.18 The increased ROS levels (such as O2, H2O2, and HOCl) promote the degradation of azurophilic granules containing myeloperoxidase (MPO) and neutrophil elastase (NE). MPO and NE are then released into the cytoplasm, where they help activate gasdermin D (GSDMD), which further contributes to the process.16
Moreover, NE interacts with F-actin to degrade actin filaments, translocates to the nucleus, and partially cleaves histones, thereby promoting chromatin decondensation.19 ROS also activate protein arginine deiminase 4 (PAD4), which, in combination with MPO, causes excessive citrullination of histones, leading to the loss of their positive charge and further chromatin condensation.20 The breakdown of the cytoskeleton causes the nuclear membrane to rupture, releasing decondensed chromatin into the cytoplasm. Throughout NETosis, GSDMD pore assembly on the plasma membrane plays a crucial role in cell death and membrane permeability. This process is triggered by PMA, LPS, and bacterial activation of caspase-11.21 Once the cytoskeleton and nuclear membrane disintegrate, chromatin decondenses, binds to proteins, and neutrophils are released following plasma membrane lysis. Some studies also suggest that NETosis is regulated by the activation of the cell cycle kinase CDK4/6, though the specific pathways remain to be fully elucidated22 (Figure 1).
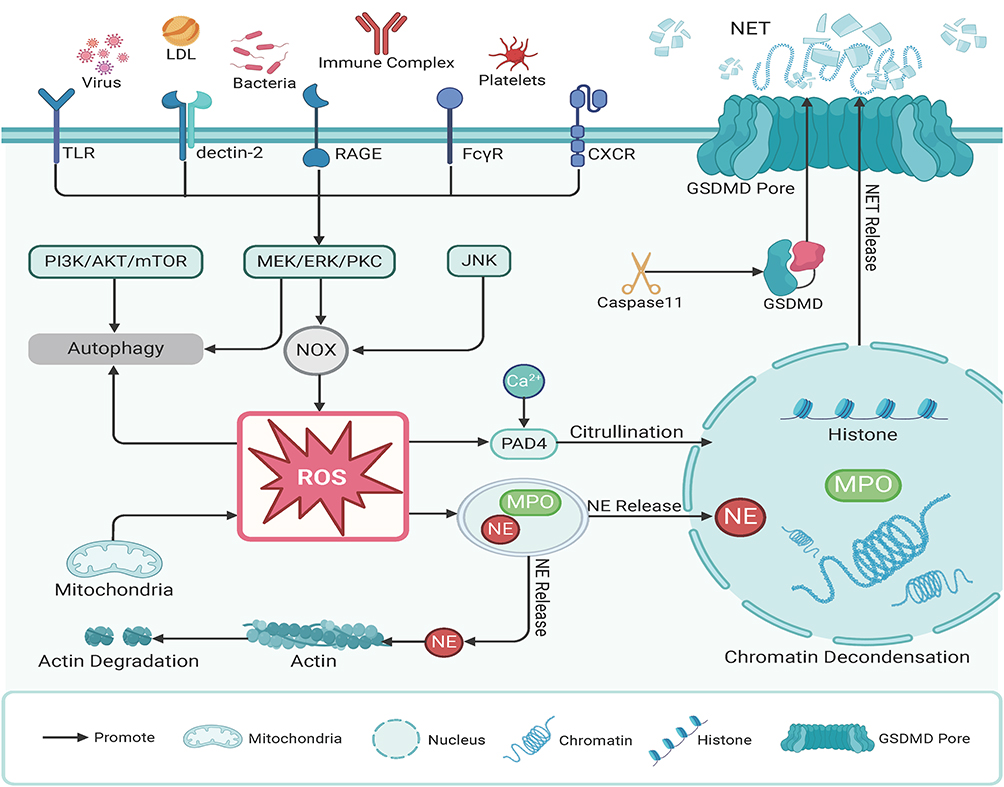 |
Figure 1 Suicidal Neutrophil Extracellular Traps (NETs). Upon activation, neutrophils trigger the NOX complex through the MAPK signaling pathway, leading to the production of reactive oxygen species (ROS). ROS promote the release of myeloperoxidase (MPO) and neutrophil elastase (NE). NE degrades actin and, together with MPO, enters the nucleus to depolymerize chromatin. Concurrently, protein arginine deiminase 4 (PAD4), activated by ROS and calcium ions, translocates to the nucleus to citrullinate histones. Ultimately, with the assistance of gasdermin D (GSDMD), the cytoskeleton and nuclear membrane disintegrate, allowing chromatin to be released into the cytoplasm. Abbreviations: AZU, Azurophilic granules; GSDMD, Gasdermin D; LDL, Low-density lipoprotein; MAPK, Mitogen-activated protein kinase; MPO, Myeloperoxidase; NOX, NADPH oxidase; NE, Neutrophil elastase; PAD4, Protein arginine deiminase 4; ROS, Reactive oxygen species; TLR, Toll-like receptor; mTOR, Mechanistic target of rapamycin; PI3K, Phosphoinositide 3-Kinase; PKC, Protein kinase C. |
NETs are implicated in the development and progression of various diseases, including sepsis, autoimmune diseases, atherosclerosis, cancer, and eclampsia, by promoting inflammation, thrombosis, immune dysregulation, and tissue damage In sepsis, NETs accelerate platelet pyroptosis, release inflammatory cytokines, and transform endothelial cells into a pro-inflammatory, pro-coagulant state, thereby inducing intravascular coagulation and organ damage.23 In acute lung injury, NETs exacerbate inflammation by damaging endothelial and epithelial cells in the lungs. In rheumatoid arthritis (RA), NETs promote the production of RA autoantibodies, citrullination of histones, and secretion of pro-inflammatory cytokines, thereby aggravating disease symptoms.24 In atherosclerosis, NETs induce oxidative stress, modify high-density lipoprotein (HDL) particles, reduce cholesterol excretion, impair endothelial function, promote cell apoptosis, and contribute to thrombosis.25 In cancer, NETs foster tumor progression primarily by amplifying inflammation, and the carcinogenic properties of neutrophils may also play a role.26 NETs capture circulating tumor cells and facilitate their implantation in distant organs, potentially awakening dormant cancer cells by remodeling the extracellular matrix.27 Additionally, NETs help create an immunosuppressive tumor microenvironment, further supporting tumor growth.26 (Figure 2).
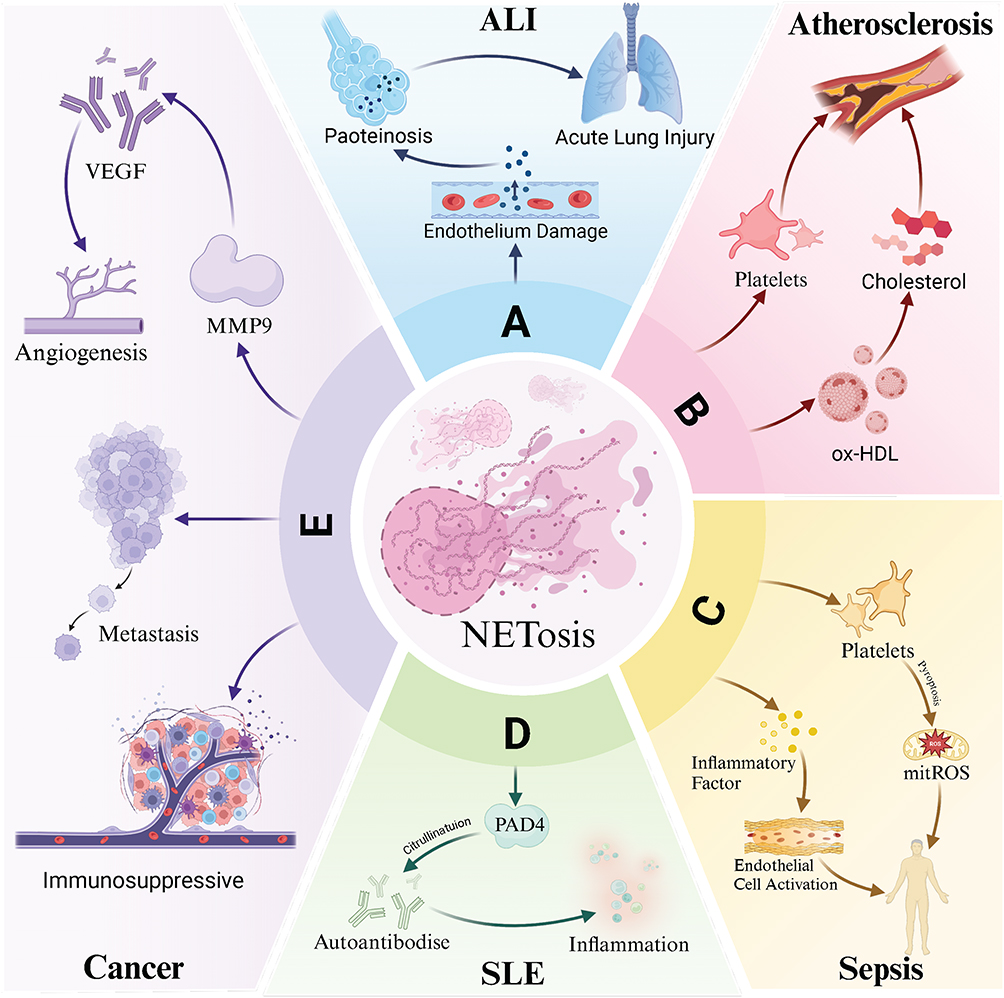 |
Figure 2 Neutrophil Extracellular Traps in Various Diseases. (A) Acute Lung Injury (ALI): ROS and other released substances cause endothelial damage and protein exudation in the alveoli, leading to the development of acute lung injury. (B) Atherosclerosis: Oxidative modification of high-density lipoprotein (HDL) reduces cholesterol excretion and induces oxidative stress, contributing to endothelial damage and platelet activation. (C) Sepsis: Inflammatory factors are released, activating endothelial cells and promoting platelet pyroptosis, which exacerbates sepsis-induced damage. (D) Systemic Lupus Erythematosus (SLE): NETs release PAD4-citrullinated autoantibodies, promoting inflammatory factor secretion and aggravating the disease. (E) Cancer: Released matrix metalloproteinase-9 (MMP-9) accelerates the release of vascular endothelial growth factor (VEGF) and promotes angiogenesis. NETs can encapsulate tumor cells to facilitate metastasis and regulate immune cells, contributing to the formation of tumor immunosuppressive niches. Abbreviations: ALI, Acute lung injury; ox-HDL, Oxidized high-density lipoprotein; mitROS, Mitochondrial reactive oxygen species; PAD4, Protein arginine deiminase 4; SLE, Systemic lupus erythematosus; MMP-9, Matrix metalloproteinase-9; VEGF, Vascular endothelial growth factor. |
Biological Characteristics of Mesenchymal Stem Cell Exosomes
Mesenchymal stem cells (MSCs) are multipotent stem cells with the capacity for self-renewal and differentiation into multiple cell lineages. They are primarily isolated from adult bone marrow and adipose tissue, though they can also be derived from embryonic stem cells (ESCs) or induced pluripotent stem cells (iPSCs). MSCs possess a variety of functions, including proliferation, pluripotency, homing, nutrition, and immunosuppression.28 The biological effects of MSCs are largely mediated through paracrine signaling, which includes the release of cytokines, growth factors, and exosomes. Exosomes are small membrane vesicles, typically less than 150 nm in diameter, secreted by cells.29 As key components of MSC paracrine signaling, exosomes carry out many of MSCs’ biological functions.
