Back to Journals » Journal of Inflammation Research » Volume 18
Immune Mechanisms and Pain in Intervertebral Disc Degeneration
Authors Cheng H ![]() , Wang L, Huang C, Yang Z, Wu H, Fang Z
, Wang L, Huang C, Yang Z, Wu H, Fang Z ![]()
Received 12 September 2025
Accepted for publication 13 November 2025
Published 1 December 2025 Volume 2025:18 Pages 16843—16855
DOI https://doi.org/10.2147/JIR.S565510
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Shouya Feng
Haozhe Cheng, Lice Wang, Chuang Huang, Zechuan Yang, Hua Wu, Zhong Fang
Department of Orthopedics, Tongji Hospital, Tongji Medical College, Huazhong University of Science and Technology, Wuhan, 430030, People’s Republic of China
Correspondence: Hua Wu, Email [email protected] Zhong Fang, Email [email protected]
Abstract: Intervertebral disc degeneration (IVDD) is a major cause of low back pain and radicular pain, posing significant socio-economic challenges. The intervertebral disc is traditionally viewed as immune-privileged. This privilege is maintained by physical barriers and molecular factors such as Fas ligand. However, when these barriers are compromised, the nucleus pulposus (NP) can be exposed to the immune system. Recent evidence underscores a critical role for immune cells, particularly macrophages, in IVDD progression. This exposure can trigger an autoimmune response, leading to inflammation that aggravates dorsal root ganglion injury and results in hyperalgesia and pain. This review aims to autoimmunity, adaptive immunity, and the origins of pain in IVDD. We conclude that understanding these immune mechanisms is crucial, as they reveal promising avenues for future targeted immunotherapies.
Keywords: intervertebral disc degeneration, immunity, macrophages, T-lymphocytes, pain, inflammation
Introduction
Low back pain (LBP) is a pervasive global health issue and a leading cause of disability, imposing a substantial socio-economic burden worldwide.1 Intervertebral disc degeneration (IVDD) is one of the most common pathological drivers of LBP, often leading to disc herniation, spinal stenosis, and radicular symptoms.2 The management of LBP, including cases stemming from IVDD, remains a significant clinical challenge, often requiring multifaceted approaches that include physical therapy and rehabilitation.3 IVDD is a prevalent degenerative condition that profoundly impacts patients’ quality of life and functional capacity. Structurally, the intervertebral disc consists of the central nucleus pulposus, surrounded by the annulus fibrosus, and capped by cartilage endplates on both superior and inferior surfaces.4,5 The nucleus pulposus, as the central component of the disc, is embedded within a gel-like matrix composed of type II collagen and proteoglycans, playing an integral role in spinal function.
The annulus fibrosus, cartilage endplates, and ligamentous barriers protect the nucleus pulposus from immune exposure.6 Typically, these physical barriers prevent interactions between immune cells and the nucleus pulposus. However, during intervertebral disc degeneration, damage to the superior and inferior cartilage endplates, rupture of the annulus fibrosus, and exposure of the nucleus pulposus can trigger the autoimmune response.7 After nucleus pulposus herniation, the immune exposure provokes a progressive infiltration of macrophages and other immune cells, initiating an inflammatory cascade. The cartilage endplates, with no blood and lymphatic vessels, consist of a dense structure formed by high concentrations of proteoglycans and resist cellular infiltration.8–10 However, once damaged, these endplates become the foci for immune cell chemotaxis and aggregation, leading to inflammation and disc degeneration, impacting disease progression and prognosis.11
Beyond the aforementioned blood-nucleus pulposus barrier, molecular immune barriers also play a pivotal role in the immune privilege of the nucleus pulposus. The concept of a molecular immune barrier fundamentally involves the exchange of substances and the relay of signals between the NP and external factors. The NP lacks vascular, lymphatic, and neural components, yet these elements are prolific around the intervertebral disc’s outer regions, serving as the foundation for interactions between macrophages and the NP. Disruption in this communication can disturb the equilibrium of the NP’s microenvironment, facilitating the attraction and invasion of macrophages. This process indirectly fosters the development of vascular, lymphatic, and neural structures within the NP, culminating in the compromise of its immune barrier.5,12 Numerous factors influence the molecular immune barrier of the intervertebral disc. The presence of immune exemption molecules, such as FasL, helps prevent immune cell infiltration.13,14 Furthermore, systemic conditions like obesity, diabetes, and aging can compromise these immune barriers through chronic, low-grade inflammation and metabolic dysregulation.15–20
While the individual roles of inflammation and immune cells in IVDD are increasingly recognized,9,21,22 a critical gap remains in comprehensively integrating the crosstalk between innate and adaptive immunity, and linking these interactions directly to the origins of discogenic pain. Current literature often treats macrophages, T cells, and neural components in isolation, lacking a unified narrative. This review aims to fill this gap by synthesizing evidence into a cohesive framework that delineates how the breakdown of immune privilege initiates a cascade involving macrophage polarization and T helper cell responses, which collectively drive inflammation, matrix degradation, and neuro-immune sensitization. By providing this integrated perspective, we seek to identify novel mechanistic insights and underscore the translational potential of targeting specific immune pathways for the treatment of IVDD.
Structure and Immune Privilege of Intervertebral Disc
In humans, there exist 23 intervertebral discs (IVDs) between adjacent vertebrae, serving as cushions to absorb axial spinal pressures and maintain spinal height, with each section harboring nucleus pulposus cell. The nucleus pulposus cells can synthesize and secrete extracellular matrix components, thereby preserving the homeostasis of discs.23 The nucleus pulposus predominantly contains type II collagen and abundant hydration, enveloped by a gel rich in proteoglycans. The annulus fibrosus consists of 15–25 concentric cartilaginous lamellae encircling the nucleus pulposus and conferring substantial tensile resilience to the disc. The outermost annulus obtains nutrients and facilitates metabolite exchange through peripheral vasculature and capillary buds related to the vertebral bodies.24 In the early stages of human life, the nucleus is primarily populated by vacuolated notochordal cells and chondrocytes. As the disc matures, the number of notochordal cells progressively diminishes, and the tissue remains mainly composed of chondrocyte-like cells.25
Nevertheless, the axial transportation via vascular buds of the endplates remains a pivotal pathway to nourish the nucleus pulposus and annulus fibrosus.26 Oxygen and nutrients, such as glucose, diffuse from vascular buds to the cartilaginous endplates to sustain the physiological activities of the nucleus pulposus cells. This avascularity, while conferring immune privilege by limiting immune cell access, also renders the disc vulnerable to degeneration. The poor clearance of metabolic waste products and the resulting low oxygen tension (hypoxia) can accelerate cell death, decrease matrix production, and exacerbate matrix degradation, thereby contributing to the initiation and progression of IVDD.27 Except for the lack of blood vessels, there exist high concentrations of proteoglycans and elevated physical pressure in the disc microenvironment.28 Both factors inhibit vascular growth, consequently limiting immune cell infiltration.
Recent studies have unveiled that multiple molecular mechanisms maintain the immune privilege property of disc. For instance, FasL, predominantly expressed on activated T lymphocytes within immune-privileged sites, can induce the apoptosis of T lymphocytes and macrophages expressing Fas. The expression of FasL has been noted in healthy nucleus pulposus tissue, suggesting its crucial role in preserving the immune privilege property of IVD.13,29 Under typical conditions, nucleus pulposus cells emit FasL, leading to the programmed cell death of immune (like macrophages and CD8+ T cells) and vascular cells. This action curtails angiogenesis and immune cell infiltration.29,30 In the IDD rabbit model, following nucleus pulposus damage (achieved through disc puncture), there was a shift in FasL function within these cells, leading to its co-expression with Fas, which consequently triggered apoptosis in disc cells.31 Additional studies have discovered that within the intervertebral disc, the expression of Fas can be inhibited by lactoferrin (LTF), thereby ameliorating the progression of intervertebral disc degeneration.32 One of the characteristics of IDD is reduced FasL expression and FasL dysfunction, leading to an imbalance in the interaction between NP cells and immune cells. Certain regulatory factors, such as miR-155, may play a role in this process.33 However, as the microenvironment changes, HIF-1a interacts with the hypoxia response element of GAL-3 in nucleus pulposus cells, elevating GAL-3 levels and diminishing the immune barrier effect by FasL.34
Destruction of the Blood-Nucleus Pulposus Barrier Leads to Immune Exposure
The annulus fibrosus, cartilaginous endplate, and immunosuppressive molecular factors together form the blood-nucleus pulposus barrier (BNB), effectively isolating the NP from the host immune system. Damage to the BNB triggers an autoimmune response in the NP, leading to various downstream cascade reactions.35 As intervertebral discs degenerate, the distinction between the annulus fibrosus and nucleus pulposus tissues becomes challenging to discern. This evident loss of demarcation worsens with age as nucleus pulposus cells lose their gel-like morphology, becoming more fibrotic, as a common characteristic of degeneration.7,36 Another significant biochemical change during disc degeneration is the loss of proteoglycans, which are essential for maintaining osmotic resistance, withstanding compressive loads, and retaining disc hydration.37–39 These notable changes in disc behavior, such as loss of water content and disc height, profoundly affect other spinal structures, potentially compromising the physiological functions of the spine.
