Back to Journals » Journal of Experimental Pharmacology » Volume 15
Pomegranate Seeds and Peel Ethanolic Extracts Anticancer Potentials and Related Genetic, Histological, Immunohistochemical, Apoptotic and Oxidative Stress Profiles: In vitro Study
Authors Nasr M ![]() , Naeem SA, El-Shenbaby I, Mohamed FMA
, Naeem SA, El-Shenbaby I, Mohamed FMA ![]() , Mahmoud SM, Abuamara TMM
, Mahmoud SM, Abuamara TMM ![]() , Abd-Elhay WM, Elbayoumy FMAE, Elkot A
, Abd-Elhay WM, Elbayoumy FMAE, Elkot A ![]() , Shikhon T, Abo-akrab M, Doma MA, Hasan A
, Shikhon T, Abo-akrab M, Doma MA, Hasan A ![]()
Received 10 January 2023
Accepted for publication 1 April 2023
Published 15 April 2023 Volume 2023:15 Pages 191—205
DOI https://doi.org/10.2147/JEP.S404321
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Roger Pinder
Mohamed Nasr,1 Suhaib Alsayed Naeem,1 Ibrahim El-Shenbaby,2 Fatma Mahmoud Abdelraheem Mohamed,3 Safinaz Moustafa Mahmoud,4 Tamer M M Abuamara,1 Wagih M Abd-Elhay,1 Fayez Mohammed Abd Elfattah Elbayoumy,5 Ahmad Elkot,6 Tarek Shikhon,7 Mostafa Abo-akrab,7 Mohamed A Doma,8 Abdulkarim Hasan9
1Histology Department, Faculty of Medicine, Al-Azhar University, Cairo, Egypt; 2Clinical Pharmacology Department, Faculty of Medicine, Mansoura University, Mansoura, Egypt; 3Histology Department, Faculty of Medicine, Assiut University, Assiut, Egypt; 4Histology Department, Faculty of Medicine, Mansoura University, Mansoura, Egypt; 5Anatomy and Embryology Department, Faculty of Medicine, Al-Azhar University, Cairo, Egypt; 6Physiology Department, Faculty of Medicine, Al-Azhar University, Cairo, Egypt; 7Medical Biochemistry Department, Faculty of Medicine, Al-Azhar University, Assiut, Egypt; 8Medical Biochemistry Department, Faculty of Medicine, Al-Azhar University, Cairo, Egypt; 9Pathology Department, Faculty of Medicine, Al-Azhar University, Cairo, Egypt
Correspondence: Abdulkarim Hasan, Department of Pathology, Faculty of Medicine, Al-Azhar University, Cairo, 11884, Egypt, Tel/Fax +20224012932, Email [email protected]
Introduction: Owing to their great quantity of hydrolyzable anthocyanins and tannins, the peel and seeds of pomegranate are edible and possess potent anti-oxidant and anti-inflammatory characteristics. This work aims to trace the pomegranate seed and peel ethanolic extracts’ anticancer activity against liver cancer cell line, namely HepG2 and related histopathological, immunohistochemical, genetic and oxidative stress profile.
Methods: In vitro study for both seed and peel extract showed the prevalence of phenols, polyphenols and acids, those have anti-proliferative potential against liver cancer cell line (HepG2) with 50% inhibitory concentration (IC50) of seed significantly reduced that of peel. Toxicity of test extracts was concentration dependent and accompanied with cell cycle arrest and cell death at theG0/G1 and S phases but not at the G2/M phase. Cell arrest was supplemented with raised ROS, MDA and decreased SOD, GSH and Catalase.
Results and discussion: Apoptosis-related genes showed significant up-expression of pro-apoptotic gene (P53), Cy-C, Bax, and casp-3 and down expression of anti-apoptotic gene (Bcl-2). Also, Casp-3 and P53 proteins were substantially expressed under the effect of test extracts. Histopathological study demonstrated that the untreated cells (control group) were regular cells with nuclear pleomorphism and hyperchromatic nuclei, while seed and peel extracts-treated cells showed necrosis, mixed euchromatin and heterochromatin, intra-nuclear eosinophilic structures, burst cell membranes, and the shrunken apoptotic cells with nuclear membranes and irregular cells. Finally, PCNA gene detected by immunohistochemistry was down regulated significantly under the effect of seed extract treatment than in case of cell medication with peel extract.
Keywords: pomegranate, HepG2 cells, cell viability, apoptosis, oxidative stress, immunohistochemistry
Introduction
Cancer is a genetic disorder characterized by uncontrolled cell division. Multiple epigenetic/environmental factors have a crucial function in the cancer’s advancement, as shown by decades of research.1,2 Numerous parameters involving tobacco, winery intake, poor diet, and obesity enhanced the development of different types of cancer.3,4 Despite the different approaches in cancer therapy, the mortality rate is increasing.3,5 It is anticipated that there would be around 20 million cancer cases by the year 2025; consequently, prevention is being emphasized as the best cancer controlling method.5–7 Presently, it is anticipated that two-thirds of cancer-associated fatalities may be averted primarily via dietary measures.7,8 Many centuries ago, Hippocrates established the food’s significance for general health. The edible substances derived from natural plants known as phytochemicals and can act as cancer chemopreventive/chemotherapeutic agents, and nearly 47% of the accessible anticancer treatments in the market are natural product alternatives.9 Numerous scientific studies have demonstrated consuming an abundance of vegetables and fruits lowers the possibility of acquiring cancer,10–12 and more recently polyphenol, vitamin, and unsaturated fatty acid constituents have been studied.13,14 Nearly 25,000 distinct phytochemicals with powerful anticancer effects have been recognized in fruits and vegetables.15 These phytochemicals are not toxic and usually affect numerous signaling cascades, nuclear factor kappa B (NF-kB), nuclear factor erythroid factor 2-related factor 2 and mitogen-activated protein kinase.16 Pomegranate (Punica granatum L.) seeds are edible and have potent anti-inflammatory and antioxidant features owing to their hydrolysable anthocyanins and tannins’ great content.17 As contrasted to the antioxidant action of β-carotene, ascorbic acid, and vitamin E, the pomegranate antioxidants have distinctive features owing to mixtures of a broad array of polyphenols that have various actions versus many types of free radicals – antioxidant activity.18 Multiple signaling cascades associated with inflammation, hyperproliferation, angiogenesis, cellular transformation, the onset of tumorigenesis, and the suppression of the last phases of metastasis and tumorigenesis have been demonstrated to be efficiently affected by pomegranate and its components.14,16 Components of pomegranate influence transcription factors, anti-apoptotic proteins, pro-apoptotic proteins, cell cycle regulator molecules, pro-inflammatory mediators, cell adhesion molecules, protein kinases, and growth factors in multiple types of cancer.19
The ongoing work’s aim was to trace the pomegranate seed and peel ethanolic extracts’ anticancer activity against liver cancer cell line, namely HepG2 and related histopathological, immunohistochemical, genetic and oxidative stress profile.
Materials and Methods
Pomegranate Peel and Seed Extraction
(Using the common techniques by soaking the dry weight in ethanol for 7 days then dried using rotary evaporator and used as crowed extract).
Peel and seed were weighed (300–400 g) washed and dried at 40°C till complete drying. The dried peel was grinded to fine powder. The dried peel and seeds were steeped in approximately 1500 mL of the extracting solvent (ethanol, 80% concentration). The extraction was completed at 80 °C for around 30 minutes before being filtered via cotton wool. Residue was extracted a second time with 1000 mL of the similar extracting solvent for about 5 minutes in a boiling water bath, stored overnight in the refrigerator, and filtered via a cotton wool plug in the funnel neck. The two extracts were vaporized until no more water could be distilled at 40 °C utilizing rotary evaporator equipment under vacuum. The extracted heavy material was measured and kept at 80°C for future studies.
The compounds from peel and seed were characterized using reverse phase-HPLC Agilent 1100 system equipped with a quaternary pump “G1311A”, a degasser “G1379A”, an ALS automatic sampler “G1329A”, a Colcom column compartment “G1316”, an ALS Therm “G1130B”, a diode array detector “DAD, G1315B” and a system controller that linked to Chem Station data handling systems “Agilent Technologies, Palo Alto, CA, USA”.
