Back to Journals » Drug Design, Development and Therapy » Volume 18
Geraniol Ameliorates Pentylenetetrazol-Induced Epilepsy, Neuroinflammation, and Oxidative Stress via Modulating the GABAergic Tract: In vitro and in vivo studies
Authors Younis NS ![]() , Almostafa MM
, Almostafa MM ![]() , Mohamed ME
, Mohamed ME
Received 20 July 2024
Accepted for publication 27 November 2024
Published 5 December 2024 Volume 2024:18 Pages 5655—5672
DOI https://doi.org/10.2147/DDDT.S481985
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Qiongyu Guo
Nancy S Younis,1,2 Mervt M Almostafa,3 Maged E Mohamed1,4
1Department of Pharmaceutical Sciences, College of Clinical Pharmacy, King Faisal University, Alhofuf, Al-Ahsa, 31982, Saudi Arabia; 2Zagazig University Hospitals, Zagazig University, Zagazig, 44519, Egypt; 3Department of Chemistry, College of Science, King Faisal University, Alhofuf, Al-Ahsa, 31982, Saudi Arabia; 4Pharmacognosy, College of Pharmacy, Zagazig University, Zagazig, 44519, Egypt
Correspondence: Nancy S Younis; Mervt M Almostafa, Email [email protected]; [email protected]
Introduction: Geraniol (Ger), a monoterpene, is a common constituent of several essential oils. This study explored the anticonvulsant effect of Ger in-vitro using nerve growth factor (NGF) prompted PC12 cell injured by Glutamate (Glu) and in-vivo using Pentylenetetrazole (PTZ)-induced kindling through the GABAergic pathway.
Materials: To assess the effect of Ger on NGF prompted PC12 cells injured by Glu, Ger at concentrations of 25, 50, 100, 200 and 400 μg/mL was used. GABA, 5-HT, IL-1β, IL-4, and TNF-α levels and the gene expressions of GABAA-Rα 1, NMDAR1, GAD 65, GAD 67, GAT 1 and GAT 3 were measured in NGF-induced PC12 cells treated with Ger (100, and 200 μg/mL). Mice were randomly separated into five groups. Normal and PTZ groups in which mice were injected with saline or PTZ, respectively. PTZ + Ger 100, PTZ + Ger 200 and PTZ + SV groups in which mice orally administered Ger or sodium valproate (SV), respectively, then injected with PTZ.
Results: Ger up to 400 μg/mL did not display any toxicity or injury in PC12 cells. Ger (100 to 200 μg/mL) reduced the injury induced by Glu, increased the gene expression of GABAA-Rα 1, GAD65 and GAD67 and decreased GAT 1, GAT 3 and NMDAR1 expression in NGF-induced PC12 cells damaged by Glu. Ger (100 to 200 μg/mL) increased GABA and reduced TNF-α, IL-4 and IL-1β levels in NGF-induced PC12 cells injured by Glu. As for the in-vivo results, Ger increased GABA, GAD, GAT 1 and 3 and lowered GABA T. Ger mitigated MDA, NO, IL-1β, IL-6, TNF-α and IFN-γ, GFAP, caspase-3, and − 9 levels and Bax gene expression and escalated GSH, SOD, catalase, BDNF and Bcl2 gene expression.
Conclusion: Ger reduced the oxidative stress status, neuroinflammation and apoptosis and activated GABAergic neurotransmission, which might clarify its anticonvulsant. Ger protects animals against PTZ prompted kindling as established by the enhancement in short term as well as long-term memory. Ger mitigated the injury induced by Glu in NGF prompted PC12 cell.
Keywords: geraniol, pentylenetetrazole, GABAergic, oxidative stress, neuroinflammation, apoptosis
Introduction
Geraniol (Ger) is a common ingredient of numerous essential oils such as ninde oil, rose oil, palmarosa oil and citronella oil.1 Ger has a distinguished rose-like odour and taste and is thus extensively utilised as a fragrance material in cosmetic and household products.2 Pavan et al3 performed a study to identify Ger’s pharmacokinetics and bioavailability and concluded that Ger has high bioavailability and is able to permeate the central nervous system following its oral administration, reaching detectable amounts in the Cerebrospinal fluid (CSF). Ger exhibits various pharmacological properties including antimicrobial,4 anti-oxidant, anticancer, anti-inflammatory5 proposing this monoterpene alcohol as a promising drug candidate. Ger exhibited neuroprotective effects in diverse neurological conditions including spinal cord injury (SCI).6 Also, Ger improves functional outcomes in cerebral ischemia/reperfusion (I/R),7 diminished the neuropathic pain after spinal cord injury (SCI)6 and attenuated behavioral and neurochemical impairments in animals exposed single-prolonged-stress (SPS) provoked post-traumatic disorder (PTSD).8 Also, Ger exerted an antidepressant action in chronic unpredictable mild stress (CUMS),9 protected against rotenone-induced PD.10
Epilepsy, a common neurological disorder, is a recurrent seizure caused by an abnormal discharge of cerebral neurons. At present, the existing antiepileptic medications have inadequate efficiency and are associated with several their adverse effects that limit their use. Thus, a key objective in epilepsy investigation is to identify new medications with greater efficacy as well as fewer toxicities.11 The disproportion between excitatory neurotransmitters (Glutamate (Glu)) and inhibitory neurotransmitters (γ-aminobutyric acid (GABA)) is a leading element in epilepsy. Multiple studies demonstrated the enhancement of endogenous GABA signaling as well as exogenous GABA delivered to the brain,12,13 which could regulate and restore the excitation/inhibition imbalance to prevent seizures in the epileptic brain.14
GABA signaling includes GABA receptors, glutamic acid decarboxylase (GADs) and GABA-transporters (GATs) mainly. Stimulation of GABA receptor leads to the opening of Cl channel, causing Cl− current to enter rapidly, leading to the hyperpolarization consequently preventing the over discharge. GABA receptors are allocated into GABAA, GABAB and GABAC, from which GABAA is utmost closely correlated to epilepsy.15 GADs, motivate synthesize of GABA, are encoded by two genes, GAD2 (GAD 65) and GAD1 (GAD 67).16 GATs specifically, GAT 1 and GAT 3 are expressed in nerve terminals and glia to remove GABA from the synaptic cleft.17
PTZ is regularly utilized to identify the neurobiology associated with epilepsy as well as to estimate the efficiency of novel treatments. Additionally, PC12 cells (pheochromocytoma cells) are generally considered as an in-vitro model for neurological diseases, including epilepsy.18 PC12 neuron-like cells promoted by nerve growth factor (NGF), which promote neuronal differentiation, and regeneration, are frequently used for examining nerve cell differentiation.19 Glu mediates excitatory neurotransmission and is involved in neural development and synaptic plasticity.20 Therefore, NGF-induced PC12 cells injured by Glu were chosen to explore neurological conditions in-vitro, such as epilepsy.18 NMDAR1, Glu receptor, is closely correlated with epilepsy, and over stimulation of NMDAR receptor is associated with epilepsy pathogenesis.
The current study explored the anticonvulsant effect of Ger in-vitro using NGF-prompted PC12 cell injured by Glu and in-vivo using PTZ-induced seizures in mice through considering the GABAergic pathway.
Materials and Methods
In-vitro Studies
Cell Culture
Pheochromocytoma-derived PC12 cells were provided by the cell culture unit of Holding Company for Biological Products and Vaccines (VACSERA), Egypt and maintained in RPMI medium mented with 5% horse serum, 5% FBS and 1% penicillin/streptomycin. PC12 cells were incubated at 37 °C in 5% CO2 atmosphere. PC12 cells were treated with 50 ng/mL of NGF for 48 h for cell differentiation.21 NGF-induced PC12 cells without both Ger and Glu were considered as normal cells, whereas NG provoked PC12 cells injured by Glu (20 mmol/L) were considered as model cells.
Cell viability assay
To estimate Geraniol (Ger) alone action on cell viability, Ger at different concentrations including 25, 50, 100, 200, and 400 μg/mL were added to cells, which were inoculated in 96-well plates for 24 h, and cultured at 37 °C for 24 h in 5% CO2. Subsequently, ten μL of CCK-8 solution were introduced to each well and maintained at 37 °C for 1 h then the absorbance was assessed at 450 nm.
To assess the effect of Ger on NGF prompted PC12 cells injured by Glu viabilities, Ger at concentrations of 25, 50, 100, 200 and 400 μg/mL for were added and cultured at 24 h at 37 °C. Subsequently, subjected to Glu (20 mmol/L) for another day to establish injure based upon the preliminary studies and previous studies.22 Then, ten μL of CCK-8 solution were added and maintained at 37 °C for 1 h followed by measuring the absorbance at 450 nm.
Assessment of GABA, 5-hydroxytryptamine receptors or serotonin (5-HT), interleukin-1 beta (IL-1β), interleukin-4 (IL-4), and Tumor necrosis factor (TNF-α) content in PC12 Cells.
NGF-induced PC12 cells were inoculated for 24 h, then Ger (100, and 200 μg/mL) was introduced and cultured for another day followed by Glu, which was cultured for 24 h. The media were collected, centrifuged, and kept at 80 °C. GABA, 5-HT, IL-1β, IL-4, and TNF-α levels were measured by ELISA kits.
Quantitative Chain Real-Time Polymerase Reaction (RT-PCR) Assay
Total RNA of the NGF-induced PC12 cells was extracted following the producer’s directions. Two μg RNA was reversely transcribed into cDNA using the Revert Aid First Strand cDNA Synthesis Kit. RT-PCR was performed using an ABI Step OnePlus System. The sequence of primers used is shown in Table 1. The gene expressions of GABAA-Rα1, NMDAR1, GAD 65, GAD 67, GAT 1 and GAT 3 were standardized to β-actin and analyzed by using 2−ΔΔCT method.
 |
Table 1 The Sequence of Primers Used |
In-vivo Studies: PTZ Prompted Kindling Model
Animals Attaining and Ethics Permission
Male Swiss albino mice (25 to 35 g) were secured from the Animal Research Centre, King Saud University, Riyadh, KSA and were continued with standard food and water in ventilated cages throughout the experiment. The Institutional Animal Care and Use Committee of King Faisal University endorsed the study procedure (KFU-REC-2024-JUN-ETHICS1883).