Immune cell infiltration primarily occurs upon the rupture of the annulus fibrosus and anterior and posterior longitudinal ligaments. The nucleus pulposus is rich in proteoglycans and type II collagen, which counteract axial loads and evenly distribute spinal pressures.40 When localized stress or shear forces exceed the load-bearing capacity of the disc, the nucleus pulposus tissue impacts surrounding structures, causing injury at the weakest points.
Furthermore, the integrity of the blood-NP barrier is also compromised by systemic metabolic and age-related factors. For instance, in obesity, adipose tissue perpetually generates pro-inflammatory cytokines (eg, leptin, TNF-α, ILs), creating a systemic inflammatory state that primes immune cells and weakens the disc’s molecular defence.15,16 Similarly, a hyperglycemic environment in diabetes can directly impair NP cell metabolism and disrupt the local immune barrier.17,18 Aging, characterized by immunosenescence and cellular senescence, alters the immune microenvironment, rendering the senescent NP highly vulnerable to vascular, neural, and lymphatic ingrowth, thereby breaching its immune privilege.19,20
Notably, the posterior aspect of the annulus fibrosus is the most vulnerable part.41,42 Johnson et al discovered that proteoglycans in the human intervertebral disc inhibit the migration of endothelial cells, partly through glycosaminoglycan side chains on the aggrecan monomer.43 Nakazawa and Yamamoto et al identified that the chronic inflammatory state of the intervertebral disc affects macrophages or macrophage-like cells, evidenced by increased expression of multiple macrophage markers associated with degeneration, especially in defective, unhealthy regions around the cartilage endplate.44,45 Given the limited self-repair capabilities of the annulus fibrosus,46,47 the acute immune infiltration following injury often fails to resolve and may instead evolve into a state of chronic inflammation. This persistent inflammatory response, characterized by continued immune cell activity and pro-inflammatory mediator release, creates a deleterious cycle that promotes extracellular matrix degradation and impedes tissue regeneration. Consequently, this self-sustaining inflammatory microenvironment is increasingly recognized as one of the core mechanisms underlying the progressive nature of intervertebral disc degeneration.48,49 The disruption of the blood nucleus pulposus tissue barrier induces autoimmunity and adaptive immunity and promotes the infiltration of immune cells, especially macrophages, into the intervertebral disc.50–52
The Role of Innate Immune Response in the Progression of IVDD
Innate immunity, or non-specific immunity, is the first line of defense against pathogens. It is a rapid response system that recognizes pathogens without previous exposure.53 The initial line of defense against infections consists of neutrophils, NK cells, monocytes/macrophages, and other innate immune cells along with chemokines and cytokines including interleukins and tumor necrosis factor.54 The adaptive immune response encompasses the entire process where T and B-lymphocytes are activated in response to foreign substances, proliferate, differentiate into effector cells, and exert various biological effects. Effector molecules of the adaptive immune system promote the response of innate immunity through positive feedback. In turn, innate immune response is an essential prerequisite for activating adaptive immunity.55,56 In the field of intervertebral disc degeneration research, Wang et al discovered certain immune cells implicated in IVDD degeneration via gene expression analysis, revealing that the infiltration of immune cells, such as Tregs and macrophages, plays a role across the spectrum of IVDD degeneration pathology.57 Ling et al conducted single-cell RNA sequencing on NP cells from patients with different stages of disc herniation and their gene set functional enrichment analysis indicated that macrophage polarization significantly influences cellular metabolism regulation.58 Research has indicated that macrophages are capable of being recruited to and infiltrating the closed nucleus pulposus within regions of the IVDD that are degenerating or have herniated. This infiltration significantly contributes to the worsening of IVDD,59,60 suggesting a pivotal role of macrophage activity in the progression of this condition (Figure 1).
 |
Figure 1 The intervertebral disc and its surrounding tissues are invaded and grown into by nerves and blood vessels, the nucleus pulposus protrudes from the damaged annulus fibrosus, and chemokines attract macrophages from the peripheral blood and tissue into the IVDD. M1 polarized macrophages have the ability to secrete pro-inflammatory molecules like TNF-α and IL-1β, which worsen IVDD. On the other hand, M2 polarized macrophages can secrete anti-inflammatory factors like IL-10 and IL-4. Intervertebral disc degeneration is caused by a vicious cycle of persistent inflammation in IVDD, which increases ECM degradation and encourages macrophages to polarize to M1 and produce additional pro-inflammatory chemicals (The MRI images of intervertebral disc degeneration are sourced from our research group’s laboratory). |
After intervertebral disc degeneration, innate immune cells, especially macrophages, release inflammatory cytokines.60 Studies have revealed that patients with disc degeneration had significantly higher amounts of basophils, eosinophils, and monocytes/macrophages in their peripheral blood.61 Notably, monocytes serve as critical markers for disease onset, progression, and resolution. They possess various pattern recognition receptors, activating pro-inflammatory factors. These cells participate directly in phagocytosis, cytokinesis, or receptor-mediated cytokinesis, and antigen presentation to T cells.62 Moreover, in the degenerated intervertebral discs, monocytes gradually differentiate into macrophages, intensifying the T-cell-mediated inflammatory response.
Macrophage Migration and Intervertebral Disc Degeneration
Macrophages exert their functional roles by migrating into the enclosed nucleus pulposus. Studies have shown that nucleus pulposus, annulus fibrosus, and endplate cells constitutively express TLR4. The expression of TLR4 is modulated by its ligands, such as LPS. Ligands binding to TLR4 initiate a cascade of signaling events upregulating pro-inflammatory mediators, including TNF-α, IL-1β, IL-6, and migration inhibitory factor (MIF), which in turn suppresses the production of extracellular matrix.63,64
MIF is a highly-conserved cytokine with multifunctional properties involved in the recruitment and migration of macrophages, leukocytes, and synovial fibroblasts. Several factors, such as TNF-α, interferons, transforming growth factors, and LPS can stimulate MIF expression by modulating TLR4 signaling.62,63 In addition to MIF, nucleus pulposus cells secrete other key chemokines that facilitate macrophage migration. For instance, TNF-α and IL-1β can induce the production of CCL3 via MAPK, NF-κB, and CCAAT-enhancer-binding protein pathways.64 CCL3 binds to its receptor CCR1, promoting macrophage migration and subsequent inflammation.65 Similarly, CCL4 is significantly upregulated in degenerated NP. Studies have shown that factors like Resistin, combined with TLR4 activation, can promote CCL4 expression via the p38-MAPK and NF-κB pathways, thereby enhancing macrophage migration.66
Macrophage Polarization During Intervertebral Disc Degeneration
Macrophages, pivotal components of the innate immune response, extensively participate in pro-inflammatory and anti-inflammatory processes and tissue remodeling through phenotypic transitions.65 In disc herniation, on the one hand, macrophages are associated with a more robust inflammatory response and more severe disc degeneration. They are also associated with the spontaneous regression of lumbar disc herniation. The high plasticity of macrophages can explain these conflicting outcomes, as they exhibit a spectrum of functional phenotypes in response to varying environmental stimuli.66,67 In response to acute injuries, macrophages swiftly shift from a pro-inflammatory phenotype (M1 type) to an anti-inflammatory phenotype (M2 type).68 The functional plasticity of macrophages is central to their role in IVDD. They can adopt a spectrum of activation states, broadly categorized into the pro-inflammatory M1 phenotype and the anti-inflammatory or pro-reparative M2 phenotype, in response to local microenvironmental cues. The balance between these phenotypes significantly influences the trajectory of disc degeneration.21
