Back to Journals » Infection and Drug Resistance » Volume 19
Antimicrobial Peptides Against Antimicrobial-Resistant Bacteria: Focus on Machine Learning
Authors Tahmasebi H ![]() , Arabestani MR
, Arabestani MR
Received 10 February 2026
Accepted for publication 9 June 2026
Published 23 June 2026 Volume 2026:19 602699
DOI https://doi.org/10.2147/IDR.S602699
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 4
Editor who approved publication: Dr Hazrat Bilal
Hamed Tahmasebi,1,2 Mohammad Reza Arabestani3,4
1School of Medicine, Shahroud University of Medical Sciences, Shahroud, Iran; 2Department of Bacteriology, Pasteur Institute of Iran, Tehran, Iran; 3Department of Microbiology, School of Medicine, Hamadan University of Medical Sciences, Hamadan, Iran; 4Nutrition Health Research Centre, Institute of Health Sciences and Technologies, Hamadan University of Medical Sciences, Hamadan, Iran
Correspondence: Mohammad Reza Arabestani, Department of Microbiology, School of Medicine, Hamadan University of Medical Sciences, Hamadan, Iran, Email [email protected]
Abstract: Antimicrobial resistance (AMR) represents a pressing public health threat of the 21st century, with an estimated ten million deaths annually from drug-resistant infections by 2050. Diminishing pipelines and the accelerating emergence of multidrug-resistant pathogens make the development of novel antibacterials more urgent than ever. Antimicrobial peptides (AMPs) are among the most promising alternatives to conventional drugs, exhibiting broad antimicrobial spectra, rapid kinetics, and mechanisms that are difficult for bacteria to circumvent. However, the problem of discovering and engineering clinically useful AMPs with desirable properties out of large sequence spaces remains unsolved by traditional approaches. Machine learning (ML) enables fast screening of millions of compounds, generation of de novo sequences with predicted therapeutic potential, and simultaneous multiobjective optimisation of efficacy, safety, stability, and manufacturability. This review provides a critical appraisal of the current advances and prospective directions in computational discovery of AMPs that can combat resistant strains, focusing on available resources for machine learning in the domain of bioinformatics, evaluation of existing approaches to modeling peptide structure, activity, and interactions ranging from classical ML algorithms to DL and generative artificial intelligence (AI) models, and a practical roadmap of how the AMP discovery pipeline could proceed towards animal studies and clinical application through the use of active learning, fine-tuned protein language models, structural graph neural networks, and other modern techniques. Finally, we discuss challenges that may hinder a successful transition from ML-assisted design to the clinic and offer actionable recommendations to overcome them. Comparison of traditional and AI methods for antimicrobial peptide synthesis.The image compares two methods for antimicrobial peptide synthesis. The traditional method involves scientists using lab equipment to select molecules, resulting in high attrition. The text reads: ′Molecules picked by traditional methods for antimicrobial peptide synthesis.′ In contrast, the predictive AI machine learning model uses AI technology to select molecules, leading to low attrition. The text reads: ′Molecules picked by predictive AI model for antimicrobial peptide synthesis.′ The comparison highlights the efficiency of AI over traditional methods.
Keywords: antimicrobial peptides, antimicrobial resistance, machine learning, DL, protein language models, drug discovery, clinical trials, geometric DL
Introduction
The Global Antimicrobial Resistance Crisis
Antimicrobial Resistance (AMR) poses an unprecedented threat to public health in the 21st century, having the potential to reverse many of the gains made in medicine in the last century. According to the World Health Organisation (WHO), AMR ranks among the top 10 global health threats of the present era.1,2 Economically, the cost of AMR is expected to reach USD 100 trillion globally by the middle of the century, driven by higher healthcare costs and lower productivity.3 The AMR is a predictable outcome of evolution driven by the widespread use of antibiotics in clinical settings, agriculture, and veterinary medicine.2,4,5 The resistance mechanisms utilised by bacteria are varied and include enzyme-based inactivation of antibiotics (eg., beta-lactamases), target modification (eg., fluoroquinolones), overexpression of efflux pumps, which reduce intracellular drug concentration, and altered membrane permeability, which reduces drug uptake into the cell.5–7 The group of bacteria known as ESKAPE pathogens (Enterococcus faecium, S. aureus, Klebsiella pneumoniae, A. baumannii, Pseudomonas aeruginosa, and Escherichia coli) is the most dangerous hospital-acquired microorganism due to their high propensity to acquire resistance determinants.8,9
The Priority Pathogens List was published by the WHO in 2017 and recommended a set of pathogens for research and development in terms of new drugs. This classification is based on 12 families of bacteria and prioritises critical, high-priority, and medium-priority pathogens according to their antibiotic resistance patterns and the urgency of innovation (Figure 1).2 Critical-priority pathogens include carbapenem-resistant A. baumannii, carbapenem-resistant Pseudomonas aeruginosa, and carbapenem-resistant Enterobacteriaceae with additional resistance to third-generation cephalosporins. These pathogens cause nosocomial infections, such as ventilator-associated pneumonia, bacteremia, and wound infections, that are increasingly resistant to available therapies.10 High-priority pathogens include methicillin-resistant Staphylococcus aureus (MRSA), vancomycin-resistant Enterococcus faecium, extensively drug-resistant Neisseria gonorrhoeae, and other types of bacteria. Unfortunately, the pipeline of antibiotics is far from adequate. Only 12 new drugs were registered between 2010 and 2020; the majority were modifications of existing antibiotic groups.7,11 Figure 1.
 |
Figure 1 WHO priority pathogen categories with projected AMR mortality timeline. |
The primary impact of AMR is far-reaching, affecting a variety of systems beyond the clinical outcomes of antibiotic-resistant patients. Examples of medical services where the absence of effective antibiotics increases the risk to patients include organ transplantation, chemotherapy, cardiothoracic surgery, and even non-abortive childbirth.12 What was once thought of as a dystopian future in which minor infections could be fatal due to the lack of effective antibiotic therapy has now become a clinical reality, experienced daily in nearly every health care facility globally. This dire need for alternative antimicrobial agents that can bypass or circumvent current mechanisms of resistance, while minimising the potential for developing new resistance to those products, has prompted many scientists to search for new antibiotics and to increase the use of current antibiotics.13
Antimicrobial Peptides as a Therapeutic Frontier
As promising candidates to replace current antibiotics, antimicrobial peptides (AMPs) represent an extremely interesting group. Known as host defence peptides (HDPs), AMPs are conserved peptides found in almost all kingdoms on Earth, including bacteria, fungi, plants, insects, amphibians, and mammals.13 These polypeptides consist of 10–100 amino acids and demonstrate broad-spectrum antimicrobial activity toward bacteria, fungi, viruses, and some protozoans.14
The mechanism of action of AMP is completely different from that of classical antibiotics, which primarily target microbial biosynthetic pathways (including cell wall synthesis, protein synthesis, and nucleic acid replication).15 Unlike conventional antibiotics, AMPs primarily bind to the membrane surface via electrostatic forces, interacting with the negative charges of the cationic peptides (such as phosphatidylglycerol, cardiolipin, lipopolysaccharides (LPS) in Gram-negative bacteria, and lipoteichoic acid in Gram-positive bacteria).16 Subsequent binding results in the formation of pores in the cell wall, leading to membrane destruction. Disruption occurs through various mechanisms, including barrel stave formation, the toroidal pore hypothesis, the carpet mechanism, and micelle formation.17 Since disruption occurs via physical mechanisms, resistance to AMPs cannot develop easily, as significant changes in membrane structure are required.
In addition to acting directly on the cell membrane, many AMPs can penetrate microbial membranes to target intracellular components that inhibit processes such as DNA replication, RNA transcription, protein biosynthesis, or enzyme catalysis.18 Moreover, some AMPs possess immunomodulatory activities that stimulate innate immunity, regulate cytokine secretion, and induce wound healing.19 The diverse range of activities makes AMPs an attractive option for treating infections in immunocompromised individuals. Lastly, some AMPs possess strong antibiofilm properties, destroying biofilm matrix structures and eliminating persister bacteria that are known to resist conventional antibiotics.20
There are several distinct benefits to using AMPs instead of conventional antibiotics. First, their broad spectrum of activity obviates the need to identify the causative agent before administering treatment, which is essential in cases of sepsis and other emergent infections.21 Unlike conventional antibiotics, AMPs exert rapid effects and can kill pathogens within a few minutes after administration, whereas antibiotics can only suppress bacterial growth for extended periods.22 The diverse mechanism of action minimises the likelihood of developing resistance to AMPs, a notion consistently validated by serial passage experiments, in which resistance to subinhibitory levels of AMPs was rarely observed.23 Lastly, AMPs enhance the efficacy of antibiotics when used in combination therapies, thereby extending the therapeutic potential of conventional medications while limiting the selection pressure for resistance.24
Despite the enormous potential benefits of therapeutic use of AMP, there have been multiple substantial barriers to the clinical translation of these candidate therapies. Many naturally occurring AMPs exhibit cytotoxicity, most notably due to their significant hemolytic activity against human erythrocytes, which greatly limits their therapeutic index.25 Due to their susceptibility to proteolytic degradation by both host and pathogen proteases, AMPs have very short plasma half-lives, typically measured in minutes rather than hours.26 Manufacturing costs for therapeutic peptides are significantly greater than for small-molecule antibiotics. As such, there are issues regarding both commercial viability and access to AMPs in resource-limited settings, where AMR typically poses the largest burden.27 Finally, because AMPs are poorly absorbed from the intestine due to degradation by digestive proteases and limited intestinal absorption, they typically require parenteral administration (as opposed to enteral routes), which contributes to the complexity of their clinical deployment.28
The Computational Revolution in Antimicrobial Peptides Discovery
The combination of rapidly growing biological sequence databases, advancements in high-performance computing, and breakthroughs in artificial intelligence (AI) and machine learning (ML) has led to major changes in AMP discovery research.29 Conventional approaches to AMP discovery have relied heavily on natural product screening, rational design based on structure–activity relationship studies, and combinatorial library construction—approaches that can be rather laborious and provide only narrow coverage of the immense search space of AMPs.30 The number of distinct peptide sequences of 20–50 residues using the standard 20-amino-acid alphabet is staggeringly large, rendering exhaustive screening experiments infeasible even with high-throughput technologies. Computationally driven strategies for AMP discovery, especially those that rely on ML, offer an alternative to experimentally intensive techniques. They use known AMP data to learn regularities and predict the antimicrobial activity of a novel sequence without the need for costly synthetic or biological experiments.31 Computational studies of early stages in the development of AMP predictors have focused on classic ML techniques such as SVM, RF, and logistic regression, which use handcrafted sequence feature representations as input.32
This article critically reviews the present and anticipated future of AMP discovery using ML. The various databases and computational tools are extensively surveyed for their applicability to AMP discovery. Furthermore, the strengths and limitations of the various ML architectures are discussed, and a translational pathway connecting computational design to animal trials to clinical studies in humans will be outlined. Lastly, key impediments to the full realisation of AI-based AMPs will be assessed. Both discriminative models for predicting in vivo activity and generative models designed de novo will be discussed, with emphasis on combating antibiotic-resistant pathogens.
Antimicrobial Peptides
AMPs have been found in all known living organisms, and the first AMP was likely found in a plant. In the 1960s, before the advent of large-scale research, it was first discovered that milk contained a protein called lactoferrin, and that frogs contained a similar protein called bromin. Although these factors were researched in the mid-1990s, and researchers originally established a significant role in insect immunity by showing that when the genetic coding for AMPs was removed, fruit flies were susceptible to potent fungal pathogens.33,34
Mechanism of Action of Antimicrobial Peptides
Previously, the specific functional mechanism of AMPs had been a point of scientific debate. The first assumption was that the role of AMPs in bacteria was limited to their ability to permeabilize and destabilise cellular membranes. In contrast, more recent studies have shown that many peptides, when added to bacteria, gained access to the bacterial membrane without damaging it, and subsequently accumulated within the bacterial cytoplasm to inhibit several essential pathways in the bacterial cell, including nucleic acid synthesis, protein synthesis, cell division, and cell wall synthesis. Here, we present examples of the various mechanisms through which peptide antibiotics exert their effects.33,34 Table 1.
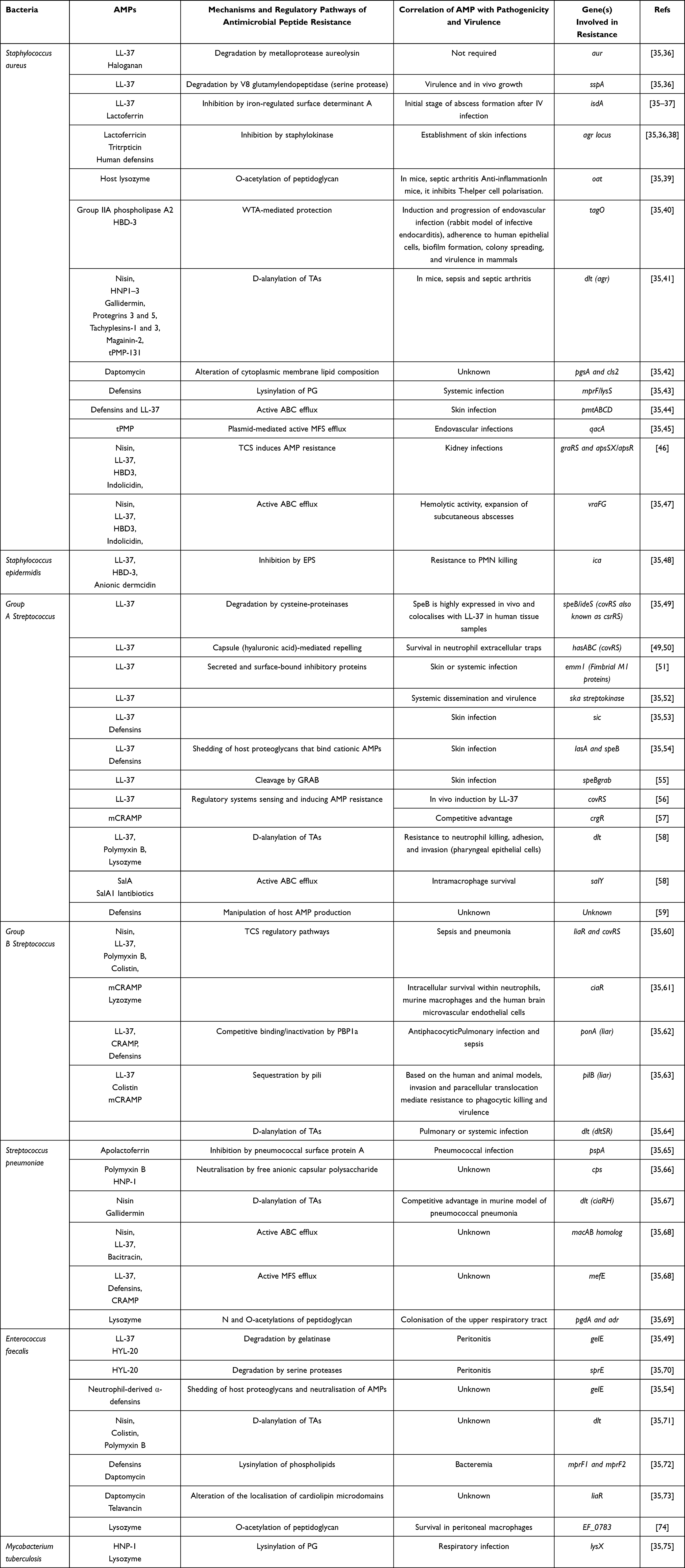 |
Table 1 Mechanisms of Human and Nan-Human AMP Resistance in Nosocomial Gram-Positive Bacteria and Mycobacterium Tuberculosis |
Cell Membrane Lysis
Most AMPs are classified as membrane-disrupting agents. During the initial phase of interaction, these molecules form electrostatic interactions with anionic lipid structures exposed on the bacterial surface, such as LPS in Gram-negative bacteria and lipoteichoic acids (LTA) in Gram-positive bacteria. This interaction initiates destabilisation of the anionic phospholipids, resulting in pore formation and ultimately cell death as cellular contents leak. The most recognised mechanisms by which AMPs disrupt cellular membranes to kill microorganisms are the barrel-stave, carpet, and toroidal models.76,77 As proposed by the barrel-stave model, peptides orient to bind to cellular membranes, leading to their aggregation into a bilayer. The greater the assembly, the more likely hydrophilic peptides will create a pathway to the interior of the membrane, while hydrophobic peptides will conjugate near the lipid core. The carpet model proposes that when peptides become immersed in the cell membrane’s outer phospholipid layer, their incorporation into the membrane causes it to break apart, as if by a “detergent,” eventually allowing them to form a relatively permanent film (ie., a carpet). Integrated peptides generate aggregation in the toroidal model, and lipid monolayers must constantly flex as they pass through the pores.10,31 The combined mechanism provides the surface of the core protein as determined by the polar regions of the lipids in conjunction with the integrated polypeptide chains.78,79
Non-Permeabilising Mechanism (Targeting Intracellular Components)
Recent findings show that AMPs kill by mechanisms distinct from membrane perturbation. Two examples may be illustrative: both peptides penetrate membranes and kill by targeting intracellular components, or by inhibiting nucleic acid and protein biosynthesis, or even cell wall biosynthesis and division.78
Inhibition of Cell Wall Synthesis
The bacterial cell envelope provides structural support and prevents osmotic lysis; its function is paramount. Antibiotics, including penicillin and cephalosporins, cause bacterial cell death by targeting the cell envelope, which confers selective toxicity because it is unique to prokaryotic cells, and (most) mammalian cells do not possess similar features. Several studies have shown that certain AMPs do not diffuse into membranes, nor do they cause inner K+ loss or ATP leakage. In contrast, research has shown that plectasin ultimately leads to bacterial death by blocking envelope turnover by binding lipid II. This bacterial death mechanism is observed with membrane-acting antibacterial peptides such as lipopeptides (daptomycin) and lantibiotics (mersacidin), as well as bacteriocins, and with nisin, a commonly used food preservative that kills bacteria by binding lipid II to form pores, leading to leakage and cell death. In other studies, nisin has been shown to competitively bind lipid III or IV and effectively prevent the formation of teichoic or lipoteichoic acids. N-acetylmuramoyl-L-alanine amidase is an autolysin that was activated in Staphylococcus simulans by nisin, and also by Pep5 (an AMP), which ultimately caused lysis of the bacteria77,80,81 Table 2.