Experimental Plan
Forty Mice were haphazardly clustered into five groups (n = 8). Normal group in which mice were given saline (0.9%). PTZ group in which mice administered PTZ (35 mg/kg,14 ip injections, every other day). PTZ + Ger 100, PTZ + Ger 200 and PTZ + SV groups in which mice were administered Ger (100 and 200 mg/kg)6 and sodium valproate (SV, 300 mg/kg) orally, respectively, and injected with PTZ.23 Mice were monitored for half an hour after PTZ injection to evaluate the seizure following Racine score.24 Mice displayed with stage 5 seizures on two continual PTZ doses were assumed to be entirely kindled and used in the behavioral tests as illustrated in Figure 1.
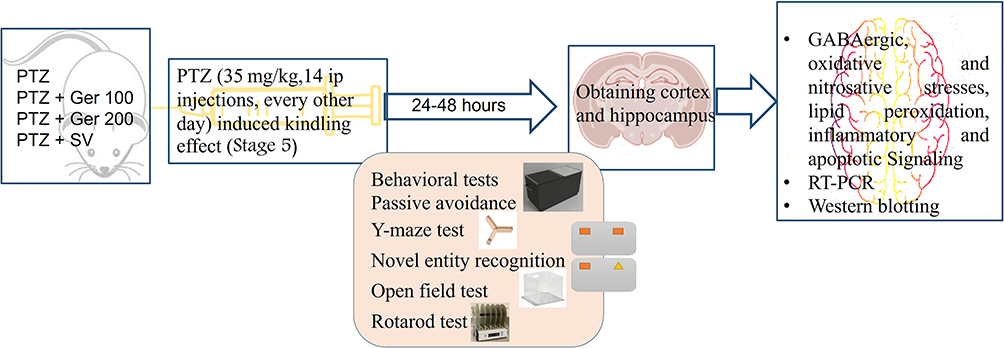 |
Figure 1 Experimental design for chronic administration of PTZ experiment. |
Behavioral Assessments
Passive Avoidance (Step-Through)
Two-compartment avoidance device was used. The sessions occurred after 1 and 24 h of animals training to evaluate the animals short-term as well as long-term memory.25 On the behavioral test first day, animals were positioned in the white start compartment. After 5 second, the sliding door was raised to allow mice to go into the darkened chamber. Then, the door was closed for 20s, and the mice were kept there for habituation. After 30 minutes, the acquisition trial was conducted. Each mice was located in the light partition; after 5 s, the door was opened. As soon as the animal entered the dark chamber, the door was locked, and the foot shock (0.5 mA for 2 s) was provided. After twenty second, the mice were located in a temporary cage for attainment training. This test was recurred after two minutes. Mice had a foot shock each time it walked into the dark compartment with all its four paws. When the animal was kept in the light room for two minutes sequentially, the learning training was completed. All the animals learned the task with 2 maximum trials in the control group. On the second day, long-term memory was evaluated by a recovery test. Each mice was positioned in the illuminated compartment for twenty second, subsequently the door was opened, and the step-through latency was documented (up to 400 s) with no electric shocks were delivered to mice throughout that time. Mice suffering from cognitive deficits tends to enter the aversive stimuli zone.
Y-Maze Test
Y-maze test appraised the spatial short-term memory in PTZ tested animals as mentioned before26 in which arms entries no and the unplanned fluctuations were calculated. The device comprised three identical arms (A, B, C) arranged at 120° angles to form a Y-shape. On the last day of the study, 1 hour after different treatment administrations, mice were positioned separately in one of the three starting arms for 8 min for exploration. The number of arms entries and the spontaneous alternations were assessed. Spontaneous alternation is the repeated entrance into the 3 different Y-maze arms in any sequence. The spontaneous alternation percentage was assessed following this equation: 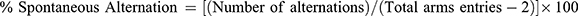 .26
.26
Novel Entity Recognition
Novel object recognition was applied to appraise mice’s recognition memory as stated before.27 This test was accomplished using an open field with 40 cm length × 40 cm width × 25 cm high. The day before the last day of the experiment, mice were placed separately in the open field arena for five minutes for place familiarization. After 1 h of the last PTZ dose and for two days, both the acquisition and retention phases were performed. In the retention part, two identical objects (A and A) were offered to each mice for five minutes as an exploration session. Exploration is considered when the animal touches the object or point its nose less than 2 cm towards the object. On the subsequent day, one of the objects that was displayed throughout the acquisition phase was exchanged into a new entity (B) and each mice was permitted to inspect the two items (A+B) for five minutes as the day before. The time needed by each animal to recognize entity A or entity B was noted as tA and tB, respectively. Lastly, discrimination index (DI) was examined 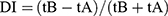 .
.
Open Field Test
Animals were located in the open field test (dimension: 40 cm length × 40 cm width × 45 cm high; divided into 16 smaller squares of equal (10 cm × 10 cm)) to assess the exploratory behavior, motor coordination, and locomotor ability.28 One-hour subsequent to the novel object recognition test, animals were submitted to the open field test. The number of lines crossed, rearing, grooming, and the time consumed in the center were documented.
Rotarod Test
The motor coordination was estimated via the rotating rod to conclude the neuroprotective actions of Ger in PTZ administered animals as stated before.29 One hour after the open field test, each mice was placed on the rotating bar of the rotarod apparatus with a speed of 16 revolutions/minutes for 60 second to evaluate animal stability, which indicates the motor coordination of mice.
Biochemical Evaluations
Subsequent to the behavioral investigations, animals were sacrificed to separate the brains, which were washed with saline. Prefrontal cortex and hippocampus regions were obtained.
Hippocampal Histopathology
Four μm hippocampal sections were obtained and stained by hematoxylin and eosin (H&E). Hippocampal sections were randomly chosen, photographed, appraised. The morphological changes in the neurons, such as cytoplasmic vacuolation, nuclear chromatin clumping, condensed cytoplasm, and fragmentation of cells, were examined by an independent pathologist to clarify the neuronal damage and scored as follows normal or no injury = 0; rare neuronal injury (< 5 clusters) = 1; occasional neuronal injury (5–15 clusters) = 2; frequent neuronal injury (> 15 clusters) = 3; diffused neuronal injury = 4 as mentioned before.30
Assessment of GABAergic Neurotransmitters
GABA content (μg/g of tissue),31 GAD and γ-aminobutyric acid transaminase (GABA-T) (both expressed as μg/min/mg of tissue) were assessed within the cortex and hippocampus homogenates.
Assessment of Oxidative and Nitrosative Stresses and Lipid Peroxidation
Glutathione (GSH, Cat. No. ab239727) content (mol/g of tissue), Catalase (CAT, Cat. No. ab118184) (mmol H2O2/min/g tissue) and superoxide dismutase (SOD, Cat. No. ab285309) activities and Malondialdehyde (MDA, Cat. No. ab238537) level, stated as µmol/g of tissue, were assessed using the specified ELISA kits obtained from Abcam Co., Eugene, OR, USA.
In addition, NO, which induces various harmful situations known as nitrosative stress, was assessed using the Griess reagent.32
Assessment of Inflammatory Mediators
Tumor necrosis factor-alpha (TNF-α, Cat. No. ab46070), interleukin-1β (IL-1β, Cat. No. ab100768), interferon-gamma (INF-γ, Cat. No. ab239425), and interleukin-6 (IL-6, Cat. No. ab100772), were calculated consuming specific ELISA kits attained from Abcam Co., USA.
Assessment of Glial Fibrillary Acidic Protein (GFAP) and Brain-Derived Neurotrophic Factor (BDNF)
GFAP (Cat. No. ab233621), brain astrocyte activation indicator, and BDNF (Cat. No. ab213899) were analyzed using their relevant ELISA kits, which were secured from Abcam Co., USA.
Hippocampal mRNA Expression Using RT-PCR
The hippocampus obtained from different groups was instantly frozen and stored to be designed for total RNA extraction, as mentioned before.33 The relative changes in gene expression were accomplished using the 2 - ΔΔCT method.34 The primers used are as follows GAT-1 (NM_178703.4) F: 5’-GAAAGCTGTCTGATTCT GAGGTG-3’, R: 3’-AGCAAACGAT GATGGAGTCCC-5’; GAT-3 (NM_172890.3) F: 5’- ATTATGTCT CCTGGCGGCCT3’, R: 3’- AAAAGATCTGCGTCCCAGCG-5’; Bcl-2 F: 5’-GTGGATGACTGAGTACC T −3′, R: 5′- CCAGGAGAAATCAAACAGAG −3′, Bax: F: 5’- CTACAGGGTTTCATCCAG −3′, R- 5′- CCA GTTCATCTCCAATTCG −3′; β actin F: 5’-ACGGCCAGGTCATCACTATTG-3’, R: 3’-CAAGAAGGAAG GCTGGAAAAGA.
Hippocampal Protein Expression by Western Blotting
Protein was separated via SDS PAGE and blotted onto PVDF membranes, which were blocked, washed with TBS, incubated with the primary antibodies (1:2000 dilution) for 2 hours, followed by the secondary antibody (goat anti-rabbit HRP-conjugated, 1:5.000 dilution). The chemiluminescence developed from the luminol reagent was detected with the C-DiGit chemiluminescence scanner, and the band intensity was analyzed using the scanner software.
Assessment of Apoptotic Signaling
Caspase- 3 (Cat. No. ab39401 and caspase 9 (Cat. No. ab65608) were evaluated in hippocampus homogenates using ELISA kits acquired from Abcam Co., USA.
Statistical Analysis
Numbers are revealed as mean ± SD. Graph Pad software was utilized to perform one-way ANOVA then Tukey–Kramer as a post hoc test for multiple comparisons using 0.05 level of probability as a significance level.
Results
In-vitro Outcomes
Outcome of Different Concentrations of Ger on NGF-Induced PC12 Cells Viability
Ger at different concentrations up to 400 μg/mL did not display any toxicity or injury in PC12 cells as displayed in Figure 2A. Ger diminished the injury induced by Glu from concentrations of 100 to 500 μg/mL, when compared to the model cells (P < 0.001) indicating that Ger had a possible neuroprotective action against the Glu prompted damage in NGF-induced PC12 cells (Figure 2B). In subsequent experiments, the current study focused on these two efficient doses of Ger which are 100 and 200 μg/mL.