Assignment for the metabolites was performed through 1H 1D and 2D NMR spectra “1H-COSY, JRES, 1H-13C HSQC, and 1H-13C HMBC” analysis.
Cell Culture
The Tissue Culture Department of the International Center for Training and Advanced Research in Egypt (ICTAR-Egypt) kindly provided the HepG-2 liver cancer cell line. At 37 ° C, HepG2 cells were cultivated in RPMI-1640 medium comprising 10% foetal bovine serum (Gibco, Thermo Fisher Scientific, Waltham, MA, USA) (Jouan SA, Saint-Herblain, Pays de la Loire, France). In accordance with the production technique, the growing media was eliminated, and the cells were rinsed with PBS (Adwia Pharmaceuticals, El Sharkeya, Egypt). At 37°C, cells were treated for 5 minutes with 0.05% (v/v) EDTA and 0.25% trypsin enzyme (GIBCO). Separated cells were subdivided based on necessity.
Cell Viability (MTT Assay)
Cytotoxicity was done as per to the prior research20 in which human liver cancer cells (HepG2 cells) were cultivated in 75 cm2 plates (SPL-Korea) as reported.20 Cells were seeded at 2×105 HepG2/mL in 96-well cell as 0.1 mL/well. Plates were incubated for 24 hrs at 37°C to attain confluence. Fresh medium comprising twofold serially diluted pomegranate seed and peel extracts was administered to the pre-cultured plate at 0.1 mL/mL. Following 24 hours, lifeless cells were eliminated utilizing phosphate-buffered saline (PBS, pH = 7.2 0.2) (Adwia, Egypt) and 50 µl of MTT stock solution (0.5 mg/mL) was injected to each well. After 4 hours of 37°C incubation, the supernatant was removed and 50 µl/well of dimethyl sulfoxide (DMSO) was applied to dissolve the precipitated formazan. Plates were incubated for 30 min at 37°C without light, and the absorbance was taken at 570 nm utilizing a microplate reader (ELx-800, Bio-Tek Instruments, Inc, Winooski, VT, USA). The formula below was employed to compute the proportion of viable cells.
Viability percentage (%) = Mean OD of test dilution x100/Mean OD of control wells.
The IC50 percentage was measured utilizing GraphPad Prism software (v. 6, GraphPad Software, La Jolla, CA, USA).
Cell Cycle Analysis
Cell cycle and apoptotic outline were done in line with Alshehri et al21 in which HepG-2 cells were pre-cultured in 75 cm2 surface area (SA). After removing the excess, the growth medium, cell culture plates were received the pomegranate peel and seed extracts’ IC50 in RPMI-1640 medium. After 24hrs, the afflicted cells were taken and subdivided for genetic and cell cycle analyses. Cells were gathered for flowcytometry analysis, fixed with 70% (v/v) ethanol in PBS, stored at 4°C overnight, and resuspended in a dark environment in PBS comprising 40 g/mL PI (propidium iodide), 0.1% (v/v) Triton X-100, and 0.1 mg/mL RNase. Following 30 min at 37°C, the cells were analysed with a flow cytometer (Becton-Dickinson, San Jose, CA, USA) accompanied with a 488 nm argon ion laser (Becton-Dickinson, San Jose, CA, USA). As priorly defined, the cell cycle and sub G1 group were measured and studied.
Molecular Biology
Apoptosis-associated genes’ expression were detected through extraction of total RNA from control, and IC50 pomegranate peel- and seed-medicated cells utilizing RNeasy Mini Kit (Qiagen, Germantown, MD, USA) as per producer’s guidelines. The isolated RNA quantity was validated utilizing a Beckman dual spectrophotometer (Beckman Instruments, Ramsey, MN, USA). The apoptosis-associated genes’ expression level; BAX (F: 5’-ATG GAC GGG TCC GGG GAG CA-3’and R: 5’-CCC AGT TGA AGT TGC CGT CA- 3’), P53 (F: 5’-TCA GAT CCT AGC GTC GAG CCC-3’ and R: 5’-GGG TGT GGA ATC AAC CCA CAG-3’), CASP3 (F 5- CTCGGTCTGGTACAGATGTCGA-3’ and R: (5- CATGGCTCAGAAGCACACAAAC-3), BCL2 (F: 5’-GTG AAC TGG GGG AGG ATT GT-3’and R: 5’-GGA GAA ATC AAA CAG AGG CC-3’), CYC (F: 5;-CCAATGAAGATCCCACATG −3; CYC R: 5;-CCAGGAAAGTAGGGGTTGAAGT −3 and housekeeping gene, ACTB (F 5’- AGCGAGCATCCCCCAAAGTT- 3’and R: 5’-GGGCACGAAGGCTCATCATT-3) were estimated utilizing real-time PCR. In which 10 ng of the extracted total RNA from each specimen was utilized for cDNA synthesis employing great ability cDNA reverse transcriptase kit (Applied Biosystems-Thermo Fisher Scientific, USA). The collected cDNA was expansion utilizing Sybr Green I PCR master kit (Thermo Fisher Scientific Inc., Lithuania) and Step One apparatus (Applied Biosystems, Thermo Fisher Scientific Inc.) as described: 10 min at 95 ° C for enzyme stimulation, accompanied by 40 cycles of 15 sec at 95 ° C, 20 sec at 55 ° C, and 30 seconds at 72 ° C for amplification. Variations in each target gene’s expression were normalised with respect to the mean critical threshold (CT) readings of the housekeeping gene β-actin utilizing the DCT method.
Hematoxylin and Eosin Stain
On clean slides, 50 µL of IC50 pomegranate peel and seed extracts-treated HepG2 cells were plated (3 for each medication). The slides were air-dried, fixed with methanol, and rehydrated in descending alcohol dosage (100%, 90%, 75%, and 50%). The slides were rinsed for 5 minutes in distilled water. The slides were waterlogged for 3 minutes in filtered hematoxylin (Hx) stain and rinsed twice with distilled water. The slides were waterlogged for 5 seconds in filtered eosin stain and then rinsed with distilled water. The slides were soaked in xylene, mounted with Canada balsam, and then allowed to dry with cover slips in place. On each slide, 10 microscopic fields were photographed utilizing a 400× magnification. This was accomplished with a digital camera (Canon, Japan) attached to a light microscope. The images were uploaded to a computer system for examination. Fields were selected dependent on the existence of the most apoptotic cells. The photomicrographs were done for qualitative analysis for the morphological apoptotic measures’ existence.21
Gas Chromatography Mass Spectrometry (GC-MS) Analysis
The pomegranate peel and seed extracts’ chemical composition was determined utilizing a Trace GC1310-ISQ mass spectrometer (Thermo Scientific, Austin, TX, USA) equipped with a direct capillary column TG–5MS (30 m × 0.25 mm × 0.25 m film thickness). The column oven temperature was originally conducted at 50 °C before being elevated by 5°C/min to 230 °C and held for 2 minutes. The column oven temperature was then increased by 30°C/min to 290 °C and preserved for 2 min. Temperatures for the MS transfer line and injector were preserved at 250 and 260°C, consecutively. At a steady flow rate of 1 mL/min, helium was utilised as the carrier gas. The solvent interval was 3 minutes, and diluted samples of 1 µL were automatically administered using an Autosampler AS1300 coupled to a GC in split mode to deliver diluted specimens. In full scan mode, EI mass spectra were obtained at ionisation voltages of 70 eV for the m/z range 40–1000. The temperature of the ion source was set at 200 °C. The constituents were identified by contrasting their retention durations and mass spectra with those of the NIST 11 and WILEY 09 mass spectral databases.
