Back to Journals » Diabetes, Metabolic Syndrome and Obesity » Volume 17
Expression Analysis of circRNAs in Human Adipogenesis
Authors Yi X, Wan X, Khan MA ![]() , Sun X, Wang Z, Chen K
, Sun X, Wang Z, Chen K ![]() , Peng L
, Peng L
Received 19 October 2023
Accepted for publication 12 December 2023
Published 3 January 2024 Volume 2024:17 Pages 45—54
DOI https://doi.org/10.2147/DMSO.S381603
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Konstantinos Tziomalos
Xuan Yi,1 Xinxing Wan,1 Md Asaduzzaman Khan,2,3 Xiaoying Sun,1 Zhouqi Wang,1 Ke Chen,1 Lin Peng4
1Department of Endocrinology, The Third Xiangya Hospital of Central South University, Changsha, People’s Republic of China; 2Research Division, Nature Study Society of Bangladesh, Dhaka, 1000, Bangladesh; 3Pulmonary Department, Chobanian & Avedisian School of Medicine, Boston University, Boston, MA, 02118, USA; 4Department of Nephrology, The First Hospital of Changsha, Changsha, People’s Republic of China
Correspondence: Lin Peng, Department of Nephrology, The First Hospital of Changsha, Changsha, People’s Republic of China, Tel +86-731-8466-7510, Email [email protected] Ke Chen, Department of Endocrinology, The Third Xiangya Hospital of Central South University, Changsha, People’s Republic of China, Tel +86-731-8861-8239, Email [email protected]
Purpose: Adipogenesis is one of the major pathways for generating obesity or overweight that can cause a range of metabolic disorders. Circular RNAs (circRNAs), a specific type of RNAs, have a significant influence on metabolic disorders. This study aims to find differentially expressed circRNAs (DECs) during human subcutaneous adipose tissue (SATs) adipogenesis.
Patients and Methods: The human adipose tissue-derived stromal cells (hADSCs) were isolated from human SATs (n = 3), and then induced into adipocytes. Total RNAs were extracted from hADSCs and adipocytes, and he DECs were detected using circRNA microarray. The GO and KEGG pathways of DECs were analyzed by bioinformatic methods, and partial DECs were further validated by quantitative polymerase chain reaction (qPCR).
Results: Our study detected a total of 1987 DECs, among which, 1134 were found upregulated and 853 were downregulated. GO analysis showed that the upregulated DECs have catalytic activity in intracellular organelle and cytoplasms, whereas downregulated DECs are enriched in organelle lumen, and are involved in positive regulation of developmental process. In addition, pathway results demonstrated that upregulated DECs are involved in platinum drug resistance and cellular senescence, and downregulated DECs are enriched in proteoglycans in cancer and focal adhesion pathway. Two circRNAs, namely has_circ_0001600 and has_circ_0001947 were validated to be significantly upregulated in adipocytes compared to hADSCs.
Conclusion: Our study explored DECs between hADSCs derived from SATs and adipocytes, and report that two circRNAs named has_circ_0001600 and has_circ_0001947 might be important factors involved in human adipogenesis, however, the molecular mechanism should be further explored.
Keywords: circRNA microarray, subcutaneous adipose tissue, human adipose tissue-derived stromal cells, adipogenesis, SAT
Introduction
Obesity is a chronic metabolic disease caused by excessive accumulation of adipose tissues and (or) abnormal fat distribution in the body.1 The human adipogenesis process includes human adipose tissue-derived stromal cells (hADSCs) differentiated to preadipocytes and further differentiated to mature adipocytes.2, 3 The abnormal hADSCs or preadipocytes proliferation and adipocytes hypertrophy lead to adipogenesis dysfunction are the pathological basis for the occurrence and development of obesity.4
Circular RNAs (circRNAs) are a category of endogenous regulatory RNA, of which, the precursor RNA forms a covalent closed-loop structure through the back-splicing site, which makes it difficult to be degraded by RNase R.5 circRNAs can act as a microRNAs(miRNAs) sponge, which in turn affects the combination with target mRNA and inhibits its activity, thus effectively changing its role in the post-transcriptional regulation of target genes expression.6, 7 In recent years, circRNAs have been recognized as key molecules in the metabolic diseases due to their regulatory effects on genes including the dynamic balance of sugars and lipids.8
There are some studies that explored circRNAs function in adipogenesis.9 However, only one study used circRNA microarray to analyze circRNAs expression profile from HPA-v cell, isolated from human visceral adipose tissues (VATs), differentiated to adipocytes.10 In addition, two other studies explored the mouse white and brown adipogenesis, respectively.11, 12 Furthermore, some studies assessed the differentially expressed circRNAs (DECs) of VATs in human obese compare to lean individuals and VATs compare to subcutaneous adipose tissue (SATs).8, 13 In other animals, for instance, the circRNAs expression of Yak and Pig of SATs adipogenesis were also evaluated.14, 15 Up to present, the circRNAs expression studies have been summarized by Zhang et al and Huang et al16, 17
Because of the single study information on explored circRNAs expression profile in human VATs adipogenesis,10 our aim was to clarify the circRNAs expression pattern in human SATs adipogenesis. Firstly, we used circRNA microarray to identify DECs from SATs derived hADSCs differentiated to adipocytes. Secondly, the expression of partially upregulated and downregulated circRNAs were verified by quantitative polymerase chain reaction (qPCR). Lastly, the bioinformatic analysis enabled the prediction of circRNA-miRNA-mRNA networks. Our research findings will offer fresh proof for studying the role of circRNAs in adipogenesis and provide novel targets for the treatment of obesity.