Exosome biogenesis begins with the endosomal pathway. Early endosomes are formed through the invagination of the plasma membrane (Figure 3). The endosomal sorting complex (ESCRT) then recruits specific proteins and lipids to these vesicles, resulting in the formation of late endosomes, which then develop intraluminal vesicles (ILVs) through further membrane invagination into the lumen.30 This process leads to the formation of multivesicular bodies (MVBs), which ultimately fuse with the plasma membrane and release ILVs as exosomes into the extracellular environment.30 Due to their biogenesis, composition, morphology, and function, exosomes are considered a subset of extracellular vesicles (EVs).29 In this article, exosomes and extracellular vesicles are discussed interchangeably.
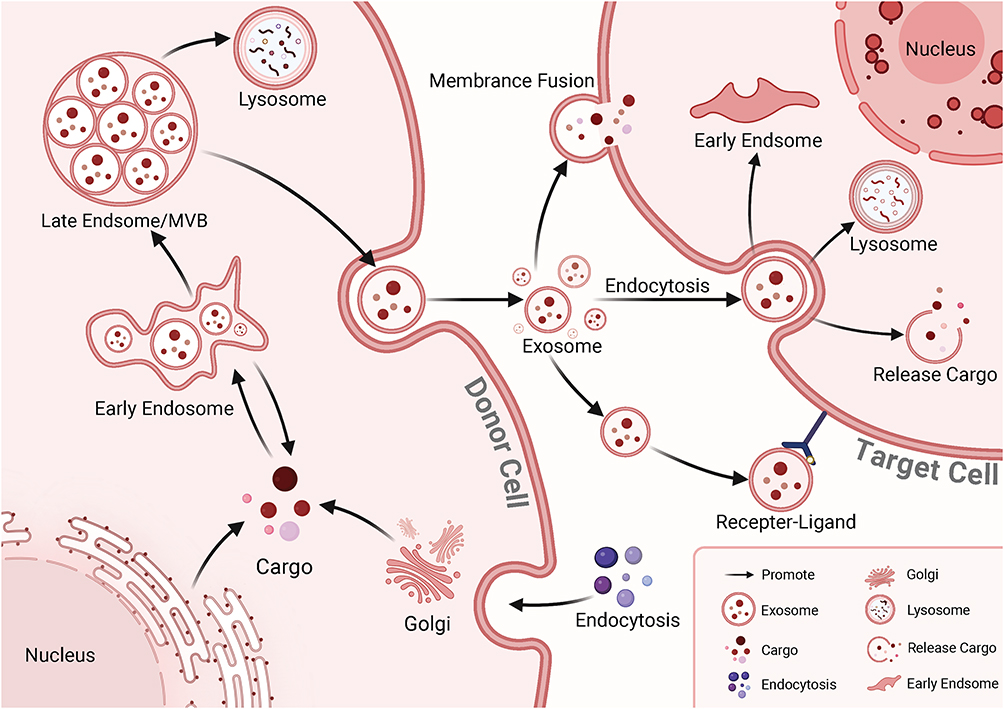 |
Figure 3 The Biogenesis and Uptake of Exosomes. Exosome formation involves several key steps: 1. The cell membrane invaginates to form early endosomes. 2. Proteins, nucleic acids, lipids, and other substances are sorted through the endosomal sorting mechanism. The endosomal membrane further invaginates to form intraluminal vesicles (ILVs), which then mature into late endosomes or multivesicular bodies (MVBs). 3. MVBs fuse with the cell membrane to release exosomes into the extracellular space. Exosomes can be internalized by recipient cells through (1) receptor binding, (2) endocytosis, or (3) direct fusion with the cell membrane. Once internalized, exosomes participate in cellular communication through three main mechanisms: (1) direct release of signaling molecules and substances, (2) participation in new endosome formation, and (3) degradation by lysosomes. |
Exosomes are rich in various proteins, including members of the tetraspanin family (CD9, CD63, CD81), membrane transport and fusion proteins (GTPases, annexins, flotillin), integrins, heat shock proteins, and endosomal sorting complex proteins such as Alix and TSG101, which are involved in protein transport.31 Exosomes also contain a variety of nucleic acids, including mRNA, miRNA, mitochondrial DNA, and lncRNA, which are crucial for intercellular communication and influence the biological functions of recipient cells. The lipid composition of the exosomal membrane includes cholesterol, phospholipids, phosphatidylethanolamine, and diacylglycerol. These lipids not only maintain the structural integrity of exosomes but also participate in various biological processes.32 In addition to standard exosome marker proteins, MSC-derived exosomes also express characteristic markers such as CD29, CD73, CD90, CD44, and CD10533 (Figure 4).
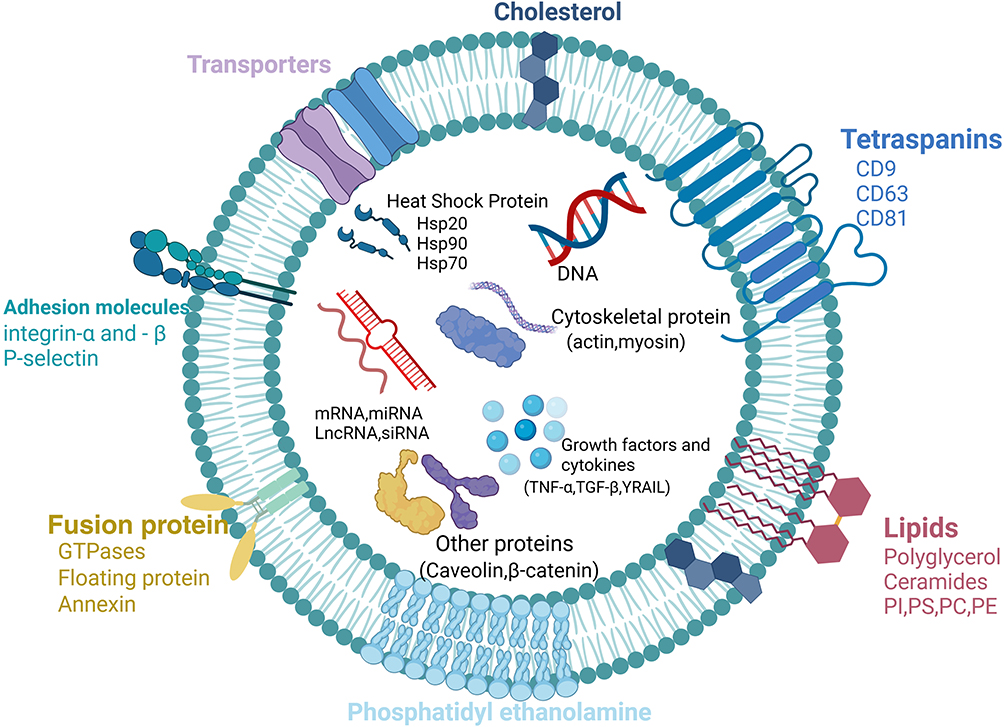 |
Figure 4 The Structure and Composition of Exosomes. Exosome proteins include the tetraspanin family (CD9, CD63, CD81), membrane transport and fusion proteins (GTPases, annexins, flotillin), integrins, heat shock proteins (Hsp20, Hsp70, Hsp90), and endosomal sorting complex proteins (Alix, TSG101). Additionally, exosomes contain various nucleic acids, such as mRNA, miRNA, mitochondrial DNA, lncRNA, and siRNA. Lipid components include cholesterol, phospholipids, phosphatidylethanolamine, and diacylglycerol. Abbreviations: PI, Phosphatidylinositol; PS, Phosphatidylserine; PC, Phosphatidylcholine; PE, Phosphatidylethanolamine. |
Mesenchymal Stem Cell Exosomes from Different Sources
Exosomes derived from MSCs from various sources share common characteristics, such as the expression of the same markers and the ability to promote anti-inflammatory, immunomodulatory, and angiogenesis-promoting effects. However, there are notable differences in their content and biological functions. Although research in this area is still limited, a systematic comparison of MSC-exosome characterization, composition, and therapeutic effects from different sources remains to be fully explored. Below, we summarize the findings from existing studies on MSC-exosomes derived from different sources.
