Back to Journals » Drug Design, Development and Therapy » Volume 19
Endoplasmic Reticulum-Targeting Natural Compounds: A Novel Frontier in Alleviating Liver Fibrosis
Authors Huang C, Hu W, Qi J, Su B, Yan H ![]() , Zhu YZ
, Zhu YZ
Received 1 September 2025
Accepted for publication 5 December 2025
Published 23 December 2025 Volume 2025:19 Pages 11465—11482
DOI https://doi.org/10.2147/DDDT.S564440
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Professor Tamer Ibrahim
Chengrong Huang,1,2,* Wei Hu,1,2,* Jia Qi,3 Bolun Su,2 Hongrui Yan,2 Yi Zhun Zhu1,2
1School of Pharmacy, Macau University of Science and Technology, Macau SAR, 999078, People’s Republic of China; 2Laboratory of Drug Discovery from Natural Resources and Industrialization, Macau University of Science and Technology, Macau SAR, 999078, People’s Republic of China; 3Department of Clinical Pharmacy, Xinhua Hospital Affiliated to Shanghai Jiaotong University School of Medicine, Shanghai, 200092, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Yi Zhun Zhu, Email [email protected]
Abstract: Liver fibrosis, a reversible yet critical stage in chronic liver disease progression, poses a significant global health burden with limited therapeutic options. This review comprehensively explores the molecular mechanisms of endoplasmic reticulum (ER) stress and its dual role in both parenchymal and non-parenchymal cells during liver fibrosis, alongside the therapeutic potential of natural compounds that target ER stress to alleviate fibrosis. Emerging evidence underscores ER stress and oxidative stress as pivotal drivers of hepatic fibrogenesis, primarily through activating hepatic stellate cells (HSCs). We systematically summarize a wide array of natural compounds, from polyphenols to terpenoids, that demonstrate potent anti-fibrotic effects by either ameliorating maladaptive ER stress in hepatocytes or selectively inducing pro-apoptotic ER stress in activated HSCs. Despite their promise, the clinical translation of these compounds is hampered by poor bioavailability and non-specific targeting. We highlight the groundbreaking potential of biomimetic nano-delivery systems, such as cell membrane-camouflaged nanoparticles, to overcome these barriers, offering precise targeting and enhanced therapeutic efficacy. Finally, we discuss current challenges and future directions, advocating for interdisciplinary efforts to advance ER stress-targeting strategies from bench to bedside.
Keywords: liver fibrosis, endoplasmic reticulum stress, unfolded protein response, natural compounds, targeted therapy
Introduction
Liver fibrosis is a pathological repair process triggered by chronic liver disease, characterized primarily by the excessive accumulation or abnormal distribution of extracellular matrix (ECM) components within the space of Disse.1 Liver fibrosis is typically caused by a various of etiological factor, including viral infections,2 metabolic abnormalities,3 alcohol use disorders,4 cholestasis,5 and autoimmune liver diseases.6 Additionally, long-term use of certain medications such as methotrexate, methyldopa, chlorpromazine, and tolbutamide has also been identified as a contributing factor to the progression of fibrosis.7–9 Without effective intervention, liver fibrosis may advance to cirrhosis or even hepatocellular carcinoma, posing a significant global health burden. According to the most recent Global Burden of Disease data, liver cirrhosis led to over 1.4 million deaths globally in 2021, with the global incidence having increased by 58% between 1990 and 2021, highlighting a growing challenge.10 Currently, treatments targeting the underlying causes, for example antiviral therapies for hepatitis B or C, can partially reverse fibrosis.11,12 Despite advancements in diagnostic techniques and therapeutic strategies, current treatments remain limited in efficacy, underscoring the urgent need for the development of novel anti-fibrotic therapies as a critical medical challenge.
Among the complex mechanisms driving liver fibrosis, endoplasmic reticulum (ER) stress has emerged as a pivotal contributor.13 Specifically, disruption of ER function triggers the unfolded protein response (UPR), which is a cellular stress response aimed at restoring ER homeostasis. However, under prolonged stress conditions, chronic ER stress leads to the sustained activation of UPR signaling pathways, ultimately resulting in inflammation, oxidative stress, and apoptosis.14 Notably, these processes play a central role in the activation of HSCs, the primary effector cells responsible for ECM deposition and fibrogenesis.15 Consequently, targeting ER stress has garnered significant attention as a promising therapeutic strategy for liver fibrosis.
Natural compounds have gained prominence in drug discovery and development due to their structural diversity, broad biological activities, and favorable safety profiles.16 Unlike synthetic drugs, many natural compounds exhibit lower toxicity and higher biocompatibility, making them attractive candidates for the treatment of chronic diseases. Recent studies have identified several natural compounds, such as curcumol, resveratrol, and chrysophanol, that mitigate ER stress by modulating UPR signaling pathways, thereby inhibiting HSC activation and attenuating fibrosis progression.17,18 These findings not only provide novel insights into the mechanisms of ER stress but also underscore the therapeutic potential of natural compounds in combating liver fibrosis. However, challenges such as poor bioavailability and limited targeting hinder their clinical application. Emerging nano-delivery systems, including liposomes and polymeric nanoparticles, offer promising solutions by enhancing stability, enabling targeted delivery to HSCs, and improving therapeutic efficacy.19 The integration of nanotechnology with natural compounds represents an advanced strategy for precise ER stress modulation in liver fibrosis treatment.
This review aims to comprehensively examine the role of ER stress in the pathogenesis of liver fibrosis and explore the therapeutic potential of natural compounds that target ER stress. First, we will elucidate the molecular mechanisms of ER stress and its contribution to liver fibrosis. Next, we will highlight key natural compounds that modulate ER stress and discuss their mechanisms of action. Finally, we will address the limitations of current research and outline future directions to advance this promising field. By providing a thorough overview, this review seeks to offer new perspectives on the treatment of liver fibrosis and to promote the development of natural compounds as viable therapeutic options.
ER Stress and UPR
The ER is a membranous organelle in eukaryotic cells, extending from the nuclear envelope and consisting of two distinct regions: the rough ER and the smooth ER. The ER is a multifunctional organelle responsible for the synthesis, processing, and transport of proteins and lipids, as well as the regulation of calcium homeostasis and cellular signaling. Under stress conditions such as nutrient deprivation or hypoxia, misfolded or unfolded proteins accumulate within the ER lumen. This accumulation disrupts protein homeostasis, triggering ER stress.20 The UPR responds to ER stress by enhancing protein folding, degrading misfolded proteins via ER-associated protein degradation (ERAD), and reducing new protein synthesis, restoring ER homeostasis and alleviating misfolded protein accumulation.21,22 The biological outcomes of ER stress activation vary depending on its severity: mild ER stress induces the UPR, lowering protein translation levels and reducing the generation of unfolded proteins, while severe ER stress triggers widespread UPR activation, including apoptotic pathways, ultimately leading to cell death.20,23
The classical UPR signaling pathways are initiated by three ER transmembrane proteins: inositol-requiring enzyme 1 (IRE1), activating transcription factor 6 (ATF6), and protein kinase R-like endoplasmic reticulum kinase (PERK). Among these, the 78 kDa glucose-regulated protein (GRP78), also known as immunoglobulin heavy chain-binding protein (BiP), is one of the most extensively studied ER chaperones.24 Under non-stress conditions, GRP78 binds to the luminal domains of these three ER transmembrane sensors. The accumulation of unfolded or misfolded proteins in the ER lumen leads to the dissociation of GRP78 from these sensors, thereby activating the UPR.25
IRE1 Pathway
IRE1 is the most evolutionarily conserved ER stress sensor and is a type I transmembrane protein found in the ER of all eukaryotes.26 It contains a serine/threonine protein kinase domain and an RNase domain. When the kinase domain of IRE1α detects misfolded or unfolded proteins, it dissociates from GRP78, dimerizes, and undergoes autophosphorylation, activating its RNase domain.27 The X-box binding protein 1 (XBP1) initially exists in an immature form but is spliced into its mature form (XBP1s) by the endonuclease activity of IRE1α under ER stress conditions. XBP1s then translocates to the nucleus, regulating the expression of genes involved in protein folding and ER-associated protein degradation (ERAD).28 Subsequently, ERAD binds to the cytoplasmic ubiquitin proteasome system, reducing the accumulation of unfolded proteins and restoring ER homeostasis. Additionally, IRE1α activation can lead to mRNA degradation through a process known as regulated IRE1α-dependent decay (RIDD), further reducing the protein load.29 Importantly, the IRE1α-XBP1s axis not only regulates genes involved in protein folding and ERAD but also directly orchestrates lipid biosynthesis. Under ER stress, XBP1s moves to the nucleus and activates transcriptional programs for fatty acid metabolism, triglyceride synthesis, and cholesterol biogenesis.30,31 Specifically, the metabolic rewiring driven by lipid metabolism is a key driver of HSC activation, a central event in the pathogenesis of liver fibrosis.32
ATF6
ATF6, a transmembrane protein that belongs to the leucine zipper transcription factor family, contains a cytoplasmic bZIP domain essential for transcription factor dimerization. It exists in two isoforms, ATF6α and ATF6β, whose activation is dependent on the dissociation of BiP and the exposure of their Golgi-targeting sequences.33,34 When misfolded or unfolded proteins accumulate, ATF6α translocates to the Golgi apparatus, where it undergoes cleavage by site-1 protease (MBTPS1) and site-2 protease (MBTPS2), releasing a truncated form of ATF6f (ATF6p50) that exhibits activated transcription factor activity.35 Subsequently, ATF6p50 migrates to the nucleus, where it binds to ER stress response elements I or II and induces the transcription and expression of genes such as GRP78, GRP94, C/EBP homologous protein (CHOP), and XBP1.36
PERK
PERK is a transmembrane protein with an N-terminal stress-sensing domain and a cytoplasmic kinase domain. During ER stress, PERK dissociates from GRP78 and is activated through dimerization and autophosphorylation.37 To alleviate ER stress, PERK phosphorylates the eukaryotic translation initiation factor 2α (eIF2α) at Ser-51, inhibiting the assembly of the 80S ribosome and preventing mRNA translation initiation, thereby reducing the load of newly synthesized proteins entering the ER lumen.38 Additionally, eIF2α enhances the expression of the transcription factor activating transcription factor 4 (ATF4), which activates CHOP and regulates the expression of growth arrest and DNA damage-inducible protein 34 (GADD34).39 This response promotes the dephosphorylation of eIF2α, restoring protein translation after ER damage. In this process, CHOP can also act as a pro-apoptotic factor by triggering caspase-8 through death receptor 5 (DR5).40 Figure 1.