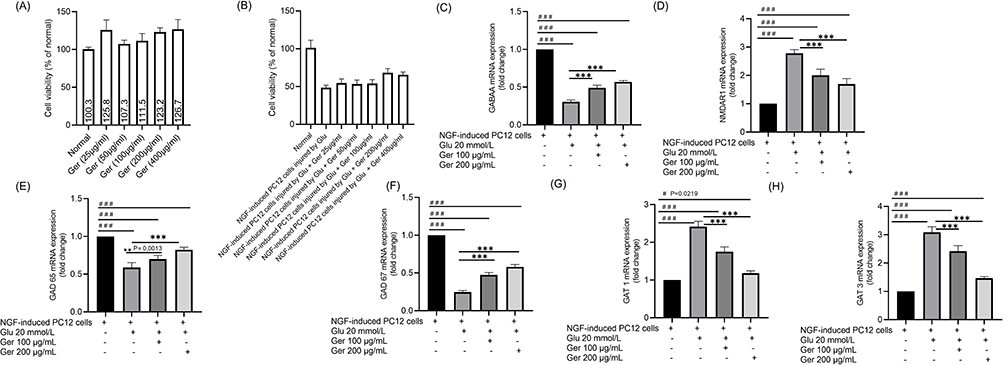 |
Figure 2 The outcome of Ger at different concentrations on the (A) normal cells viability (B) NGF-induced PC12 cells injured by Glu (20 mmol/L) viability. Effects of Ger (100 and 200 μg/mL) on the gene expressions of (C) GABAA-Rα1, (D) NMDAR1, (E) GAD 65, (F) GAD 67, (G) GAT 1 and (H) GAT 3 in NGF prompted PC12 cells damage by Glu. PC12 cells (induced by 50 ng/mL NGF for 48 h) were treated with Ger (100 or 200 μg/mL) for 24 h, then Glu (20 mmol/L) for another 24 h. Normal cells are NGF-induced PC12 cells without Ger and Glu. #P≤0.0219 vs normal cells ###P≤ 0.001 vs normal cells; **P≤ 0.0013 vs model cells; ***P≤ 0.001 vs model cells. |
Influences of Ger on Gene Expressions of GABA Signaling Molecules in NGF Prompted PC12 Cells Damaged by Glu
To scrutinize the shielding action of Ger on NGF prompted PC12 cells damaged by Glu, gene expressions of GABAA-Rα1, NMDAR1, GAD 65, GAT 1, GAT 3 and GAD 67 were identified. The model cells showed elevated gene expressions of NMDAR1, GAT 1, and GAT 3 as well as diminished GABAA-Rα1, GAD 65 and GAD 67 expression (P < 0.001). On the other hand, Ger (100 and 200 μg/mL) considerably amplified GABAA-Rα1, GAD65 and GAD67 and decreased GAT 1, GAT 3 and NMDAR1 expressions relative to the model cells (Figure 2C–H).
Influences of Ger on GABA, 5-HT, IL-1β, IL-4 and TNF-α Levels in NGF Prompted PC12 Cells Damaged by Glu
As revealed in Figure 3, GABA and 5-HT levels were substantially declined (P < 0.001) in model cells. In Ger-treated cells (100 and 200 μg/mL), GABA level was clearly amplified compared with the model cells. As for 5-HT, only Ger (200 μg/mL) instigated significant elevation related to the model cells. IL-1β, IL-4 and TNF-α were augmented in model cells, while Ger (100 and 200 μg/mL) considerably reduced them (P < 0001).
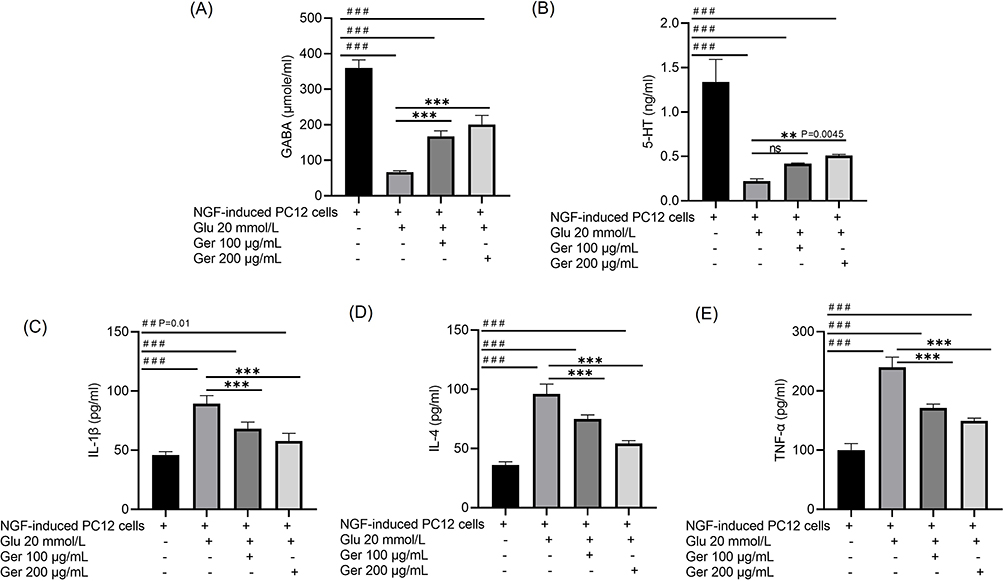 |
Figure 3 Outcomes of Ger on (A) GABA, (B) 5-HT, (C) IL-1β, (D) IL-4 and (E) TNF-α in model cells. PC12 cells (induced by 50 ng/mL NGF for 48 h) were treated with Ger (100 or 200 μg/mL) for 24 h, then Glu (20 mmol/L) for another 24 h. ##P≤0.01 vs normal cells; ###P≤ 0.001 vs normal cells; ***P≤ 0.001 vs model cells. Normal cells are NGF prompted cells without Ger and Glu; Model cells are NGF prompted cells damaged by Glu (20 mmol/L). |
In vivo Results
Behavioral Assessments
Assessment of Ger Effects on PTZ Kindling-Induced Seizures
No seizures were witnessed in the control group. Kindling (score 5/5) was attained after repetitive PTZ injections (35 mg/kg, 14 ip injections, every other day) showing Racine score of 5. Ger administration (100 or 200 mg/kg) with PTZ injections suppressed the development of kindling considerably (P < 0.001) as established by the reduction in seizure scores in correlation with the PTZ alone (Figure 4A).
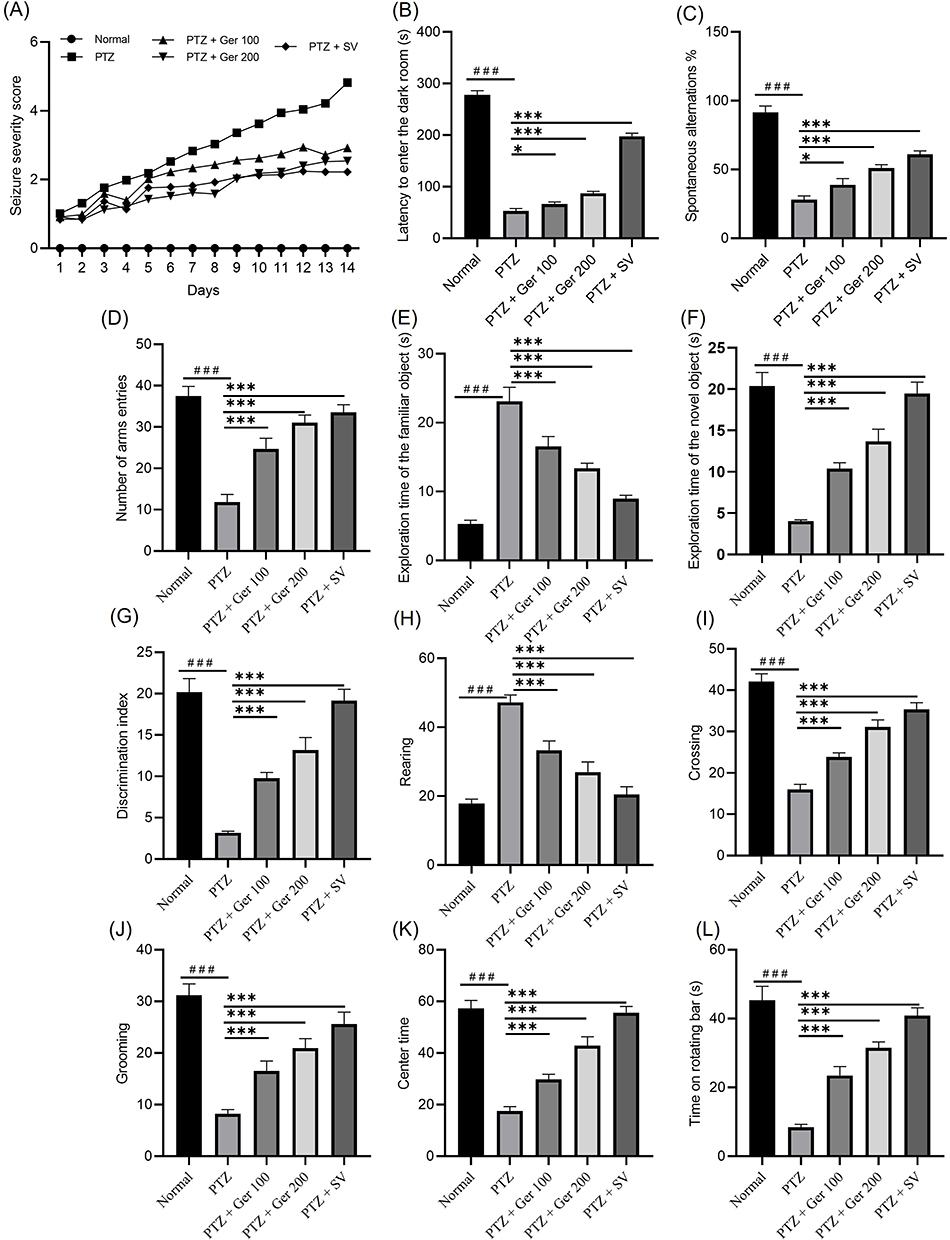 |
Figure 4 The outcomes of Ger (100 and 200 mg/kg) and SV (300 mg/kg) administration in PTZ (35 mg/kg, 14 ip injections, every other day) kindled mice on different behavioral assessments including (A) Racine seizures scale, (B) latency to enter the dark room, (C) % of spontaneous alternations, (D) number of arms entries, (E) exploration time of the familiar object, (F) exploration time of the novel object (G) discrimination index, (H) rearing, (I) crossing, (J) grooming (K) center time and (L) time on the rotating bar. Data are expressed as mean ± SD (n=6); ###P<0.001 compared to the normal group, *P<0.002 compared to the PTZ group, P<0.001 compared to the PTZ group. |
Assessment of Ger Effects on Passive Avoidance and Y-Maze Tests
Repetitive seizures were associated with significant comorbidities, such as, learning and memory insufficiency. Kindling prompted by PTZ (35 mg/kg, 14 ip injections, every other day) drastically declined memory retrieval, which can be assessed by passive avoidance and Y maze tests. In passive avoidance tests, the latency to go in the dark room was considerably reduced in full-kindled animals, whereas Ger administration improved the latency to enter the dark room compared to the PTZ (P < 0.001) (Figure 4B). In the Y maze, kindling induction by PTZ considerably lessened the % of spontaneous alternations, reaching 28.17± 2.64% from 91.5 ± 4.73% in the normal group. Ger and SV increased spontaneous alternations percentage, reaching 38.98 ± 4.45%, 51.18 ± 2.35%, and 61 ± 2.56%, respectively, when related with the PTZ, as illustrated in Figure 4C. Similarly, arms entries escalated from 11.81 ± 1.85 in the PTZ administered animals to 24.73 ± 2.52, 31.06 ± 1.84, and 33.56 ± 1.82 in mice administered Ger and SV, respectively (Figure 4D). These outcomes suggest that Ger modified the memory impairments occurring in PTZ prompted kindling.