Western Blotting
Upon completion of the experiment, the cells were lysed in cold lysis buffer. The cells were then promptly kept at −20 °C for 1 h for additional lysis, gathered by cell scraper, sonicated 2×105 time, then spun at 4000 rpm for 10 min while cooled. Before proceeding with Western blotting, total protein levels were measured colorimetrically in the supernatant via the Bradford assay. Western blotting assay was performed as per,22,23 in which equal quantities (20 µg) of protein specimens were combined and heated with SDS Loading buffer for 10 min, allowed to cool on ice and subsequently was loaded onto SDS-polyacrylamide gel and separated by Cleaver electrophoresis unit (Cleaver, UK), transferred onto polyvinylidene fluoride (PVDF) membranes (BioRad) for 30 min. The membrane was blocked for 2 hours at room temperature with 5% nonfat dry milk in TBS-T to reduce non-specific protein interactions between the antibody and the membrane. The membrane was treated overnight at 4°C with Cell Signaling Technology’s P53 and Caspase 3 primary antibodies and Sigma’s -actin. The blots were then cleansed with TBS-T triplicates for 10 minutes each time. The membrane was then incubated with secondary antibodies conjugated to horse radish peroxidase (HRP) (Dako, USA) at room temperature for an additional hour, followed by three 10-minute washes with TBS-T. Following the producer’s instructions, the chemiluminescent Western ECL substrate device (Perkin Elmer, Waltham, MA) was applied to the blot. In the end, membranes were incubated for 1 minute with a mixture of ECL solution A and ECL solution B of equal proportion. The chemiluminescent signals were recorded using a CCD camera-based imager (Chemi Doc imager, Biorad, USA), and the band intensities were examined utilizing ImageLab (Biorad). On all gels, protein size markers were used to define the gel transfer zones for specific proteins and evaluate the transfer efficiency.
Statistical Analysis
Each experiment was conducted three times independently. Results were examined statistically with Graph Pad Prism v. 6.07 (Graph Pad Software, San Diego, CA, USA) utilizing the subsequent assays: t-test between two subgroups, regression analysis, one-way analysis of variance (ANOVA) between more than two subgroups, and Spearman correlation assay. All findings were provided as the mean ±SD. At P < 0.05, the difference was found statistically significant.
Results
Compounds generated from polyphenols, phenols, and acids in purified pomegranate seed and peel extracts were chemically recognized, and their MW, formulae, and RT were tabulated and illustrated (Table 1 and Table 2).
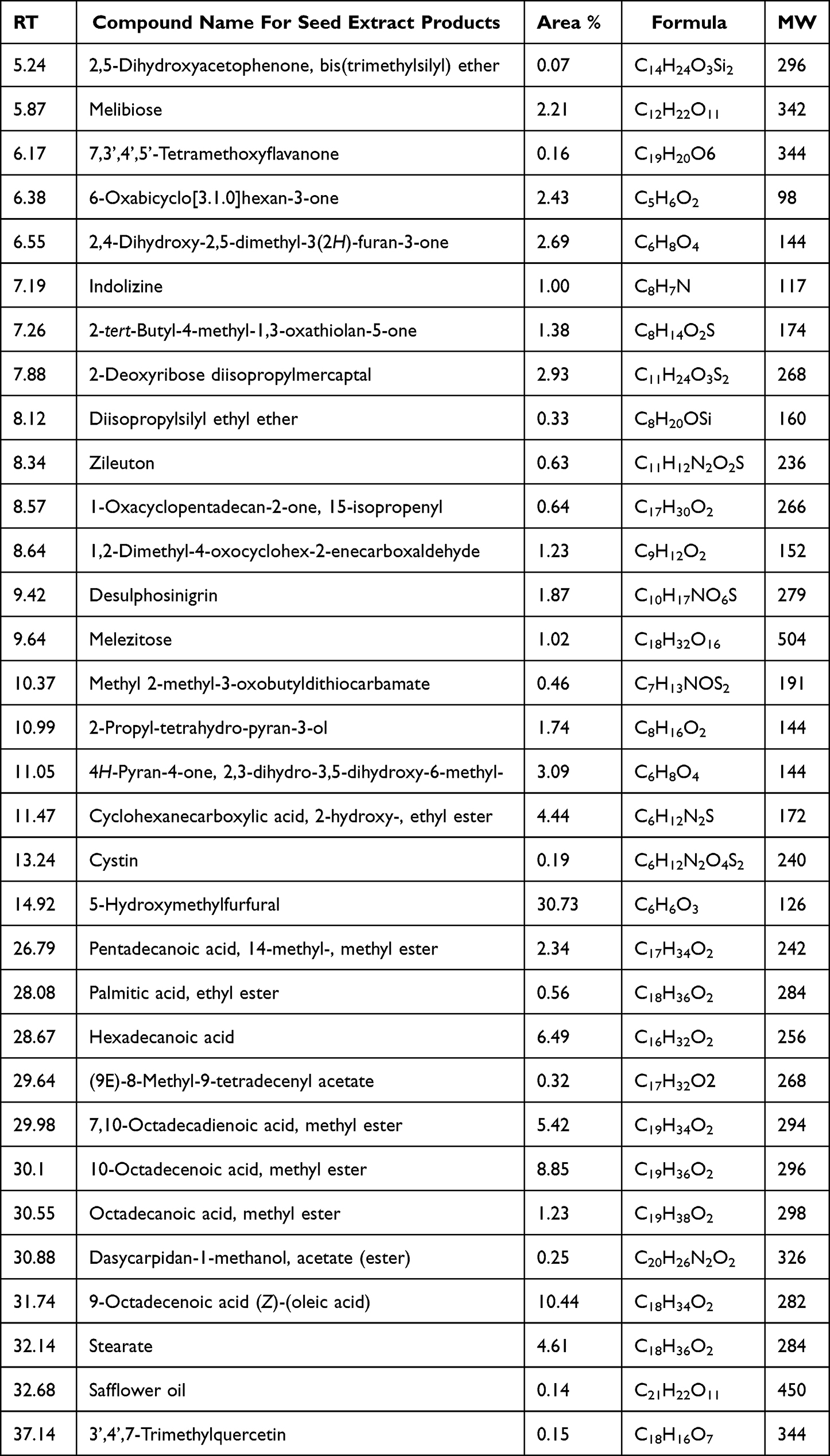 |
Table 1 Identification of Seed-Extracted Compounds Using GCMS |
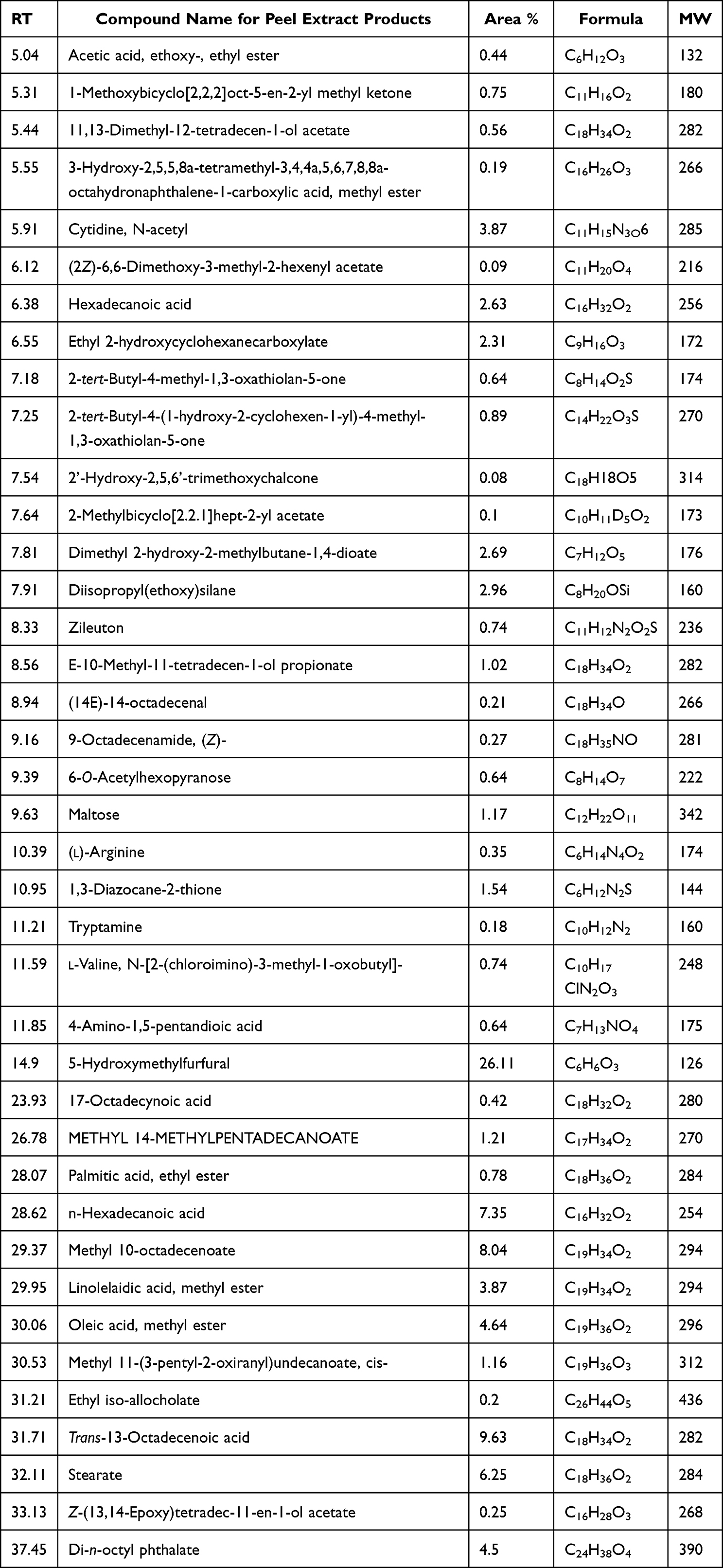 |
Table 2 Identification of Peel Extracted Compounds Using GCMS |
Cytotoxicity (MTT Assay)
Viability assay of liver cancer namely HepG2 performed against peel and seed pomegranate extracts showed a concentrate-dependent viability as viability % increased as long as the extracts concentrations decreased. ALso, there was a significant toxic effect of pomegranate seed compared with that of peel extract (P<0.05). The toxicity was glory regarding the inhibitory concentration (IC50) recording (7.8 and 1.95 µgm/mL) consecutively (Figures 1 and 2).
