Back to Journals » Breast Cancer: Targets and Therapy » Volume 18
CYP4B1 Expression in Breast Cancer: An Immunohistochemical Study with Complementary in silico Analyses
Authors Al-saraireh YM ![]() , Alshammari FO, Hassan HA, Jaradat JH, Al-abadleh AA, Satari AO, Al-sarayreh SA, Al-dalain SM, Al-Qtaitat A, Al Shuneigat JM
, Alshammari FO, Hassan HA, Jaradat JH, Al-abadleh AA, Satari AO, Al-sarayreh SA, Al-dalain SM, Al-Qtaitat A, Al Shuneigat JM ![]() , Hareedy MS
, Hareedy MS
Received 28 March 2026
Accepted for publication 26 June 2026
Published 7 July 2026 Volume 2026:18 612743
DOI https://doi.org/10.2147/BCTT.S612743
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Harikrishna Nakshatri
Yousef M Al-saraireh1, Fatemah OFO Alshammari2, Heba A Hassan1,3, Jaber H Jaradat1, Ahmed A Al-abadleh1,4, Anas O Satari1,5, Sameeh A Al-sarayreh6, Sa’ed M Al-dalain1, Aiman Al-Qtaitat7, Jehad M Al Shuneigat6, Mohammad Salem Hareedy1
1Department of Pharmacology, Faculty of Medicine, University of Mutah, Al-Karak, Jordan; 2Department of Medical Laboratory Technology, Faculty of Health Sciences, The Public Authority for Applied Education and Training, Shuwaikh, Kuwait; 3Department of Clinical Pharmacology, Faculty of Medicine, Zagazig University, Zagazig, Egypt; 4Department of Urology, Faculty of Medicine, MUST University, Cairo, Egypt; 5Department of General Pediatrics, Maternity and Children’s Hospital at Al Bashir Hospital, Amman, Jordan; 6Department of Biochemistry and Molecular Biology, Faculty of Medicine, University of Mutah, Al-Karak, Jordan; 7Department of Anatomy and Histology, Faculty of Medicine, University of Mutah, Al-Karak, Jordan
Correspondence: Yousef M Al-saraireh, Email [email protected]
Purpose: Cytochrome 4B1 (CYP4B1) is a monooxygenase extrahepatic enzyme, recently found expressed in several malignancies. However, the role of CYP4B1 in breast cancer is still undetermined.
Patients and Methods: The study consisted of two complementary components. First, CYP4B1 protein expression was evaluated by immunohistochemistry using a breast cancer tissue microarray (n = 207 tumors; n = 8 normal breast tissues). Second, in silico analyses were performed using publicly available databases and included transcriptomic expression profiling, survival analysis, protein–protein interaction analysis (PPI), functional enrichment analysis, drug-sensitivity correlations, and AlphaFold-based structural assessment.
Results: Immunohistochemical analysis demonstrated focal and heterogeneous CYP4B1 protein expression, which was detected in 18.3% of breast tumors and was significantly associated with lower histological grade. No CYP4B1 immunoreactivity was observed in the sampled normal breast tissues under the experimental conditions. In silico analyses demonstrated reduced CYP4B1 mRNA expression in breast cancer at the population level, compared with normal breast tissue. Survival analysis showed no statistically significant differences between CYP4B1 expression groups. According to PPI analysis, CYP4B1 is part of a closely related redox-regulatory and xenobiotic-metabolizing module enriched in CYPs, GSTs, and SULTs. Structural analysis provided contextual support for the previously reported association of the P427S meander-region substitution with reduced catalytic competence. Weak exploratory correlations were observed between CYP4B1 expression and drug sensitivity metrics; however, these findings were descriptive and did not indicate clinically meaningful resistance.
Conclusion: Immunohistochemical analysis demonstrated that CYP4B1 protein expression is focal and heterogeneous, being retained in only a small subset of breast tumors and associated with lower histological grade. Bioinformatics analyses showed reduced CYP4B1 mRNA expression in breast cancer and identified CYP4B1 within xenobiotic metabolism and redox-regulatory networks. Collectively, these findings provide a descriptive multilevel characterization of CYP4B1 in breast cancer; however, its prognostic and therapeutic significance remains exploratory and requires further functional validation.
Keywords: breast cancer, Cytochrome 4B1, immunohistochemistry, structural modeling, proteomics, transcriptomics
Introduction
Breast cancer is a significant global health issue. Approximately 316,950 new cases of invasive breast cancer in females are anticipated in the United States alone in 2025, along with approximately 42,170 fatalities from this type of cancer.1,2 Worldwide, it is estimated that there will be up to 3.2 million new cases of breast cancer and 1.1 million annual deaths by 2050.3 Breast cancer remains the most prevalent cancer among females even though the huge steps made in early diagnostic and therapeutic methods.2,3 Therefore, there is still a need for further diagnostic biomarkers and therapeutic research to advance and enhance patients’ survival and quality of life.
The cytochrome P450 superfamily consists of a vast set of monooxygenase enzymes that catalyze various enzymatic reactions, One important subfamily is CYP4 (eg, CYP4A, CYP4F, and CYP4B), which mediates the ω-hydroxylation of fatty acids and eicosanoids, thereby affecting lipid signaling, vascular tone, and inflammation.4–7 Aberrant regulation of these enzymes is associated with several pathological conditions, such as carcinogenesis, cardiovascular diseases, and inflammation.8 Therefore, the CYP4 family offers a potential biological link between oxidative stress, cellular metabolism reprogramming and cancer biology.
Among the CYP4 members CYP4B1, which has recently gained attention in cancer research. Despite its low constitutive activity in the human enzyme, CYP4B1 is expressed in extrahepatic tissues (such as the bladder, lung, and adrenal glands) and has been shown to be capable of xenobiotic activation and ω-hydroxylation.9 Many reports have linked aberrant expression or polymorphism of CYP4B1 with cancers of the colon, lung, and urothelium, implying its potential role in tumor development or response to therapy.10 For example, CYP4B1 genetic polymorphisms were associated with breast cancer susceptibility and tumor stage, suggesting a potential contribution of CYP4B1 to breast tumor biology.11 Additionally, CYP4B1 was found to be a potential therapeutic target in cancer due to its ability to bioactivate specific prodrugs and modulate xenobiotic metabolism.10 More recently, Calaf et al analyzed CYP4 family enzyme expression in breast cancer tissues and reported altered expression patterns including CYP4B1.8 While these studies suggest potential relevance of CYP4B1 in breast cancer, the available evidence remains limited and fragmented. Existing knowledge is mostly confined to expression profiling and studies of genetic association, but the biological relevance of CYP4B1 expression inside breast cancers remains unclear. Specifically, it’s unknown if CYP4B1 expression is linked to particular clinicopathological characteristics, reflects unique metabolic states of breast cancers, or plays a key part in molecular mechanisms important to the development of breast cancer.8,11 Consequently, comprehensive profiling of protein expression with multilevel bioinformatics analyses including transcriptomic profiling, structural modeling, and systems-level network analysis remains lacking.