Materials and Methods
Human Tissue Specimens
The SATs were collected from three normal weight individuals (BMI:18.5–23.9 kg/m2) who underwent abdominal surgery at the Department of Gastrointestinal Surgery, Third Xiangya Hospital, Central South University. The individuals were eliminated from conditions like severe contagious diseases, autoimmune conditions, hematological disorders, malignant tumors, and more. The Ethical Committee of the Third Xiangya Hospital of Central South University in Changsha, China, approved this study, which followed the guidelines stated in the Declaration of Helsinki during its conduction. The consent of every patient was obtained in writing.
hADSCs Isolation and Differentiation
Approximately 10g of SATs were first washed with phosphate buffer saline (PBS) for three times. Then the sample underwent fragmentation into fragments less than 1 mm3, and digested at 37°C with 0.1% (w/v) collagenase I solution (Gibco, Life Technology, China) for 1.5 hours. The digestion was terminated with DMEM/F12 (Gibco, Life Technology, China) and filtered by 100μm cell strainer (NEST Biotechnology Co., Ltd., China), centrifuged at 800 rpm for 5 min, gently poured out the supernatant. Subsequently, 3mL red blood cell lysis buffer (Merck, China) was used to remove red blood cells, and after centrifuged at 800 rpm for 5 min again, the isolated cells were suspended in DMEM/F12 supplemented with 10% fetal bovine serum (FBS, Gibco, Life Technology, Australia), 100U/mL penicillin and 100 μg/mL streptomycin were added to the culture medium. The cells were then incubated at 37°C in an environment containing 5% CO2. In order to induce the hADSCs differentiated to adipocytes, when hADSCs grew to almost 100% confluency, the hADSCs were cultured in complete DMEM/F12 medium which contained 1μM dexamethasone, 10μM insulin, 0.5mM isobutylmethylxanthine (IBMX), and 200μM indomethacin (four reagents were purchased from Merck, China); the culture medium was replaced every 48 hours.
circRNA Microarray Analysis
The circRNA microarray analysis was performed in accordance with our previous studies.18, 19 Briefly, total RNA was obtained from human hADSCs and adipocytes using the Trizol (Invitrogen, Carlsbad, CA, USA) method, and then, the linear RNA samples were removed by treating with RNase R (Epicentre, Inc., WI, USA), the circRNAs were collected and amplified. The fluorescent complementary RNAs (cRNAs) were transcribed and labeled using a random primer approach (Arraystar, MD, USA). The presence of cRNAs in the Agilent G2565CA Microarray Scanner System (Agilent Technologies, CA, USA) was confirmed. Fold-change filtering identified DECs between two groups. The Gene ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway analysis were carried out.