M1 macrophages release various pro-inflammatory mediators, including TNF-α and IL-1.69 TNF-α plays a central role in several intracellular processes that lead to cell death.70 TNF-α released by M1 macrophages stimulates nucleus pulposus cells to produce chemokines like CCL2 and CCL3. These chemokines, in turn, spurs IL-8 secretion. Both CCL2 and CCL3 can provoke CCR1+ and CCR2+ M1 macrophages, expediting their migration.71 The pro-inflammatory effects of TNF-α and IL-1β are largely mediated through the activation of canonical signaling pathways, most notably NF-κB and MAPK, which collectively lead to the expression of various mediators, including CCL4, and further amplify the inflammatory cascade.72,73 IL-1β, a notable pro-inflammatory agent, is predominantly produced by M1 macrophages. In M1 macrophages co-cultured with nucleus pulposus cells, HMGB1/MyD88/NFkB axis and NLRP3 pathway were activated and induced IL-1β secretion.74 Zhang et al discovered that in activated M1 macrophages, there is an upregulation of heat shock protein 90 and a downregulation of heat shock protein 70, which in turn activates the JAK2-STAT3 pathway, along with the NF-kB and MAPK pathways, leading to a series of inflammatory responses.75 In intervertebral disc degradation, the presence of M1 macrophages signifies a shift in the cellular milieu. Yang et al highlighted that in intervertebral disc degeneration, the α, β, and δ isoforms of p38 MAPK are mainly expressed. NP cells expressing p38 affect macrophage polarization by producing GM-CSF and IFNγ. Of these, either P38a or p38b provokes GM-CSF and IFNG secretion, shifting macrophages toward M1 polarization.76
M2 macrophages, known for their anti-inflammatory properties, functionally counteract M1 macrophages. M2 macrophages can phagocytose and remove tissue debris and apoptotic cells and they can suppress the apoptosis and aging of nucleus pulposus cells induced by TNF-α.77,78 IL-4 and IL-13 can prompt macrophages to polarize toward the M2 type.21 M2 macrophages participate in Th2 response, mitigate inflammatory reactions and promote tissue remodeling and repair, angiogenesis, and tumorigenesis.79 Li et al investigated the effects of conditioned medium from M2 macrophages on nucleus pulposus cells, identifying 975 upregulated genes and 930 downregulated genes. Among these, the most significant KEGG pathway was the cell cycle, which mitigated the progression of intervertebral disc degeneration.80 Liu et al demonstrated in vitro the promotive effect of M2c macrophages on the anabolic metabolism of nucleus pulposus cells and found that M2c-derived exosomes improved ECM metabolic imbalance in IVDD through the miR-124/CILP/TGF-β regulatory axis.81 Yang et al found that following DNMT1 silencing or overexpression of SIRT6, there was an overexpression of M2 macrophage-specific markers CD163, Arg-1, and MR, which provided protective effects on the nucleus pulposus.73 Long et al used siRNA to inhibit the transcription of CHI3L1, confirming that the CHI3L1 protein mediates the effects of M2a macrophages on NP cells.82 Collectively, these functional attributes suggest that the M2 phenotype plays a protective and reparative role in IVDD. Consequently, we interpret these findings as a strong rationale for exploring therapeutic strategies that promote M2 polarization. When macrophages are stimulated by Th2-derived cytokines IL-4 and IL-13, they activate cell membrane surface receptor IL-4 receptor alpha (IL-4Rα) and downstream JAK/STAT6 and PI3K/Akt signals, leading to M2 macrophage polarization.83 Besides IL-4 and IL-13, several cytokines, such as IL-10, participate in M2 polarization by activating IL-10R/STAT3 pathway.84 In addition, IL-33, which is produced by Th2 cells, can also induce M2 polarization.85,86 Furthermore, JAK/STAT is a crucial regulatory pathway inducing macrophage polarization toward M1 and M2.87 STAT6 and STAT1 counteract each other. STAT1 and Th1-mediated immune responses are associated with M1 polarization, whereas STAT6 and Th2 cell-mediated immune responses are related to M2 polarization.88 STAT6 plays a crucial role in mediating the effects of IL-4 and IL-13 in the M2 polarization of macrophages. IL-4 and IL-13 activate JAK1 and JAK3 through IL-4 receptor alpha (IL-4Rα), resulting in the activation and translocation of STAT6, subsequently polarizing macrophages to the M2 type.89 A summary of the key characteristics and roles of M1 and M2 macrophages in IVDD, with supporting references, is provided in Table 1.
 |
Table 1 Summary of Macrophage Phenotypes in Intervertebral Disc Degeneration |
Collectively, these studies suggest that the dynamic balance between M1 and M2 macrophage polarization is a critical determinant in IVDD progression. M1 macrophages drive inflammation and tissue catabolism, while M2 macrophages promote resolution of inflammation and tissue repair. The prevailing polarization state is regulated by a complex network of cytokines and signaling pathways, notably the JAK/STAT axis. Therefore, understanding and therapeutically modulating this balance represents a promising strategy for mitigating IVDD. However, there are still many aspects of the molecular mechanisms of macrophage polarization that need to be explored. Particularly, the effects of different subtypes of Akt and various signaling pathways in macrophage polarization deserve further studies.
The Role of Adaptive Immune Response in the Progression of IVDD
The adaptive immune response, mediated by T and B lymphocytes, is activated upon exposure to NP antigens and plays a decisive role in chronic inflammation and IVDD progression. The interplay between different T helper (Th) cell subsets and their cytokine networks is critical. An imbalance skewed towards pro-inflammatory responses exacerbates disc degeneration, while regulatory or anti-inflammatory responses may confer protection.57,92
After activation by NP tissue, T lymphocytes differentiate into subgroups, such as Th1, Th2, and Th17, each with unique interactions and cytokine secretion.93 Th1 and Th2 cells represent counterbalancing forces in IVDD immunity. Th1 cells typically produce pro-inflammatory cytokines (eg, IFN-γ, IL-2, IL-12) that activate macrophages and promote cell-mediated immunity.10,93–95 In contrast, Th2 cells secrete cytokines like IL-4, IL-5, IL-10, and IL-13, which stimulate humoral immunity and generally suppress cellular inflammation.96–100 Critically, in IVDD, this balance is disrupted. The interaction with macrophages surrounding a protruded disc amplifies the Th1 response while inhibiting the Th2 response, leading to a net pro-inflammatory state that fuels degeneration and pain. This Th1-skewed response is further mediated by immunoglobulin antibodies produced by plasma cells, which activate complement pathways and accumulate inflammatory cells at the site of injury.101
Th17 cells are the major source of pro-inflammatory cytokines, including IL-17A, TNF-α, IL-22, and IL-26.102,103 In particular, increased IL-17A levels have been noted in degenerated and herniated IVD tissues. Activated by IL-23, Th17 lymphocytes primarily generate IL-17, which exacerbates inflammation through various pathways. Some of the aforementioned pathways are influenced by products such as IFN-γ.104 This interaction stimulates the production of other cytokines, proteases, nitric oxide (NO), and prostaglandin E2 (PGE2).105 IL-17A upregulates nitric oxide synthase and cyclooxygenase-2, enhances NO, PGE2, and IL-6 production.105 Significant populations of CD4+ IL-17A+ and CD4+ CCR6+ cells have been found in IVD. Pene et al have suggested that CD4+ CCR6+ might be a characteristic cellular marker of IL-17-secreting cells.106 As disc degeneration progresses, substantial amounts of CCL20 are produced, attracting peripheral blood Th17 cells expressing CCR6.107 After chemotactic migration to the lesion site, Th17 cells secrete multiple inflammatory cytokines, forming an inflammatory milieu.103 IL-17A can also activate the JAK/STAT and PI3K/AKT pathways.108,109 IL-17A upregulates VEGF expression in NPs via the JAK/STAT pathway, promoting angiogenesis in disc lesions and mediating immune and inflammatory responses.110 Conversely, IL-17A inhibits autophagy in NP cells through PI3K activation. Autophagy is a protective mechanism in IVDD; thus, activating IL-17A/PI3K/AKT pathway, which inhibits autophagy, might exacerbate disc degeneration.111 The key features and functions of the major T helper cell subsets discussed in this section, with supporting references, are summarized in Table 2.