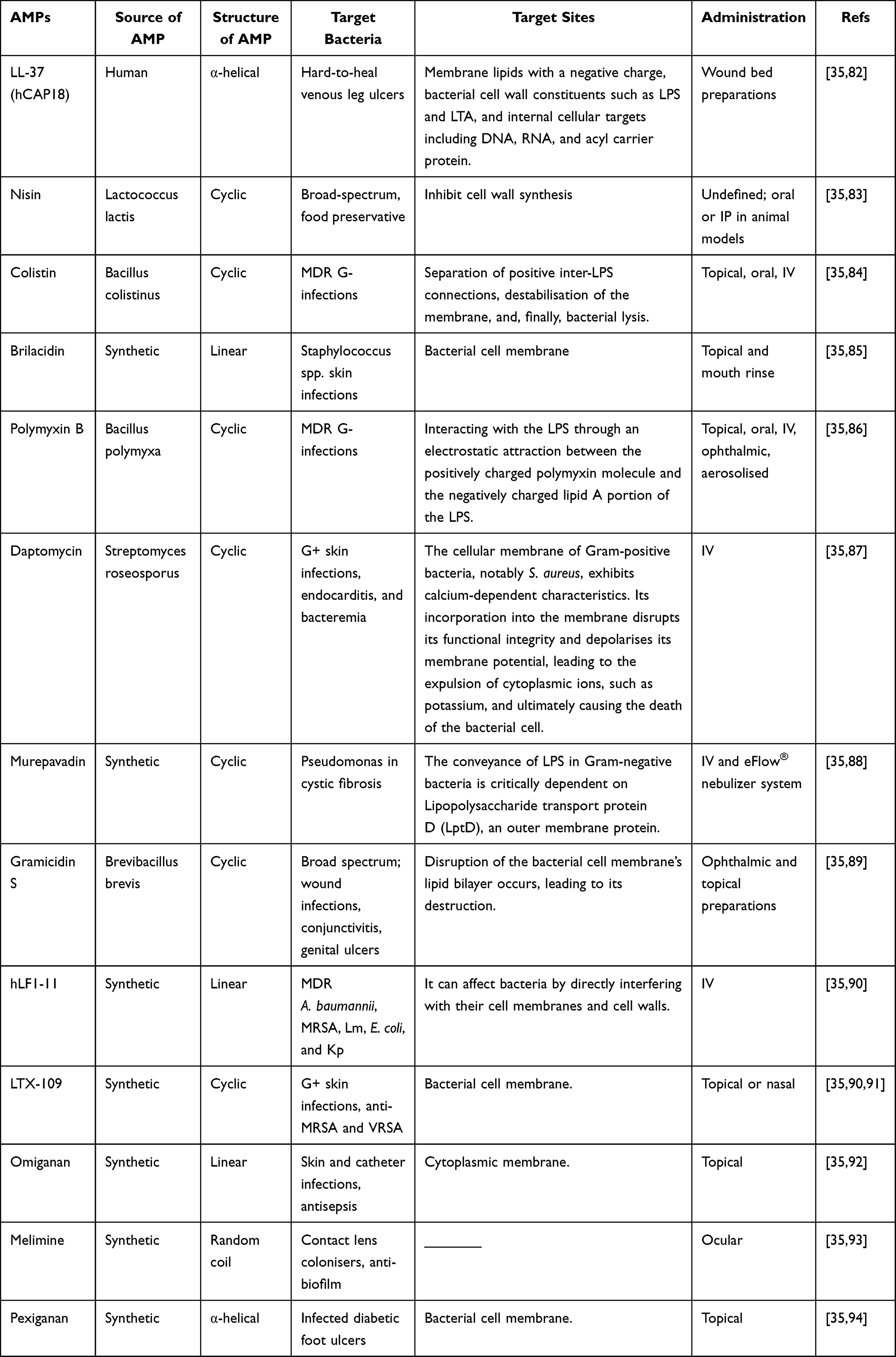 |
Table 2 Selected AMP with Bacterial Membrane Depolarisation |
Inhibition of Nucleic Acid and Protein Synthesis
Peptides that translocate through bacterial membranes inhibit cellular DNA, RNA, and protein synthesis. The short 21-residue peptide buforin II gains access and accumulates inside bacteria without disrupting their cell membranes. The buforin peptide translocates via the Pro11 hinge, which can transport otherwise non-cell-permeable peptides into cells. Internalisation of buforin II led to cell death, given its affinity for bacterial DNA and RNA, which affected their cellular functions. The robust binding affinity with nucleic acids indicates that buforin II is similar to the N-terminal of histone H2A, as peptoids like the dermaseptin, defensin, and pleurocidin have also been shown to inhibit bacterial nucleic acid and protein synthesis through impeding radioactive precursors (thymidine, uridine, and leucine) during macromolecular biosynthesis.79,95 Insects contain proline-rich AMP such as pyrrhocoricin, drosocin, and apidaecin. These peptides act non-lytically to kill bacteria; they then use SbmA to enter bacterial cells, where they accumulate and target or bind to their structures. Proline-rich peptides are found to bind DnaK from bacterial lysates with high affinities at low micromolar concentrations. However, DnaK is not the principal target of proline-rich peptides, as DnaK null mutants were shown to be more sensitive to proline peptides than wild-type strains, indicating DnaK was not the target of proline-rich peptides. Proline-rich peptides have been shown to bind strongly to the 70S ribosomal subunit and, at nanomolar concentrations, inhibit protein translation efficiently. Crystallography has recently reported an oncocin derivative that inhibits translation at the 70S ribosome. It was shown that the peptide acts, apparently, in the ribosomal exit tunnel to disrupt the binding of aminoacyl-tRNA.96,97 Figure 2.
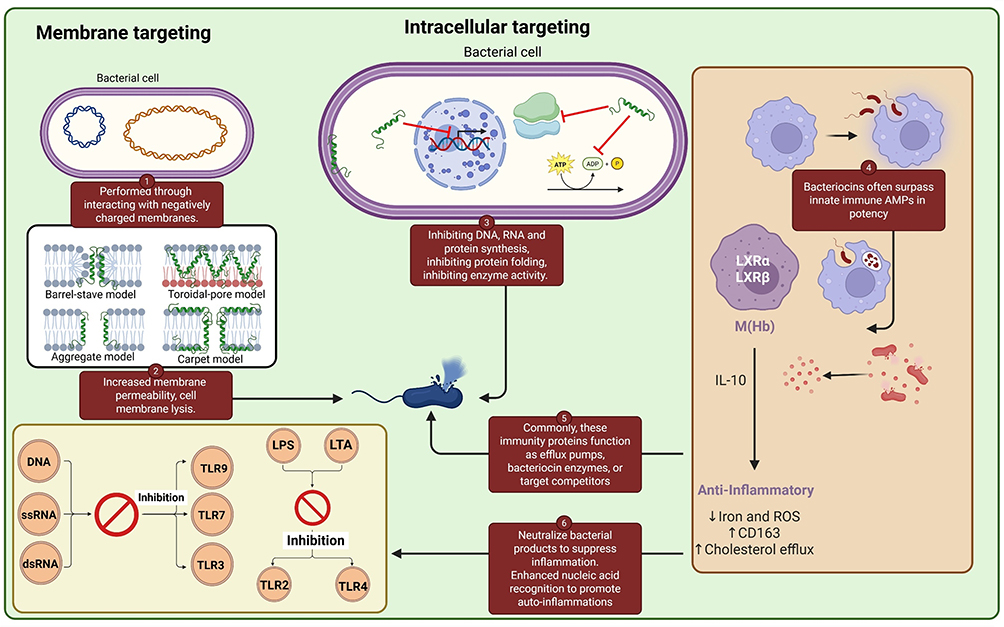 |
Figure 2 How AMP fight bacteria: proposed models. Through interactions with membranes, AMPs exert their direct germ-killing mechanism by increasing membrane permeability, rupturing the cell membrane, or causing leakage of intracellular material, ultimately leading to cell death. Membrane-pore formation is primarily explained by four models, specifically the barrel-stave, toroidal-pore, carpet, and aggregate models. After the AMPs pass through the phospholipid membrane, their hydrophobic components associate with the inner hydrophobic region of the bilayer, while their hydrophilic components remain on the exterior. AMPs employ another bactericidal mechanism by permeating the cytoplasm and interacting with internal cellular components, including obstructing DNA, RNA, and protein synthesis; disrupting protein folding; inhibiting enzyme function and cell wall synthesis; and promoting the release of lytic enzymes to demolish cellular architecture. |
Antimicrobial Peptides-Producing Bacteria
The production of AMP most likely gives the bacteria a competitive advantage by eliminating competing bacteria from their niche. These bacteriocins are often more potent than innate immune AMPs. The resistance gene is normally located close to the biosynthesis genes, and is generally involved in self-protection by producers. Resistance genes can be found in chromosomes and/or mobile genetic elements. These immunity proteins have efflux pumps, bacteriocin enzymes, or target competitors when acting in immunity. Staphylococcus epidermidis has an example of epidermin immunity. The 54-kb plasmid encodes a peptide that has production and immunity functions. The peptide is exported from the cytoplasmic membrane by the EpiFEG ABC transporter, providing immunity.98,99
Nisin is a widely studied bacteriocin used as a food preservative, as shown in Table 3. The nisin resistance genes in its producer, Lactococcus lactis, can be transferred to other bacteria, conferring immunity to nisin. Foodborne bacteria can also develop nisin resistance. The frequency of spontaneous nisin resistance in Listeria monocytogenes ranges from 10–2 to 10–7, depending on the strain and conditions. Mechanisms of nisin resistance can involve changes to the bacterial cell wall/membrane composition, protection of lipid II, and secretion of enzymes. These examples demonstrate that bacteria can develop resistance to AMPs and indicate the potential for existing natural determinants for resistance to spread if selected.76,100 Table 3.
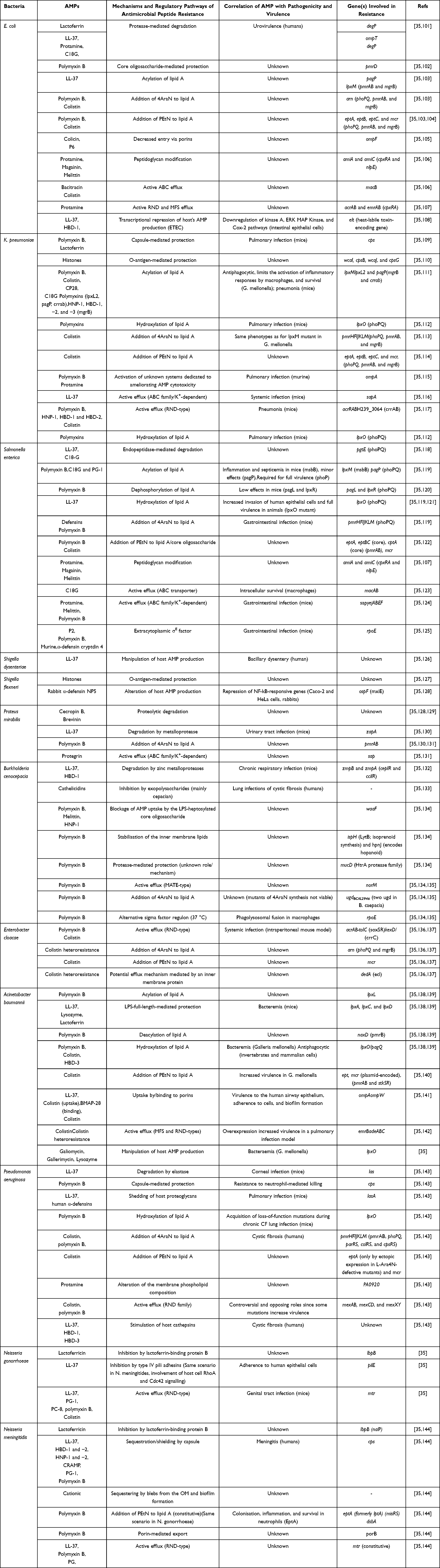 |
Table 3 Mechanisms of Human and Nan-Human AMP Resistance in Nosocomial Gram-Negative Bacteria (Enterobacteriaceae and Non-Enterobacteriaceae) and Gram-Negative Cocci |
Advantages of Antimicrobial Peptides
Broad Spectrum, Rapid Killing, and Synergism with Antibiotics
Antimicrobial peptides have demonstrated efficacy against many Gram-positive and Gram-negative bacteria, and some are also active against viruses, parasites, and fungi. Its broad activity is particularly useful for polymicrobial infections, particularly among hosts with compromised immune systems who regularly acquire multiple infections. These peptide groups also exhibit rapid killing activity at their MICs, unlike a conventional antibiotic.95,96 Many fast-acting AMPs possess attributes that limit the development of resistance, improve prognoses, contain infections, or shorten treatment duration. Both antibiotics and peptides quickly weaken bacterial cell membranes cooperatively, allowing greater antibiotic access to bind bacterial targets. There is a range of benefits from the synergistic combination of antibiotics and peptides. These benefits might include reducing the likelihood of resistance, allowing a lower antibiotic dose with lower toxicity, or broadening the range of pathogens effectively treated.4,80,145
Active Against Drug-Resistant Pathogens and Low Emergence of Bacterial Resistance to Antimicrobial Peptides
The mechanism of killing associated with AMPs does not allow for cross-resistance with traditional antibiotics. Furthermore, AMPs can kill multidrug- and pan-drug-resistant bacterial strains. The occurrence of resistance to AMPs is rarely associated with a significant risk of resistance development compared to traditional antibiotics, likely because the mechanisms of action of AMPs may involve the cellular membrane or target vital cellular processes; thus, these compounds are often assessed as “dirty” drugs. The likelihood of developing a mutation resistant to AMPs is low; it would require either modifying the lipids that comprise the bacterial cell membrane or multiple mutations in targets essential for bacterial fitness or survival.146 Others have demonstrated that bacteria do not develop resistance after 7 passages of subinhibitory pexiganan concentrations. In clinical trials, authors reported observations consistent with those previously reported in the literature and did not demonstrate significant development of pexiganan resistance in clinical patients. Resistance to ofloxacin was reported by some individuals in trials. Researchers conducting long-term studies with other AMPs reported either a complete absence of resistance to AMPs or a lower likelihood of bacterial resistance compared with standard antibiotics.147,148
Anti-Endotoxic Activity
LPS is a unique pathogenic factor for Gram-negative bacteria. The activation of mononuclear phagocytes to secrete proinflammatory cytokines such as TNF-α and IL-6 occurs when LPS is liberated into the host during bacterial death, replication, or antibiotic exposure. Excessive amounts of LPS cause excessive quantities of proinflammatory cytokines to be secreted and may result in fatal sepsis. Each year, around 18 million people are affected by septic shock worldwide, and approximately 30% of those affected die. Numerous AMPs prevent toll-like receptor 4 activation by binding LPS and blocking LPS association with LBP. These peptides subsequently inhibit proinflammatory cytokine secretion and the secretion of other mediators of septic shock. LL37 is a human cathelicidin that inhibits the binding of LPS to CD14 on mononuclear phagocytes and the inhibition of TNF-α expression. In models of murine endotoxin shock, LL-37 was reported to be a potent inhibitor of proinflammatory cytokines. Furthermore, LL-37 interfered with the assembly of the LPS receptor complex (CD14/TLR4) and blocked JNK-mediated apoptosis phosphorylation in endothelial cells following LPS exposure. AMPs have been identified as inhibitors of serious consequences such as sepsis and endotoxaemia, working by neutralising LPS, preventing substantial loss of life worldwide. Additionally, it offers a unique advantage in treating Gram-negative infections, as conventional antibiotics (especially cell wall inhibitors) can lead to greater LPS release and cell lysis during treatment.149,150
Immune-Modulatory Properties
Numerous AMPs possess strong antimicrobial activity and directly modulate immunity to enhance bacterial infection clearance. LL-37 induces chemotaxis, modulates the release of pro- and anti- inflammatory cytokines, regulates the differentiation of immune cell populations, and stimulates dendritic cell functions. In addition, LL-37 is implicated in the generation of neutrophil extracellular traps (NETs), in which neutrophil DNA forms extracellular fibres that bind to the pathogen. Cathelicidin stimulates both NET induction and protects NETs from degradation by bacterial nucleases. The biochemical facilitators are hydrophobicity and cationicity. LL-37 is chemotactic and induces purified human neutrophils, CD4 T cells, and monocytes to migrate.151,152 IDR-1 lacked direct in vitro antimicrobial activity, yet it protected in a mouse model of bacterial infection. IDR-1 enhances the anti-inflammatory cytokine IL-10 in the blood and reduces the pro-inflammatory cytokines TNF- and IL-6 at the site of infection. As a chemotactic agent, IDR-1 attracts monocytes and macrophages to the site of infection by enhancing the production of chemokines such as MCP-1 and RANTES, which are critical for protection. The tremendous immunomodulatory properties of IDR-1 suggest it will be an effective candidate for the clearance of bacterial infections. In summary, the immunomodulatory functions of AMPs effectively control inflammation in the host, which must be potent enough to preserve key aspects of the immune responses required for the resolution of a bacterial infection.151 Table 4.