circRNAs Verification
The 4 most upregulated and 2 most downregulated circRNAs in DECs data were validated using RT-qPCR. Briefly, RNA was isolated from hADSCs and adipocytes, and then was reverse transcribed into cDNA (Invitrogen, Carlsbad, CA, USA). The relative quantification of circRNAs expression using qPCR was calculated by 2−∆∆ct method. The primers were designed with circinteractome software (https://circinteractome.irp.nia.nih.gov/index.html), and all premiers are listed in Supplementary Table 1.
circRNA-miRNA-mRNA Networks Construction
At first, the circRNAs and miRNAs interaction was analyzed by TargetScan and miRanda. Then, the DIANA tools enabled the prediction of the interaction between miRNAs and mRNA. And finally, the Cytoscape 2.8.2 software was utilized to generate a visual representation of the circRNA-miRNA-mRNA network.
Statistics Analysis
The data underwent analysis utilizing GraphPad Prism 10 software, after the data was verified to conform to a normal distribution; the data from two groups were compared using a t-test, where the significance was defined as p<0.05.
Results
The DECs Analysis of hADSCs and Adipocytes
The DECs profiles between hADSCs and adipocytes were investigated by human circRNA microarray. The Box Plot result revealed that the log2 ratio distribution was similar in 6 samples (Figure 1A). There were 1987 DECs in two group, among them, 1134 DECs were upregulated and 853 DECs were downregulated in adipocytes compare to hADSCs. The most significantly upregulated and downregulated 10 circRNAs are showed in Table 1 and Table 2, respectively. The Hierarchical Clustering showed partially significant difference in two groups (Figure 1B). In addition, the partial DECs were filtered according to the fold change ≥0.5 and p values <0.05. The results are showed in Scatter Plot and Volcano Plot (Figure 1C and D). Furthermore, the chromosome distribution of DECs is displayed in Figure 1E.
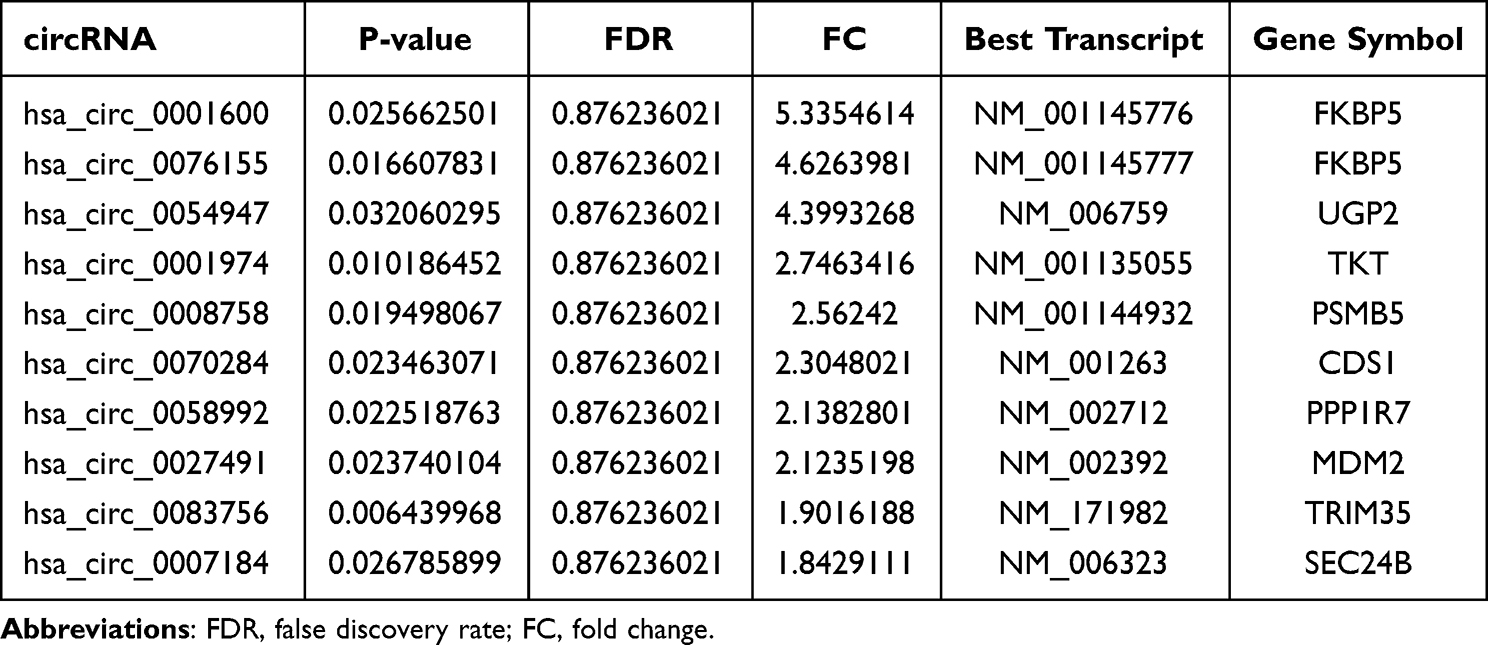 |
Table 1 Upregulated of circRNAs in Human Adipocytes Vs hADSCs |
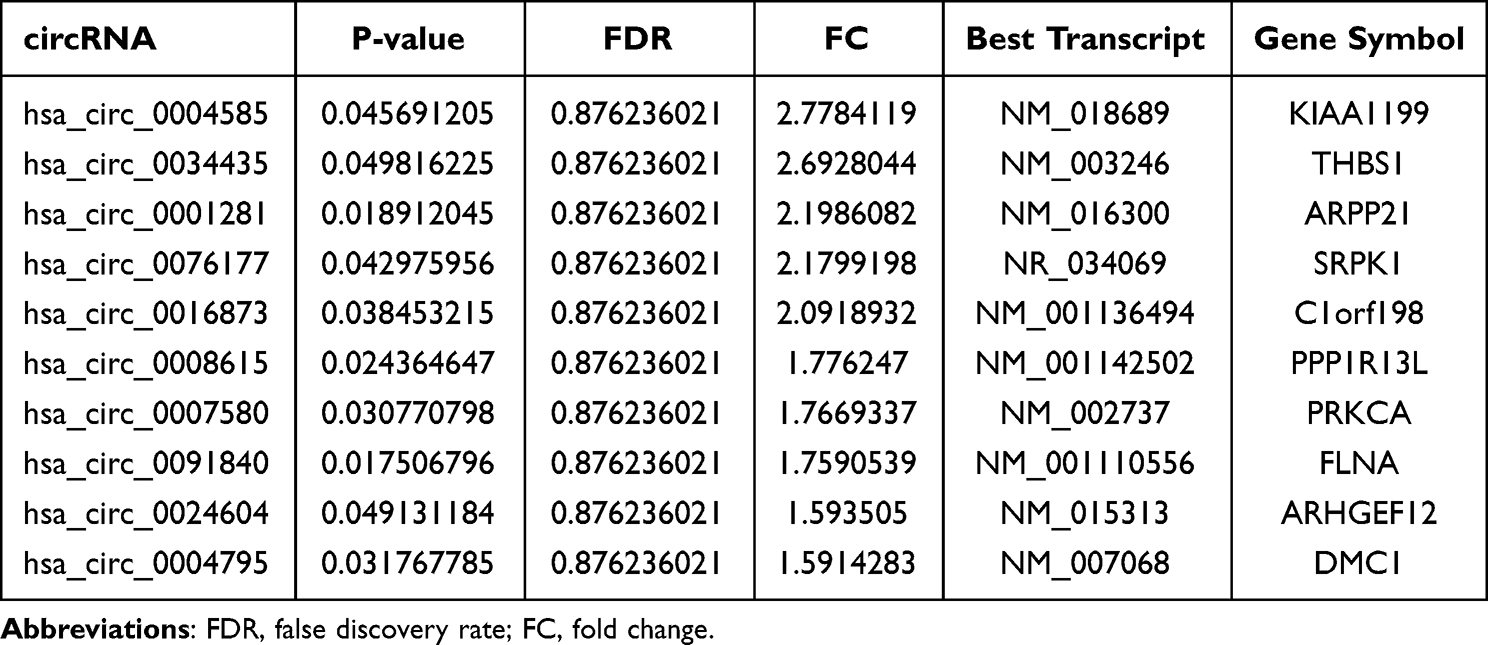 |
Table 2 Downregulated of circRNAs in Human Adipocytes Vs hADSCs |
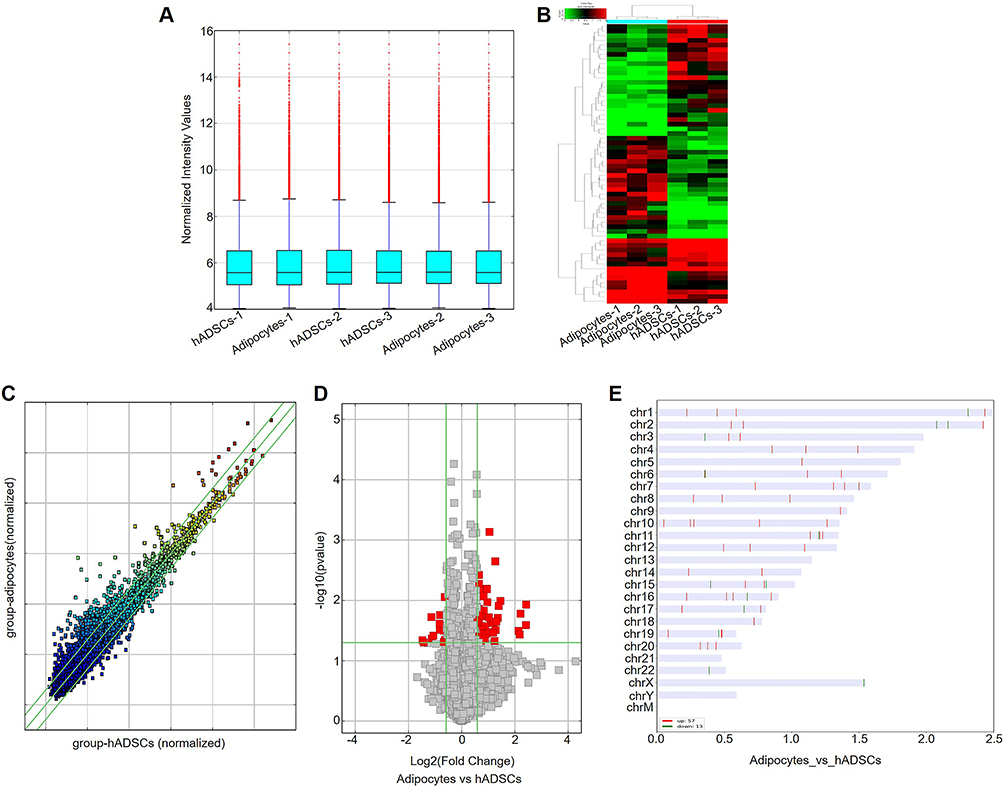 |