Breast cancer is a physiologically diverse disease with several molecular subtypes that differ significantly in terms of hormone receptor status, metabolic reprogramming, redox homeostasis and responsiveness to treatment. As CYP enzymes have well-known roles in metabolism of xenobiotics and lipids, and regulation of oxidative stress, CYP4B1 expression may differ amongst physiologically distinct breast cancer subgroups. Such variability may contribute to the diverse expression patterns seen in earlier investigations, highlighting the significance of assessing CYP4B1 particularly within the context of breast cancer rather than extrapolating results from other cancers.7,8
Despite emerging evidence linking CYP4B1 dysregulation to several malignancies and limited observations suggesting potential involvement in breast cancer, its biological role in breast cancer remains largely undefined. In particular, it is unclear whether CYP4B1 expression is associated with specific clinicopathological characteristics or reflects distinct biological states within breast tumors. To address these knowledge gaps, the present study first investigated CYP4B1 protein expression in breast cancer using immunohistochemistry on a tissue microarray cohort, with the aim of characterizing its expression pattern and evaluating its association with clinicopathological features. In parallel, a complementary bioinformatics framework was employed to examine CYP4B1 at the molecular and systems levels. Publicly available transcriptomic datasets were used to assess gene expression and survival associations, while protein–protein interaction network analysis, functional enrichment analysis, drug-sensitivity correlations, and AlphaFold-based structural assessment were performed to explore the biological context of CYP4B1 in breast cancer. Unlike previous studies that focused primarily on genetic associations or broad CYP family expression patterns, the present work integrates laboratory-based and computational approaches to provide a multilevel characterization of CYP4B1. Importantly, the objective of this study was exploratory and descriptive rather than to establish CYP4B1 as a prognostic biomarker or therapeutic target. Together, these complementary experimental and bioinformatics analyses provide a comprehensive framework for understanding CYP4B1 expression patterns, molecular associations, and potential biological relevance in breast cancer.
Materials and Methods
Experimental Analysis
Study Cohort and Tissue Microarray Characteristics
The Institutional Review and Ethics Committee of the University of Mutah’s Faculty of Medicine approved the study, which was conducted in compliance with the Declaration of Helsinki (Reference No. EC6-2326, date: March 2, 2026). Patient consent was waived due to an exemption issued by the Institutional Review and Ethics Committee of the Faculty of Medicine, University of Mutah (Reference No. EC6-2326, date: March 2, 2026), because de-identified, commercially purchased TMAs were used in the study.
A fully annotated and commercially accessible tissue microarray (TMAs) comprising cylindrical cores measuring 1.0 mm in diameter, with 216 cores obtained from each case according to the manufacturer’s design (BR2161, TissueArray.com LLC). This microarray consisted of 207 female breast cancer samples, one male breast cancer sample, and eight normal breast tissue samples which served as qualitative reference controls to establish baseline CYP4B1 expression, rather than as a statistically powered comparison group. The one male breast cancer sample was excluded from the study. Where present, adjacent non-neoplastic breast epithelium within tumor cores was also evaluated for CYP4B1 staining. All patient data, including age, sex, tumor size, histological grade, and stage, were provided by the manufacturer.
The mean age was 48.8 years with a standard deviation of 9.9 years for the 207 breast cancer cases included in the tissue microarray cohort. Furthermore, eight normal breast tissue samples were included as qualitative reference controls for immunohistochemical evaluation (Table 1). Of the total, 60.9% were aged 50 years or younger (n = 126), and 39.1% were older than 50 years (n = 81). The most common histological type was invasive ductal carcinoma, which accounted for 86.6% of cases (n = 188), followed by medullary carcinoma (2.8%), invasive lobular carcinoma (1.8%), and other rare types (4.1%). 136 patients (65.7%) had a tumor size of T2, while the remaining cases were T1 (6.3%), T3 (15%), and T4 (13%). Moreover, 22.7% of the patients presented with lymph node metastasis, whereas no cases were with distant metastasis. Additionally, most patients had grade II (70.1%), while the remaining patients were equally distributed between grades I and III (15%). According to the clinical stage, 6.3% were at stage I, 78.7% at stage II, 14.0% at stage III, and 1.0% at stage IV stages. These clinicopathologic variables were examined to characterize the study cohort and to provide a clinical framework for interpreting CYP4B1 expression patterns. In particular, stratification by tumor grade, stage, and histological subtype enables assessment of whether CYP4B1 expression is associated with tumor differentiation, disease progression, and biologically distinct tumor subsets that inform subsequent molecular and systems-level analyses. These clinicopathologic stratifications are further integrated with molecular findings in Section 2.2, where CYP4B1 protein expression is evaluated to determine whether its distribution corresponds with tumor differentiation and clinicopathologic heterogeneity.