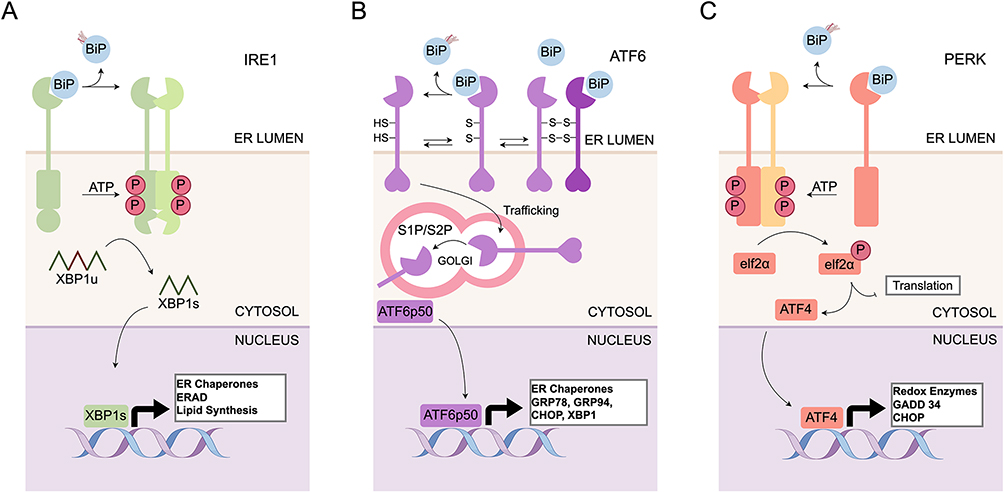 |
Figure 1 The unfolded protein response pathway. (A) IRE1 Pathway: BiP dissociation triggers IRE1 dimerization and RNase activation. IRE1 splices XBP1u mRNA to produce the active transcription factor XBP1s, which drives expression of ER cghaperones, ERAD and lipid metabolism genes. (B) ATF6 Pathway: Released ATF6 translocates to the Golgi for proteolytic cleavage. The cytosolic fragment (ATF6p50) moves to the nucleus to upregulate chaperones like GRP78 and XBP1. (C) PERK Pathway: Active PERK phosphorylates eIF2α, attenuating global protein synthesis while selectively translating ATF4. ATF4 induces genes for stress response and, under prolonged stress, the pro-apoptotic factor CHOP. |
ER Stress and Liver Fibrosis
The homeostasis of the liver is maintained through the synergistic action of diverse cell populations, primarily categorized into parenchymal and non-parenchymal cells. Approximately 65% of liver cells are parenchymal cells, primarily hepatocytes with a minor population of cholangiocytes, which are essential for liver regeneration following injury. Non-parenchymal cells, on the other hand, include HSCs, liver sinusoidal endothelial cells, and various immune components such as macrophages, natural killer cells, and dendritic cells, which collectively contribute to immune surveillance, inflammation regulation, and tissue repair.41–43
ER Stress in Parenchymal Cells
In parenchymal cells, particularly hepatocytes, ER stress serves as a critical molecular mechanism linking various hepatic injuries to fibrogenesis. Hepatocytes, being the primary metabolic units of the liver, are exceptionally vulnerable to ER stress due to their high protein synthesis burden and involvement in detoxification processes. Multiple pathogenic pathways converge on ER stress in these cells. For instance, QRICH1 enhances HBV-induced hepatic fibrosis by promoting ER stress-mediated high-mobility group box 1 (HMGB1) translocation and secretion, establishing a pro-fibrotic microenvironment.44 Similarly, GP73 upregulation during HBV infection mediates transforming growth factor β 1 (TGF-β1)-induced fibrogenic effects through the Smad2-ER stress axis in hepatocyte models.45
In metabolic contexts, CYP2E1 promotes liver fibrosis by inducing oxidative stress and ER stress in response to high-cholesterol diets, creating a vicious cycle of hepatocyte damage and inflammation.46 The loss of protective mechanisms also contributes significantly; PCSK9 deficiency exacerbates hepatic steatosis and fibrosis through CD36-mediated lipid accumulation, leading to ER stress and inflammatory activation.47 Conversely, several protective factors mitigate these processes. Fibroblast growth factor 21 (FGF21) demonstrates remarkable hepatoprotective effects by attenuating steatosis, lipotoxicity, and ER stress, thereby reducing the activation of downstream fibrogenic pathways.48 GDF15 similarly suppresses fibrosis-related genes and alleviates NASH phenotypes through modulation of ER stress responses.49
The transcription factor CHOP, a key mediator of ER stress-induced apoptosis, is upregulated in various fibrotic models and promotes fibrosis not only through hepatocyte death but also by facilitating alternative activation of pro-fibrotic macrophages in schistosomiasis models.50 Additionally, TRC8 deficiency exacerbates NASH progression by enhancing ER stress and apoptosis, while its overexpression ameliorates these effects, highlighting its crucial role in maintaining ER protein homeostasis in hepatocytes.51 These findings collectively demonstrate that ER stress in parenchymal cells acts as both a sensor of hepatic injury and an active contributor to the fibrotic cascade through multiple interconnected mechanisms.
ER Stress in Non-Parenchymal Cells
The role of ER stress in non-parenchymal cells, particularly HSCs and immune cells, represents a crucial mechanism in fibrosis progression. In HSCs, ER stress is not merely a consequence of activation but actively drives their transition to pro-fibrotic myofibroblasts. The thiol-disulfide oxidoreductase TXNDC5 is significantly upregulated in activated HSCs, where it promotes cell viability and collagen production through ER stress modulation, making it a promising therapeutic target.52 Similarly, Derlin-1 accelerates NASH progression by enhancing ERAD activity and ER stress, leading to RIPK3-mediated necroptosis in HSCs.53
The collagen secretion machinery in HSCs is intimately connected to ER homeostasis. TANGO1, an ER-resident protein essential for collagen transport, is upregulated during HSC activation in an XBP1-dependent manner. Its depletion causes procollagen I retention, inducing severe UPR activation and ultimately HSC apoptosis.54 Ion channels play a pivotal role in modulating ER stress and its downstream effects in HSCs, thereby influencing liver fibrosis. For instance, the acid-sensing ion channel 1a (ASIC1a) promotes HSC activation primarily by facilitating proton and calcium influx. This disruption of ionic homeostasis, particularly calcium overload within the ER lumen, impairs chaperone function and protein folding, thereby activating the UPR. The ensuing ER stress then drives fibrogenesis through the PI3K/AKT pathway.55,56 The UPR branches play distinct roles, with IRE1α signaling typically promoting fibrogenesis, while AT2R attenuates fibrosis by suppressing the IRE1α-XBP1 pathway in HSCs.57
In immune cells, particularly macrophages, ER stress significantly influences fibrotic progression. IRE1α activation in macrophages promotes pyroptosis and accelerates MASH progression through the IRE1α-GSDMD pathway, with its inhibition showing protective effects.58 CHOP promotes liver fibrosis in schistosomiasis by facilitating alternative macrophage activation and M2 polarization.50 Additionally, adipocyte-derived factors contribute to this network; PCPE1 functions as a BAT-derived profibrotic adipokine that promotes liver fibrosis through ER stress-induced secretion.59
The interplay between different non-parenchymal cells further complicates the ER stress-fibrosis axis. HMGB1 released from injured hepatocytes activates HSCs via toll-like receptor 4(TLR4)/RAGE-mediated ER stress, creating a paracrine loop that sustains fibrogenesis.60 Meanwhile, protective mechanisms also exist; Atp6v0d2 maintains hepatic lipid homeostasis and suppresses MASH progression by inhibiting XBP1-regulated ER stress in macrophages.61 These findings highlight the complex, cell-type-specific roles of ER stress in non-parenchymal cells and emphasize the potential of targeted therapies that modulate specific ER stress pathways in different cell populations to combat liver fibrosis.
