Back to Journals » Infection and Drug Resistance » Volume 16
High Prevalence of Group III-Like Mutations Among BLPACR and First Report of Haemophilus influenzae ST95 Isolated from Blood in China
Authors Lin J ![]() , Wang Y, Lin C, Li R, Wang G
, Wang Y, Lin C, Li R, Wang G
Received 4 December 2022
Accepted for publication 14 February 2023
Published 17 February 2023 Volume 2023:16 Pages 999—1008
DOI https://doi.org/10.2147/IDR.S400207
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 4
Editor who approved publication: Prof. Dr. Héctor Mora-Montes
Jiansheng Lin,1,2 Yinna Wang,2 Chunli Lin,2 Ran Li,3 Gaoxiong Wang1,4
1School of Public Health of Fujian Medical University, Fuzhou, People’s Republic of China; 2Microbiology Laboratory, Quanzhou Women’s and Children’s Hospital, Quanzhou, People’s Republic of China; 3Department of Anesthesiology, The Second Affiliated Hospital of Fujian Medical University, Quanzhou, People’s Republic of China; 4Research Administration Office, Quanzhou Women’s and Children’s Hospital, Quanzhou, People’s Republic of China
Correspondence: Ran Li, Department of Anesthesiology, The Second Affiliated Hospital of Fujian Medical University, 34 Zhongshan North Road, Licheng District, Quanzhou, Fujian Province, 350122, People’s Republic of China, Tel +86 595 22791140, Email [email protected] Gaoxiong Wang, Research Administration Office, Quanzhou Women’s and Children’s Hospital, 700 Fengze Street, Fengze District, Quanzhou, Fujian Province, 350122, People’s Republic of China, Tel +86 595 22131685, Email [email protected]
Purpose: We aimed to evaluate antibiotic resistance and molecular epidemiological characteristics of non-invasive Haemophilus influenzae (H. influenzae) from pneumonia patients and analyze the whole genome of one invasive H. influenzae isolated from blood in pediatric patients.
Methods: Antibiotic susceptibility was tested using the turbidimetric method. β-lactamase-producing and serotyping genes were evaluated via multiplex polymerase chain reaction (PCR), and ftsI was amplified using high-fidelity PCR. Lastly, whole genome sequencing (WGS) was conducted using Illumina HiSeq and PacBio sequencing technology.
Results: We observed that the ampicillin (AMP) and amoxicillin/clavulanate (AMC) resistance rates of non-invasive H. influenzae were as high as 99.06% (after adjustment) and 49.53%, respectively. The β-lactamase gene of 106 AMP-resistant strains was blaTEM-1. Group III-like mutation accounted for 71.15% of β-lactamase-positive, AMC-resistant (BLPACR) strain mutants. The novel Asn-526→His mutation was present in one β-lactamase-negative AMP-susceptible (BLNAS) strain. Non-invasive H. influenzae strains all belonged to non-typeable H. influenzae (NTHi). In contrast, the invasive H. influenzae 108 isolated from blood in China belonged to H. influenzae type b (Hib). It belonged to sequence typing ST95 and exhibited sensitivity to all 11 antibiotics. Three prophages were identified, and the capb loci of the H. influenzae strain 108 revealed regions I–III exist in duplicate; however, complete deletion of IS1016 was only present in one of the copies.
Conclusion: Non-invasive H. influenzae NTHi with β-lactamase-positive was highly prevalent. Notably, group III-like mutations had increased prevalence among BLPACR strains. H. influenzae belonging to Hib and ST95 was first reported to cause sepsis in China.
Keywords: group III-like mutations, BLPACR, prevalence, invasive Haemophilus influenzae, genome
Introduction
H. influenzae, a gram-negative bacillus that commonly colonizes the nasopharynx of humans, is an opportunistic pathogen that can cause upper respiratory tract infection, pneumonia, asthma, other respiratory diseases, and severe systemic infections such as purulent meningitis and sepsis.1,2 Invasive H. influenzae disease is defined as a H. influenzae infection isolated from sterile sites such as blood, pleural fluid, peritoneal fluid, cerebrospinal fluid etc. Children under five years of age, especially those under one year of age, are more likely to develop invasive H. influenzae disease. It can also occur in individuals without any underlying diseases. It is more often severe, with a high mortality rate or various sequelae left in survivors.2,3 Non-invasive H. influenzae disease has a better prognosis; however, the disease burden remains high owing to its high occurrence in children.
H. influenzae is divided into two types: encapsulated type and non-encapsulated type. There are six serotypes: a, b, c, d, e, and f, often abbreviated as Hia, Hib, Hic, Hid, Hie, and Hif, respectively. NTHi is non-encapsulated and does not agglutinate with either type of serum. In a few cases, the encapsulated strains ceased producing capsules to adapt to the environment. Therefore, simple serotyping will be misjudged as NTHi, and capsular gene testing is required. In recent years, Hib infections have become rare with the widespread introduction of the Hib vaccine. Previous population-based studies in Hefei and Nanning, China, found that the incidence of Hib meningitis was 10.66 and 0.98 cases/100,000 children <5 years of age in the pre- and post-vaccine eras, respectively.4,5 However, the proportion of Hia and Hif in the infected population in some regions has increased,3,6 and NTHi has become the most common pathogen of H. influenzae disease globally.7 The gene sequence IS1016 has been associated with an increased capacity to cause severe infections. It is usually present in encapsulated strains but is sometimes harbored by NTHi.8 In nasopharyngeal colonization, NTHi can easily exchange large DNA fragments between strains. This exchange is done to obtain new virulence factors to enhance its ability to resist human neutrophils and complement-mediated bactericidal effects.9,10 Consequently, NTHi is already the most common cause of invasive H. influenzae disease in children under one year of age.11
The preferred drugs for children infected with H. influenzae include oral ampicillin (AMP), amoxicillin/clavulanate (AMC), and an intravenous infusion of cefotaxime (CTX) and ceftriaxone. With the widespread use of antibiotics, the increasing trend of the H. influenzae resistance rate to various antibacterial drugs has received extensive clinical attention. There are two main mechanisms of β-lactam antibiotic resistance in H. influenzae. One mechanism is the inactivation of AMP by hydrolysis of β-lactamase (TEM-1 or ROB-1 type encoded by blaTEM-1 or blaROB-1 gene, respectively). H. influenzae with β-lactamase-positive AMP resistance is called β-lactamase-positive, AMP-resistant (BLPAR). The other mechanism is that an amino acid site mutation of the penicillin-binding protein (PBP3) encoded by ftsI causes changes in the spatial conformation, which reduces its affinity for AMP. H. influenzae with ftsI mutation for AMP resistance without producing β-lactamase is called β-lactamase negative, AMP resistant (BLNAR).12 AMC-resistant strain with β-lactamase-positive is called β-lactamase-producing AMC-resistant (BLPACR), often occurring in ftsI mutations.13
A recent multicenter study in China showed that the resistance rates of H. influenzae to AMP was increasing year by year, and there were statistically significant differences in the resistance rates of all antibiotics in different regions except chloramphenicol.14 Since the introduction of the Hib vaccine in Fujian, China, in 2003, there has been no research on antibiotic resistance and molecular epidemiological characteristics of H. influenzae. Therefore, in this study, we investigated the antibiotic resistance and molecular characterization of H. influenzae isolated from pediatric children in Quanzhou, China, to provide a basis for the prevention and treatment of H. influenzae disease.