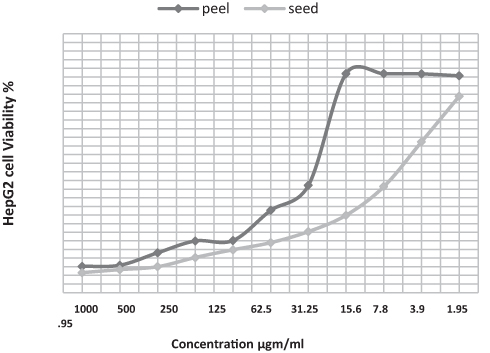 |
Figure 1 Evaluation of HepG2 viability % post cell treatment with pomegranate seed and peel ethanolic extract relative to concentration. |
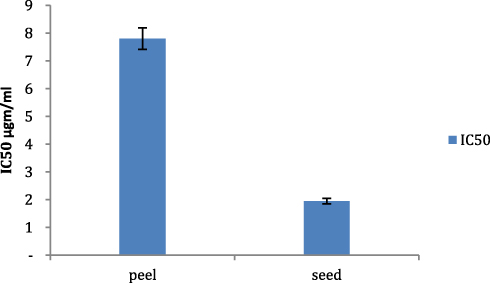 |
Figure 2 Evaluation of Inhibitory concentration (IC50) of both pomegranate seed and peel ethanolic extract. |
Biochemistry Assay
Concerning the antioxidant profile post HepG2 cells treatment with pomegranate peel and seed extracts, it was noticed that ROS level detected a significant (P<0.05) rise contrasted with ROS values contrasted with its value in untreated cell control (506.3, 56812 and 323 Pg/mL) respectively, while there was an insignificant (P>0.05) difference of ROS values (506.3 and 568.12 Pg/mL) post cell treatment with both of peel and seeds. At the same time, malondialdehyde (MDA) showed a significant (P<0.05) elevation post HepG2 cell medication with pomegranate seed extract than in case of cells medication with peel extract and its value I untreated cell control (2.531, 1.817 and 1.575 nmol/mL), respectively. Also, MDA-detected post cells treatment with seed extract demonstrated an insignificant (P>0.05) rise than that detected in untreated cell control (1.817 and 1.575 nmol/mL). On the contrary glutathione reductase (GR/GSH) indicated a significant (P<0.05) decreased values in both pomegranate seed- and peel extracts-medicated HepG2 cells (1.18, 1.868 and 3.93 µg/mL). Similarly, superoxide dismutase (SD) showed a similar profile of GSH/GR where there was a significant (P<0.05) depleted values of SOD post HepG2 cells treatment with pomegranate seed and peel extracts contrasted with its values in untreated cell, control (1.18, 1.868, and 3.93 ng/mL), respectively. Also, peel extract-treated cells showed a significantly depleted value of SOD contrasted with that of untreated cell control (1.868 and 3.93ng/mL) respectively. Finally, catalase activity showed a significantly (P<0.05) decrease activity post HepG2 cell medication with pomegranate peel and seed extracts (0.00731, 0.00952 and 0.0194 nmol/mL) respectively. Also, seed extract-treated HepG2 cells showed insignificant depleted catalase activity (P>0.05) contrasted with its value in peel-treated cells.
Flow Cytometry
Cell cycle profile of liver cancer cells treated with both pomegranate peel and seeds extracts demonstrated a mild substantial elevated (P<0.05) cell arrest/DNA accumulation at the G/G1 phase contrasted with its value in untreated cell control (55.24%, 57.41% and 47.55%) respectively, where there was a nonsignificant (P>0.05) difference in DNA accumulation % between pomegranate peel- and seed-treated cells (55.24%, 57.41%). On the contrary, there was a significant (P<0.05) DNA accumulation % throughout the S phase post HepG2 cell treatment with pomegranate peel- than seed-treated cells (39.15%, 27.52%), while both peel- and seed-treated HepG2 cells demonstrated nonsignificant (P>0.05) elevated DNA accumulation throughout the S phase contrasted with DNA accumulation in the untreated cell control (39.15%, 27.52% and 34.09%) but not in case of seed-treated HepG2 cells. Also, there was a significant (P<0.05) elevated DNA accumulation post HepG2 cell treatment with pomegranate seed than in case of application of peel extract, despite both showed an insignificant (p>0.05) change contrasted with DNA accumulation detected in case of untreated cell control (15.07, 5.61% and 18.36%) respectively. In the same way, there was a significant (P<0.05) raised overall apoptosis of pomegranate peel- and seed-medicated cells contrasted with total apoptosis detected in untreated cell control (24.75%, 33.69% and 2.16%) accompanied with a significant (P<0.05) difference of total apoptosis detected in case on cell treatment with both pomegranate peel and seed extracts (24.75%, 33.69% and 2.13%) respectively. Also, early and late apoptosis in peel- and seed-treated HEpG2 cells was a significantly (P<0.05) raised than that detected in unmedicated cell control (19.17%, 12.66% and 0.32%) consecutively. Early apoptosis of peel-treated cells was significantly (P<0.05) raised than that in seed-treated cells (19.17% and 12.66%), while seed-treated cells demonstrated a significantly (P<0.05) raised late apoptosis than that in peel-treated cells (17.51% and 4.04%). Finally, pomegranate seed-treated cells demonstrated significant (P<0.05) elevated necrotic cell % than in peel-treated and untreated negative control cells (3.52%, 1.54% and 1.67%), respectively (Figure 3).
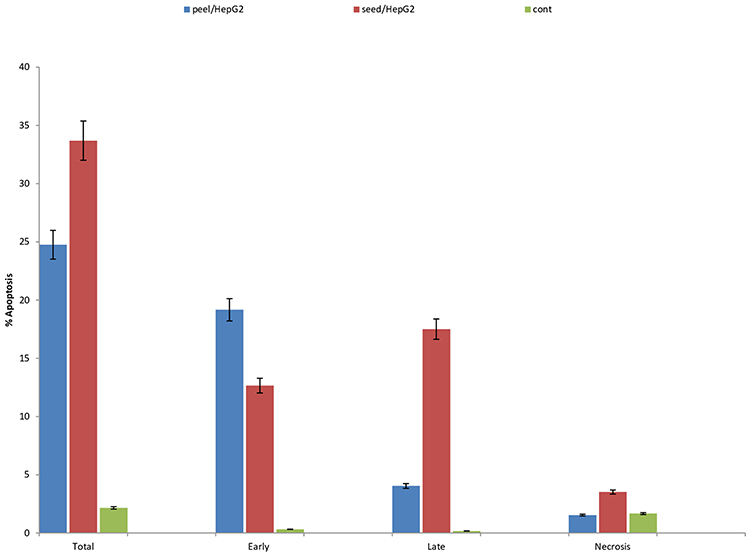 |
Figure 3 Apoptotic phases: evaluation of cell cycle profile and cell arrest phases and apoptosis profile post HepG2 cell treatment with the IC50 of pomegranate seed and peel ethanolic extract. |
Molecular Biology
Regarding the apoptotic related genes, as shown in Figure 4, it was noticed that there was a significant (P<0.05) upregulation of Bax, P53 Casp-3 and cytochrome-c in seed- and peel-medicated cells contrasted with that detected in untreated cell control (7.944, 5.293, 2.976, 5.556 and 1), respectively. At the same time, there was a significant down expression of anti-apoptotic gene (Bcl-2) recording (0.328, 0.211 and 1).