In summary, ER stress serves as a pivotal pathogenic link that connects diverse hepatic injuries to the progression of liver fibrosis. It orchestrates fibrogenesis through cell-type-specific mechanisms: in parenchymal cells, such as hepatocytes, ER stress promotes apoptosis, inflammatory signaling, and the release of pro-fibrotic factors. Concurrently, in non-parenchymal cells, it directly activates HSCs and modulates immune responses, including macrophage polarization. The intricate cross-talk between these cell populations, particularly between injured hepatocytes and HSCs, as shown in Figure 2. This multifaceted involvement highlights the therapeutic potential of targeting cell-specific ER stress pathways as a promising strategy to mitigate liver fibrosis. Table 1.
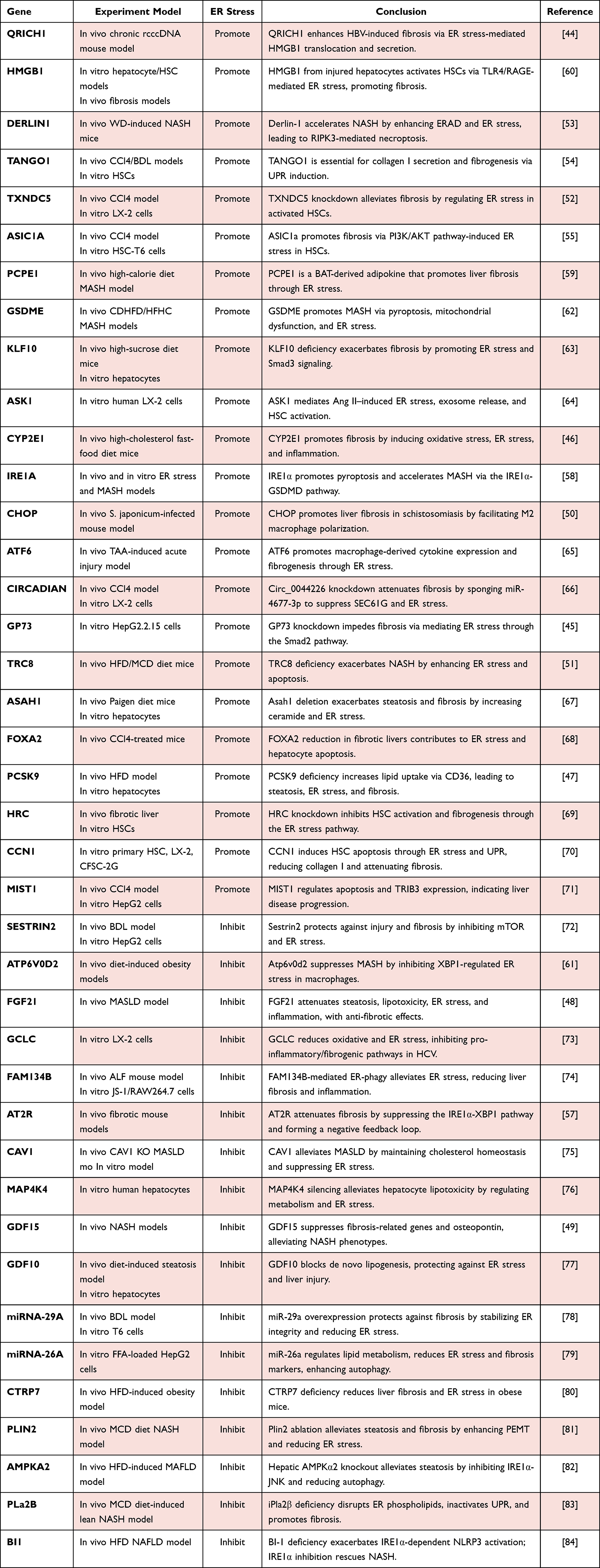 |
Table 1 Genetic Regulators of Liver Fibrosis via ER Stress Pathways |
 |
Figure 2 Cross-talk between hepatocytes and stellate cells in the formation of fibrosis. Injured hepatocytes release HMGB1, TGF-β1, and ROS, activating HSCs into myofibroblasts. Activated HSCs secrete collagen, cytokines (eg, IL-6, TNF-α), and ROS, exacerbating hepatocyte damage and ER stress. Key pathways include TLR4/RAGE signaling, UPR activation, and oxidative stress. ER stress in hepatocytes promotes apoptosis and inflammatory signaling, while in HSCs, it drives collagen production and myofibroblast transition. The paracrine loop sustains fibrosis through HMGB1, TGF-β1, and ROS. Therapeutic targets include TXNDC5, IRE1α, and ASIC1a to disrupt this pathological interaction, highlighting the complex interplay between hepatocytes and HSCs in liver fibrosis progression. |
ER Stress-Targeting Natural Compound
ER stress is increasingly recognized as a central mechanism driving hepatic fibrogenesis, drawing significant interest as a therapeutic target. A wide array of natural compounds demonstrates potent anti-fibrotic efficacy by modulating ER stress pathways. These substances can be functionally grouped into two strategic categories: one that suppresses detrimental ER stress to preserve hepatocyte function and inhibit HSCs activation, and another that selectively induces pro-apoptotic ER stress in activated HSCs, promoting their elimination.
Compounds That Ameliorate ER Stress
Numerous natural compounds alleviate liver fibrosis primarily by inhibiting maladaptive unfolded protein response (UPR) signaling, which in turn reduces hepatocyte apoptosis, inflammation, and HSC activation. Among these, polyphenols and flavonoids constitute a prominently effective class. For example, resveratrol downregulates pivotal ER stress markers such as CHOP, Bip, and caspases-3/7/12, while concurrently inhibiting pro-inflammatory pathways like TGF-β and NF-κB in carbon tetrachloride (CCl4)-induced rodent models and in vitro HSC cultures.85 Similarly, didymin exerts a broad inhibitory effect across all three UPR sensors, such as ATF6, IRE1α, and PERK, thereby mitigating apoptotic signaling and abnormal glycerophospholipid metabolism.86 Additional flavonoids, including naringenin and tetrahydropalmatine, suppress critical UPR axes such as PERK-eIF2α-ATF4-CHOP and IRE1α-XBP1, significantly attenuating fibrotic progression.87,88
Beyond polyphenols, several alkaloids and organic acids also play crucial roles. Berberine specifically targets the ATF6/SREBP-1c pathway, reducing lipid synthesis and ER stress, which leads to improved hepatic steatosis and fibrosis in metabolic models.89 Likewise, p-coumaric acid and oxymatrine alleviate oxidative and ER stress, with oxymatrine notably enhancing calcium homeostasis in HSCs through regulation of Sarco/endoplasmic reticulum Ca2⁺-ATPase 2 (SERCA2).90,91 Among terpenoids and other structurally distinct compounds, additional mechanisms emerge. Irisin modulates PERK phosphorylation to stabilize the RNA-binding protein HNRNPA1, thereby conferring protection against fibrotic signaling.92 Meanwhile, lycopene inhibits the apoptosis signal-regulating kinase 1 (ASK1)-JNK pathway, and tanshinone IIA activates the PPARα/FGF21 axis, both resulting in suppression of ER stress and fibrosis in non-alcoholic steatohepatitis (NASH) models.93,94 Furthermore, salvianolic acid A activates SIRT1, leading to HSF1 deacetylation and subsequent downregulation of the UPR, offering another layer of protection against HSC activation.95
Compounds That Induce Pro-Apoptotic ER Stress in Activated HSCs
In contrast, a complementary anti-fibrotic strategy employs natural compounds to selectively induce lethal levels of ER stress in activated HSCs, triggering their apoptosis and facilitating the clearance of these key fibrogenic cells. Quinones and terpenoids are notably effective in this regard. Cryptotanshinone promotes activated HSC apoptosis by upregulating ER stress markers including CHOP and GRP78 and activating UPR-related molecules such as PERK, IRE1α, and ATF4.96 Curcumol, a sesquiterpenoid, uniquely provokes necroptosis in activated HSCs via RIP3, a process modulated through Sirt1/Notch signaling under ER stress conditions.97
Flavonoids also contribute significantly to this targeted approach. Quercetin and oroxylin A effectively initiate ER stress pathway activation, resulting in increased CHOP expression, altered Bcl-2-associated X protein (Bax)/ B-cell lymphoma 2 (Bcl-2) ratios, and caspase activation, collectively driving activated HSC apoptosis.96,98 Similarly, malvidin induces programmed cell death through both ER stress and mitochondrial pathways.99 Even some synthetic agents, such as etoposide, operate through this mechanism, inducing ER stress and JNK-mediated apoptosis in human HSC line LX-2.100
Novel Mechanisms and Emerging Compounds
Recent research continues to uncover natural compounds with innovative anti-fibrotic mechanisms centered on ER stress modulation. For instance, 12-deoxyphorbol 13-palmitate represents a novel strategy by specifically targeting the protein Apolipoprotein L2 (APOL2), disrupting its interaction with SERCA2, and thereby inhibiting the pro-fibrotic PERK–HES1 signaling cascade.101 Another compound, senkyunolide A, exhibits specificity for cholestatic fibrosis models. It binds to CLCC1 and promotes its ubiquitination, which inhibits ER calcium release and suppresses ER autophagy, ultimately reducing cholangiocyte proliferation and ductular reaction.102 Additionally, arctigenin mitigates HSC activation through dual mechanisms: not only does it suppress ER stress, but it also restores lipid homeostasis via modulation of the ERAD pathway, underscoring the interplay between proteostasis and metabolic regulation in liver fibrosis.103 Table 2
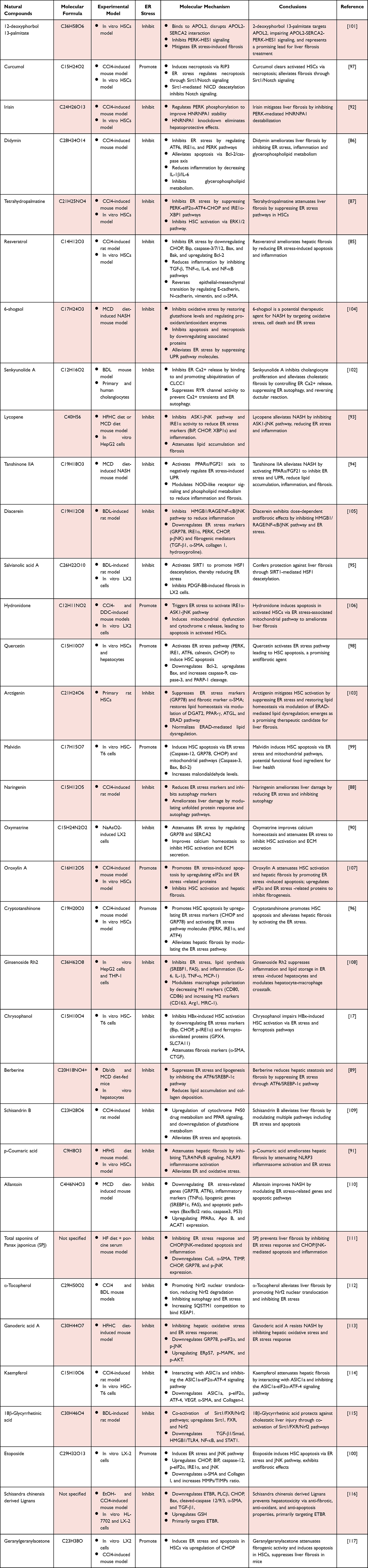 |
Table 2 Natural Compounds Targeting ER Stress for Liver Fibrosis |
Current Research and Future Perspective
