Back to Journals » Drug Design, Development and Therapy » Volume 16
Integrating Network Pharmacology and Experimental Verification to Explore the Pharmacological Mechanisms of Aloin Against Gastric Cancer
Authors Gao J, Yang S, Xie G ![]() , Pan J, Zhu F
, Pan J, Zhu F ![]()
Received 1 March 2022
Accepted for publication 30 May 2022
Published 20 June 2022 Volume 2022:16 Pages 1947—1961
DOI https://doi.org/10.2147/DDDT.S360790
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Prof. Dr. Tin Wui Wong
Jia Gao,1,* Sifu Yang,2,* Guanqun Xie,3 Jieli Pan,4 Feiye Zhu4
1The Second Clinical Medical College, Zhejiang Chinese Medical University, Hangzhou, Zhejiang, 310053, People’s Republic of China; 2Medical Oncology Department, Zhejiang Provincial People’s Hospital, Hangzhou, Zhejiang, 310014, People’s Republic of China; 3College of Basic Medical Science, Zhejiang Chinese Medical University, Hangzhou, Zhejiang, 310053, People’s Republic of China; 4Academy of Chinese Medical Sciences, Zhejiang Chinese Medical University, Hangzhou, Zhejiang, 310053, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Jieli Pan; Feiye Zhu, Academy of Chinese Medical Sciences, Zhejiang Chinese Medical University, Hangzhou, Zhejiang, 310053, People’s Republic of China, Email [email protected]; [email protected]
Purpose: This study was designed to evaluate the pharmacological mechanisms of Aloin against gastric cancer (GC) via network pharmacology analysis combined with experimental verification.
Methods: Using network pharmacology methods, the potential targets of Aloin and targets related to GC were screened from public databases. The protein–protein interaction (PPI) network, Gene Ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway enrichment analysis were performed to predict the core targets and pathways of Aloin against GC. The expressions of major targets predicted by network pharmacology in normal stomach tissues and GC tissues and their relationships with overall survival of GC were searched in GEPIA, HPA and DriverDBv3 database. The results of network pharmacology analysis were verified by in vitro experiments.
Results: A total of 129 potential targets were retrieved by searching the intersection of Aloin and GC targets. PPI network analysis indicated that 10 targets, including AKT1 and CASP3, were hub genes. GO enrichment analysis involved 93 biological processes, 19 cellular components, and 37 molecular functions. KEGG enrichment analysis indicated that the anti-cancer effect of Aloin was mediated through multiple pathways, such as PI3K-AKT, FoxO and Ras signaling pathway. Among them, the PI3K-AKT signaling pathway, which contained the largest number of enriched genes, may play a greater role in the treatment of GC. The validation of key targets in GEPIA, HPA and DriverDBv3 database showed that the verification results for most core genes were consistent with this study. Then, the results of in vitro experiment indicated that Aloin could inhibit proliferation of NCI-N87 cells and induce cell apoptosis. The results also showed that Aloin could decrease the mRNA and protein expressions of PI3K and AKT, suggesting that Aloin can treat GC by inducing cell apoptosis and regulating the PI3K-AKT signaling pathway.
Conclusion: This study identified the potential targets of Aloin against GC using network pharmacology and in vitro verification, which provided a new understanding of the pharmacological mechanisms of Aloin in treatment of GC.
Keywords: gastric cancer, Aloin, pharmacological mechanisms, network pharmacology, experimental verification
Introduction
Gastric cancer (GC) is the most prevalent digestive tract malignant tumor that originates from gastric mucosal epithelial cells. The deaths related to GC account for approximately 23% of all cancer deaths. In addition, the prognosis of advanced GC was poor, with the brutal 5-year survival rate still less than 20%.1 Chemotherapy, one of the treatment methods of GC, has serious side effects and can result in drug resistance. Natural anti-cancer compound is a promising medicine for tumor treatment due to its better efficacy, fewer side effects and lower toxicity. Therefore, the study on natural anti-cancer drugs is of great significance in tumor treatment and prognosis.
Aloin, an anthraquinone compound extracted from the aloe that employed as a dietary supplement in a variety of foods, has the effects of anti-oxidation and anti-inflammation. In recent years, an increasing number of studies extended the pharmacological activity of Aloin to tumor treatment.2–4 Tao et al reported that Aloin induced gastric cancer cell apoptosis by down-regulating expressions of HMGB1 and RAGE, inhibiting HMGB1 release and suppressing rhHMGB1-induced activation of Akt-mTOR-P70S6K and ERK-P90RSK-CREB signalling pathways.5 Wang et al found Aloin inhibited the proliferation and migration of GC cells by regulating NOX2-ROS-mediated pro-survival signal pathways.6 However, the mechanisms of Aloin in the treatment of GC are multi-targets, unilateral factors and single target cannot uncover the complex mechanisms of Aloin.
Network pharmacology is a new valuable approach that integrates systems biology and bioinformatics to elaborate the complex mechanisms of drugs.7 It provides a theoretical basis for further research of natural medicine.8 Therefore, we applied network pharmacology to determine the potential targets of Aloin and reveal its complicated mechanisms in the treatment of GC. The detailed process of this study is shown in Figure 1.
 |
Figure 1 Detailed process of present study. |
Materials and Methods
Network Pharmacology Analysis
Potential Targets of Aloin
The molecular structure (sdf format) of Aloin was obtained from PubChem database (https://pubchem.ncbi.nlm.nih.gov/). The potential targets of Aloin were screened from the following databases: PharmMapper (http://lilab.ecust.edu.cn/pharmmapper/), Swiss Target Prediction (http://www.swisstargetprediction.ch/) and Traditional Chinese Medicine Systems Pharmacology (TCMSP) database (https://www.tcmsp-e.com/). Then, we submitted the collected targets into Uniprot database (https://www.uniprot.org/) and limited the species to “Homo sapiens”. The official symbols of all gene targets were obtained.