 |
Table 2 Summary of T Helper Cell Subsets in Intervertebral Disc Degeneration |
The Role of Neural-Immune Interactions in the Pain-Producing Process
Inflammation, immune reactions, mechanical stress, and hypoxia trigger low back and leg pain by damaging spinal tissues. And surgical procedures, such as discectomy or spinal fusion, inevitably cause tissue trauma, which can instigate a pronounced adjacent inflammatory response.120,121 This exposure triggers a robust local immune response, characterized by the infiltration and activation of macrophages and T cells, which in turn release a cascade of pro-inflammatory cytokines such as TNF-α, IL-1β, IL-6, and IL-17.60,122,123
These cytokines play a dual role in the genesis of pain. First, they directly and indirectly sensitize sensory neurons within the dorsal root ganglia (DRG), leading to hyperalgesia.90,91 Second, they create a chemotactic and permissive microenvironment that promotes the ingrowth of sensory nerve fibers from the DRG into the normally aneural regions of the annulus fibrosus and even the nucleus pulposus.124 Key factors in this neurotrophic process include nerve growth factor (NGF) and brain-derived neurotrophic factor (BDNF), whose levels are elevated in degeneration.125–127
Consequently, the initially immune-privileged disc is transformed into a pain generator. The confluence of persistent inflammation (from immune cells), neuronal sensitization (in the DRG), and aberrant innervation (within the disc) establishes a self-sustaining cycle of neuro-immune interaction that underlies chronic discogenic pain.90,128,129
This neuro-immune cascade is further amplified at the systemic level. Following nerve root injury or compression, activated macrophages and T lymphocytes migrate to the affected DRG.130,131 Here, macrophage-derived TNF-α augments neurogenesis and intensifies DRG damage, leading to spinal pain hypersensitivity.132 Similarly, products from autoimmune lymphocytes, such as IFN-γ and IL-17, enhance the production of inflammatory mediators like nitric oxide (NO) and prostaglandin E2 (PGE2) in the disc, further fueling the inflammatory milieu.123,133,134 Beyond the intrinsic pathophysiology of IVDD, iatrogenic factors can significantly modulate the local immune environment. Surgical procedures, such as discectomy or spinal fusion, inevitably cause tissue trauma, which can instigate a pronounced adjacent inflammatory response.121,135,136 This postsurgical inflammation involves the release of pro-inflammatory cytokines and chemokines, which can further sensitize dorsal root ganglia, promote neurite sprouting, and potentially contribute to persistent postoperative pain. This underscores that the neuro-immune axis described herein can be activated not only by degenerative processes but also by therapeutic interventions, highlighting the need for perioperative anti-inflammatory strategies. An overview of these processes, highlighting the key interactions and pathways involved in disc-related pain, is depicted in Figure 2.
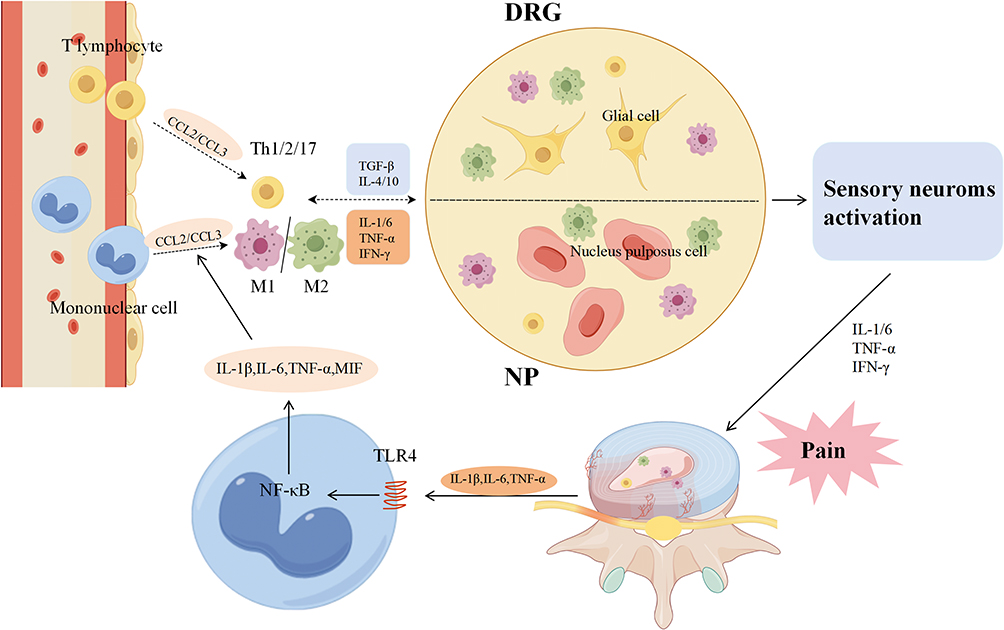 |
Figure 2 The figure illustrates how intervertebral disc-related pain is produced by the immune cascade. Upon breaching the immune barrier and being identified by the immune system, the projecting nucleus pulposus triggers the activation and release of immune cells (CCL2/CCL3) into the bloodstream, which are then gathered at the NP and DRG. Immune cells in NP and DRG tissues were stimulated concurrently by the release of inflammatory mediators (TNF-α, IFN-γ, IL-1/6, etc) and inhibitory mediators (TGF-β, IL-4/10, etc). Ultimately, inflammatory cytokines (IL-1/6, TNF-α, IFN-γ, etc) are concurrently released by two distinct types of immune cells, activate sensory neurons, and cause pain. |
Conclusion
The rising prevalence of IVDD necessitates a shift from viewing it as a passive mechanical process to recognizing it as an active, immune-mediated disorder. We propose that IVDD pathogenesis and discogenic pain are driven by a sustained dysregulation between innate and adaptive immunity, characterized by an imbalance in M1/M2 macrophage polarization and a skewed T-cell response favoring Th1/Th17 activity. This review identifies key therapeutic targets across this immune network, spanning chemokine axes (eg, CCL3/CCR1), signaling hubs (eg, JAK/STAT, NF-κB), and effector cytokines (eg, IL-17A, TNF-α). Consequently, the most promising therapeutic strategy appears to be the precise recalibration of this immune response, rather than broad immunosuppression. It is important to acknowledge the limitations of the current evidence. Our understanding relies heavily on data from animal models, and key mechanistic insights await validation in human systems. The scarcity of clinical data directly linking these immune profiles to patient outcomes remains a significant gap. However, future efforts aimed at therapeutically targeting this defined immune circuitry may hold the key to achieving the dual goals of halting degeneration and alleviating pain.
Literature Search Strategy
A comprehensive literature search was conducted to identify relevant articles for this narrative review. Electronic databases including PubMed and Web of Science were queried for publications from January 2000 to September 2025. The search utilized a combination of the following key terms and their variants: (“intervertebral disc degeneration” OR “IVDD”) AND (“immune response” OR “inflammation” OR “macrophage” OR “T cell” OR “autoimmunity”) AND (“pain” OR “neuroinflammation” OR “dorsal root ganglion”) AND (“immunotherapy” OR “treatment”). The selection prioritized original research articles and high-impact reviews in English, encompassing both human and animal studies to provide a comprehensive mechanistic overview.
Data Sharing Statement
Data sharing is not applicable to this article as no data were created or analysed in this study.
Acknowledgments
This work was financially supported by the National Natural Science Foundation of China (2022CFB924).
Author Contributions
H.C.: Writing – original draft, conceptualization and visualization. L.W.: Writing – original draft and data curation. C.H.: Writing – original draft and investigation. Z.Y: Writing – original draft and visualization. H.W.: Writing – original draft, supervision, methodology, writing – review and editing and project administration. Z.F.: Writing – original draft, supervision, conceptualization, writing – review and editing and fund acquisition. All authors gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Disclosure
The authors declare that they have no known financial or interpersonal conflict that might have appeared to influence the research presented in this paper.