Materials and Methods
Strains
A total of 108 H. influenzae cases were collected from hospitalized children in Quanzhou Women’s and Children’s Hospital from July 2021 to August 2022. The mean age of the children was 1.56±0.16 years. There were 77 boys and 31 girls, and the sex ratio was 2.48. The non-invasive H. influenzae inclusion criteria were <10 squamous epithelium and >25 white blood cells (WBC) in each low-power field of view to avoid the collection of colonized H. influenzae. Gram-negative bacilli were seen in the WBC. In addition, there were evident clinical features of H. influenzae community-acquired pneumonia. Invasive H. influenzae inclusion criteria was that H. influenzae was cultured from double blood culture bottles. Repeated isolates from the same infection sites of the same patients were excluded. Among the 108 H. influenzae strains, 106 (98.15%) non-invasive strains were isolated from sputum, one (0.92%) non-invasive strain from bronchoalveolar lavage fluid, and one (0.92%) named invasive H. influenzae 108 from the blood of an unvaccinated pediatric patient. This pediatric patient was clinically characterized by fever, headache, and vomiting. During the first three days of hospitalization, ceftazidime (CAZ) was used for anti-infection. According to 1.151×109/L WBC in cerebrospinal fluid, positive culture for H. influenzae and the 135.12 mg/L value of C-reactive protein, vancomycin + meropenem were given for the next 3 days. After 6 days of treatment, the symptoms improved significantly and he was discharged from the hospital. H. influenzae strains were stored at −80℃ in BHI-sheep blood-20% glycerol storage solution. This study complied with the Declaration of Helsinki, and ethical permission for this study was approved by the Quanzhou Women’s and Children’s Hospital ethics committee (2021 Ethical Review No. 16). Guardians provided written informed consent on behalf of the minors. H. influenzae strains were identified by VITEK 2 bioMérieux bacterial identification instrument. H. influenzae ATCC 49247 was the quality control strain.
Antibiotic Susceptibility Test
Antibiotic susceptibility testing was performed using the turbidimetric method (similar to microdilution) of the ATB HAEMO CLSI (12) strip from bioMérieux. Antibiotics that were evaluated include AMP, AMC, Cefoxitin (CEP), Cefaclor (CEC), CAZ, Cefotaxime (CTX), Ofloxacin (OFX), Rifampicin (Rif), Chloramphenicol (CHL), Tetracycline (TCY), and Cotrimoxazole (SXT). First, H. influenzae colonies were cultured for 24 hours, and the bacterial suspension was adjusted to 0.5 McF concentration (0.85% NaCl). Subsequently, 50 µL of bacterial suspension was transferred to ATB S medium. After mixing, 135 µL of suspension was added to each well of the ATB HAEMO CLSI (12) reagent strip. The strip was then placed in a CO2 incubator for 20 h. Finally, the results were read automatically using an ATB instrument. The results were interpreted based on the breakpoints recommended by the CLSI M100-S23 2013 committee for H. influenzae. The quality control strain used was H. influenzae ATCC 49247.
Detection of β-Lactamase-Producing Genes and Serotype Capsular Genes
β-lactamase-producing genes and serotype capsular genes were assessed by the Multiplex PCR method. Bacterial DNA was extracted according to the instructions of the TIANamp Bacteria DNA Kit. The PCR amplification primer sequences and product lengths of β-lactamase-producing genes (blaTEM-1 and blaROB-1) and serotype capsular genes (a–f) are listed in Table S1. Multiplex PCR reagents were purchased from Vazyme. The total reaction volume of 50 µL consisted of 7 µL RNase-free ddH2O, 25 µL 2× Multiplex buffer, 5 µL 10× primer mix (Premix all amplification primers so that the concentration of each primer is 1 µM), 2 µL template DNA, 1 µL Multiplex DNA polymerase, and 10 µL 5× Multiplex GC Enhancer. The thermal cycle was programmed for 5 min at 95℃ for pre-denaturation, followed by 35 cycles of 30s at 95℃ for denaturation, 90s at 60℃ for annealing, and 90s at 72℃ for extension; a final extension was conducted for 10 min at 72℃. The PCR products were observed using agarose gel electrophoresis.
Amplification and Sequencing of fstI Gene
The ftsI gene was amplified by high-fidelity PCR. The PCR amplification primer sequences and product lengths of ftsI are listed in Table S1. High-fidelity PCR was purchased from Vazyme. The total reaction volume of 50 µL consisted of 20 µL ddH2O, 25 µL 2× Phanta Max Master Mix, 2 µL primer F, 2 µL primer R, and 1 µL template DNA. The thermal cycle was programmed for 3 min at 95℃ for pre-denaturation, followed by 32 cycles of 15s at 95℃ for denaturation, 15s at 58℃ for annealing, 60s at 72℃ for extension; a final extension was conducted for 5 min at 72℃. The PCR products were observed using agarose gel electrophoresis.