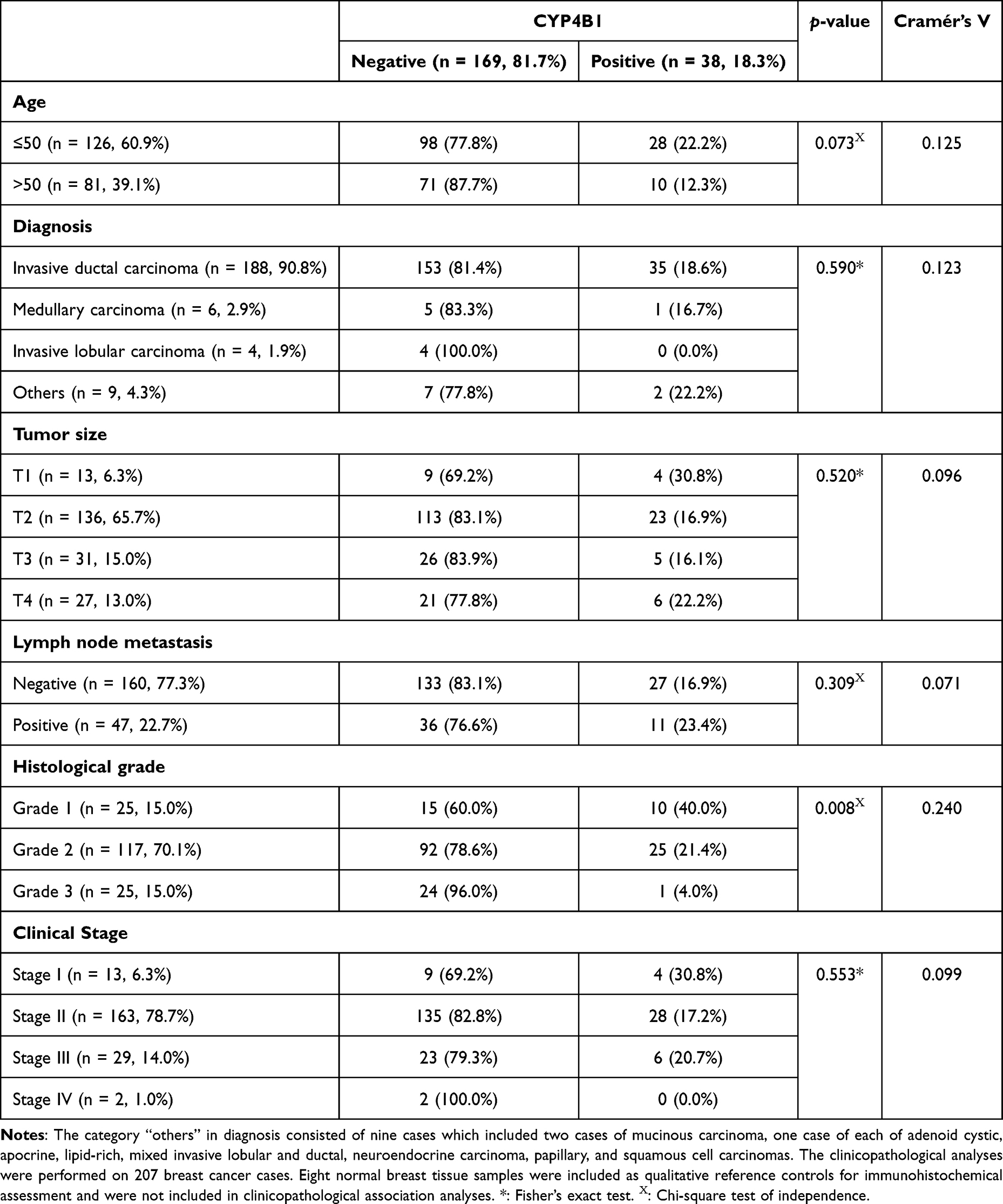 |
Table 1 Clinical and Sociodemographic Features of Breast Cancer Patients |
Immunohistochemistry and Scoring
The TMAs were dewaxed and rehydrated using alcohol-xylene solutions. The tissues were then treated with 5% hydrogen peroxide solution for five minutes to inhibit endogenous peroxidase activity. After that, the heat-induced epitope retrieval (HIER) method was used by microwaving (650 W) the TMAs in citrate buffer (pH 6.0) for 20 min. Subsequently, to inhibit non-specific binding, 5% normal goat serum was added to the tissues for 20 min. Microarray tissue sections were then incubated at room temperature for one hour with CYP4B1 primary antibody (NBP1-86142, Novus Biologicals, USA) at a concentration of 5 μg/mL. To check primary antibody specificity, several experimental controls were included in the study. Antibody specificity was validated by appropriate positive (colon cancer tissue) and negative controls and was further confirmed in breast tissue by demonstrating reproducible cytoplasmic staining in CYP4B1-positive breast tumor tissue and absence of staining in control sections incubated without primary antibody or with neutralized antibody with the corresponding CYP4B1 blocking peptide (NBP1-86142PEP, Novus Biologicals, USA). After several washes, the tissues were incubated with a peroxidase polymer for 30 min (MP-7451, Vector Laboratories, Burlingame, CA, USA). Following several washes, tissues were incubated with 3,3′-diaminobenzidine (DAB) substrate for 5 min at room temperature to develop immunoreactivity. Subsequently, the tissue microarrays were stained with hematoxylin, dehydrated, and mounted using a permanent mounting medium.
The Allred scoring system was used to semi-quantitatively analyze and score the staining of CYP4B1.4,12–16 This system is composed of a score that results in the sum of two elements: the first was the proportion of cells showing positive staining, which ranged from 0 to 5 (0 = none; 1 = <1%; 2 = 1–10%; 3 = 11–33%; 4 = 34–66%; 5 = >66%). The second component is the intensity score, which is given a rating of 0 (no staining), 1 (weak staining), 2 (moderate staining), or 3 (strong staining), determined by the average staining intensity in cancer cells. The total of these two variables yields the final Allred score, which ranges from 0 to 8. A value between 0 and 2 indicates negative biomarker expression, whereas a score between 3 and 8 indicates positive expression. The cutoff of Allred score ≥3 was selected based on established immunohistochemical literature and prior applications in exploratory biomarker studies; however, this threshold has not been specifically validated for CYP4B1 and should be interpreted accordingly. Immunohistochemical scoring was independently performed by two experienced pathologists who were blinded to all clinicopathological data. Inter-rater reliability was formally assessed following independent evaluation using the Intraclass Correlation Coefficient (ICC), which demonstrated good agreement (ICC = 0.842, 95% CI: 0.794–0.879). Bland–Altman analysis further confirmed scoring consistency, with most differences falling within the 95% limits of agreement.
Statistical Analysis
Statistical analyses were performed using IBM SPSS Statistics software (version v26; IBM Corp., Armonk, NY, USA). Descriptive statistics were used to summarize clinicopathological characteristics and CYP4B1 expression patterns. Associations between CYP4B1 protein expression and categorical clinicopathological variables were evaluated using Pearson’s chi-square test or Fisher’s exact test when appropriate. Effect sizes for significant associations were estimated using Cramér’s V. To identify independent clinicopathological predictors of CYP4B1 expression, multivariable binary logistic regression analysis was performed with CYP4B1 positivity as the dependent variable. Odds ratios (ORs) and corresponding 95% confidence intervals (CIs) were calculated. Inter-observer agreement between the two pathologists who independently evaluated immunohistochemical staining was assessed using the Intraclass Correlation Coefficient (ICC). Bland–Altman analysis was additionally performed to evaluate agreement and identify potential systematic scoring differences. For all statistical analyses, p-values < 0.05 were considered statistically significant.
Bioinformatics Analyses
In silico Analysis of CYP4B1 Gene Expression
The expression of CYP4B1 in breast cancer (BRCA) was analyzed using TIMER3 (Tumor Immune Estimation Resource, version 3; https://timer.cistrome.org/), an online platform for the systematic evaluation of tumor-infiltrating immune cells across The Cancer Genome Atlas (TCGA) datasets.17 The Gene DE module was employed to compare CYP4B1 expression between tumor and adjacent normal breast tissues. Raw RNA-seq counts from TCGA were normalized, and differential expression was assessed using the edgeR statistical package. Significance was determined based on p-values.