Identification of GC-Related Gene Targets
GC-related targets were retrieved from Online Mendelian Inheritance in Man (OMIM, https://omim.org/), GeneCards (https://www.genecards.org/) and DisGeNET (https://disgenet.org/). The keyword “gastric cancer” was applied as search term and duplicate target genes were removed. Venn diagram was drawn for analyzing the overlapping genes to obtain potential targets of Aloin against GC.
Protein–Protein Interaction (PPI) Network Construction
The overlapping genes were imported into STRING database (https://string-db.org/) to construct the PPI network. The Cytoscape (version 3.8.2) was used to analyze the complicated relationships among genes visually.
Enrichment Analysis and “Disease Target-Pathway” Network Construction
Gene Ontology (GO) Enrichment and Kyoto Encyclopedia of Genes and Genomes (KEGG) Pathway Enrichment were conducted using DAVID database (https://david.ncifcrf.gov/). GO Enrichment included biological process (BP), cellular component (CC) and molecular function (MF). P<0.05 and FDR <0.05 were recognized as significant of GO Enrichment. P<0.01 and FDR <0.01 were recognized as significant of KEGG Enrichment. The “Disease target-Pathway” network was performed using Cytoscape (version 3.8.2).
Validation of Key Targets in GEPIA, HPA and DriverDBv3 Database
The online tool of GEPIA (http://gepia.cancer-pku.cn/index.html) was used to verify the expressions of 10 key targets in TCGA-STAD. The protein expressions and distribution of 10 key targets in normal stomach and GC tissues were retrieved in HPA database (https://www.proteinatlas.org/). The overall survival of 10 core genes was searched in DriverDBv3database (http://driverdb.tms.cmu.edu.tw/).
Experimental Verification in vitro
Cells and Reagents
Human GC cell line NCI-N87 was purchased from Shanghai Institute of Cell Biology, Chinese Academy of Sciences (Shanghai, China). Aloin (purity ≥95%, CID 12305761) was obtained from Shanghai Source Leaf Biological Technology Co., Ltd (Shanghai, China). Fetal bovine serum (FBS) and Roswell Park Memorial Institute-1640 (RPMI-1640) medium were bought from Gibco (Carlsbad, CA, USA). Bicinchoninic acid (BCA) protein assay kit and RIPA lysis buffer were purchased from Beyotime Institute of Biotechnology (Shanghai, China). Antibodies against Cleaved Caspase-3, AKT, p-AKT (Ser473), and GAPDH were obtained from Cell signaling technology (Danvers, MA, USA). Antibodies against PI3K and p-PI3K were provided by Abcam (Cambridge, UK). Wes separation kit was obtained from ProteinSimple (California, USA). Cell Counting Kit-8 (CCK8) was bought from Dojindo Laboratories (Tokyo, Japan). The total RNA rapid extraction kit, HiFiScript Quick gDNA Removal cDNA Synthesis kit and SYBR green PCR Master Mix were purchased from Beijing Biotek Biotechnology Co., Ltd (Beijing, China). The PCR primers were synthesized by the Sangon Biotech (Shanghai) Co., Ltd (Shanghai, China).
Cell Culture and Viability Analysis
NCI-N87 cells were cultured in RPMI-1640 medium containing 10% FBS and 1% penicillin-streptomycin in a 5% CO2 humidified incubator at 37°C.
Aloin was dissolved in dimethyl sulfoxide (DMSO) and diluted into series concentrations with culture medium (the final concentration of DMSO was 0.1%). To detect the inhibitory effect of Aloin on cell proliferation, NCI-N87 cells in logarithmic growth phase were seeded in 96-well plate at a density of 5×103 cells per well and treated with Aloin at multiple concentrations (100, 200, 300, 400, 500 μg/mL) or vehicle (medium containing 0.1% DMSO) for 24h. After treatment, each well was added with 10 μL of CCK8 solution and incubated at 37°C for another 2h. The absorbance at 450nm was detected by a microplate reader (Bio-Tek, Winooski, VT, USA).
RT-qPCR Analysis
By using total RNA rapid extraction kit, total RNA was extracted from cells that had been treated with different concentrations of Aloin for 24h. The concentration of total RNA was determined by a NanoDrop One Spectrophotometer (Thermo Scientific, USA). The total RNA was reverse transcribed to cDNA by the HiFiScript gDNA Removal cDNA Synthesis Kit. The PCR reaction system was prepared using the SYBR green PCR Master Mix (Low ROX) kit, the cDNA was amplified and the expression levels were detected by the Roche LightCycler 480 (Roche Diagnostics, Basel, Switzerland). The relative expressions of the target genes were computed by 2−ΔΔCt method. Table 1 shows the primer sequences.
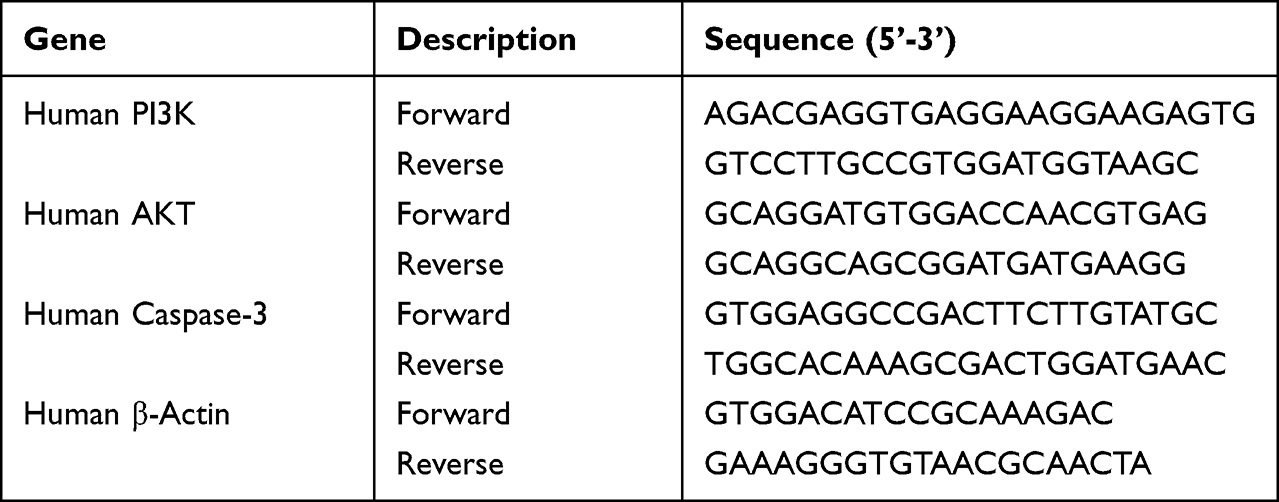 |
Table 1 Primer Sequences in RT-qPCR |
Transmission Electron Microscope (TEM)
NCI-N87 cells (5×106 cells per dish) were collected and fixed with 2.5% glutaraldehyde overnight at 4°C after treatment with Aloin at different concentrations for 24h, then fixed with 1% (w/v) osmium acid for 1h, dehydrated through gradient ethanol-acetone. Samples were packed with different proportions of embedding-dehydrating agent and then immersed in pure embedding agent overnight. After polymerizing at 70°C for 24h the next day, samples were cut into ultrathin sections of 70nm by ultramicrotome (Leica, Germany), and double-stained with uranyl acetate and lead citrate. Then, the ultrastructure of cells was observed under the TEM (Hitachi, Tokyo, Japan).