References
1. Knezevic NN, Candido KD, Vlaeyen JWS, Van Zundert J, Cohen SP. Low back pain. Lancet. 2021;398:78–92. doi:10.1016/S0140-6736(21)00733-9
2. Kocaman H, Yıldırım H, Gökşen A, Arman GM. An investigation of machine learning algorithms for prediction of lumbar disc herniation. Med Biol Eng Comput. 2023;61:2785–2795. doi:10.1007/s11517-023-02888-x
3. Kuzu Ş, Canli M, Valamur İ, Özüdoğru A, Alkan H, Hartavi A. Effects of aerobic exercise in addition to core stabilization exercises on functional capacity, physical performance and fall risk in geriatric individuals with chronic non-specific low back pain. BMC Sports Sci Med Rehabil. 2025;17:218. doi:10.1186/s13102-025-01271-7
4. Chen X, Wang Z, Deng R, Yan H, Liu X, Kang R. Intervertebral disc degeneration and inflammatory microenvironment: expression, pathology, and therapeutic strategies. Inflamm Res. 2023;72:1811–1828. doi:10.1007/s00011-023-01784-2
5. Gao B, Jiang B, Xing W, Xie Z, Luo Z, Zou W. Discovery and application of postnatal nucleus pulposus progenitors essential for intervertebral disc homeostasis and degeneration. Adv Sci. 2022;9:e2104888. doi:10.1002/advs.202104888
6. Zhou T, Chen Y, Liao Z, et al. Spatiotemporal characterization of human early intervertebral disc formation at single‐cell resolution. Adv Sci. 2023;10:e2206296. doi:10.1002/advs.202206296
7. Roberts S, Evans H, Trivedi J, Menage J. Histology and Pathology of the Human Intervertebral Disc. J Bone Joint Surg Am. 2006;88:10–14. doi:10.2106/JBJS.F.00019
8. Koroth J, Buko EO, Abbott R, et al. Macrophages and intervertebral disc degeneration. IJMS. 2023;24:1367. doi:10.3390/ijms24021367
9. Ochi M, Wan B, Bao Q, et al. Contribution of immune cells to intervertebral disc degeneration and the potential of immunotherapy. Connective Tissue Res. 2023;64:413–427. doi:10.1080/03008207.2023.2212051
10. Barter MJ, Gomez R, Hyatt S, et al. A new immunometabolic perspective of intervertebral disc degeneration. Nat Rev Rheumatol. 2022;18:47–60. doi:10.1038/s41584-021-00713-z
11. Feng P, Che Y, Gao C, Zhu L, Gao J, Vo NV. Immune exposure: how macrophages interact with the nucleus pulposus. Front Immunol. 2023;14:1155746. doi:10.3389/fimmu.2023.1155746
12. Tu J, Li W, Yang S, et al. Single‐cell transcriptome profiling reveals multicellular ecosystem of nucleus pulposus during degeneration progression. Adv Sci. 2022;9:e2103631. doi:10.1002/advs.202103631
13. Liu Z-H, Sun Z, Wang H-Q, et al. FasL expression on human nucleus pulposus cells contributes to the immune privilege of intervertebral disc by interacting with immunocytes. Int J Med Sci. 2013;10:1053–1060. doi:10.7150/ijms.6223
14. Sun Z, Liu Z-H, Chen Y, et al. Molecular immunotherapy might shed a light on the treatment strategies for disc degeneration and herniation. Med Hypotheses. 2013;81:477–480. doi:10.1016/j.mehy.2013.06.014
15. Zhou J, Mi J, Peng Y, Han H, Liu Z. Causal associations of obesity with the intervertebral degeneration, low back pain, and sciatica: a two-sample mendelian randomization study. Front Endocrinol. 2021;12:740200. doi:10.3389/fendo.2021.740200
16. Tseng C, Han Y, Lv Z, et al. Glucose-stimulated PGC-1α couples with CBP and Runx2 to mediate intervertebral disc degeneration through transactivation of ADAMTS4/5 in diet-induced obesity mice. Bone. 2023;167:116617. doi:10.1016/j.bone.2022.116617
17. Jin P, Xing Y, Xiao B, et al. Diabetes and intervertebral disc degeneration: a Mendelian randomization study. Front Endocrinol. 2023;14:1100874. doi:10.3389/fendo.2023.1100874
18. Cannata F, Vadalà G, Ambrosio L, et al. Intervertebral disc degeneration: a focus on obesity and type 2 diabetes. Diabetes Metabolism Res. 2020;36:e3224. doi:10.1002/dmrr.3224
19. Yousefzadeh MJ, Flores RR, Zhu Y, et al. An aged immune system drives senescence and ageing of solid organs. Nature. 2021;594:100–105. doi:10.1038/s41586-021-03547-7
20. Saberi M, Zhang X, Mobasheri A. CB2R attenuates intervertebral disc degeneration by delaying nucleus pulposus cell senescence through AMPK/GSK3β pathway. Aging Dis. 2022;13:552. doi:10.1007/s11357-021-00341-1
21. Witherel CE, Sao K, Brisson BK, et al. Regulation of extracellular matrix assembly and structure by hybrid M1/M2 macrophages. Biomaterials. 2021;269:120667. doi:10.1016/j.biomaterials.2021.120667
22. Ji X, Shao H, Li X, et al. Injectable immunomodulation-based porous chitosan microspheres/HPCH hydrogel composites as a controlled drug delivery system for osteochondral regeneration. Acta Biomater. 2022;285:121530. doi:10.1016/j.biomaterials.2022.121530
23. Lee J, Lee CY, Park J-H, et al. Mechanical aspects of intervertebral disc injury and implications on biomechanics. Spine. 2020;45:E457–E464. doi:10.1097/BRS.0000000000003291
24. Zhang A, Cheng Z, Chen Y, Shi P, Gan W, Zhang Y. Emerging tissue engineering strategies for annulus fibrosus therapy. Acta Biomater. 2023;167:1–15. doi:10.1016/j.actbio.2023.06.012
25. Harfe BD. Intervertebral disc repair and regeneration: insights from the notochord. Semin Cell Dev Biol. 2022;127:3–9. doi:10.1016/j.semcdb.2021.11.012
26. Ding S-L, Zhang T-W, Zhang Q-C, et al. Excessive mechanical strain accelerates intervertebral disc degeneration by disrupting intrinsic circadian rhythm. Exp Mol Med. 2021;53:1911–1923. doi:10.1038/s12276-021-00716-6
27. Li G, Zhang W, Liang H, Yang C. Epigenetic regulation in intervertebral disc degeneration. Trends Mol Med. 2022;28:803–805. doi:10.1016/j.molmed.2022.07.007
28. Hee HT, Chuah YJ, Tan BHM, Setiobudi T, Wong HK. Vascularization and morphological changes of the endplate after axial compression and distraction of the intervertebral disc. Spine. 2011;36:505–511. doi:10.1097/BRS.0b013e3181d32410
29. Sun Z, Wan Z-Y, Guo Y-S, Wang H-Q, Luo Z-J. FasL on human nucleus pulposus cells prevents angiogenesis in the disc by inducing Fas-mediated apoptosis of vascular endothelial cells. Int J Clin Exp Pathol. 2013;6:2376–2385.
30. Zeng Y, Danielson KG, Albert TJ, et al. Fas ligand exists on intervertebral disc cells: a potential molecular mechanism for immune privilege of the disc. Spine. 2002;27:1526–1530. doi:10.1097/00007632-200207150-00009
31. Wang J, Tang T, Yang H, et al. The expression of Fas ligand on normal and stabbed-disc cells in a rabbit model of intervertebral disc degeneration: a possible pathogenesis. SPI. 2007;6:425–430. doi:10.3171/spi.2007.6.5.425
32. Zhang X-B, Xu S-Q, Hui Y-G, et al. Lactotransferrin promotes intervertebral disc degeneration by regulating Fas and inhibiting human nucleus pulposus cell apoptosis. Aging. 2022;14:4572–4585. doi:10.18632/aging.204100
33. Ma C-J, Liu X, Che L, Liu Z-H, Samartzis D, Wang H-Q. Stem cell therapies for intervertebral disc degeneration: immune privilege reinforcement by Fas/FasL regulating machinery. CSCR. 2015;10:285–295. doi:10.2174/1574888X10666150416114027
34. Zeng Y, Danielson KG, Albert TJ, Shapiro IM, Risbud MV. HIF-1α is a regulator of Galectin-3 expression in the intervertebral disc. J Bone Mineral Res. 2007;22:1851–1861. doi:10.1359/jbmr.070620
35. Sun Z, Liu B, Luo Z-J. The immune privilege of the intervertebral disc: implications for intervertebral disc degeneration treatment. Int J Med Sci. 2020;17:685–692. doi:10.7150/ijms.42238
36. Zheng H-D, Sun Y-L, Kong D-W, et al. Deep learning-based high-accuracy quantitation for lumbar intervertebral disc degeneration from MRI. Nat Commun. 2022;13:841. doi:10.1038/s41467-022-28387-5