In silico Survival Analysis of CYP4B1
We GEPIA 2. (Gene Expression Profiling Interactive Analysis, http://gepia.cancer-pku.cn/index.html) web tool was utilized to evaluate the prognostic value of CYP4B1 in patient of breast cancer (accessed on 10 January 2026).18 This web tool is a publicly available database using data for gene expression obtained from Gene Expression Omnibus (GEO), European Genome-Phenome Archive (EGA) and The Cancer Genome Atlas (TCGA). From GEPIA webpage, the analysis for overall and disease free survival for breast cancer patients expressing CYP4B1 were conducted by calculating hazard ratio (HR) with 95% confidence intervals (CI), and log-rank p-values. Here, patients were dichotomized into high and low CYP4B1 expression groups using the platform’s default median expression value as the cut-off. Results were considered significant if p-value was less than 0.05.
In silico Protein–Protein Interaction Network Analysis of CYP4B1
Protein–protein interaction (PPI) network analysis was performed to evaluate the functional connectivity of CYP4B1. Network construction and topological analysis were performed using the STRING database (version 12.0; https://string-db.org) with a confidence interaction score threshold of 0.4 (medium confidence) with a maximum of 10 interactor. This threshold was selected to balance network coverage and biological reliability, allowing identification of coordinated metabolic interactions while minimizing inclusion of low-confidence associations, which is appropriate for exploratory systems-level analysis.
In silico Functional Enrichment Analysis
Functional enrichment analysis was subsequently conducted using STRING database to identify over-represented Kyoto Encyclopedia of Genes and Genomes (KEGG) pathways among the identified PPI networks. Enrichment significance was assessed using Benjamini–Hochberg false discovery rate (FDR) correction, and pathways with FDR < 0.05 were considered statistically significant.
In silico Drug Sensitivity Analysis of CYP4B1 Expression
We employed a drug sensitivity analysis using the Gene Set Cancer Analysis platform (GSCA) to explore the correlation between CYP4B1 expression and drug sensitivity in human cancer cell lines.19,20 Drug sensitivity data were obtained from the Genomics of Drug Sensitivity in Cancer (GDSC), which includes the half-maximal inhibitory concentration (IC50) values of 265 small molecules tested in 860 cancer cell lines. The corresponding mRNA expression profiles were integrated with the IC50 values. Pearson’s correlation analysis was used to evaluate the association between CYP4B1 mRNA expression and the drug IC50 values. We used p-values adjusted for false discovery rate (FDR).
In silico Structural Analysis of CYP4B1 Using AlphaFold
The three-dimensional structure of human CYP4B1 was examined using the publicly available AlphaFold structural model retrieved from the AlphaFold Protein Structure Database (UniProt entry: P13584).21,22 No de novo structural prediction or independent AlphaFold modeling was performed in this study. Instead, the deposited AlphaFold model was analyzed descriptively to assess structural organization and residue-level confidence metrics. We inspected the structural accuracy and reliability using the various metrics provided by the model. The metrics were assessed directly through the database as it provides interactive re-sidual level visualization and our assessment focused on just reporting the descriptive overall metrics. One metric is the predicted local distance difference test (pLDDT) which gives a simple code with score >90 that corresponds to very high reliability, 70–90 corresponds to high, 50–70 low, and less than 50 very low. Structural confidence was assessed using AlphaFold-specific metrics (pLDDT and PAE), which are the recommended validation measures for AI-based structure predictions and supersede traditional stereochemical validation approaches. These metrics were used to evaluate residue-level structural reliability and inter-domain consistency.
Results
Results of Experimental Analysis
CYP4B1 Protein Expression in Breast Cancer and Association with Clinicopathologic Data
Immunohistochemical analysis indicated that CYP4B1 protein expression was heterogeneous and focal, being absent in the majority of breast cancer samples (81.7%, 169/207), but retained in a restricted subset of tumors 18.3% (38/207) (Table 1 and Figure 1). The distribution of Allred scores was as follows: 0 (46.5%), 1 (26.3%), 2 (9.7%), 3 (9.7%), 4 (6.0%), and 5 (1.8%), confirming that most tumors exhibited absent or low CYP4B1 staining. Conversely, no CYP4B1 protein immunoreactivity was observed in the sampled normal breast tissues under the experimental conditions, which served as qualitative reference controls rather than a statistically powered comparator group. Moreover, adjacent non-neoplastic epithelium, when present, showed no detectable CYP4B1 staining, consistent with its use as a qualitative reference rather than a quantitative comparator. CYP4B1 protein expression was mainly localized in cell cytoplasm. Such expression was validated by proper experimental controls where positive control (colon cancer tissue) exhibited CYP4B1 immunoreactivity. Furthermore, negative control tissues and both colon and breast cancer tissues treated with neutralized antibody exhibited weak to no immunoreactivity (Figure 2).
 |
Figure 1 CYP4B1 expression in normal versus different types of breast cancer. (A) Normal breast tissue displayed no CYP4B1 expression. CYP4B1 expression was detected in invasive ductal carcinoma (B), medullary carcinoma (C), papillary (D), and squamous cell carcinomas (E). There was no CYP4B1 expression in other types of breast cancer including invasive lobular (F), mucinous (G), apocrine (H), adenoid cystic (I), neuroendocrine (J) and lipid-rich carcinomas (K). Arrows indicate representative CYP4B1-positive cells. All images were captured at ×400 magnification, Scale bar = 50 μm. |
 |
Figure 2 CYP4B1 expression in different experimental controls. (A) Breast cancer tissue incubated with normal goat serum (negative control) displayed no immunoreactivity; (B) Breast cancer tissue incubated with CYP4B1 primary antibody developed cytoplasmic immunoreactivity; (C) Breast cancer tissue incubated with neutralized CYP4B1 primary antibody developed minimal CYP4B1 immunoreactivity; (D) Colon cancer tissue incubated with CYP4B1 primary antibody developed strong CYP4B1 immunoreactivity (positive control); and (E) Colon cancer tissue incubated with neutralized CYP4B1 primary antibody developed minimal CYP4B1 immunoreactivity. Arrows indicate representative CYP4B1-positive cells. All images were captured at ×400 magnification, Scale bar = 50 μm. |
Interestingly, CYP4B1 protein expression was only found in invasive ductal carcinoma, medullary carcinoma, papillary and squamous cell carcinomas between CYP4B1 protein expression and histological tumor grade (p = 0.008, Cramér’s V = 0.240). A high expression rate was observed in grade 1 tumors (40%), followed by grade 2 (21.4%), and the lowest in grade 3 tumors (4%), showing a tendency for CYP4B1 expression to decline as the histological grade increased.
