Back to Journals » Journal of Inflammation Research » Volume 18
Exploration Progress on Inflammatory Responses and Immune Regulatory Mechanisms in Diabetic Retinopathy
Authors Cheng S ![]() , Chen Z
, Chen Z ![]() , Peng F, Deng P, Wang M, Liu S, Du Y, Zuo G
, Peng F, Deng P, Wang M, Liu S, Du Y, Zuo G ![]()
Received 12 March 2025
Accepted for publication 20 August 2025
Published 28 August 2025 Volume 2025:18 Pages 11895—11909
DOI https://doi.org/10.2147/JIR.S527784
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 5
Editor who approved publication: Dr Justin Jacob
Shizhou Cheng,1,* Zijian Chen,2,* Fan Peng,1,* Pan Deng,1 Mengni Wang,1 Sili Liu,1 Yanhua Du,3 Guojin Zuo1
1Department of Ophthalmology, the First Affiliated Hospital of Yangtze University, Jingzhou, Hubei Province, 434000, People’s Republic of China; 2Department of Ophthalmology, The Affiliated Eye Hospital,Jiangxi Medical College, Nanchang University, Nanchang City, Jiangxi Province, 330000, People’s Republic of China; 3Physical Examination Department, the First Affiliated Hospital of Yangtze University, Jingzhou, Hubei Province, 434000, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Yanhua Du, Email [email protected] Guojin Zuo, Email [email protected]
Abstract: Diabetic retinopathy (DR) is one of the most common microvascular complications of diabetes. In recent years, with the number of diabetic patients increasing annually, the number of DR patients has also been rising, and it has become one of the major blinding eye diseases worldwide. The pathogenesis of DR has not been fully clarified but includes microvascular lesions and metabolic factors. However, with the continuous exploration of the pathogenesis of DR, immune system disorders and inflammation have also been found to be important pathogenic mechanisms of DR and research on inflammation and immune mechanisms in DR have received increasing attention. This article reviews aspects such as the activation of inflammatory cells, the expression of inflammatory factors, and the complement system and explores the important roles of inflammation and immune mechanisms in the pathogenesis of DR, providing a theoretical basis for a deeper understanding of DR and potential explorations of new therapeutic strategies for DR.
Keywords: diabetic retinopathy, diabetes, inflammation, immune mechanism, research progress, macular degeneration, proliferative diabetic retinopathy
Introduction
Diabetes is currently the most common metabolic disease globally.1,2 Diabetic retinopathy (DR) is a serious and common complication of diabetes and a leading cause of blindness.3 It affects people of all ages worldwide. The prevalence rate is 34.6% (93 million) among adults aged 40 and above.4 A systematic review focusing on population-based studies estimated that the annual incidence rate of DR increased from 2.2% to 12.7%.5 Almost all patients with type 1 diabetes will experience some degree of DR within 20 years after disease onset. In contrast, almost 60% of patients with type 2 diabetes will experience DR within the same time frame.6 Failure to treat DR in a timely fashion will lead to its progression from a mild, nonproliferative form to moderate and severe forms. These, in turn, will further develop into proliferative DR (PDR), and severe cases will progress to neovascular glaucoma and other serious complications.7,8
To date, the pathogenesis of DR has not been completely elucidated. Previous studies attributed it to microvascular lesions and metabolic factors.9 Meanwhile, various factors, including oxidative stress, mitochondrial dysfunction, epigenetic modifications of genes, and aberrant expression of non-coding RNAs, also contribute substantially to the onset and progression of DR.10 Retinal laser photocoagulation, vitrectomy, and intravitreal injections of anti-vascular endothelial growth factor (VEGF) drugs are the main pharmacological options used to treat DR. Although the prognosis of patients with DR has improved significantly, problems such as a lack of responsiveness to treatment, recurrence, and long-term decline in vision continue to pose a challenge. Current studies indicated the important role of inflammation and immune mechanisms in DR.11–13 This review focuses on exploring the role of immunity, inflammation-related cell activation, systemic inflammatory biomarkers, and the upregulation of inflammation-related factors in the pathogenesis of DR, providing new insight and ideas for treating DR.
Microvascular Changes and Blood-Retinal Barrier Injury
DR is a microvascular complication of diabetes (Figure 1). Blood glucose levels play an important role in the occurrence and progression of DR.14 The presence of DR indicates a compromise in microcirculation due to the diabetic environment, especially cardiovascular and cerebrovascular diseases, and other peripheral complications.15 While poor blood glucose control is directly related to the long-term development and deterioration of DR, the initial onset of DR results from a persistent state of hyperglycemia. Significant improvements have been made in the treatment and management of diabetes; however, DR remains the main cause of visual impairment and blindness.16 The retina is protected by a highly complex immune mechanism, and the blood-retinal barrier (BRB) is the first line of defense, offering retinal immune protection. The BRB is composed of the blood-retinal inner barrier (tight connections between retinal endothelial cells) and the blood-retinal outer barrier (tight connection between retinal pigment epithelial cells located on Bruch’s membrane).17 The BRB isolates the retina from exogenous pathogens. Retinal immunosuppression is triggered once the BRB is compromised and the second and third lines of immune protection are initiated.18 Inflammation is a symptom in the pathogenesis of DR that presents in different stages of DR, eventually leading to capillary atrexia, retinal nonperfusion, and hypoxia. These events finally lead to endothelial cell damage and, eventually, extensive BRB damage. The function of the retinal immune system is also severely damaged. Numerous circulating immune cells infiltrate the retina, leading to progressive degeneration and the death of retinal neurons.19,20 DR in patients with diabetes is not only associated with vision problems; these patients are also at high risk of developing other microvascular and macrovascular complications. DR is a condition that is often not considered when developing personalized treatment plans for diabetes. Coordinated activities between ophthalmologists and diabetes specialists in a clinical setting are particularly important to ameliorate the devastating outcomes of DR.
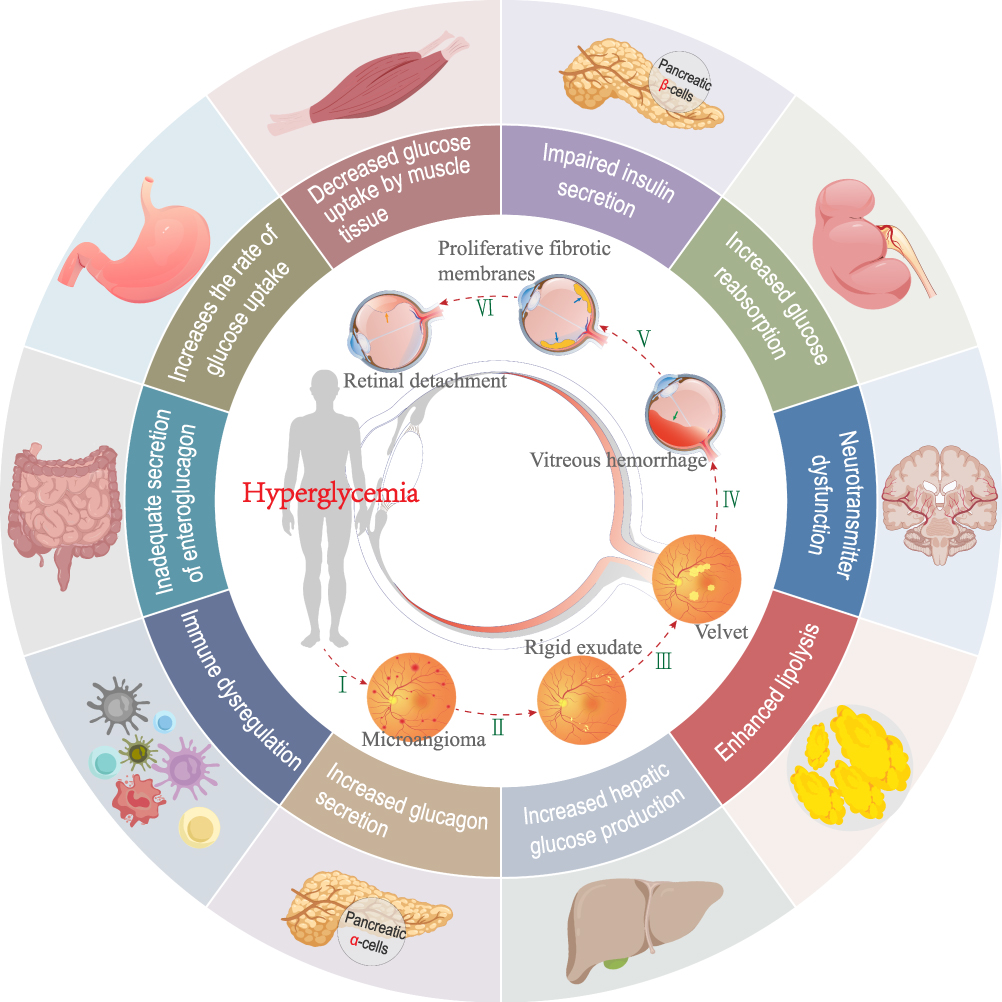 |
Figure 1 Pathogenesis of diabetes mellitus and the stages of progression of diabetic retinopathy. |
Activation of Inflammatory Cells
Infiltration of Macrophages
The main functions of macrophages in the pathological state include phagocytosis, foreign antigen presentation, and several regulatory functions, such as the production of cytokines and growth factors.21 Macrophage polarization is closely related to the pathogenesis of eye diseases22,23 Inflammatory cells, including macrophages and monocytes, accumulate around blood vessels in response to retinal ischemia and hypoxia.24 Macrophages are involved in inflammation, oxidative stress, pathological angiogenesis, and tissue healing in the pathogenesis of DR (Figure 2).21 Macrophages and other circulating immune cells infiltrate the retina during the course of DR, and the infiltrated immune cells and activated microglia are collectively responsible for retinopathy.19
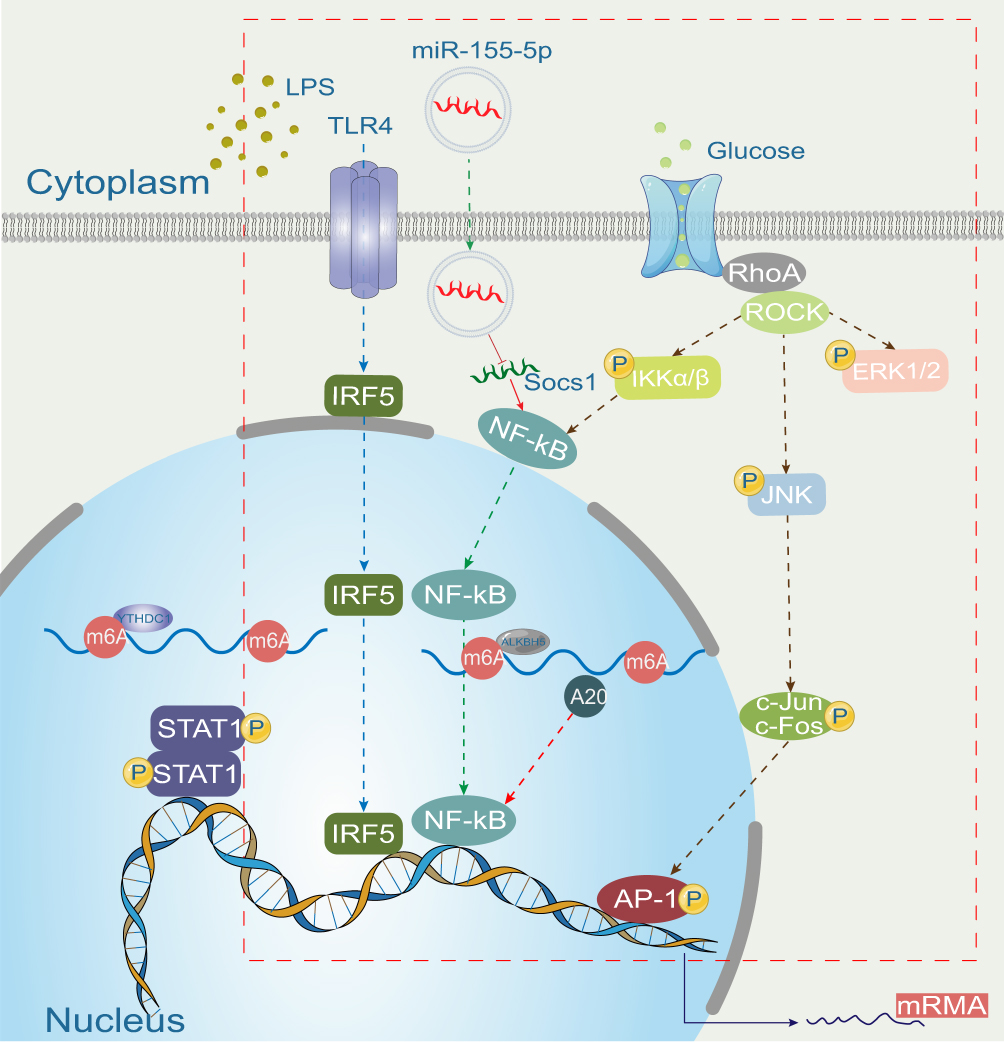 |
Figure 2 Mechanism of macrophage polarization in diabetic retinopathy (marked with a red-dashed box). |