Assessment of Ger Effects in the Novel Object Recognition Test
PTZ amplified the exploration time of the familiar entity and diminished the exploration time of the novel entity as well as DI, as presented in Figure 4E and F. On the other hand, Ger considerably lowered the exploration time of the familiar entity and augmented the time of the novel entity and DI (Figure 4G).
Assessment of Ger Effects on the Open Field and the Rotarod Tests of PTZ-Kindled Animals
PTZ amplified the number of rearing and reduced the number of crossing and grooming and the center time in the open field, as depicted in Figure 4H and K. Whereas Ger-administered mice showed lowered number of rearing as well as increased number of crossings, grooming, and the center time in the open field. In addition, the period consumed on the rotating bar was considerably diminished in the PTZ-administered animals. An effect that was inverted by Ger and SV (Figure 4L).
Assessment of Ger Effects on the GABAergic Pathway
As GAD is one of the chief regulators in GABA synthesis and GAT-1 and GAT-3 are responsible for the regulation of extracellular GABA levels.35 Therefore, in the current study, GABA, GAD and GABA T were evaluated in hippocampus and cortex as well as GAT 1 and GAT 3 gene and protein expression were measured (Figure 5). In PTZ induced kindled animals, GABA and GAD were suppressed reaching a reduction percentage of 61.30% and 64.13% for GABA and in 70% and 62.97% for GAD in both hippocampus and cortex, respectively, whereas GABA T was increased in both hippocampus and cortex. These outcomes indicate that PTZ is associated with lowered GABA synthesis and availability in synapse. On the contrary, Ger significantly restored GABA level and GAD activity and lowered GABA T in both hippocampus and cortex as presented in Figure 5.
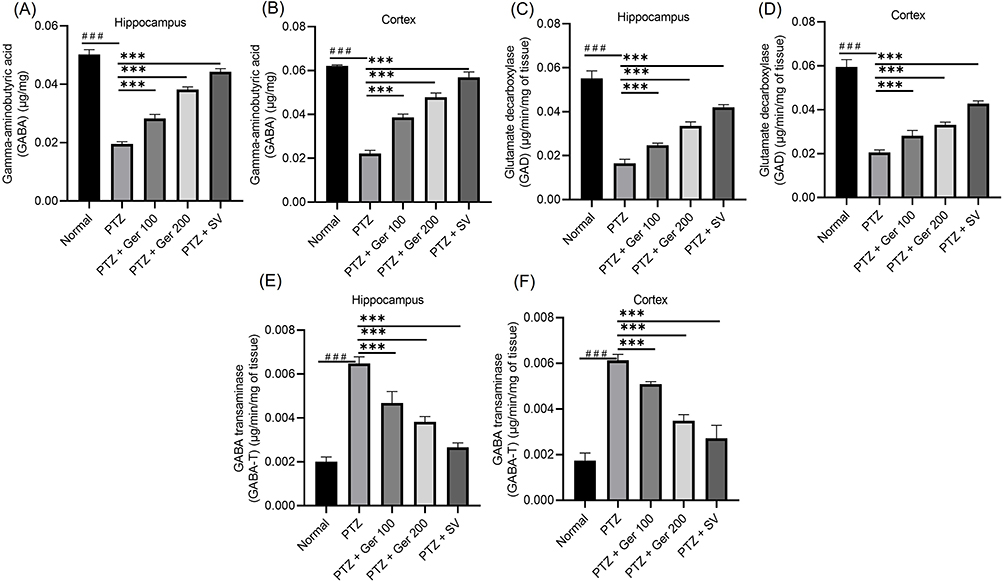 |
Figure 5 The outcomes of Ger (100 and 200 mg/kg) and SV (300 mg/kg) administration in PTZ (35 mg/kg, 14 ip injections, every other day) kindled mice on GABAergic pathway specifically (A and B) GABA, (C and D) GAD and (E and F) GABA T levels in the hippocampus and cortex respectively. Data are expressed as mean ± SD (n=6); ###compared to normal group (P<0.001), ***Compared to PTZ group (P<0.001) using One-Way ANOVA tailed by Tukey’s post hoc test. |
Additionally, PTZ induced kindled animals, GAT 1 and 3 gene and protein expression were suppressed, whereas Ger significantly increased GAT 1 and 3 gene and protein expression as presented in Figure 6.
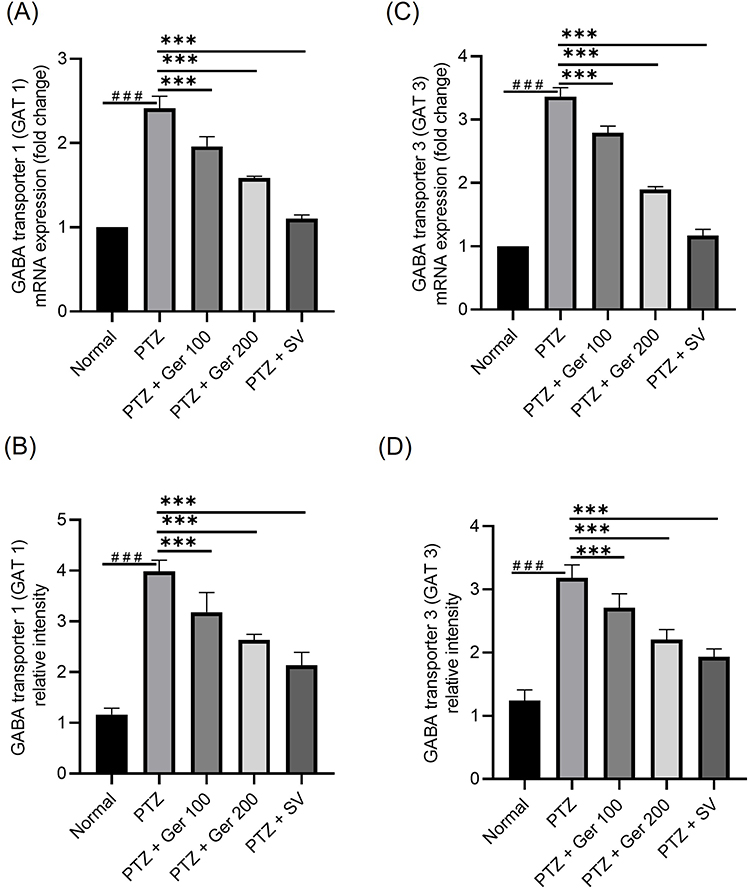 |
Figure 6 The outcomes of Ger (100 and 200 mg/kg) and SV (300 mg/kg) administration in PTZ (35 mg/kg, 14 ip injections, every other day) kindled mice on GABAergic pathway specifically on the gene and protein expression (A and B) GAT 1, and (C and D) GAT 3 in the hippocampus. Data are expressed as mean ± SD (n=6); ###compared to normal group (P<0.001), ***Compared to PTZ group (P<0.001) using One-Way ANOVA tailed by Tukey’s post hoc test. |
Assessment of Ger Effects on Oxidative/Nitrosative Stress and Lipid Peroxidation in PTZ-Kindled Animals
PTZ induced a considerable escalation in the MDA level. However, management with Ger (100 and 200 mg/kg) lessened MDA when linked with PTZ-group (Figure 7A and B). Likewise, NO levels were extensively amplified in PTZ. However, Ger (100 and 200 mg/kg) and SV exhibited a substantial reduction in NO, causing a % reduction of 22.6%, 51.78%, and 60% in the hippocampus and 20.65%, 42.63% and 57.8% in the cortex, respectively (Figure 7C and D).