Although not statistically significant, several trends were observed in the clinicopathological features. A trend of higher frequency of CYP4B1 expression in people aged ≤50 years was observed, although this was insignificant (22.2% vs 12.3%, p = 0.073). Additionally, CYP4B1 expression was not significantly associated with tumor size, lymph node status, distant metastasis, histological type, or tumor stage. Although CYP4B1 protein expression was detected in a minority of breast cancer cases (18.3%), its non-uniform distribution and significant association with histological grade suggest that CYP4B1 expression characterizes a biologically distinct subset of tumors rather than serving as a ubiquitous feature of breast cancer.
To further evaluate clinicopathologic determinants of CYP4B1 expression, a multivariable logistic regression model was performed using CYP4B1 positivity as the dependent variable (Table 2). Variables entered into the model included age, tumor size, lymph node status, and histological grade. Clinical stage and histologic type were not included due to redundancy and reduced degrees of freedom. The analysis demonstrated that higher histological grade was independently associated with reduced likelihood of CYP4B1 positivity. Compared with grade 1 tumors, grade 3 tumors showed significantly lower odds of CYP4B1 expression (OR = 0.068, 95% CI: 0.008–0.601, p = 0.016). No statistically significant associations were observed for age, tumor size, or lymph node metastasis. Model diagnostics indicated high accuracy in predicting CYP4B1-negative cases (99.2%) but limited sensitivity for CYP4B1-positive cases (2.8%), reflecting the relatively low prevalence of CYP4B1 expression in the cohort. Accordingly, the regression findings should be interpreted cautiously, as the limited number of CYP4B1-positive tumors reduces statistical power and may restrict the ability to detect additional independent associations.
 |
Table 2 Multivariable Logistic Regression Model Was Performed Using CYP4B1 Positivity as the Dependent Variable |
Results of Bioinformatics Analyses
In silico Analysis of CYP4B1 Gene Expression
CYP4B1 mRNA expression differed significantly (p < 0.001) between normal and breast cancer samples in the TCGA-BRCA dataset (Figure 3). CYP4B1 mRNA expression was higher in the normal than in breast cancer tissues. These findings indicate downregulation of CYP4B1 at the transcript level in breast cancer.
 |
Figure 3 CYP4B1 gene expression in breast cancer tissues compared to that in normal breast tissues. ***: p-value <0.001. |
In silico Survival Analysis of CYP4B1
Disease-free survival (DFS) was assessed in 1069 patients with breast cancer, who were classified into high CYP4B1 expression (n = 534) and low CYP4B1 expression (n = 535) groups. Kaplan–Meier analysis demonstrated no statistically significant difference in disease-free survival between high and low CYP4B1 expression groups (HR = 0.73, log-rank p = 0.092). These findings do not support CYP4B1 as a prognostic biomarker for disease-free survival (Figure 4). Another analysis of overall survival (OS) was conducted in a distinct cohort (n = 536), which was evenly divided into high- and low-expression groups. No statistically significant difference in overall survival was detected between CYP4B1 expression groups (HR = 1.50, log-rank p = 0.078) (Figure 4). Because these analyses were derived from publicly available datasets and no statistically significant associations were identified, the results should be considered exploratory and not supportive of a prognostic role for CYP4B1 in breast cancer.
 |
Figure 4 Kaplan-Meier graphs showing overall survival (OS) and disease-free survival (DFS). |
In silico Protein–Protein Interaction (PPI) Network Analysis of CYP4B1
Using a medium-confidence STRING interaction score threshold of 0.4 (selected to balance interaction coverage and biological relevance), the constructed PPI network comprised 11 nodes and 30 edges, with an average node degree of 5.45 and an average local clustering coefficient of 0.833, indicating a highly interconnected network architecture. The nodes represent the proteins, and the edges represent the connections between these nodes, such as the connection between CYP4B1 and AASDH. The observed edges were significantly higher than expected, indicating significant interactions within the network (Figure 5). Unsupervised clustering was performed using k-means with three clusters selected. Cluster 1 included 14 genes, and it included the majority of the genes in the network and consisted mainly of cytochrome P450 enzymes and glutathione transferases that play roles in fatty acid oxidation, xenobiotic metabolism, and steroid hormone processing, emphasizing a coordinated enzymatic network involved in drug detoxification and metabolism pathways. Cluster 2 was composed of a sole protein, a beta-alanine-activating enzyme (AASDH), that seems to be involved in post-translational modification pathways, and amino acids metabolism. Cluster 3 also included only SULT1C2, a sulfotransferase associated with drug and xenobiotic metabolism, including the activation of carcinogenic hydroxylamines. The PPI network showed more than expected connections within the network, which reflects that the proteins are not independent of each other and they interact and influence each other in a way or another and have cooperative roles. Moreover, higher confidence thresholds were also explored but resulted in a sparsely connected network that limited systems-level interpretation, whereas lower thresholds increased network density at the expense of interaction reliability.
 |
Figure 5 Protein–protein interaction (PPI) network of CYP4B1 and associated proteins generated using the STRING database. Nodes represent proteins and edges represent known or predicted interactions. Clustering highlights coordinated metabolic and xenobiotic-processing modules. This network is presented as a descriptive, systems-level analysis to illustrate functional context and connectivity rather than direct mechanistic or causal relationships. The network is color coded, where the red nodes represent cluster 1, green nodes represent cluster 2, and blue nodes represent cluster 3. |
In silico Functional Enrichment Analysis
KEGG pathway enrichment analysis demonstrated significant over-representation of metabolic pathways, including Metabolism of xenobiotics by cytochrome P450, Drug metabolism – cytochrome P450, Arachidonic acid metabolism, Linoleic acid metabolism, Steroid hormone biosynthesis, and Retinol metabolism (FDR < 0.05). These findings indicate that the CYP4B1 interaction network is functionally enriched for lipid and xenobiotic metabolic processes, consistent with the established biological roles of cytochrome P450 enzymes (Figure 6).
 |
Figure 6 The KEGG pathway overrepresentation among the PPI networks for CYP4B1. |
In silico Drug Sensitivity Analysis of CYP4B1 Expression
We investigated drug sensitivity correlations with CYP4B1 using publicly available GDSC datasets; all observed associations were weak, with absolute correlation coefficients remaining below 0.2. Collectively, these weak correlations are hypothesis-generating only and reflect descriptive metabolic associations rather than predictors of drug response or clinically relevant resistance. Figure 7 illustrates the top 30 CYP4B1-drug correlations, where it primarily shows that higher gene expression may be associated with more resistance across several drug families, most notably with kinase inhibitors and chemotherapeutic agents.