The compelling preclinical evidence supporting ER stress as a therapeutic target in liver fibrosis has not yet translated into effective clinical treatments. A primary obstacle lies in the suboptimal pharmacokinetic profiles of many promising natural compounds, including poor aqueous solubility, metabolic instability, and, crucially, non-specific biodistribution, which collectively result in low bioavailability and limited efficacy at the target site.118 Therefore, the development of advanced delivery strategies that can enhance the stability of these compounds and direct them precisely to specific liver cell populations is paramount for clinical translation.
Nanoparticle-based delivery systems have emerged as a transformative strategy to overcome these pharmacological hurdles.119,120 The core advantage of these systems is their ability to be engineered for cell-specific targeting within the complex liver microenvironment. For instance, targeting hepatocytes can be achieved by functionalizing nanocarriers with galactose or lactose ligands, which are recognized by the asialoglycoprotein receptor highly expressed on hepatocytes.121 This approach allows for the precise delivery of therapeutic agents to alleviate ER stress induced by metabolic insults or viral infections within parenchymal cells. However, the success of anti-fibrotic therapy hinges even more critically on targeting non-parenchymal cells. A well-established strategy exploits the vitamin A storage characteristic of HSCs by using retinol-coupled liposomes or polymeric nanoparticles, which enables the selective delivery of pro-apoptotic or anti-fibrotic agents directly to activated HSCs. Vitamin A-coupled nanoparticles, such as retinol-modified crosslinking nanopolyplexes, hijack retinol-binding protein for precise activated HSC targeting and ROS-responsive siRNA release, effectively reducing collagen deposition and liver injury in preclinical models.122 Furthermore, other targeting ligands, such as those binding to the platelet-derived growth factor receptor β or integrins upregulated on activated HSCs, are under active investigation to achieve high specificity.123,124
Beyond conventional active targeting, the frontier of drug delivery involves biomimetic nanocarriers.125,126 These innovative systems, such as those camouflaged with membranes derived from platelets or even activated HSCs themselves, inherit the complex surface proteins and “self” markers of the source cells. This biomimicry offers unparalleled targeting precision to the fibrotic niche and enhances immune evasion, representing a significant breakthrough for site-specific drug release. Moreover, the development of “smart” nanocarriers, such as the CXCR4-targeted ROS-responsive platform AMD-Dex-ROS-sorafenib, leverages high intracellular ROS levels in activated HSCs for thioketal rupture and on-demand drug release.127
Notwithstanding the promise of advanced delivery systems, several formidable challenges remain. Firstly, the vast majority of evidence is derived from rodent models, creating a critical gap in clinical validation. There is an urgent and unmet need for well-designed Phase I and II clinical trials that rigorously evaluate the safety and efficacy of the most promising candidates, such as curcumin or berberine, in patients with chronic liver diseases.128 Another significant challenge lies in the dualistic nature of the UPR. While inhibiting chronic, maladaptive ER stress is beneficial, uncontrolled suppression can disrupt the UPR’s essential adaptive functions in protein homeostasis.
To successfully bridge the translational gap, future research should prioritize several key directions. Advanced human-relevant preclinical models, including patient-derived organoids and 3D-bioprinted liver tissues, will be crucial for better predicting drug efficacy and toxicity before costly clinical trials.129,130 Concurrently, multi-omics technologies such as genomics and proteomics, along with high-throughput screening, will accelerate the discovery of novel natural compounds and precisely define their molecular targets within the complex UPR network.131 Finally, exploring rational combination therapies that synergize ER stress-targeting natural compounds delivered via sophisticated nanocarriers with existing anti-fibrotic agents or first-line treatments like antivirals represents a highly promising strategic direction. In summary, by addressing these challenges through interdisciplinary collaboration integrating pharmacology, nanotechnology, and clinical science, the field can move closer to harnessing the full potential of ER stress modulation as a viable and precise therapeutic strategy for patients with liver fibrosis.
Conclusion
This review establishes the unequivocal role of ER stress as a central mechanism in the pathogenesis of liver fibrosis, orchestrating a complex interplay of cellular responses in both hepatocytes and HSCs. The extensive catalogue of natural compounds presented herein not only validates ER stress as a druggable target but also offers a rich reservoir of potential therapeutic agents capable of modulating the UPR to halt or reverse fibrogenesis. However, the inherent limitations of these compounds, particularly their pharmacokinetic drawbacks, underscore a critical translational challenge. The advent of sophisticated biomimetic nanotechnologies, exemplified by cell membrane-camouflaged platforms, provides a transformative solution to these obstacles, enabling targeted delivery and maximizing on-site therapeutic efficacy while minimizing off-target effects. Looking forward, the future of anti-fibrotic therapy lies in the convergence of pharmacology, nanotechnology, and precision medicine. Prioritizing the development of intelligent nano-formulations for lead natural compounds, validating their efficacy in human-relevant models, and exploring synergistic combination therapies will be paramount. By harnessing these innovative strategies, the field can successfully translate the compelling preclinical promise of ER stress modulation into tangible clinical benefits for patients suffering from liver fibrosis.
Abbreviations
APOL2, Apolipoprotein L2; ASK1, Apoptosis Signal-regulating Kinase 1; ATF4, Activating Transcription Factor 4; ATF6, Activating Transcription Factor 6; Bax, Bcl-2-Associated X Protein; Bcl-2, B-Cell Lymphoma 2; BiP, Immunoglobulin Heavy Chain-Binding Protein; CHOP, C/EBP Homologous Protein; ECM, Extracellular Matrix; eIF2α, Eukaryotic Initiation Factor 2 Alpha; ER, Endoplasmic Reticulum; ERAD, Endoplasmic Reticulum-Associated Protein Degradation; ER stress, Endoplasmic Reticulum Stress; FGF21, Fibroblast Growth Factor 21; GADD34, Growth Arrest and DNA Damage-inducible protein 34; GSDMD, Gasdermin D; GRP78, 78-kDa Glucose-Regulated Protein; HSCs, Hepatic Stellate Cells; HMGB1, High-Mobility Group Box 1; PERK, Protein Kinase R-like Endoplasmic Reticulum Kinase; RIDD, Regulated IRE1α-Dependent Decay; SERCA2, Sarco/Endoplasmic Reticulum Ca2⁺-ATPase 2; TGF-β1, Transforming Growth Factor Beta 1; TLR4, Toll-Like Receptor 4; UPR, Unfolded Protein Response; XBP1, X-Box Binding Protein 1.