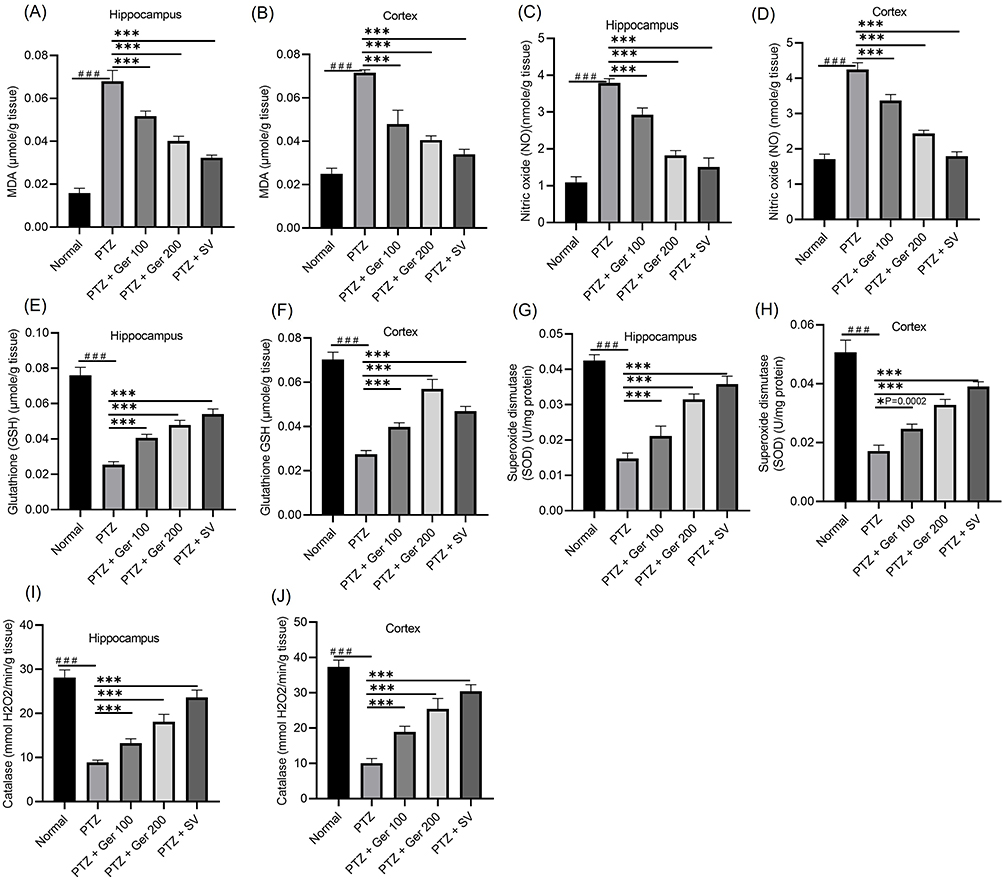 |
Figure 7 The outcomes of Ger (100 and 200 mg/kg) and SV (300 mg/kg) administration in PTZ (35 mg/kg, 14 ip injections, every other day) kindled mice on lipid peroxidation and oxidative/nitrosative stress specifically on (A and B) MDA, (C and D) NO, (E and F) GSH, (G and H) SOD and (I and J) Catalase in the hippocampus and cortex respectively. Data are expressed as mean ± SD (n=6); ###Compared to normal group (P<0.001), *Compared to PTZ group (P<0.0002), ***Compared to PTZ group (P<0.001) using One-Way ANOVA tailed by Tukey’s post hoc test. |
Oral administration of Ger reversed the GSH insufficiency produced by PTZ, as illustrated in Figure 7E and F. PTZ produced a % decline of 66.45% and 60.9% in GSH in hippocampus and cortex respectively. Whereas Ger (100 and 200mg) amplified GSH in the hippocampus and cortex, when compared to PTZ. In the same way, PTZ prompted a % deficiency in the SOD and catalase activities in the hippocampus and cortex when compared with normal animals, while Ger (100 and 200 mg/kg) augmented SOD catalase activities (Figure 7).
Assessment of Ger Effects on Inflammatory Markers in PTZ-Kindled Animals
PTZ alone administered to mice experienced a boost in IL-1ß. However, Ger (100 and 200 mg/kg) exhibited an attenuation in IL-1β, attaining a percentage decline of 12.70% and 32.3% in the hippocampus and 21.47% and 34.53% in the cortex, respectively (Figure 8). Whereas Ger (100 and 200 mg/kg) administration significantly lowered IL-6 levels.
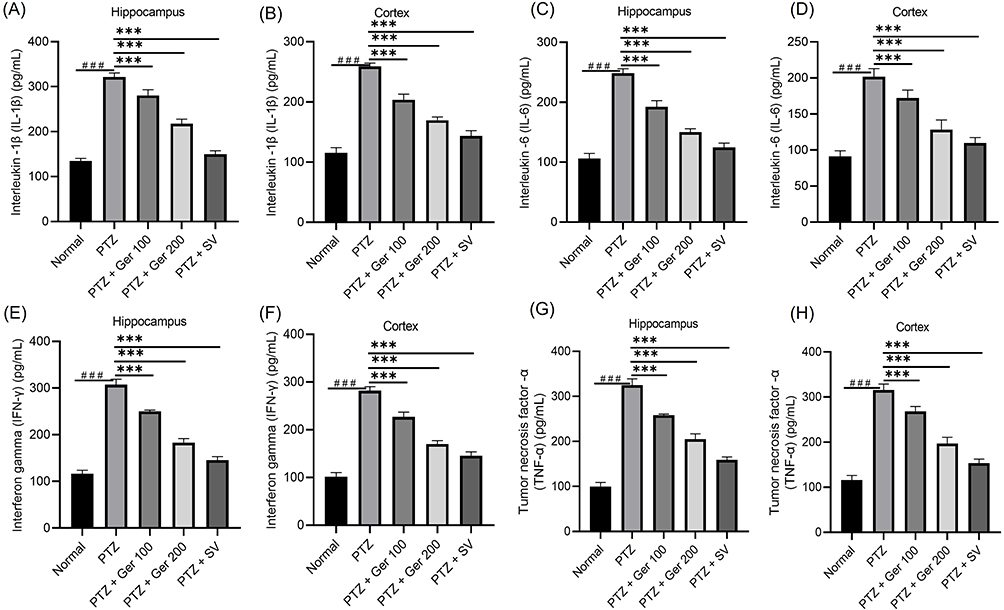 |
Figure 8 The outcomes of Ger (100 and 200 mg/kg) and SV (300 mg/kg) administration in PTZ (35 mg/kg, 14 ip injections, every other day) kindled mice on inflammatory markers, specifically (A and B) IL- 1β, (C and D) IL-6, (E and F) IFN-γ and (G and H)TNF-α levels in the hippocampus and cortex respectively. Data are expressed as mean ± SD (n=6); ###Compared to normal group. (P<0.001), ***Compared to PTZ group (P<0.001) using One-Way ANOVA tailed by Tukey’s post hoc test. |
In the same way, TNF-α level was augmented by PTZ application alone when compared to the normal animals. PTZ administered mice that were treated with Ger (100 and 200 mg/kg) exhibited a percentage decline of 20.49% and 36.92% (P < 0.001) in the hippocampus and 14.97%, and 37.53% (P < 0.05) in cortex, respectively, when related with PTZ alone group.
Alike, PTZ animals displayed an elevation in IFN-γ in the hippocampus from 116.03 ± 7.67 pg/mL in normal group to 307.08 ± 11.9 pg/mL in PTZ given mice and in the cortex from 101.54 ± 8.57 pg/mL in normal animals to 281.75 ± 8.46 pg/mL in the PTZ group. On the other hand, Ger (100 and 200 mg/kg) exhibited a percentage decline of 18.55% and 40.31% (P < 0.001) in the hippocampus and 19.54%, and 39.56% (P < 0.001) in cortex, respectively.
Assessment of Ger Effects on Brain Astrocytes and Microglia Biomarkers Levels in PTZ-Kindled Animals
GFAP and BDNF levels were presented in Figure 9A and B. PTZ- group exposed a noteworthy upsurge in GFAP levels, indicating astrocyte triggering, and a diminishing in BDNF protein levels. On the contrary, Ger (100 and 200 mg/kg) administration significantly lowered GFAP by 16.63% and 30.54%, respectively, and increased BDNF levels by 46.55% and 86.02%, respectively, in the hippocampus.
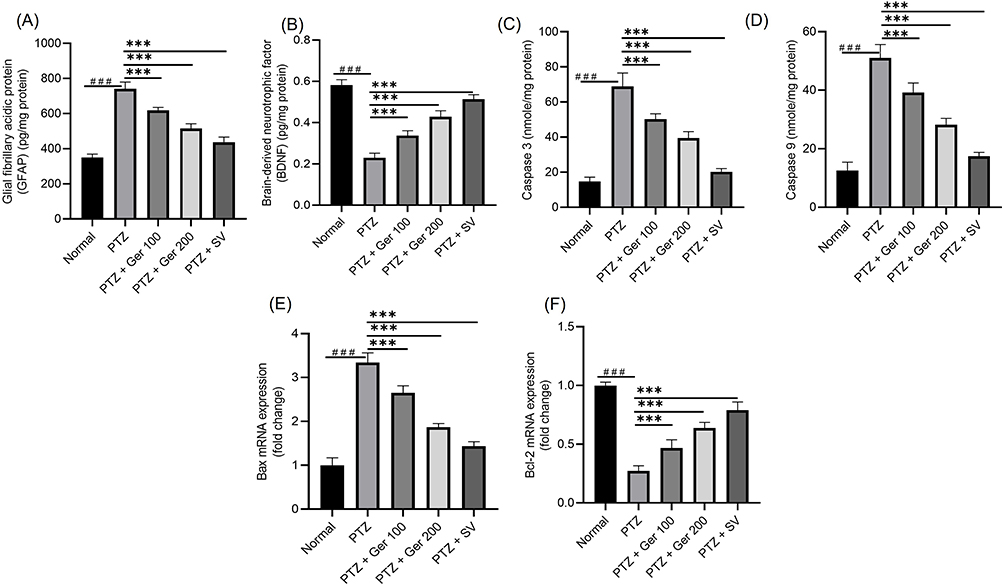 |
Figure 9 The outcomes of Ger (100 and 200 mg/kg) and SV (300 mg/kg) administration in PTZ (35 mg/kg, 14 ip injections, every other day) kindled mice on (A) GFAP and (B) BDNF levels and on apoptotic markers specifically (C) caspase 3, (D) caspase 9, the gene expression of (E) BAX, and (F) Bcl2 levels in the hippocampus. Data are expressed as mean ± SD (n=6); ###Compared to normal group (P<0.001), ***Compared to PTZ group (P<0.001) using One-Way ANOVA tailed by Tukey’s post hoc test. |
Assessment of Ger Effects on Apoptotic Signaling Markers in PTZ-Kindled Animals
PTZ increased caspase-3 and −9 levels and Bax gene expression and diminished Bcl2 gene expression (Figure 9). Alternatively, Ger (100 and 200 mg/kg) significantly reduced caspase-3 and −9 levels and Bax gene expression and escalated Bcl2 gene expression.
Assessment of Ger Effects on Histopathological Alteration
The hippocampal CA1 region in PTZ kindling mice displayed fragmented, contracted, and pyknotic neurons complemented with perineuronal edema, which presented apparent regular hippocampal layers. Ger-administered animals exhibited inferior neural degenerative as well as intercellular edema with a decline in the scoring when compared to the PTZ group (Figure 10).