 |
Figure 7 Exploratory correlation analysis between CYP4B1 expression and drug sensitivity across cancer cell lines in the GDSC dataset. Bubble plot summarizing the top 30 gene–drug correlations, where each bubble represents the association between CYP4B1 mRNA expression and the half-maximal inhibitory concentration (IC50) of a drug in 860 cancer cell lines. Bubble color indicates correlation direction (red = positive correlation; blue = negative correlation), and bubble size reflects statistical significance after false discovery rate adjustment. These correlations are weak and are presented for hypothesis generation and systems-level context only, without implying therapeutic prediction or clinically relevant drug resistance. Bubble size reflects the significance of the association after false discovery rate adjustment, and bubbles outlined in black indicate statistically significant correlations (FDR ≤ 0.05). |
In silico Structural Analysis of CYP4B1 Using AlphaFold
The AlphaFold-model of CYP4B1 (511 amino acids) showed an overall very high confidence, with an average pLDDT score of 94.04. A residue-level breakdown indicated that 88.3% of the protein was predicted with very high confidence (pLDDT >90), 7.4% with high confidence (70–90), 3.1% with low confidence (50–70), and 1.2% with very low confidence (<50) (Figure 8). The analysis preformed was using the publicly available AlphaFold model without performing new structural prediction or refinement.
 |
Figure 8 Predicted 3D structure of human Cytochrome P450 4B1 (CYP4B1) generated by AlphaFold. |
The PAE plot demonstrated low error values across most regions, indicating strong reliability of residue positioning. Structural confidence was consistent across the heme-binding domain, providing contextual support for annotation of CYP4B1 as a heme-thiolate monooxygenase, without implying functional activity or clinical relevance in breast cancer. Detailed pLDDT and PAE confidence profiles were assessed using the interactive AlphaFold database interface, which provides higher interpretative value than static representations for residue-level structural assessment.
Discussion
Despite its established physiological roles in extrahepatic tissues such as the lung and adrenal cortex, the oncological role of CYP4B1 remains incompletely understood. Some studies have previously suggested a potential relationship between CYP4B1 and breast cancer biology. For instance, Yin et al reported associations between CYP4B1 polymorphisms and breast cancer susceptibility and tumor stage,11 while Calaf et al observed altered expression of CYP4 family enzymes, including CYP4B1, in breast cancer tissues.8 Additionally, Lim et al highlighted CYP4B1 as a candidate therapeutic target in cancer due to its role in xenobiotic metabolism and prodrug activation.10 However, these studies did not comprehensively examine CYP4B1 across multiple biological levels in breast cancer. In the current study, immunohistochemical analysis demonstrated focal and heterogeneous CYP4B1 protein expression, which was detected in only a subset of breast cancers and was significantly associated with lower histological grade. These findings suggest that CYP4B1 expression may characterize a biologically distinct subgroup of breast tumors. Moreover, such findings correspond with earlier observations of CYP4B1 silencing in various cancer types, indicating that its suppression may be associated with cancer dedifferentiation or metabolic reprogramming.9,10,23 However, the present findings should be interpreted within the framework of an exploratory characterization study designed to describe CYP4B1 expression patterns and molecular associations rather than establish prognostic, predictive, or causal roles.
Our immunohistochemical analysis revealed that CYP4B1 protein was expressed in only 18.3% of breast cancer samples, and its expression decreased with increasing tumor grade. Although this restricted expression pattern may reflect biologically distinct associated features, the relatively small number of CYP4B1-positive tumors limits statistical power and warrants cautious interpretation of subgroup analyses. Consistent with this pattern, multivariable logistic regression identified higher histological grade as an independent predictor of reduced CYP4B1 expression. While this limited prevalence indicates that CYP4B1 is not a universal feature of breast cancer, it does not preclude biological relevance. Instead, the focal retention of CYP4B1 protein in a restricted subset of predominantly low-grade tumors suggests that it represents a differentiation-associated metabolic feature rather than a broadly active oncogenic determinant. Similar patterns of CYP4B1 protein downregulation have also been found in lung adenocarcinoma and bladder cancer, supporting the notion that loss of CYP4B1 protein may accompany tumor dedifferentiation and metabolic reprogramming rather than act as a primary oncogenic driver.24–26 Collectively, these findings suggest that post-transcriptional regulation, altered protein stability, or intratumoral metabolic heterogeneity may contribute to the heterogeneous protein expression pattern observed across breast tumors. Beyond the immunohistochemical findings, several complementary in silico analyses were performed to further characterize CYP4B1 at the transcriptomic, prognostic, systems, and structural levels.
Consistent with the immunohistochemical findings, transcriptomic analyses demonstrated reduced CYP4B1 mRNA expression in breast cancer at the population level compared with normal breast tissue. This notion is further supported by transcriptomic datasets demonstrating progressive CYP4B1 mRNA downregulation in more aggressive disease states.8,9,24 Regarding the survival analysis, no statistically significant differences in disease-free or overall survival were observed between CYP4B1 expression groups. While minor non-significant trends were observed, these do not support CYP4B1 as a prognostic biomarker in breast cancer. Although CYP4B1 expression is reduced in breast cancer and declines with increasing tumor grade, the observed non-significant trend toward poorer survival in CYP4B1-high groups does not imply a direct adverse effect; rather, it likely reflects tumor heterogeneity, context-dependent metabolic roles, or subtype-specific biology, and should be interpreted cautiously given the lack of statistical significance. In this framework, this pattern is not contradictory, as focal CYP4B1 protein retention in a metabolically distinct tumor subset may not follow a linear relationship with overall tumor aggressiveness or clinical outcome. Importantly, these considerations remain speculative and hypothesis-generating, as survival differences were not statistically significant and analyses were not stratified by molecular subtype.
Our PPI network analysis revealed that CYP4B1 is a hub protein connecting several metabolic functions and pathways, including fatty acid oxidation, glutathione conjugation, and xenobiotic metabolism. Prominent collaborators, such as glutathione S-transferases (GSTs), CYP2E1, and CYP3A5, jointly promote cellular defense against electrophilic and peroxidized lipids. Disruptions in this network may alter redox balance and trigger mutagenesis.27–29 The clustering of CYP4B1 with AASDH and SULT1C2 may establish crosstalk between amino acid metabolism and sulfate conjugation, consistent with the well-known roles of AASDH and SULT1C2 in these processes, reflecting previous evidence of CYP-SULT crosstalk in the bioactivation of carcinogens.30,31 These network-level associations are further supported by functional enrichment analysis, which demonstrated significant over-representation of metabolic pathways including xenobiotic metabolism by cytochrome P450, drug metabolism–cytochrome P450, arachidonic acid metabolism, linoleic acid metabolism, steroid hormone biosynthesis, and retinol metabolism. Collectively, these findings reinforce a contextual, systems-level role for CYP4B1 within coordinated lipid and xenobiotic metabolic networks, rather than implying direct mechanistic or causal effects in breast cancer progression. Accordingly, these network associations should be interpreted as biologically informative but hypothesis-generating, requiring experimental validation before mechanistic conclusions can be drawn.