37. Ferreira J, Teixeira G, Neto E, et al. IL-1β-pre-conditioned mesenchymal stem/stromal cells’ secretome modulates the inflammatory response and aggrecan deposition in intervertebral disc. eCM. 2021;41:431–543. doi:10.22203/eCM.v041a28
38. Watanabe H. Aggrecan and versican: two brothers close or apart. Am J Physiol Cell Physiol. 2022;322:C967–C976. doi:10.1152/ajpcell.00081.2022
39. Zheng J, Wang J, Liu H, et al. Alarmins S100A8/A9 promote intervertebral disc degeneration and inflammation-related pain in a rat model through toll-like receptor-4 and activation of the NF-κB signaling pathway. Osteoarthritis Cartilage. 2022;30:998–1011. doi:10.1016/j.joca.2022.03.011
40. Xing H, Zhang Z, Mao Q, et al. Injectable exosome-functionalized extracellular matrix hydrogel for metabolism balance and pyroptosis regulation in intervertebral disc degeneration. J Nanobiotechnol. 2021;19:264. doi:10.1186/s12951-021-00991-5
41. Yang R, Xu W, Zheng H, et al. Involvement of oxidative stress‐induced annulus fibrosus cell and nucleus pulposus cell ferroptosis in intervertebral disc degeneration pathogenesis. J Cell Physiol. 2021;236:2725–2739. doi:10.1002/jcp.30039
42. Yu Q, Han F, Yuan Z, et al. Fucoidan-loaded nanofibrous scaffolds promote annulus fibrosus repair by ameliorating the inflammatory and oxidative microenvironments in degenerative intervertebral discs. Acta Biomater. 2022;148:73–89. doi:10.1016/j.actbio.2022.05.054
43. Johnson WEB, Caterson B, Eisenstein SM, Roberts S. Human intervertebral disc aggrecan inhibits endothelial cell adhesion and cell migration in vitro. Spine. 2005;30:1139–1147. doi:10.1097/01.brs.0000162624.95262.73
44. Nakazawa KR, Walter BA, Laudier DM, et al. Accumulation and localization of macrophage phenotypes with human intervertebral disc degeneration. Spine J. 2018;18:343–356. doi:10.1016/j.spinee.2017.09.018
45. Yamamoto Y, Kokubo Y, Nakajima H, Honjoh K, Watanabe S, Matsumine A. Distribution and Polarization of Hematogenous Macrophages Associated with the Progression of Intervertebral Disc Degeneration. Spine. 2021;47:E149–E158. doi:10.1097/BRS.0000000000004222
46. Peredo AP, Gullbrand SE, Smith HE, Mauck RL. Putting the pieces in place: mobilizing cellular players to improve annulus fibrosus repair. Tissue Eng Part B Rev. 2021;27:295–312. doi:10.1089/ten.teb.2020.0196
47. Tavakoli J, Diwan AD, Tipper JL. Advanced strategies for the regeneration of lumbar disc annulus fibrosus. IJMS. 2020;21:4889. doi:10.3390/ijms21144889
48. Yurube T, Takada T, Suzuki T, et al. The role of the immune microenvironment in bone, cartilage, and soft tissue regeneration: from mechanism to therapeutic opportunity. Military Med Res. 2022;9:65. doi:10.1186/s40779-022-00426-8
49. Salahi A, Rane A, Xiao L, et al. Skeletal infections: microbial pathogenesis, immunity and clinical management. Nat Rev Microbiol. 2022;20:385–400. doi:10.1038/s41579-022-00686-0
50. Fan C, Wang W, Yu Z, et al. M1 macrophage-derived exosomes promote intervertebral disc degeneration by enhancing nucleus pulposus cell senescence through LCN2/NF-κB signaling axis. J Nanobiotechnol. 2024;22:301. doi:10.1186/s12951-024-02556-8
51. Dou Y, Zhang Y, Liu Y, et al. Role of macrophage in intervertebral disc degeneration. Bone Res. 2025;13:15. doi:10.1038/s41413-024-00397-7
52. Liu X-W, Xu H-W, Yi -Y-Y, Zhang S-B, Wang S-J. Role of ferroptosis and immune infiltration in intervertebral disc degeneration: novel insights from bioinformatics analyses. Front Cell Dev Biol. 2023;11:1170758. doi:10.3389/fcell.2023.1170758
53. Sun L, Wang X, Saredy J, Yuan Z, Yang X, Wang H. Innate-adaptive immunity interplay and redox regulation in immune response. Redox Biol. 2020;37:101759. doi:10.1016/j.redox.2020.101759
54. Bekkering S, Domínguez-Andrés J, Joosten LAB, Riksen NP, Netea MG. Trained immunity: reprogramming innate immunity in health and disease. Annu Rev Immunol. 2021;39:667–693. doi:10.1146/annurev-immunol-102119-073855
55. Cao Z, Wang H, Chen J, et al. Trained immunity — basic concepts and contributions to immunopathology. Nat Rev Nephrol. 2023;19:23–37. doi:10.1038/s41581-022-00633-5
56. Zhang Q, Cao X. Epigenetic remodeling in innate immunity and inflammation. Annu Rev Immunol. 2021;39:279–311. doi:10.1146/annurev-immunol-093019-123619
57. Wang L, He T, Liu J, et al. Revealing the immune infiltration landscape and identifying diagnostic biomarkers for lumbar disc herniation. Front Immunol. 2021;12:666355. doi:10.3389/fimmu.2021.666355
58. Ling Z, Liu Y, Wang Z, et al. Single-cell RNA-Seq analysis reveals macrophage involved in the progression of human intervertebral disc degeneration. Front Cell Dev Biol. 2021;9:833420. doi:10.3389/fcell.2021.833420
59. Hu X, Wang Z, Zhang H, et al. Single-cell sequencing: new insights for intervertebral disc degeneration. Biomed Pharmacother. 2023;165:115224. doi:10.1016/j.biopha.2023.115224
60. Risbud MV, Shapiro IM. Role of cytokines in intervertebral disc degeneration: pain and disc content. Nat Rev Rheumatol. 2014;10:44–56. doi:10.1038/nrrheum.2013.160
61. Li G, Yun X, Ye K, et al. Chemokine profile of herniated intervertebral discs infiltrated with monocytes and macrophages. Spine. 2002;27:1511–1516. doi:10.1097/00007632-200207150-00006
62. Shao Z, Wang B, Shi Y, et al. Monocyte conversion during inflammation and injury. ATVB. 2017;37:35–42. doi:10.1161/ATVBAHA.116.308198
63. Zhang Q, Weng Y, Jiang Y, Zhao S, Zhou D, Xu N. Overexpression of miR-140-5p inhibits lipopolysaccharide-induced human intervertebral disc inflammation and degeneration by downregulating toll-like receptor 4. Front Bioeng Biotechnol. 2018;40:793–802. doi:10.3892/or.2018.6488
64. Rajan NE, Bloom O, Maidhof R, et al. Toll-Like Receptor 4 (TLR4) expression and stimulation in a model of intervertebral disc inflammation and degeneration. Spine. 2013;38:1343–1351. doi:10.1097/BRS.0b013e31826b71f4
65. Silva AJ, Ferreira JR, Cunha C, et al. Macrophages down-regulate gene expression of intervertebral disc degenerative markers under a pro-inflammatory microenvironment. Front Immunol. 2019;10:1508. doi:10.3389/fimmu.2019.01508
66. Ohba T, Haro H. TWEAK and TSLP in disc degeneration and spontaneous hernia resorption. JOR Spine. 2020;3:e1068. doi:10.1002/jsp2.1068
67. Kühl B, Beyerbach M, Baumgärtner W, Gerhauser I. Characterization of microglia/macrophage phenotypes in the spinal cord following intervertebral disc herniation. Front Vet Sci. 2022;9:942967. doi:10.3389/fvets.2022.942967
68. Vizcaíno Revés N, Mogel HM, Stoffel M, Summerfield A, Forterre F. Polarization of macrophages in epidural inflammation induced by canine intervertebral disc herniation. Front Vet Sci. 2020;7:32. doi:10.3389/fvets.2020.00032
69. Nakao Y, Fukuda T, Zhang Q, et al. Exosomes from TNF-α-treated human gingiva-derived MSCs enhance M2 macrophage polarization and inhibit periodontal bone loss. Acta Biomater. 2021;122:306–324. doi:10.1016/j.actbio.2020.12.046
70. Jang D, Lee A-H, Shin H-Y, et al. The role of Tumor Necrosis Factor Alpha (TNF-α) in autoimmune disease and current TNF-α inhibitors in therapeutics. IJMS. 2021;22:2719. doi:10.3390/ijms22052719
71. Nakawaki M, Uchida K, Miyagi M, et al. Sequential CCL2 expression profile after disc injury in mice. J Orthop Res. 2020;38:895–901. doi:10.1002/jor.24522