M1 macrophages produce pro-inflammatory cytokines25 and are effector cells in the Th1 immune response. They kill intracellular pathogens, remove foreign substances, and participate in acute pro-inflammatory reactions.26–28 M1 macrophages, while eliminating pathogens, may also exacerbate inflammatory processes, which are harmful to tissues. An increased abundance of M2 macrophages has been reported in the vitreous and fibrovascular membranes of patients with PDR.29 M2 macrophages promote angiogenesis and fibrosis, which may be exacerbated and prolonged by innate immune disorders.19 An increase in blood glucose levels is the hallmark of diabetes. Changes in blood glucose levels affect the polarization of macrophages and the secretion of inflammatory cytokines.21 In healthy individuals, the polarization of M1 and M2 macrophages is balanced in the circulation; however, the lack of anti-inflammatory cells leads to a significant decrease in the polarization of M2 macrophages, resulting in an increase in the M1/M2 polarization ratio in the peripheral blood of patients with type 2 diabetes.30 Recent studies have evaluated glucose tolerance and insulin sensitivity using animal models and in vitro experiments. Macrophage phenotypes and inflammation were analyzed. The results showed that the polarization of the M2 macrophage phenotype could reduce insulin resistance and stabilize glucose/lipid metabolism to a certain extent. This may become one of the most promising treatment options for diabetes. This therapy could also help diabetic patients reduce endothelial dysfunction and prevent diabetic complications.31–33 However, in-depth research on macrophages in DR has not been conducted, and macrophages are multifunctional. Only by implementing different measures based on the specific conditions of the retinal microenvironment in different patients can personalized treatment be applied. Therefore, from the perspective of the therapeutic potential that macrophages bring to the clinical treatment of DR, difficulties remain. This is also a key point requiring further study.
Leukocyte Adhesion and Stasis
Studies have reported that the number of neutrophils in the retina and choroid of patients with diabetes is significantly increased.34 In the pathogenesis of DR, leukocyte adhesion is related to capillary endothelial cell injury and a series of microvascular lesions such as vascular leakage, BRB damage, vascular endothelial cell injury/death, and the formation of capillary nonperfusion areas.35 Notably, the increase in white blood cell stasis parallels increases in diabetes-related metabolic abnormalities.36 The latter results in a loss of components covering the surface of endothelial cells, decreasing capillary density and increasing cell-free capillaries in the later stages of DR.37 Many researchers believe that leukocyte adhesion and leukocyte stasis play a key role in inducing chronic, low-grade inflammation in DR; however, others are of the opinion that the increase in leukocyte stasis is the result of nonspecific endothelial cell dysfunction rather than a key specific step in the development of DR. Therefore, leukocyte stasis may be an epiphenomenon or a secondary effect in the pathogenesis of DR.38 Considerable controversy surrounds the role of white blood cell adhesion and stasis in DR. Animal model experiments should be further applied to verify the specific mechanism of action in the occurrence and development of DR, such as whether the potential molecular mechanism of activating white blood cells and enhancing the interaction between white blood cells and endothelial cells can reduce white blood cell stasis and BRB destruction. If this research can achieve breakthrough progress based on this, it will provide new therapeutic strategies for treating DR.
Activation of Microglia
Microglia are phagocytes in the innate immune system of the central nervous system. The retina and optic nerve, as the crucial components of the eye, play an important role in the immunological pathogenesis of eye diseases.39 Microglia are mainly located in the retinal ganglion cell (RGC) layer, inner plexiform layer, and outer plexiform layer and participate in immune responses as immune cells to maintain retinal homeostasis.40 During conditions such as hyperglycemia, ischemia, and hypoxia, microglia are abnormally activated and further proliferate, migrate, and polarize, therefore playing a dual role in protecting, as well as damaging, retinal tissues (Figure 3).41 The density of microglia in each layer of the retina was reported to increase significantly in rats 4 months after the onset of streptozotocin-induced diabetes; the branches became thicker and shorter, and microglia were activated.42 Another study found that the number of activated microglia in the retina of diabetic rats increased significantly, indicating enhanced proliferation and migration.43
 |
Figure 3 Mechanisms of microglia in the pathogenesis of diabetic retinopathy. Microglia are activated in diabetic retinopathy; microglial cell proliferation is enhanced and their numbers are increased; migration from the inner retina to the outer retina is enhanced and microglia can migrate to the subretinal cavity and retinal pigment epithelial (RPE) layer; activated microglia can penetrate the basement membrane of capillaries, phagocytose endothelial cells, and destroy the BRB (pathway shown with a green dotted line). At the same time, activated microglia release inflammatory factors such as TNF-α, IL-1β, and inducible nitric oxide synthase (iNOS) to damage retinal neurons and vascular endothelial cells, exacerbating BRB damage (pathway shown in blue dashes). Retinal neurons control the activation of microglia via the CD200-CD200R and CX3CL1-CX3CR1 pathways (pathway shown in purple dashes). Neuronal degeneration leads to pathway dysfunction, resulting in the uncontrolled activation of microglia (pathway shown in red dashes). Increased neuronal apoptosis leads to decreased fractalkine (FKN) expression and, therefore, the insufficient inhibition of microglia, leading to the activation of microglia. In a rat model of diabetes, an intravitreal injection of FKN reduced the expression of retinal inflammatory factors, as well as the intracellular production of reactive oxygen species, thereby inhibiting the activation of microglia (pathway shown in Orange dashes). |
Microglia may participate in the occurrence and development of DR in the following ways. (1) Microglia secrete cytotoxic substances, and inflammatory factors mediate the apoptosis and death of RGCs.11 (2) Microglia participate in the pathogenesis of DR by disrupting the structure and function of the BRB. Studies have found that microglia can activate the STAT3 pathway in mice by secreting interleukin (IL)-6 and decrease the expression of tight junction proteins zonula occludens-1 and occludin to increase retinal endothelial permeability.44 Other studies have reported the aggregation of microglia in the retinal pigment epithelial (RPE) layer in different mouse models of DR, suggesting that activated microglia may migrate outside the retina in RPE cells.45 (3) RGCs are lost by microglia through phagocytosis.41,46,47 (4) Microglia can induce retinal neovascularization.40 In a mouse model of oxygen-induced retinopathy, microglia with proliferative and migratory phenotypes were found to aggregate in the ischemic and neovascular areas of the retina.48,49
Müller Cell Activation
Müller cells are an important source of multiple inflammatory mediators and play a key role in initiating inflammatory processes.50 Müller cells are also a type of macroglial cells of the retina, which are unique to the fundus and play an important role in maintaining retinal homeostasis.51 DR is considered to be a neurovascular disease due to the involvement of neurovascular units (neurons, glial cells, vascular endothelial cells, vascular smooth muscle cells, and pericytes) rather than only a microvascular complication of diabetes.52 Degenerative changes in neurons precede changes in microvessels, and neuronal apoptosis is observed within a few weeks of the onset of diabetes.53
Abnormal metabolic pathways resulting from diabetes can lead to cellular dysfunction, causing pathological reactions such as oxidative stress, inflammatory response, and the disruption of the water-liquid balance.54 Müller cells are crucial for maintaining ion and water homeostasis, metabolic coupling, and neurotransmitter recovery (Figure 4A).55–57 They also participate in macular drainage and the maintenance of BRB integrity and are closely related to the pathogenesis of DR.51,58,59 Initial metabolic abnormalities resulting from hyperglycemia can affect retinal cells. Due to their special structure and function, Müller cells can, in turn, aggravate metabolic abnormalities and participate in other pathological mechanisms, such as drainage dysfunction, BRB damage, and inflammation, ultimately contributing to the formation of a complex, interacting network of pathological mechanisms in DR (Figure 4B).