Figure 1 DECs in hADSCs and adipocytes. (A) The box plot depicted the distribution of normalized intensity for the entire data set. (B) Hierarchical clustering analysis was performed to generate maps of DECs in hADSCs and adipocytes. Red indicates a relatively elevated level of expression. Green indicates a relatively low level of expression. (C) Scatter plot was utilized to evaluate alterations in the expression of circRNAs between hADSCs and adipocytes. The presence of circRNAs above the upper green threshold and below the lower green threshold signifies a quantity greater than 1.5-fold DECs between the two samples being compared. (D) The volcano plot revealed a substantial fold change indicating statistically significant differential expression comparisons between hADSCs and adipocytes (fold change >0.5, P<0.05). (E) Characterization of the classification and distribution of DEGs across human chromosomes. |
GO and KEGG Pathway Analysis of the DECs
The GO and KEGG pathway were analyzed according to the host genes of circRNAs. The GO results of upregulation of DECs demonstrated that intracellular organelle, cytoplasm and catalytic activity were significantly enriched. In addition, in Biological Process (BP) analysis, some host genes of circRNAs were found related to steroid metabolic process, regulation of steroid biosynthetic process, cellular response to fatty acid, etc. (Figure 2A). In downregulation of DECs, organelle lumen, positive regulation of developmental process and membrane-enclosed lumen, etc. were enriched (Figure 2B). The pathway analysis of upregulated circRNAs were enriched in platinum drug resistance, cellular senescence and bladder cancer (Figure 2C), however, in downregulated circRNAs, proteoglycans in cancer, focal adhesion and PI3K-Akt signaling pathway were significantly enriched (Figure 2D).
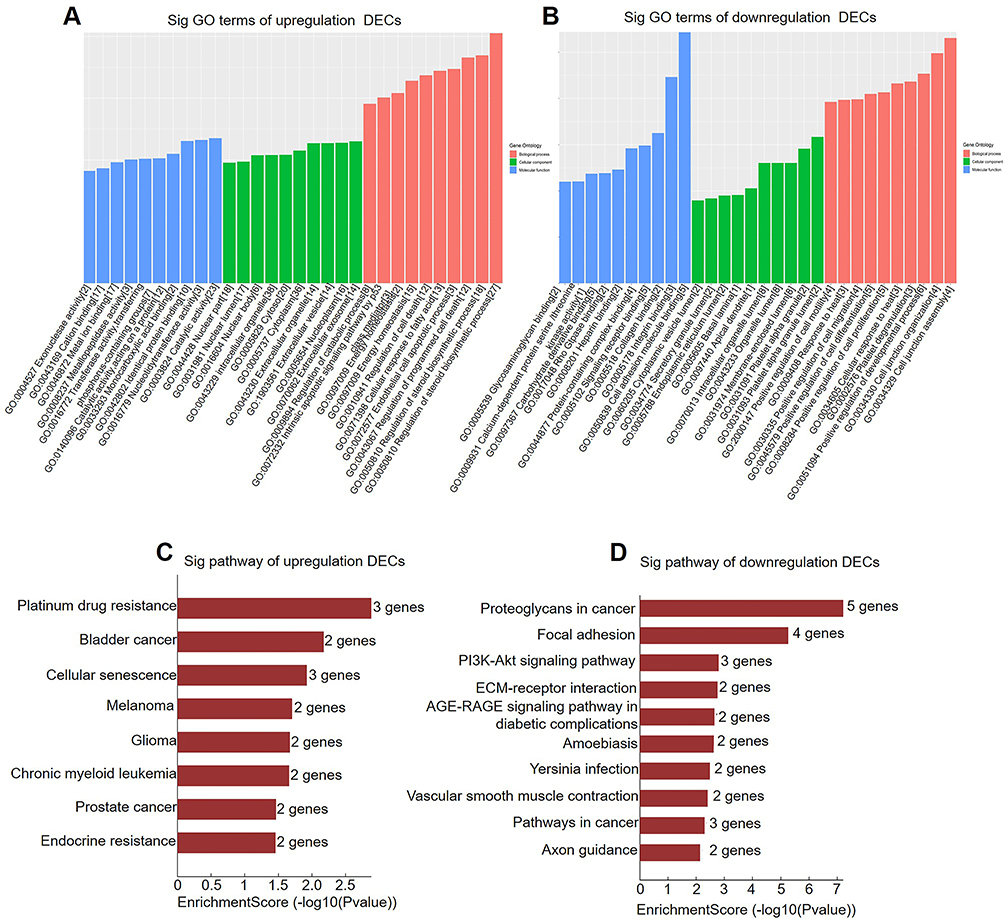 |
Figure 2 GO and KEGG pathway analysis of DECs according to host genes in hADSCs and adipocytes. GO analysis was used to compare the expression of upregulated (A) and downregulated (B) DECs in adipocytes versus hADSCs. KEGG pathway analysis of upregulated (C) and downregulated (D) DECs in adipocytes versus hADSCs. |
Partial DECs Validation by qPCR
To confirm our circRNA microarray results, the 4 most upregulated and 2 most downregulated circRNAs were chosen for qPCR validation. hsa_circ_0001600 and hsa_circ_0001974 were significantly upregulated in adipocytes compare to hADSCs (Figure 3A and B), however, there was no notable difference observed for hsa_circ_0076155 and hsa_circ_0054947 (Figure 3C and D). Among downregulated circRNAs, hsa_circ_0004585 and hsa_circ_0034435 also showed no significant difference (Figure 3E and F). The back-splice junction sequence of hsa_circ_0001600 and hsa_circ_0001974 were further verified using Sanger sequencing of PCR product (Figure 3G and H).