Subsequently, the PCR products were sent to Fuzhou Shangya Biological Co., Ltd. for Sanger sequencing. Amino acids 330–530 of PBP3, including the highly conserved motifs Ser-Thr-Val-Lys (STVK), Lys-Thr-Gly (KTG), and Ser-Ser-Asn (SSN), were compared with the corresponding sequence of H. influenzae Rd KW20. The PBP3 mutation patterns were determined according to the rules reported in the literature:15 Group I was defined as Arg-517→His-517. Group II as Asn-526→Lys-526). Group II was further divided into four subgroups. IIa: only Asn-526→Lys-526; IIb: Asn-526→Lys-526 and Ala-502→Val-502; group IIc: Asn-526→Lys-526 and Ala-502→Thr-502; group IId: Asn-526→Lys-526 and Ile-449→Val. Group III was defined as Met-377→Ile, Ser-385→Thr, Leu-389→Phe, and Asn-526→Lys). Group III-like was defined as Met-377→Ile, Ser-385→Thr, and/or Leu-389→Phe in the SSN motif, Arg-517→His and Thr-532→Ser in the KTG motif, Asp-350→Asn in the STVK motif; and finally, Ser-357→Asn.
Whole Genome Sequencing (WGS)
The H. influenzae 108 isolated from blood was sent to Majorbio Bio-Pharm Technology Co., Ltd. for WGS. In this study, whole genome de novo sequencing was performed using the second-generation Illumina HiSeq and third-generation PacBio sequencing methods. No less than 100× Illumina sequencing data of the genome and 100× PacBio sequencing data were provided to ensure a complete and accurate assembly while avoiding the loss of small plasmid (<15 kb) information. Further, this information ensured that the complete genome contained the plasmid. The genome coverage of H. influenzae 108 strain was 100% for this sequencing. The Hicap tool (https://github.com/scwatts/hicap), online bioinformatic tools (http://www.genomicepidemiology.org/), the comprehensive antibiotic resistance database (https://card.mcmaster.ca/), and Abricate v.0.8 (https://github.com/tseemann/abricate) with the VFDB database (http://www.mgc.ac.cn/Vfs/) were used to analyze the serotypes, sequence types, resistomes, and virulomes. A >80% for gene identity was considered as the threshold for predicting virulence genes. In addition, PHASTER (https://phaster.ca/) was used to predict the prophage regions. A CLC sequence viewer was used to investigate single nucleotide polymorphisms (SNPs) of the cap loci, the complete deletion of the putative virulence-enhancing IS1016, and the partial deletion of bexA. 16S rRNA was utilized for performing phylogenetic analyses using megax software. The evolutionary relationship was inferred using maximum likelihood method.
Nucleotide Sequence
The complete sequences of H. influenzae 108 was deposited in GenBank under the accession number: PRJNA900629.
Results
Analysis of Drug Resistance in H. influenzae
We observed that the AMP resistance rate of non-invasive H. influenzae was as high as 91.59%, followed by the SXT resistance rate of 71.96%, CEC of 60.74%, CAZ of 56.07%, AMC of 49.53%, and CEP of 46.73%. CTX, OFX, Rif, and CHL were not resistant, as presented in Table 1. Only 2 non-invasive H. influenzae strains (1.87%) showed multidrug-resistance (MDR). Their MDR pattern was resistance to β-lactam antibiotics (AMP, AMC, CEP, CEC, CAZ), TCY and SXT. One BLNAS strain and the invasive H. influenzae 108 was sensitive to all 11 antibiotics.
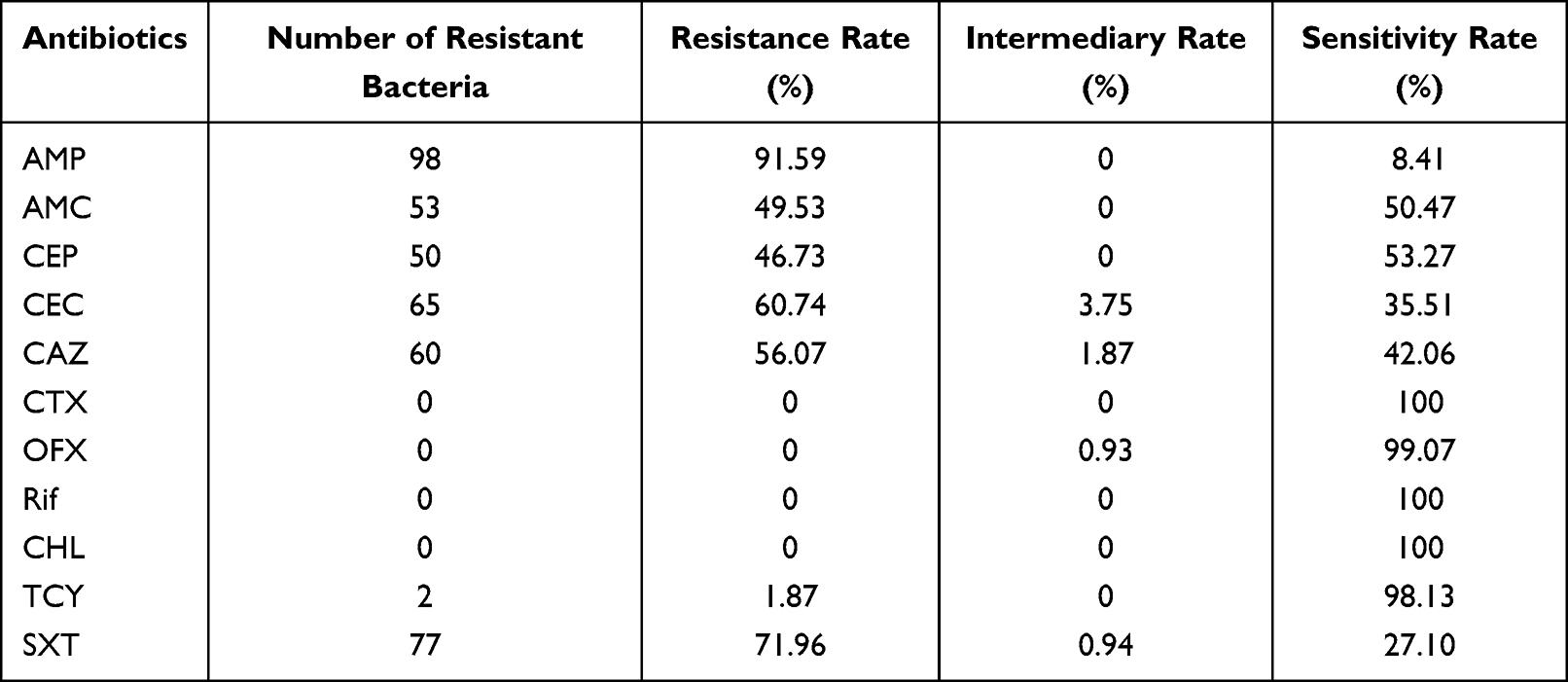 |
Table 1 Antibiotic Resistance of 107 Non-Invasive H. influenzae Strains |
Analysis of β-Lactamase-Producing Genes in 107 Non-Invasive H. influenzae Strains
The β-lactamase gene of 98 AMP-resistant strains were all blaTEM-1 genes but not blaROB-1. BLNAR strains were not detected. The instructions of the H. influenzae drug susceptibility test kit recommend verifying the AMP-susceptible strains to confirm the absence of β-lactamase. Therefore, the β-lactamase-producing gene of AMP-sensitive strains was detected. Notably, the blaTEM-1 gene was detected in eight of the nine isolates. Only one strain was confirmed as β-lactamase-producing gene negative. Finally, the AMP resistance rate was corrected to 99.06%.