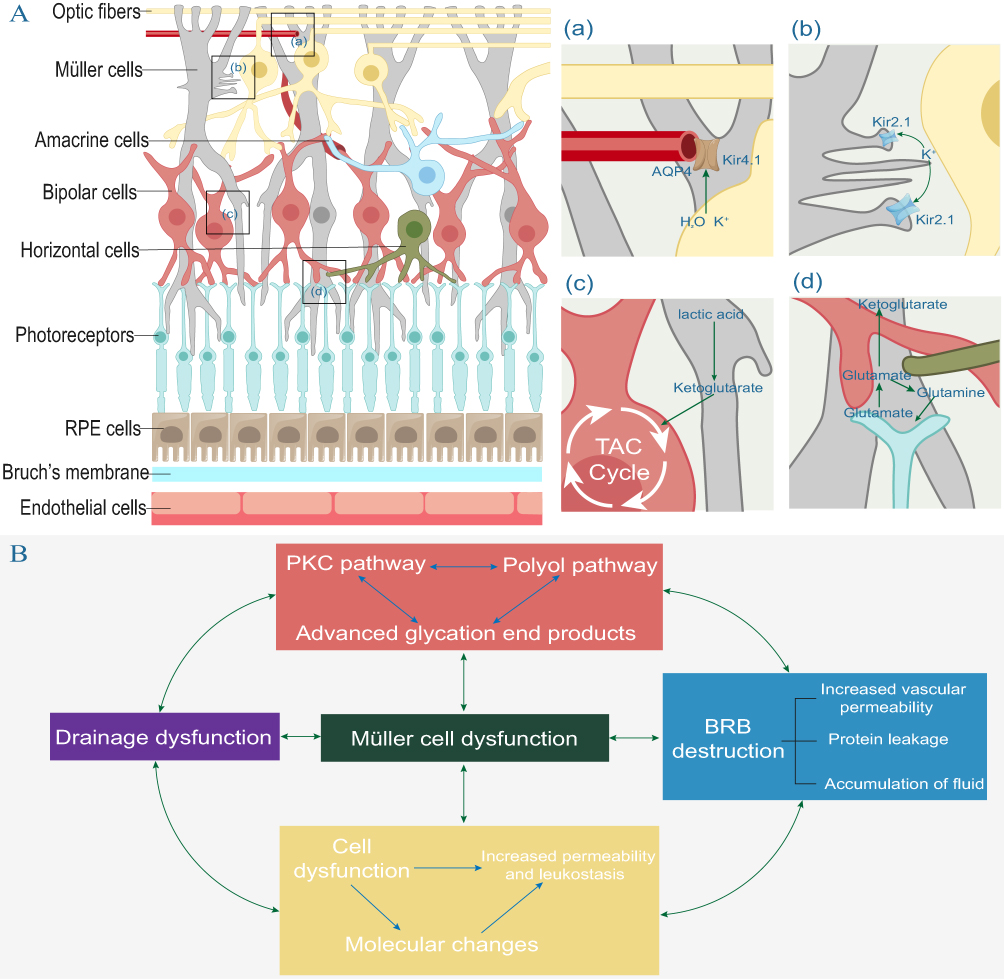 |
Figure 4 (A) Functions of Müller cells. (a)(b) Müller cells can maintain water and ion homeostasis. Water and ions enter cells mainly through Kir2.1 and water channel proteins on the membrane, which form synapses with retinal neurons. Water and ions are transported out of Müller cells through Kir4.1 and aquaporin (AQP)4 on the cell membrane near the vascular plexus and vitreous cavity. (c) Lactate produced during glycolysis can be converted to pyruvate, which then enters neurons as a metabolic substrate for the tricarboxylic acid (TCA) cycle. (d) Retinal neurons release glutamate while transmitting visual signals, and Müller cells take up neurotransmitters including glutamate and direct them into two different metabolic pathways: glutamate is converted to glutamine by glutamate synthetase (GS), which is then released from Müller cells via the sodium-coupled neutral amino acid transporter protein (SNAT), and finally transported back to neurons. (B) Metabolic abnormalities in diabetes, BRB damage, and inflammatory and immune mechanisms are associated with cellular dysfunction and are important pathological mechanisms of DR. |
Activated Müller cells synthesize acute-phase response proteins and several growth factors and cytokines,60 including VEGF, IL-1β, IL-6, tumor necrosis factor (TNF)-α, and monocyte attractant protein (MCP)-1, in a hyperglycemic environment.61 Studies have shown that cluster of differentiation (CD40) (a member of the TNF receptor superfamily) is expressed in several hematopoietic and nonhematopoietic cells, as well as retinal endothelial cells and Müller cells and that the interaction of CD40 with its ligand, CD154, regulates cellular and humoral immunity and enhances inflammation by activating macrophages/microglia.62 As unique glial cells in the fundus, Müller cells play a unique role in inflammation of the fundus. Therefore, further studies on the role of Müller cells in DR can promote our understanding of DR and help develop suitable therapies. Furthermore, with recent advances in gene therapy, the targeting of Müller cells may provide a new direction for treating DR and lay a more solid foundation for its applications in treating eye diseases.