Acknowledgments
Figure were created in https://www.figdraw.com and https://BioRender.com.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
Macau Science and Technology Development fund (FDCT (0012/2021/AMJ, 0001/2024/RDP, 0001/2024/AKP, 0092/2022/A2, 0144/2022/A3).
Disclosure
The authors declare no competing interest.
References
1. Kisseleva T, Brenner D. Molecular and cellular mechanisms of liver fibrosis and its regression. Nat Rev Gastroenterol Hepatol. 2021;18(3):151–166.
2. Cardoso AC, Figueiredo-Mendes C, Villela-Nogueira CA, Marcellin P. Staging fibrosis in chronic viral hepatitis. Viruses. 2022;14(4):660. doi:10.3390/v14040660
3. Fu J, Yi Y, Li L, Shu X, Chang Y. Association of metabolic abnormalities and the risk of hepatic fibrosis. Npj Gut and Liver. 2025;2(1):13. doi:10.1038/s44355-025-00025-z
4. Caputo F, Penitenti F, Bergonzoni B, et al. Alcohol use disorders and liver fibrosis: an update. Minerva Med. 2024;115(3):354–363. doi:10.23736/S0026-4806.24.09203-6
5. Negi SS, Sakhuja P, Malhotra V, Chaudhary A. Factors predicting advanced hepatic fibrosis in patients with postcholecystectomy bile duct strictures. Arch Surg. 2004;139(3):299–303. doi:10.1001/archsurg.139.3.299
6. Pellicano R, Ferro A, Cicerchia F, Mattivi S, Fagoonee S, Durazzo M. Autoimmune hepatitis and fibrosis. J Clin Med. 2023;12(5):1979. doi:10.3390/jcm12051979
7. Bichenapally S, Khachatryan V, Muazzam A, et al. Risk of liver fibrosis in methotrexate-treated patients: a systematic review. Cureus. 2022;14(10):e30910. doi:10.7759/cureus.30910
8. Tiwari V, Shandily S, Albert J, et al. Insights into medication-induced liver injury: understanding and management strategies. Toxicol Rep. 2025;14:101976. doi:10.1016/j.toxrep.2025.101976
9. Ghabril M, Vuppalanchi R, Chalasani N. Drug-induced liver injury in patients with chronic liver disease. Liver Int. 2025;45(3):e70019. doi:10.1111/liv.70019
10. Luo X, He Y, Jiang Z, Liao J. Global burden of liver cirrhosis: trends from 1990–2021 and projections to 2060. J Health Popul Nutr. 2025;44(1):370. doi:10.1186/s41043-025-01109-5
11. Liang H, Zheng X, Liu Y, et al. Effects of different antiviral treatments on liver inflammation and fibrosis in patients with chronic hepatitis B. J Viral Hepat. 2025;32(4):e70019. doi:10.1111/jvh.70019
12. Lee SH, Shin HP, Lee JI. Real-world single-center experience with direct-acting antivirals for improvement of the liver fibrosis after chronic hepatitis C treatment. Antivir Chem Chemother. 2020;28:2040206620974835. doi:10.1177/2040206620974835
13. Ajoolabady A, Kaplowitz N, Lebeaupin C, et al. Endoplasmic reticulum stress in liver diseases. Hepatology. 2023;77(2):619–639. doi:10.1002/hep.32562
14. Zheng Z, Shang Y, Tao J, Zhang J, Sha B. Endoplasmic reticulum stress signaling pathways: activation and diseases. Curr Protein Pept Sci. 2019;20(9):935–943. doi:10.2174/1389203720666190621103145
15. Mannaerts I, Thoen LFR, Eysackers N, et al. Unfolded protein response is an early, non-critical event during hepatic stellate cell activation. Cell Death Dis. 2019;10(2):98. doi:10.1038/s41419-019-1327-5
16. Singh K, Gupta JK, Chanchal DK, et al. Natural products as drug leads: exploring their potential in drug discovery and development. Naunyn. 2025;398(5):4673–4687. doi:10.1007/s00210-024-03622-6
17. Kuo C-Y, Chiu V, Hsieh P-C, et al. Chrysophanol attenuates hepatitis B virus X protein-induced hepatic stellate cell fibrosis by regulating endoplasmic reticulum stress and ferroptosis. J Pharmacol Sci. 2020;144(3):172–182. doi:10.1016/j.jphs.2020.07.014
18. Ho CH, Huang JH, Sun MS, Tzeng IS, Hsu YC, Kuo CY. Wild bitter melon extract regulates LPS-induced hepatic stellate cell activation, inflammation, endoplasmic reticulum stress, and ferroptosis. Evid Based Complement Alternat Med. 2021;2021:6671129. doi:10.1155/2021/6671129
19. Yuan Y, Li J, Chen M, et al. Nano-encapsulation of drugs to target hepatic stellate cells: toward precision treatments of liver fibrosis. J Control Release. 2024;376:318–336. doi:10.1016/j.jconrel.2024.10.012
20. Chen X, Shi C, He M, Xiong S, Xia X. Endoplasmic reticulum stress: molecular mechanism and therapeutic targets. Signal Transduct Target Ther. 2023;8(1):352. doi:10.1038/s41392-023-01570-w
21. Hetz C, Zhang K, Kaufman RJ. Mechanisms, regulation and functions of the unfolded protein response. Nat Rev Mol Cell Biol. 2020;21(8):421–438. doi:10.1038/s41580-020-0250-z
22. Keerthiga R, Pei DS, Fu A. Mitochondrial dysfunction, UPR(mt) signaling, and targeted therapy in metastasis tumor. Cell Biosci. 2021;11(1):186. doi:10.1186/s13578-021-00696-0
23. Zhang J, Guo J, Yang N, Huang Y, Hu T, Rao C. Endoplasmic reticulum stress-mediated cell death in liver injury. Cell Death Dis. 2022;13(12):1051. doi:10.1038/s41419-022-05444-x
24. Gonzalez-Gronow M, Gopal U, Austin RC, Pizzo SV. Glucose-regulated protein (GRP78) is an important cell surface receptor for viral invasion, cancers, and neurological disorders. IUBMB Life. 2021;73(6):843–854. doi:10.1002/iub.2502
25. Xu F, Wang L. Deciphering ER stress-unfolded protein response relationship by visualizing unfolded proteins in the ER. Cell Rep. 2024;43(6):114358. doi:10.1016/j.celrep.2024.114358
26. Liu S, Zhang X, Yao X, et al. Mammalian IRE1α dynamically and functionally coalesces with stress granules. Nat Cell Biol. 2024;26(6):917–931. doi:10.1038/s41556-024-01418-7
27. Mori K. Evolutionary aspects of the unfolded protein response. Cold Spring Harb Perspect Biol. 2022;14(12). doi:10.1101/cshperspect.a041262
28. Tang Y, Yao T, Tian X, et al. Hepatic IRE1alpha-XBP1 signaling promotes GDF15-mediated anorexia and body weight loss in chemotherapy. J Exp Med. 2024;221(7). doi:10.1084/jem.20231395
29. Almanza A, Mnich K, Blomme A, et al. Regulated IRE1alpha-dependent decay (RIDD)-mediated reprograming of lipid metabolism in cancer. Nat Commun. 2022;13(1):2493. doi:10.1038/s41467-022-30159-0
30. Park SM, Kang TI, So JS. Roles of XBP1s in transcriptional regulation of target genes. Biomedicines. 2021;9(7):791. doi:10.3390/biomedicines9070791
31. Moncan M, Mnich K, Blomme A, Almanza A, Samali A, Gorman AM. Regulation of lipid metabolism by the unfolded protein response. J Cell & Mol Med. 2021;25(3):1359–1370. doi:10.1111/jcmm.16255
32. Silva CM, Ferrari GD, Alberici LC, Malaspina O, Moraes KCM. Cellular and molecular effects of silymarin on the transdifferentiation processes of LX-2 cells and its connection with lipid metabolism. Mol Cell Biochem. 2020;468(1–2):129–142. doi:10.1007/s11010-020-03717-7
33. Hien LT, Back SH. Establishment of a reporter system for monitoring activation of the ER stress transducer ATF6beta. Biochem Biophys Res Commun. 2021;558:1–7. doi:10.1016/j.bbrc.2021.04.052
34. Stauffer WT, Arrieta A, Blackwood EA, Glembotski CC. Sledgehammer to scalpel: broad challenges to the heart and other tissues yield specific cellular responses via transcriptional regulation of the ER-stress master regulator ATF6alpha. Int J Mol Sci. 2020;21(3):1134. doi:10.3390/ijms21031134
35. Lei Y, Yu H, Ding S, Liu H, Liu C, Fu R. Molecular mechanism of ATF6 in unfolded protein response and its role in disease. Heliyon. 2024;10(5):e25937. doi:10.1016/j.heliyon.2024.e25937
36. Keramidas P, Pitou M, Papachristou E, Choli-Papadopoulou T. Insights into the activation of unfolded protein response mechanism during coronavirus infection. Curr Issues Mol Biol. 2024;46(5):4286–4308. doi:10.3390/cimb46050261