Mutation Analysis of ftsI in 53 BLPACR Strains and One BLNAS Strain, and Serotype Analysis of 107 Non-Invasive H. influenzae Strains
We identified eight mutational patterns from 52 BLPACR strains; however, one BLPACR strain expressed no mutations (Table 2). Thirty-seven strains were group III-like, accounting for 71.15% of BLPACR strain mutants. Eleven strains belonged to group III, of which four were combined with group IIb or group IIc. Two strains belonged to group IIc. The untyped patterns of the two BLPACR strains were Asp-350→Asn, Ser-357→Asn, Met-377→Ile, and Ser-385→Thr. In one BLNAS strain, the mutation pattern was Asp-350→Asn, Ser-385→Thr, Leu-389→Phe, and Asn-526→His.
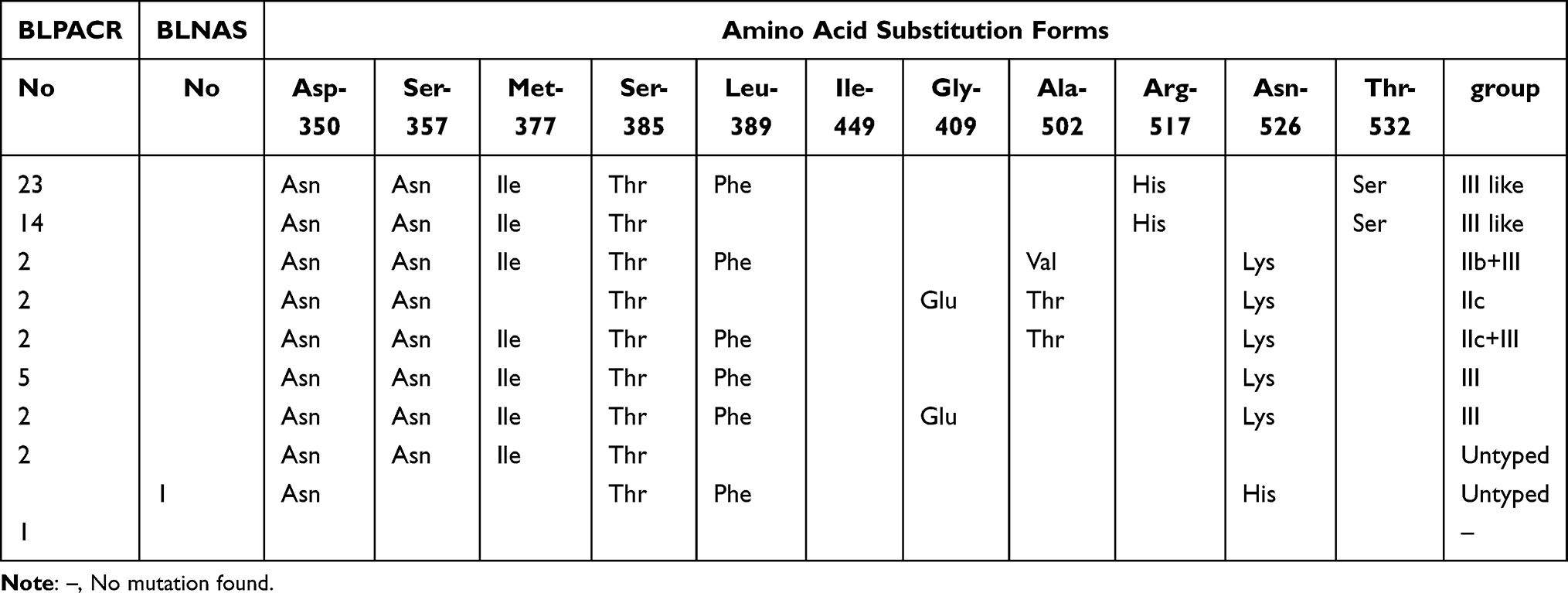 |
Table 2 Different Amino Acid Substitution Patterns Identified in Part of PBP3 from 53 BLPACR Strains and One BLNAS Strain |
All 107 non-invasive H. influenzae strains belonged to NTHi because the six serotype genes were negative.
Whole Genome Analysis of Invasive H. influenzae 108
WGS of invasive H. influenzae 108 revealed the genome size to be 1,903,516 bp, comprising 1798 protein-coding sequences, 20 rRNAs, and 57 tRNAs, with a GC content of 38.1%. No plasmids were present in the strains. Three prophages were identified: one intact, two questionable (Figure 1). The prophages 1 (questionable), located at 478,683–513,213, is 34.5 Kb in length and contains 54 proteins. The prophages 2 (intact), located at 1,089,401–1,145,733, is 56.3 Kb in full length and contains 61 proteins. The prophages 3 (questionable), located at 1,303,040–1,319,103, is 16 Kb in full length and contains 22 proteins. No exogenous antimicrobial resistance or putative virulence genes were present in the prophage regions.