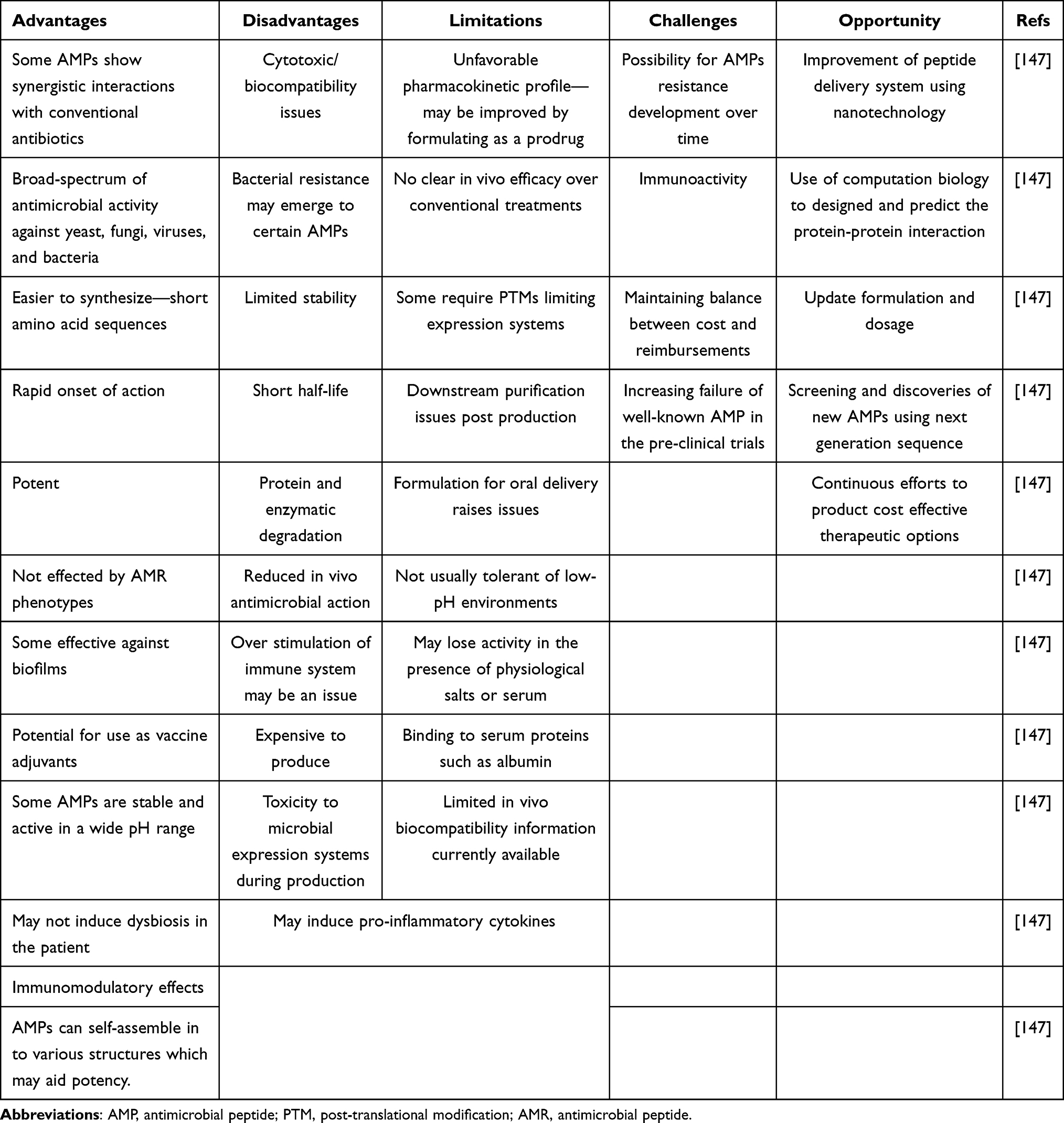 |
Table 4 Advantages, Disadvantages, and Limitations of AMP |
Mechanisms of Antimicrobial Peptide Resistance in Bacteria
Given bacteria’s adaptation and resistance to AMPs, the extensive use of AMPs in healthcare settings should promote the emergence of novel resistance mechanisms or the dissemination of existing resistance elements. This speculation is supported by the observations discussed in more detail below. After millennia of co-evolution with hosts, bacteria have developed defences against host AMPs that are lethal to other forms of life. The combinations of cellular processes that encode these mechanisms protect bacteria during host colonisation by either exploiting other pathways or by directly responding to host AMPs. We consider these mechanisms intrinsic resistance mechanisms because they do not require further genetic adaptation. Examples of acquired resistance show that bacteria have adapted to tolerating AMPs better because of these peptides; acquired resistance is observed despite the phylogenetic proximity of the parent strain. Genes encoding self-protective immunity in bacteria that tolerate AMPs are often located on their plasmids or transposons.35,153 An overview of all these mechanisms is depicted in Figure 3
 |
Figure 3 Bacterial resistance mechanisms to antibiotics and the mechanisms of AMP in bacteria. The majority of AMPs demonstrated bactericidal activity against microbial cells by destabilising the plasma membrane, regardless of the presence of potential antibiotic resistance mechanisms. Bacteria can develop resistance to AMPs when exposed to sub-lethal doses, which are subsequently removed by highly effective efflux pumps. Membrane interactions significantly influence the direct antimicrobial efficacy of AMPs. There are four antimicrobial peptide membrane models. Specifically, these models include the barrel-stave, toroidal-pore, and carpet models. Also, four key molecular strategies that enable bacterial survival against antibiotic treatments are shown in this figure. These include changes in drug targets, efflux pumps, and antibiotic enzymes that degrade or alter antibiotics. When these resistance mechanisms occur together in one bacterium, it results in high resistance to diverse antibiotics. |
Intrinsic Resistance
Numerous examples have shown that innate AMP resistance exists. AMP hyper-susceptibility has been found in many organisms using various mutagenesis techniques. Bacteria commonly develop AMP resistance through cell wall or outer membrane modifications that reduce permeability or the net negative charge. This will likely minimise the attraction of AMPs to these, therefore hindering interaction or absorption. Deletion of these genes will increase AMP susceptibility and decrease virulence in mouse models. While these modifications have been observed in many Bacillus species, it is important to note that multiple genes encode them. For example, the dltABCD gene products attach the positively charged D-alanine to anionic teichoic acids within the cell walls of Gram-positive bacteria. S. aureus has the MprF enzyme, which modifies phosphatidylglycerol with D-lysine and is the only known positively charged phospholipid.151,152 Furthermore, many Gram-negative bacterial species modify the outer membrane LPS molecules with aminoarabinose. The PmrA-PmrB two-component pathway controls many genes in this process. The two-component system, the PhoP-PhoQ system, well characterised in Salmonella, detects environmental Mg2+ levels and regulates various virulence and AMP-resistance genes. The pagP gene adds an acyl chain to lipid A of LPS, reducing membrane permeability.119,121
Acquired Resistance
The body of research on the ingenuity of bacteria in acquiring increased AMP resistance and its implications for physiology and virulence is limited; however, we will provide a compilation of studies on this topic. A team of researchers conducted a significant laboratory experiment demonstrating that bacteria developed high resistance to an AMP in a fully controlled in vitro laboratory environment. They escalated exposure to the pharmaceutical AMP pexiganan (a synthetic magainin analogue) in populations of P. fluorescens and E. coli. The authors found that most bacterial lineages (22 of 24 populations) developed high resistance to the AMP after approximately 600–700 bacterial generations, regardless of the mutation type. The resistance was found to be stable, and pexiganan did not affect log-phase bacterial growth. A complete fitness assessment regarding resistance was not conducted; therefore, the specific resistance determinants were not assessed. Another study produced 20 Group A Streptococcus isolates that showed greater resistance to cathelicidin peptides using a combination of transposon mutagenesis and repeated exposure to the peptide. Through these techniques, they isolated a resistant clone, more closely analysed it, and found that a gene was disrupted, with similarity to GntR regulators.154
After subcutaneous inoculation, this strain exhibited increased virulence, producing larger, more persistent lesions than the parent strain. Another example describes an S. aureus mutant that showed increased tPMP resistance due to a transposon insertion mutation. The transposon was found in the snoD (mnhD) gene, which encodes the complex I NADH: ubiquinone oxidoreductase. When the gene was disrupted, membrane fluidity increased, transmembrane potential decreased, and mutant survival increased in rabbits. Moreover, tPMP-resistant strains emerged from serial exposure to low doses of the peptides.154,155 A fourth example demonstrated that E. coli developed resistance to specific proline-rich AMPs through mutations in the sbmA gene, leading to decreased peptide uptake. Spontaneous SbmA mutants of Salmonella typhimurium exhibited increased AMP-resistance from the same cause. Mutations that allow constitutive expression of resistance gene homologs can confer resistance, as in Haemophilus influenzae, which develops increased cationic drug resistance (eg., colistin, LL-37) from specific mutations in phoP-phoQ or pmrA-pmrB. Interestingly, constitutive expression of phosphorylcholine, resulting from the action of LPS-oligosaccharides, greatly increases resistance to LL-37.154,156
All approaches to combating AMR are fundamentally based on infection prevention, stewardship initiatives, or novel therapies like AMPs. The use of ML involves using WGS data to generate predictions of resistance patterns, enabling preemptive policy intervention. AMPs and ML represent a glimmer of hope for developing long-term solutions to AMR; however, ultimately addressing AMR requires a multidisciplinary approach.
Machine Learning Approaches for AMP Discovery
The application of machine learning to antimicrobial peptide discovery has evolved substantially over the past decade, progressing from simple binary classifiers based on hand-engineered features to sophisticated DL architectures capable of generating entirely novel peptide sequences with predicted therapeutic properties. This section provides a systematic review of the major ML approaches employed in AMP research, organised by methodological paradigm. We examine classical machine learning methods, DL architectures, protein language models, geometric DL, and generative models, analysing the strengths, limitations, and appropriate use cases for each approach. Figure 4.
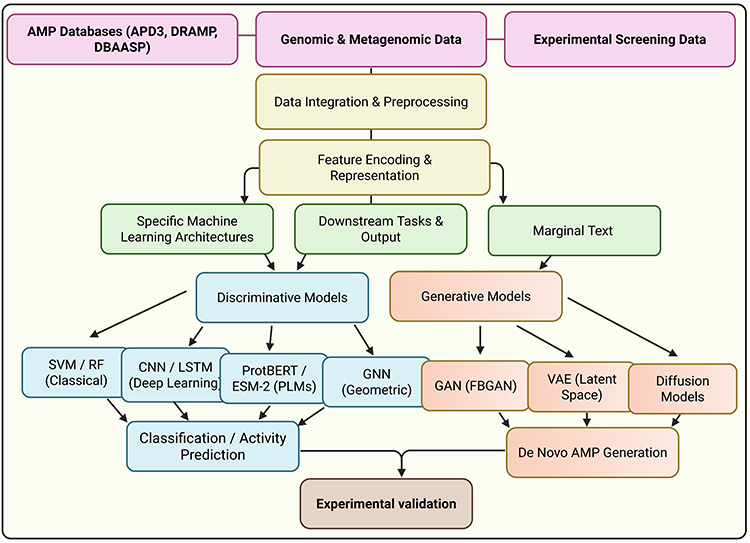 |
Figure 4 End-to-end machine learning pipeline for antimicrobial peptide discovery. |
Classical Machine Learning Approaches
Classical machine learning techniques, including support vector machine (SVM), random forest (RF), k-nearest neighbour (k-NN), logistic regression (LR), and gradient boosting algorithms (XGBoost, LightGBM), have formed the prevailing approach towards AMP prediction during the early era of computational AMP studies (around 2010–2018) and remain useful approaches for particular purposes.157 In general, classical algorithms process information encoded as peptide feature vectors, thereby being easy to understand and computationally efficient.157
Support vector machines employ kernel transformations to project input features into a higher-dimensional space where linearly separable decision boundaries emerge.158 The kernel transformation used (linear, polynomial, radial basis function, sigmoid) greatly affects the model’s performance; the RBF kernel has proven more efficient for predicting AMPs due to the flexibility it offers in generating decision boundaries. AMP prediction algorithms based on SVMs include the aforementioned AntiBP2, iAMP-2L, and CAMP-SVM models; these techniques have demonstrated the feasibility of AMP prediction, achieving accuracy of 75–85% on early benchmarks.159 On the other hand, SVMs are constrained by computational complexity (quadratic training time relative to the data volume), hyperparameter sensitivity, and difficulty handling unbounded-length sequences.160
Random forests ensemble decision tree learners using bootstrap sampling techniques and randomly selected features to provide reliable results requiring limited hyperparameter tuning.161 RF-based classifiers have shown significant promise for AMP prediction, with models such as AMPEP, CAMP-RF, and AMPScanner achieving promising accuracy levels.162 The built-in feature-scoring mechanism of random forests provides useful interpretability for identifying the physicochemical properties that best predict antimicrobial effects.163 The use of random forests is generally less prone to overfitting than that of decision trees alone, and they handle imbalanced classes reasonably well by employing weight adjustment or balancing techniques. Like SVM classifiers, random forests rely on predetermined feature vectors.