37. Romine IC, Wiseman RL. PERK signaling regulates extracellular proteostasis of an amyloidogenic protein during endoplasmic reticulum stress. Sci Rep. 2019;9(1):410. doi:10.1038/s41598-018-37207-0
38. Uppala JK, Ghosh C, Sathe L, Dey M. Phosphorylation of translation initiation factor eIF2α at Ser51 depends on site- and context-specific information. FEBS Lett. 2018;592(18):3116–3125. doi:10.1002/1873-3468.13214
39. Shacham T, Patel C, Lederkremer GZ. PERK pathway and neurodegenerative disease: to inhibit or to activate? Biomolecules. 2021;11(3):354. doi:10.3390/biom11030354
40. Song G, Weng F, Zou B, et al. Potential therapeutic action of tauroursodeoxycholic acid against cholestatic liver injury via hepatic Fxr/Nrf2 and CHOP-DR5-caspase-8 pathway. Clin Sci. 2023;137(7):561–577. doi:10.1042/CS20220674
41. Wei Y, Bingyu W, Lei Y, Xingxing Y. The antifibrotic role of natural killer cells in liver fibrosis. Exp Biol Med. 2022;247(14):1235–1243. doi:10.1177/15353702221092672
42. Zhang Y, Wu Y, Shen W, Wang B, Yuan X. Crosstalk between NK cells and hepatic stellate cells in liver fibrosis (Review). Mol Med Rep. 2022;25(6). doi:10.3892/mmr.2022.12724
43. Wang B, Yang L, Yuan X, Zhang Y. Roles and therapeutic targeting of dendritic cells in liver fibrosis. J Drug Target. 2024;32(6):647–654. doi:10.1080/1061186X.2024.2347365
44. Feng Y, Geng Y, Liu Z, et al. QRICH1, as a key effector of endoplasmic reticulum stress, enhances HBV in promoting HMGB1 translocation and secretion in hepatocytes. Immunobiology. 2025;230(3):152913. doi:10.1016/j.imbio.2025.152913
45. He Y, Cai L, Liu L, et al. HBV infection upregulates GP73 expression to promote liver fibrosis by enhancing ER stress via the Smad2 pathway. Immunol Res. 2025;73(1):100. doi:10.1007/s12026-025-09656-z
46. Abdelmegeed MA, Choi Y, Godlewski G, et al. Cytochrome P450-2E1 promotes fast food-mediated hepatic fibrosis. Sci Rep. 2017;7:39764. doi:10.1038/srep39764
47. Lebeau PF, Byun JH, Platko K, et al. Pcsk9 knockout exacerbates diet-induced non-alcoholic steatohepatitis, fibrosis and liver injury in mice. JHEP Rep. 2019;1(6):418–429. doi:10.1016/j.jhepr.2019.10.009
48. Cui X, Sun Q, Wang H. Targeting fibroblast growth factor (FGF)-21: a promising strategy for metabolic dysfunction-associated steatotic liver disease treatment. Front Pharmacol. 2025;16:1510322. doi:10.3389/fphar.2025.1510322
49. Kim KH, Kim SH, Han DH, Jo YS, Lee YH, Lee MS. Growth differentiation factor 15 ameliorates nonalcoholic steatohepatitis and related metabolic disorders in mice. Sci Rep. 2018;8(1):6789. doi:10.1038/s41598-018-25098-0
50. Duan M, Yang Y, Peng S, et al. C/EBP Homologous Protein (CHOP) activates macrophages and promotes liver fibrosis in schistosoma japonicum-infected mice. J Immunol Res. 2019;2019:5148575. doi:10.1155/2019/5148575
51. Chang PC, Tsai HW, Chiang MT, Huang PL, Shyue SK, Chau LY. TRC8 downregulation contributes to the development of non-alcoholic steatohepatitis by exacerbating hepatic endoplasmic reticulum stress. Biochim Biophys Acta. 2015;1852(11):2339–2351. doi:10.1016/j.bbadis.2015.08.022
52. Zhang L, Zeng J, Wu H, et al. Knockdown of TXNDC5 alleviates CCL4-induced hepatic fibrosis in mice by enhancing endoplasmic reticulum stress. Am J Med Sci. 2023;366(6):449–457. doi:10.1016/j.amjms.2023.08.016
53. Wang T, Wang D, Kuang G, et al. Derlin-1 promotes diet-induced non-alcoholic fatty liver disease via increasing RIPK3-mediated necroptosis. Free Radic Biol Med. 2024;217:29–47. doi:10.1016/j.freeradbiomed.2024.03.014
54. Maiers JL, Kostallari E, Mushref M, et al. The unfolded protein response mediates fibrogenesis and collagen I secretion through regulating TANGO1 in mice. Hepatology. 2017;65(3):983–998. doi:10.1002/hep.28921
55. Zuo L, Zhu Y, Hu L, et al. PI3-kinase/Akt pathway-regulated membrane transportation of acid-sensing ion channel 1a/Calcium ion influx/endoplasmic reticulum stress activation on PDGF-induced HSC activation. J Cell Mol Med. 2019;23(6):3940–3950. doi:10.1111/jcmm.14275
56. Kong L, Huang H, Luan S, Liu H, Ye M, Wu F. Inhibition of ASIC1a-mediated ERS improves the activation of HSCs and copper transport under copper load. Front Pharmacol. 2021;12:653272. doi:10.3389/fphar.2021.653272
57. An Y, Xu C, Liu W, et al. Angiotensin II type-2 receptor attenuates liver fibrosis progression by suppressing IRE1α-XBP1 pathway. Cell Signal. 2024;113:110935. doi:10.1016/j.cellsig.2023.110935
58. Zeng X, Wu T, Xu Q, et al. Inhibition of IRE-1α alleviates pyroptosis and metabolic dysfunction-associated steatohepatitis by suppressing gasdermin D. Liver Int. 2025;45(2):e16234. doi:10.1111/liv.16234
59. Hsiao YT, Yoshida Y, Okuda S, et al. PCPE-1, a brown adipose tissue-derived cytokine, promotes obesity-induced liver fibrosis. EMBO J. 2024;43(21):4846–4869. doi:10.1038/s44318-024-00196-0
60. He Q, Fu Y, Ding X, et al. High-mobility group box 1 induces endoplasmic reticulum stress and activates hepatic stellate cells. Lab Invest. 2018;98(9):1200–1210. doi:10.1038/s41374-018-0085-9
61. Wang Z, Chen X, Li J, et al. Macrophages Atp6v0d2 regulates XBP1-mediated cholesterol metabolism to suppress metabolic dysfunction-associated steatohepatitis progression. Int Immunopharmacol. 2025;161:115088. doi:10.1016/j.intimp.2025.115088
62. Zhu JH, Ouyang SX, Zhang GY, et al. GSDME promotes MASLD by regulating pyroptosis, Drp1 citrullination-dependent mitochondrial dynamic, and energy balance in intestine and liver. Cell Death Differ. 2024;31(11):1467–1486. doi:10.1038/s41418-024-01343-0
63. Lee J, Oh AR, Lee HY, Moon YA, Lee HJ, Cha JY. Deletion of KLF10 leads to stress-induced liver fibrosis upon high sucrose feeding. Int J Mol Sci. 2020;22(1):331. doi:10.3390/ijms22010331
64. Fang PP, Pan CW, Lin W, et al. ASK1 enhances angiotensin ii-induced liver fibrosis in vitro by mediating endoplasmic reticulum stress-dependent exosomes. Mediators Inflamm. 2020;2020:8183713. doi:10.1155/2020/8183713
65. Wang Q, Zhu X, Li Z, Feng M, Liu X. ATF6 promotes liver fibrogenesis by regulating macrophage-derived interleukin-1α expression. Cell Immunol. 2021;367:104401. doi:10.1016/j.cellimm.2021.104401
66. Yuan S, Liu J, Yang L, Zhang X, Zhuang K, He S. Knockdown of circ_0044226 promotes endoplasmic reticulum stress-mediated autophagy and apoptosis in hepatic stellate cells via miR-4677-3p/SEC61G axis. J Bioenerg Biomembr. 2024;56(3):261–271. doi:10.1007/s10863-024-10007-0
67. Zuo R, Wang M, Wang YT, et al. Ablation of hepatic Asah1 gene disrupts hepatic lipid homeostasis and promotes fibrotic nonalcoholic steatohepatitis in mice. Am J Pathol. 2025;195(3):542–560. doi:10.1016/j.ajpath.2024.11.003
68. Wang W, Yao LJ, Shen W, et al. FOXA2 alleviates CCl(4)-induced liver fibrosis by protecting hepatocytes in mice. Sci Rep. 2017;7(1):15532. doi:10.1038/s41598-017-15831-6
69. Liu J, Li M, Gong J, et al. Knockdown of histidine-rich calcium-binding protein (HRC) suppresses liver fibrosis by inhibiting the activation of hepatic stellate cells. Biol Open. 2017;6(1):29–34. doi:10.1242/bio.019828
70. Borkham-Kamphorst E, Steffen BT, Van de Leur E, et al. CCN1/CYR61 overexpression in hepatic stellate cells induces ER stress-related apoptosis. Cell Signal. 2016;28(1):34–42. doi:10.1016/j.cellsig.2015.10.013