Release of Inflammatory Factors by Astrocytes
Astrocytes are in contact with retinal blood vessels and neurons and play a key role in maintaining the integrity of the BRB.63 Retinal astrocytes are closely related to blood vessels and are involved in BRB damage in DR. In early diabetes, there is evidence that astrocytes are activated, and several pro-inflammatory cytokines, including IL-6, IL1β, IL-8, cyclooxygenase-2, tumor growth factor-β, epidermal growth factor, macrophage inflammatory protein 2α, and VEGF, are produced.64 Reactive astrocytes can also secrete chemokines that recruit microglia, monocytes/macrophages, and T cells, thereby amplifying the inflammatory response.65
Retinal Pigment Epithelium Is Involved in the Inflammatory Response of DR
RPE is a single layer of cells that undergoes polarization after mitosis. It plays a crucial role in maintaining the integrity of the BRB and the physiological functions of the retina.66 Previous studies found that RPE is involved in the pathogenesis of DR.45 Experimental evidence showed that the RPE structure of diabetic animals or cells became disordered, and gene expression changed under high glucose conditions.67,68 Studies have shown that high sugar conditions may cause structural and functional damage to RPE cells, thereby contributing to the pathogenesis of DR.69 The absence of ZO-1 and occludin expression in RPE cells may lead to the disruption of the RPE barrier, resulting in vascular leakage, retinal edema, and exudative retinal detachment, as described in DR.70 The changes in the growth factors and inflammatory cytokines secreted by RPE also play a role in inflammation and angiogenesis during DR.71
The Role of Systemic Inflammatory Markers and Inflammatory Factors in the Occurrence and Development of DR
At present, the new blood inflammatory markers that are easily accessible in clinical practice, including the systemic immune inflammation index (SII), neutrophil-lymphocyte ratio (NLR), and platelet-lymphocyte ratio (PLR), are closely related to various inflammation-related diseases such as cardiovascular diseases72 and cancer.73 Recent research reports showed that an increase in SII values was closely related to the existence of diabetic macular edema (DME).74 The NLR and SII levels were significantly increased in DME patients with subretinal fluid (SRF).75 These studies revealed an association between systemic inflammatory markers, ophthalmic treatment, and DME. As promising systemic inflammatory markers, SII, the NLR, and the PLR have been studied in a variety of chronic diseases. SII is regarded to be a predictive and prognostic marker in various cancers,76–78 autoimmune diseases,79 cardiovascular diseases,80,81 and metabolic diseases.82 The NLR is now widely applied in all medical disciplines and is regarded to reflect the body’s response to various external or internal stimuli. Similarly, the PLR has also demonstrated potential value in multiple diseases.83 The systemic neutrophil count is elevated in patients with DM and DR, suggesting that neutrophil-mediated inflammation may play a significant role in the pathogenesis of DM and DR.84 Currently, platelets are increasingly recognized as being associated with inflammation. Platelet adhesion, aggregation, and secretion are all involved in the inflammatory response.85 These indicators are correlated with the imaging features of DME, further supporting the inflammatory origin of DME and revealing the relationship between different imaging features of DME. Thus, the inflammatory response underlying the pathogenesis of DME may be mediated by multiple blood cells.86
In addition to the activation of inflammatory cells, the upregulation of inflammatory factors is involved in the occurrence and development of DR (Table 1). Numerous studies have shown that patients with diabetes have elevated levels of inflammatory factors in the eye fluid, including hypoxia-inducing factor (HIF)-1α, VEGF, placental growth factor (PlGF), insulin-like growth factor 1, basic fibroblast growth factor, pigment epithelium-derived factor, IL-1β, IL-2, IL-4, IL-6, IL-8, IL-10, MCP-1/2, intercellular adhesion molecule (ICAM)-1, VCAM-1, TNF-α, interferon-γ, interferon-γ–induced protein 10, complement components 3a and 5a, and CD40.87,88 In patients with DR, the levels of inflammatory cytokines, including VEGF, ICAM-1, IL-1β, IL-6, TNF-α, IL-8, and MCP-1, were significantly increased in the vitreous humor, suggesting the involvement of inflammatory factors in the pathogenesis of DR.89
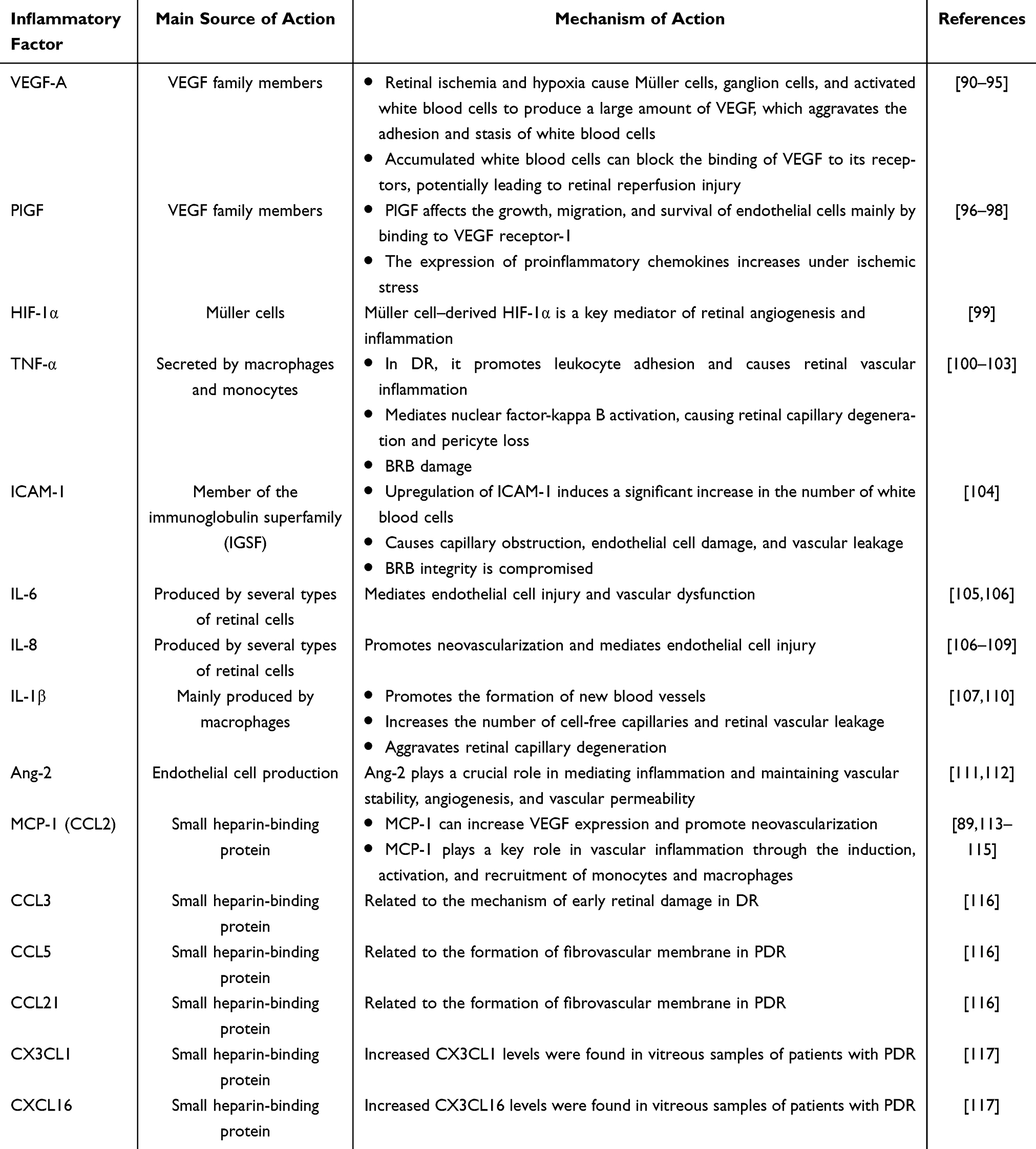 |
Table 1 Mechanisms of the Various Inflammatory Factors in Diabetic Retinopathy |
Role of the Complement System in DR
An unknown heat-resistant cracking substance found in the blood at the end of the 19th century was found to be active against various bacteria and later named “complement”.118 Studies have shown that retinal vascular damage in DR is closely related to the activation of the complement system.19 Other studies found that Bruch’s membrane and the choroidal capillaries of patients with DR contain high levels of C3d and C5b-9 complexes, as well as deposits of the membrane-attack complex.119 High levels of complement C3 are associated with an increased risk of DR.120 Polymorphism of the complement C5 gene is associated with PDR.121
Activation of the complement system may promote the development of DR via the following pathways. (1) Classical pathway: Only a few studies have attempted to elucidate the classical pathway proteins in the retina of patients with DR; thus, their role in the pathogenesis of DR needs to be further determined. Some studies have reported that C4b, C3, C9, and factor B-related complement proteins in the vitreous body of patients with PDR were significantly higher than those in nondiabetic individuals. Western blotting of C3 and factor B revealed results that were consistent with the quantitative and timed determination of mRNA expression using RT-PCR. These findings collectively indicate that the incidence of DR is closely related to complement activation through classical pathways.122 The absence of immunoglobulin G (IgG) in the exosomes of diabetic mice can decrease the extent of retinal blood vessel damage. Further C1-activation experiments revealed that exosomes containing IgG in plasma might activate proteolysis of the C1 complex by binding to it, thereby activating the classical complement pathway and leading to downstream retinal blood vessel damage.123 (2) Alternative pathways: The complement-replacement pathway is overactivated in the early stages in patients with DR, suggesting that complement-mediated inflammation may accelerate DR.124 Studies involving humans and mice demonstrated reduced levels of the two membrane-binding inhibitors of the complement system, namely CD55 and CD59, suggesting the activation of the complement-replacement pathway in DR.125 In addition, the levels of 16 complement components in the vitreous body of patients with PDR were significantly higher than in nondiabetic individuals, and the C3a/C3, C5a/C5, and Ba/factor B ratios were higher in patients with diabetes than in the control group. The local and intraocular activation of C3, C5, and factor B indicate activation of the complement-replacement pathway in patients with PDR.126 (3) Lectin pathway: A cross-sectional study found that serum mannose-binding lectin (MBL) levels were higher in patients with DR than in healthy subjects. Furthermore, MBL levels have been reported to increase with increases in the severity of DR, suggesting the involvement of the complement lectin activation pathway in DR.127 Other studies have shown that high levels of serum lectin pathway proteins MAp44 and MASP-2 were correlated with DR, whereas high levels of H-ficolin were correlated with the occurrence of simple retinopathy, suggesting that lectin pathway complement activation may play a prominent role in the early stages of DR.128 BRB damage in DR leads to the infiltration of serum proteins such as complements, cytokines, and chemokines into retinal tissues, aggravating tissue damage. The deactivation of complement is associated with DR, and systemic complement activation is positively correlated with DR progression.127
Concluding Remarks and Future Perspectives
Immune system disorders and inflammation are important factors for the occurrence and development of DR. Among them, the BRB is the first line of immune defense of the retina. Microglia, as immune cells, participate in immune responses, maintain retinal homeostasis, and exert dual protective and damaging effects on retinal tissues. Müller cells play an important role in maintaining the in vivo balance of ions and water, metabolic coupling, neurotransmitter resuscitation, maintain macular drainage, and BRB integrity. The degree of injury to the complement system is positively correlated with DR progression, and the interaction of multiple cytokines affects immune regulation and other processes. The main approaches to treating DR currently include retinal laser photocoagulation and intravitreal injections of anti-VEGF drugs, corticosteroids, and vitrectomy; however, these approaches have limitations, which may be related to individual differences in immune responses and anti-VEGF drug treatment that does not target inflammation. Therefore, recognizing the nature of DR is the fundamental starting point in treating DR. Elucidating the immune mechanisms has become a hotspot in DR research; however, their specific mechanisms in DR need to be further understood to provide a basis and strategy for treating DR in clinical settings.