Bone Marrow Mesenchymal Stem Cells (BMSCs): BMSCs, the first MSCs to be discovered and extensively studied, are adult stem cells isolated from bone marrow. Exosomes derived from BMSCs are rich in miRNAs. RNA sequencing has shown that the most abundant miRNAs in these exosomes are miRNA-143-3p, miRNA-10b-5p, miRNA-486-5p, miRNA-22-3p, and miRNA-21-5p, which together account for nearly half of the total miRNAs. These miRNAs play a key role in regulating MSC differentiation, migration, immune modulation, and angiogenesis.34 Proteomic analysis of BMSC-derived exosomes revealed that the most abundant membrane proteins include ADAM9, ADAM10, CD81, CACNA2D1, NOTCH2, and HLA-A, all of which are associated with bone remodeling and nerve repair.35 For example, Notch2, which enhances bone remodeling in osteoprogenitor cells, highlighting its potential in regenerative medicine.35 Clinical trials involving BMSC-EXO remain limited. Notably, Vikram Sengupta et al conducted the first clinical application of BMSC-EXO in hospitalized patients, reporting that approximately 71% of COVID-19 patients showed recovery following intravenous administration of BMSC-EXO.36
Human Umbilical Cord Mesenchymal Stem Cells (hUC-MSCs): hUC-MSCs are derived from the amniotic membrane, umbilical cord lining, Wharton’s jelly, and perivascular areas. Compared to BMSCs, hUC-MSCs offer several advantages, including painless collection and faster self-renewal, making them more accessible.37 Proteomic analysis of hUC-MSC-exosomes revealed an enrichment of PAI-1, which helps maintain endothelial homeostasis and promotes wound healing.38 Additionally, hUC-MSC-exosomes have been shown to inhibit inflammation,39 educe oxidative stress,40 and decrease cell apoptosis.41 Comparative studies show that the proteome of hUC-MSC-EXO, relative to iPSC-EXO and ESC-EXO, is more focused on immune regulation pathways, including complement activation, infection responses, and NF-κB signaling. Transcriptomic profiling indicates that miRNA-146a-5p and miRNA-320a-3p are among the most abundant miRNAs, with the miRNA profile predominantly involved in regulating immune cell activity and TNF, JAK-STAT, and NF-κB pathways.42 Due to the ease of acquisition, hUC-MSC-EXO is more widely used in clinical applications. Intrathecal injection of hUC-MSC-EXO has been reported to improve spinal cord injury outcomes without significant adverse effect,43 while aerosolized inhalation of hUC-MSC-EXO has demonstrated safety and efficacy in treating COVID-19.44
Adipose-Derived Mesenchymal Stem Cells (ASCs): ASCs are derived from adipose tissue, which provides an abundant, easily accessible source for cell collection. ASCs are more ethically acceptable than BMSCs and hUC-MSCs and can be collected on a larger scale.45 Nucleic acid analysis of ASC-derived exosomes revealed that the most abundant miRNAs include miRNA-486-5p, miRNA-10a-5p, miRNA-10b-5p, miRNA-191-5p, and miRNA-222-3p, which are associated with cell proliferation, differentiation, and angiogenesis, similar to the miRNAs found in BMSC-derived exosomes.34 In proteomic analysis, ASCs exosomes highly express antioxidant proteins such as PRDX1, PRDX2, and PRDX6.35 ASC-derived exosomes (ASC-EXO) is more frequently utilized for skin-related conditions due to its homology with skin tissues and superior absorption. Clinical studies have reported significant benefits of ASC-EXO in treating skin hyperpigmentation and acne scars.46
iPSC-MSCs are derived by reprogramming iPSCs into mesenchymal stem cells. Studies suggest that iPSC-MSC-derived exosomes (iPSC-MSC-EXO) outperform those from adult MSCs in terms of expansion potential, immune modulation, biological activity, and microenvironment regulation—advantages largely attributed to the enhanced differentiation capacity of iPSCs.47 For example, Yu Zhu et al reported that iPSC-MSC-EXO promotes greater chondrocyte migration and proliferation compared to synovial MSC-derived exosomes, resulting in superior therapeutic effects in osteoarthritis.48 Jiaqi Chen et al found that iPSC-MSC-EXO outperforms BMSC-EXO in improving cardiac function, potentially due to higher levels of miRNA-202-5p, which targets the TRAF3IP2/JNK axis to inhibit NLRP3-related pyroptosis in cardiomyocytes.49 Nonetheless, iPSC-MSCs have limitations. First, various reprogramming techniques may lead to differentiation into unintended cell types or alter exosome yield and composition. Second, there is a notable lack of clinical trials investigating iPSC-MSC-EXO, and its safety and efficacy remain to be validated. Moreover, the reported advantages of iPSC-MSC-EXO over MSC-EXO are largely theoretical, with limited experimental validation, underscoring the need for further comparative studies.
Transmission electron microscopy(TEM) revealed no significant morphological differences amongthese exosome types, all displaying the classic cup-shaped vesicular structure.35 Proteomic and transcriptomic analyses indicate that most proteins and miRNAs are shared among these exosomes, contributing to biological functions such as immunomodulation and angiogenesis. However, a small subset of unique proteins and miRNAs may underlie the distinct therapeutic effects of MSC-derived exosomes (MSC-EXO) from different sources.35,50
Mechanism of Mesenchymal Stem Cell Exosomes in Inhibiting Neutrophil Extracellular Traps
Since the pathological role of NETs has been identified, increasing attention has been given to targeting NETs as a therapeutic strategy for various diseases. However, traditional anti-NET drugs often suffer from low targeting specificity and significant side effects. This highlights the need for safer, more effective treatments. Mesenchymal stem cell exosomes have garnered interest as an alternative for anti-NET therapy due to their targeting ability, low immunogenicity, and multifunctionality. Below, we summarize the current research on MSC-exosome-based anti-NET treatment and discuss the mechanisms involved (Table 1 and Figure 5).
 |
Table 1 Mesenchymal Stem Cell Exosome Anti-Neutrophil Extracellular Traps Therapy |
 |
Figure 5 The Mechanism of Mesenchymal Stem Cell Exosomes in Anti-Neutrophil Extracellular Trap (NET) Formation. (A) Bone Marrow MSC Exosomes: These exosomes inhibit NLRP3 inflammasome activation through miRNA-199. They also prevent neutrophil activation by binding to CD64 mRNA in macrophages via miRNA-127-5p, reducing its expression. (B) Embryonic stem cells-derived mesenchymal stem cells Exosomes: These exosomes specifically bind to the C5b-9 complex, the terminal component of the NET complement activation pathway, to inhibit NET formation. (C) Human Amniotic MSC Exosomes: By inhibiting the TLR-NF-κB pathway through miRNA-125a-3p, these exosomes suppress NET formation. (D) Human-umbilical Cord MSC Exosomes: (1) These exosomes transfer mitochondria to neutrophils, restoring mitochondrial function and inhibiting NET formation. (2) They also promote neutrophil apoptosis, reducing NET formation. Abbreviations: NLRP3, NOD-like receptor pyrin domain-containing 3; TLR, Toll-like receptor; NF-κB, Nuclear factor kappa-B. |
Inhibition of Neutrophil Activation by MSC-Exosomes
Neutrophil activation and recruitment to inflammatory sites are crucial for the initiation of NETosis. Inflammatory stimuli, such as foreign pathogens and the release of inflammatory mediators, activate macrophages and neutrophils. This activation leads to the formation of NETs through the recognition of damage-associated molecular patterns (DAMPs) or pathogen-associated molecular patterns (PAMPs). Conversely, the absence of such stimuli or the lack of NETosis-associated receptors (eg, IL-1, TLRs, GPCRs, TNF, Fc receptors) prevents NET formation.57
In sepsis, a condition marked by immune dysregulation and organ dysfunction due to infection, NET levels are significantly elevated and contribute to tissue and organ damage.23 In a study by Xing-Long Zheng et al, BMSC-derived exosomes were injected into mice with sepsis-related acute lung injury, leading to a reduction in tissue damage. They found that exosomes derived from BMSCs are enriched in miRNA-127-5p, which can reach the lungs via circulation and enter target cells such as neutrophils. Within neutrophils, miRNA-127-5p binds to CD64 mRNA through complementary base pairing, inhibiting CD64 transcription and translation. CD64, a critical surface receptor on neutrophils, is closely linked to neutrophil immune responses and serves as a diagnostic and prognostic marker for sepsis.51,52 Downregulation of CD64 reduces neutrophil activation, thereby inhibiting the formation of neutrophil extracellular traps (NETs). Reduced NET formation limits the release of toxic components such as histones, MPO, and NE, mitigates inflammatory damage and pulmonary edema, and ultimately improves sepsis-induced acute lung injury (ALI) (Figure 6).