Automated Western Immunoblotting Analysis
NCI-N87 cells were collected and lysed in extraction buffer containing protease inhibitor and PMSF for 2h on ice after treatment with Aloin at different concentrations for 24h. Centrifugation was operated on the lysates at 12,000 rpm for 15 min. BCA protein assay kit was used to determine the concentration of protein. The Wes separation kit was used to perform simple Western immunoblotting on a Peggy Sue system (ProteinSimple, California, USA) and detect the expression of protein in the light of the manufacturer’s instructions. GAPDH was added as an internal amplification control.
Statistical Analysis
The experimental data were presented as mean ±standard deviation. Software SPSS16.0 and GraphPad Prism 8, method One-way ANOVA were used to analyze the result of data. A level of P<0.05 was deemed significant.
Results
Acquisition of Potential Targets of Aloin Against GC
Based on PharmMapper database, Swiss Target Prediction and TCMSP database, a total of 213 potential targets were identified for Aloin. GC-related gene targets were obtained from OMIM, GeneCards and DisGeNET. A total of 4034 targets were selected. We intersected the 213 target genes of Aloin with the 4034 potential GC-related target genes. There were 129 intersections considered as potential candidate targets of Aloin against GC (Figure 2, Supplementary Table 1).
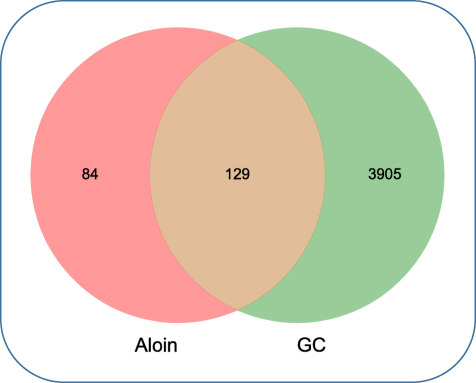 |
Figure 2 Venn diagram for the overlap analysis of Aloin targeted genes and GC-related genes. |
PPI Network Construction and Key Targets Screening
After submitting all predicted target genes into the STRING database, the medium confidence (0.4) was set for analysis. The PPI network contained 126 nodes and 1078 edges was constructed by Cytoscape (version 3.8.2) (Figure 3A). In the PPI network, the top 10 hub genes with Cytohubba were screened by Degree method: ALB (degree = 79), AKT1 (degree = 75), EGFR (degree = 64), MAPK1 (degree = 59), CASP3 (degree = 57), HSP90AA1 (degree = 56), MAPK8 (degree = 54), SRC (degree = 54), HRAS (degree =53) and ESR1 (degree = 53) (Figure 3B and C). MCODE was used to reveal seven gene clusters of Aloin against GC (Figure 4, Supplementary Table 2). The highest score was cluster 1 containing 27 nodes and 235 edges.
 |
Figure 3 PPI network of Aloin targets against GC. (A) The PPI network is constructed by Cytoscape. The node size and color stand for the size of the degree. Node size is proportional to its degree. The node color is from yellow to purple and the corresponding degree is gradually larger. (B) The top 10 hub genes cluster generated from (A). (C) Hub genes list for Aloin on GC by Degree method. |
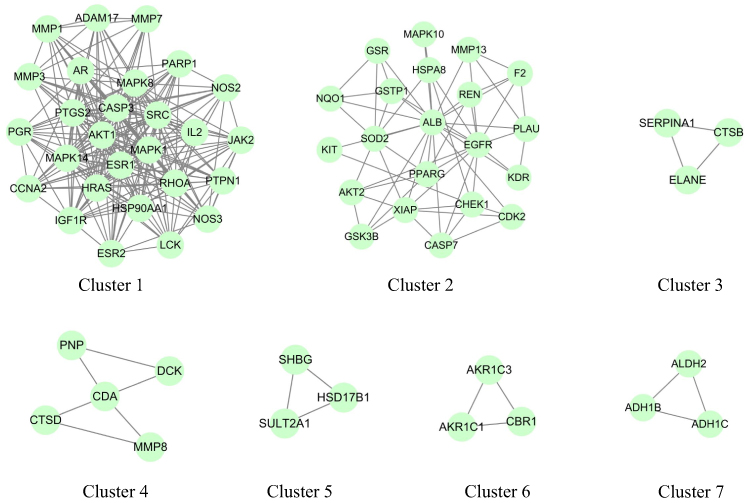 |
Figure 4 The gene clusters of Aloin against GC PPI network. The cluster from 1 to 7 is based on their cluster score; genes in Cluster 1 has the highest score. |
GO Enrichment and KEGG Enrichment
GO and KEGG pathway enrichment analyses were performed to explore the biological functions of the 129 predicted genes in DAVID database. The result showed that the 129 potential target genes of Aloin against GC were significantly enriched in 93 biological processes, 19 cellular components, and 37 molecular functions (P<0.05 and FDR <0.05) (Figure 5, supplementary Tables 3–5). The top 5 terms of biological processes were mainly enriched in peptidyl-serine phosphorylation (GO: 0018105), protein autophosphorylation (GO: 0046777), peptidyl-threonine phosphorylation (GO: 0018107), negative regulation of apoptotic process (GO: 0043066) and protein phosphorylation (GO: 0006468). The top 5 terms of cellular components were cytosol (GO: 0005829), extracellular exosome (GO: 0070062), extracellular space (GO: 0005615), extracellular region (GO: 0005576) and extrinsic component of cytoplasmic side of plasma membrane (GO: 0031234). The top 5 terms of molecular functions included serine-type endopeptidase activity (GO: 0004252), steroid hormone receptor activity (GO: 0003707), protein tyrosine kinase activity (GO: 0004713), kinase activity (GO: 0016301) and ATP binding (GO: 0005524).