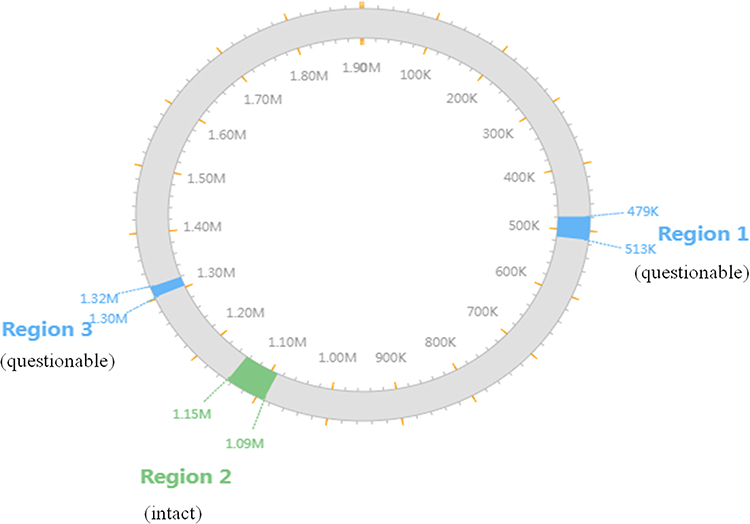 |
Figure 1 The locations of the prophages on the chromosome of the strain Regions 1 to 3 represent prophages 1 to 3; Blue, red and green represent questionable, incomplete, and intact, respectively. |
The invasive H. influenzae 108 were discovered to belong to Hib, and its sequence type belonged to ST95. β-lactamase-producing genes and ftsI mutations were not detected. However, the LpsA gene for intrinsic peptide antibiotic resistance and the EF-Tu R234F amino acid mutation, which mediates felfamycin resistance, were present. According to the analysis of 16S rRNA phylogenetic tree, H. influenzae 108 was the closest to H. influenzae NCTC11394 and H. influenzae M3918 (Figure 2).
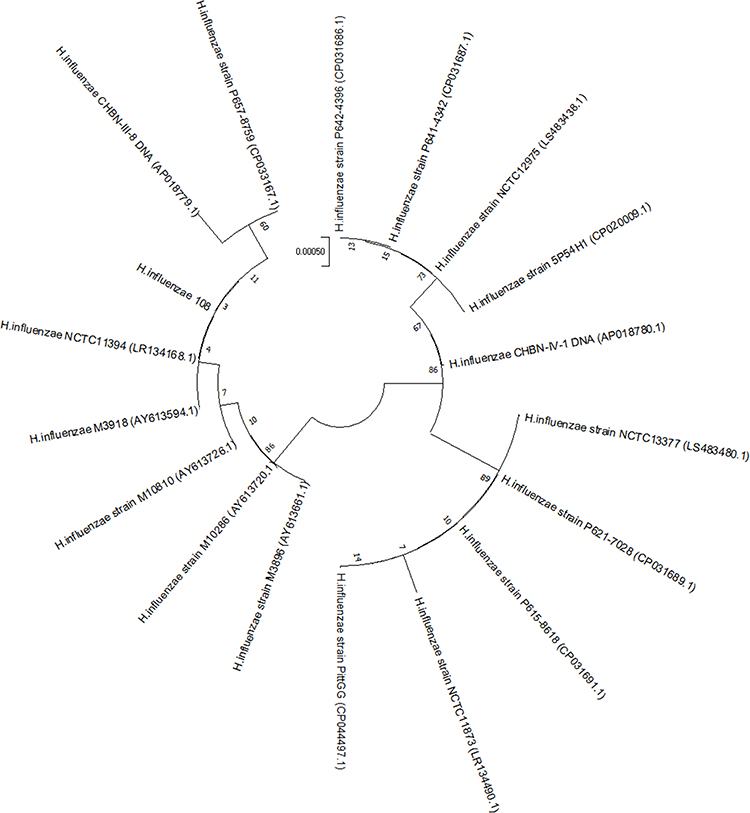 |
Figure 2 Phylogenetic analysis between the H. infuenzae strains The 17 H. infuenzae genomes available on the NCBI database and H. influenzae strain 108 isolated in this study. |
Comparative analysis of the capb loci of the H. influenzae 108 and the H. influenzae 146 references (GenBank accession number: PQJI00000000.1) revealed regions I, II, and III exist in duplicates. However, IS1016 complete deletion rather than partial deletion was identified upstream of the region I in only one of the duplicate copies (Figure 3). Compared with the multiple mutations in the SNPs analysis of the capb locus from reference H. influenzae 146, no mutations were identified in the H. influenzae 108.
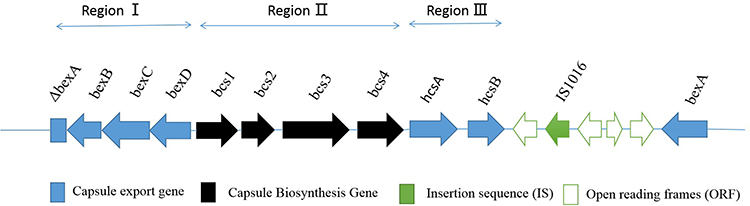 |
Figure 3 Cap locus structure identified in H. influenzae strain 108 Arrows indicate the direction of transcription and translation, and squares indicate the deletion of a part of the gene. |
There are 79 predicted related virulence genes, as displayed in Table S2. These virulence genes can be divided into four categories: capsule gene (20 genes), lipopolysaccharide (LPS) biosynthesis-related genes (45 genes), nonspecific virulence factors (6 genes), defensive virulence factors (3 genes), and offensive virulence factors (5 genes).