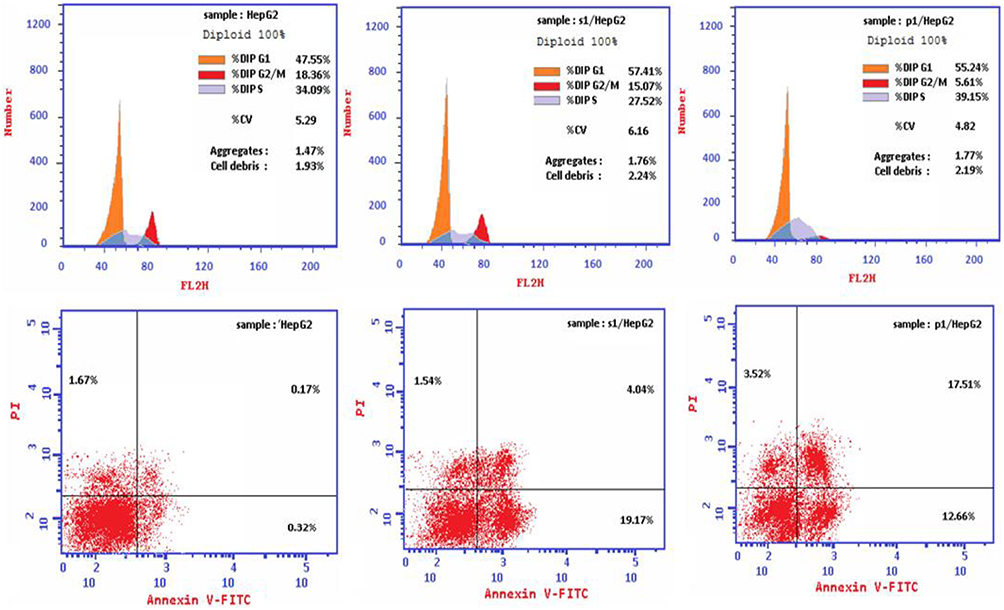 |
Figure 4 Assessment of pro-apoptotic and anti-apoptotic genes post HepG2 cells therapy with the IC50 values of pomegranate seed and peel extracts using rt-PCR. |
Histopathology
Histopathological changes in detected in liver cancer cells treated with both pomegranate seeds and peel extracts (Figure 5) showed that untreated cell control were regular cells with nuclear enlargement and hyperchromatic nuclei. While seed-treated cells were necrotic with combined heterochromatin and euchromatin (red arrows), Intranuclear eosinophilic structures (orange arrows), Ruptured cell membranes (green arrows), Shrunken apoptotic cells with irregular cell and nuclear membranes (black arrows) and peel-medicated cells were necrotic with mixed euchromatin and heterochromatin (red arrows) as shown in Figure 5.
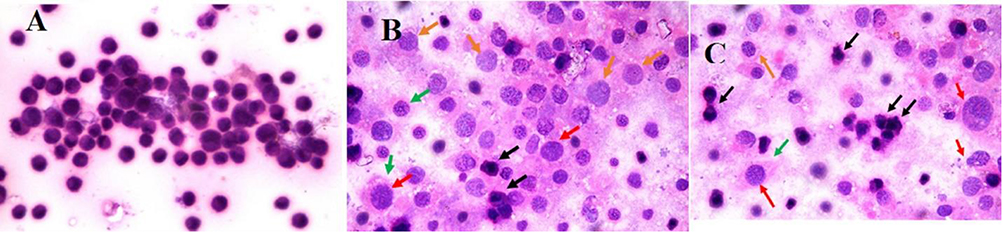 |
Figure 5 (A) Photomicrograph demonstrated regular cells with nuclear pleomorphism and hyperchromatic nuclei. (B) A photomicrograph showed necrotic cells with combined euchromatin and heterochromatin (red arrows), Intranuclear eosinophilic structures (orange arrows), Ruptured cell membranes (green arrows), Shrunken apoptotic cells with nuclear membranes and irregular cell (black arrows) and (C) A photomicrograph demonstrating necrotic cells with combined euchromatin and heterochromatin (red arrows), Intranuclear eosinophilic structures (orange arrows), Ruptured cell membranes (green arrows), Shrunken apoptotic cells with nuclear membranes and irregular cell (black arrows) (H and E, Original magnification 100×, Oil). |
Western Blot
Apoptosis (Figure 6) was detected in a different way compared with that of the traditional rt-PCR, and it was noticed that the optical density of expressed caspase-3 and P53 gene proteins was detected using Western blot method. It was noticed that HepG2c cells medicated with pomegranates seed extract enhanced the caspase-3 and P53 gene expressed proteins significantly (P<0.05) contrasted with its OD in case of HepG2 cell treatment with pomegranate peel. Also, the optical density of the two genes was significantly (P<0.05) elevated than the optical density detected in untreated cell control.
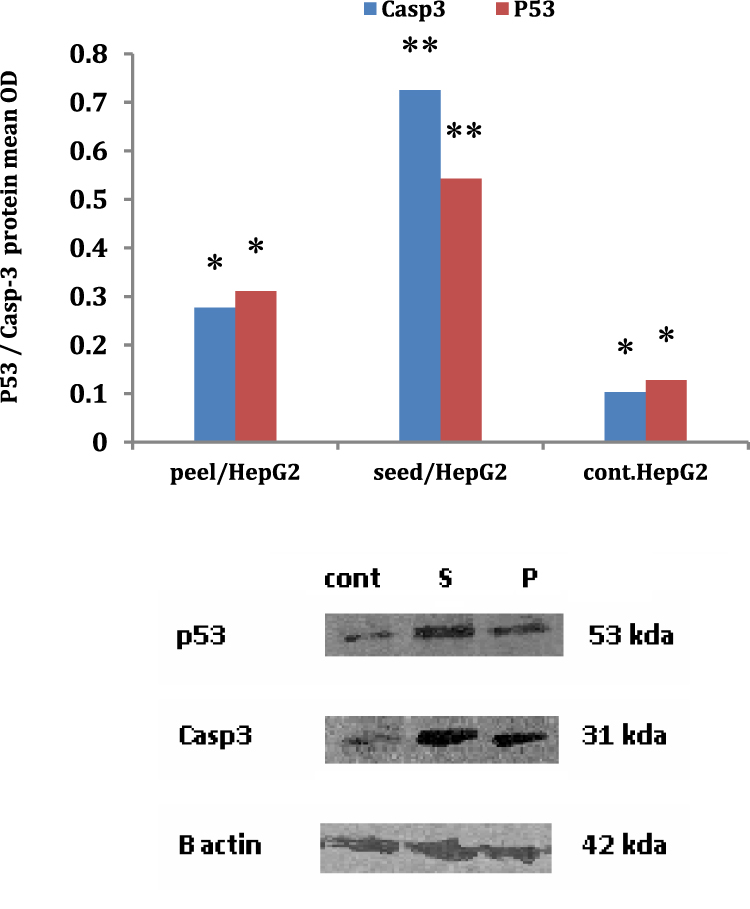 |
Figure 6 Evaluation of apoptotic genes using Western blot assay post HepG2cells treatment with pomegranate seed and peel extracts. |
Immunohistochemistry
Immunohistochemistry analysis (Figure 7) was performed for detection of PCNA in HepG2cells treated with both pomegranate peel and seed extracts. Data revealed that the protein expression was down regulated in seed extract-treated cells significantly that that detected in peel extract-treated cells.