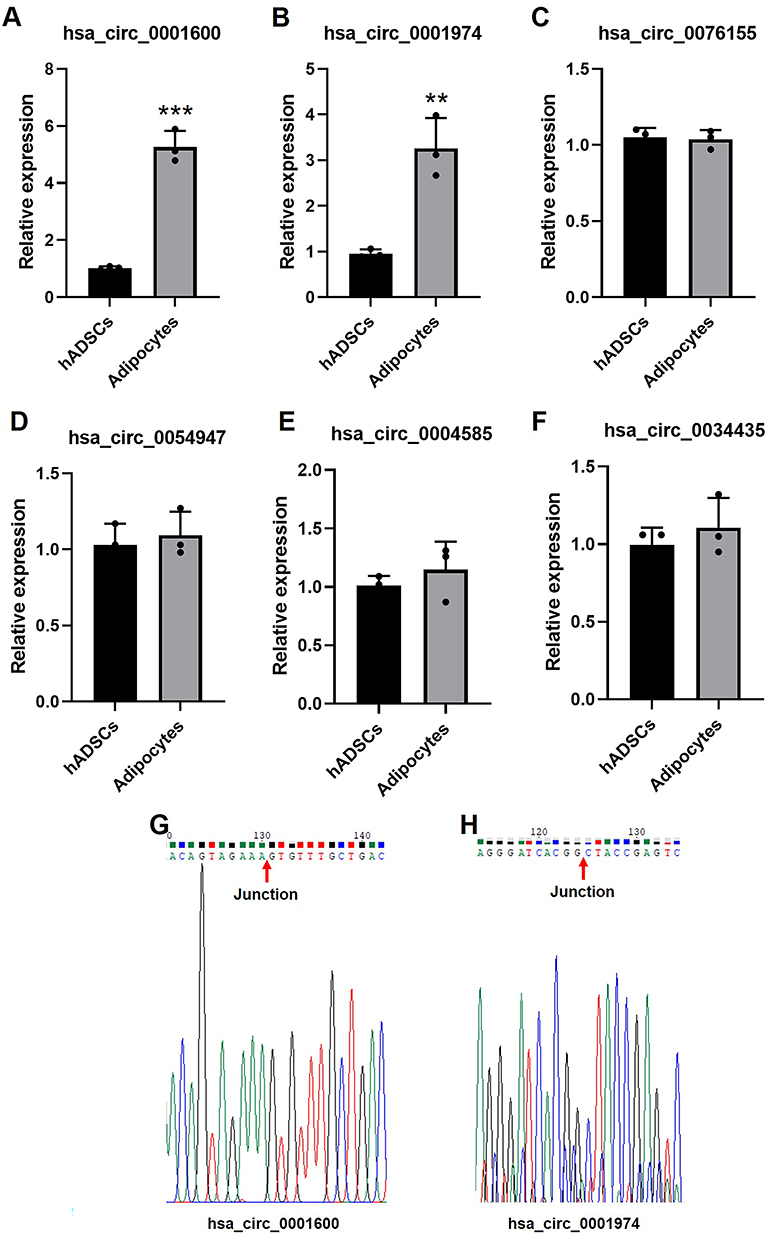 |
Figure 3 Validate of partial DECs by qPCR. The expression of has_circ_0001600 (A), has_circ_0001947 (B), has_circ_0076155 (C), has_circ_0054947 (D), has_circ_0004585 (E) and has_circ_0034435 (F) in hADSCs and adipocytes, n = 3, **p<0.01, ***p<0.001. Sanger sequencing map of hsa_circ_0001600 (G) and hsa_circ_0001974 (H). The red arrow represents back-splice junction sequence of circRNAs. |
Prediction of circRNA-miRNA-mRNA Interaction Networks
In order to gain a deeper comprehension of the potential possible molecular mechanism of has_circ_0001600 and has_circ_0001947 in adipogenesis, a construction of circRNA-miRNA-mRNA interaction networks was carried out using TargetScan, miRanda, and DIANA TOOLS software. The results indicated that certain miRNAs (hsa-miR-619-5p, hsa-miR-20b-3p, hsa-miR-367-3p, hsa-miR-153-5p, and hsa-miR-665) potentially target has_circ_0001600. Moreover, hsa-miR-668-3p, hsa-miR-194-3p, hsa-miR-182-5p, hsa-miR-588 and hsa-miR-494-5p were predicted as target miRNAs of has_circ_0001947 (Figure 4).