71. Hur S, Jeong H, Kim K, et al. MIST1 regulates endoplasmic reticulum stress-induced hepatic apoptosis as a candidate marker of fatty liver disease progression. Cell Death Dis. 2024;15(11):805. doi:10.1038/s41419-024-07217-0
72. Han D, Kim H, Kim S, et al. Sestrin2 protects against cholestatic liver injury by inhibiting endoplasmic reticulum stress and NLRP3 inflammasome-mediated pyroptosis. Exp Mol Med. 2022;54(3):239–251. doi:10.1038/s12276-022-00737-9
73. Fu N, Li D, Li W, et al. Glutamate-cysteine ligase catalytic subunit attenuated hepatitis C virus-related liver fibrosis and suppressed endoplasmic reticulum stress. Front Mol Biosci. 2020;7:199. doi:10.3389/fmolb.2020.00199
74. Wang T, Li X, Xia G, Gong M, Lv X. FAM134B-mediated ER-phagy alleviates alcohol-related liver fibrosis by reducing endoplasmic reticulum stress. Int J Biol Macromol. 2025;308(Pt 2):142395. doi:10.1016/j.ijbiomac.2025.142395
75. Xu H, Li Y, Guo N, et al. Caveolin-1 mitigates the advancement of metabolic dysfunction-associated steatotic liver disease by reducing endoplasmic reticulum stress and pyroptosis through the restoration of cholesterol homeostasis. Int J Biol Sci. 2025;21(2):490–506. doi:10.7150/ijbs.100794
76. Anand SK, Caputo M, Xia Y, et al. Inhibition of MAP4K4 signaling initiates metabolic reprogramming to protect hepatocytes from lipotoxic damage. J Lipid Res. 2022;63(7):100238. doi:10.1016/j.jlr.2022.100238
77. Platko K, Lebeau PF, Byun JH, et al. GDF10 blocks hepatic PPARγ activation to protect against diet-induced liver injury. Mol Metab. 2019;27:62–74. doi:10.1016/j.molmet.2019.06.021
78. Huang YH, Yang YL, Huang FC, et al. MicroRNA-29a mitigation of endoplasmic reticulum and autophagy aberrance counteracts in obstructive jaundice-induced fibrosis in mice. Exp Biol Med. 2018;243(1):13–21. doi:10.1177/1535370217741500
79. Ali O, Darwish HA, Eldeib KM, Abdel Azim SA. Abdel Azim SA: miR-26a potentially contributes to the regulation of fatty acid and sterol metabolism in vitro human HepG2 cell model of nonalcoholic fatty liver disease. Oxid Med Cell Longev. 2018;2018:8515343. doi:10.1155/2018/8515343
80. Petersen PS, Lei X, Wolf RM, et al. CTRP7 deletion attenuates obesity-linked glucose intolerance, adipose tissue inflammation, and hepatic stress. Am J Physiol Endocrinol Metab. 2017;312(4):E309–e325. doi:10.1152/ajpendo.00344.2016
81. Najt CP, Senthivinayagam S, Aljazi MB, et al. Liver-specific loss of Perilipin 2 alleviates diet-induced hepatic steatosis, inflammation, and fibrosis. Am J Physiol Gastrointest Liver Physiol. 2016;310(9):G726–738. doi:10.1152/ajpgi.00436.2015
82. Zhao S, Fang W, Wei Y, Meng J, Jin Q, Fang W. Role of AMPKα2 in regulating the IRE1α-JNK pathway in metabolic dysfunction-associated fatty liver disease. Zhong Nan Da Xue Xue Bao Yi Xue Ban. 2024;49(12):1891–1901. doi:10.11817/j.issn.1672-7347.2024.240474
83. Ming Y, Zhu X, Tuma-Kellner S, et al. iPla2β deficiency suppresses hepatic ER UPR, Fxr, and phospholipids in mice fed with MCD diet, resulting in exacerbated hepatic bile acids and biliary cell proliferation. Cells. 2019;8(8):879. doi:10.3390/cells8080879
84. Lebeaupin C, Vallée D, Rousseau D, et al. Bax inhibitor-1 protects from nonalcoholic steatohepatitis by limiting inositol-requiring enzyme 1 alpha signaling in mice. Hepatology. 2018;68(2):515–532. doi:10.1002/hep.29847
85. Ma Z, Sheng L, Li J, et al. Resveratrol alleviates hepatic fibrosis in associated with decreased endoplasmic reticulum stress-mediated apoptosis and inflammation. Inflammation. 2022;45(2):812–823. doi:10.1007/s10753-021-01586-w
86. Li Y, Li C, Xiong Y, Fang B, Lin X, Huang Q. Didymin ameliorates liver fibrosis by alleviating endoplasmic reticulum stress and glycerophospholipid metabolism: based on transcriptomics and metabolomics. Drug Des Devel Ther. 2022;16:1713–1729. doi:10.2147/DDDT.S351092
87. Xuan Y, Chen S, Ding X, et al. Tetrahydropalmatine attenuates liver fibrosis by suppressing endoplasmic reticulum stress in hepatic stellate cells. Chin Med J. 2021;135(5):628–630. doi:10.1097/CM9.0000000000001883
88. Ustuner D, Kolac UK, Ustuner MC, et al. Naringenin ameliorate carbon tetrachloride-induced hepatic damage through inhibition of endoplasmic reticulum stress and autophagy in rats. J Med Food. 2020;23(11):1192–1200. doi:10.1089/jmf.2019.0265
89. Zhang Z, Li B, Meng X, et al. Berberine prevents progression from hepatic steatosis to steatohepatitis and fibrosis by reducing endoplasmic reticulum stress. Sci Rep. 2016;6:20848. doi:10.1038/srep20848
90. Wang H, Han B, Wang N, et al. Oxymatrine attenuates arsenic-induced endoplasmic reticulum stress and calcium dyshomeostasis in hepatic stellate cells. Ann Transl Med. 2020;8(18):1171. doi:10.21037/atm-20-5881
91. Truong TMT, Seo SH, Chung S, Kang I. Attenuation of hepatic fibrosis by p-Coumaric acid via modulation of NLRP3 inflammasome activation in C57BL/6 mice. J Nutr Biochem. 2023;112:109204. doi:10.1016/j.jnutbio.2022.109204
92. Liao X, Zhan W, Li R, Tian T, Yu L, Yang Q. Irisin ameliorates endoplasmic reticulum stress and liver fibrosis through inhibiting PERK-mediated destabilization of HNRNPA1 in hepatic stellate cells. Biol Chem. 2021;402(6):703–715. doi:10.1515/hsz-2020-0251
93. Song X, Sun J, Liu H, et al. Lycopene alleviates endoplasmic reticulum stress in steatohepatitis through inhibition of the ASK1-JNK signaling pathway. J Agric Food Chem. 2024;72(14):7832–7844. doi:10.1021/acs.jafc.3c08108
94. Pi D, Liang Z, Pan J, et al. Tanshinone IIA inhibits the endoplasmic reticulum stress-induced unfolded protein response by activating the PPARα/FGF21 axis to ameliorate nonalcoholic steatohepatitis. Antioxidants. 2024;13(9). doi:10.3390/antiox13091026
95. Zhu J, Wang R, Xu T, et al. Salvianolic acid A attenuates endoplasmic reticulum stress and protects against cholestasis-induced liver fibrosis via the SIRT1/HSF1 pathway. Front Pharmacol. 2018;9:1277. doi:10.3389/fphar.2018.01277
96. Hou XX, Li YW, Song JL, et al. Cryptotanshinone induces apoptosis of activated hepatic stellate cells via modulating endoplasmic reticulum stress. World J Gastroenterol. 2023;29(17):2616–2627. doi:10.3748/wjg.v29.i17.2616
97. Sun S, Huan S, Li Z, et al. Curcumol alleviates liver fibrosis by inducing endoplasmic reticulum stress-mediated necroptosis of hepatic stellate cells through Sirt1/NICD pathway. PeerJ. 2022;10:e13376. doi:10.7717/peerj.13376
98. He L, Hou X, Fan F, Wu H. Quercetin stimulates mitochondrial apoptosis dependent on activation of endoplasmic reticulum stress in hepatic stellate cells. Pharm Biol. 2016;54(12):3237–3243. doi:10.1080/13880209.2016.1223143
99. Ma Y, Li Y, Zhang H, Wang Y, Wu C, Huang W. Malvidin induces hepatic stellate cell apoptosis via the endoplasmic reticulum stress pathway and mitochondrial pathway. Food Sci Nutr. 2020;8(9):5095–5106. doi:10.1002/fsn3.1810
100. Wang C, Zhang F, Cao Y, et al. Etoposide induces apoptosis in activated human hepatic stellate cells via ER stress. Sci Rep. 2016;6:34330. doi:10.1038/srep34330
101. Gan L, Jiang Q, Huang D, et al. A natural small molecule alleviates liver fibrosis by targeting apolipoprotein L2. Nat Chem Biol. 2025;21(1):80–90. doi:10.1038/s41589-024-01704-3
102. Li YJ, Guo MY, Qin WQ, et al. Senkyunolide A ameliorates cholestatic liver fibrosis by controlling CLCC1-mediated endoplasmic reticulum Ca(2+) release. Acta Pharmacol Sin. 2025:1–6.