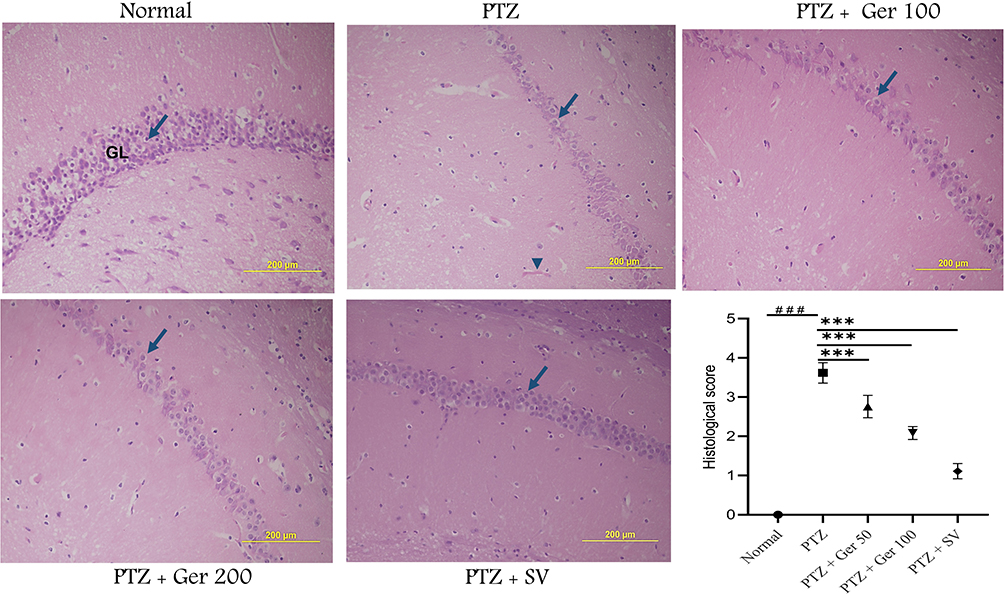 |
Figure 10 The action of Ger on the histopathological alteration of hippocampus stained with H&E stain showing the GL includes 4–6 cell layers (long arrow), PTZ showing apparent reduction in the number of GL cell layers with swelling cells (long arrow) and dilated blood vessel (arrow head), Ger (100 and 200 mg/kg) and SV (300 mg/kg) administration with PTZ revealed amendment in the hippocampus structure, with a noticeable increase in the number of GL cell layers and normally sized blood vessels, and histopathological alteration scoring. ###Compared to normal group (P<0.001), ***Compared to PTZ group (P<0.001). |
Discussion
Several plant extracts that contain Ger were assessed for anticonvulsant activity such as Cymbopogon winterianus Jowitt (Poaceae) leaf essential oil36 and Ocimum basilicum leaf essential oil,37 hydro-alcoholic extract of R. damascena and Cymbopogon citratus Stapf leaves essential oil and hydroalcoholic extract in zebra fish.38 However, robust is known about the anticonvulsant effect of Ger alone. Therefore, the current study tried to scrutinize the antiepileptic effect of Ger using both in-vitro via NGF-prompted PC12 cell injured by Glu and in-vivo models via PTZ-induced seizures in mice through with regard to the GABAergic pathway.
The neuron-like PC12 cell is extensively utilized to develop neuroprotective agents and identify the underlying molecular mechanisms.39 Ger reduced the injury induced by Glu, indicating that Ger had a possible defensive action in Glu prompted injury in PC12 cells. The next step was to explore GABA signaling pathway. In brain tissues, the GABA accessibility is synchronized by GABA synthesizing glutamate decarboxylases (GAD65 and GAD67) activity, the GABA degrading GABA aminotransferase (GABAT), and the GABA transport proteins (GAT1–3 and BGT-1).40 Loss-of-function mutations in GABAAR genes are an essential mechanism in the pathophysiology of familial idiopathic generalized epilepsy.41 GABAAR α-subunit changes play a significant role in the process of epileptogenesis.42 GAT 1 and GAT 3, as GATs, are expressed in nerve terminals and glia to remove GABA from the synaptic cleft in the brain, and lack of expression of GATs would enhance the ability of GABA to spread through the extracellular space.17 In PTZ-induced kindling was associated with an upregulated gene expression of GAT-1 and GAT-3 indicating that PTZ might affect GABA homeostasis by regulating the GABA transporters.43 GAD65−/− mice develop spontaneous seizures associated with increased mortality,40 whereas GAD67−/− mice showed a 93% reduction in GABA concentration in the cerebral cortex.44 Furthermore, clinical confirmations further showed that lowered GAD activity is associated with several forms of epilepsy.45
The results showed Ger (100 and 200 μg/mL) amplified the gene expressions of GABAA-Rα1, GAD65 and GAD67 and decreased GAT 1, GAT 3 and NMDAR1 expression in NGF prompted PC12 cells damaged by Glu. Additionally, GABA and 5-HT levels were increased in Ger-treated cells (100 and 200 μg/mL). These outcomes signify that Ger escalates the GABA signaling pathway, which is a major inhibitory neurotransmitter.
Regarding inflammatory mediators, TNF-α, IL-4 and IL-1β contents were increased in NGF prompted PC12 cells damaged by Glu whereas Ger reduced these inflammatory cytokines by demonstrating the anti-inflammatory effect.
The in-vitro outcomes achieved displayed that PTZ chronic administration caused a intense alteration of memory. These effects are in line with preceding outcomes that disclosed that PTZ induced kindling in mice disturbs short-term as well as long-term memory. The Y-maze and the novel object recognition task are extensively applied to recognize the implication of diverse brain areas in cognitive abilities.39 The passive avoidance test estimates the capability of the animal to learn and to maintain an associative rule.46 In the current study, Ger increased the percentage of spontaneous alternations suggesting that it may protect mice from memory impairment. Also, Ger mitigated PTZ prompted spatial and recognition memory deficiency signifying anti-amnesic effects of Ger in epileptic animals. Moreover, the outcomes of the open field and rotarod tests propose that locomotion, exploratory behavior and motor coordination are enhanced by Ger in PTZ kindled mice. Earlier studies showed that Ger abated SPS-induced locomotor and memory diminishing, as well as behavioral despair in SPS-induced PTSD.8 Also, Ger improved passive avoidance memory and hippocampal synaptic plasticity deficits in Alzheimer’s disease47 and enriched neuronal construction and spatial memory in D-galactose (D-gal) generated cognitive diminishing.48
In the current study, Ger diminished PTZ induced escalations in GABA-transaminase activity and reductions GABA, GAD and GABA receptor in hippocampus and cortex. These effects advocate that Ger has anticonvulsant, antiepileptogenic, as well as anti-amnesic properties via affecting GABAergic pathway.
Formerly, Ger increased DA, 5-HT concentrations and GAD enzyme activity in SPS-induced PTSD.8 Also, Ger showed antipruritic abilities of on acute and chronic itch via modifying spinal GABA/GRPR signaling.49 A prior study investigated the neuroprotective actions of C. citratus essential oil and hydroalcoholic extract, as well as of its correlated ingredients citral and geraniol in PTZ provoked seizures in zebrafish. The increase in the latency time for the first seizure, was reduced when zebrafish were pretreated with Flumazenil, signifying the contribution of GABAA receptor in the anticonvulsant actions of Ger in zebrafish.38
Oxidative stress is another contributor in the pathogenesis of seizures.50 The existing study exposed that PTZ persuaded oxidative stress, manifested as provoked MDA and NO levels and reduced GSH level, and CAT and SOD activities. Whereas Ger inhibited NO and lipid peroxidation as well as increased GSH level and CAT and SOD activities, proposing that Ger protected brain cells against the destructive actions of reactive oxygen and nitrogen species. Previous studies revealed that Ger exhibited antioxidant effects through which it exerted neuroprotective potential in the cerebral I/R injury,7 acrylamide,51 and cisplatin52 induced neurotoxicity. Furthermore, Ger efficiently amended aging-related memory weakening through intensifying GSH, BDNF, Ach levels, and SOD activity in D-gal prompted cognitive diminishing.48
Another participant involved in the epilepsy pathophysiology is the neuroinflammatory response to epileptogenic insults.53 PTZ-induced kindling triggered neuroinflammatory response causing an elevation in the brain cytokines. Remarkably, Ger prevented the escalation in brain cytokines prompted by PTZ implying that Ger exerts anti-inflammatory properties in epileptic mice. Previous reports revealed that Ger exhibited anti-inflammatory against lipopolysaccharide (LPS) induced inflammation,54 carbon tetrachloride (CCl4) instigated in liver fibrosis,55 trinitrobenzene sulfonic acid (TNBS) induced colitis.56 As for neurological actions, Ger exhibited anti-inflammatory potential against SPS-induced PTSD.8 Prior studies revealed the molecular mechanisms of the potent anti-inflammatory effect of Ger. For instance, Ger protected against traumatic SCI by downregulating the NF-κB and p38 MAPK pathways in SCI rats.6 A another study demonstrated that in Ox-LDL-stimulated HUVECs, Ger suppressed the nuclear translocation and activity of NF-ᴋB as well as phosphorylation of IkBα.57 Also, Ger effectively reduces the expression of COX-2, mPGES-1 (produces PGE2), PTGDS (produces PGD2) and MMP-1 expressions in complete Freund’s adjuvant (CFA)-induced inflammatory arthritis (IA) in rats.5
Both oxidative stress and neuroinflammatory are associated with apoptosis. In the current, Ger reduced caspase-3, and −9 levels and Bax gene expression and escalated Bcl2 gene expression, indicating deterring PTZ associated apoptosis. Similarly, Ger relieved LPS-induced acute lung damage via deterring lung inflammation and apoptosis.58 Also, Ger defended against oxidative stress-induced apoptosis in liver I/R damage.59
In the existing study, SV was administered as a positive control in PTZ kindling-induced seizures. SV associated antioxidant, anti-inflammatory, antiapoptotic, and neuronal damage-preventing effects in mice were due to its ability in preventing seizures and subsequent lower neuronal damage and therefore lesser oxidative stress, inflammation and apoptosis occurring in the animals’ brains. On the other hand, valproic acid alone can be used to induce brain damage60–64 and autism-like symptoms in animals,65–68 and in zebra fish.69
The limitations of the current study include the lack of pharmacokinetics and bioavailability experiments for Ger in the CNS and the absence of cell death-specific assays such as TUNEL assay.
Conclusions
Outcomes of the existing study showed that Ger protects animals against PTZ prompted kindling as established by the enhancement in short term as well as the long-term memory. Ger activated GABAergic neurotransmission, which might elucidate its anticonvulsant effect. Furthermore, Ger reduced the oxidative stress and neuroinflammation in PTZ kindled animals.
Data Sharing Statement
The datasets generated and/or analyzed during the current study are available from the corresponding author on reasonable request.