Gradient boosting algorithms, such as XGBoost, LightGBM, and CatBoost, have consistently delivered high-quality results across multiple AMP prediction tasks.164 These algorithms build a collection of weak learners, such as shallow decision trees, where subsequent learners are designed to address the mistakes made by their predecessors. Gradient boosting classifier methods, including XGB-AMP and CatBoost-AMP, have achieved accuracies above 90%, outperforming or matching many competing DL algorithms, especially when only a small number of training examples are available.165 Using gradient boosting combined with engineered feature sets based on physicochemical properties and evolutionary information has shown particular success.
Traditional ML methods (ie., machine learning methods) are hindered in their ability to predict AMP due to their need for handcrafted (or manually created) feature representations (eg., input data, characteristics of the data, etc).166 Developing optimal features for AMP prediction requires substantial domain expertise. As a result, using a handcrafted feature representation may unintentionally eliminate information relevant to predicting AMP activity that is very difficult to encode manually. Classical ML methods cannot adequately model higher-order interactions among amino acid residues that may be important for membrane targeting. However, traditional ML practices still have advantages: they are computationally efficient, require smaller training datasets than deep learning techniques, allow for interpretable models via feature importance analysis, and consistently perform competitively when implemented with high-quality handcrafted features.167 Therefore, traditional ML practices are generally considered an effective and viable solution for researchers with limited computational resources or small training datasets.
Deep Learning Architectures
Deep learning algorithms have emerged as the dominant paradigm for predicting AMPs because of their ability to automatically learn hierarchical representations of input sequences, without any need for manual feature selection or extraction.168 There have been three main classes of DL frameworks used in AMP prediction applications: convolutional neural networks (CNNs), recurrent neural networks (RNNs), and transformers that utilise attention mechanisms. CNNs use sliding-window operations on one-hot or embedding-encoded AMP sequences to detect local motifs responsible for antimicrobial activity.169 First-layer convolutional filters identify simple motifs, such as clusters of positively charged amino acids or hydrophobic segments; deeper layers combine these basic motifs to form more complex structure-function associations.170 Several AMP predictor models based on CNNs, such as Deep-AMPEP30, AMPScannerV2, and ACP-DL, achieve AUC-ROC values greater than 0.90 on benchmark datasets.171 The translation invariance of convolutional operations makes CNNs a good choice for detecting motifs regardless of their location within a sequence. However, conventional CNNs struggle to model long-range residue interactions, which might be important for maintaining secondary structures required for AMP membrane insertion.172
RNNs and their variants, such as LSTM and BiLSTM networks, sequentially operate on input sequences, storing information of previous residues through their internal state vectors.173 Such a framework is especially well-suited to modelling sequential dependencies, ie., how AMP activity emerges from the sequential arrangement of amino acids. BiLSTM networks operate on input sequences in two directions, allowing the network to use sequence information from previous and subsequent positions to predict each position in the sequence.174 RNNs have been successfully used to build several AMP predictors, such as BiLSTM-AMP and deep-AMPpred, which outperform other sequence encoding algorithms by virtue of the bidirectional sequence processing capabilities of BiLSTM models.175 GRU networks offer an alternative to LSTMs with improved efficiency without compromising performance.176
One such limitation is the ability of attention models to overcome the challenges posed by modelling dependencies, using RNNs and the fixed receptive field in CNNs. Attention methods dynamically emphasise important positions in the sequence to make accurate predictions. The key characteristic of self-attention is the calculation of weighted associations between all pairs of positions in the sequence, allowing the method to capture dependencies regardless of their distance. Architectures using only self-attention, including transformers – purely feed-forward neural networks that do not have any recurrent or convolutional elements – demonstrate impressive results in natural language processing and can be applied for bioinformatics purposes.177 Transformer-based AMP predictors treat peptides as sequences of tokens, where each amino acid corresponds to an embedding vector learned during training. Self-attention layers calculate association scores between each pair of positions, allowing detection of key residue-residue contacts responsible for antimicrobial action. Transformer-based AMP predictors achieve state-of-the-art results on the benchmark dataset, with AUC exceeding 0.95.178,179 dsAMP represents an approach to designing AMP predictors by combining CNN layers for local feature extraction with attention-augmented BiLSTM layers for sequential representation, achieving over 95% accuracy through transfer learning on small datasets.180 Another example of a successful AMP predictor is the AMP-EBiLSTM framework, which utilises embedding layers, bidirectional LSTM networks, and attention mechanisms.181 Figure 5.
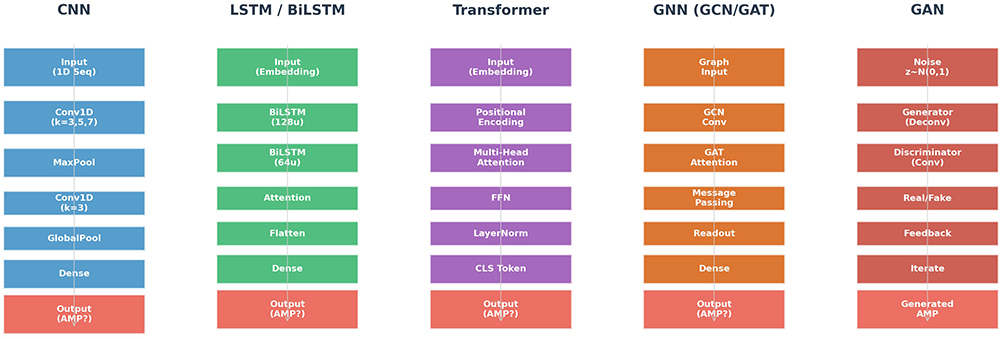 |
Figure 5 Comparative DL architectures for AMP prediction. |
Protein Language Models and Transfer Learning
Protein language models (PLMs) represent a paradigm shift in computational biology, leveraging techniques from natural language processing to learn rich representations of protein sequences through self-supervised pre-training on massive sequence databases.182 Unlike task-specific models trained from scratch on limited AMP datasets, PLMs are pre-trained on millions of protein sequences from UniProt, Pfam, and other resources, using objectives such as masked language modelling (predicting masked amino acids from context) and next-token prediction. This pre-training enables PLMs to capture evolutionary patterns, structural constraints, and functional relationships embedded in protein sequence data, generating contextualised embeddings that serve as powerful input features for downstream tasks.183
ProtBERT, a BERT-based model pre-trained on over 2 billion protein sequences, generates 1024-dimensional contextualised embeddings for each amino acid position.184 When fine-tuned for AMP prediction, ProtBERT achieves AUC values exceeding 0.95 on multiple benchmark datasets, substantially outperforming models trained from scratch with limited training data.185 The self-attention mechanism in ProtBERT enables the model to capture long-range interactions between residues, including contacts that stabilise antimicrobial-relevant secondary structures.
ESM-2 (Evolutionary Scale Model 2), developed by Meta AI, is one of the most powerful protein language models, trained on 65 million protein sequences with the masked language modelling objective.186 The ESM-2 architecture family includes various models with 8 million to 15 billion parameters; the larger models provide better representations. For AMP prediction, ESM-2_650M (with 650 million parameters) provides a good trade-off between representation quality and computational cost, achieving about 0.96 AUC on AMP benchmarks.187 ESM-2 produces representations that include evolutionary, structural, and functional features and enables zero-shot predictions of protein characteristics without task-specific fine-tuning.188
The ProtT5 model uses the text-based T5 (Text-to-Text Transfer Transformer) architecture, treating protein sequences as text and training it to restore corrupted sequences using an encoder–decoder paradigm.189 It was shown that ProtT5 representations can be used for the AMP prediction problem, as fine-tuning ProtT5 models on AMP data using transfer learning achieves state-of-the-art results.190 One of the main benefits of ProtT5 is its encoder–decoder architecture, which enables it to be applied not only to sequence classification but also to sequence generation tasks. The use of transfer learning strategies to predict AMP typically involves one of two methods: feature extraction or fine-tuning the pretrained language model (PLM). When employing the feature extraction method, peptide sequences are fed into the pre-trained PLM, which generates embeddings that serve as input to downstream classifiers (eg., logistic regression, gradient boosting), but the PLM’s parameters are not modified.191 In contrast, with the fine-tuning method, the pre-trained PLM is modified by adding task-specific layers, and both the new and old layers are trained on AMP-specific datasets, allowing the new representations to adapt to the idiosyncratic representations relevant to antimicrobial activity. Fine-tuning usually has higher accuracy than feature extraction, but also has a higher requirement for training data and computational resources; whereas, feature extraction can achieve very good performance without requiring as large an amount of training data, making it a more efficient method of extracting relevant features from the data than the fine-tuning method.192
Geometric Deep Learning
Geometric DL models using graph neural networks (GNNs) offer an appealing approach to AMP prediction, particularly because they incorporate three-dimensional structural data.193 Sequence-based models, while assuming that antimicrobial behaviour is influenced solely by the primary amino acid sequence, overlook the crucial aspects of peptide folding and the formation of structures that enable membrane disruption and insertion.194 Peptide molecules used for prediction via GNNs can be represented as graphs, where each node corresponds to a specific amino acid residue, and edges correspond to either spatial proximity (a distance of 6–8 Å between heavy atoms) or chemical bonds.195 Each node represents not only an amino acid but also its physicochemical characteristics; edge features may include distance, bonding types, etc. Convolutional neural networks operating with graph data (GCNs) transfer information between nodes through message-passing algorithms.196
In this regard, the sAMPpred-GAT approach highlights the use of GNNs for AMP prediction, specifically graph attention networks (GATs) for recognising relevant spatial connections associated with antimicrobial activity.197 By incorporating sequence-based features (one-hot encoding, positional encoding, PSSM profiles) and structural features derived from contact map predictions, sAMPpred-GAT has been shown to significantly outperform sequence-based methods across eight independent test datasets using AUC, MCC, accuracy, sensitivity, and specificity metrics.198 AMPs-Net uses a graph representation of peptides, where nodes represent atoms and edges represent bonds, to apply graph convolutional networks with 20 layers of message passing to learn the graph representations and combining these graph embeddings with physicochemical property annotations results in improved precision by 8.80% to 19.02% and accuracy by 5.74% to 24.23% when compared with alternative DL techniques.198,199
LABAMPsGCN employs a heterogeneous graph model with nodes of amino acids, dipeptides, and tripeptides, with the edges representing co-occurrence information, resulting in accuracies for independent tests between 0.913 and 0.938. By using a heterogeneous graph, this model learns multi-scale sequence information at three levels: individual amino acid residues, dipeptides, and tripeptides, to incorporate sequence information for AMP discovery.200 One major challenge in applying GNNs to peptide prediction is their reliance on structural information, which can only be predicted when the structures have not yet been determined. While structures predicted by AlphaFold2 are accurate enough in most cases, there is still potential for error propagation due to incorrect structural predictions, especially for disordered peptides and novel folds. Another challenge with GNNs is the increased computational demand of graph construction and message passing. Despite these challenges, advancements in both structure prediction and computational capacity ensure that geometric DL will continue to grow.201,202
Generative Models for De Novo AMP Design
While discriminative models classify or score existing peptide sequences, generative models create entirely novel sequences with predicted antimicrobial properties, offering a powerful approach to exploring the vast sequence space of potential AMPs.203 Generative models learn the underlying distribution of antimicrobial peptide sequences and can sample new sequences from this learned distribution, potentially discovering peptides with improved properties that differ substantially from known sequences. Generative adversarial networks (GANs) consist of two competing neural networks: a generator that produces candidate peptide sequences and a discriminator that distinguishes between real (from the training data) and fake (generated) sequences.204 Through adversarial training, the generator progressively learns to produce increasingly realistic AMP-like sequences that can fool the discriminator. The Feedback GAN (FBGAN) framework integrates a property predictor into the GAN training loop, guiding generation toward sequences with predicted antimicrobial activity.205 The dsAMPGAN model extends this approach by incorporating multi-label classification for multiple antimicrobial activities (antibacterial, antifungal, antiviral) alongside a regression model for MIC prediction, generating peptides with predicted broad-spectrum activity.180
VAE encodes peptide sequences into a continuous latent space and then learns to reconstruct peptide sequences using a decoder network.206 By sampling points within the latent space and subsequently reconstructing, a VAE can generate novel peptide sequences through interpolation in the latent space.207 Since minor variations in latent vector values result in slight changes to generated peptides, VAEs allow optimisation of peptide properties via gradient ascent/descent in the latent space.208 The AMP Generator represents a Variational autoencoder (VAE) trained on AMP peptide sequences, enabling interpolations between peptide types belonging to different AMP families and, hence, generating peptides with intermediate properties.206 Modern approaches to VAEs include combining VAE training with various optimisation strategies, such as Bayesian Optimisation or Genetic Algorithms, to discover peptides with optimal activities and safety properties.206
Diffusion models are the latest trend in machine learning for AMP peptide design, gradually applying noise to peptide sequences and learning to reverse the process for generating peptides.209 AMP-Diffusion is a framework that relies on a latent diffusion model and protein language models to generate diverse AMP peptides, allowing decoupling of sequence generation from peptide property control and providing users with the flexibility to tune the length and diversity of AMPs.210 While diffusion models still trail behind both GANs and VAEs in terms of stability and generated peptide quality, this type of model is expected to dominate AMP peptide design soon. As mentioned above, one approach to peptide property optimisation involves reinforcement learning, in which an RL algorithm learns to construct a peptide amino acid by amino acid, receiving a reward based on predicted peptide activity and properties.211 RL methods allow encoding multiple objectives (eg., activity, toxicity, stability) into the reward function to generate sequences with specific target properties.211 MCTS combined with a neural network value function is another technique used for navigating the huge combinatorial space of peptides. Figure 6.
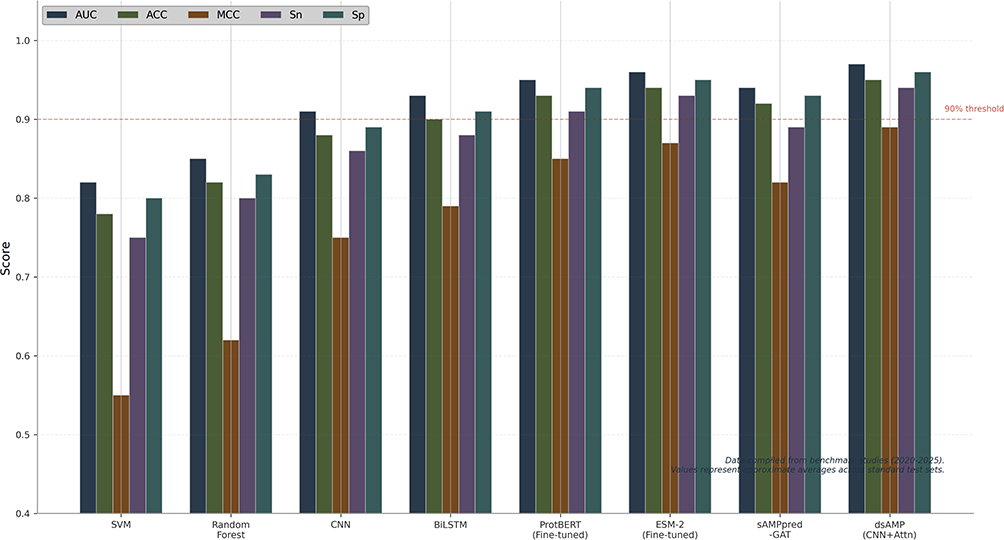 |
Figure 6 Performance benchmarking of ML methods for AMP prediction. |
Choosing the most suitable generative model depends on your design objectives. GANs can generate a wide variety of candidate sequences for experimental screening.206 VAEs can help you find intermediate sequences and optimise property values in the latent space. Diffusion models can generate very high-quality sequences while allowing precise control over specific characteristics.206 Reinforcement learning (RL) methods are ideal for optimising specific multi-objective property profiles. Once again, it is crucial, no matter which of these methods you choose to use to generate your new sequence(s), that you filter your new sequence(s) through computational means (ie. predicting activity, toxicity and stability) prior to synthesising them in order to increase the likelihood of finding a clinically relevant candidate from your new sequence generation.212
ML-Guided AMP Design and Optimisation
Activity Prediction Models
Activity prediction corresponds to one of the fundamental aims of computational AMP research, involving binary classification (separating AMPs from non-AMPs), multi-class classification (forecasting activity toward certain pathogen classes), and regression (quantitative predictions of minimum inhibitory concentration, MIC, for individual bacterial strains).213 All of the mentioned tasks pose different issues, which require appropriate evaluation criteria.