Discussion
H. influenzae, a common pathogen of community-acquired pneumonia, was majority isolated from the sputum of the lower respiratory tract. In this study, we observed a 99.06% AMP resistance rate, which was higher than the 61.4% reported by a Chinese multicenter study in 2016,16 and the 13% in Paraguay17 but similar to the 91.1% reported by another Chinese study in 2022.18 This increasing trend may be attributed to the 99.06% detection rate of the β-lactamase gene in this study. Except for AMC, the resistance rates of CEC, CTX, OFX, Rif, TCY, and SXT were consistent with those reported in another Chinese study in 2022.18 Compared with other reports of CHL low resistance,18 no CHL resistance was observed in this study. A multi-center study in China showed that the major multidrug resistance pattern was resistant to β-lactams, macrolides, and sulfonamides,14 which was inconsistent with the MDR pattern in this study. The MDR detection rate in this study was much lower than 12.8% reported in China.19 MDR may exhibit a unique resistance pattern in a low prevalence state at Quanzhou.
Given that CHL, OFX, and TCY are contraindicated in clinical use for pediatric patients, CTX and Rif, with a 100% sensitivity rate, can be the first choice for empirical medication. The high AMC resistance rate of 49.53% observed in this study was associated with the reduced drug sensitivity caused by the high PBP3 mutation rate. Therefore, it is not recommended that AMC be the drug of choice for treatment.
Chromosome-mediated overproduction of dihydrofolate reductase is the primary resistance mechanism of SXT. SXT resistance rate is also high, close to 72%; therefore, it is not recommended for empirical treatment.
The β-lactamase genes identified in this study were all blaTEM genes, consistent with a previous Chinese study.20 Furthermore, eight strains carrying β-lactamase-producing blaTEM-1 genes were detected from nine AMP-sensitive strains to clarify the detection efficiency of the ATB HAEMO CLSI (12) strip for AMP sensitivity. The missed detection rate was 8/107 (7.48%), which exhibits effective false negative prevention.
The primary resistance-conferring mutations are in the transpeptidase domain of PBP3. These mutations decrease the binding of β-lactam antibiotics to PBP3 under the selective pressures of different β-lactam antibiotics.21,22 BLNAR strains had reduced sensitivity to AMC and its combinations with beta-lactamase inhibitors and second-generation cephalosporins.23 However, no BLNAR strains were identified in this study. The detection rate of BLPACR strains was 49.53%, which was higher than the 15.4% reported in another Chinese study,24 23% in a British Columbian study,25 and 32.1% in a Korean study.26 The MICs of CTX for group III-like strains were at least 10 times higher than fully susceptible isolates.22 Moreover, group III and III-like H. influenzae strains were more resistant to β-lactam antibiotics than group I and II strains.26,27 BLPACR strains were the main group III and III-like strains in this study, which is consistent with previous literature.26,28 However, group III-like BLPACR strains exhibited a high prevalence. One BLPACR strain was observed to have no variation in the amino acid sequence 350–532 of PBP3. We speculated that this strain might carry resistance mutations in other regions of PBP3 or other β-lactam resistance genes. Asn-526→His in the KTG motif, with Asp-350→Asn, Ser-385→Thr, and Leu-389→Phe were first reported in one BLNAS strain, which may slightly reduce or not affect the MIC values of β-lactam antibiotics.
With the introduction of the H. influenzae vaccine, the serotypes of strains causing respiratory diseases are mainly NTHi.28 In this study, we observed that all non-invasive strains were NTHi, consistent with the strain isolated from the lower respiratory tract previously reported in China.29 However, BLPACR NTHi strains are highly prevalent in Korea.26 We also observed that BLPACR NTHi strains were popular in China, thereby drawing attention to the increasing β-lactam resistance.
Compared with the reference genome of H. influenzae 146 belonging to Hib, this H. influenzae 108 strain’s genome is relatively large but does not carry a plasmid containing the blaTEM-1B gene and has no mutations in ftsI, which may explain the lack of β-lactam antibiotic resistance. Diversification due to genome rearrangement has been reported to contribute to the global spread of H. influenzae.30 Compared with the strains reported to carry four complete prophages and one incomplete prophage,31 the strains in this study carried fewer prophages, resulting in less phenotype diversity. In the pre-vaccination era, H. influenzae ST95, which belonged to Hib and was transferred by healthy Japanese children, exhibited resistance to AMP and TCY.32 In the post-Hib vaccine era, some invasive H. influenzae Hib and ST-95 isolated from Canada exhibited no resistance, whereas some demonstrated resistance to AMP owing to β-lactamase production.33 H. influenzae Hib and ST-95 isolated from children with bacterial meningitis are typical for strains circulating in Russia.34 H. influenzae Hib and ST-95 have been isolated from the respiratory tract in China.35 In this study, we identified H. influenzae Hib and ST-95 isolated from Chinese children’s blood for the first time, which demonstrated no resistance to eleven antibiotics. H. influenzae NCTC11394 and H. influenzae M3918 isolated from USA also belonged to Hib, which is the same as H. influenzae 108 serotype. Although H. influenzae NCTC11394 belonged to ST6 through MLST analysis by us, the scores of 6 of the 7 housekeeping genes of its ST were consistent with those of H. influenzae 108 and H. influenzae 108 belonged to ST-6 complex. This further indicates that the two strains are related in evolution.
The capsular polysaccharide of H. influenzae Hib is a major virulence factor and a crucial antigen for Hib vaccination.36 The capb locus, which consists of three functionally distinct regions (I to III), is responsible for generating the type b capsule.37 Multiple copies of the capb locus exhibit strong virulence and can result in increased polysaccharide production. Another large category of virus-related genes are those involved in LPS synthesis.38 The LPS of H. influenzae may play an important role in the pathogenesis of meningitis by destroying the blood-brain barrier39 and septicemia.40 The strains in this study also comprised invasiveness LPS biosynthesis genes, lic1 and lic2, consistent with Cardosoa et al, suggesting that these genes exist in invasive strains.37 We also observed that H. influenzae Hib and ST-95 caused severe septicemia in unvaccinated children, highlighting the indispensable and crucial role of vaccines.