 |
Figure 6 The mechanism by which exosomes inhibit NETs. BMSC-EXO inhibits inflammatory factors S1000A8/A9 and IL-1β through the TLR4-NLRP3 pathway by miRNA-199. In addition, they can inhibited neutrophil mobilization by reducing CXCL1 and CXCL2, and thus inhibit NETs. The BMSC-EXO also specifically binds CD64 mRNA to inhibit neutrophil activation and thus inhibit NET through mi RNA 127–5p. HUC-MSC-EXO inhibits NET formation by transferring fused mitochondria and reducing mitROS production. Abbreviations: BMSC-EXO, Bone Marrow Mesenchymal Stem Cell-exosome; TLR, Toll-like receptor; NLRP3, NOD-like receptor pyrin domain-containing 3; CXCL, Chemokine (C-X-C motif) ligand 1; hUC-MSC-EXO, human umbilical cord mesenchymal stem cell-exosome. |
Mesenchymal Stem Cell Exosomes Inhibit Neutrophil Extracellular Trap Formation via TLR-NLRP3 Signaling Pathway Suppression
Pattern recognition receptors (PRRs) are molecules present on immune cells that recognize pathogen-associated molecular patterns (PAMPs) and damage-associated molecular patterns (DAMPs). Neutrophils express several PRRs, including TOLL-like receptors (TLRs), C-type lectin receptors (CLRs), and NOD-like receptors (NLRs), which have been implicated in the process of NETosis.58 Activation of NLRP3 inflammasomes via these PRRs initiates a cascade of signaling events that ultimately activate caspases and trigger gasdermin D (GSDMD)-dependent pore formation, leading to NET formation.54,59
In myocardial reperfusion injury, excessive neutrophil infiltration and NET release promote microthrombus formation through platelet binding, exacerbating microvascular obstruction and inflammation.55 S100A8/A9 released from NETs further activates NLRP3 inflammasomes, creating a vicious cycle of “NETs-NLRP3-S100A8/A9” that aggravates myocardial injury.60 Yuting Feng et al investigated the effects of BMSC-derived exosomes in a myocardial ischemia/reperfusion (I/R) injury model. They observed a reduction in neutrophil infiltration and a alleviation of the inflammatory response.53 BMSC-derived exosomes mitigate these effects through two mechanisms: they reduce neutrophil mobilization by decreasing chemokine secretion (eg, CXCL1 and CXCL2), and miRNA-199 within the exosomes downregulates the release of pro-inflammatory cytokines such as IL-1β and S100A8/A9 by targeting NLRP3 inflammasome activation. Lower S100A8/A9 levels interrupt its positive feedback on NLRP3 activation, thereby inhibiting NET formation. (Figure 6). Notably, exosomes played a crucial role in the early stages of I/R injury but did not affect later repair processes. Long-term pathological observations revealed that exosomes alleviated ventricular remodeling and fibrosis.53 Another study found that exosomes from human umbilical cord mesenchymal stem cells (hUC-MSCs) also target NLRP3 to reduce inflammation, with miRNA-378a-5p playing a central role. Additional miRNAs such as miRNA-7, miRNA-20b, miRNA-223, and miRNA-495 may help alleviate tissue damage and inflammation by targeting NLRP3, though their direct relation to NETs requires further investigation.61
Yutaka Morishima et al demonstrated that intravenous injection of exosomes derived from amniotic mesenchymal stem cells (AMSC-exo) alleviated spinal cord injury by inhibiting NET formation. The study suggested that the abundant miRNA-125a-3p in AMSC-exo might mediate this effect by inhibiting TLR-mediated NF-κB signaling. However, further research is needed to fully elucidate the mechanisms and pathways involved in AMSC-exo’s action on NETs.62
Mesenchymal Stem Cell Exosomes Inhibit Neutrophil Extracellular Trap Formation by Modulating Complement Activation
Complement system activation plays a pivotal role in NET formation. In C3- or C3 receptor-deficient mice, NET formation is impaired.63 Key complement components such as C3b and C5a are involved in neutrophil activation and the subsequent formation of the membrane attack complex (MAC, C5b-9), which promotes NET release.64
In a study on psoriasis mice, intravenous injection of MSC exosomes reduced psoriasis-associated inflammation. Specifically, MSC exosome-derived CD59 inhibited complement activation, reducing levels of C5b-9 and IL-17, thereby diminishing NETosis in neutrophils. CD59, a membrane-anchored protein, inhibits MAC formation and membrane perforation.65 This reduction in NETosis also led to the suppression of IL-17 and IL-23, cytokines crucial to psoriasis pathogenesis.66 Similarly, Loh, J.T. et al demonstrated that the initiation of complement activation led to NET release and IL-17 production in neutrophils, with MSC exosomes specifically inhibiting C5b-9 and attenuating neutrophil activation and NET formation in vitro67 (Figure 7).
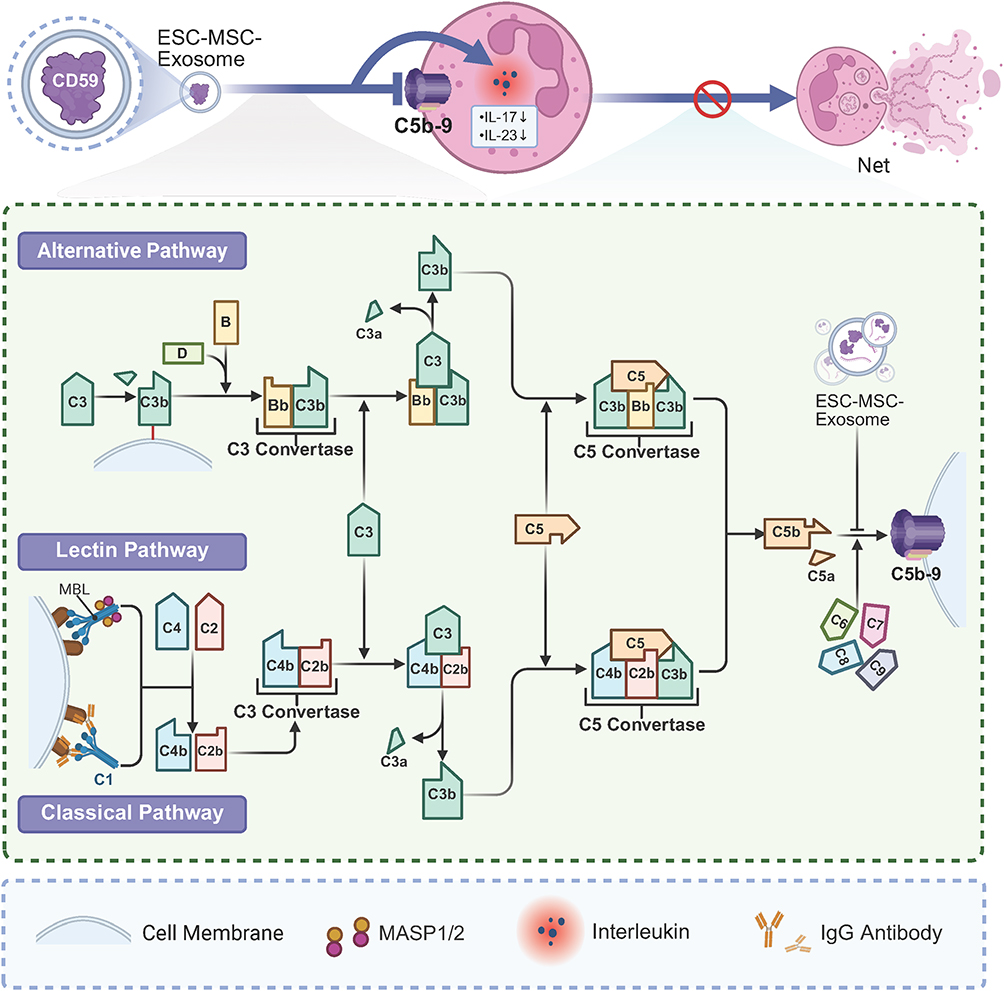 |
Figure 7 ESC-MSC-EXO modulating complement activation. ESC-MSC-EXO inhibits NET formation by inhibiting complement activation of the membrane attack complex (C5b-9) and inhibiting the release of inflammatory factors such as IL-17 and IL-23. Abbreviations: ESC-MSC-EXO, Embryonic stem cells-derived mesenchymal stem cell exosome; MASP, MBL (Mannose-Binding Lectin)-associated serine protease. |
Mesenchymal Stem Cell Exosomes Play an Anti-NET Role by Protecting Mitochondrial Function
Mitochondria play a crucial role in NETosis. They contribute to the production of mitochondrial reactive oxygen species (mitROS), which promote NET formation. Additionally, mitochondrial DNA can serve as a substrate for NET formation. Excessive mitROS production leads to mitochondrial dysfunction, which is considered a key driver of NET formation and inflammation. MSC extracellular vesicles (EVs) containing functional mitochondria have been shown to protect mitochondrial function and reduce inflammation by transferring mitochondria to recipient cells.68,69
Tongyu Lu et al first demonstrated that MSC-derived extracellular vesicles not only reduce inflammatory cytokine release from neutrophils but also inhibit NET formation by restoring mitochondrial function. Functional mitochondria within hUC-MSC-exosomes are transferred to intrahepatic neutrophils, inducing a morphological shift from fragmentation to elongation, promoting mitochondrial fusion, and restoring mitochondrial function. This fusion closes the mitochondrial permeability transition pore (mPTP) and re-establishes membrane potential, leading to reduced mitDNA, mitROS levels, and lactate production in neutrophils, thereby reducing NET formation and hepatic inflammation.70
In diabetic wounds, mitochondrial dysfunction in neutrophils drives excessive NET formation, promoting ferroptosis and impairing angiogenesis by suppressing the PI3K/AKT pathway in endothelial cells. Lu Wei et al showed that hUC-MSC-EVs transfer functional mitochondria to wound-infiltrating neutrophils, promoting mitochondrial fusion, limiting mPTP opening, restoring membrane potential, and reducing mitROS levels. This process repairs mitochondrial damage in neutrophils, decreases NET formation, prevents endothelial ferroptosis, and enhances angiogenesis and wound healing, as evidenced by the diminished inhibitory effects of MSC exosomes on NETs following rhodamine 6G-induced mitochondrial damage71 (Figure 6).