Binary classification involves assessing peptide sequences to determine whether a given sequence displays AMP activity, thereby reducing the problem to two-class discrimination.214 Despite its simplicity in principle, binary classification faces issues such as class imbalance (where positive samples are predominant in training sets) and the lack of negative samples (randomly generated peptides cannot be considered representative of inactive ones). Binary classification models can be evaluated using accuracy, sensitivity (true positive rate), specificity (true negative rate), precision, F1 score, Matthews correlation coefficient (MCC), and area under the ROC curve (AUC).215 Among the evaluation criteria, MCC is particularly valuable because it uses all four elements of confusion matrices.216 Recent advances in binary classifiers include fine-tuning the ESM-2 model and transformers, resulting in MCC values of 0.85–0.90 and AUC > 0.95.203
Activity prediction against specific bacterial species or strains constitutes a step beyond generalised antimicrobial activity prediction, which has practical application since different pathogens require differential treatment.217 It poses additional difficulties due to insufficient labelled data for each species and to intricate amino acid sequence motifs responsible for species-specificity. Transfer learning techniques that fine-tune a generalised AMP classifier on individual species datasets are successful. Notably, the dsAMP approach demonstrates that transfer learning can achieve more than 90% classification accuracy when predicting species-specific activity using small, species-specific datasets (up to 200 peptides) across the four bacteria, P. aeruginosa, E. coli, S. aureus, and S. pneumoniae.180 Binary VAE combined with quantum annealing has also been applied to species-specific optimisation, although it is too computationally intensive for practical use.218 Table 5.
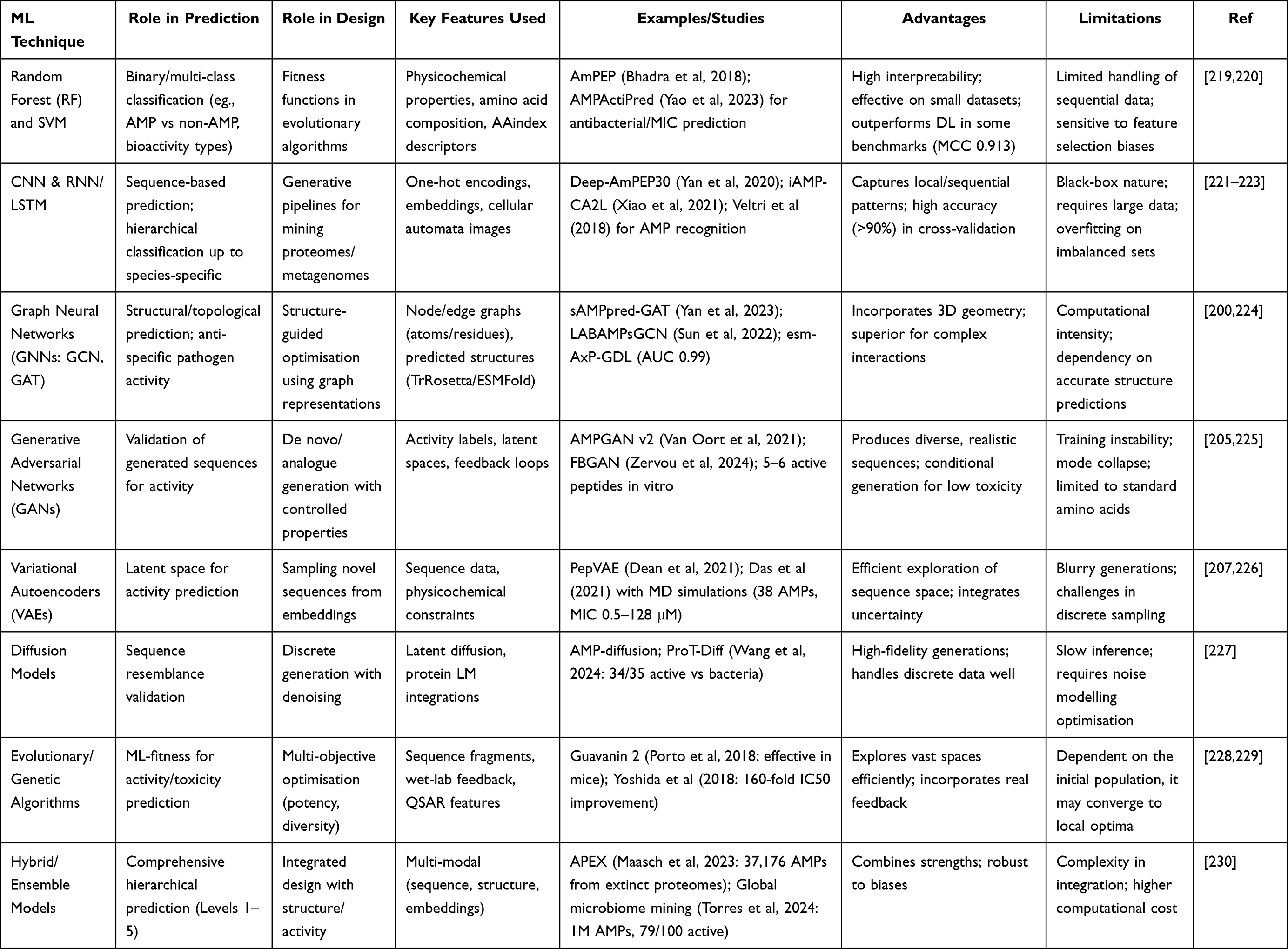 |
Table 5 ML in Predicting and Designing AMP |
The second type of quantitative predictive modelling concerns the estimation of the minimal inhibitory concentration (MIC), or the minimal amount of peptide required to inhibit bacterial growth to a certain degree. MIC regression provides clinically useful information, but it poses even greater challenges than binary classification. First, it demands precision, as rough estimates are not useful for drug discovery. Second, MIC values depend on the laboratory methods used to measure them and therefore exhibit great variability. BERT-AmPEP represents an innovative step toward MIC regression, fine-tuning a BERT classifier to predict MIC values for peptides targeting E. coli and S. aureus using self-attention-based encoding of long-range amino acid motifs.231 Deep learning MIC regression models that combine knowledge encoded in the physical and chemical properties of peptides with a learned representation achieve high Pearson correlation coefficients between predicted and experimentally measured MIC values (0.7–0.8). However, a small number of MIC datasets (a few hundred to a thousand peptides) limits the development of a higher-accuracy model.232,233
The classification of peptides based on their activity spectra is an extension of species-level spectrum prediction. It includes the classification of peptides that display an activity against either a narrow spectrum (active only against a certain type of Gram-positive or Gram-negative pathogens) or a broad spectrum (active against both types), or extended-spectrum (show activity against fungi, viruses, and parasites).234 Multi-label classification systems make simultaneous predictions of multiple activity labels to help identify peptides with appropriate spectral profiles. Recently, GNNs that model the evolutionary relationships among organisms targeted by these peptides have demonstrated promise for spectrum prediction, allowing the evolutionary relatedness of pathogens to be generalised across species in which they were predicted.212
Property Prediction and Multi-Objective Optimisation
In addition to antimicrobial activity, peptides must demonstrate acceptable safety profiles, stability, and pharmaceutical properties for successful clinical translation.235 Predictors for these key peptide characteristics have been developed using ML algorithms, enabling multi-objective optimisation of both the activity and safety of potential AMPs.
Hemolysis prediction models evaluate peptide hemolytic activity toward human erythrocytes, a critical parameter of AMP safety.236 Hemolytic activity is often positively correlated with antimicrobial activity because peptides share the ability to perturb membranes. However, the therapeutic potential of AMPs depends not on their activity but on the therapeutic index, defined as the ratio of the hemolytic concentration to the MIC. Data on hemolytic activity can be found in the HemoPI and DBAASP databases, together with activity information,41 which allows the development of specialised hemolysis predictors. In particular, DL architectures based on multi-head cross-attention-based feature fusion, such as HemoFuse, achieved predictive performance of >90% for predicting peptide hemolytic activity.237
Prediction of cytotoxicity and immunogenicity helps evaluate broader safety concerns beyond hemolysis.238 The cytotoxic activity toward mammalian cell lines (HEK293, HepG2) determines the maximal tolerated dose of peptides, while immunogenicity associated with activation of adaptive immunity precludes their further use. Cell viability data used to train cytotoxicity predictors are less abundant than hemolysis data, which hampers the availability of public training data sets.239 Prediction of immunogenicity was traditionally performed by predicting MHC binding affinity using NetMHC and NetMHCpan.240
Stability prediction provides information on the sensitivity of peptides to enzymatic digestion, which helps estimate plasma half-life and oral bioavailability.241 Cleavage site predictors of proteases provide insight into amino acid positions on a peptide chain that may serve as cleavage sites of host proteases such as trypsin, chymotrypsin, pepsin, or bacterial proteases. Predicted cleavage sites can be eliminated through mutations to produce stable peptides.242 Other methods to enhance peptide stability include cyclisation (head-to-head and head-to-tail, or via disulfide linkages), substitution of D-amino acids, and incorporation of non-canonical amino acids; their impact on stability can be assessed computationally using structural modelling.243
The multi-objective optimisation framework considers several goals, including optimising antimicrobial activity, minimising potential toxicity, maximising stability, and optimising physicochemical properties.244 The problem can be posed either as a constrained optimisation problem, where antimicrobial activity is maximised while satisfying constraints on toxicity, stability, and other properties, or as a Pareto optimisation problem, which describes all possible solutions that represent an optimal balance between the objectives.245 A DL generative model that incorporates predictors of multiple properties into the loss function can simultaneously optimise multiple properties during sequence generation.246 Bayesian optimisation techniques applied in the latent space of variational autoencoders (VAE) have proven successful in handling multi-objective problems and producing sequences with optimised compromises between activities and safety.247
Structure-Activity Relationship Modelling
Insights into the mechanisms by which peptide structure determines antimicrobial activity are important for the rational development of AMPs.248 SAR studies analyse structural properties such as the presence of specific secondary structures, hydrophobic moment, charge distribution, and amphipathy that correlate with activity and the spectrum of action.
The alpha-helix is the largest group among AMPs that have been extensively studied and form amphipathic helices in a membrane-like environment, allowing membrane binding and insertion.249 The hydrophobic moment, which indicates the separation of hydrophobic and hydrophilic amino acids along two sides of the helix, is an important property determining the ability of peptides to bind to the membrane and antimicrobial activity.250 Computational models that predict helical structure and the hydrophobic moment from sequence are widely used to optimise peptide amphipathy. QSAR models based on molecular descriptors calculated from three-dimensional peptide structures have yielded successful results in predicting the activity of alpha-helical peptides. However, their accuracy is limited by the requirement for reliable structure prediction.251
Alternative secondary structures, such as beta-sheets and cyclic AMPs, exhibit unique strengths. Beta-sheet structures held together by disulfide bridges (defensins) and cyclic backbones (bacteriocins) display increased proteolytic stability, and the latter can achieve high selectivity with specific binding surfaces.252 Designing AMPs using computational approaches such as molecular docking and molecular dynamics simulation is possible by identifying optimal replacements for amino acids.252 ML-assisted molecular dynamics utilises simulation data to learn force-field corrections or better sampling techniques, either.253 3D-QSAR extends QSAR by leveraging structural alignment and the use of fields, namely electrostatic, steric, and hydrophobic. CoMFA and CoMSIA have been used on the AMP series with a common structural core. However, the application of this method has a limitation, as training molecules must be structurally related, thereby limiting sufficient diversity among sequences.254,255
Integration of Experimental Validation
Predictions generated by computational methods, however advanced, must always be confirmed experimentally. Incorporating experimental results into the iterative design process is an integral component of AMP discovery via machine learning, enabling the model to learn from real-world data and refine its predictive capabilities.256
Active learning, also known as optimal experimental design, selects the most valuable candidates for experimentation to extract the maximum information from each trial.257 In the context of AMP discovery, active learning strategies choose the most informative sequences in terms of uncertainty (ie., those that are most unpredictable according to the model; uncertainty sampling) or those whose properties will provide the greatest benefit to the model’s performance; query-by-committee).258 Active learning approaches have been shown to reduce the number of required experiments to identify active peptides by 50–80% compared to random screening.259
In closed-loop discovery, the iterative process between design and experiment becomes fully automated.260 In this type of system, machine learning models generate sequence candidates, which are then synthesised and screened by automated laboratory processes (eg., liquid-handling robotics and high-throughput screening systems).185 The information gathered from the experiments can be incorporated into further model refinement. Companies such as Recursion Pharmaceuticals and Insilico Medicine have already proven closed-loop drug discovery for small molecules, while similar techniques have been applied for peptides.261 One key problem with closed-loop discovery is time – usually, computational design takes from hours to days, while synthesis and tests span from days to weeks.261 Testing and validation are done through successive tiers of tests.262 The first in silico filtration tests the prediction of the activity, toxicity and stability of potential peptides. Those that do not appear in any of the parameters mentioned proceed to further in vitro tests. These include MIC tests for activity, hemolysis tests for toxicity evaluation, and protease resistance test for pharmacokinetics. Potential candidates which passed in vitro screening will go on to animal trials for further testing of their biological effects.263 This method ensures efficient spending since the costly in vivo experimentation is undertaken only for the most promising sequences.248
Animal Model Studies
In vivo Efficacy Studies
Infection models in animals are a critical stepping stone between in vitro assessments of antimicrobial activity and subsequent clinical trials, as they demonstrate whether peptides that exhibit significant antimicrobial activity in vitro retain it in the complex environment of a living organism.264 There are numerous infection models used to examine the efficacy of AMPs, each with strengths and weaknesses specific to the model. Murine sepsis models remain the most common strategy for investigating systemic AMP efficacy.265 Such models involve infecting mice with the target bacteria via intraperitoneal injection, followed by treatment of the infection with the AMP of interest, either prophylactically (prior to infection) or therapeutically (afterwards). Evaluation endpoints include survival rates and bacterial load in the blood, peritoneum, or organs. A murine sepsis model is a highly challenging evaluation of AMP efficacy, as systemic infection requires AMP delivery to achieve therapeutic levels in both the bloodstream and other tissues, along with being able to function despite interference from serum proteins and lipids, as well as numerous host defence molecules capable of neutralising the activity of cationic peptides.266 Several ML-designed peptides have been shown to exhibit therapeutic efficacy in murine sepsis models, delivering survival rates ranging from 60 to 80% against multidrug-resistant pathogens, compared to 0 to 20% in untreated groups.267
Models of wound infection represent another widely used animal infection model assessing the efficiency of AMPs in localised tissue infections.268 Such models include creating full-thickness skin ulcers on the dorsal side of a mouse or rat, inoculating the ulcers with the target pathogens, and applying an AMP topically or systemically, followed by analysis of bacterial load in the tissue after the specified treatment period. Wound infection models are particularly relevant for AMPs designed for topical application (eg., wound, burn, and surgical treatments), as they also provide information on local tissue distribution and biofilm penetration.269 Numerous investigations into the activity of ML-optimised peptides have confirmed their efficacy in reducing bacterial colonisation in wound infections and in outperforming untreated controls and conventional antibiotic groups.270
Murine pneumonia models are used in testing the effectiveness of AMPs in combating respiratory infections.271 Bacterial inoculation in this model involves either intranasal instillation or aerosol exposure, while peptide administration is performed intranasally, intravenously, or by nebulization. Parameters measured in this model include bacterial colonisation of the lungs and bronchoalveolar lavage (BAL) fluid, as well as lung histopathology analysis. This pneumonia model is essential in evaluating AMPs as treatments against the pathogenic bacteria causing respiratory infections, including Pseudomonas aeruginosa, Klebsiella pneumoniae, and A. baumannii – the top causes of ventilator-associated pneumonia in the ICU.272
Zebrafish infection models represent an excellent tool, in addition to mammalian models, as an alternative to animal testing for toxicity and efficacy screening during early stages of development.273 The zebrafish innate immune system shows a high degree of similarity to the mammalian innate immune system; moreover, the optical clarity of zebrafish larvae provides an opportunity to visualise and observe infection dynamics and immune responses in action. This kind of screening uses relatively small quantities of compounds (in the nanogram to microgram range), making it appropriate when peptide amounts are insufficient for evaluation.274 Using this model, machine-learning-designed peptides exhibited their effectiveness against S. aureus and Escherichia coli, and interesting correlations between zebrafish and mouse data were observed.275
Research on how AMP’s behaviour in living systems is measured through Pharmacokinetic studies in animals, and this information is used to develop the key components (ADME) of AMP use.276 Blood is collected following injection (intravenous, subcutaneous, or intraperitoneal) at specific time points, and the concentrations of the AMPs are then measured using methods such as LC-MS/MS or ELISA. The pharmacokinetic components measured are the maximum plasma concentration (Cmax), time to achieve the maximum plasma concentration (Tmax), area under the plasma concentration-time curve (AUC), elimination half-life (t1/2), clearance (CL), and volume distribution (Vd).277 The majority of natural AMPs are found to have very short half-lives in blood (minutes to hours). This is caused by the breakdown of AMP (through proteolysis) and by its clearance from the body (through the kidneys). Therefore, calcium stabilisation is done by using various methods to improve circulation time, including structural modifications (adding a ring, changing D-amino acids, or adding PEG to amplified peptides) and by using different formulations.278
ML-Designed AMPs Validated in Animal Models
Multiple recent studies have shown that peptide designs optimised using ML techniques can achieve notable efficacy in animal infection models, providing proof-of-principle evidence that peptides can be designed computationally.267 For example, the breakthrough work by Wang et al (2025) used an explainable DL approach, combined with virtual evolution, to identify AMPs effective against MDR human pathogens.267 Training a DL model on an initial dataset of AMPs, it was predicting their activity against pathogens and evolving AMP sequences using optimisation algorithms to increase their ability to combat specific MDR pathogens. Finally, the designed peptides were tested in murine infection models, showing activity equivalent to or higher than that of antibiotics against carbapenem-resistant A. baumannii and colistin-resistant P. aeruginosa. Importantly, the authors demonstrated that the ML-designed peptides were nontoxic to the animals at therapeutic concentrations and did not induce resistance to the treatment when passed through multiple generations.267
Similarly, the rapid discovery of antimicrobials by Das et al used generative ML models along with molecular dynamics simulations to generate AMPs and subsequently synthesised and tested these peptides.279 Namely, the generative ML model suggested new sequences, molecular dynamics simulations predicted binding conformations with pathogen membranes, whereas high-throughput synthesis and testing confirmed the antiviral activity of the peptides. Further experiments involving promising peptides included testing their activity in murine sepsis models; specifically, several peptides demonstrated a dose-dependent reduction in MRSA infections, and one candidate achieved 80% survival at 10 mg/kg, compared with only 10% in controls.279
Deep learning was used by Torres et al to identify antibiotics with peptide structures from archaeal proteomes, thereby opening unexplored regions of sequence space for novel antimicrobial sequences.280 The predicted peptides were synthesised and tested in infection mouse models using Enterococcus faecium and S. aureus, in which multiple peptides exhibited significant decreases in bacterial load in peritoneal wash and spleen tissue samples. This study highlights the potential of machine learning-guided genomic mining to generate novel AMP from sources beyond the usual suspects.280 Several studies comparing peptides generated by machine learning algorithms to those generated by traditional or natural methods indicate that ML-guided optimisation improves the therapeutic index (ratio of efficacy to toxicity), likely due to sequence modifications that boost antimicrobial activity without increasing hemolytic activity.21 This increased efficiency is attributable to machine learning algorithms’ superior ability to navigate complex multidimensional sequence spaces.