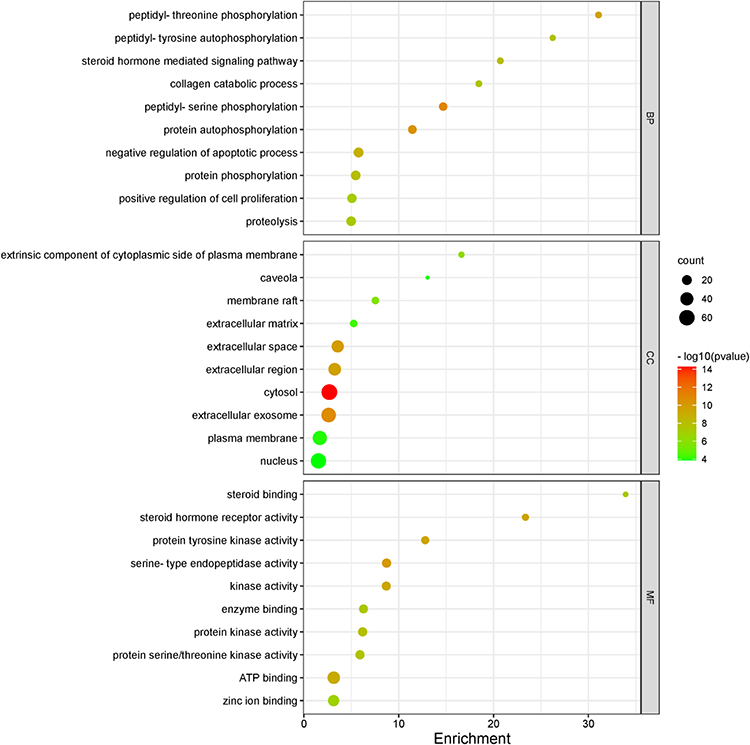 |
Figure 5 GO enrichment analysis of target genes of Aloin against GC. The number of genes enriched in each GO term is shown as the circle size and the P-value is shown by different colors. |
Then, 55 pathways associated with anti-gastric cancer effect of Aloin were identified by KEGG enrichment analysis (P<0.01 and FDR <0.01) (Figure 6, Supplementary Table 6). The following processes had the highest number of genes: Pathways in cancer (28), PI3K-AKT signaling pathway (21), Proteoglycans in cancer (16), FoxO signaling pathway (15), and Ras signaling pathway (15). The results distinctly suggested that the effect of Aloin against GC was closely related to the PI3K-AKT, FoxO and Ras signaling pathways. Among them, the PI3K-AKT signal pathway, which contained the largest number of enriched genes, may play a greater role in the treatment of GC. The “Disease target-Pathway” was drawn with the top 20 KEGG pathways (Figure 7). From the top 20 KEGG pathways, the significantly enriched genes were MAPK1, AKT2, AKT1, HRAS, MAPK8, MAPK10, MAPK14, PDPK1, EGFR, and IGF1R. This result indicated that Aloin affected multiple targets and multiple pathways in treating GC.
 |
Figure 6 KEGG enrichment analysis of target genes of Aloin against GC. The number of genes enriched in each KEGG pathway term is shown as the circle size and the P-value is shown by different colors. |
 |
Figure 7 The “Disease target-Pathway” network of Aloin targets against GC. The network is constructed by Cytoscape. The node with red color and diamond-type stands for pathway and the green color and circle stands for genes. The node size represents the size of the degree. Node size is proportional to its degree. |
Validation of the Key Targets in GEPIA, HPA and DriverDBv3 Database
The expressions of hub genes in the STAD samples were investigated in GEPIA and HPA database. The results showed that mRNA expressions of HSP90AA1 and SRC were higher in GC tissues than in normal stomach tissues (P<0.05) (Figure 8). As shown in Figure 9, the protein expressions of SRC, MAPK8 and EGFR were higher in GC tissues than in normal gastric tissues. The protein expression of CASP3 was medium in normal gastric tissues, but not detected in GC tissues. The expressions of other genes were no significant differences between GC tissues and normal tissues. The overall survival was searched in DriverDBv3 database. The results indicated that the high expressions of EGFR and ESR1 were associated with a significantly poor survival (P<0.05) (Figure 10).