Institutional Review Board Statement
The Institutional Animal Care and Use Committee of King Faisal University allowed the experimental protocol (KFU-REC-2023-SEP-ETHICS1321). All the experiments were accomplished in agreement with the relevant procedures and regulations of the Ethical Conduct for the Use of Animals in Research at King Faisal University.
Acknowledgments
The authors extend their appreciation to the King Salman center for Disability Research for funding this work.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
The authors extend their appreciation to the King Salman center for Disability Research for funding this work through Research Group no KSRG-2023-401.
Disclosure
The authors declare no conflicts of interest.
References
1. Chen W, Viljoen AM. Geraniol — a review of a commercially important fragrance material. S Afr J Bot. 2010;76(4):643–651. doi:10.1016/j.sajb.2010.05.008
2. Lei Y, Fu P, Jun X, Cheng P. Pharmacological properties of geraniol–a review. Planta med. 2019;85(01):48–55. doi:10.1055/a-0750-6907
3. Pavan B, Dalpiaz A, Marani L, et al. Geraniol pharmacokinetics, bioavailability and its multiple effects on the liver antioxidant and xenobiotic-metabolizing enzymes. Front Pharmacol. 2018;9:9. doi:10.3389/fphar.2018.00009
4. Lira MH, Andrade Júnior FP, Moraes GF, Macena G, Pereira F, Lima IO. Antimicrobial activity of geraniol: an integrative review. J Essent Oil Res. 2020;32(3):187–197. doi:10.1080/10412905.2020.1745697
5. Malik MNH, Tahir MN, Alsahli TG, et al. Geraniol suppresses oxidative stress, inflammation, and interstitial collagenase to protect against inflammatory arthritis. ACS omega. 2023;8(40):37128–37139. doi:10.1021/acsomega.3c04684
6. Lv Y, Zhang L, Li N, Mai N, Zhang Y, Pan S. Geraniol promotes functional recovery and attenuates neuropathic pain in rats with spinal cord injury. Can J Physiol Pharmacol. 2017;95(12):1389–1395. doi:10.1139/cjpp-2016-0528
7. Buch P, Sharma T, Airao V, et al. Geraniol protects hippocampal CA1 neurons and improves functional outcomes in global model of stroke in rats. Chem Biol Drug Des. 2023;102(3):523–535. doi:10.1111/cbdd.14260
8. Ben-Azu B, Adebayo OG, Moke EG, et al. Geraniol attenuates behavioral and neurochemical impairments by inhibitions of HPA-axis and oxido-inflammatory perturbations in mice exposed to post-traumatic stress disorder. J Psychiatr Res. 2023;168:165–175. doi:10.1016/j.jpsychires.2023.10.057
9. Deng X-Y, Xue J-S, Li H-Y, et al. Geraniol produces antidepressant-like effects in a chronic unpredictable mild stress mice model. Physiol Behav. 2015;152:264–271. doi:10.1016/j.physbeh.2015.10.008
10. Rekha KR, Inmozhi Sivakamasundari R. Geraniol protects against the protein and oxidative stress induced by rotenone in an in vitro model of Parkinson’s disease. Neurochem Res. 2018;43(10):1947–1962. doi:10.1007/s11064-018-2617-5
11. Lins LC, Santos IM, de MELO MS, et al. The anticonvulsant effect of geraniol and inclusion complex geraniol: β-cyclodextrin. Bol latinoam Caribe plantas med. 2014;13(6):557–565.
12. Chandran D, Hegde S, Upadhya R, et al. Cell-specific extracellular vesicle-encapsulated exogenous GABA controls seizures in epilepsy. Stem Cell Res Ther. 2024;15(1):108. doi:10.1186/s13287-024-03721-4
13. Slezia A, Proctor CM, Kaszas A, Malliaras GG, Williamson A, Electrophoretic delivery of γ-aminobutyric acid (GABA) into epileptic focus prevents seizures in mice. J Vis Exp. 2019;147:e59268. doi:10.3791/59268-v
14. Upadhya D, Attaluri S, Liu Y, et al. Grafted hPSC-derived GABA-ergic interneurons regulate seizures and specific cognitive function in temporal lobe epilepsy. NPJ Regen Med. 2022;7(1):38. doi:10.1038/s41536-022-00234-7
15. Stork O, Ji F-Y, Kaneko K, et al. Postnatal development of a GABA deficit and disturbance of neural functions in mice lacking GAD65. Brain Res. 2000;865(1):45–58. doi:10.1016/S0006-8993(00)02206-X
16. Soghomonian -J-J, Martin DL. Two isoforms of glutamate decarboxylase: why? Trends Pharmacol Sci. 1998;19(12):500–505. doi:10.1016/S0165-6147(98)01270-X
17. Dalby NO. Inhibition of γ-aminobutyric acid uptake: anatomy, physiology and effects against epileptic seizures. Eur J Pharmacol. 2003;479(1–3):127–137. doi:10.1016/j.ejphar.2003.08.063
18. He L-Y, Hu M-B, Li R-L, et al. The effect of protein-rich extract from bombyx batryticatus against glutamate-damaged PC12 cells via regulating γ-Aminobutyric acid signaling pathway. Molecules. 2020;25(3):553. doi:10.3390/molecules25030553
19. Ma K, Yan N, Huang Y, Cao G, Deng J, Deng Y. Effects of nerve growth factor on nerve regeneration after corneal nerve damage. Int J Clin Exp Med. 2014;7(11):4584.
20. Zeng X, Hu K, Chen L, et al. The effects of ginsenoside compound K against epilepsy by enhancing the γ-aminobutyric acid signaling pathway. Front Pharmacol. 2018;9:1020. doi:10.3389/fphar.2018.01020
21. Lin K-H, Li C-Y, Hsu Y-M, et al. Oridonin, A natural diterpenoid, protected NGF-differentiated PC12 cells against MPP+- and kainic acid-induced injury. Food Chem Toxicol. 2019;133:110765. doi:10.1016/j.fct.2019.110765
22. Ravizza T, Vezzani A. Status epilepticus induces time-dependent neuronal and astrocytic expression of interleukin-1 receptor type I in the rat limbic system. Neuroscience. 2006;137(1):301–308. doi:10.1016/j.neuroscience.2005.07.063
23. Zhen J, Qu Z, Fang H, et al. Effects of grape seed proanthocyanidin extract on pentylenetetrazole-induced kindling and associated cognitive impairment in rats. Int J Mol Med. 2014;34(2):391–398. doi:10.3892/ijmm.2014.1796
24. Racine RJ. Modification of seizure activity by electrical stimulation: II. Motor seizure. Electroencephalogr Clin Neurophysiol. 1972;32(3):281–294. doi:10.1016/0013-4694(72)90177-0
25. Malik H, Javaid S, Fawad Rasool M, et al. Amelioration of scopolamine-induced amnesic, anxiolytic and antidepressant effects of Ficus benghalensis in behavioral experimental models. Medicina. 2020;56(3):144. doi:10.3390/medicina56030144
26. Kraeuter A-K, Guest PC, Sarnyai Z. The Y-maze for assessment of spatial working and reference memory in mice. Pre-Clinical Models. 2019;105–111.
27. Ennaceur A, Delacour J. A new one-trial test for neurobiological studies of memory in rats. 1: behavioral data. Behav Brain Res. 1988;31(1):47–59. doi:10.1016/0166-4328(88)90157-X
28. Royce JR. On the construct validity of open-field measures. Psychol Bull. 1977;84(6):1098. doi:10.1037/0033-2909.84.6.1098
29. Taïwe GS, Bum EN, Talla E, et al. Nauclea latifolia Smith (Rubiaceae) exerts antinociceptive effects in neuropathic pain induced by chronic constriction injury of the sciatic nerve. J Ethnopharmacol. 2014;151(1):445–451. doi:10.1016/j.jep.2013.10.068
30. Li X, Liu N, Wu D, et al. Hippocampal transcriptomic analyses reveal the potential antiapoptotic mechanism of a novel anticonvulsant agent Q808 on pentylenetetrazol-induced epilepsy in rats. Biomed Pharmacother. 2024;175:116746. doi:10.1016/j.biopha.2024.116746
31. Lowe IP, Robins E, Eyerman GS. The fluorimetric measurement of glutamic decarboxylase and its distribution in brain. J Neurochem. 1958;3(1):8–18. doi:10.1111/j.1471-4159.1958.tb12604.x
32. Cortas NK, Wakid NW. Determination of inorganic nitrate in serum and urine by a kinetic cadmium-reduction method. Clin Chem. 1990;36(8):1440–1443. doi:10.1093/clinchem/36.8.1440
33. Barakat W, Safwet N, El-Maraghy NN, Zakaria MN. Candesartan and glycyrrhizin ameliorate ischemic brain damage through downregulation of the TLR signaling cascade. Eur J Pharmacol. 2014;724:43–50. doi:10.1016/j.ejphar.2013.12.032
34. Livak KJ, Schmittgen TD. Analysis of relative gene expression data using real-time quantitative PCR and the 2− ΔΔCT method. methods. 2001;25(4):402–408. doi:10.1006/meth.2001.1262
35. Jin XT, Galvan A, Wichmann T, Smith Y. Localization and function of GABA transporters GAT-1 and GAT-3 in the basal Ganglia. Front Syst Neurosci. 2011;5:63. doi:10.3389/fnsys.2011.00063
36. Quintans-Júnior LJ, Souza T, Leite B, et al. Phythochemical screening and anticonvulsant activity of Cymbopogon winterianus Jowitt (Poaceae) leaf essential oil in rodents. Phytomedicine. 2008;15(8):619–624. doi:10.1016/j.phymed.2007.09.018
37. Oliveira JS, Porto LA, Estevam C, et al. Phytochemical screening and anticonvulsant property of Ocimum basilicum leaf essential oil. Bol latinoam Caribe plantas med. 2009;8(3):195–202.