Human CYP4B1 possesses an evolutionarily acquired mutation (P427S) in the meander region that destabilizes the heme environment and significantly decreases its catalytic activity. Previous biochemical and mutagenesis studies have demonstrated that the P427S substitution contributes to reduced catalytic competence of the native human enzyme.32–35 In the present study, AlphaFold-based structural modeling is used solely as contextual background to explain this intrinsic limitation and to support interpretation of the weak functional associations observed at the systems level. However, the present study does not experimentally evaluate enzyme reactivation, substrate turnover, or therapeutic manipulation. Consequently, all structural interpretations herein are confined to native CYP4B1 biology and serve to contextualize observed expression patterns rather than to support therapeutic reactivation strategies. Engineered CYP4B1-based suicide-gene or prodrug-activation strategies are conceptually distinct from native CYP4B1 biology and fall outside the scope of the present study.
Taken together, the limited catalytic competence of native human CYP4B1 provides a plausible biological context for the weak and non-predictive pharmacogenomic correlations observed in this study. Therefore, these correlations should not be interpreted as evidence of clinically relevant drug resistance or treatment response prediction. Accordingly, CYP4B1 is best interpreted as a differentiation-associated metabolic enzyme embedded within xenobiotic and redox networks, rather than as a dominant determinant of drug response or clinical outcome. These findings should be regarded as exploratory and hypothesis-generating, and do not support the use of CYP4B1 as a standalone prognostic or predictive biomarker. While this remains experimentally attractive, indication-specific optimization and rigorous safety evaluation will be required before any clinical translation can be considered.
This study has several limitations. The number of normal breast tissues in the immunohistochemical analysis was limited, consistent with commercial tissue microarray design, and these samples were used as qualitative reference controls rather than a statistically powered comparator. Consequently, the small number of normal controls, together with the limited prevalence of CYP4B1-positive tumors, reduces statistical precision and restricts the strength of clinicopathological comparisons. Moreover, the Allred scoring cut-off applied to define CYP4B1 positivity is not specifically validated for this marker, and thus the categorization should be interpreted as descriptive rather than clinically standardized. In addition, the observational design precludes causal inference, survival associations were not statistically significant, and pharmacogenomic correlations were weak. Furthermore, because only 18.3% of tumors exhibited CYP4B1 positivity, subgroup analyses and logistic regression modeling were performed on a limited number of positive events, reducing statistical power and increasing uncertainty around effect estimates. Analyses based on public datasets lacked subtype-specific stratification, and no functional validation experiments were performed. Therefore, the findings should be interpreted as descriptive and hypothesis-generating, and should not be considered evidence of clinical utility, mechanistic causality, or therapeutic relevance without further validation.
Conclusion
In summary, the principal finding of this study was the identification of focal and heterogeneous CYP4B1 protein expression in breast cancer, with expression retained in only a small subset of tumors and significantly associated with lower histological grade. These immunohistochemical findings suggest that CYP4B1 may represent a differentiation-associated metabolic feature of a biologically distinct subset of breast cancers rather than a universal characteristic of the disease.
Complementary in silico analyses demonstrated reduced CYP4B1 mRNA expression in breast cancer and provided systems-level insights through survival, protein–protein interaction, functional enrichment, pharmacogenomic, and structural analyses. However, these computational findings were exploratory in nature and should be interpreted as hypothesis-generating rather than evidence of prognostic, predictive, mechanistic, or therapeutic relevance.
Overall, the present study provides a descriptive multilevel characterization of CYP4B1 in breast cancer. Further experimental and functional studies are required to clarify the biological significance of CYP4B1 and determine whether it has clinical relevance in specific breast cancer subgroups.
Acknowledgments
The authors thank the Deanship of Scientific Research at Mutah University for supporting this project (No: 743/2023).
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Disclosure
The authors report no conflicts of interest in this work.
References
1. American Cancer S. Breast cancer statistics: how common is breast cancer? cancerorg. 2025.
2. Breast Cancer Research F. Breast cancer statistics and resources. BCRF. 2022.
3. International Agency for Research on C. Breast cancer cases and deaths are projected to rise globally to 2050. IARC. 2025.
4. Al-Saraireh YM, Alshammari F, Youssef AMM, et al. Cytochrome 4Z1 expression is associated with poor prognosis in colon cancer patients. Onco Targets Ther. 2021;14:5249–17. doi:10.2147/OTT.S332037
5. Al-Saraireh YM, Alshammari F, Abu-Azzam OH, et al. Targeting Cytochrome P450 enzymes in ovarian cancers: new approaches to tumor-selective intervention. Biomedicines. 2023;11(11):2898. doi:10.3390/biomedicines11112898
6. Al-Saraireh YM, Alshammari F, Satari AO, et al. Cytochrome 4Z1 expression connotes unfavorable prognosis in ovarian cancers. Medicina. 2022;58(9):1263. doi:10.3390/medicina58091263
7. Zhou M, Li J, Xu J, Zheng L, Xu S. Exploring human CYP4 enzymes: physiological roles, function in diseases and focus on inhibitors. Drug Discov Today. 2023;28(5):103560. doi:10.1016/j.drudis.2023.103560
8. Calaf GM, Crispin LA, Ossandon-Acosta F, Perez-Tapia S, Ardiles LN. The Cytochrome CYP4 in breast and other cancers. Biology. 2025;14(7):812. doi:10.3390/biology14070812
9. Röder A. Spotlight on CYP4B1. Int J Mol Sci. 2023;24(3):2038. doi:10.3390/ijms24032038
10. Lim S, Alshagga M, Ong C, Chieng J, Pan Y. Cytochrome P450 4B1 (CYP4B1) as a target in cancer treatment. Hum Exp Toxicol. 2020;39(6):785–796. doi:10.1177/0960327120905959
11. Yin Y, Tong L, Wan Z, et al. CYP4B1 polymorphisms and the risk of breast cancer in Chinese women: a case-control study. BMC Cancer. 2023;23(1):1177. doi:10.1186/s12885-023-11477-y
12. Allred DC, Harvey JM, Berardo M, Clark GM. Prognostic and predictive factors in breast cancer by immunohistochemical analysis. Mod Pathol. 1998;11(2):155–168.