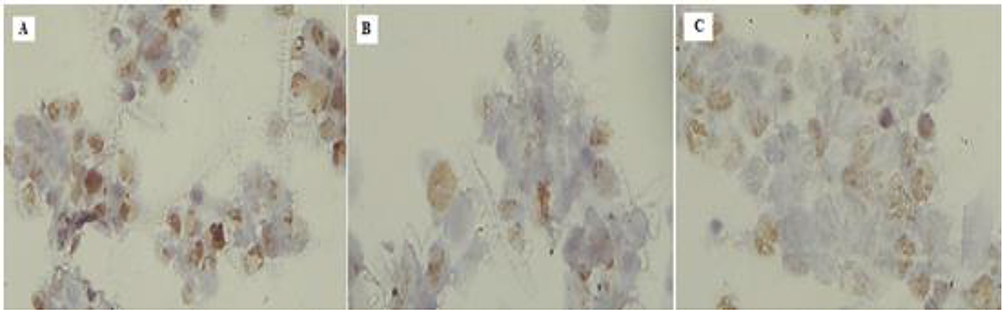 |
Figure 7 Evaluation of PCNA protein immunohistochemical expression in HepG2 cell post treatment showing downregulation of the staining (from A to C). |
Discussion
Pomegranates (Punica granatum L.) have traditional health benefits, as it contains variable remarkable bioactive derivatives have a wide variety of medicinal therapeutic potentials.24 Various studies have proven that various pomegranate pieces have antibacterial properties vs a broad spectrum of infections. The compounds with the greatest biological activity are ellagic acid and hydrolysable tannins, including punicalagin.25 Use of pomegranate peel extract was linked to anticancer effects, including reduction of tumor growth, invasion, and angiogenesis. The pharmacological action of pomegranate derivatives demonstrated an extensive array of clinical uses for cancer avoidance and the therapy of chronic inflammatory diseases.26 During the last few years, several scientific works on pomegranate constituents have been suggested to have antioxidant, anti-carcinogenic activities, which are effective against different types of cancer.27–29 The present study indicated that both peel and seed extract have an anti-proliferative activity, and cell viability was extract type dependent, as viability increased as long as the concentration deceased. Also, the inhibitory concentration of both extracts varies as seed extract was significantly decreased compared with its value of peel extract, that was in accordance with.26,30 Also, Pomegranate as anticancer agent against different types of cancers24,31–34 showed anti-proliferative and anti-cancer actions versus colon, oral, prostate, and breast-cancer cell lines. Cancer-avoiding and cancer-treating properties of pomegranate have been identified, as well as its ability to protect diabetic rats from oxidative stress-induced damage. And according to our reported data, pomegranate peel and seed extracts indicated promising anticancer prospective versus liver cancer cell line (HepG2) that was in agreement with the report of several studies,24,35–40 recording that pomegranate have shown anticancer activities against different organ cancers. Anticancer properties were related to the presence of polyphenols, which demonstrated anticarcinogenic properties in vivo and in vitro. Pomegranate extract caused apoptosis and decreased proliferation in cancer cells; also, pomegranate juice caused considerable apoptosis in myeloid and lymphoid leukemia cell lines, in conjunction with cell cycle arrest.41 Despite the various types of cancers studied,41 found that pomegranate extracts inhibited the breast, colon, lung, and prostate cancer cells’ growth in vitro. In preclinical animal tests, pomegranate extract suppressed skin, colon, lung, and prostate tumor growth. In an initial Phase II clinical research, prostate cancer patients who received pomegranate juice had a significantly longer prostate specific antigen doubling time. In laboratory experiments, pomegranate seed oil has been demonstrated to suppress the growth of many kinds of tumour cells42 and to reduce skin carcinogenesis in mice and mouse mammary organ culture models of carcinogenesis.43 The activity of pomegranate juice suggests additive or synergistic impacts from the other phytochemicals included in pomegranate juice.17 Extracts of pomegranate peel have been shown to suppress cell growth in various cancer cell lines.42,44,45 Anti-carcinogenic features of various mixtures of the phytochemicals in pomegranate were evaluated before.46 Tang et al47 discovered that punicalagin inhibits the β-catenin signaling pathway, hence possessing chemopreventive and chemotherapeutic properties against human cervical cancer. Ellagic acid was contrasted to its antecedent punicalagin and the overall tannin content of pomegranate juice. All of them decreased the proportion of alive human colon and oral cancer cells, however pomegranate juice was the most powerful antioxidant sample examined. Concerning the apoptosis related protein expression we suggest the apoptosis may be via mitochondrial path way as pro apoptotic proteins (P53 and Caspase- 3) expression were noticed utilizing Western blot methods and the expression rate was extract dependent that was in agreement with48 reported that the apoptosis-related proteins’ expression (p53, Bcl-2, Cyt-c, Bax, and PARP) utilizing western-blot was significantly elevated in EA, PC, and PPPs-treated liver cancer cells (HepG2) and the alterations in apoptosis-associated protein expression in HepG2 cells recommends that EA and PC could cause HepG2 cells apoptosis and its apoptosis may be through the intracellular mitochondrial cascade. Antioxidants are compounds that react with and neutralize free radicals, avoiding them from resulting in tissue destruction. Antioxidants are often commonly referred to as “free radical scavengers”. Some of the antioxidants used to neutralize endogenous free radicals are created via the body. Nevertheless, the body obtains the remaining antioxidants from external (exogenous) sources, primarily from the diet. Typically, these exogenous antioxidants are referred to as dietary antioxidants. Vegetables, fruits, and grains are significant dietary sources of antioxidants. In addition, several antioxidants are accessible as nutritional supplements.49,50 The antioxidants beta-carotene, vitamins A, C, and E, and lycopene are examples of dietary antioxidants (alpha-tocopherol). Selenium is commonly believed to be a dietary antioxidant; however, its antioxidant benefits are likely due to the antioxidant action of proteins that contain this element as an important portion (eg Selenium containing proteins), and not to selenium itself.51 Also, and in agreement with the data of the present work concerning the apoptosis parameters including, cell cycle, gene profile and antioxidant profile, it was reported by52 that pomegranate peel extracted polyphenols (PPPs) affected the growth and apoptosis mechanisms of HepG2 cells and the HepG2 cells cycle was arrested at the S-phase via PPPs that was in agreement with our data as cells were arrested at the Go/G1 and S phases and % of arrest was extract dependent and the quantity of the early apoptotic cells and ROS level were clearly enhanced, the Cyt-c level and the action of Caspase-3/9 increased greatly and also did the proportion of Bax/Bcl-2 and the P53 protein expressions. PPPs inhibited the development of HepG2 cells by inhibiting the cell cycle and triggering the mitochondrial apoptotic cascade in a dose-dependent approach. And similar with our findings,48 demonstrated that Pomegranate peel polyphenols (PPPs) extracts exhibited anti-proliferation and apoptosis-inducing actions in human HepG2 hepatoma cells, especially PC and EA and their action mechanisms. And the outcome revealed that PC and EA induced dose-dependent S-phase and G0/G1-phase cell cycle arrest and death in HepG2 cells. Furthermore, apoptosis-associated protein action and reactive oxygen species quantities indicated that apoptosis-related protein activation and intracellular H2O2 production were enhanced. PC was more efficient than EA at inhibiting cell expansion and inducing apoptosis in HepG2 cells. In addition, their engagement in the mitochondrial apoptosis cascade is an essential apoptosis-inducing mechanism. Concurrently,53 found that the majority of pomegranate peel polyphenols consist of PC (65.75%) of the total phenol.54 Consequently, there are normally two types of pomegranates: one is a natural extract of pomegranate peel polyphenols and the other is pomegranate juice. PC and EA induce apoptosis in hepatoma cells, and their mechanism affects cell morphology and viability, cell-cycle distribution, caspase-9 and caspase-3 actions, apoptosis, and intracellular reactive oxygen species (ROS) generation in HepG2 cells. In addition, both PC and EA were found to strongly limit the growth of HepG2 cells and cause apoptosis. PC and EA may eventually be associated with engagement in the mitochondrial apoptotic cascade. EA is the hydrolysis product of PC; its molecular structure is less complex.54 PC derivative predominantly inhibited the cell cycle during the S phase, whereas EA did so at the G0/G1 phase. At present, it was believed that phytochemicals contribute via many mechanisms by detecting distinct targets.55 Regarding the impact of pomegranate-derived phenolic compounds as PC and EA on cell cycle arrest at the S and G/G1 phases55 demonstrated that PC showed a retardation influence on HepG2-treated cells inducing cell cycle arrest at the S phase and EA-induced cell arrest at the G0/G1 phase as well, that was attributed to the hypothesis that the Phytochemical contribution in cell cycle arrest done via different cascades by recognizing various targets. The cell cycle’s regulation is hypothetically via the retention’s G1 phase. The G0 phase occurs when the cell temporarily exits the cell cycle and stops dividing.56
Conclusion
Finally, it can be concluded that both pomegranate seed and peel extracts showed cytotoxic effect on liver cancer cell line. Cell proliferation inhibitory concentration was much effective in case of seed extract tan in peel extract. Also, cell cycle arrest was significantly detected at the S and G0/G1 phases according to the other references, apoptotic genes were significantly expressed post HepG2 cell treatment with seed extract than peel. Antioxidant profile indicates the good performance of free radical scavenging action and lipid peroxidation. Also, histopathological changes were detected using H&E stain in addition to the expression of apoptotic gene proteins recognized by Western blot and PCNA.