DR is a moderate, low-grade, chronic inflammatory disease characterized by the activation of inflammatory cells and the production and expression of inflammatory factors. The entire retina is affected by inflammatory factors, leading to BRB damage, retinal neuronal death, and aggravation of the occurrence and development of DR. The current DME treatments mainly include intraocular injections of anti-VEGF drugs. As some patients are insensitive or unresponsive to anti-VEGF treatment, it is likely that other factors, especially inflammatory factors, play a role in affecting the onset of DR. In view of the different pathogeneses of DR, individualized anti-inflammatory treatment is needed for patients to derive treatment benefits. In clinical applications, relevant intraocular inflammatory factors can be analyzed in DR patients, such as the detection of intraocular fluid and systemic inflammatory markers (eg, SII, NLR, and PLR), multimodal imaging, such as blood flow optical coherence tomography (OCT) for the detection of high reflectance points, and SND to distinguish and determine the role of inflammation in DR and DME. Only in this way is it possible to achieve truly individualized anti-inflammatory treatment.
From inevitable vision loss to the continuous pursuit of long-term vision improvement, large-scale clinical trials have laid the foundation for today’s DR treatment, with laser, vitrectomy, and anti-VEGF drugs being the main approaches. Although the prognosis has improved significantly, problems such as a lack of response to conventional treatment, recurrence, and long-term vision loss remain. Thus, exploring new disease management methods and discovering new therapeutic targets is urgently needed. As mentioned, Müller cells are a unique type of retinal glial cells, and their role in the pathogenesis of DR has received extensive attention. The rapid development of gene therapy, especially the emergence of gene editing technology, has driven the development of treatment methods targeting Müller cells, presenting new treatment strategies for DR. Whether it is anti-VEGF drug treatment, individualized anti-inflammatory treatment, or gene-targeted therapy, some patients may develop drug resistance. Therefore, individualized, combined, and intelligent strategies need to be adopted, including changing or upgrading drugs, precise combined treatment, optimizing drug administration methods, developing new therapies, and using biomarkers to further guide decision-making. The current challenge lies in the fact that the roles and treatments of many inflammatory and immune mechanisms in DR need further research. Future findings can provide a deeper understanding of the biological functions and related targeted therapies and promote the transfer of the latest research results to clinical applications.
List of Abbreviations
AQP, aquaporin; BRB, blood–retinal barrier; CD, cluster of differentiation; DME, diabetic macular edema; DR, diabetic retinopathy; FKN, fractalkine; GS, glutamate synthetase; HIF, hypoxia-inducing factor; ICAM, intercellular adhesion molecule; IgG, immunoglobulin G; IL, interleukin; IGSF, immunoglobulin superfamily; iNOS, inducible nitric oxide synthase; MBL, mannose-binding lectin; SRF, subretinal fluid; MCP, monocyte attractant protein; PDR, proliferative DR; PlGF, placental growth factor; RGC, retinal ganglion cell; SII, systemic immune inflammation index; NLR, neutrophil-lymphocyte ratio; PLR, platelet-lymphocyte ratio; RPE, retinal pigment epithelial; SNAT, sodium-coupled neutral amino acid transporter protein; TCA, tricarboxylic acid; TNF, tumor necrosis factor; VEGF, vascular endothelial growth factor.
Data Sharing Statement
All figures and tables are original and are not taken from other publications. Data sharing is not applicable to this article, as no new data were created or analysed in this study.
Ethics Approval and Consent to Participate
Ethics approval and consent to participate is not applicable for this review article.
Consent for Publication
Consent for publication from participants is not applicable for this review study.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
No relevant funding was received for this study.
Disclosure
The authors declare no competing interests.
References
1. Bohler F, Bohler L, Taranikanti V. Targeting pericyte retention in diabetic retinopathy: a review. Ann Med. 2024;56(1):2398200. doi:10.1080/07853890.2024.2398200
2. Shaw JE, Sicree RA, Zimmet PZ. Global estimates of the prevalence of diabetes for 2010 and 2030. Diabetes Res Clin Pract. 2010;87(1):4–14. doi:10.1016/j.diabres.2009.10.007
3. Mohamed Q, Gillies MC, Wong TY. Management of diabetic retinopathy: a systematic review. JAMA. 2007;298(8):902–916. doi:10.1001/jama.298.8.902
4. Cheung N, Mitchell P, Wong TY. Diabetic retinopathy. Lancet. 2010;376(9735):124–136. doi:10.1016/S0140-6736(09)62124-3
5. Sabanayagam C, Banu R, Chee ML, et al. Incidence and progression of diabetic retinopathy: a systematic review. Lancet Diabetes Endocrinol. 2019;7(2):140–149. doi:10.1016/S2213-8587(18)30128-1
6. Cole JB, Florez JC. Genetics of diabetes mellitus and diabetes complications. Nat Rev Nephrol. 2020;16(7):377–390. doi:10.1038/s41581-020-0278-5
7. Jiang F, Lei C, Chen Y, et al. The complement system and diabetic retinopathy. Surv Ophthalmol. 2024;69(4):575–584. doi:10.1016/j.survophthal.2024.02.004
8. Kour V, Swain J, Singh J, et al. A review on diabetic retinopathy. Curr Diabetes Rev. 2024;20(6):e201023222418. doi:10.2174/0115733998253672231011161400
9. Lechner J, O’Leary OE, Stitt AW. The pathology associated with diabetic retinopathy. Vision Res. 2017;139:7–14. doi:10.1016/j.visres.2017.04.003
10. Li H, Liu X, Zhong H, et al. Research progress on the pathogenesis of diabetic retinopathy. BMC Ophthalmol. 2023;23(1):372. doi:10.1186/s12886-023-03118-6
11. Altmann C, Schmidt MHH. The role of microglia in diabetic retinopathy: inflammation, microvasculature defects and neurodegeneration. Int J Mol Sci. 2018;19(1):110. doi:10.3390/ijms19010110
12. Tonade D, Liu H, Palczewski K, et al. Photoreceptor cells produce inflammatory products that contribute to retinal vascular permeability in a mouse model of diabetes. Diabetologia. 2017;60(10):2111–2120. doi:10.1007/s00125-017-4381-5
13. Whitehead M, Wickremasinghe S, Osborne A, et al. Diabetic retinopathy: a complex pathophysiology requiring novel therapeutic strategies. Expert Opin Biol Ther. 2018;18(12):1257–1270. doi:10.1080/14712598.2018.1545836
14. Simo-Servat O, Hernandez C, Simo R. Diabetic retinopathy in the context of patients with diabetes. Ophthalmic Res. 2019;62(4):211–217. doi:10.1159/000499541
15. Pearce I, Simo R, Lovestam-Adrian M, et al. Association between diabetic eye disease and other complications of diabetes: implications for care. A systematic review. Diabetes Obes Metab. 2019;21(3):467–478. doi:10.1111/dom.13550
16. Trento M, Durando O, Lavecchia S, et al. Vision related quality of life in patients with type 2 diabetes in the EUROCONDOR trial. Endocrine. 2017;57(1):83–88. doi:10.1007/s12020-016-1097-0
17. Tawfik A, Samra YA, Elsherbiny NM, et al. Implication of hyperhomocysteinemia in blood retinal barrier (BRB) dysfunction. Biomolecules. 2020;10(8):1119. doi:10.3390/biom10081119
18. Chen M, Luo C, Zhao J, et al. Immune regulation in the aging retina. Prog Retin Eye Res. 2019;69:159–172. doi:10.1016/j.preteyeres.2018.10.003
19. Xu H, Chen M. Diabetic retinopathy and dysregulated innate immunity. Vision Res. 2017;139:39–46. doi:10.1016/j.visres.2017.04.013
20. Kastelan S, Oreskovic I, Biscan F, et al. Inflammatory and angiogenic biomarkers in diabetic retinopathy. Biochem Med (Zagreb). 2020;30(3):030502. doi:10.11613/BM.2020.030502
21. Yao Y, Li J, Zhou Y, et al. Macrophage/microglia polarization for the treatment of diabetic retinopathy. Front Endocrinol (Lausanne). 2023;14:1276225. doi:10.3389/fendo.2023.1276225
22. Yu J, Chai P, Xie M, et al. Histone lactylation drives oncogenesis by facilitating m(6)A reader protein YTHDF2 expression in ocular melanoma. Genome Biol. 2021;22(1):85. doi:10.1186/s13059-021-02308-z
23. Qu R, Zhou M, Qiu Y, et al. Glucocorticoids improve the balance of M1/M2 macrophage polarization in experimental autoimmune uveitis through the P38MAPK-MEF2C axis. Int Immunopharmacol. 2023;120:110392. doi:10.1016/j.intimp.2023.110392
24. Zeng Y, Wen F, Mi L, et al. Changes in macrophage-like cells characterized by en face optical coherence tomography after retinal stroke. Front Immunol. 2022;13:987836. doi:10.3389/fimmu.2022.987836
25. Costantini A, Viola N, Berretta A, et al. Age-related M1/M2 phenotype changes in circulating monocytes from healthy/unhealthy individuals. Aging (Albany NY). 2018;10(6):1268–1280. doi:10.18632/aging.101465
26. Shapouri-Moghaddam A, Mohammadian S, Vazini H, et al. Macrophage plasticity, polarization, and function in health and disease. J Cell Physiol. 2018;233(9):6425–6440. doi:10.1002/jcp.26429
27. Geng P, Zhu H, Zhou W, et al. Baicalin inhibits influenza a virus infection via promotion of M1 macrophage polarization. Front Pharmacol. 2020;11:01298. doi:10.3389/fphar.2020.01298
28. Wang X, Li H, Chen S, et al. P300/CBP-associated factor (PCAF) attenuated M1 macrophage inflammatory responses possibly through KLF2 and KLF4. Immunol Cell Biol. 2021;99(7):724–736. doi:10.1111/imcb.12455
29. Kobayashi Y, Yoshida S, Nakama T, et al. Overexpression of CD163 in vitreous and fibrovascular membranes of patients with proliferative diabetic retinopathy: possible involvement of periostin. Br J Ophthalmol. 2015;99(4):451–456. doi:10.1136/bjophthalmol-2014-305321
30. Fadini GP, de Kreutzenberg SV, Boscaro E, et al. An unbalanced monocyte polarisation in peripheral blood and bone marrow of patients with type 2 diabetes has an impact on microangiopathy. Diabetologia. 2013;56(8):1856–1866. doi:10.1007/s00125-013-2918-9
31. Sun J, Huang Q, Li S, et al. miR-330-5p/Tim-3 axis regulates macrophage M2 polarization and insulin resistance in diabetes mice. Mol Immunol. 2018;95:107–113. doi:10.1016/j.molimm.2018.02.006
32. Gao J, Cheng Y, Hao H, et al. Decitabine assists umbilical cord-derived mesenchymal stem cells in improving glucose homeostasis by modulating macrophage polarization in type 2 diabetic mice. Stem Cell Res Ther. 2019;10(1):259. doi:10.1167/iovs.12-9446
33. Chen X, Zhuo S, Zhu T, et al. Fpr2 deficiency alleviates diet-induced insulin resistance through reducing body weight gain and inhibiting inflammation mediated by macrophage chemotaxis and M1 polarization. Diabetes. 2019;68(6):1130–1142. doi:10.2337/db18-0469
34. Lutty GA, Cao J, McLeod DS. Relationship of polymorphonuclear leukocytes to capillary dropout in the human diabetic choroid. Am J Pathol. 1997;151(3):707–714.