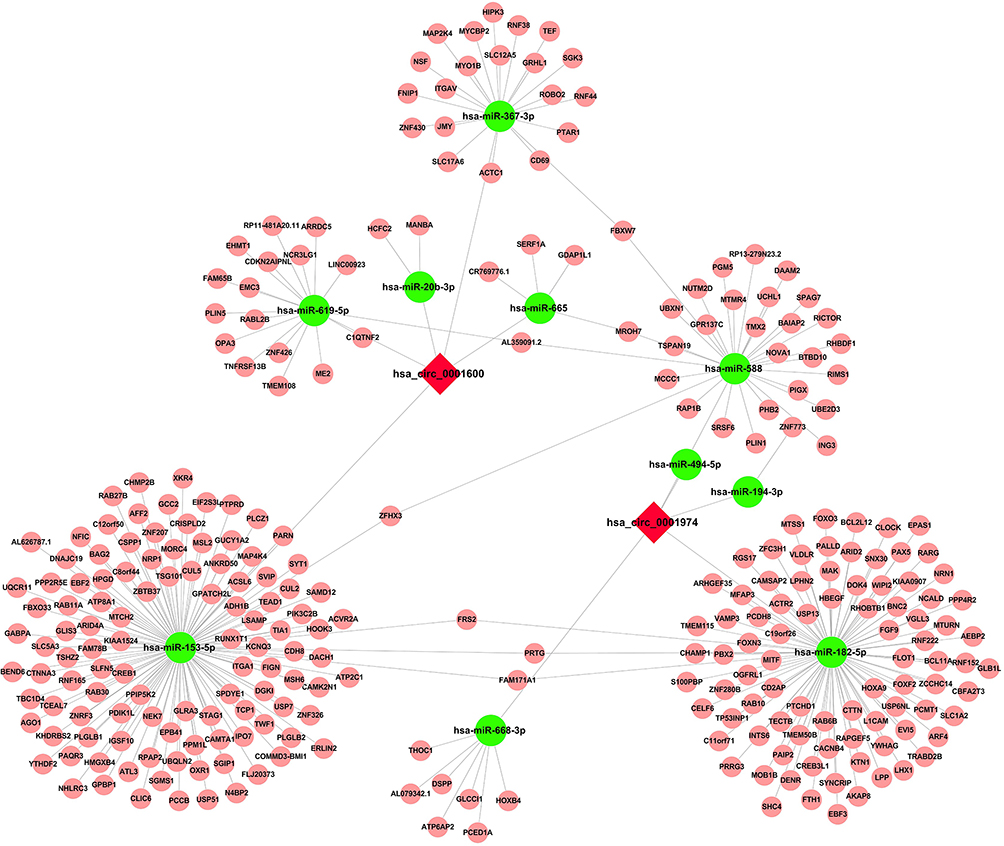 |
Figure 4 Prediction of hsa_circ_0001600 and hsa_circ_0001974 -miRNA-mRNA interaction network. Predictions were made about the interaction of circRNAs and miRNAs using TargetScan and miRanda software, while the interaction between miRNA and mRNA was predicted using DIANA software. |
Discussion
Increased adipose tissues are strongly associated with some metabolic diseases, for instance, hypertension, diabetes, hyperlipidemia, etc.20 In a previous study, Sun et al have detected circRNAs expression during the human preadipocytes derived from VATs differentiated to adipocytes,10 and in this study, we explored human SATs adipogenesis according to hADSCs differentiated to adipocytes.
Our findings indicate significant difference of circRNAs expression between SATs and VATs; the data differences might be due to different race, gender, age, or limited sample size. In our study, 1987 DECs between hADSCs and adipocytes were identified, and among them, 1134 DECs were found upregulated and 853 DECs were found downregulated. In GO analysis, intracellular organelle, cytoplasm and catalytic activity were found significantly enriched, especially in BP, some lipid metabolic related processes including steroid metabolic process, regulation of steroid biosynthetic process, cellular response to fatty acid, etc. also were enriched. Thus, our results demonstrated that some circRNAs might play important role in adipogenesis or lipogenesis.
A series of studies have confirmed the molecular mechanisms of some circRNAs in adipogenesis, for instances, hsa_circH19 is highly expressed in blood of metabolic syndrome patients, and silencing of hsa_circH19 promoted hADSCs adipogenesis via regulation of PTBP1.21 Moreover, circSAMD4A showed positive correlation and upregulated in VATs of obese patients, and knockdown of circSAMD4A blocked human preadipocytes adipogenesis through sponging with miR-138-5p and further repressing EZH2.13 Besides, decrease of circRNA CDR1as attenuate human bone marrow mesenchymal stem cells (BMSCs) adipogenesis but enhance osteogenic differentiation through regulation of miR-7-5p/WNT5B signaling.22 In addition, in bovine model, circFUT10 prevented adipogenesis and circFLT1 accelerated adipogenesis via regulation of let-7c/let-e and miR-93, respectively.23, 24 Overall, the recent studies have been summarized by Zhang et al and Huang et al16, 17
In our study, hsa_circ_0001600 (circBank ID: hsa_circFKBP5_002) and hsa_circ_0001974 (circBank ID: hsa_circTKT_008) were validated and found to be dramatically upregulated in adipocytes compared to hADSCs. In mouse model, both of circ_FKBP5 and its host gene, FKBP prolyl isomerase 5 (FKBP5), were validated to be upregulated during adipogenesis,11 although it was not clear whether hsa_circ_0001600 participate in adipogenesis, but FKBP5 was an important factor to regulate lipolysis, lipogenesis and adipogenesis. FKBP5 was found upregulated in SATs of Type 2 diabetes compared to non-diabetic individuals and positive correlation with blood sugar level.24 FKBP5 of SATs has significant positive relation with adipocytes size and HOMA-IR and blood insulin level.25 Knockout of FKBP5 inhibited mice adipogenesis and fat-diet induced obesity in normoxia and hypoxic stress state.26
In miRNA prediction target of hsa_circ_0001600, miR-20b was found increased in obesity with normal blood sugar compared to obesity with type 2 diabetes in VATs.27 miR-20b-5p derived from serum exosomal repress 3T3-L1 adipogenesis,28 however, the expression of hsa-miR-665 was found decreased during adipogenesis.29 So, we speculate that hsa_circ_0001600 might regulate adipogenesis via miR-20b-5p or hsa-miR-665.