Mesenchymal Stem Cell Exosomes Redirect Apoptosis to Inhibit Neutrophil Extracellular Trap Formation
Apoptosis and NETosis are distinct forms of cell death. Redirecting neutrophils toward apoptosis can reduce NET formation. NETs have been shown to induce ferroptosis in various cell types.72 In a mouse model of abdominal aortic aneurysm, the injection of MSC-derived extracellular vesicles (MSC-EVs) significantly reduced NET formation and alleviated elastin degeneration and arterial dilation. Additionally, studies have shown that MSC-EVs can inhibit NET formation by delivering specific cargos, thereby preventing NET-induced ferroptosis in vascular smooth muscle cells (VSMCs) and slowing abdominal aortic aneurysm (AAA) progression.73 Neutrophil activation during AAA triggers NETosis,56 with released NETs suppressing the PI3K/AKT pathway, downregulating GPX4 and SLC7A11 while upregulating ACSL4 and TFR1. This cascade leads to iron accumulation, glutathione depletion, lipid peroxidation, and VSMC ferroptosis, disrupting vascular wall homeostasis.74 hUC-MSC-EVs can redirect neutrophil death, inhibit NADPH oxidase-mediated ROS bursts and PAD4-induced histone citrullination, and reduce NET release. Concurrently, they restore PI3K/AKT signaling, upregulate GPX4 and SLC7A11, suppress lipid peroxidation, and protect VSMCs from ferroptosis, thereby mitigating AAA progression.73 Notably, MSC-EVs were only effective in vivo, suggesting that exosomes may indirectly contribute to the inhibition of NETs. 59 However, the precise components of the exosomes responsible for this effect remain unidentified, and further studies are needed to assess the impact of excessive apoptosis on neutrophil function.
Overall, human ESC-derived MSC exosomes inhibit neutrophil extracellular traps (NETs) by suppressing complement activation, primarily through high levels of the membrane-anchored protein CD59. hUC-MSC-EXO mitigates mitochondrial ROS by delivering exogenous mitochondria. Shanshan Zhang et al further confirmed that hUC-MSC-EXO reduces mitochondrial oxidative stress.75 AMSC-EXO is enriched in miRNA-125a-3p, while BMSC-EXO contains high levels of miRNA-127-5p and miRNA-199, which inhibit NET formation by downregulating NET-associated signaling pathways.
The evidence overwhelmingly supports the ability of MSC-derived exosomes to exert anti-NET effects. These exosomes can inhibit NET formation or mitigate the damage caused by NETs through various mechanisms, including modulation of the TLR-NLRP3 signaling pathway, inhibition of complement activation, mitochondrial protection, and redirection of neutrophils toward apoptosis. Exosomes from different MSC sources contain varying therapeutic components and mechanisms, but due to the complex nature of exosome content, it is difficult to definitively identify the therapeutic agents responsible for these effects. miRNAs, proteins, mRNA, mitochondria, and other molecules may all contribute to the anti-NET efficacy. It is likely that a combination of mechanisms underlies the overall effects of MSC exosomes, warranting further research to more comprehensively elucidate their therapeutic potential.
Discussion
Prospects and Current Challenges of Mesenchymal Stem Cell Exosome Therapy
Exosomes are small extracellular vesicles that can carry a variety of biomolecules, offering protection from degradation in the bloodstream. They have demonstrated therapeutic efficacy in numerous animal disease models.46 Compared to mesenchymal stem cell (MSC) therapy, exosome therapy offers several advantages, including lower immunogenicity, greater biological stability, the ability to cross the blood-brain barrier, and reduced risks of pulmonary embolism and tumor formation.76 Additionally, MSC-derived exosomes (MSC-exo) can serve as drug carriers, facilitating the targeted delivery of specific therapeutic molecules. Ongoing research is focused on modifying exosome surface molecules to enhance their targeting capabilities. Wang et al recently reported that by fusing a myocardial-targeting peptide with the exosomal membrane protein gene Lamp2b and transfecting this construct alongside an adenovirus carrying miRNA-26a, they successfully generated cardiac-targeted exosomes encapsulating miRNA-26a, thereby enhancing delivery specificity to the heart.77
Despite these advantages, MSC-exosome therapy faces several challenges. One major issue is the low yield of exosomes; less than 1μg of exosome protein is typically produced per milliliter of culture medium. However, in vivo studies suggest that an effective dose of exosomes has generally been established at 10–500 μg of exosomal protein, which requires a large number of cells to generate a sufficient yield for experiments.78 Another challenge is the lack of standardization in exosome classification and quantification. Accurately determining the content and characterization of exosomes remains an ongoing issue.29 Additionally, exosomes have a short half-life in vivo, typically lasting only six hours, necessitating multiple injections to maintain effective concentrations in the body.79 Although a few MSC-exosome therapies have been tested in clinical settings, further research is needed to determine the optimal therapeutic dose, administration routes, and safety profiles for clinical use.46
Potential Strategies to Enhance Mesenchymal Stem Cell Exosome Therapy
Preclinical studies suggest that MSC-exosomes hold significant promise for anti-NET (neutrophil extracellular trap) therapy, but as noted, they have inherent limitations. To maximize their therapeutic potential, bioengineering strategies should be explored to improve exosome production and functionality. One approach is enhancing the yield of exosomes by optimizing the isolation and purification processes, such as using 3D cell culture systems. Additionally, pre-treating MSCs under hypoxic conditions may enhance the therapeutic potential of their exosomes.
3D cell culture systems offer an environment that better mimics the natural physiological conditions by promoting cell-to-cell and cell-extracellular matrix interactions. These systems also simulate more realistic biochemical reactions, which can result in more accurate therapeutic outcomes.80 Two primary types of 3D culture are scaffold-free and scaffold-based methods. Scaffold-free systems, such as spheroids, are aggregates of cells that retain the natural extracellular matrix and cellular phenotypes, better replicating in vivo conditions. Scaffold-based systems use exogenous materials, like fibrin, hyaluronic acid, or polylactic-co-glycolic acid, to support cell growth, simulating specific in vivo environments and enhancing cell yield.81 Other methods, such as cell sheet engineering, microcarriers, organ chips, organoids, and 3D bioprinting, can also be employed in 3D cell culture.
Studies have shown that human pluripotent stem cells cultured on microcarriers can secrete 17–23 times more extracellular vesicles (EVs), which are enriched with proteins and miRNAs that promote cell proliferation and reduce apoptosis.82 Similarly, adipose-derived MSCs cultured in spheroids show significantly higher exosome yields and increased expression of chondrogenesis-related miRNAs.83 Moreover, exosomes from 3D-cultured MSCs have a longer maintenance period after administration, enhanced blood-brain barrier penetration, and improved targeting abilities, which can reduce the frequency of clinical dosing.84 Overall, 3D-cultured exosomes show higher yields, enhanced therapeutic potential, and greater clinical applicability than those produced in traditional 2D cultures.
Hydrogels are emerging as promising exosome delivery platforms. Their excellent biocompatibility and adjustable physicochemical properties make them ideal for controlled exosome release. Using hydrogels allows for more precise control over the delivery of exosomes, enhancing their therapeutic effects.79 One innovative approach involves the use of hydrogen peroxide-responsive hydrogels, designed to target the site of NETosis, where hydrogen peroxide is a key mediator. These hydrogels could release exosomes directly at the site of inflammation, minimizing the wasteful consumption of exosomes elsewhere in the body or avoiding unwanted side effects.85 A recent study demonstrated that BMSC-exosomes loaded in GMOCS hydrogels could effectively disrupt NETs via the Nrf2 pathway. More notably, the hydrogel-loaded exosomes showed sustained therapeutic effects compared to controls, suggesting that these hydrogels can provide prolonged exosome release for more lasting treatments.86
MSC-exosomes pretreated with hypoxia represent another potential strategy. Hypoxia mimics the physiological environment of MSCs in vivo, enhancing their therapeutic effects, particularly in inflammatory regulation and tissue repair.87 Research has shown that EVs from hypoxic hUC-MSCs contain higher levels of miRNA-17–5p, which suppresses NET formation in diabetic wounds by inhibiting the TLR4/ROS/MAPK pathway.88 This suggests that hypoxic preconditioning could enhance MSC-exosome therapy for anti-NET applications. However, determining the specific content and the variations in exosome components following hypoxic preconditioning remains a significant challenge.87
Conclusion
MSC-exosome-based therapy offers significant promise for anti-NET treatment by regulating neutrophil activation, inhibiting NET formation signaling pathways, and protecting mitochondria. These mechanisms can effectively reduce excessive NET formation, alleviating inflammation and tissue damage in related diseases. While challenges such as exosome yield, targeting, and clinical application remain, the continued development of technology and optimization of exosome production methods suggest that MSC-exosomes could become a novel and effective treatment for NET-related conditions. Future research should focus on enhancing exosome production processes, integrating biomaterials to improve targeting, and exploring clinical applications to unlock their full therapeutic potential.