72. Li Z, Wang X, Pan H, et al. Resistin promotes CCL4 expression through toll-like receptor-4 and activation of the p38-MAPK and NF-κB signaling pathways: implications for intervertebral disc degeneration. Osteoarthritis Cartilage. 2017;25:341–350. doi:10.1016/j.joca.2016.10.002
73. Pinkosky SL, Filippov S, Srivastava RAK, et al. DNMT1 regulates polarization of macrophage-induced intervertebral disc degeneration by modulating SIRT6 expression and promoting pyroptosis in vivo. Aging. 2023;15:4288–4303. doi:10.18632/aging.204729
74. Zhao F, Guo Z, Hou F, Fan W, Wu B, Qian Z. Magnoflorine alleviates “M1” polarized macrophage-induced intervertebral disc degeneration through repressing the HMGB1/Myd88/NF-κB pathway and NLRP3 inflammasome. Front Pharmacol. 2021;12:701087. doi:10.3389/fphar.2021.701087
75. Zhang S, Wang P, Hu B, et al. HSP90 inhibitor 17-AAG attenuates nucleus pulposus inflammation and catabolism induced by M1-polarized macrophages. Front Cell Dev Biol. 2021;9:796974. doi:10.3389/fcell.2021.796974
76. Mi B, Xiong W, Xu N, et al. Differential expression of p38 MAPK α, β, γ, δ isoforms in nucleus pulposus modulates macrophage polarization in intervertebral disc degeneration. Sci Rep. 2016;6:22182. doi:10.1038/srep22182
77. Lu Y-F, Liu Y, Fu W-M, et al. Macrophages during the fibrotic process: M2 as friend and foe. Front Immunol. 2017;6:602. doi:10.3389/fimmu.2015.00602
78. Li X-C, Luo S-J, Fan W, Zhou T-L, Huang C-M, Wang M-S. M2 macrophage-conditioned medium inhibits intervertebral disc degeneration in a tumor necrosis factor-α-rich environment. J Orthop Res. 2022;40:2488–2501. doi:10.1002/jor.25292
79. Song G, Ouyang G, Bao S. The activation of Akt/PKB signaling pathway and cell survival. J Cell Mol Med. 2005;9:59–71. doi:10.1111/j.1582-4934.2005.tb00337.x
80. Li X-C, Luo S-J, Fan W, et al. Macrophage polarization regulates intervertebral disc degeneration by modulating cell proliferation, inflammation mediator secretion, and extracellular matrix metabolism. Front Immunol. 2022;13:922173. doi:10.3389/fimmu.2022.922173
81. Liu Y, Xue M, Han Y, et al. Exosomes from M2c macrophages alleviate intervertebral disc degeneration by promoting synthesis of the extracellular matrix via MiR‐124/CILP/TGF‐β. Bioeng Transl Med. 2023;8:e10500. doi:10.1002/btm2.10500
82. Li L, Wei K, Ding Y, et al. M2a macrophage-secreted CHI3L1 promotes extracellular matrix metabolic imbalances via activation of IL-13Rα2/MAPK pathway in rat intervertebral disc degeneration. Front Immunol. 2021;12:666361. doi:10.3389/fimmu.2021.666361
83. Byles V, Covarrubias AJ, Ben-Sahra I, et al. The TSC-mTOR pathway regulates macrophage polarization. Nat Commun. 2013;4:2834. doi:10.1038/ncomms3834
84. Mahon OR, Browe DC, Gonzalez-Fernandez T, et al. Nano-particle mediated M2 macrophage polarization enhances bone formation and MSC osteogenesis in an IL-10 dependent manner. Biomaterials. 2020;239:119833. doi:10.1016/j.biomaterials.2020.119833
85. Kurowska-Stolarska M, Stolarski B, Kewin P, et al. IL-33 amplifies the polarization of alternatively activated macrophages that contribute to airway inflammation. J Immunol. 2009;183:6469–6477. doi:10.4049/jimmunol.0901575
86. Porta C, Riboldi E, Ippolito A, Sica A. Molecular and epigenetic basis of macrophage polarized activation. Semin Immunopathol. 2015;27:237–248. doi:10.1016/j.smim.2015.10.003
87. Lawrence T, Natoli G. Transcriptional regulation of macrophage polarization: enabling diversity with identity. Nat Rev Immunol. 2011;11:750–761. doi:10.1038/nri3088
88. Tao Y, Xu L, Liu X, et al. Chitosan-coated artesunate protects against ulcerative colitis via STAT6-mediated macrophage M2 polarization and intestinal barrier protection. Int J Biol Macromol. 2024;254:127680. doi:10.1016/j.ijbiomac.2023.127680
89. Ohmori Y, Hamilton TA. IL-4-induced STAT6 suppresses IFN-gamma-stimulated STAT1-dependent transcription in mouse macrophages. J Immunol. 1997;159:5474–5482.
90. Lu X, Chen L, Jiang C, Cao K, Gao Z, Wang Y. Microglia and macrophages contribute to the development and maintenance of sciatica in lumbar disc herniation. Pain. 2023;164:362–374. doi:10.1097/j.pain.0000000000002708
91. Ibeakanma C, Vanner S. TNF is a key mediator of the pronociceptive effects of mucosal supernatant from human ulcerative colitis on colonic DRG neurons. Gut. 2010;59:612–621. doi:10.1136/gut.2009.190439
92. Shamji MF, Guha D, Paul D, Shcharinsky A. Systemic inflammatory and Th17 immune activation among patients treated for lumbar radiculopathy exceeds that of patients treated for persistent postoperative neuropathic pain. Neurosurgery. 2017;81:537–544. doi:10.1093/neuros/nyx052
93. Peeters M, Detiger SEL, Karfeld-Sulzer LS, et al. T helper cell differentiation, heterogeneity, and plasticity. Cold Spring Harb Perspect Biol. 2018;10:a030338. doi:10.1101/cshperspect.a030338
94. Jansen K, Cevhertas L, Ma S, Satitsuksanoa P, Akdis M, van de Veen W. Regulatory B cells, A to Z. Allergy. 2021;76:2699–2715. doi:10.1111/all.14763
95. Song C, Zhou D, Cheng K, et al. Bioinformatics‐based discovery of intervertebral disc degeneration biomarkers and immune‐inflammatory infiltrates. JOR Spine. 2024;7:e1311. doi:10.1002/jsp2.1311
96. Wang X, Ma X, Li J, et al. Precision prediction of intervertebral disc degeneration in ankylosing spondylitis using a nomogram model reveals the pivotal role of Th2-type immune dysregulation. Front Immunol. 2025;16:1556738. doi:10.3389/fimmu.2025.1556738
97. Ruterbusch M, Pruner KB, Shehata L, Pepper M. In vivo CD4+T cell differentiation and function: revisiting the Th1/Th2 paradigm. Annu Rev Immunol. 2020;38:705–725. doi:10.1146/annurev-immunol-103019-085803
98. Wang W, Sung N, Gilman-Sachs A, Kwak-Kim J. T Helper (Th) cell profiles in pregnancy and recurrent pregnancy losses: th1/Th2/Th9/Th17/Th22/Tfh cells. Adv Funct Mater. 2022;11:2025. doi:10.3389/fimmu.2020.02025
99. Rajasekaran S, Soundararajan DCR, Nayagam SM, et al. Modic changes are associated with activation of intense inflammatory and host defense response pathways – molecular insights from proteomic analysis of human intervertebral discs. Spine J. 2022;22:19–38. doi:10.1016/j.spinee.2021.07.003
100. Freidin MB, Keser T, Gudelj I, et al. The association between low back pain and composition of IgG glycome. Sci Rep. 2016;6:26815. doi:10.1038/srep26815
101. Li Z, Li X, Chen C, et al. An in‐depth analysis of the immunomodulatory mechanisms of intervertebral disc degeneration. JOR Spine. 2022;5:e1233. doi:10.1002/jsp2.1233
102. Cheng L, Fan W, Liu B, Wang X, Nie L. Th17 lymphocyte levels are higher in patients with ruptured than non-ruptured lumbar discs, and are correlated with pain intensity. Injury. 2013;44:1805–1810. doi:10.1016/j.injury.2013.04.010
103. Suyama K, Sakai D, Watanabe M. The role of IL-17-mediated inflammatory processes in the pathogenesis of intervertebral disc degeneration and herniation: a comprehensive review. Front Cell Dev Biol. 2022;10:857164. doi:10.3389/fcell.2022.857164
104. Patel DD, Kuchroo VK. Th17 cell pathway in human immunity: lessons from genetics and therapeutic interventions. Immunity. 2015;43:1040–1051. doi:10.1016/j.immuni.2015.12.003
105. Miljkovic D, Trajkovic V. Inducible nitric oxide synthase activation by interleukin-17. Cytokine Growth Factor Rev. 2004;15:21–32. doi:10.1016/j.cytogfr.2003.10.003
106. Pène J, Chevalier S, Preisser L, et al. Chronically inflamed human tissues are infiltrated by highly differentiated Th17 lymphocytes. J Immunol. 2008;180:7423–7430. doi:10.4049/jimmunol.180.11.7423