 |
Figure 8 The expression of hub genes in GEPIA database. The red box plots represent tumor, and the gray box plots represent normal. |
 |
Figure 9 The protein expressions of hub genes in the HPA. The representative immunohistochemistry images of ALB, AKT1, EGFR, MAPK1, CASP3, HSP90AA1, MAPK8, SRC, HRAS and ESR1 in normal gastric tissues and GC tissues. The staining strengths are annotated as high, medium, low and not detected. The bar plots show the number of samples with different staining strengths. |
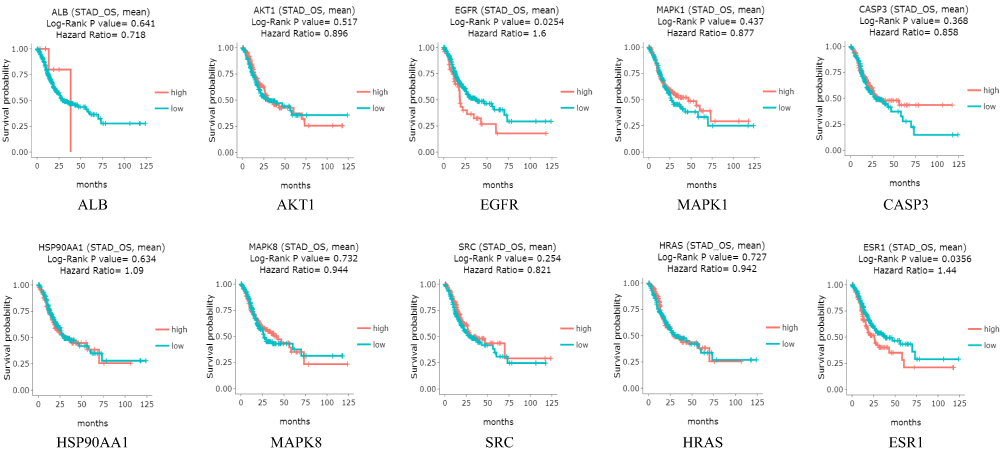 |
Figure 10 The overall survival of hub genes in DriverDBv3 database. The blue represents low expressions, and the red represents high expression. |
Aloin Inhibits Proliferation of NCI-N87 Cells
To verify the anti-proliferative effect of Aloin on GC, CCK8 was used to determine the cell viability after treatment with Aloin for 24h. As the concentrations of Aloin were gradually increased from 100 to 500 μg/mL, the survival of GC cells decreased with dose (Figure 11A), suggesting that Aloin had a significant inhibitory effect on proliferation of GC cells. Aloin inhibited the growth of NCI-N87 cells with an IC50 of 327.1μg/mL. Based on the experimental results within CCK8 assay, the concentrations of 100μg/mL, 200μg/mL and 400μg/mL were chosen for subsequent experimental program.
 |
Figure 11 Aloin inhibits NCI-N87 cell proliferation and induces cell apoptosis. (A) Inhibition of NCI-N87 cell proliferation was assessed by CCK8 assay after 24 h of treatment with the different concentrations of Aloin. **P < 0.01 versus 0μg/mL of Aloin treatment group, *P < 0.05 versus 0μg/mL of Aloin treatment group. (B) The mRNA expression of Caspase-3 was examined by qPCR. β-actin was used as an internal control. Data are expressed as mean±SEM. **P < 0.01 versus 0μg/mL of Aloin treatment group, *P < 0.05 versus 0μg/mL of Aloin treatment group. (C) The protein expression of Cleaved Caspase-3 was evaluated by automated Western immunoblotting. (D) The ultrastructure of NCI-N87 cells of each group was detected by TEM. Bar = 1μm. |
Aloin Induces NCI-N87 Cells Apoptosis
Caspase-3, which can be activated into Cleaved Caspase-3, plays a vital role in cell apoptosis. To detect the effect of Aloin on apoptosis of NCI-N87 cells, the mRNA and protein expressions of Caspase-3 were analyzed. Compared with the untreated group, the mRNA expression of Caspase-3 increased significantly in the groups of treatment with Aloin in concentrations of 200 μg/mL and 400 μg/mL, respectively (P<0.05 or P<0.01). However, there were no significant differences between treatment with 100 μg/mL of Aloin group and untreated group (P>0.05). The protein expression of Cleaved Caspase-3 underwent a similar tendency as the mRNA expression of Caspase-3 (Figure 11B and C).
The ultrastructures of NCI-N87 cells were performed via TEM. In the untreated group, the cells were full of microvilli on their membrane surface. The cell structure was intact with abundant normal organelles. Nuclear division can be seen in a number of cells. Cells in the treatment with 100 μg/mL of Aloin group were similar to the untreated group. Most of cells were in a good state. Microvilli of cells in the treatment with 200 μg/mL of Aloin group were reduced compared to the untreated cells and there were some vacuoles in the cells. Cells in the treatment with 400 μg/mL of Aloin group showed that the nuclear/cytoplasmic ratio decreased significantly and the nucleus chromatin exhibited the condensation and edge-set phenomenon. Mitochondria were swollen, and even displayed vacuolization with the cristae broke or disappeared. There were some autophagic vacuoles in the cells and microvilli were disappeared on the membrane surface. A few cells were in the late stage of apoptosis, presenting with secondary necrosis, discontinuity of the cell membrane, nuclear lysis, and disappearance of organelles (Figure 11D).
Effects of Aloin on the mRNA and Protein Expressions of Related Genes in PI3K-AKT Signaling Pathway
According to the results of the network pharmacology analysis, the PI3K-AKT signaling pathway is involved in the mechanism of Aloin against GC. Therefore, we evaluated the effect of Aloin on regulating the PI3K-AKT signaling pathway. The results showed that after the intervention of different concentrations of Aloin, the mRNA expression of PI3K decreased significantly compared with the untreated group (P< 0.01). The mRNA expression of AKT also decreased significantly in the treatment with Aloin in the concentrations of 200 μg/mL and 400 μg/mL (P< 0.05 or P< 0.01). Although the intervention of 100 μg/mL of Aloin decreased the mRNA expression of AKT, there was no remarkable difference in comparison with the untreated group (P>0.05) (Figure 12A and B). The protein expressions of p-PI3K and p-AKT exhibited a similar tendency as the mRNA levels (Figure 12C).
 |
Figure 12 Aloin inhibits the expressions of PI3K and AKT. The mRNA expressions of PI3K (A), AKT (B) were examined by qPCR. β-actin was used as an internal control. Data are expressed as mean±SEM. *P < 0.05 versus 0μg/mL of Aloin treatment group, **P < 0.01 versus 0μg/mL of Aloin treatment group. The protein expressions of PI3K, p-PI3K, AKT and p-AKT (C) were evaluated by automated Western immunoblotting. |
Discussion
The research of natural products for cancer treatment has attracted increasing attention in recent years.9 A number of studies have demonstrated that Aloin could inhibit proliferation and induce apoptosis in various kinds of tumor cells such as colorectal cancer, hepatic carcinoma, lung cancer and melanoma.2–4,10 The mechanisms of Aloin in the treatment of GC are diverse. One or two certain signaling pathways cannot reveal the multiple targets of Aloin against GC. Based on network pharmacology analysis, the complex interactions between drug and its targets and the potential mechanisms can be effectively revealed at system level.11 The present study aimed to elucidate the effects of Aloin on the GC and verify the mechanisms predicted by network pharmacology using experiment in vitro.