Abbreviations
AAA, abdominal aortic aneurysm; ASC, adipose tissue-derived mesenchymal Stem Cells; BMSC, bone marrow mesenchymal stem cells; CDK, cyclin-dependent kinases; CLR, C-type lectin receptors; DAMP, damage associated molecular patterns; DNase I, Deoxyribonuclease I; ERK, extracellular regulatedprotein kinase; ESC, embryonic stem cell; ESCRT, Endosomal Sorting Complexes Required for Transport; EV, extracellular vesicle; EXO, exosome; GSDMD, gasderminD; HDL, high-density lipoprotein; hUC-MSC, human umbilical cord mesenchymal stem cells; I/R, ischemia/reperfusion; IL-8, interleukin-8; ILV, intraluminal vesicles; IPSC, induced pluripotent stem cells; LPS, lipopolysaccharides; MAC, Membrane attack complex; MAP, mitogen-activated protein kinase; MEK, MAPK/ERK kinase; mitDNA, mitochondria DNA; mitROS, mitochondria ROS; MPO, myeloperoxidase; mPTP, mitochondrial permeability transition pore; MSC, Mesenchymal stem cells; MVB, multivesicular body; NADPH, nicotinamide adenine dinucleotide phosphate; NE, neutrophil elastase; NET, neutrophil extracellular trap; NLPR3, NOD-like receptor thermal protein domain associated protein 3; NLR, nucleotide oligomerization domain(NOD)-like receptors; NOX, NADPH oxidase; PAD4, protein arginine deiminase 4; PAMP, pathogen-associated molecular patterns; PKC, protein kinase C; PMA, phorbol 12-myristate 13-acetate; PRR, pattern recognition receptor; RA, rheumatoid arthritis; Raf, rapidly accelerated fibrosarcoma; ROS, Reactive oxygen species; TEM, Transmission electron microscopy; TLR, toll-like receptor; VSMC, vascular smooth muscle cells.
Acknowledgments
We would like to acknowledge the contributions of the individuals involved in this study.
Author Contributions
YY, YY, MT, YZ, ZG, CJ, YZ, and SD were responsible for the collection and analysis of literature, as well as the preparation of figures and manuscript writing. YY, MT, ZG, YZ were involved in the collecting literature and writing manuscript. YY, YY, MT, CJ were responsible for drawing pictures. SD and YZ provided final approval for the submitted version of the manuscript. We also confirm that all authors have reviewed and agreed to the final published version of the manuscript. All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This study is supported in part by National Natural Science Foundation of China (Grant No 82072161), Zhejiang Provincial Natural Science Foundation of China (Grant No Z22H158124), National Traditional Chinese Medicine Comprehensive Reform Demonstration Zone Science and Technology Co-construction Project (Grant No GZY-KJS-ZJ-2025-066).
Disclosure
The author(s) report no conflicts of interest in this work.
References
1. Borregaard N. Neutrophils, from marrow to microbes. Immunity. 2010;33(5):657–670. doi:10.1016/j.immuni.2010.11.011
2. Brinkmann V, Reichard U, Goosmann C, et al. Neutrophil extracellular traps kill bacteria. Science. 2004;303(5663):1532–1535. doi:10.1126/science.1092385
3. Papayannopoulos V. Neutrophil extracellular traps in immunity and disease. Nat Rev Immunol. 2018;18(2):134–147. doi:10.1038/nri.2017.105
4. Wang Y, Du C, Zhang Y, Zhu L. Composition and function of neutrophil extracellular traps. Biomolecules. 2024;14(4). doi:10.3390/biom14040416
5. Jorch SK, Kubes P. An emerging role for neutrophil extracellular traps in noninfectious disease. Nat Med. 2017;23(3):279–287. doi:10.1038/nm.4294
6. Zhang Y, Wu D, Sun Q, et al. Atorvastatin combined with imipenem alleviates lung injury in sepsis by inhibiting neutrophil extracellular trap formation via the ERK/NOX2 signaling pathway. Free Radic Biol Med. 2024;220:179–191. doi:10.1016/j.freeradbiomed.2024.05.006
7. Xu C, Ye Z, Jiang W, Wang S, Zhang H. Cyclosporine A alleviates colitis by inhibiting the formation of neutrophil extracellular traps via the regulating pentose phosphate pathway. Mol Med. 2023;29(1):169. doi:10.1186/s10020-023-00758-8
8. Menegazzo L, Scattolini V, Cappellari R, et al. The antidiabetic drug metformin blunts NETosis in vitro and reduces circulating NETosis biomarkers in vivo. Acta Diabetol. 2018;55(6):593–601. doi:10.1007/s00592-018-1129-8
9. Jarrahi A, Khodadadi H, Moore NS, et al. Recombinant human DNase-I improves acute respiratory distress syndrome via neutrophil extracellular trap degradation. J Thromb Haemost. 2023;21(9):2473–2484. doi:10.1016/j.jtha.2023.04.044
10. Herre M, Cedervall J, Mackman N, Olsson AK. Neutrophil extracellular traps in the pathology of cancer and other inflammatory diseases. Physiol Rev. 2023;103(1):277–312. doi:10.1152/physrev.00062.2021
11. Chamardani TM, Amiritavassoli S. Inhibition of NETosis for treatment purposes: friend or foe? Mol Cell Biochem. 2022;477(3):673–688. doi:10.1007/s11010-021-04315-x
12. Shao J, Zaro J, Shen Y. Advances in exosome-based drug delivery and tumor targeting: from tissue distribution to intracellular fate. Int J Nanomed. 2020;15:9355–9371. doi:10.2147/ijn.S281890
13. Pedrazza L, Cunha AA, Luft C, et al. Mesenchymal stem cells improves survival in LPS-induced acute lung injury acting through inhibition of NETs formation. J Cell Physiol. 2017;232(12):3552–3564. doi:10.1002/jcp.25816
14. Xiao B, Zhu Y, Liu M, et al. miR-340-3p-modified bone marrow mesenchymal stem cell-derived exosomes inhibit ferroptosis through METTL3-mediated m(6)A modification of HMOX1 to promote recovery of injured rat uterus. Stem Cell Res Ther. 2024;15(1):224. doi:10.1186/s13287-024-03846-6
15. Hade MD, Suire CN, Suo Z. Mesenchymal stem cell-derived exosomes: applications in regenerative medicine. Cells. 2021;10(8). doi:10.3390/cells10081959
16. Hidalgo A, Libby P, Soehnlein O, Aramburu IV, Papayannopoulos V, Silvestre-Roig C. Neutrophil extracellular traps: from physiology to pathology. Cardiovasc Res. 2022;118(13):2737–2753. doi:10.1093/cvr/cvab329
17. de Bont CM, Koopman WJH, Boelens WC, Pruijn GJM. Stimulus-dependent chromatin dynamics, citrullination, calcium signalling and ROS production during NET formation. Biochim Biophys Acta Mol Cell Res. 2018;1865(11 Pt A):1621–1629. doi:10.1016/j.bbamcr.2018.08.014
18. Hakkim A, Fuchs TA, Martinez NE, et al. Activation of the Raf-MEK-ERK pathway is required for neutrophil extracellular trap formation. Nat Chem Biol. 2011;7(2):75–77. doi:10.1038/nchembio.496
19. Metzler KD, Goosmann C, Lubojemska A, Zychlinsky A, Papayannopoulos V. A myeloperoxidase-containing complex regulates neutrophil elastase release and actin dynamics during NETosis. Cell Rep. 2014;8(3):883–896. doi:10.1016/j.celrep.2014.06.044
20. Wang Y, Li M, Stadler S, et al. Histone hypercitrullination mediates chromatin decondensation and neutrophil extracellular trap formation. J Cell Biol. 2009;184(2):205–213. doi:10.1083/jcb.200806072
21. Sollberger G, Choidas A, Burn GL, et al. Gasdermin D plays a vital role in the generation of neutrophil extracellular traps. Sci Immunol. 2018;3(26). doi:10.1126/sciimmunol.aar6689
22. Amulic B, Knackstedt SL, Abu Abed U, et al. Cell-cycle proteins control production of neutrophil extracellular traps. Dev Cell. 2017;43(4):449–462.e5. doi:10.1016/j.devcel.2017.10.013
23. Zhang H, Wang Y, Qu M, et al. Neutrophil, neutrophil extracellular traps and endothelial cell dysfunction in sepsis. Clin Transl Med. 2023;13(1):e1170. doi:10.1002/ctm2.1170
24. Chen J, Cao Y, Xiao J, Hong Y, Zhu Y. The emerging role of neutrophil extracellular traps in the progression of rheumatoid arthritis. Front Immunol. 2024;15:1438272. doi:10.3389/fimmu.2024.1438272
25. Moschonas IC, Tselepis AD. The pathway of neutrophil extracellular traps towards atherosclerosis and thrombosis. Atherosclerosis. 2019;288:9–16. doi:10.1016/j.atherosclerosis.2019.06.919