Although hsa_circ_0001974 is not being reported to participate in adipogenesis, its host gene, transketolase (TKT), is a critical molecule to regulate lipolysis, and knockout of TKT in mice adipose tissue decreased fat-diet induced obesity through increase lipolysis.30 In addition, TKT was found downregulated in omental tissues of obese vs non-obese and upregulated during 3T3-L1 adipogenesis.31, 32 In miRNA target prediction of hsa_circ_0001974, hsa-miR-194 impedes adipogenesis through COUP-TFII axis in murine C3H10T1/2 mesenchymal cells and primary bone marrow stromal cells.33 Moreover, miR-182 acts as negative regulatory factor that prevented 3T3-L1 and human preadipocytes adipogenesis through combining with CCAAT/enhancer-binding protein α (C/EBPα).34 In another study, miR-182 impaired adipogenesis in Graves’ orbitopathy through regulation of thyrotropin receptor.35 miR-182 also could increase beige adipose tissue thermogenesis and block obesity.36 In mice beige adipocytes, miR-494-3p was downregulated after cold exposure and enhance of miR-494-3p decreased thermogenesis.37 So, we postulate that hsa_circ_0001974 might decrease hsa-miR-194 or miR-182 to promote adipogenesis and inhibit miR-494-3p to increase thermogenesis. The more clarified molecular mechanism should be further explored.
Conclusion
In this study, we performed circRNA microarray analysis, and detected hsa_circ_0001600 and hsa_circ_0001974 elevated in adipocytes vs hADSCs. hsa_circ_0001600 might promote adipogenesis via decreasing miR-20b-5p or hsa-miR-66, and hsa_circ_0001974 might induce adipogenesis through inhibiting hsa-miR-194 or miR-182. Our study provides a couple of novel targets for treating adipogenesis or obesity.
Abbreviations
circRNAs, circular RNAs; DECs, differentially expressed circRNAs; SATs, subcutaneous adipose tissue; hADSCs, human adipose tissue-derived stromal cells; qPCR, quantitative polymerase chain reaction; miRNAs, microRNAs; cRNAs, fluorescent complementary RNAs; GO, gene ontology; KEGG, Kyoto encyclopedia of genes and genomes; BMSCs, human bone marrow mesenchymal stem cells; C/EBPα, CCAAT/enhancer-binding protein α.
Funding
This study received partial funding from the Hunan Province Natural Science Foundation of China (Grant no. 2022JJ30881 and 2022JJ70045).
Disclosure
The authors affirm that there are no competing interests relating to this study.
References
1. Xiao Y, Liu D, Cline MA, et al. Chronic stress, epigenetics, and adipose tissue metabolism in the obese state. Nutr Metab. 2020;17(1):88. doi:10.1186/s12986-020-00513-4
2. Kurylowicz A. microRNAs in human adipose tissue physiology and dysfunction. Cells. 2021;10(12):3342. doi:10.3390/cells10123342
3. Dufau J, Shen JX, Couchet M, et al. In vitro and ex vivo models of adipocytes. Am J Physiol Cell Physiol. 2021;320(5):C822–C841. doi:10.1152/ajpcell.00519.2020
4. He X, Guan H, Liang W, et al. Exendin-4 modifies adipogenesis of human adipose-derived stromal cells isolated from omentum through multiple mechanisms. Int J Obes. 2018;42(5):1051–1061. doi:10.1038/s41366-018-0024-2
5. Li J, Sun D, Pu W, et al. Circular RNAs in cancer: biogenesis, function, and clinical significance. Trends Cancer. 2020;6(4):319–336. doi:10.1016/j.trecan.2020.01.012
6. Memczak S, Jens M, Elefsinioti A, et al. Circular RNAs are a large class of animal RNAs with regulatory potency. Nature. 2013;495(7441):333–338. doi:10.1038/nature11928
7. Huang A, Zheng H, Wu Z, et al. Circular RNA-protein interactions: functions, mechanisms, and identification. Theranostics. 2020;10(8):3503–3517. doi:10.7150/thno.42174
8. Arcinas C, Tan W, Fang W, et al. Adipose circular RNAs exhibit dynamic regulation in obesity and functional role in adipogenesis. Nat Metab. 2019;1(7):688–703. doi:10.1038/s42255-019-0078-z
9. Chen C, Zhang X, Deng Y, et al. Regulatory roles of circRNAs in adipogenesis and lipid metabolism emerging insights into lipid related diseases. FEBS J. 2021;288(12):3663–3682. doi:10.1111/febs.15525
10. Sun W, Sun X, Chu W, et al. CircRNA expression profiles in human visceral preadipocytes and adipocytes. Mol Med Rep. 2020;21(2):815–821. doi:10.3892/mmr.2019.10886
11. Zhang PP, Han Q, Sheng MX, et al. Identification of circular RNA expression profiles in white adipocytes and their roles in adipogenesis. Front Physiol. 2021;12:728208. doi:10.3389/fphys.2021.728208
12. Zhang P, Sheng M, Du C, et al. Assessment of circRNA expression profiles and potential functions in brown adipogenesis. Front Genet. 2021;12:769690. doi:10.3389/fgene.2021.769690
13. Liu Y, Liu H, Li Y, et al. Circular RNA SAMD4A controls adipogenesis in obesity through the miR-138-5p/EZH2 axis. Theranostics. 2020;10(10):4705–4719. doi:10.7150/thno.42417
14. Zhang Y, Guo X, Pei J, et al. CircRNA expression profile during yak adipocyte differentiation and screen potential circRNAs for adipocyte differentiation. Genes. 2020;11(4):414. doi:10.3390/genes11040414