38. Hacke ACM, Miyoshi E, Marques JA, Pereira RP. Cymbopogon citratus (DC.) stapf, citral and geraniol exhibit anticonvulsant and neuroprotective effects in pentylenetetrazole-induced seizures in zebrafish. J Ethnopharmacol. 2021;275:114142. doi:10.1016/j.jep.2021.114142
39. Jeong JH, Kim HJ, Park SK, et al. An investigation into the ameliorating effect of black soybean extract on learning and memory impairment with assessment of neuroprotective effects. BMC Complement Altern Med. 2014;14(1):1–11. doi:10.1186/1472-6882-14-482
40. Conti F, Minelli A, Melone M. GABA transporters in the mammalian cerebral cortex: localization, development and pathological implications. Brain Res Rev. 2004;45(3):196–212. doi:10.1016/j.brainresrev.2004.03.003
41. Han D-Y, Guan B-J, Wang Y-J, Hatzoglou M, Mu T-W. L-type calcium channel blockers enhance trafficking and function of epilepsy-associated α1 (D219N) subunits of GABAA receptors. ACS Chem Biol. 2015;10(9):2135–2148. doi:10.1021/acschembio.5b00479
42. Grabenstatter HL, Russek SJ, Brooks-Kayal AR. Molecular pathways controlling inhibitory receptor expression. Epilepsia. 2012;9(53 Suppl):71–78. doi:10.1111/epi.12036
43. Javaid S, Alqahtani F, Ashraf W, et al. Tiagabine suppresses pentylenetetrazole-induced seizures in mice and improves behavioral and cognitive parameters by modulating BDNF/TrkB expression and neuroinflammatory markers. Biomed Pharmacother. 2023;160:114406. doi:10.1016/j.biopha.2023.114406
44. Asada H, Kawamura Y, Maruyama K, et al. Mice lacking the 65 kDa isoform of glutamic acid decarboxylase (GAD65) maintain normal levels of GAD67 and GABA in their brains but are susceptible to seizures. Biochem Biophys Res Commun. 1996;229(3):891–895. doi:10.1006/bbrc.1996.1898
45. Lloyd K, Bossi L, Morselli P, Munari C, Rougier M, Loiseau H. Alterations of GABA-mediated synaptic transmission in human epilepsy. Adv Neurol. 1986;44:1033–1044.
46. Tsuji M, Takeda H, Matsumiya T. Modulation of passive avoidance in mice by the 5-HT1A receptor agonist flesinoxan: comparison with the benzodiazepine receptor agonist diazepam. Neuropsychopharmacology. 2003;28(4):664–674. doi:10.1038/sj.npp.1300080
47. Bagheri S, Rashno M, Salehi I, Karimi SA, Raoufi S, Komaki A. Geraniol improves passive avoidance memory and hippocampal synaptic plasticity deficits in a rat model of alzheimer’s disease. Eur J Pharmacol. 2023;951:175714. doi:10.1016/j.ejphar.2023.175714
48. Atef MM, Emam MN, Abo E, et al. Mechanistic insights into ameliorating effect of geraniol on D-Galactose induced memory impairment in rats. Neurochem Res. 2022;47(6):1664–1678. doi:10.1007/s11064-022-03559-3
49. Yang YY, Du LX, Zhu JY, et al. Antipruritic effects of geraniol on acute and chronic itch via modulating spinal GABA/GRPR signaling. Phytomedicine. 2023;119:154969. doi:10.1016/j.phymed.2023.154969
50. Kilinc E, Ankarali S, Ayhan D, Ankarali H, Torun IE, Cetinkaya A. Protective effects of long-term probiotic mixture mentation against pentylenetetrazole-induced seizures, inflammation and oxidative stress in rats. J Nutr Biochem. 2021;98:108830. doi:10.1016/j.jnutbio.2021.108830
51. Prasad SN. Mitigation of acrylamide-induced behavioral deficits, oxidative impairments and neurotoxicity by oral ments of geraniol (a monoterpene) in a rat model. Chem Biol Interact. 2014;223:27–37. doi:10.1016/j.cbi.2014.08.016
52. Kandeil MA, Mahmoud MO, Abdel-Razik AH, Gomaa SB. Thymoquinone and geraniol alleviate cisplatin-induced neurotoxicity in rats through downregulating the p38 MAPK/STAT-1 pathway and oxidative stress. Life Sci. 2019;228:145–151. doi:10.1016/j.lfs.2019.04.065
53. Vezzani A, Friedman A, Dingledine RJ. The role of inflammation in epileptogenesis. Neuropharmacology. 2013;69:16–24. doi:10.1016/j.neuropharm.2012.04.004
54. El Azab EF, Saleh AM, Yousif SO, et al. New insights into geraniol’s antihemolytic, anti-inflammatory, antioxidant, and anticoagulant potentials using a combined biological and in silico screening strategy. Inflammopharmacology. 2022;30(5):1811–1833. doi:10.1007/s10787-022-01039-2
55. Elguindy NM, Elguindy NM, Yacout GA, Elgamal DA. Hepatoprotective impact of geraniol against CCl(4)-induced liver fibrosis in rats. Pak J Biol Sci. 2020;23(12):1650–1658. doi:10.3923/pjbs.2020.1650.1658
56. Soubh AA, Abdallah DM, El-Abhar HS. Geraniol ameliorates TNBS-induced colitis: involvement of Wnt/β-catenin, p38MAPK, NFκB, and PPARγ signaling pathways. Life Sci. 2015;136:142–150. doi:10.1016/j.lfs.2015.07.004
57. Ben Ammar R, Mohamed ME, Alfwuaires M, et al. Anti-inflammatory activity of geraniol isolated from lemon grass on Ox-LDL-stimulated endothelial cells by upregulation of heme oxygenase-1 via PI3K/Akt and Nrf-2 signaling pathways. Nutrients. 2022;14(22):4817. doi:10.3390/nu14224817
58. Jiang K, Zhang T, Yin N, et al. Geraniol alleviates LPS-induced acute lung injury in mice via inhibiting inflammation and apoptosis. Oncotarget. 2017;8(41):71038–71053. doi:10.18632/oncotarget.20298
59. El-Emam SZ, Soubh AA, Al-Mokaddem AK, Abo El-Ella DM. Geraniol activates Nrf-2/HO-1 signaling pathway mediating protection against oxidative stress-induced apoptosis in hepatic ischemia-reperfusion injury. Naunyn Schmiedebergs Arch Pharmacol. 2020;393(10):1849–1858. doi:10.1007/s00210-020-01887-1
60. Terim Kapakin KA, Bolat İ, Iskender H, et al. Protective effects of curcumin against valproic acid induced brain kidney and liver damage in rats. Drug Chem Toxicol. 2024;1–16. doi:10.1080/01480545.2024.2391868
61. Hussein AM, Awadalla A, Abbas KM, et al. Chronic valproic acid administration enhances oxidative stress, upregulates IL6 and downregulates Nrf2, Glut1 and Glut4 in rat’s liver and brain. Neuroreport. 2021;32(10):840–850. doi:10.1097/WNR.0000000000001663
62. Aranarochana A, Sirichoat A, Pannangrong W, Wigmore P, Welbat JU, Hrnčić D. Melatonin ameliorates valproic acid-induced neurogenesis impairment: the role of oxidative stress in adult rats. Oxid Med Cell Longev. 2021;2021(1):9997582. doi:10.1155/2021/9997582
63. Turkyilmaz IB, Altas N, Arisan I, Yanardag R. Effect of vitamin B6 on brain damage in valproic acid induced toxicity. J Biochem Mol Toxicol. 2021;35(9):e22855. doi:10.1002/jbt.22855
64. Hamza R, El-Shenawy N. The beneficial effects of l-cysteine on brain antioxidants of rats affected by sodium valproate. Hum Exp Toxicol. 2017;36(11):1212–1221. doi:10.1177/0960327117695634
65. Saadat M, Taherian AA, Aldaghi MR, Raise-Abdullahi P, Sameni HR, Vafaei AA. Prangos ferulacea (L.) ameliorates behavioral alterations, hippocampal oxidative stress markers, and apoptotic deficits in a rat model of autism induced by valproic acid. Brain Behav. 2023;13(11):e3224. doi:10.1002/brb3.3224
66. Viana CE, Bortolotto VC, Araujo SM, et al. Lutein-loaded nanoparticles reverse oxidative stress, apoptosis, and autism spectrum disorder-like behaviors induced by prenatal valproic acid exposure in female rats. NeuroToxicology. 2023;94:223–234. doi:10.1016/j.neuro.2022.12.006
67. Jian J, Li LG, Zhao PJ, et al. TCHis mitigate oxidative stress and improve abnormal behavior in a prenatal valproic acid-exposed rat model of autism. Physiol Genomics. 2022;54(9):325–336. doi:10.1152/physiolgenomics.00104.2021
68. Mirza R, Sharma B. Benefits of Fenofibrate in prenatal valproic acid-induced autism spectrum disorder related phenotype in rats. Brain Res Bull. 2019;147:36–46. doi:10.1016/j.brainresbull.2019.02.003
69. Wang J, Zou L, Jiang P, et al. Vitamin A ameliorates valproic acid-induced autism-like symptoms in developing zebrafish larvae by attenuating oxidative stress and apoptosis. Neurotoxicology. 2024;101:93–101. doi:10.1016/j.neuro.2023.12.015
 © 2024 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2024 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Eriochloa villosa Alleviates Progression of Benign Prostatic Hyperplasia in vitro and in vivo
Baek EB, Hwang YH, Park S, Hong EJ, Won YS, Kwun HJ
Research and Reports in Urology 2022, 14:313-326
Published Date: 24 September 2022
Selenium Nanoparticles-Enriched Lactobacillus casei ATCC 393 Prevents Cognitive Dysfunction in Mice Through Modulating Microbiota-Gut-Brain Axis

Qiao L, Chen Y, Song X, Dou X, Xu C
International Journal of Nanomedicine 2022, 17:4807-4827
Published Date: 13 October 2022
Zinc Oxide Particles Can Cause Ovarian Toxicity by Oxidative Stress in Female Mice Model

Xu Y, Zhao Y, Liu S, Lv S, Chen L, Wang W, Feng Y, Fu F, Xu H
International Journal of Nanomedicine 2022, 17:4947-4960
Published Date: 19 October 2022
Methanolic Extract of Aerva javanica Leaves Prevents LPS-Induced Depressive Like Behavior in Experimental Mice
Arshad HM, Ahmad FUD, Lodhi AH
Drug Design, Development and Therapy 2022, 16:4179-4204
Published Date: 7 December 2022
Pomegranate Seeds and Peel Ethanolic Extracts Anticancer Potentials and Related Genetic, Histological, Immunohistochemical, Apoptotic and Oxidative Stress Profiles: In vitro Study
Nasr M, Naeem SA, El-Shenbaby I, Mohamed FMA, Mahmoud SM, Abuamara TMM, Abd-Elhay WM, Elbayoumy FMAE, Elkot A, Shikhon T, Abo-akrab M, Doma MA, Hasan A
Journal of Experimental Pharmacology 2023, 15:191-205
Published Date: 15 April 2023