26. Adrover JM, McDowell SAC, He XY, Quail DF, Egeblad M. NETworking with cancer: the bidirectional interplay between cancer and neutrophil extracellular traps. Cancer Cell. 2023;41(3):505–526. doi:10.1016/j.ccell.2023.02.001
27. Ma Y, Wei J, He W, Ren J. Neutrophil extracellular traps in cancer. MedComm. 2024;5(8):e647. doi:10.1002/mco2.647
28. Naji A, Eitoku M, Favier B, Deschaseaux F, Rouas-Freiss N, Suganuma N. Biological functions of mesenchymal stem cells and clinical implications. Cell Mol Life Sci. 2019;76(17):3323–3348. doi:10.1007/s00018-019-03125-1
29. Welsh JA, Goberdhan DCI, O’Driscoll L, et al. Minimal information for studies of extracellular vesicles (MISEV2023): from basic to advanced approaches. J Extracell Vesicles. 2024;13(2):e12404. doi:10.1002/jev2.12404
30. Teng F, Fussenegger M. Shedding light on extracellular vesicle biogenesis and bioengineering. Adv Sci. 2020;8(1):2003505. doi:10.1002/advs.202003505
31. Zou Z, Li H, Xu G, Hu Y, Zhang W, Tian K. Current knowledge and future perspectives of exosomes as nanocarriers in diagnosis and treatment of diseases. Int J Nanomed. 2023;18:4751–4778. doi:10.2147/ijn.S417422
32. Liang Y, Duan L, Lu J, Xia J. Engineering exosomes for targeted drug delivery. Theranostics. 2021;11(7):3183–3195. doi:10.7150/thno.52570
33. Zhao W, Li K, Li L, et al. Mesenchymal stem cell-derived exosomes as drug delivery vehicles in disease therapy. Int J Mol Sci. 2024;25(14). doi:10.3390/ijms25147715
34. Baglio SR, Rooijers K, Koppers-Lalic D, et al. Human bone marrow- and adipose-mesenchymal stem cells secrete exosomes enriched in distinctive miRNA and tRNA species. Stem Cell Res Ther. 2015;6(1):127. doi:10.1186/s13287-015-0116-z
35. Wang ZG, He ZY, Liang S, Yang Q, Cheng P, Chen AM. Comprehensive proteomic analysis of exosomes derived from human bone marrow, adipose tissue, and umbilical cord mesenchymal stem cells. Stem Cell Res Ther. 2020;11(1):511. doi:10.1186/s13287-020-02032-8
36. Sengupta V, Sengupta S, Lazo A, Woods P, Nolan A, Bremer N. Exosomes derived from bone marrow mesenchymal stem cells as treatment for severe COVID-19. Stem Cells Dev. 2020;29(12):747–754. doi:10.1089/scd.2020.0080
37. Ding DC, Chang YH, Shyu WC, Lin SZ. Human umbilical cord mesenchymal stem cells: a new era for stem cell therapy. Cell Transplant. 2015;24(3):339–347. doi:10.3727/096368915x686841
38. Zhang Z, Mi T, Jin L, et al. Comprehensive proteomic analysis of exosome mimetic vesicles and exosomes derived from human umbilical cord mesenchymal stem cells. Stem Cell Res Ther. 2022;13(1):312. doi:10.1186/s13287-022-03008-6
39. Xiang E, Han B, Zhang Q, et al. Human umbilical cord-derived mesenchymal stem cells prevent the progression of early diabetic nephropathy through inhibiting inflammation and fibrosis. Stem Cell Res Ther. 2020;11(1):336. doi:10.1186/s13287-020-01852-y
40. Che J, Wang H, Dong J, et al. Human umbilical cord mesenchymal stem cell-derived exosomes attenuate neuroinflammation and oxidative stress through the NRF2/NF-κB/NLRP3 pathway. CNS Neurosci Ther. 2024;30(3):e14454. doi:10.1111/cns.14454
41. Li Z, Tian Y, Liu Y, et al. Human umbilical cord mesenchymal stem cell‑derived exosomes improve ovarian function in natural aging by inhibiting apoptosis. Int J Mol Med. 2023;52(4). doi:10.3892/ijmm.2023.5297
42. Malik SZA, Muhilan Y, Nordin F, et al. Stem cell derived exosome trilogy: an epic comparison of human MSCs, ESCs and iPSCs. Stem Cell Res Ther. 2025;16(1):318. doi:10.1186/s13287-025-04440-0
43. Akhlaghpasand M, Tavanaei R, Hosseinpoor M, et al. Safety and potential effects of intrathecal injection of allogeneic human umbilical cord mesenchymal stem cell-derived exosomes in complete subacute spinal cord injury: a first-in-human, single-arm, open-label, Phase I clinical trial. Stem Cell Res Ther. 2024;15(1):264. doi:10.1186/s13287-024-03868-0
44. Chu M, Wang H, Bian L, et al. Nebulization therapy with umbilical cord mesenchymal stem cell-derived exosomes for COVID-19 pneumonia. Stem Cell Rev Rep. 2022;18(6):2152–2163. doi:10.1007/s12015-022-10398-w
45. Bunnell BA. Adipose tissue-derived mesenchymal stem cells. Cells. 2021;10(12). doi:10.3390/cells10123433
46. Lotfy A, AboQuella NM, Wang H. Mesenchymal stromal/stem cell (MSC)-derived exosomes in clinical trials. Stem Cell Res Ther. 2023;14(1):66. doi:10.1186/s13287-023-03287-7
47. Zhou AK, Jou E, Lu V, et al. Using pre-clinical studies to explore the potential clinical uses of exosomes secreted from induced pluripotent stem cell-derived mesenchymal stem cells. Tissue Eng Regen Med. 2023;20(6):793–809. doi:10.1007/s13770-023-00557-6
48. Zhu Y, Wang Y, Zhao B, et al. Comparison of exosomes secreted by induced pluripotent stem cell-derived mesenchymal stem cells and synovial membrane-derived mesenchymal stem cells for the treatment of osteoarthritis. Stem Cell Res Ther. 2017;8(1):64. doi:10.1186/s13287-017-0510-9
49. Chen J, Liang X, Han Q, et al. Exosomal miR-202-5p derived from iPSC-MSCs protects against myocardial infarction through inhibition of cardiomyocyte pyroptosis. Stem Cell Res Ther. 2025;16(1):282. doi:10.1186/s13287-025-04390-7
50. Soni N, Gupta S, Rawat S, Krishnakumar V, Mohanty S, Banerjee A. MicroRNA-enriched exosomes from different sources of mesenchymal stem cells can differentially modulate functions of immune cells and neurogenesis. Biomedicines. 2021;10(1):69. doi:10.3390/biomedicines10010069
51. Pham HM, Nguyen DLM, Duong MC, et al. Neutrophil CD64-a prognostic marker of sepsis in intensive care unit: a prospective cohort study. Front Med Lausanne. 2023;10:1251221. doi:10.3389/fmed.2023.1251221
52. Yin WP, Li JB, Zheng XF, An L, Shao H, Li CS. Effect of neutrophil CD64 for diagnosing sepsis in emergency department. World J Emerg Med. 2020;11(2):79–86. doi:10.5847/wjem.j.1920-8642.2020.02.003
53. Feng Y, Bao X, Zhao J, Kang L, Sun X, Xu B. MSC-derived exosomes mitigate myocardial ischemia/reperfusion injury by reducing neutrophil infiltration and the formation of neutrophil extracellular traps. Int J Nanomed. 2024;19:2071–2090. doi:10.2147/ijn.S436925
54. Chen KW, Monteleone M, Boucher D, et al. Noncanonical inflammasome signaling elicits gasdermin D-dependent neutrophil extracellular traps. Sci Immunol. 2018;3(26). doi:10.1126/sciimmunol.aar6676
55. Perdomo J, Leung HHL, Ahmadi Z, et al. Neutrophil activation and NETosis are the major drivers of thrombosis in heparin-induced thrombocytopenia. Nat Commun. 2019;10(1):1322. doi:10.1038/s41467-019-09160-7
56. Spinosa M, Su G, Salmon MD, et al. Resolvin D1 decreases abdominal aortic aneurysm formation by inhibiting NETosis in a mouse model. J Vasc Surg. 2018;68(6s):93s–103s. doi:10.1016/j.jvs.2018.05.253
57. Thiam HR, Wong SL, Wagner DD, Waterman CM. Cellular mechanisms of NETosis. Annu Rev Cell Dev Biol. 2020;36:191–218. doi:10.1146/annurev-cellbio-020520-111016
58. Huang J, Hong W, Wan M, Zheng L. Molecular mechanisms and therapeutic target of NETosis in diseases. MedComm. 2022;3(3):e162. doi:10.1002/mco2.162
59. Münzer P, Negro R, Fukui S, et al. NLRP3 inflammasome assembly in neutrophils is supported by PAD4 and promotes netosis under sterile conditions. Front Immunol. 2021;12:683803. doi:10.3389/fimmu.2021.683803
60. Sreejit G, Abdel-Latif A, Athmanathan B, et al. Neutrophil-derived S100A8/A9 amplify granulopoiesis after myocardial infarction. Circulation. 2020;141(13):1080–1094. doi:10.1161/circulationaha.119.043833