35. Joussen AM, Murata T, Tsujikawa A, et al. Leukocyte-mediated endothelial cell injury and death in the diabetic retina. Am J Pathol. 2001;158(1):147–152. doi:10.1080/14712598.2018.1545836
36. Roy S, Kern TS, Song B, et al. Mechanistic insights into pathological changes in the diabetic retina: implications for targeting diabetic retinopathy. Am J Pathol. 2017;187(1):9–19. doi:10.1016/j.ajpath.2016.08.022
37. Forrester JV, Kuffova L, Delibegovic M. The role of inflammation in diabetic retinopathy. Front Immunol. 2020;11:583687. doi:10.3389/fimmu.2020.583687
38. van der Wijk AE, Hughes JM, Klaassen I, et al. Is leukostasis a crucial step or epiphenomenon in the pathogenesis of diabetic retinopathy? J Leukoc Biol. 2017;102(4):993–1001. doi:10.1189/jlb.3RU0417-139
39. Madore C, Yin Z, Leibowitz J, et al. Microglia, lifestyle stress, and neurodegeneration. Immunity. 2020;52(2):222–240. doi:10.1016/j.immuni.2019.12.003
40. Kinuthia UM, Wolf A, Langmann T. Microglia and inflammatory responses in diabetic retinopathy. Front Immunol. 2020;11:564077. doi:10.3389/fimmu.2020.564077
41. Karlstetter M, Scholz R, Rutar M, et al. Retinal microglia: just bystander or target for therapy? Prog Retin Eye Res. 2015;45:30–57. doi:10.1016/j.preteyeres.2014.11.004
42. Zeng XX, Ng YK, Ling EA. Neuronal and microglial response in the retina of streptozotocin-induced diabetic rats. Vis Neurosci. 2000;17(3):463–471. doi:10.1017/s0952523800173122
43. Chen X, Zhou H, Gong Y, et al. Early spatiotemporal characterization of microglial activation in the retinas of rats with streptozotocin-induced diabetes. Graefes Arch Clin Exp Ophthalmol. 2015;253(4):519–525. doi:10.1007/s00417-014-2727-y
44. Yun JH, Park SW, Kim KJ, et al. Endothelial STAT3 activation increases vascular leakage through downregulating tight junction proteins: implications for diabetic retinopathy. J Cell Physiol. 2017;232(5):1123–1134. doi:10.1002/jcp.25575
45. Jo DH, Yun JH, Cho CS, et al. Interaction between microglia and retinal pigment epithelial cells determines the integrity of outer blood-retinal barrier in diabetic retinopathy. Glia. 2019;67(2):321–331. doi:10.1002/glia.23542
46. Brown GC, Neher JJ. Microglial phagocytosis of live neurons. Nat Rev Neurosci. 2014;15(4):209–216. doi:10.1038/nrn3710
47. Anderson SR, Zhang J, Steele MR, et al. Complement targets newborn retinal ganglion cells for phagocytic elimination by microglia. J Neurosci. 2019;39(11):2025–2040. doi:10.1523/JNEUROSCI.1854-18.2018
48. Boeck M, Thien A, Wolf J, et al. Temporospatial distribution and transcriptional profile of retinal microglia in the oxygen-induced retinopathy mouse model. Glia. 2020;68(9):1859–1873. doi:10.1002/glia.23810
49. Xu W, Hu Z, Lv Y, et al. Microglial density determines the appearance of pathological neovascular tufts in oxygen-induced retinopathy. Cell Tissue Res. 2018;374(1):25–38. doi:10.1007/s00441-018-2847-5
50. Rubsam A, Parikh S, Fort PE. Role of inflammation in diabetic retinopathy. Int J Mol Sci. 2018;19(4):942. doi:10.3390/ijms19040942
51. Lai D, Wu Y, Shao C, et al. The role of muller cells in diabetic macular edema. Invest Ophthalmol Vis Sci. 2023;64(10):8. doi:10.1167/iovs.64.10.8
52. Gardner TW, Davila JR. The neurovascular unit and the pathophysiologic basis of diabetic retinopathy. Graefes Arch Clin Exp Ophthalmol. 2017;255(1):1–6. doi:10.1007/s00417-016-3548-y
53. Barber AJ, Lieth E, Khin SA, et al. Neural apoptosis in the retina during experimental and human diabetes. Early onset and effect of insulin. J Clin Invest. 1998;102(4):783–791. doi:10.1172/JCI2425
54. Curtis TM, Hamilton R, Yong PH, et al. Muller glial dysfunction during diabetic retinopathy in rats is linked to accumulation of advanced glycation end-products and advanced lipoxidation end-products. Diabetologia. 2011;54(3):690–698. doi:10.1007/s00125-010-1971-x
55. Gao F, Xu LJ, Zhao Y, et al. K+ channels of muller glial cells in retinal disorders. CNS Neurol Disord Drug Targets. 2018;17(4):255–260. doi:10.2174/1871527317666180202114233
56. Toft-Kehler AK, Skytt DM, Kolko M. A perspective on the muller cell-neuron metabolic partnership in the inner retina. Mol Neurobiol. 2018;55(6):5353–5361. doi:10.1007/s12035-017-0760-7
57. Jo AO, Ryskamp DA, Phuong TT, et al. TRPV4 and AQP4 channels synergistically regulate cell volume and calcium homeostasis in retinal muller glia. J Neurosci. 2015;35(39):13525–13537. doi:10.1523/JNEUROSCI.1987-15.2015
58. Ji N, Guo Y, Liu S, et al. MEK/ERK/RUNX2 pathway-mediated IL-11 autocrine promotes the activation of muller glial cells during diabetic retinopathy. Curr Eye Res. 2022;47(12):1622–1630. doi:10.1080/02713683.2022.2129070
59. Carpi-Santos R, de Melo Reis RA, Gomes FCA, et al. Contribution of müller cells in the diabetic retinopathy development: focus on oxidative stress and inflammation. Antioxidants (Basel). 2022;11(4). doi:10.3390/antiox11040617
60. Zong H, Ward M, Madden A, et al. Hyperglycaemia-induced pro-inflammatory responses by retinal Müller glia are regulated by the receptor for advanced glycation end-products (RAGE). Diabetologia. 2010;53(12):2656–2666. doi:10.1007/s00125-010-1900-z
61. Mu H, Zhang XM, Liu JJ, et al. Effect of high glucose concentration on VEGF and PEDF expression in cultured retinal muller cells. Mol Biol Rep. 2009;36(8):2147–2151. doi:10.1007/s11033-008-9428-8
62. Portillo JC, Lopez Corcino Y, Miao Y, et al. CD40 in retinal muller cells induces P2X7-dependent cytokine expression in macrophages/microglia in diabetic mice and development of early experimental diabetic retinopathy. Diabetes. 2017;66(2):483–493. doi:10.2337/db16-0051
63. Ridet JL, Malhotra SK, Privat A, et al. Reactive astrocytes: cellular and molecular cues to biological function. Trends Neurosci. 1997;20(12):570–577. doi:10.1016/s0166-2236(97)01139-9
64. Pekny M, Wilhelmsson U, Pekna M. The dual role of astrocyte activation and reactive gliosis. Neurosci Lett. 2014;565:30–38. doi:10.1016/j.neulet.2013.12.071
65. Rothhammer V, Quintana FJ. Control of autoimmune CNS inflammation by astrocytes. Semin Immunopathol. 2015;37(6):625–638. doi:10.1007/s00281-015-0515-3
66. Lakkaraju A, Umapathy A, Tan LX, et al. The cell biology of the retinal pigment epithelium. Prog Retin Eye Res. 2020:100846. doi:10.1016/j.preteyeres.2020.100846
67. Tarchick MJ, Bassiri P, Rohwer RM, et al. Early functional and morphologic abnormalities in the diabetic Nyx nob mouse retina. Invest Ophthalmol Vis Sci. 2016;57(7):3496–3508. doi:10.1167/iovs.15-18775
68. Zhang Y, Xi X, Mei Y, et al. High-glucose induces retinal pigment epithelium mitochondrial pathways of apoptosis and inhibits mitophagy by regulating ROS/PINK1/Parkin signal pathway. Biomed Pharmacother. 2019;111:1315–1325. doi:10.1016/j.biopha.2019.01.034
69. Chen Q, Tang L, Xin G, et al. Oxidative stress mediated by lipid metabolism contributes to high glucose-induced senescence in retinal pigment epithelium. Free Radic Biol Med. 2019;130:48–58. doi:10.1016/j.freeradbiomed.2018.10.419
70. Tonade D, Kern TS. Photoreceptor cells and RPE contribute to the development of diabetic retinopathy. Prog Retin Eye Res. 2021;83:100919. doi:10.1016/j.preteyeres.2020.100919