First, we identified 129 potential targets of Aloin against GC. Then, the PPI network was constructed and 10 hub genes were found. The core genes were associated with the pathogenesis of GC. ALB is a gene that encodes the most abundant protein in human blood. This protein functions in the regulation of blood plasma colloid osmotic pressure and acts as a carrier protein for a wide range of endogenous molecules including hormones, fatty acids, and metabolites, as well as exogenous drugs. An elevated serum ALB has been reported to be associated with a favorable prognosis for GC.12 AKT1 is a serine/threonine specific protein kinase, which plays a vital role in many biological processes such as glucose metabolism, cell proliferation and apoptosis, cell migration and so on.13,14 As an active kinase, AKT1 contributes to the progression of GC and promotes proliferation of GC cells.15 EGFR participates in regulation of cell growth and proliferation in many tumor cells. EGFR highly expresses in gastric cancer patients that acts as a poor prognostic biomarker. In this study, we found high expression of EGFR was related to a significantly poor survival in DriverDBv3 database. Interfering with the expression of EGFR can inhibit the growth of gastric cancer cells and increase the sensitivity of chemotherapy.16–18 MAPKs are involved in the regulation of cell growth and survival in physiological and pathological processes. Aberrant MAPK signaling, such as MAPK/ERK pathway, is related to the production and metastasis of cancer cells.19,20 We found MAPK8 highly expressed in GC patients, suggesting it is vital for GC progression. CASP3 is a main executor of apoptosis that must be converted into activated Caspase-3 (Cleaved caspase-3) finally to execute the program of apoptosis,21,22 which is also confirmed in our study. HSP90AA1 is an isoform of the molecular chaperone HSP90. High expression of HSP90AA1 is associated with disease progression and poor survival in patients with GC.23 The database validation showed that the expression of HSP90AA1 was higher in GC tissues than in normal tissues. SRC, identified as the first cancer-causing oncogene in 1976, plays a significant role in tumor growth and progression and its activity is associated with poor patient survival.24 In this study, we found it was highly expressed in GC tissues, indicating it may be an important biomarker in GC diagnosis and treatment. HRAS, an oncogene of the RAS family, is closely related to the proliferation, migration, invasion, and proangiogenic activity of gastric cancer cells.25 ESR1 encodes estrogen receptor α (ER α) which can interact with other signaling molecules through multiple pathways, such as PI3K/AKT or MAPK signaling pathway. The ESR1 mutation was first reported in breast cancer, and it is associated with cell proliferation, metastasis and invasion in gastric cancer.26 In this study, the ESR1 was found as a biomarker for a poor survival in GC patients. In brief, these core genes exert an important role in the occurrence, development and prognosis of GC, which can be potential targets for Aloin therapy. We have verified the role of some core genes in GC through public databases; however, some other genes still have not been verified due to many reasons such as the limited sample of the database, etc. We will do some research to further clarify the role of these genes in GC and the effect of Aloin on them.
KEGG pathway analysis revealed Aloin treatment of GC primarily involved the PI3K-AKT signaling pathway, FoxO signaling pathway and Ras signaling pathway, which were mainly associated with tumor-related disease. According to the numbers of targets corresponding to their respective pathways, we focused on the PI3K-AKT signaling pathway, which was a key pathway that may be involved in the anti-gastric cancer mechanism of Aloin. Mounting evidence has shown PI3K-AKT pathway is of crucial importance for cell proliferation, migration, apoptosis, and survival of GC.27–29 In addition, PI3K-AKT pathway is implicated in distant metastasis, chemo-resistance, and contributes to epithelial-mesenchymal transition (EMT) which occurs in drug-resistant and metastatic human cancer cells.30–33 Therefore, our study focused on the PI3K-AKT signaling pathway to explore the mechanism underlying the effects of Aloin on GC.
Meanwhile, we performed the experiment in vitro to verify the result of network pharmacology analysis and clarify the mechanisms of Aloin in the treatment of GC. Through the CCK8 assays, we found that the inhibitory effect of Aloin on the proliferation of NCI-N87 cells in a dose-dependent manner. Cell apoptosis is a programmed cell death to respond to the stimulation from internal or external factors. As a protective mechanism against cancer development, apoptosis is the key for anti-cancer treatment.34 In this study, the result showed that Aloin significantly increased mRNA level of Caspase-3 and the protein level of Cleaved Caspase-3 in NCI-N87 cells. The ultrastructures of NCI-N87 cells also showed that Aloin effectively induced apoptosis. These results revealed that Aloin had a promotion effect on apoptosis of GC cells. PI3K-AKT pathway is a negative regulator of apoptosis. Once PI3K-AKT signaling is over activated, the cell proliferation and migration will be promoted and lead to inhibition of apoptosis.35 It is reported that blocking of Akt activation obviously enhanced Aloin-induced gastric cancer cell apoptosis.5 Thus, inhibition of this pathway could be potential target for GC therapy. Then we found the Aloin significantly decreased mRNA levels of PI3K and AKT, and reduced protein expressions of p-PI3K and p-AKT, suggesting the Aloin may inhibit cell proliferation and promote apoptosis of NCI-N87 cells by inhibiting the PI3K-AKT pathway.