107. Zhang W, Nie L, Wang Y, et al. CCL20 secretion from the nucleus pulposus improves the recruitment of CCR6-expressing Th17 cells to degenerated IVD tissues. PLoS One. 2013;8:e66286. doi:10.1371/journal.pone.0066286
108. Ritchlin C, Adamopoulos IE. Axial spondyloarthritis: new advances in diagnosis and management. BMJ. 2021;372:m4447. doi:10.1136/bmj.m4447
109. Ding T, Zhao S, Gu Y, et al. IL-17A regulates airway remodelling in COPD through the PI3K/AKT/mTOR pathway. Sci Rep. 2025;15:16546. doi:10.1038/s41598-025-00458-9
110. Hu B, Wang J, Wu X, Chen Y, Yuan W, Chen H. Interleukin-17 upregulates vascular endothelial growth factor by activating the JAK/STAT pathway in nucleus pulposus cells. Joint Bone Spine. 2017;84:327–334. doi:10.1016/j.jbspin.2016.05.014
111. Xu K, Chen W, Wang X, et al. Autophagy attenuates the catabolic effect during inflammatory conditions in nucleus pulposus cells, as sustained by NF-κB and JNK inhibition. Int J Mol Med. 2015;36:661–668. doi:10.3892/ijmm.2015.2280
112. Zielinski CE. T helper cell subsets: diversification of the field. Eur J Immunol. 2023;53:e2250218. doi:10.1002/eji.202250218
113. Sowerby JM, Rao DA. T cell–B cell interactions in human autoimmune diseases. Curr Opin Immunol. 2025;93:102539. doi:10.1016/j.coi.2025.102539
114. Osum KC, Jenkins MK. Toward a general model of CD4+ T cell subset specification and memory cell formation. Immunity. 2023;56:475–484. doi:10.1016/j.immuni.2023.02.010
115. Phan AT, Goldrath AW, Glass CK. Metabolic and epigenetic coordination of T cell and macrophage immunity. Immunity. 2017;46:714–729. doi:10.1016/j.immuni.2017.04.016
116. Wesolowski M, Can P, Warzecha K, et al. Long-term changes of Th17 and regulatory T cells in peripheral blood of dogs with spinal cord injury after intervertebral disc herniation. BMC Vet Res. 2023;19:90. doi:10.1186/s12917-023-03647-8
117. Zhou L, Cai F, Zhu H, et al. Immune-defensive microspheres promote regeneration of the nucleus pulposus by targeted entrapment of the inflammatory cascade during intervertebral disc degeneration. Bioact Mater. 2024;37:132–152. doi:10.1016/j.bioactmat.2024.03.020
118. Wu S, Jiang J, Wang D, et al. JAK/STAT3 signaling promotes pain and depression-like behaviors in rats with bone cancer pain by regulating Th17 cell differentiation. Brain Res Bull. 2025;221:111218. doi:10.1016/j.brainresbull.2025.111218
119. Kanno T, Nakajima T, Miyako K, Endo Y. Lipid metabolism in Th17 cell function. Pharmacol Ther. 2023;245:108411. doi:10.1016/j.pharmthera.2023.108411
120. Lyu F-J, Cui H, Pan H, et al. Painful intervertebral disc degeneration and inflammation: from laboratory evidence to clinical interventions. Bone Res. 2021;9:7. doi:10.1038/s41413-020-00125-x
121. Moldovan F. Correlation between peripheric blood markers and surgical invasiveness during humeral shaft fracture osteosynthesis in young and middle-aged patients. Diagnostics. 2024;14:1112. doi:10.3390/diagnostics14111112
122. Finnerup NB, Kuner R, Jensen TS. Neuropathic pain: from mechanisms to treatment. Physiol Rev. 2021;101:259–301. doi:10.1152/physrev.00045.2019
123. Shamji MF, Setton LA, Jarvis W, et al. Proinflammatory cytokine expression profile in degenerated and herniated human intervertebral disc tissues. Arthritis Rheum. 2010;62:1974–1982. doi:10.1002/art.27444
124. Kang L, Zhang H, Jia C, Zhang R, Shen C. Epigenetic modifications of inflammation in intervertebral disc degeneration. Ageing Res Rev. 2023;87:101902. doi:10.1016/j.arr.2023.101902
125. La Binch A, Cole AA, Breakwell LM, et al. Expression and regulation of neurotrophic and angiogenic factors during human intervertebral disc degeneration. Arthritis Res Ther. 2014;16:416. doi:10.1186/s13075-014-0416-1
126. Murata K, Sawaji Y, Alimasi W, et al. PGE1 attenuates IL-1β-induced NGF expression in human intervertebral disc cells. Spine. 2016;41:E710–E716. doi:10.1097/BRS.0000000000001379
127. Kusakabe T, Sawaji Y, Endo K, et al. DUSP-1 induced by PGE2 and PGE1 attenuates IL-1β-activated MAPK signaling, leading to suppression of NGF expression in human intervertebral disc cells. IJMS. 2021;23:371. doi:10.3390/ijms23010371
128. Yan M, Song Z, Kou H, et al. New progress in basic research of macrophages in the pathogenesis and treatment of low back pain. Front Cell Dev Biol. 2022;10:866857. doi:10.3389/fcell.2022.866857
129. Sun K, Jiang J, Wang Y, et al. The role of nerve fibers and their neurotransmitters in regulating intervertebral disc degeneration. Ageing Res Rev. 2022;81:101733. doi:10.1016/j.arr.2022.101733
130. Hamidi A, Song J, Thakur N, et al. Detailed characterization of neuro-immune responses following neuropathic injury in mice. Brain Res. 2011;1405:95–108. doi:10.1016/j.brainres.2011.06.022
131. McLachlan EM, Hu P. Inflammation in dorsal root ganglia after peripheral nerve injury: effects of the sympathetic innervation. Auton Neurosci. 2014;182:108–117. doi:10.1016/j.autneu.2013.12.009
132. Pan H, Li H, Guo S, et al. The mechanisms and functions of TNF-α in intervertebral disc degeneration. Exp Gerontology. 2023;174:112119. doi:10.1016/j.exger.2023.112119
133. Park J-B, Chang H, Kim Y-S. The pattern of interleukin-12 and T-helper types 1 and 2 cytokine expression in herniated lumbar disc tissue. Spine. 2002;27:2125–2128. doi:10.1097/00007632-200210010-00009
134. Gabr MA, Jing L, Helbling AR, et al. Interleukin‐17 synergizes with IFNγ or TNFα to promote inflammatory mediator release and intercellular adhesion molecule‐1 (ICAM‐1) expression in human intervertebral disc cells. J Orthop Res. 2011;29:1–7. doi:10.1002/jor.21206
135. Wilke H-J, Sciortino V. The past, present, and the future of disc nucleus replacement. A systematic review of a large diversity of ideas and experiences. Biomaterials. 2025;312:122717. doi:10.1016/j.biomaterials.2024.122717
136. Janssen M, Garcia R, Miller L, et al. Challenges and solutions for lumbar total disc replacement implantation. Spine. 2017;42:S108–S111. doi:10.1097/BRS.0000000000002454
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Engeletin Alleviates the Inflammation and Apoptosis in Intervertebral Disc Degeneration via Inhibiting the NF-κB and MAPK Pathways
Li B, Yang X, Zhang P, Guo J, Rong K, Wang X, Cao X, Zhou T, Zhao J
Journal of Inflammation Research 2022, 15:5767-5783
Published Date: 10 October 2022
Shensong Yangxin Capsule Reduces the Susceptibility of Arrhythmia in db/db Mice via Inhibiting the Inflammatory Response Induced by Endothelium Dysfunction
Zhang J, Li H, Wang D, Gu J, Hou Y, Wu Y
Drug Design, Development and Therapy 2023, 17:313-330
Published Date: 5 February 2023
The Temporal Changes in Ankle Joint Pathology, Pain and Secondary Osteoporosis in Collagen-Induced Arthritis Rats
Liu Q, Nan N, Li W, Dong M, Pu W, Liu Y, Zhao J, Hao H
Journal of Inflammation Research 2024, 17:9795-9804
Published Date: 26 November 2024
Thymosin β4 Regulates Tissue Inflammatory Response in Mouse Nonalcoholic Fatty Liver Disease by Promoting Macrophage M2-Type Polarization
Zhu Z, Liao Y, Mou Q, Liu H, Shen Y, Zhu L, Cong S
Journal of Inflammation Research 2025, 18:5791-5809
Published Date: 29 April 2025
Application of Multidisciplinary Rehabilitation Nursing in Patients with Brucellar Spondylitis and Its Effects on Pain, Inflammation, and Immune Function
Zhang Y, Zuo X, Yang X, Li R, Wang H, Yu J, Yang X
Journal of Multidisciplinary Healthcare 2025, 18:3813-3826
Published Date: 2 July 2025