71. Ponnalagu M, Subramani M, Jayadev C, et al. Retinal pigment epithelium-secretome: a diabetic retinopathy perspective. Cytokine. 2017;95:126–135. doi:10.1016/j.cyto.2017.02.013
72. Bhat T, Teli S, Rijal J, et al. Neutrophil to lymphocyte ratio and cardiovascular diseases: a review. Expert Rev Cardiovasc Ther. 2013;11(1):55–59. doi:10.1586/erc.12.159
73. Hirahara T, Arigami T, Yanagita S, et al. Combined neutrophil-lymphocyte ratio and platelet-lymphocyte ratio predicts chemotherapy response and prognosis in patients with advanced gastric cancer. BMC Cancer. 2019;19(1):672. doi:10.1186/s12885-019-5903-y
74. Elbeyli A, Kurtul BE, Ozcan SC, et al. The diagnostic value of systemic immune-inflammation index in diabetic macular oedema. Clin Exp Optom. 2022;105(8):831–835. doi:10.1080/08164622.2021.1994337
75. Ozata Gundogdu K, Dogan E, Celik E, et al. Serum inflammatory marker levels in serous macular detachment secondary to diabetic macular edema. Eur J Ophthalmol. 2022;32(6):3637–3643. doi:10.1177/11206721221083465
76. Hu B, Yang XR, Xu Y, et al. Systemic immune-inflammation index predicts prognosis of patients after curative resection for hepatocellular carcinoma. Clin Cancer Res. 2014;20(23):6212–6222. doi:10.1158/1078-0432.CCR-14-0442
77. Xie QK, Chen P, Hu WM, et al. The systemic immune-inflammation index is an independent predictor of survival for metastatic colorectal cancer and its association with the lymphocytic response to the tumor. J Transl Med. 2018;16(1):273. doi:10.1186/s12967-018-1638-9
78. Li QX, Shi DJ, Zhang LX, et al. Association of body mass and systemic immune-inflammation indices with endocrine therapy resistance in luminal breast cancers. J Int Med Res. 2019;47(5):1936–1947. doi:10.1177/0300060519831570
79. Dincer Rota D, Tanacan E. The utility of systemic-immune inflammation index for predicting the disease activation in patients with psoriasis. Int J Clin Pract. 2021;75(6):e14101. doi:10.1111/ijcp.14101
80. Bagci A, Aksoy F. Systemic immune-inflammation index predicts new-onset atrial fibrillation after ST elevation myocardial infarction. Biomark Med. 2021;15(10):731–739. doi:10.2217/bmm-2020-0838
81. Erdogan M, Erdol MA, Ozturk S, et al. Systemic immune-inflammation index is a novel marker to predict functionally significant coronary artery stenosis. Biomark Med. 2020;14(16):1553–1561. doi:10.2217/bmm-2020-0274
82. Wang J, Zhou D, Dai Z, et al. Association between systemic immune-inflammation index and diabetic depression. Clin Interv Aging. 2021;16:97–105. doi:10.2147/CIA.S285000
83. Wang G, Mivefroshan A, Yaghoobpoor S, et al. Prognostic value of platelet to lymphocyte ratio in sepsis: a systematic review and meta-analysis. Biomed Res Int. 2022;2022(1):9056363. doi:10.1155/2022/9056363
84. Woo SJ, Ahn SJ, Ahn J, et al. Elevated systemic neutrophil count in diabetic retinopathy and diabetes: a hospital-based cross-sectional study of 30,793 Korean subjects. Invest Ophthalmol Vis Sci. 2011;52(10):7697–7703. doi:10.1167/iovs.11-7784
85. Schwertz H, Rowley JW, Portier I, et al. Human platelets display dysregulated sepsis-associated autophagy, induced by altered LC3 protein-protein interaction of the vici-protein EPG5. Autophagy. 2022;18(7):1534–1550. doi:10.1080/15548627.2021.1990669
86. Zhou J, Song S, Zhang Y, et al. OCT-based biomarkers are associated with systemic inflammation in patients with treatment-naive diabetic macular edema. Ophthalmol Ther. 2022;11(6):2153–2167. doi:10.1007/s40123-022-00576-x
87. Vujosevic S, Lupidi M, Donati S, et al. Role of inflammation in diabetic macular edema and neovascular age-related macular degeneration. Surv Ophthalmol. 2024;69(6):870–881. doi:10.1016/j.survophthal.2024.07.006
88. Noma H, Mimura T, Yasuda K, et al. Role of inflammation in diabetic macular edema. Ophthalmologica. 2014;232(3):127–135. doi:10.1159/000364955
89. Taghavi Y, Hassanshahi G, Kounis NG, et al. Monocyte chemoattractant protein-1 (MCP-1/CCL2) in diabetic retinopathy: latest evidence and clinical considerations. J Cell Commun Signal. 2019;13(4):451–462. doi:10.1007/s12079-018-00500-8
90. Peach CJ, Mignone VW, Arruda MA, et al. Molecular pharmacology of VEGF-A Isoforms: binding and signalling at VEGFR2. Int J Mol Sci. 2018;19(4):1264. doi:10.3390/ijms19041264
91. Shibuya M, Claesson-Welsh L. Signal transduction by VEGF receptors in regulation of angiogenesis and lymphangiogenesis. Exp Cell Res. 2006;312(5):549–560. doi:10.1016/j.yexcr.2005.11.012
92. Bai Y, Ma JX, Guo J, et al. Muller cell-derived VEGF is a significant contributor to retinal neovascularization. J Pathol. 2009;219(4):446–454. doi:10.1002/path.2611
93. Liu Y, Shen J, Fortmann SD, et al. Reversible retinal vessel closure from VEGF-induced leukocyte plugging. JCI Insight. 2017;2(18). doi:10.1172/jci.insight.95530
94. Bressler NM, Beaulieu WT, Glassman AR, et al. Persistent macular thickening following intravitreous aflibercept, bevacizumab, or ranibizumab for central-involved diabetic macular edema with vision impairment: a secondary analysis of a randomized clinical trial. JAMA Ophthalmol. 2018;136(3):257–269. doi:10.1001/jamaophthalmol.2017.6565
95. Bromberg-White JL, Glazer L, Downer R, et al. Identification of VEGF-independent cytokines in proliferative diabetic retinopathy vitreous. Invest Ophthalmol Vis Sci. 2013;54(10):6472–6480. doi:10.1167/iovs.13-12518
96. Van Bergen T, Etienne I, Cunningham F, et al. The role of placental growth factor (PlGF) and its receptor system in retinal vascular diseases. Prog Retin Eye Res. 2019;69:116–136. doi:10.1016/j.preteyeres.2018.10.006
97. Perelman N, Selvaraj SK, Batra S, et al. Placenta growth factor activates monocytes and correlates with sickle cell disease severity. Blood. 2003;102(4):1506–1514. doi:10.1182/blood-2002-11-3422
98. Ando R, Noda K, Namba S, et al. Aqueous humour levels of placental growth factor in diabetic retinopathy. Acta Ophthalmol. 2014;92(3):e245–246. doi:10.1111/aos.12251
99. Lin M, Chen Y, Jin J, et al. Ischaemia-induced retinal neovascularisation and diabetic retinopathy in mice with conditional knockout of hypoxia-inducible factor-1 in retinal Muller cells. Diabetologia. 2011;54(6):1554–1566. doi:10.1007/s00125-011-2081-0
100. Demircan N, Safran BG, Soylu M, et al. Determination of vitreous interleukin-1 (IL-1) and tumour necrosis factor (TNF) levels in proliferative diabetic retinopathy. Eye (Lond). 2006;20(12):1366–1369. doi:10.1038/sj.eye.6702138
101. Behl Y, Krothapalli P, Desta T, et al. Diabetes-enhanced tumor necrosis factor-alpha production promotes apoptosis and the loss of retinal microvascular cells in type 1 and type 2 models of diabetic retinopathy. Am J Pathol. 2008;172(5):1411–1418. doi:10.2353/ajpath.2008.071070
102. Huang H, Gandhi JK, Zhong X, et al. TNFalpha is required for late BRB breakdown in diabetic retinopathy, and its inhibition prevents leukostasis and protects vessels and neurons from apoptosis. Invest Ophthalmol Vis Sci. 2011;52(3):1336–1344. doi:10.1167/iovs.10-5768
103. Joussen AM, Doehmen S, Le ML, et al. TNF-alpha mediated apoptosis plays an important role in the development of early diabetic retinopathy and long-term histopathological alterations. Mol Vis. 2009;15:1418–1428.