Toxicity and Safety Assessment
Comprehensive toxicity assessment is essential before advancing AMP candidates to human clinical trials.281 While computational toxicity prediction models provide initial filtering, animal studies are required to characterise dose-limiting toxicities and establish safety margins. Hemolysis and cytotoxicity in animal models extend beyond in vitro measurements by assessing toxicity in the context of systemic circulation and tissue distribution.282 Acute toxicity studies in mice involve single-dose administration escalating to identify the maximum tolerated dose (MTD) and lethal dose (LD50). Most ML-designed AMPs that pass initial in silico toxicity filters demonstrate acceptable safety profiles in animal studies, though unexpected toxicities (hypersensitivity reactions, complement activation) occasionally emerge that were not predicted by computational models.283
Immunogenicity assessment evaluates whether repeated administration of the peptide triggers adaptive immune responses that could limit clinical utility.278 Even though most AMPs are too short to function as T-cell-dependent antigens, they may bind to MHC molecules and elicit T-cell responses or form aggregates that activate B cells. Animal immunogenicity studies measure anti-peptide antibody titers following repeated dosing and assess hypersensitivity reactions.284 Studying the development of resistance focuses on the ability of pathogens to rapidly acquire resistance to ML-designed AMPs under selective pressure. Serial passage studies subject bacteria to subinhibitory concentrations of peptides over many generations, with MIC changes monitored over time. One of the primary benefits of many AMPs (particularly those that act through membrane disruption) is that, compared to conventional antibiotics, they have a relatively low potential to promote the development of resistant bacterial strains.285–287 Studies have shown that ML-designed AMPs that disrupt membranes exhibit unchanging MIC values even after 20–30 serial passages, whereas control antibiotics exhibit rapid increases in MIC.282 This property of ML-designed AMPs is critical for clinical applications where the therapeutic regimen must be maintained for a long period. Figure 7.
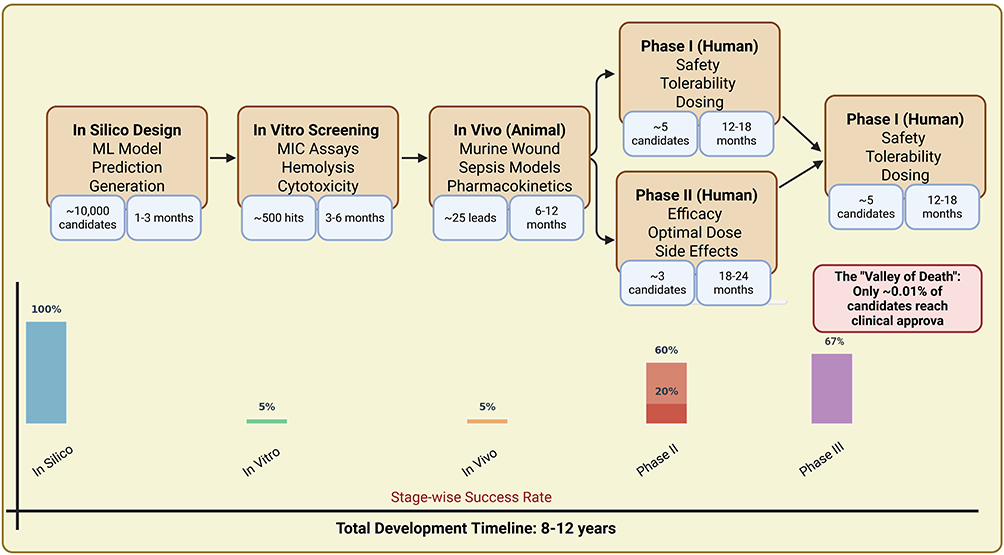 |
Figure 7 Complete translational pipeline from computational design through clinical trials. |
Human Clinical Trials
Approved AMP-Based Therapeutics
Even with decades of research on and identification of AMP, very few AMP-based drugs have received regulatory approval for use.282 The lag between the large number of identified AMPs and the small number of FDA-approved drugs can be attributed to significant hurdles in developing AMPs for human use.
The most advanced AMP drug to date was Pexiganan (MSI-78). It was a 22-amino-acid analogue of magainin, found in Xenopus laevis. It was developed as a topically applied antibiotic for the treatment of infected diabetic foot ulcers. Pexiganan demonstrated broad-spectrum activity against Gram-positive and Gram-negative bacteria in preclinical testing. In Phase III testing, however, it failed to show superiority over ofloxacin as a treatment option. As a result, the FDA issued a not-approvable letter because it did not demonstrate efficacy or safety in humans.288 Although it did not receive FDA approval as an antibiotic, it still has a role as a topical antiseptic, especially after a lawsuit settlement between Bayer and Novartis.252
Another lantibiotic in use today is Nisin. It is a 34-amino acid antibacterial peptide produced by Lactococcus lactis. It has been used safely as a food preservative since the late nineteenth century in many countries, including the United States (FDA) and the European Union (EFSA).289 Even though Nisin has not been approved for use as a drug, there is scientific literature on its activity against various cancer cells and pathogens.290 It is under clinical trial for the treatment of squamous cell carcinoma of the mouth, where it serves as both an antimicrobial and an anticancer agent. Gramicidin, a mixture of pentadecapeptides produced by Bacillus brevis, was among the first AMPs used clinically and remains available as a topical antibiotic in combination products (often with polymyxin B and neomycin).291 However, its narrow spectrum (primarily Gram-positive), mammalian cell toxicity, and limited topical use illustrate the challenges that have constrained broader clinical deployment of early AMPs.
Antimicrobial Peptides in Clinical Development
Several AMP-based drugs are at various stages of clinical development and stand out as the most promising for upcoming entry into clinical practice.292 hLF1-11 is a synthetic AMP isolated from the N-terminus of human lactoferrin exhibiting broad-spectrum antibacterial properties towards Gram-positive and Gram-negative microorganisms, as well as fungal species such as those causing infections by MRSA, Klebsiella pneumoniae, and Listeria monocytogenes.293 Of note, the drug shows minimal in vitro antimicrobial activity under physiologic conditions but demonstrates high efficacy in vivo and might exert immunomodulatory effects in addition to its direct anti-infective mechanisms.294 In particular, hLF1-11 stimulates monocyte differentiation and cytokine release, promoting innate host immunity to infectious agents.295 Some Phase I and Phase II clinical trials have already been performed, demonstrating favourable safety and tolerability profiles, with ongoing evaluation of clinical efficacy in specific infection indications.
EA-230 is a synthetic AMP consisting of a linear tetrapeptide derived from human chorionic gonadotropin, which acts predominantly as an immunomodulatory substance without significant direct antimicrobial effects.296 EA-230 demonstrates renoprotective properties in animal experiments and shows a favourable pharmacological profile in Phase I and Phase II clinical trials.297 This AMP is not considered a typical antimicrobial agent but rather represents other AMP-based compounds that exert immunomodulatory effects in infected hosts, particularly important for immunosuppressed individuals.
C16G2, a specific antimicrobial peptide (STAMP) from C3 Jian Inc., targets Streptococcus mutans, the primary causative agent of dental caries.297 This peptide incorporates a targeting domain that binds to S. mutans, in conjunction with the killing domain derived from the antimicrobial peptide novispirin G10.298 It allows for the specific killing of S. mutans while leaving other oral microbes intact. C16G2 has received FDA approval to proceed with human clinical trials for caries prevention, based on promising data from Phase II testing in healthy volunteers.297
Novexatin(R) (NP213) is a newly developed antifungal peptide designed based on host defence peptide structure templates.299 NP213 is indicated for topical use in the treatment of onychomycosis.37 Two Phase IIa human trials showed the agent’s effectiveness in treating fungal nails and demonstrated a safer side-effect profile compared to oral antifungals, which can cause liver damage or interact with concomitantly taken drugs. As such, NP213 appears superior to existing options due to limited absorption.
Challenges in Clinical Translation
The clinical translation of AMPs from preclinical development to commercial products faces several challenges that increase the risk of high attrition rates during clinical development.300 The high cost and scalability associated with production remain two of the major challenges to commercial success.301 The cost of solid-phase peptide synthesis is considerably higher than that of producing antibiotics, which may range from hundreds to thousands of dollars per gram depending on the peptide’s length and complexity. Recombinant expression using bacteria and yeast provides a more cost-effective method of producing longer peptides; however, issues of expression, purification, and folding can be problematic.302 To compete with cheap generic antibiotics available at a few cents per unit dosage, the cost of producing AMPs needs to be reduced by optimising synthesis processes or adopting new technologies.303
Issues surrounding bioavailability and mode of delivery limit the possible methods of administration for most AMPs.304 Proteolytic breakdown of the AMPs in the GI tract limits their potential for oral administration, thereby restricting their delivery route to either parenteral injection (intravenous, intramuscular, subcutaneous) or topical application. In the case of parenteral injection, formulation must ensure sterility; parenteral injection will often require hospital administration, thus restricting outpatient use.305 Various strategies being explored include liposomal delivery, polymer nanoparticles, hydrogels, and modification (eg., PEGylation, cyclisation, lipidation).306
AMPs face additional regulatory hurdles that complicate their development. In contrast to small molecules, which have established regulatory procedures, peptide drugs require validation of immunogenicity, consistency of manufacturing processes (including critical quality attributes such as purity, impurities, and aggregation state), and stability data that may differ from those expected for ordinary antibiotics.307,308 The lack of precedent for regulatory approval of AMPs as antimicrobial agents suggests that drug manufacturers must invest considerable effort in discussing regulatory guidelines with regulators to determine appropriate endpoints and tolerable safety limits.309
Economic viability is the key overarching issue to be addressed. The combination of high production costs, limited delivery methods, and competition from low-cost generic antibiotics results in a poor economic outlook for AMPs unless they target narrow indications that cannot be treated with currently available drugs due to antibiotic resistance. The most realistic strategies are focused on treating hospital-acquired infections with multidrug-resistant pathogens, where treatment expenses are significant and new solutions are desperately needed, and on topical treatments, where production costs are reduced, and less rigorous formulations are possible.248,310,311
Key Challenges and Future Perspectives
Data and Modelling Challenges
The small size and bias of the training datasets remain the main limitations in developing machine learning models capable of predicting AMPs.203 While several AMP databases have been developed, the number of sequences (3,000 to 5,000) in these databases, which have undergone experimental validation of their antimicrobial activities, represents a very small portion of the extensive sequence space of AMPs.312 In addition, the collected data sets exhibit a considerable bias towards peptides from particular sources (eg., amphibian skin secretions, mammalian HDPs) and pathogens. As a result, models are likely to fail when encountering new classes of AMPs and pathogens.313 The quality of the data set continues to represent a serious issue that negatively affects the performance of machine learning models. Diverse approaches and criteria used in experiments for activity measurement, as well as the lack of standardisation in data annotation, introduce noise into the data, making it difficult to identify patterns. Efforts to establish guidelines for data collection and reporting are necessary to improve data quality.175,314,315
Generalizability to novel sequence families is an important test of the model’s usefulness.63 While most benchmark studies focus on identifying sequences similar to those seen during training (at around 30–40% sequence identity), real-world applications require detecting AMPs derived from novel sequence families.316 Methods such as few-shot and zero-shot learning, which leverage transfer learning via pre-trained protein language models, show promise for generalizability beyond the training distribution, but significant performance drops are typically observed for highly dissimilar sequences.317 The mechanistic understanding of machine learning predictions and results remains limited.318 While the use of DL models allows for accurate predictions, understanding how specific sequence features contribute to antimicrobial properties is crucial for rational design.319 Using explainable AI (XAI) techniques such as attention maps, SHAP values, and saliency maps helps uncover insights into the model’s workings, but turning them into design principles poses challenges.320
Experimental Validation Bottlenecks
The high-throughput production of synthesised peptides is a major limiting step in the drug discovery pipeline.29 While computational algorithms can screen several thousand to several million sequences per hour, peptide synthesis is limited to tens to hundreds of sequences per day using automated solid-phase peptide synthesis (SPPS) and costs a lot of money.321 Despite promising developments in DNA-encoded libraries and in cell-free protein expression as alternative approaches that would accelerate the process, these methods have not yet been widely applied to AMP discovery.28 Differences in translation, both in vivo and in vitro, create significant problems in predicting peptide activity.322 For instance, peptides that exhibit interesting antimicrobial activity in vitro by MIC assay may fail to work in vivo due to interactions with plasma proteins, proteolysis, lipid binding, etc.323 Predicting in vivo activity of the sequences based on in vitro results remains one of the major unresolved issues in AMP drug discovery.319 Limited budgets and time for preclinical studies do not allow testing many compounds.175 Indeed, evaluation of each candidate in vitro and in vivo can cost tens to hundreds of thousands of dollars and take months to years.324 Therefore, it is crucial to improve computational approaches and minimise candidates for further experiments.