Ethical Approval
Ethical approval was obtained from Al-Azhar University, Faculty of Medicine in Cairo.
Funding
The authors have no financial or non-financial interests to disclose.
Disclosure
The authors declare that they have no competing interests.
References
1. Pogribny IP, Rusyn I. Environmental toxicants, epigenetics, and cancer. Adv Exp Med Biol. 2013;754:215–232.
2. Oh ES, Petronis A. Origins of human disease: the chrono-epigenetic perspective. Nat Rev Genet. 2021;22(8):533–546. doi:10.1038/s41576-021-00348-6
3. Torre LA, Bray F, Siegel RL, Ferlay J, Lortet-Tieulent J, Jemal A. Global cancer statistics, 2012. CA Cancer J Clin. 2015;65:87–108. doi:10.3322/caac.21262
4. Khan N, Afaq F, Mukhtar H. Lifestyle as risk factor for cancer: evidence from human studies. Cancer Lett. 2010;293:133–143. doi:10.1016/j.canlet.2009.12.013
5. Siegel RL, Miller KD, Jemal A. Cancer statistics, 2016. CA Cancer J Clin. 2016;66:7–30. doi:10.3322/caac.21332
6. DiMarco-Crook C, Xiao H. Diet-based strategies for cancer chemoprevention: the role of combination regimens using dietary bioactive components. Annu Rev Food Sci Technol. 2015;6:505–526. doi:10.1146/annurev-food-081114-110833
7. Key TJ, Schatzkin A, Willett WC, Allen NE, Spencer EA, Travis RC. Diet, nutrition and the prevention of cancer. Public Health Nutr. 2004;7:187–200. doi:10.1079/PHN2003588
8. Barnard RJ. Prevention of cancer through lifestyle changes. Evid Based Complement Altern Med. 2004;1:233–239. doi:10.1093/ecam/neh036
9. Newman DJ, Cragg GM. Natural products as sources of new drugs over the last 25 years. J Nat Prod. 2007;70:461–477. doi:10.1021/np068054v
10. Boeing H, Bechthold A, Bub A, et al. Critical review: vegetables and fruit in the prevention of chronic diseases. Eur J Nutr. 2012;51:637–663. doi:10.1007/s00394-012-0380-y
11. Sauvaget C, Nagano J, Hayashi M, Spencer E, Shimizu Y, Allen N. Vegetables and fruit intake and cancer mortality in the Hiroshima/Nagasaki Life Span Study. Br J Cancer. 2003;88:689–694. doi:10.1038/sj.bjc.6600775
12. Gundgaard J, Nielsen JN, Olsen J, Sørensen J. Increased intake of fruit and vegetables: estimation of impact in terms of life expectancy and healthcare costs. Public Health Nutr. 2003;6:25–30. doi:10.1079/PHN2002355
13. Huang WY, Cai YZ, Zhang Y. Natural phenolic compounds from medicinal herbs and dietary plants: potential use for cancer prevention. Nutr Cancer. 2010;62:1–20. doi:10.1080/01635580903191585
14. Khan N, Afaq F, Mukhtar H. Cancer chemoprevention through dietary antioxidants: progress and promise. Antioxid Redox Signal. 2008;10:475–510. doi:10.1089/ars.2007.1740
15. Anand P, Kunnumakkara AB, Sundaram C, et al. Cancer is a preventable disease that requires major lifestyle changes. Pharm Res. 2008;25:2097–2116. doi:10.1007/s11095-008-9661-9
16. Maphetu N, Unuofin JO, Masuku NP, Olisah C, Lebelo SL. Medicinal uses, pharmacological activities, phytochemistry, and the molecular mechanisms of Punica granatum L. (pomegranate) plant extracts: a review. Biomed Pharmacother. 2022;1(153):113256. doi:10.1016/j.biopha.2022.113256
17. Viuda-Martos M, Fernández-López J, Pérez-álvarez JA. Pomegranate and its many functional components as related to human health: a review. Compr Rev Food Sci Food Saf. 2010;9:635–654. doi:10.1111/j.1541-4337.2010.00131.x
18. Aviram M, Kaplan M, Rosenblat M, Fuhrman B. Dietary antioxidants and paraoxonases against LDL oxidation and atherosclerosis development. Handb Exp Pharmacol. 2005;170:263–300.
19. Debib A, Menadi S, Sahnouni„ F, Boukhatem MN, Ouahchia C, Kaced A. Bacterial inhibitory effect of Algerian pomegranate (Punica Granatum L.) extracts (Peel, Juice, And Seed) against multidrug resistant bacteria. J Microbiol Biotechnol Food Sci. 2022;11(4):e4622. doi:10.55251/jmbfs.4622
20. Lim HK, Asharani PV, Hande MP, et al. Cytotoxicity and genotoxicity of silver nanoparticles in human cells. ACS Nano. 2009;3(2):279–290. Pubmed: 19236062. doi:10.1021/nn800596w
21. Alshehri MA, Wierzbicki PM, Hassan F, et al. Mohamed. In vitro evaluation of electroporated gold nanoparticles and extremely-low frequency electromagnetic field anticancer activity against Hep-2 laryngeal cancer cells. Folia Histochem Cytobiol. 2019;57(4):159–167. doi:10.5603/FHC.a2019.0018
22. Burnette WN. ”Western blotting”: electrophoretic transfer of proteins from sodium dodecyl sulfate--polyacrylamide gels to unmodified nitrocellulose and radiographic detection with antibody and radioiodinated protein A. Anal Biochem. 1981;112:195–203. doi:10.1016/0003-2697(81)90281-5
23. Sambrook J, Fritsch EF, Maniatis T. Molecular cloning: a laboratory manual. In: Cold Spring Harbor Laboratory. New York: Cold Spring Harbor; 1989.
24. Longtin R. The pomegranate: nature’s power fruit? JNCI J Natl Cancer Inst. 2003;95:346–348. doi:10.1093/jnci/95.5.346
25. Howell AB, D’Souza DH. The pomegranate: effects on bacteria and viruses that influence human health. Evid Based Complement Alternat Med. 2013;2013:606212. doi:10.1155/2013/606212
26. Lansky EP, Newman RA. Punica granatum (pomegranate) and its potential for prevention and treatment of inflammation and cancer. J Ethnopharmacol. 2007;19(109):177–206. doi:10.1016/j.jep.2006.09.006
27. Ismail T, Sestili P, Akhtar S. Pomegranate peel and fruit extracts: a review of potential anti-inflammatory and antiinfective effects. J Ethnopharmacol. 2012;143:397–405. doi:10.1016/j.jep.2012.07.004
28. Tehran: Soroush publications 1362. Avicenna (Abue-Alisina). Cannon of Medicine (Qanon) II; 2021:306–307.
29. Lloyd JU. Punica granatum. Chicago, IL: The Western Druggist; 1897. Available from: http://www.swsbm.com/.ManualsOther/Punicagranatum-Lloyd.