Conclusions
In this study, AMP and AMC were not suitable for empiric medication due to high drug resistance rate. MDR was in a very low prevalence state at this region. BLPACR NTHi strains dominated by group III-like showed high prevalence. H. influenzae Hib and ST-95 was isolated from blood for the first time in China. In addition, genomic surveillance study conducted to monitor changes in the molecular epidemiology of invasive H. influenzae infection was important.
Acknowledgments
We expressed our appreciation to the patient and her parents in this study.
Funding
This research was funded by Quanzhou Municipal Science and Technology Plan Project (Grant No: 2021N089S).
Disclosure
The authors declare that they have no competing interests.
References
1. Zhang J, Zhu Z, Zuo X, Pan H, Gu Y, Yuan Y. The role of NTHi colonization and infection in the pathogenesis of neutrophilic asthma. Respir Res. 2020;21(1):170. doi:10.1186/s12931-020-01438-5
2. Soeters HM, Oliver SE, Plumb ID, et al. Epidemiology of invasive Haemophilus influenzae serotype a disease—United States, 2008–2017. Clin Infect Dis. 2021;73(2):e371–e379. doi:10.1093/cid/ciaa875
3. Eton V, Schroeter A, Kelly L, Kirlew M, Tsang RSW, Ulanova M. Epidemiology of invasive pneumococcal and Haemophilus influenzae diseases in Northwestern Ontario, Canada, 2010–2015. Int J Infect Dis. 2017;65:27–33. doi:10.1016/j.ijid.2017.09.016
4. Yang Y, Leng D, Lu D. Pediatric Haemophilus influenzae type b meninngitis in Hefei city: an epidemiologic study. Zhonghua Yi Xue Za Zhi. 1998;78(4):251–253.
5. Dong B-Q, Tang -Z-Z, Li M, et al. Epidemiologic surveillance for bacterial meningitis in 140 000 children under 5 years of age in Nanning district, Guangxi province. Zhonghua Liu Xing Bing Xue Za Zhi. 2004;25(5):391–395.
6. Van Eldere J, Slack MPE, Ladhani S, Cripps,AW. Non-typeable Haemophilus influenzae, an under-recognised pathogen. Lancet Infect Dis. 2014;14(12):1281–1292. doi:10.1016/S1473-3099(14)70734-0
7. Slack Mary PE. The evidence for non-typeable Haemophilus influenzae as a causative agent of childhood pneumonia. Pneumonia. 2017;9(1):9. doi:10.1186/s41479-017-0033-2
8. Karlsson E, Melhus Å. Nontypeable Haemophilus influenzae strains with the capsule-associated insertion element IS1016 may mimic encapsulated strains. APMIS. 2006;114(9):633–640. doi:10.1111/j.1600-0463.2006.apm_333.x
9. Mell JC, Lee JY, Firme M, Sinha S, Redfield,RJ. Extensive cotransformation of natural variation into chromosomes of naturally competent Haemophilus influenzae. G3. 2014;4(4):717–731. doi:10.1534/g3.113.009597
10. Langereis JD, Weiser JN. Shielding of a lipooligosaccharide IgM epitope allows evasion of neutrophil-mediated killing of an invasive strain of nontypeable Haemophilus influenzae. mBio. 2014;5(4):e01478–01414. doi:10.1128/mBio.01478-14
11. Takla A, Schönfeld V, Claus H. Invasive Haemophilus influenzae infections in Germany after the introduction of routine childhood immunization, 2001––2016. Open Forum Infect Dis. 2020;7(10):ofaa444. doi:10.1093/ofid/ofaa444
12. Hotomi M, Fujihara K, Billal DS, Suzuki K, Nishimura T. Genetic characteristics and clonal dissemination of β-lactamase-negative ampicillin-resistant Haemophilus influenzae strains isolated from the upper respiratory tract of patients in Japan. Antimicrob Agents Chemother. 2007;51(11):3969–3976. doi:10.1128/AAC.00422-07
13. Ubukata K, Shibasaki Y, Yamamoto K, Chiba N, Hasegawa K. Association of amino acid substitutions in penicillin-binding protein 3 with β-lactam resistance in β-lactamase-negative ampicillin-resistant Haemophilus influenzae. Antimicrob Agents Chemother. 2001;45(6):1693–1699. doi:10.1128/AAC.45.6.1693-1699.2001
14. Zhou M, Fu P, Fang C, et al. Antimicrobial resistance of Haemophilus influenzae isolates from pediatric hospitals in Mainland China: report from the ISPED program, 2017–2019. Indian J Med Microbiol. 2021;39(4):434–438. doi:10.1016/j.ijmmb.2021.09.001
15. Dabernat H, Delmas C, Seguy M, Pelissier R, Faucon G, Bennamani S. Diversity of β-lactam resistance-conferring amino acid substitutions in penicillin-binding protein 3 of Haemophilus influenzae. Antimicrob Agents Chemother. 2002;46(7):2208–2218. doi:10.1128/AAC.46.7.2208-2218.2002
16. Wang H-J, Wang C-Q, Hua C-Z, et al. Antibiotic resistance profiles of Haemophilus influenzae isolates from children in 2016: a multicenter study in China. Can J Infect Dis Med Microbiol. 2019;2019:6456321. doi:10.1155/2019/6456321
17. León ME, Kawabata A, Nagai M, et al. Estudio epidemiológico de Haemophilus influenzae causante de enfermedad invasiva y no invasiva en Paraguay (1999–2017) [Epidemiological study of Haemophilus influenzae causing invasive and non-invasive disease in Paraguay]. Enferm Infecc Microbiol Clin. 2021;39(2):59–64. doi:10.1016/j.eimc.2020.02.020
18. Zhu X, Ye T, Zhong H, et al. Distribution and drug resistance of bacterial pathogens associated with lower respiratory tract infection in children and the effect of COVID-19 on the distribution of pathogens. Can J Infect Dis Med Microbiol. 2022;2022:1181283. doi:10.1155/2022/1181283
19. Shen XZ, Lu Q, Deng L, et al. Resistance of Haemophilus influenzae isolates in children under 5 years old with acute respiratory infections in china between 2000 and 2002. J Int Med Res. 2007;35(4):554–563. doi:10.1177/147323000703500416
20. Wang GL, Hua CZ, Yang LH, et al. Clinical characteristics of 84 children with invasive Haemophilus influenzae infection from 2014 to 2018. Zhonghua Er Ke Za Zhi. 2019;57(8):592–596. doi:10.3760/cma.j.issn.0578-1310.2019.08.004
21. Kaczmarek FS, Gootz TD, Dib-Hajj F, Shang W, Hallowell S, Cronan M. Genetic and molecular characterization of β-lactamase-negative ampicillin-resistant Haemophilus influenzae with unusually high resistance to ampicillin. Antimicrob Agents Chemother. 2004;48(5):1630–1639. doi:10.1128/AAC.48.5.1630-1639.2004
22. García-Cobos S, Campos J, Lázaro E, et al. Ampicillin-resistant non-β-lactamase-producing Haemophilus influenzae in Spain: recent emergence of clonal isolates with increased resistance to cefotaxime and cefixime. Antimicrob Agents Chemother. 2007;51(7):2564–2573. doi:10.1128/AAC.00354-07
23. Kostyanev TS, Sechanova LP. Virulence factors and mechanisms of antibiotic resistance of Haemophilus influenzae. Folia Med. 2012;54(1):19–23. doi:10.2478/v10153-011-0073-y
24. Yuan M, Shu JT, Li J, Zhang JY, Kong N. Capsular typing and drug resistance patterns of Haemophilus influenzae strains isolated from children in Yunnan. Chin J Nosocomiology. 2021;31(18):2826–2830.