104. Rangasamy S, McGuire PG, Franco Nitta C, et al. Chemokine mediated monocyte trafficking into the retina: role of inflammation in alteration of the blood-retinal barrier in diabetic retinopathy. PLoS One. 2014;9(10):e108508. doi:10.1371/journal.pone.0108508
105. Yao Y, Li R, Du J, et al. Interleukin-6 and diabetic retinopathy: a systematic review and meta-analysis. Curr Eye Res. 2019;44(5):564–574. doi:10.1080/02713683.2019.1570274
106. Sonoda S, Sakamoto T, Shirasawa M, et al. Correlation between reflectivity of subretinal fluid in OCT images and concentration of intravitreal VEGF in eyes with diabetic macular edema. Invest Ophthalmol Vis Sci. 2013;54(8):5367–5374. doi:10.1167/iovs.13-12382
107. Sui A, Chen X, Shen J, et al. Inhibiting the NLRP3 inflammasome with MCC950 ameliorates retinal neovascularization and leakage by reversing the IL-1beta/IL-18 activation pattern in an oxygen-induced ischemic retinopathy mouse model. Cell Death Dis. 2020;11(10):901. doi:10.1038/s41419-020-03076-7
108. Mesquida M, Drawnel F, Fauser S. The role of inflammation in diabetic eye disease. Semin Immunopathol. 2019;41(4):427–445. doi:10.1007/s00281-019-00750-7
109. Sbardella D, Tundo GR, Mecchia A, et al. A novel and atypical NF-KB pro-inflammatory program regulated by a CamKII-proteasome axis is involved in the early activation of Muller glia by high glucose. Cell Biosci. 2022;12(1):108. doi:10.1186/s13578-022-00839-x
110. Vincent JA, Mohr S. Inhibition of caspase-1/interleukin-1beta signaling prevents degeneration of retinal capillaries in diabetes and galactosemia. Diabetes. 2007;56(1):224–230. doi:10.2337/db06-0427
111. Augustin HG, Koh GY, Thurston G, et al. Control of vascular morphogenesis and homeostasis through the angiopoietin-Tie system. Nat Rev Mol Cell Biol. 2009;10(3):165–177. doi:10.1038/nrm2639
112. Scholz A, Plate KH, Reiss Y. Angiopoietin-2: a multifaceted cytokine that functions in both angiogenesis and inflammation. Ann N Y Acad Sci. 2015;1347(1):45–51. doi:10.1111/nyas.12726
113. Dong N, Li X, Xiao L, et al. Upregulation of retinal neuronal MCP-1 in the rodent model of diabetic retinopathy and its function in vitro. Invest Ophthalmol Vis Sci. 2012;53(12):7567–7575. doi:10.1167/iovs.12-9446
114. Dong N, Xu B, Wang B, et al. Study of 27 aqueous humor cytokines in patients with type 2 diabetes with or without retinopathy. Mol Vis. 2013;19:1734–1746.
115. Jiang Z, Hennein L, Xu Y, et al. Elevated serum monocyte chemoattractant protein-1 levels and its genetic polymorphism is associated with diabetic retinopathy in Chinese patients with Type 2 diabetes. Diabet Med. 2016;33(1):84–90. doi:10.1111/dme.12804
116. Zeng Y, Cao D, Yu H, et al. Comprehensive analysis of vitreous humor chemokines in type 2 diabetic patients with and without diabetic retinopathy. Acta Diabetol. 2019;56(7):797–805. doi:10.1007/s00592-019-01317-6
117. Abu El-Asrar AM, Nawaz MI, Ahmad A, et al. Evaluation of proteoforms of the transmembrane chemokines CXCL16 and CX3CL1, their receptors, and their processing metalloproteinases ADAM10 and ADAM17 in proliferative diabetic retinopathy. Front Immunol. 2020;11:601639. doi:10.3389/fimmu.2020.601639
118. Ling M, Murali M. Analysis of the complement system in the clinical immunology laboratory. Clin Lab Med. 2019;39(4):579–590. doi:10.1016/j.cll.2019.07.006
119. Gerl VB, Bohl J, Pitz S, et al. Extensive deposits of complement C3d and C5b-9 in the choriocapillaris of eyes of patients with diabetic retinopathy. Invest Ophthalmol Vis Sci. 2002;43(4):1104–1108.
120. Rasmussen KL, Nordestgaard BG, Nielsen SF. Complement C3 and risk of diabetic microvascular disease: a cohort study of 95202 individuals from the general population. Clin Chem. 2018;64(7):1113–1124. doi:10.1373/clinchem.2018.287581
121. Xu D, Yi H, Yu S, et al. Association of complement C5 gene polymorphisms with proliferative diabetic retinopathy of type 2 diabetes in a chinese han population. PLoS One. 2016;11(3):e0149704. doi:10.1371/journal.pone.0149704
122. Garcia-Ramirez M, Canals F, Hernandez C, et al. Proteomic analysis of human vitreous fluid by fluorescence-based difference gel electrophoresis (DIGE): a new strategy for identifying potential candidates in the pathogenesis of proliferative diabetic retinopathy. Diabetologia. 2007;50(6):1294–1303. doi:10.1007/s00125-007-0627-y
123. Huang C, Fisher KP, Hammer SS, et al. Plasma exosomes contribute to microvascular damage in diabetic retinopathy by activating the classical complement pathway. Diabetes. 2018;67(8):1639–1649. doi:10.2337/db17-1587
124. Zhang J, Gerhardinger C, Lorenzi M. Early complement activation and decreased levels of glycosylphosphatidylinositol-anchored complement inhibitors in human and experimental diabetic retinopathy. Diabetes. 2002;51(12):3499–3504. doi:10.2337/diabetes.51.12.3499
125. Xu H, Chen M. Targeting the complement system for the management of retinal inflammatory and degenerative diseases. Eur J Pharmacol. 2016;787:94–104. doi:10.1016/j.ejphar.2016.03.001
126. Mandava N, Tirado-Gonzalez V, Geiger MD, et al. Complement activation in the vitreous of patients with proliferative diabetic retinopathy. Invest Ophthalmol Vis Sci. 2020;61(11):39. doi:10.1167/iovs.61.11.39
127. Geng P, Ding Y, Qiu L, et al. Serum mannose-binding lectin is a strong biomarker of diabetic retinopathy in Chinese patients with diabetes. Diabetes Care. 2015;38(5):868–875. doi:10.2337/dc14-1873
128. Holt CB, Hoffmann-Petersen IT, Hansen TK, et al. Association between severe diabetic retinopathy and lectin pathway proteins - an 18-year follow-up study with newly diagnosed type 1 diabetes patients. Immunobiology. 2020;225(3):151939. doi:10.1016/j.imbio.2020.151939
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Telehealth Encourages Patients with Diabetes in Racial and Ethnic Minority Groups to Return for in-Person Ophthalmic Care During the COVID-19 Pandemic
Ramsey DJ, Lasalle CC, Anjum S, Marx JL, Roh S
Clinical Ophthalmology 2022, 16:2157-2166
Published Date: 4 July 2022
miR-17-5p Promotes Glucose Uptake of HTR8/SVneo Trophoblast Cells by Inhibiting TXNIP/NLRP3 Inflammasome Pathway
Jiang Y, Wei L, Zhang H, Chen Y, Gao P, Zhang J, Zhou X, Zhu S, Du Y, Fang C, Li J, Feng L, He M, Wang S, Yu J
Diabetes, Metabolic Syndrome and Obesity 2022, 15:3361-3374
Published Date: 31 October 2022
Accelerated Wound Healing in Diabetic Rat by miRNA-185-5p and Its Anti-Inflammatory Activity
Wang KX, Zhao LL, Zheng LT, Meng LB, Jin L, Zhang LJ, Kong FL, Liang F
Diabetes, Metabolic Syndrome and Obesity 2023, 16:1657-1667
Published Date: 7 June 2023
Exploring the Correlation Between the Systemic Immune Inflammation Index (SII), Systemic Inflammatory Response Index (SIRI), and Type 2 Diabetic Retinopathy

Wang S, Pan X, Jia B, Chen S
Diabetes, Metabolic Syndrome and Obesity 2023, 16:3827-3836
Published Date: 24 November 2023
Targeting Senescence, Oxidative Stress, and Inflammation: Quercetin-Based Strategies for Ocular Diseases in Older Adults
Medoro A, Davinelli S, Scuderi L, Scuderi G, Scapagnini G, Fragiotta S
Clinical Interventions in Aging 2025, 20:791-813
Published Date: 7 June 2025