In summary, the results of this study elucidated the therapeutic mechanisms of Aloin against GC using network pharmacology and validated the results by pharmacological experiment in vitro. Aloin has the potential to become an effective drug for the treatment of GC, which makes new prospects for the research of natural compounds on anti-tumor treatment. Furthermore, the experiment in vivo is still needed to validate the anti-gastric cancer mechanisms of Aloin in the present study.
Conclusion
In conclusion, the present study revealed the underlying mechanisms of multi-target and multi-pathway of Aloin in the treatment of GC. Our results suggest that Aloin is a promising candidate drug for the treatment of GC.
Ethical Approval
This study is involving human data from public databases GEPIA, HPA and DriverDBv3. Due to GEPIA, HPA and DriverDBv3 belong to public databases and users can download relevant data for free for research and publish relevant articles, the ethics committee of Zhejiang Chinese Medical University confirms that this study would have had the need for ethics approval waived.
Acknowledgments
This work was supported by Zhejiang Provincial Natural Science Foundation of China (grant no. LY18H270013 and grant no. LQ17H270001), and the Science Research Fund of Academy of Traditional Chinese Medicine of Zhejiang Chinese Medical University (grant no.2020J04 and grant no.2020J03).
Disclosure
The authors declare that there are no conflicts of interest.
References
1. Ferlay J, Shin HR, Bray F, et al. Estimates of worldwide burden of cancer in 2008: GLOBOCAN 2008. Int J Cancer. 2010;127(12):2893–2917. doi:10.1002/ijc.25516
2. Pan Q, Pan H, Lou H, et al. Inhibition of the angiogenesis and growth of Aloin in human colorectal cancer in vitro and in vivo. Cancer Cell Int. 2013;13(1):69. doi:10.1186/1475-2867-13-69
3. Wan L, Zhang L, Fan K, et al. Aloin promotes A549 cell apoptosis via the reactive oxygen species-mitogen activated protein kinase signaling pathway and p53 phosphorylation. Mol Med Rep. 2017;16(5):5759–5768. doi:10.3892/mmr.2017.7379
4. Sun R, Zhai R, Ma C, et al. Combination of aloin and metformin enhances the antitumor effect by inhibiting the growth and invasion and inducing apoptosis and autophagy in hepatocellular carcinoma through PI3K/AKT/mTOR pathway. Cancer Med. 2020;9(3):1141–1151. doi:10.1002/cam4.2723
5. Tao H, Tang T, Wang S, et al. The molecular mechanisms of Aloin induce gastric cancer cells apoptosis by targeting High Mobility Group Box 1. Drug Des Devel Ther. 2019;13:1221–1231. doi:10.2147/DDDT.S201818
6. Wang Z, Tang T, Wang S, et al. Aloin inhibits the proliferation and migration of gastric cancer cells by regulating NOX2-ROS-mediated pro-survival signal pathways. Drug Des Devel Ther. 2020;14:145–155. doi:10.2147/DDDT.S219247
7. Li S, Zhang B. Traditional Chinese medicine network pharmacology: theory, methodology and application. Chin J Nat Med. 2013;11(2):110–120. doi:10.1016/S1875-5364(13)60037-0
8. Hao da C, Xiao PG. Network pharmacology: a Rosetta Stone for traditional Chinese medicine. Drug Dev Res. 2014;75(5):299–312. doi:10.1002/ddr.21214
9. Hiebl V, Ladurner A, Latkolik S, et al. Natural products as modulators of the nuclear receptors and metabolic sensors LXR, FXR and RXR. Biotechnol Adv. 2018;36(6):1657–1698. doi:10.1016/j.biotechadv.2018.03.003
10. Li P, Ren K, Liang YY, et al. Aloin promotes cell apoptosis by targeting HMGB1-TLR4-ERK axis in human melanoma cells. EXCLI J. 2020;19:641–651.
11. Berger SI, Iyengar R. Network analyses in systems pharmacology. Bioinformatics. 2009;25(19):2466–2472. doi:10.1093/bioinformatics/btp465
12. Chen J, Zhou Y, Xu Y, et al. Low pretreatment serum globulin may predict favorable prognosis for gastric cancer patients. Tumour Biol. 2016;37(3):3905–3911. doi:10.1007/s13277-015-3778-3
13. Balasuriya N, McKenna M, Liu X, et al. Phosphorylation-dependent inhibition of Akt1. Genes. 2018;9(9):450. doi:10.3390/genes9090450
14. Martínez-Rodríguez OP, Thompson-Bonilla MDR, Jaramillo-Flores ME. Association between obesity and breast cancer: molecular bases and the effect of flavonoids in signaling pathways. Crit Rev Food Sci Nutr. 2020;60(22):3770–3792. doi:10.1080/10408398.2019.1708262
15. Han Z, Wu K, Shen H, et al. Akt1/protein kinase B alpha is involved in gastric cancer progression and cell proliferation. Dig Dis Sci. 2008;53(7):1801–1810. doi:10.1007/s10620-007-9824-2
16. Gao M, Liang XJ, Zhang ZS, et al. Relationship between expression of EGFR in gastric cancer tissue and clinicopathological features. Asian Pac J Trop Med. 2013;6(4):260–264. doi:10.1016/S1995-7645(13)60054-1
17. Sigismund S, Avanzato D, Lanzetti L. Emerging functions of the EGFR in cancer. Mol Oncol. 2018;12(1):3–20. doi:10.1002/1878-0261.12155
18. Higaki E, Kuwata T, Nagatsuma AK, et al. Gene copy number gain of EGFR is a poor prognostic biomarker in gastric cancer: evaluation of 855 patients with bright-field dual in situ hybridization (DISH) method. Gastric Cancer. 2016;19(1):63–73. doi:10.1007/s10120-014-0449-9