25. Shuel M, Hoang L, Law DKS, Tsang R. Invasive Haemophilus influenzae in British Columbia: non-hib and non-typeable strains causing disease in children and adults. Int J Infect Dis. 2011;15(3):e167e173. doi:10.1016/j.ijid.2010.10.005
26. Han MS, Jung HJ, Lee HJ, Choi,EH. Increasing prevalence of group III penicillin-binding protein 3 mutations conferring high-level resistance to beta-lactams among nontypeable Haemophilus influenzae isolates from children in Korea. Microb Drug Resist. 2019;25(4):567–576. doi:10.1089/mdr.2018.0342
27. Honda H, Sato T, Shinagawa M, et al. Multiclonal expansion and high prevalence of β-lactamase-negative Haemophilus influenzae with high-level ampicillin resistance in Japan and susceptibility to quinolones. Antimicrob Agents Chemother. 2018;62(9):e00851–00818. doi:10.1128/AAC.00851-18
28. Li -X-X, Xiao S-Z, Gu -F-F, He W-P, Ni Y-X, Han,L-Z. Molecular epidemiology and antimicrobial resistance of Haemophilus influenzae in adult patients in Shanghai, China. Front Public Health. 2020;8:95. doi:10.3389/fpubh.2020.00095
29. Wang XL, Xie J, Guo YB, et al. Lower respiratory tract isolates of non-typeable Haemophilus influenzae in Western Sichuan, China: antimicrobial susceptibility, mechanisms of β-lactam resistance and decade changes. J Glob Antimicrob Resist. 2020;21:324–330. doi:10.1016/j.jgar.2019.10.026
30. Leavis HL, Willems RJL, van Wamel WJB, Schuren FH, Caspers MP, Martien PM. Insertion Sequence–driven diversification creates a globally dispersed emerging multiresistant subspecies of E. faecium. PLoS Pathog. 2007;3(1):e7. doi:10.1371/journal.ppat.0030007
31. Tanaka E, Wajima T, Nakaminami H, Noguchi N. Whole-genome sequence of Haemophilus influenzae ST422 outbreak clone strain 2018-Y40 with low quinolone susceptibility isolated from a paediatric patient. J Glob Antimicrob Resist. 2020;22:759–761. doi:10.1016/j.jgar.2020.06.024
32. Otsuka T, Fujii K, Okazaki M, Komiyama K, Yoshida K, Ishikawa Y. Genotyping of Haemophilus influenzae type b in pre-vaccination era. J Infect Chemother. 2012;18(2):213–218. doi:10.1007/s10156-011-0323-1
33. Tsang RSW, Li A, Mullen A, et al. Laboratory characterization of invasive Haemophilus influenzae isolates from Nunavut, Canada, 2000–2012. Int J Circumpolar Health. 2016;75(1):29798. doi:10.3402/ijch.v75.29798
34. Mironov KO, Tagachenkova TA, Platonov AE, Iakovenko ML, Koroleva IS, Shipulin,GA. Genetic characteristics of Haemophilus influenzae type b isolated from children with bacterial meningitis in Moscow in 2007–2009. Zh Mikrobiol Epidemiol Immunobiol. 2010;4:3–8.
35. Wang FS, Yang TT, Yao PP, Xu BX, Ye JL. Comparison of typing among 4 Haemophilus influenzae isolates with capsule. Chin J Health Lab Technol. 2012;22(12):2892–2894.
36. Moxon ER, Kroll JS. Type b capsular polysaccharide as a virulence factor of Haemophilus influenzae. Vaccine. 1988;6(2):113–115. doi:10.1016/S0264-410X(88)80011-2
37. Cardoso B, Fontana H, Esposito F, et al. Genomic insights of international clones of Haemophilus influenzae causing invasive infections in vaccinated and unvaccinated infants. Microb Pathog. 2021;150:104644. doi:10.1016/j.micpath.2020.104644
38. Ali TR, Kroll JS, Langford,PR. Haemophilus influenzae microarrays: virulence and vaccines. Comp Funct Genomics. 2002;3(4):358–361. doi:10.1002/cfg.194
39. Burroughs M, Cabellos C, Prasad S, Tuomanen E. Bacterial components and the pathophysiology of injury to the blood-brain barrier: does cell wall add to the effects of endotoxin in gram-negative meningitis? J Infect Dis. 1992;Suppl 165(Supplement 1):S82–S85. doi:10.1093/infdis/165-Supplement_1-S82
40. Griffin R, Bayliss CD, Herbert MA, et al. Digalactoside expression in the lipopolysaccharide of Haemophilus influenzae and its role in intravascular survival. Infect Immun. 2005;73(10):7022–7026. doi:10.1128/IAI.73.10.7022-7026.2005
 © 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.