15. Liu X, Liu K, Shan B, et al. A genome-wide landscape of mRNAs, lncRNAs, and circRNAs during subcutaneous adipogenesis in pigs. J Anim Sci Biotechnol. 2018;9(1):76. doi:10.1186/s40104-018-0292-7
16. Zhang Y, Tian Z, Ye H, et al. Emerging functions of circular RNA in the regulation of adipocyte metabolism and obesity. Cell Death Discov. 2022;8(1):268. doi:10.1038/s41420-022-01062-w
17. Huang CJ, Choo KB. Circular RNA- and microRNA-mediated post-transcriptional regulation of preadipocyte differentiation in adipogenesis from expression profiling to signaling pathway. Int J Mol Sci. 2023;24(5):4549. doi:10.3390/ijms24054549
18. Sun X, Wan X, Khan MA, et al. Comprehensive analysis of circRNA expression profiles in human brown adipose tissue. Diabetes Metab Syndr Obes. 2023;16:469–478. doi:10.2147/DMSO.S398620
19. Zhang K, Wan X, Khan MA, et al. Peripheral blood circRNA microarray profiling identities hsa_circ_0001831 and hsa_circ_0000867 as two novel circRNA biomarkers for early type 2 diabetic nephropathy. Diabetes Metab Syndr Obes. 2022;15:2789–2801. doi:10.2147/DMSO.S384054
20. Koenen M, Hill MA, Cohen P, et al. Obesity, adipose tissue and vascular dysfunction. Circ Res. 2021;128(7):951–968. doi:10.1161/CIRCRESAHA.121.318093
21. Zhu Y, Gui W, Lin X, et al. Knock-down of circular RNA H19 induces human adipose-derived stem cells adipogenic differentiation via a mechanism involving the polypyrimidine tract-binding protein 1. Exp Cell Res. 2020;387(2):111753. doi:10.1016/j.yexcr.2019.111753
22. Chen G, Wang Q, Li Z, et al. Circular RNA CDR1as promotes adipogenic and suppresses osteogenic differentiation of BMSCs in steroid-induced osteonecrosis of the femoral head. Bone. 2020;133:115258. doi:10.1016/j.bone.2020.115258
23. Jiang R, Li H, Yang JM, et al. circRNA profiling reveals an abundant circFUT10 that promotes adipocyte proliferation and inhibits adipocyte differentiation via sponging let-7. Mol Ther Nucleic Acids. 2020;20:491–501. doi:10.1016/j.omtn.2020.03.011
24. Sidibeh CO, Pereira MJ, Abalo XM, et al. FKBP5 expression in human adipose tissue: potential role in glucose and lipid metabolism, adipogenesis and type 2 diabetes. Endocrine. 2018;62(1):116–128. doi:10.1007/s12020-018-1674-5
25. Pereira MJ, Palming J, Svensson MK, et al. FKBP5 expression in human adipose tissue increases following dexamethasone exposure and is associated with insulin resistance. Metabolism. 2014;63(9):1198–1208. doi:10.1016/j.metabol.2014.05.015
26. Zhang L, Qiu B, Wang T, et al. Loss of FKBP5 impedes adipocyte differentiation under both normoxia and hypoxic stress. Biochem Biophys Res Commun. 2017;485(4):761–767. doi:10.1016/j.bbrc.2017.02.126
27. Gentile AM, Lhamyani S, Coín-Aragüez L, et al. miR-20b, miR-296, and let-7f expression in human adipose tissue is related to obesity and type 2 diabetes. Obesity. 2019;27(2):245–254. doi:10.1002/oby.22363
28. Hong Y, Wu J, Yu S, et al. Serum-derived exosomal microRNAs in lipid metabolism in polycystic ovary syndrome. Reprod Sci. 2022;29(9):2625–2635. doi:10.1007/s43032-022-00930-1
29. Yi X, Liu J, Wu P, et al. The key microRNA on lipid droplet formation during adipogenesis from human mesenchymal stem cells. J Cell Physiol. 2020;235(1):328–338. doi:10.1002/jcp.28972
30. Tian N, Liu Q, Li Y, et al. Transketolase deficiency in adipose tissues protects mice from diet-induced obesity by promoting lipolysis. Diabetes. 2020;69(7):1355–1367. doi:10.2337/db19-1087
31. Pérez-Pérez R, García-Santos E, Ortega-Delgado FJ, et al. Attenuated metabolism is a hallmark of obesity as revealed by comparative proteomic analysis of human omental adipose tissue. J Proteomics. 2012;75(3):783–795. doi:10.1016/j.jprot.2011.09.016
32. Calzadilla P, Gómez-Serrano M, García-Santos E, et al. N-acetylcysteine affects obesity-related protein expression in 3T3-L1 adipocytes. Redox Rep. 2013;18(6):210–218. doi:10.1179/1351000213Y.0000000066
33. Jeong BC, Kang IH, Hwang YC, et al. MicroRNA-194 reciprocally stimulates osteogenesis and inhibits adipogenesis via regulating COUP-TFII expression. Cell Death Dis. 2014;5(11):e1532. doi:10.1038/cddis.2014.485
34. Dong M, Ye Y, Chen Z, et al. MicroRNA 182 is a novel negative regulator of adipogenesis by targeting ccaat/enhancer-binding protein α. Obesity. 2020;28(8):1467–1476. doi:10.1002/oby.22863
35. Wang S, Chen L, Xu B. ELF3-induced miR-182 inhibits adipogenic differentiation in by targeting thyrotropin receptor. Cent Eur J Immunol. 2022;47(4):308–322. doi:10.5114/ceji.2022.124087
36. Meng W, Xiao T, Liang X, et al. The miR-182-5p/FGF21/acetylcholine axis mediates the crosstalk between adipocytes and macrophages to promote beige fat thermogenesis. JCI Insight. 2021;6(17):e150249. doi:10.1172/jci.insight.150249
37. Lemecha M, Morino K, Imamura T, et al. MiR-494-3p regulates mitochondrial biogenesis and thermogenesis through PGC1-α signalling in beige adipocytes. Sci Rep. 2018;8(1):15096. doi:10.1038/s41598-018-33438-3
 © 2024 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2024 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.