Future Directions
Multimodal learning that combines sequence information, structural data, and functions is another area of research to explore. ML models using embeddings of protein sequences, predicted 3D structures, and functional information (activity profile, toxicity) can generate higher-quality multimodal representations than unimodal models.325,326 Transformers that take multiple input modalities, mapped using modality-specific encoders followed by cross-modality attention layers, are expected to make significant progress on this task.326 Protein sequence foundation models, such as ESM-3 and ProGen2, are becoming larger and more sophisticated in capturing the relationship between a protein sequence and its function.182 These foundation models, which are pre-trained on billions of protein sequences, might have emergent capabilities to predict AMPs and design them without explicit training on that task.327 The application of prompt-based and in-context learning from natural language processing to protein sequences represents an untapped direction that warrants investigation.328
Closed-loop discovery platforms that incorporate computational design and automated protein sequence synthesis are expected to greatly expedite AMP discovery.65 The recent advancements in automated chemistry, microfluidics, and laboratory robotics enable iterative optimisation through computational design and experimentation without human interaction.329 Several companies, including Recursion, Exscientia, and Insilico Medicine, have successfully applied a closed-loop discovery approach to designing small molecules, and the use of that platform for peptide drugs is currently under development. Personalised antimicrobial therapy involves a futuristic concept where machine learning models will generate personalised AMP depending on the pathogen causing the disease, its location in the body, and the immunological and physiological status of the patient.330,331 By quickly decoding the pathogen’s DNA, followed by computational AMP design, on-demand, highly personalised antimicrobial therapy can be provided.332 Though we are far from realising this scenario, technological advances in rapid diagnostics, computational peptide drug design, and automated synthesis can make it possible.261
Collaboration on data sharing and benchmarking is needed to advance the field.333 Challenges that benchmark different standardised methods against standardised datasets (eg., CASP for protein structure prediction) allow a fair comparison and identify the most promising techniques.334 Federated learning tools that allow model training across diverse datasets without sharing data belonging to a single set of genes could be an effective way to advance the field while respecting data ownership.335
Discussion
Critical Synthesis of ML Approaches
The space of ML techniques for AMPs has witnessed tremendous advancements, evolving from the use of classic classifiers employing hand-crafted physicochemical features to modern techniques such as deep learning (DL), protein language modelling, geometric DL, and generative AI. Each methodology paradigm comes with its strengths and weaknesses and must be considered in relation to particular applications. Classical ML techniques (eg., support vector machines, random forests, gradient boosting), although no longer state-of-the-art in terms of predictive power due to their inefficiency, remain valuable options for problems with limited data available; however, in such cases, efficiency and interpretability become more important than the representational capability of DL. Classical ML algorithms enable researchers to easily visualise decision boundaries and extract feature importance scores from models, providing valuable insights into which physicochemical features contribute to AMPs being antimicrobial rather than non-AMPs. However, by using only predefined features, these algorithms are unable to discover new patterns, a key strength of DL architectures. The latter have set a new bar for AMP classification performance, with transformer-based models consistently outperforming other architectures in classification accuracy. The transformer architecture captures long-range residue interactions through attention mechanisms that play a crucial role in an antimicrobial function and cannot be represented by traditional features. For example, the protein language model ESM-2 achieves the best-known AUC values greater than 0.96 for AMP classification tasks when fine-tuned.
Geometric DL approaches that incorporate three-dimensional structural information via GNNs offer a promising direction, particularly for structure-activity relationship modelling. However, their performance advantage over sequence-based methods remains modest on most benchmarks, suggesting that current structure representation strategies have not yet fully exploited the information available in three-dimensional conformations.110 The integration of AlphaFold2-predicted structures with GNN encoders is an active area of development, and improvements in structure-prediction accuracy are expected to directly translate into better AMP prediction performance.
Generative models for de novo AMP design have advanced from proof-of-concept demonstrations to systems capable of generating libraries of candidate sequences with predicted therapeutic properties. GANs, VAEs, diffusion models, and reinforcement learning each offer distinct trade-offs among generation diversity, controllability of properties, and computational efficiency. Diffusion models currently represent the most promising generative approach, offering high-quality generation with fine-grained control over properties, though their computational requirements remain substantial. The integration of multiple generative strategies within unified frameworks that can adapt the generation approach to specific design objectives represents a promising direction.
The Translation Gap
Notwithstanding the computational successes achieved thus far, the major problem plaguing the development of AMPs designed using machine learning is bringing in silico-predicted peptides to market as approved medicines. The attrition rates are very high, with about 10,000 computationally designed peptides being reduced to only about 500 showing promise in vitro, ~25 of which will be found to have biological activity in animal studies, resulting in 1–2 peptides for clinical trials, each with a mere 10%–20% chance of getting approved.
Several reasons account for this high attrition rate. The problem is that if machine learning models are trained on small and/or unbalanced data sets, their predictions of peptide activity may suffer from overfitting, in which spurious correlations are mistaken for true structure-activity relationships. In addition, there is always the problem of transferring knowledge from peptides with in vitro activity (tested in simple buffer solutions) to peptides exhibiting in vivo activity in the more complicated environment of living animals. The “valley of death” between preclinical development and clinical trials is particularly deep for AMP therapeutics, due to high manufacturing costs, limited bioavailability, and competition from inexpensive generic antibiotics. The most viable path forward likely involves targeting specific clinical niches where existing therapies fail due to resistance and where premium pricing can support development costs: multidrug-resistant nosocomial infections, chronic wound infections, topical applications, and adjunctive therapy in combination with existing antibiotics.
Recommendations for the Field
Based on the results presented in this review, we make the following recommendations to improve ML-driven AMP discovery:
Improve data quality and accessibility: Using identical protocols across communities, requiring minimal information for database deposition, and using the same data would substantially increase the quality and quantity of training data available for model development. Negative examples (confirmed inactive peptides) should be collected and shared to improve class imbalance. Quantitative activity measurements (MIC values) under standardised conditions should be preferred over binary activity annotations. Develop clinically relevant benchmarks: Current benchmarks focus on binary classification accuracy, which is only weakly associated with clinical utility. Multi-objective performance should include activity, toxicity, stability, and pharmacokinetic properties. The use of time-holdout data (train on older data, test on new data) would better inform model generalisation to newly discovered sequences.
Strengthen experimental validation. Predictions should be combined with experimental validation on representative candidates to achieve real-world prediction accuracy.131 The model should be trained via active learning to continually improve its accuracy at minimal cost. Reporting the validation results, both positive and negative, is needed so the model can be evaluated with confidence. Foster interdisciplinary collaboration: Computational scientists, microbiologists, immunologists, medicinal chemists, and clinicians must collaborate to advance further. Computational scientists must gain a better understanding of antimicrobial biology; experiments should use computational methods and validate their results with high-quality data. Agency funding agencies should support collaborative teams consisting of computational and experimental specialists.
Address manufacturing and delivery challenges. Investment in cost-effective peptide manufacturing technologies (recombinant production, novel synthesis methods) is essential for commercial viability. Formulation science to enable oral administration or extended-release parenteral delivery would dramatically expand clinical applications. Platform approaches that leverage common manufacturing and formulation technologies across multiple peptide candidates could reduce per-compound development costs.
Conclusion
The confluence between antimicrobial peptide studies and machine learning is undoubtedly one of the most exciting fields in the worldwide war against AMR. According to what was stated above, machine learning-assisted discovery of AMP has evolved from its infancy, with the use of simplistic classification techniques, to a sophisticated endeavour that now employs protein language models, geometric DL, and generative AI for the prediction, design, and optimisation of new AMPs in a way that has never been seen before. The problem of AMR persists, with WHO priority pathogens such as carbapenem-resistant A. baumannii, Pseudomonas aeruginosa, and Enterobacteriaceae causing hundreds of thousands of fatalities per year, while the antibiotic pipeline lags in addressing the rising demand. AMPs can offer an attractive treatment option because of their broad-spectrum activity, rapid bactericidal effect, ability to act via multiple mechanisms, making it difficult for bacteria to evolve against them, and the potential to serve as immunomodulators.
Current state-of-the-art machine-learning approaches for AMP prediction exhibit very high computational efficiency. Transformers and geometric DL models achieve AUCs above 0.95 on benchmark datasets, whereas generative approaches generate new peptide sequences capable of displaying antimicrobial properties both in vivo and in vitro. Transfer learning from protein language models enables accurate predictions even with limited training data. Moreover, multi-objective optimisation methods are currently used to achieve a trade-off among antimicrobial effect, hemolytic potential, proteolytic stability, and immunogenicity.
Notwithstanding these achievements, the transition from computational success to clinical success has been considerably constrained. AMPs designed using machine learning have shown survival benefits and low levels of resistance to the respective pathogens in animal model experiments. However, few therapeutic agents based on AMP have advanced beyond preclinical evaluation and entered clinical trials, and the process from computational design to clinical validation is long and demanding, with more than 99% of designs failing before reaching the market. Problems of manufacturing, delivery, regulation, and economic viability need to be overcome via consistent investment in the development of peptide formulations and affordable manufacturing techniques. Several factors indicate future advancements in computational design of AMPs. Multimodal learning models that leverage the synergy among sequence information, structural data, and functional properties promise improved representational capacity. Larger protein language models exhibit emergent capabilities, including increased scalability and predictive accuracy in AMP design. Finally, closed-loop automated systems that seamlessly combine peptide design, synthesis, and high-throughput testing may reduce development time by orders of magnitude. This may pave the way for an integrated approach to personalised antimicrobial therapy by leveraging the synergistic use of rapid diagnostics, computational design, and on-site peptide synthesis.
Realising this vision will require sustained interdisciplinary collaboration between computational scientists, experimental biologists, medicinal chemists, clinicians, and regulatory specialists. Community initiatives to standardise data collection, establish meaningful benchmarks, and share both positive and negative validation results are essential for collective progress. The AMR crisis demands urgent action, and the tools of machine learning - properly developed, rigorously validated, and thoughtfully translated - offer a powerful weapon in this existential struggle against resistant pathogens. The future of antimicrobial therapy will be increasingly computational, and the integration of AI-driven peptide design with experimental validation and clinical development represents our best hope for outpacing the relentless evolution of bacterial resistance.
Abbreviations
AAC, Amino Acid Composition; ACC, Accuracy; ADME, Absorption, Distribution, Metabolism, Excretion; AI, Artificial Intelligence; AMR, Antimicrobial Resistance; AMP, Antimicrobial Peptide; APD3, Antimicrobial Peptide Database version 3; AUC, Area Under the ROC Curve; BiLSTM, Bidirectional Long Short-Term Memory; CAMPR3, Collection of Anti-Microbial Peptides version 3; CASP, Critical Assessment of Protein Structure Prediction; CLS, Clinical Laboratory Standard; CNN, Convolutional Neural Network; CoMFA, Comparative Molecular Field Analysis; CoMSIA, Comparative Molecular Similarity Indices Analysis; CTD, Composition-Transition-Distribution; DBAASP, Database of Antimicrobial Activity and Structure of Peptides; DL, Deep Learning; DPC, Dipeptide Composition; DRAMP, Data Repository of Antimicrobial Peptides; EFA, European Food Safety Authority; EMA, European Medicines Agency; ESM, Evolutionary Scale Model; FBGAN, Feedback Generative Adversarial Network; FDA, U.S. Food and Drug Administration; GAN, Generative Adversarial Network; GAT, Graph Attention Network; GCN, Graph Convolutional Network; GNN, Graph Neural Network; GRU, Gated Recurrent Unit.
Data Sharing Statement
The data used and/or analysed during the current study are available from the corresponding author on reasonable request.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Disclosure
The authors report no conflicts of interest in this work.
References
1. Murray CJL, Ikuta KS, Sharara F, et al. Global burden of bacterial antimicrobial resistance in 2019: a systematic analysis. Lancet. 2022;399(10325):629–50. doi:10.1016/S0140-6736(21)02724-0
2. Leung E, Weil DE, Raviglione M, et al. The WHO policy package to combat antimicrobial resistance. Bull World Health Organ. 2011;89(5):390–392. doi:10.2471/blt.11.088435
3. Holmes AH, Moore LSP, Sundsfjord A, et al. Understanding the mechanisms and drivers of antimicrobial resistance. Lancet. 2016;387(10014):176–187. doi:10.1016/s0140-6736(15)00473-0
4. Bahar A, Porbaran M, Khazaei M, et al. Antimicrobial peptides for anticancer and antiviral therapy: last promising update. Discover Oncology. 2025;16(1):1991. doi:10.1007/s12672-025-03855-8
5. Tahmasebi H, Dehbashi S, Jahantigh M, et al. Relationship between biofilm gene expression with antimicrobial resistance pattern and clinical specimen type based on sequence types (STs) of methicillin-resistant S. aureus. Mol Biol Rep. 2020;47(2):1309–1320. doi:10.1007/s11033-019-05233-4
6. Porbaran M, Tahmasebi H, Arabestani M, Falkinham J. A comprehensive study of the relationship between the production of β -lactamase enzymes and iron/siderophore uptake regulatory genes in clinical isolates of Acinetobacter baumannii. Int. J. Microbiol. 2021;2021:1–8. doi:10.1155/2021/5565537
7. Dehbashi S, Tahmasebi H, Sedighi P, et al. Development of high-resolution melting curve analysis in rapid detection of vanA gene, Enterococcus faecalis, and Enterococcus faecium from clinical isolates. Trop. Med. Health. 2020:48. doi:10.1186/s41182-020-00197-9.