61. Cai X, Zhang ZY, Yuan JT, et al. hucMSC-derived exosomes attenuate colitis by regulating macrophage pyroptosis via the miR-378a-5p/NLRP3 axis. Stem Cell Res Ther. 2021;12(1):416. doi:10.1186/s13287-021-02492-6
62. Morishima Y, Kawabori M, Yamazaki K, et al. Intravenous administration of mesenchymal stem cell-derived exosome alleviates spinal cord injury by regulating neutrophil extracellular trap formation through exosomal miR-125a-3p. Int J Mol Sci. 2024;25(4):2406. doi:10.3390/ijms25042406
63. Yipp BG, Petri B, Salina D, et al. Infection-induced NETosis is a dynamic process involving neutrophil multitasking in vivo. Nat Med. 2012;18(9):1386–1393. doi:10.1038/nm.2847
64. de Bont CM, Boelens WC, Pruijn GJM. NETosis, complement, and coagulation: a triangular relationship. Cell Mol Immunol. 2019;16(1):19–27. doi:10.1038/s41423-018-0024-0
65. Couves EC, Gardner S, Voisin TB, et al. Structural basis for membrane attack complex inhibition by CD59. Nat Commun. 2023;14(1):890. doi:10.1038/s41467-023-36441-z
66. Zhang B, Lai RC, Sim WK, Choo ABH, Lane EB, Lim SK. Topical application of mesenchymal stem cell exosomes alleviates the imiquimod induced psoriasis-like inflammation. Int J Mol Sci. 2021;22(2). doi:10.3390/ijms22020720
67. Loh JT, Zhang B, Teo JKH, et al. Mechanism for the attenuation of neutrophil and complement hyperactivity by MSC exosomes. Cytotherapy. 2022;24(7):711–719. doi:10.1016/j.jcyt.2021.12.003
68. Thomas MA, Fahey MJ, Pugliese BR, Irwin RM, Antonyak MA, Delco ML. Human mesenchymal stromal cells release functional mitochondria in extracellular vesicles. Front Bioeng Biotechnol. 2022;10:870193. doi:10.3389/fbioe.2022.870193
69. Dutra Silva J, Su Y, Calfee CS, et al. Mesenchymal stromal cell extracellular vesicles rescue mitochondrial dysfunction and improve barrier integrity in clinically relevant models of ARDS. Eur Respir J. 2021;58(1):2002978. doi:10.1183/13993003.02978-2020
70. Lu T, Zhang J, Cai J, et al. Extracellular vesicles derived from mesenchymal stromal cells as nanotherapeutics for liver ischaemia-reperfusion injury by transferring mitochondria to modulate the formation of neutrophil extracellular traps. Biomaterials. 2022;284:121486. doi:10.1016/j.biomaterials.2022.121486
71. Lu W, Li X, Wang Z, et al. Mesenchymal stem cell-derived extracellular vesicles accelerate diabetic wound healing by inhibiting NET-induced ferroptosis of endothelial cells. Int J Biol Sci. 2024;20(9):3515–3529. doi:10.7150/ijbs.97150
72. Zhang H, Wu D, Wang Y, et al. METTL3-mediated N6-methyladenosine exacerbates ferroptosis via m6A-IGF2BP2-dependent mitochondrial metabolic reprogramming in sepsis-induced acute lung injury. Clin Transl Med. 2023;13(9):e1389. doi:10.1002/ctm2.1389
73. Chen L, Liu Y, Wang Z, et al. Mesenchymal stem cell-derived extracellular vesicles protect against abdominal aortic aneurysm formation by inhibiting NET-induced ferroptosis. Exp Mol Med. 2023;55(5):939–951. doi:10.1038/s12276-023-00986-2
74. Zhang X, Li LX, Ding H, Torres VE, Yu C, Li X. Ferroptosis promotes cyst growth in autosomal dominant polycystic kidney disease mouse models. J Am Soc Nephrol. 2021;32(11):2759–2776. doi:10.1681/asn.2021040460
75. Zhang S, Zou X, Feng X, et al. Exosomes derived from hypoxic mesenchymal stem cell ameliorate premature ovarian insufficiency by reducing mitochondrial oxidative stress. Sci Rep. 2025;15(1):8235. doi:10.1038/s41598-025-90879-3
76. Tan F, Li X, Wang Z, Li J, Shahzad K, Zheng J. Clinical applications of stem cell-derived exosomes. Signal Transduct Target Ther. 2024;9(1):17. doi:10.1038/s41392-023-01704-0
77. Wang H, Jiang R, Zhong F, et al. Ultrasound-triggered targeted delivery of engineered ADSCs-derived exosomes with high SDF-1α levels to promote cardiac repair following myocardial infarction. Int J Pharm. 2025;681:125786. doi:10.1016/j.ijpharm.2025.125786
78. Kimiz-Gebologlu I, Oncel SS. Exosomes: large-scale production, isolation, drug loading efficiency, and biodistribution and uptake. J Control Release. 2022;347:533–543. doi:10.1016/j.jconrel.2022.05.027
79. Fan MH, Pi JK, Zou CY, et al. Hydrogel-exosome system in tissue engineering: a promising therapeutic strategy. Bioact Mater. 2024;38:1–30. doi:10.1016/j.bioactmat.2024.04.007
80. Chen W, Wu P, Jin C, Chen Y, Li C, Qian H. Advances in the application of extracellular vesicles derived from three-dimensional culture of stem cells. J Nanobiotechnol. 2024;22(1):215. doi:10.1186/s12951-024-02455-y
81. Casajuana Ester M, Day RM. Production and utility of extracellular vesicles with 3d culture methods. Pharmaceutics. 2023;15(2):663. doi:10.3390/pharmaceutics15020663
82. Urrata V, Toia F, Cammarata E, et al. Characterization of the secretome from spheroids of adipose-derived stem cells (SASCs) and its potential for tissue regeneration. Biomedicines. 2024;12(8):1842. doi:10.3390/biomedicines12081842
83. Chen J, Zhang E, Wan Y, Huang T, Wang Y, Jiang H. A quick and innovative pipeline for producing chondrocyte-homing peptide-modified extracellular vesicles by three-dimensional dynamic culture of hADSCs spheroids to modulate the fate of remaining ear chondrocytes in the M1 macrophage-infiltrated microenvironment. J Nanobiotechnol. 2024;22(1):300. doi:10.1186/s12951-024-02567-5
84. Lin G, Pan W, He Y, Yi X, Zhou P, Lu J. Opportunities and perspectives for three dimensional culture of mesenchymal stem cell-derived exosomes. Tissue Eng Part B Rev. 2024;30(4):462–476. doi:10.1089/ten.TEB.2023.0253
85. Pan S, Yin Z, Shi C, et al. Multifunctional injectable hydrogel microparticles loaded with miR-29a abundant BMSCs derived exosomes enhanced bone regeneration by regulating osteogenesis and angiogenesis. Small. 2024;20(16):e2306721. doi:10.1002/smll.202306721
86. Zhou R, Guo J, Jin Z. Advancing osteoarthritis therapy with GMOCS hydrogel-loaded BMSCs-exos. J Nanobiotechnol. 2024;22(1):493. doi:10.1186/s12951-024-02713-z
87. Zhuo H, Chen Y, Zhao G. Advances in application of hypoxia-preconditioned mesenchymal stem cell-derived exosomes. Front Cell Dev Biol. 2024;12:1446050. doi:10.3389/fcell.2024.1446050
88. Chu Z, Huang Q, Ma K, et al. Novel neutrophil extracellular trap-related mechanisms in diabetic wounds inspire a promising treatment strategy with hypoxia-challenged small extracellular vesicles. Bioact Mater. 2023;27:257–270. doi:10.1016/j.bioactmat.2023.04.007
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Neutrophil Extracellular Traps Aggravate Contrast-Induced Acute Kidney Injury by Damaging Glomeruli and Peritubular Capillaries
Wang H, Gao T, Zhang R, Hu J, Gao S, Wang Y, Qi X, Zhou Y, Zheng G, Dong H
Journal of Inflammation Research 2023, 16:5629-5646
Published Date: 28 November 2023
Identification and Validation of Hub Genes Related to Neutrophil Extracellular Traps-Mediated Cell Damage During Myocardial Infarction

Ke D, Ni J, Yuan Y, Cao M, Chen S, Zhou H
Journal of Inflammation Research 2024, 17:617-637
Published Date: 1 February 2024
Neutrophil Extracellular Traps Enhance Procoagulant Activity and Predict Poor Prognosis in Patients With Metastatic Breast Cancer
Gong Y, Chen B, Tan Q, Wei W
International Journal of General Medicine 2025, 18:1247-1259
Published Date: 4 March 2025
The Role of Neutrophils in Non-Alcoholic Fatty Liver Disease: Mechanisms and Clinical Significance
Zhang F, Li W
Journal of Inflammation Research 2026, 19:585447
Published Date: 4 March 2026
Neutrophil Extracellular Traps in Dry Eye: A Comprehensive Review of Pathogenic Mechanisms and Implications for Targeted Therapy
Guo X, Wang C, Liu W, Sun Y, Su L, Wang H, Liu B
International Journal of General Medicine 2026, 19:583609
Published Date: 23 March 2026