30. Sharma P, McClees SF, Afaq F. Pomegranate for prevention and treatment of cancer: an update. Molecules. 2017;22:E177. doi:10.3390/molecules22010177
31. Adhami VM, Khan N, Mukhtar H. Cancer chemoprevention by pomegranate: laboratory and clinical evidence. Nutr Cancer. 2009;61:811–815. doi:10.1080/01635580903285064
32. Menezes SM, Cordeiro LN, Viana GS. Punica granatum (pomegranate) extract is active against dental plaque. J Herb Pharmacother. 2006;6:79–92. doi:10.1080/J157v06n02_07
33. Malik A, Mukhtar H. Prostate cancer prevention through pomegranate fruit. Cell Cycle. 2006;5:371–373.
34. Malik A, Afaq F, Sarfaraz S, et al. Pomegranate fruit juice for chemoprevention and chemotherapy of prostate cancer. Proc Natl Acad Sci U S A. 2005;102:14813–14818. doi:10.1073/pnas.0505870102
35. Amakura Y, Okada M, Tsuji S, Tonogai Y. High-performance liquid chromatographic determination with photodiode array detection of ellagic acid in fresh and processed fruits. J Chromatogr A. 2000;896:87–93. doi:10.1016/S0021-9673(00)00414-3
36. West T, Atzeva M, Holtzman DM. Pomegranate polyphenols and resveratrol protect the neonatal brain against hypoxic ischemic injury. Dev Neurosci. 2007;29:363–372. doi:10.1159/000105477
37. Quirynen M, Teughels W, Haake SK, Newman MG. Microbiology of periodontal diseases. In: Giannobile WV, editor. Carranza’s Clinical Periodontology.
38. Mehta VV, Rajesh G, Rao A, et al. Antimicrobial efficacy of Punica granatum mesocarp, Nelumbo nucifera Leaf, Psidium guajava Leaf and Coffea Canephora extract on common oral pathogens: an in vitro study. J Clin Diagn Res. 2014;8:ZC65–ZC68. doi:10.1152/ajplegacy.1975.229.3.570
39. Dahlawi H, Jordan-Mahy N, Clench M, et al. Polyphenols are responsible for the proapoptotic properties of pomegranate juice on leukemia cell lines. Food Sci Nutr. 2013;1:196–208. doi:10.1002/fsn3.26
40. Kim ND, Mehta R, Yu W, et al. Chemopreventive and adjuvant therapeutic potential of pomegranate (Punica granatum) for human breast cancer. Breast Cancer Res Treat. 2002;71:203–217. doi:10.1023/A:1014405730585
41. Adhami VM, Malik A, Zaman N, et al. Combined inhibitory effects of green tea polyphenols and selective cyclooxygenase-2 inhibitors on the growth of human prostate cancer cells both in vitro and in vivo. Clin Cancer Res. 2007;13:1611–1619.
42. Kawaii S, Lansky EP. Differentiation-promoting activity of pomegranate (Punica granatum) fruit extracts in HL-60 human promyelocytic leukemia cells. J Med Food. 2004;7:13–18. doi:10.1089/109662004322984644
43. Mehta R, Lansky EP. Breast cancer chemopreventive properties of pomegranate (Punica granatum) fruit extracts in a mouse mammary organ culture. Eur J Cancer Prev. 2004;13:345–348. doi:10.1097/01.cej.0000136571.70998.5a
44. Cortez-Trejo MC, Olivas-Aguirre FJ, Dufoo-Hurtado E, et al. Potential anticancer activity of pomegranate (Punica granatum L) fruits of different color: in vitro and in silico evidence. Biomolecules. 2022;12:1649. doi:10.3390/biom12111649
45. Mavlyanov SM, Islambekov SY, Karimdzhanov AK, Ismailov AI. Polyphenols of pomegranate peels show marked anti-tumor and anti-viral action. Chem Nat Compounds. 1997;33:98–99. doi:10.1007/BF02273936
46. Adams LS, Seeram NP, Aggarwal BB, et al. Pomegranate juice, total pomegranate ellagitannins, and punicalagin suppress inflammatory cell signaling in colon cancer cells. J Agric Food Chem. 2006;54:980–985. doi:10.1021/jf052005r
47. Tang Y, Zhao DY, Elliott S, et al. Epigallocatechin-3 gallate induces growth inhibition and apoptosis in human breast cancer cells through survivin suppression. Int J Oncol. 2007;31:705–711.
48. Jia L, Wang G, Hou C, Jianke L, Luo Y, Baicun L. Punicalagin and ellagic acid from pomegranate peel induce apoptosis and inhibits proliferation in human HepG2 hepatoma cells through targeting mitochondria. Food Agric Immunol. 2019;30(1):897–912. doi:10.1080/09540105.2019.1642857
49. Diplock AT, Charleux JL, Crozier-Willi G, et al. Functional food science and defence against reactive oxygen species. Br J Nutr. 1998;80(Suppl 1):S77–S112. doi:10.1079/BJN19980106
50. Tang Q, Huang G. Improving method, properties and application of charide as emulsifier. Food Chem. 2022;15(376):131937. doi:10.1016/j.foodchem.2021.131937
51. Bouayed J, Bohn T. Exogenous antioxidants—double-edged swords in cellular redoc state: health beneficial effects at physiologic doses versus deleterious effects at high doses. Oxid Med Cell Longev. 2010;3(4):228–237. doi:10.4161/oxim.3.4.12858
52. Song B, Jianke L, Jianke L. Pomegranate peel extract polyphenols induced apoptosis in human hepatoma cells by mitochondrial pathway. Food Chem Toxicol. 2016;93:158–166. doi:10.1016/j.fct.2016.04.0
53. He XY, Li MY, Zhao W, Liu L, Kong XH, Li JK. Chemical fingerprint and quantitative analysis for quality control of polyphenols extracted from pomegranate peel by HPLC. Food Chem. 2015;176:7–11. doi:10.1016/j.foodchem.2014.12.040
54. Larrosa M, Tomas-Barberan FA, Espin JC. The dietary hydrolysable tannin punicalagin releases ellagic acid that induces apoptosis in human colon adenocarcinoma Caco-2 cells by using the mitochondrial pathway. J Nutr Biochem. 2006;17:611–625. doi:10.1016/j.jnutbio.2005.09.004
55. Adamu BA, Emiru YK, Sintayehu B, Araya EM, Periasamy G, Hiben MG. In vivo hepatoprotective and in vitro radical scavenging activities of extracts of Rumex abyssinicus jacq Rhizome. J Exp Pharmacol. 2020;12:221. doi:10.2147/JEP.S258566
56. Alberts B, Bray D, Hopkin K, et al. Essential Cell Biology.
 © 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Eriochloa villosa Alleviates Progression of Benign Prostatic Hyperplasia in vitro and in vivo
Baek EB, Hwang YH, Park S, Hong EJ, Won YS, Kwun HJ
Research and Reports in Urology 2022, 14:313-326
Published Date: 24 September 2022
Zinc Oxide Particles Can Cause Ovarian Toxicity by Oxidative Stress in Female Mice Model

Xu Y, Zhao Y, Liu S, Lv S, Chen L, Wang W, Feng Y, Fu F, Xu H
International Journal of Nanomedicine 2022, 17:4947-4960
Published Date: 19 October 2022
TEAD1 Silencing Regulates Cell Proliferation and Resistance to 5-Fluorouracil in Cutaneous Squamous Cell Carcinoma
Wang Z, Liu M, Lei H, Xiao S, Zheng Y
Clinical, Cosmetic and Investigational Dermatology 2022, 15:2685-2692
Published Date: 13 December 2022
miR-98-5p Prevents Hippocampal Neurons from Oxidative Stress and Apoptosis by Targeting STAT3 in Epilepsy in vitro
Guo Z, Zhong W, Zou Z
Neuropsychiatric Disease and Treatment 2023, 19:2319-2329
Published Date: 30 October 2023
Remimazolam Suppresses Oxidative Stress and Apoptosis in Cerebral Ischemia/Reperfusion Injury by Regulating AKT/GSK-3β/NRF2 Pathway
Duan M, Yu N, Liu J, Zhao Y, Zhang J, Song S, Wang S
Drug Design, Development and Therapy 2025, 19:111-128
Published Date: 8 January 2025