19. Low HB, Zhang Y. Regulatory roles of MAPK phosphatases in cancer. Immune Netw. 2016;16(2):85–98. doi:10.4110/in.2016.16.2.85
20. Guo YJ, Pan WW, Liu SB, et al. ERK/MAPK signalling pathway and tumorigenesis. Exp Ther Med. 2020;19(3):1997–2007. doi:10.3892/etm.2020.8454
21. Lin B, Zhu M, Wang W, et al. Structural basis for alpha fetoprotein-mediated inhibition of caspase-3 activity in hepatocellular carcinoma cells. Int J Cancer. 2017;141(7):1413–1421. doi:10.1002/ijc.30850
22. Lakhani SA, Masud A, Kuida K, et al. Caspases 3 and 7: key mediators of mitochondrial events of apoptosis. Science. 2006;311(5762):847–851. doi:10.1126/science.1115035
23. Ba MC, Ba Z, Gong YF, et al. Knockdown of lncRNA ZNRD1-AS1 suppresses gastric cancer cell proliferation and metastasis by targeting the miR-9-5p/HSP90AA1 axis. Aging. 2021;13(13):17285–17301. doi:10.18632/aging.203209
24. Caner A, Asik E, Ozpolat B. SRC signaling in cancer and tumor microenvironment. Adv Exp Med Biol. 2021;1270:57–71. doi:10.1007/978-3-030-47189-7_4
25. Wang Y, Chu F, Lin J, et al. Erianin, the main active ingredient of Dendrobium chrysotoxum Lindl, inhibits precancerous lesions of gastric cancer (PLGC) through suppression of the HRAS-PI3K-AKT signaling pathway as revealed by network pharmacology and in vitro experimental verification. J Ethnopharmacol. 2021;279:114399. doi:10.1016/j.jep.2021.114399
26. Ur Rahman MS, Cao J. Estrogen receptors in gastric cancer: advances and perspectives. World J Gastroenterol. 2016;22(8):2475–2482. doi:10.3748/wjg.v22.i8.2475
27. Porta C, Paglino C, Mosca A. Targeting PI3K/Akt/mTOR signaling in cancer. Front Oncol. 2014;4:64. doi:10.3389/fonc.2014.00064
28. Jafari M, Ghadami E, Dadkhah T, et al. PI3k/AKT signaling pathway: erythropoiesis and beyond. J Cell Physiol. 2019;234(3):2373–2385. doi:10.1002/jcp.27262
29. Lee HJ, Venkatarame Gowda Saralamma V, Kim SM, et al. Pectolinarigenin induced cell cycle arrest, autophagy, and apoptosis in gastric cancer cell via PI3K/AKT/mTOR signaling pathway. Nutrients. 2018;10(8):1043. doi:10.3390/nu10081043
30. Le Rhun E, Bertrand N, Dumont A, et al. Identification of single nucleotide polymorphisms of the PI3K-AKT-mTOR pathway as a risk factor of central nervous system metastasis in metastatic breast cancer. Eur J Cancer. 2017;87:189–198. doi:10.1016/j.ejca.2017.10.006
31. Xu W, Yang Z, Lu N. A new role for the PI3K/Akt signaling pathway in the epithelial-mesenchymal transition. Cell Adh Migr. 2015;9(4):317–324. doi:10.1080/19336918.2015.1016686
32. Yu HG, Ai YW, Yu LL, et al. Phosphoinositide 3-kinase/Akt pathway plays an important role in chemoresistance of gastric cancer cells against etoposide and doxorubicin induced cell death. Int J Cancer. 2008;122(2):433–443. doi:10.1002/ijc.23049
33. Yu LL, Dai N, Yu HG, et al. Akt associates with nuclear factor kappaB and plays an important role in chemoresistance of gastric cancer cells. Oncol Rep. 2010;24(1):113–119. doi:10.3892/or_00000835
34. Wang H, Zhang T, Sun W, et al. Erianin induces G2/M-phase arrest, apoptosis, and autophagy via the ROS/JNK signaling pathway in human osteosarcoma cells in vitro and in vivo. Cell Death Dis. 2016;7(6):e2247. doi:10.1038/cddis.2016.138
35. Jia X, Wen Z, Sun Q, et al. Apatinib suppresses the proliferation and apoptosis of gastric cancer cells via the PI3K/Akt signaling pathway. J BUON. 2019;24(5):1985–1991.
 © 2022 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2022 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Pharmacological Mechanism of Sancao Yuyang Decoction in the Treatment of Oral Mucositis Based on Network Pharmacology and Experimental Validation
Liu Y, Ye Y, Xie G, Xu Y, Cheng M, Li C, Qu M, Zhu F
Drug Design, Development and Therapy 2023, 17:55-74
Published Date: 13 January 2023
Effect and Mechanism of Aloin in Ameliorating Chronic Prostatitis/Chronic Pelvic Pain Syndrome: Network Pharmacology and Experimental Verification
Li R, Wang Y, Lao Y, You C, Qing L, Guan X, Wang J, Li X, Li Q, Liu S, Dong Z
Drug Design, Development and Therapy 2025, 19:1945-1969
Published Date: 14 March 2025
The Mechanism of Sijunzi Decoction Suppresses Gastric Cancer Metastasis via the m6A Methyltransferase METTL14 Based on Untargeted Metabolomics Studies and Network Pharmacology Analysis
Li X, Zhao L, Wang J, Ma T, Zhou J, Bian Y, Guo J
Drug Design, Development and Therapy 2025, 19:2369-2392
Published Date: 31 March 2025
ITGA4 Contributes to 5-Fluorouracil Resistance by Up-Regulating PI3K/AKT Signaling: Evidence from Network Pharmacology, Molecular Docking and Experimental Verification

Yan S, Hu X, Wu Y, Ye W, Zhu Y, He Y, Zhan F, Wu W, Ma Z
Drug Design, Development and Therapy 2025, 19:4105-4122
Published Date: 20 May 2025
Cornus officinalis Protects Against Steroid-Induced Osteonecrosis of the Femoral Head Through Inhibiting Inflammatory Responses and Apoptosis via Network Pharmacology and Experimental Validation
Wang Y, Fan Y, Li M, Song Z, Wang P, Xie F, Miao Y, Wang Y, Zhang P, Zhan Q
Drug Design, Development and Therapy 2025, 19:5871-5898
Published Date: 9 July 2025