13. Al-Saraireh YM, Alboaisa NS, Alrawashdeh HM, et al. Screening of cytochrome 4Z1 expression in human non-neoplastic, pre-neoplastic and neoplastic tissues. Ecancermedicalscience. 2020;14:1114. doi:10.3332/ecancer.2020.1114
14. Alshammari FO, Satari AO, Aljabali AS, et al. Glypican-3 differentiates intraductal carcinoma and Paget’s disease from other types of breast cancer. Medicina. 2023;59(1):86–100. doi:10.3390/medicina59010086
15. Al-Saraireh Y, Alrawashdeh F, Al-Shuneigat J, Alsbou M, Alnawaiseh N, Al-Shagahin H. Screening of Glypican-3 expression in human normal versus benign and malignant tissues: a comparative study glypican-3 expression in cancers. Biosci Biotechnol Res Asia. 2016;13(2):687–692. doi:10.13005/bbra/2085
16. Al-Saraireh YM, Alshammari FO, Youssef AM, et al. Screening of glypican-6 expression in Benign, primary and metastatic colon cancers. Clin Med Insights Oncol. 2021;15:11795549211036419. doi:10.1177/11795549211036419
17. Cui H, Zhao G, Lu Y, et al. TIMER3: an enhanced resource for tumor immune analysis. Nucleic Acids Res. 2025;53(W1):W534–W541. doi:10.1093/nar/gkaf388
18. Tang Z, Li C, Kang B, Gao G, Li C, Zhang Z. GEPIA: a web server for cancer and normal gene expression profiling and interactive analyses. Nucleic Acids Res. 2017;45(W1):W98–W102. doi:10.1093/nar/gkx247
19. Liu CJ, Hu FF, Xie GY, et al. GSCA: an integrated platform for gene set cancer analysis at genomic, pharmacogenomic and immunogenomic levels. Briefings Bioinf. 2023;24(1). doi:10.1093/bib/bbac558
20. Liu CJ, Hu FF, Xia MX, Han L, Zhang Q, Guo AY. GSCALite: a web server for gene set cancer analysis. Bioinformatics. 2018;34(21):3771–3772. doi:10.1093/bioinformatics/bty411
21. Jumper J, Evans R, Pritzel A, et al. Highly accurate protein structure prediction with AlphaFold. Nature. 2021;596(7873):583–589. doi:10.1038/s41586-021-03819-2
22. Fleming J, Magana P, Nair S, et al. AlphaFold protein structure database and 3D-Beacons: new data and capabilities. J Mol Biol. 2025;437(15):168967. doi:10.1016/j.jmb.2025.168967
23. Mohr L, Rainov NG, Mohr UG, Wands JR. Rabbit cytochrome P450 4B1: a novel prodrug activating gene for pharmacogene therapy of hepatocellular carcinoma. Cancer Gene Ther. 2000;7(7):1008–1014. doi:10.1038/sj.cgt.7700190
24. Liu X, Jia Y, Shi C, et al. CYP4B1 is a prognostic biomarker and potential therapeutic target in lung adenocarcinoma. PLoS One. 2021;16(2):e0247020. doi:10.1371/journal.pone.0247020
25. Li Q, Liu X-L, Jiang N, et al. A new prognostic model for RHOV, ABCC2, and CYP4B1 to predict the prognosis and association with immune infiltration of lung adenocarcinoma. J Thorac Dis. 2023;15(4):1919–1934. doi:10.21037/jtd-23-265
26. Lin J-T, Chan T-C, Li C-F, et al. Downregulation of the cytochrome P450 4B1 protein confers a poor prognostic factor in patients with urothelial carcinomas of upper urinary tracts and urinary bladder. APMIS. 2019;127(4):170–180. doi:10.1111/apm.12939
27. Sharma R, Yang Y, Sharma A, Awasthi S, Awasthi YC. Antioxidant role of glutathione S-transferases: protection against oxidant toxicity and regulation of stress-mediated apoptosis. Antioxid Redox Signal. 2004;6(2):289–300. doi:10.1089/152308604322899350
28. Valencia A, Morán J, Camacho-Carranza R, Prospero-García O, Espinosa-aguirre J. CYP2E1 induction leads to oxidative stress and cytotoxicity in glutathione-depleted cerebellar granule neurons. Toxicol In Vitro. 2014;28.
29. Fan HH, Li BQ, Wu KY, et al. Polymorphisms of Cytochromes P450 and Glutathione S-Transferases synergistically modulate risk for Parkinson’s disease. Front Aging Neurosci. 2022;14:888942. doi:10.3389/fnagi.2022.888942
30. Daniels J, Kadlubar S. Sulfotransferase genetic variation: from cancer risk to treatment response. Drug Metab Rev. 2013;45(4):415–422. doi:10.3109/03602532.2013.835621
31. Liu Y, Glatt H. Human cytochrome P450 2E1 and sulfotransferase 1A1 coexpressed in Chinese hamster V79 cells enhance spontaneous mutagenesis. Environ Mol Mutagen. 2010;51(1):23–30. doi:10.1002/em.20503
32. Hüsken S, Röder A, Ptok J, et al. Functional analysis of CYP4B1 enzymes from apes and humans uncovers evolutionary hot spots for adaptations of the catalytical function. PLoS Genet. 2025;21(6):e1011750. doi:10.1371/journal.pgen.1011750
33. Röder A, Hutter MC, Heitzer E, et al. Exploration of CYP4B1 Substrate Promiscuity Across Three Species. Catalysts. 2025;15(5):454. doi:10.3390/catal15050454
34. Wiek C, Schmidt E, Roellecke K, et al. Identification of amino acid determinants in CYP4B1 for optimal catalytic processing of 4-Ipomeanol. Biochem J. 2014;465.
35. Wiek C, Schmidt EM, Roellecke K, et al. Identification of amino acid determinants in CYP4B1 for optimal catalytic processing of 4-ipomeanol. Biochem J. 2015;465(1):103–114. doi:10.1042/BJ20140813
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
The Prognostic Role of HuR Varies Between Different Subtypes of Breast Cancer Patients: Data Mining and Retrospective Analysis
Liao Y, Liao Y, Li J, Li Y, Fan Y
Breast Cancer: Targets and Therapy 2023, 15:135-146
Published Date: 11 February 2023
Genetic Alterations in HER2-Positive and Equivocal Breast Cancer by Immunohistochemistry
Tsai YF, Hsu CY, Chiu YN, Huang CC, Chou SH, Lin YS, Chao TC, Liu CY, Chiu JH, Tseng LM
Breast Cancer: Targets and Therapy 2025, 17:253-263
Published Date: 12 March 2025
Understanding the Molecular Toxicity of Metal-Based Nanoparticles Through Nanotoxicomics
González-Vega JG, Fabian-Ortiz E, Hernández-Pérez J, Hinojosa Alvarez S, Chavez Santoscoy RA
International Journal of Nanomedicine 2025, 20:13293-13315
Published Date: 4 November 2025
HER2 Overexpression in Invasive Breast Carcinoma in the Democratic Republic of Congo and its Association with Selected Clinicopathological Characteristics
Ilunga Nday G, Ilunga Banza M, Umpungu AN, Kabamba Nzaji M, Moyambe JNT
Breast Cancer: Targets and Therapy 2026, 18:600288
Published Date: 15 April 2026