103. Xia M, Li J, Martinez Aguilar LM, et al. Arctigenin attenuates hepatic stellate cell activation via Endoplasmic Reticulum-Associated Degradation (ERAD)-mediated restoration of lipid homeostasis. J Agric Food Chem. 2025;73(22):13918–13933. doi:10.1021/acs.jafc.5c01366
104. Yang AY, Kim K, Kwon HH, Leem J, Song JE. 6-shogaol ameliorates liver inflammation and fibrosis in mice on a methionine- and choline-deficient diet by inhibiting oxidative stress, cell death, and endoplasmic reticulum stress. Molecules. 2024;29(2). doi:10.3390/molecules29020419
105. Abdelfattah AM, Mahmoud SS, El-Wafaey DI, Abdelgeleel HM, Abdelhamid AM. Diacerein ameliorates cholestasis-induced liver fibrosis in rat via modulating HMGB1/RAGE/NF-κB/JNK pathway and endoplasmic reticulum stress. Sci Rep. 2023;13(1):11455. doi:10.1038/s41598-023-38375-4
106. Sun Z, Guo Y, Xu X, et al. Hydronidone induces apoptosis in activated hepatic stellate cells through endoplasmic reticulum stress-associated mitochondrial apoptotic pathway. J Gastroenterol Hepatol. 2024;39(8):1695–1703. doi:10.1111/jgh.16635
107. Bian M, He J, Jin H, et al. Oroxylin A induces apoptosis of activated hepatic stellate cells through endoplasmic reticulum stress. Apoptosis. 2019;24(11–12):905–920. doi:10.1007/s10495-019-01568-2
108. Park S, Jeong I, Kim OK. Ginsenoside Rh2 mitigates endoplasmic reticulum stress-induced apoptosis and inflammation and through inhibition of hepatocyte-macrophage inflammatory crosstalk. Nutrients. 2025;17(10):1682. doi:10.3390/nu17101682
109. Zhang H, Chen Q, Dahan A, et al. Transcriptomic analyses reveal the molecular mechanisms of schisandrin B alleviates CCl(4)-induced liver fibrosis in rats by RNA-sequencing. Chem Biol Interact. 2019;309:108675. doi:10.1016/j.cbi.2019.05.041
110. Komeili Movahhed T, Moslehi A, Golchoob M, Ababzadeh S. Allantoin improves methionine-choline deficient diet-induced nonalcoholic steatohepatitis in mice through involvement in endoplasmic reticulum stress and hepatocytes apoptosis-related genes expressions. Iran J Basic Med Sci. 2019;22(7):736–744. doi:10.22038/ijbms.2019.33553.8012
111. Yuan D, Xiang T, Huo Y, et al. Preventive effects of total saponins of Panax japonicus on fatty liver fibrosis in mice. Arch Med Sci. 2018;14(2):396–406. doi:10.5114/aoms.2016.63260
112. Fang R, Wang X, Zhang H, et al. α-tocopherol ameliorates liver fibrosis by inhibiting hepatic stellate cell activation by promoting nrf2 nuclear translocation. Antioxid Redox Signal. 2025;43(16–18):833–848. doi:10.1177/15230864251364900
113. Zhu J, Ding J, Li S, Jin J. Ganoderic acid A ameliorates non-alcoholic streatohepatitis (NASH) induced by high-fat high-cholesterol diet in mice. Exp Ther Med. 2022;23(4):308. doi:10.3892/etm.2022.11237
114. Cao R, Cao C, Hu X, et al. Kaempferol attenuates carbon tetrachloride (CCl(4))-induced hepatic fibrosis by promoting ASIC1a degradation and suppression of the ASIC1a-mediated ERS. Phytomedicine. 2023;121:155125. doi:10.1016/j.phymed.2023.155125
115. Pan PH, Wang YY, Lin SY, et al. 18β-glycyrrhetinic acid protects against cholestatic liver injury in bile duct-ligated rats. Antioxidants. 2022;11(5). doi:10.3390/antiox11050961
116. Xu JB, Gao GC, Yuan MJ, et al. Lignans from Schisandra chinensis ameliorate alcohol and CCl(4)-induced long-term liver injury and reduce hepatocellular degeneration via blocking ETBR. J Ethnopharmacol. 2020;258:112813. doi:10.1016/j.jep.2020.112813
117. Senoo T, Sasaki R, Akazawa Y, et al. Geranylgeranylacetone attenuates fibrogenic activity and induces apoptosis in cultured human hepatic stellate cells and reduces liver fibrosis in carbon tetrachloride-treated mice. BMC Gastroenterol. 2018;18(1):34. doi:10.1186/s12876-018-0761-7
118. Zhu Y, Ouyang Z, Du H, et al. New opportunities and challenges of natural products research: when target identification meets single-cell multiomics. Acta Pharmaceutica Sinica B. 2022;12(11):4011–4039. doi:10.1016/j.apsb.2022.08.022
119. Bai Y, Chen J, Zhang S, et al. Inflammation-responsive cell membrane-camouflaged nanoparticles against liver fibrosis via regulating endoplasmic reticulum stress and oxidative stress. Adv Mater. 2024;36(19):e2310443. doi:10.1002/adma.202310443
120. Huang L, Huang X-H, Yang X, et al. Novel nano-drug delivery system for natural products and their application. Pharmacol Res. 2024;201:107100. doi:10.1016/j.phrs.2024.107100
121. Ramírez-Cortés F, Ménová P. Hepatocyte targeting via the asialoglycoprotein receptor. RSC Med Chem. 2025;16(2):525–544. doi:10.1039/D4MD00652F
122. Huang J, Huang H, Wang Y, et al. Retinol-binding protein-hijacking nanopolyplex delivering siRNA to cytoplasm of hepatic stellate cell for liver fibrosis alleviation. Biomaterials. 2023;299:122134. doi:10.1016/j.biomaterials.2023.122134
123. Ezhilararasan D, Lakshmi T, Raut B. Novel nano-based drug delivery systems targeting hepatic stellate cells in the fibrotic liver. J Nanomater. 2021;2021(1):4674046. doi:10.1155/2021/4674046
124. Li Y, Pu S, Liu Q, et al. An integrin-based nanoparticle that targets activated hepatic stellate cells and alleviates liver fibrosis. J Control Release. 2019;303:77–90. doi:10.1016/j.jconrel.2019.04.022
125. Vijayan VM, Unagolla J, Panchal D, JE J, SS M, JU M. Biomimetic nanoparticles for targeted therapy of liver disease. RSC Pharmaceutics. 2025;2(4):667–682. doi:10.1039/D5PM00044K
126. Du Y, Ding H, Chen Y, Gao B, Mao Z, Wang W. Ding Y: a Genetically Engineered Biomimetic Nanodecoy for the Treatment of Liver Fibrosis. Adv Sci. 2024;11(40):2405026. doi:10.1002/advs.202405026
127. Sun L, Luo X, Zhou C, Zhou Z, Sun M. Natural polysaccharide-based smart CXCR4-targeted nano-system for magnified liver fibrosis therapy. Chin Chem Lett. 2024;35(2):108803. doi:10.1016/j.cclet.2023.108803
128. Sun H, Liu T, Wang Z, et al. Role of curcumin in chronic liver diseases: a comprehensive review. Drug Des Devel Ther. 2025;19:3395–3406. doi:10.2147/DDDT.S518547
129. Frankowski J, Kurzątkowska M, Sobczak M, Piotrowska U. Utilization of 3D bioprinting technology in creating human tissue and organoid models for preclinical drug research – state-of-the-art. Int J Pharm. 2023;644:123313. doi:10.1016/j.ijpharm.2023.123313
130. Abbasian MH, Sobhani N, Sisakht MM, D’Angelo A, Sirico M, Roudi R. Patient-derived organoids: a game-changer in personalized cancer medicine. Stem Cell Rev Rep. 2025;21(1):211–225. doi:10.1007/s12015-024-10805-4
131. Reich S, Nguyen CDL, Has C, et al. A multi-omics analysis reveals the unfolded protein response regulon and stress-induced resistance to folate-based antimetabolites. Nat Commun. 2020;11(1):2936. doi:10.1038/s41467-020-16747-y
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
The Role of Adipocyte Endoplasmic Reticulum Stress in Obese Adipose Tissue Dysfunction: A Review
Xu S, Xi J, Wu T, Wang Z
International Journal of General Medicine 2023, 16:4405-4418
Published Date: 27 September 2023
Nanomedicines in the Treatment of Liver Fibrosis: A Review
Du X, Niu R, Liu X, Wu F, Yang X, Ma X, Zhang J, Zhou H, Shao L, Wang S
International Journal of Nanomedicine 2025, 20:9641-9665
Published Date: 5 August 2025
Endoplasmic Reticulum Stress as a Stage-Dependent Regulatory Hub in Rheumatoid Arthritis
Hu N, Song Y, Yu T, Li H, Wang X, Lv Q
Journal of Inflammation Research 2026, 19:606604
Published Date: 28 May 2026