8. Rice LB. Federal funding for the study of antimicrobial resistance in nosocomial pathogens: no ESKAPE. J Infect Dis. 2008;197(8):1079–1081. doi:10.1086/533452
9. Asghar MU, Zaidi AH, Tariq M, et al. Seasonal and hospital settings variations in antimicrobial resistance among clinical isolates from cardiac patients: insights from a 7-Year study. BMC Infect Dis. 2025;25(1):936. doi:10.1186/s12879-025-11171-3
10. Tacconelli E, Carrara E, Savoldi A, et al. Discovery, research, and development of new antibiotics: the WHO priority list of antibiotic-resistant bacteria and tuberculosis. Lancet Infect Dis. 2018;18(3):318–327. doi:10.1016/s1473-3099(17)30753-3
11. Butler MS, Paterson DL. Antibiotics in the clinical pipeline in. J Antibiot. 2020;73:329–364. doi:10.1038/s41429-020-0291-8
12. Laxminarayan R, Matsoso P, Pant S, et al. Access to effective antimicrobials: a worldwide challenge. Lancet. 2016;387(10014):168–175. doi:10.1016/s0140-6736(15)00474-2
13. Zasloff M. Antimicrobial peptides of multicellular organisms. Nature. 2002;415(6870):389–395. doi:10.1038/415389a
14. Wang G, Li X, Wang Z. APD3: the antimicrobial peptide database as a tool for research and education. Nucleic Acids Res. 2016;44(D1):D1087–93. doi:10.1093/nar/gkv1278
15. Butler MS, Gigante V, Sati H, et al. Analysis of the clinical pipeline of treatments for drug-resistant bacterial infections: despite progress, more action is needed. Antimicrob Agents Chemother. 2022;66(3):e0199121. doi:10.1128/aac.01991-21
16. Yeaman MR, Yount NY. Mechanisms of antimicrobial peptide action and resistance. Pharmacol Rev. 2003;55(1):27–55. doi:10.1124/pr.55.1.2
17. Teixeira V, Feio MJ, Bastos M. Role of lipids in the interaction of antimicrobial peptides with membranes. Prog Lipid Res. 2012;51(2):149–177. doi:10.1016/j.plipres.2011.12.005
18. Lehrer RI, Lu W. α-Defensins in human innate immunity. Immunol Rev. 2012;245(1):84–112. doi:10.1111/j.1600-065X.2011.01082.x
19. Hancock RE, Nijnik A, Philpott DJ. Modulating immunity as a therapy for bacterial infections. Nat Rev Microbiol. 2012;10:243–254. doi:10.1038/nrmicro2745
20. Batoni G, Maisetta G, Esin S. Antimicrobial peptides and their interaction with biofilms of medically relevant bacteria. Biochim Biophys Acta. 2016;1858(5):1044–1060. doi:10.1016/j.bbamem.2015.10.013
21. Huan Y, Kong Q, Mou H, et al. Antimicrobial peptides: classification, design, application and research progress in multiple fields. Front Microbiol. 2020;11:582779. doi:10.3389/fmicb.2020.582779
22. Matsuzaki K. Why and how are peptide–lipid interactions utilized for self-defense? Magainins and tachyplesins as archetypes. Biochim Biophys Acta. 1999;1462(1–2):1–10. doi:10.1016/s0005-2736(99)00197-2
23. Antunes B, Zanchi C, Johnston PR, et al. The evolution of antimicrobial peptide resistance in Pseudomonas aeruginosa is severely constrained by random peptide mixtures. PLOS Biology. 2024;22(7):e3002692. doi:10.1371/journal.pbio.3002692
24. Donaghy C, Javellana JG, Hong YJ, et al. The synergy between zinc and antimicrobial peptides: an insight into unique bioinorganic interactions. Molecules. 2023:28. doi:10.3390/molecules28052156.
25. Antonietti M, Kim CK, Granack S, et al. An analysis of intrinsic protein disorder in antimicrobial peptides. Protein J. 2025;44(2):175–191. doi:10.1007/s10930-025-10253-0
26. French NP, Thomas KM, Amani NB, et al. Population structure and antimicrobial resistance in campylobacter jejuni and c. coli isolated from humans with diarrhea and from poultry, East Africa. Emerg Infect Dis. 2024;30(10):2079–2089. doi:10.3201/eid3010.231399
27. Zhu Y, Hao W, Wang X, et al. Antimicrobial peptides, conventional antibiotics, and their synergistic utility for the treatment of drug-resistant infections. Med Res Rev. 2022;42(4):1377–1422. doi:10.1002/med.21879
28. Zheng B, Wang X, Guo M, et al. Therapeutic peptides: recent advances in discovery, synthesis, and clinical translation. Int J Mol Sci. 2025;26(11):5131. doi:10.3390/ijms26115131
29. Silver D, Hubert T, Schrittwieser J, et al. A general reinforcement learning algorithm that masters chess, shogi, and Go through self-play. Science. 2018;362(6419):1140–1144. doi:10.1126/science.aar6404
30. Agüero-Chapin G, Antunes A, Marrero-Ponce Y. A 2022 update on computational approaches to the discovery and design of antimicrobial peptides. Antibiotics. 2023;12. doi:10.3390/antibiotics12061011
31. Zhang K, Yang X, Wang Y, et al. Artificial intelligence in drug development. Nature Med. 2025;31(1):45–59. doi:10.1038/s41591-024-03434-4
32. Gawde U, Chakraborty S, Waghu FH, et al. CAMPR4: a database of natural and synthetic antimicrobial peptides. Nucleic Acids Res. 2023;51(D1):D377–d383. doi:10.1093/nar/gkac933
33. Owliaee I, Khaledian M, Shojaeian A, et al. Antimicrobial peptides against arboviruses: mechanisms, challenges, and future directions. Probiotics Antimicrob Proteins. 2025;17(4):2058–2085. doi:10.1007/s12602-024-10430-0
34. Kiper E, Ben Hur D, Alfandari D, et al. Antimicrobial peptides selectively target malaria parasites by a cholesterol-dependent mechanism. J Biol Chem. 2025;301(4):108298. doi:10.1016/j.jbc.2025.108298
35. Tajer L, Paillart J-C, Dib H, et al. Molecular mechanisms of bacterial resistance to antimicrobial peptides in the modern era: an updated review. Microorganisms. 2024;12(7):1259. doi:10.3390/microorganisms12071259
36. Seo S, Jung J, Kim CY, et al. Antimicrobial peptides encounter resistance of aureolysin during their action on Staphylococcus aureus biofilm. Biotechnol Bioprocess Eng. 2021;26(2):216–222. doi:10.1007/s12257-020-0384-z
37. Clarke SR, Foster SJ. IsdA Protects Staphylococcus aureus against the Bactericidal Protease Activity of Apolactoferrin. Infect Immun. 2008;76(4):1518–1526. doi:10.1128/iai.01530-07
38. Recsei P, Kreiswirth B, O’Reilly M, et al. Regulation of exoprotein gene expression in Staphylococcus aureus by agr. Mol Gen Genet. 1986;202(1):58–61. doi:10.1007/bf00330517
39. Jones CS, Anderson AC, Clarke AJ. Mechanism of Staphylococcus aureus peptidoglycan O-acetyltransferase A as an O-acyltransferase. Proc Natl Acad Sci U S A. 2021;118. doi:10.1073/pnas.2103602118
40. Koprivnjak T, Weidenmaier C, Peschel A, et al. Wall teichoic acid deficiency in Staphylococcus aureus confers selective resistance to mammalian group iia phospholipase a 2 and human β-defensin 3. Infect Immun. 2008;76(5):2169–2176. doi:10.1128/iai.01705-07
41. Collins LV, Kristian SA, Weidenmaier C, et al. Staphylococcus aureus strains lacking D-alanine modifications of teichoic acids are highly susceptible to human neutrophil killing and are virulence attenuated in mice. J Infect Dis. 2002;186:214–219. doi:10.1086/341454
42. Peleg AY, Miyakis S, Ward DV, et al. Whole genome characterization of the mechanisms of daptomycin resistance in clinical and laboratory derived isolates of Staphylococcus aureus. PLoS One. 2012;7(1):e28316. doi:10.1371/journal.pone.0028316
43. Peschel A, Jack RW, Otto M, et al. Staphylococcus aureus resistance to human defensins and evasion of neutrophil killing via the novel virulence factor mprf is based on modification of membrane lipids with l -lysine. J Exp Med. 2001;193(9):1067–1076. doi:10.1084/jem.193.9.1067
44. Cheung GYC, Fisher EL, McCausland JW, et al. Antimicrobial peptide resistance mechanism contributes to Staphylococcus aureus infection. J Infect Dis. 2018;217:1153–1159. doi:10.1093/infdis/jiy024
45. Kupferwasser LI, Skurray RA, Brown MH, et al. Plasmid-mediated resistance to thrombin-induced platelet microbicidal protein in staphylococci: role of the qaca locus. Antimicrob Agents Chemother. 1999;43(10):2395–2399. doi:10.1128/aac.43.10.2395
46. Falord M, Karimova G, Hiron A, et al. GraXSR proteins interact with the VraFG ABC transporter to form a five-component system required for cationic antimicrobial peptide sensing and resistance in Staphylococcus aureus. Antimicrob Agents Chemother. 2012;56(2):1047–1058. doi:10.1128/aac.05054-11
47. Cao R, Su H, Wei Z, et al. An induced mutation of ABC-transporter component VraF(K84E) contributes to vancomycin resistance and virulence in Staphylococcus aureus strain MW2. Int J Med Microbiol. 2024;315:151624. doi:10.1016/j.ijmm.2024.151624
48. Vuong C, Voyich JM, Fischer ER, et al. Polysaccharide intercellular adhesin (PIA) protects Staphylococcus epidermidis against major components of the human innate immune system. Cell Microbiol. 2004;6(3):269–275. doi:10.1046/j.1462-5822.2004.00367.x
49. Schmidtchen A, Frick IM, Andersson E, et al. Proteinases of common pathogenic bacteria degrade and inactivate the antibacterial peptide LL-37. Mol Microbiol. 2002;46:157–168. doi:10.1046/j.1365-2958.2002.03146.x
50. Cole JN, Pence MA, von Köckritz-Blickwede M, et al. M protein and hyaluronic acid capsule are essential for in vivo selection of covRS mutations characteristic of invasive serotype M1T1 group A Streptococcus. mBio. 2010:1. doi:10.1128/mBio.00191-10.
51. Lauth X, von Köckritz-Blickwede M, McNamara CW, et al. M1 protein allows Group A streptococcal survival in phagocyte extracellular traps through cathelicidin inhibition. J Innate Immun. 2009;1(3):202–214. doi:10.1159/000203645
52. Hollands A, Gonzalez D, Leire E, et al. A bacterial pathogen co-opts host plasmin to resist killing by cathelicidin antimicrobial peptides. J Biol Chem. 2012;287(49):40891–40897. doi:10.1074/jbc.M112.404582
53. Frick IM, Akesson P, Rasmussen M, et al. SIC, a secreted protein of Streptococcus pyogenes that inactivates antibacterial peptides. J Biol Chem. 2003;278:16561–16566. doi:10.1074/jbc.M301995200
54. Schmidtchen A, Frick IM, Björck L. Dermatan sulphate is released by proteinases of common pathogenic bacteria and inactivates antibacterial alpha-defensin. Mol Microbiol. 2001;39:708–713. doi:10.1046/j.1365-2958.2001.02251.x
55. Nyberg P, Rasmussen M, Björck L. alpha2-Macroglobulin-proteinase complexes protect Streptococcus pyogenes from killing by the antimicrobial peptide LL-37. J Biol Chem. 2004;279:52820–52823. doi:10.1074/jbc.C400485200
56. Froehlich BJ, Bates C, Scott JR. Streptococcus pyogenes CovRS mediates growth in iron starvation and in the presence of the human cationic antimicrobial peptide LL-37. J Bacteriol. 2009;191:673–677. doi:10.1128/jb.01256-08
57. Nizet V, Ohtake T, Lauth X, et al. Innate antimicrobial peptide protects the skin from invasive bacterial infection. Nature. 2001;414(6862):454–457. doi:10.1038/35106587
58. Kristian SA, Datta V, Weidenmaier C, et al. D-alanylation of teichoic acids promotes group a streptococcus antimicrobial peptide resistance, neutrophil survival, and epithelial cell invasion. J Bacteriol. 2005;187:6719–6725. doi:10.1128/jb.187.19.6719-6725.2005
59. Dinulos JG, Mentele L, Fredericks LP, et al. Keratinocyte expression of human beta defensin 2 following bacterial infection: role in cutaneous host defense. Clin Diagn Lab Immunol. 2003;10(1):161–166. doi:10.1128/cdli.10.1.161-166.2003
60. Klinzing DC, Ishmael N, Hotopp JCD, et al. The two-component response regulator LiaR regulates cell wall stress responses, pili expression and virulence in group B Streptococcus. Microbiology. 2013;159:1521–1534. doi:10.1099/mic.0.064444-0
61. Quach D, van Sorge NM, Kristian SA, et al. The CiaR response regulator in group b streptococcus promotes intracellular survival and resistance to innate immune defenses. J Bacteriol. 2009;191(7):2023–2032. doi:10.1128/jb.01216-08
62. Hamilton A, Popham DL, Carl DJ, et al. Penicillin-binding protein 1a promotes resistance of group B streptococcus to antimicrobial peptides. Infect Immun. 2006;74(11):6179–6187. doi:10.1128/iai.00895-06
63. Maisey HC, Doran KS, Nizet V. Recent advances in understanding the molecular basis of group B Streptococcus virulence. Expert Rev Mol Med. 2008;10:e27. doi:10.1017/s1462399408000811
64. Poyart C, Lamy MC, Boumaila C, et al. Regulation of d -alanyl-lipoteichoic acid biosynthesis in streptococcus agalactiae involves a novel two-component regulatory system. J Bacteriol. 2001;183:6324–6334. doi:10.1128/jb.183.21.6324-6334.2001
65. Shaper M, Hollingshead SK, Benjamin WH, et al. PspA protects Streptococcus pneumoniae from killing by apolactoferrin, and antibody to PspA enhances killing of pneumococci by apolactoferrin [corrected]. Infect Immun. 2004;72:5031–5040. doi:10.1128/iai.72.9.5031-5040.2004
66. Llobet E, Tomás JM, Bengoechea JA. Capsule polysaccharide is a bacterial decoy for antimicrobial peptides. Microbiology. 2008;154(12):3877–3886. doi:10.1099/mic.0.2008/022301-0
67. Kovács M, Halfmann A, Fedtke I, et al. A functional dlt operon, encoding proteins required for incorporation of d-alanine in teichoic acids in gram-positive bacteria, confers resistance to cationic antimicrobial peptides in Streptococcus pneumoniae. J Bacteriol. 2006;188:5797–5805. doi:10.1128/jb.00336-06
68. Majchrzykiewicz JA, Kuipers OP, Bijlsma JJ. Generic and specific adaptive responses of Streptococcus pneumoniae to challenge with three distinct antimicrobial peptides, bacitracin, LL-37, and nisin. Antimicrob Agents Chemother. 2010;54:440–451. doi:10.1128/aac.00769-09
69. Davis KM, Akinbi HT, Standish AJ, et al. Resistance to mucosal lysozyme compensates for the fitness deficit of peptidoglycan modifications by Streptococcus pneumoniae. PLoS Pathog. 2008;4(12):e1000241. doi:10.1371/journal.ppat.1000241
70. Nešuta O, Budešínský M, Hadravová R, et al. How proteases from Enterococcus faecalis contribute to its resistance to short α-helical antimicrobial peptides. Pathog Dis. 2017:75. doi:10.1093/femspd/ftx091.
71. Fabretti F, Theilacker C, Baldassarri L, et al. Alanine esters of enterococcal lipoteichoic acid play a role in biofilm formation and resistance to antimicrobial peptides. Infect Immun. 2006;74(7):4164–4171. doi:10.1128/iai.00111-06
72. Bao Y, Sakinc T, Laverde D, et al. Role of mprF1 and mprF2 in the pathogenicity of Enterococcus faecalis. PLoS One. 2012;7(6):e38458. doi:10.1371/journal.pone.0038458
73. Reyes J, Panesso D, Tran TT, et al. A liaR deletion restores susceptibility to daptomycin and antimicrobial peptides in multidrug-resistant Enterococcus faecalis. J Infect Dis. 2015;211(8):1317–1325. doi:10.1093/infdis/jiu602
74. Hébert L, Courtin P, Torelli R, et al. Enterococcus faecalis constitutes an unusual bacterial model in lysozyme resistance. Infect Immun. 2007;75(11):673–677. doi:10.1128/JB.01256-08
75. Maloney E, Stankowska D, Zhang J, et al. The two-domain LysX protein of Mycobacterium tuberculosis is required for production of lysinylated phosphatidylglycerol and resistance to cationic antimicrobial peptides. PLoS Pathog. 2009;5(7):e1000534. doi:10.1371/journal.ppat.1000534
76. Kumar P, Kizhakkedathu JN, Straus SK. Antimicrobial peptides: diversity, mechanism of action and strategies to improve the activity and biocompatibility in vivo. Biomolecules. 2018;8. doi:10.3390/biom8010004
77. Huang HW. DAPTOMYCIN, its membrane-active mechanism vs. that of other antimicrobial peptides. Biochim Biophys Acta Biomembr. 2020;1862(10):183395. doi:10.1016/j.bbamem.2020.183395
78. Urmi UL, Vijay AK, Kuppusamy R, et al. A review of the antiviral activity of cationic antimicrobial peptides. Peptides. 2023;166:171024. doi:10.1016/j.peptides.2023.171024
79. Gani Z, Kumar A, Raje M, et al. Antimicrobial peptides: an alternative strategy to combat antimicrobial resistance. Drug Discov Today. 2025;30(2):104305. doi:10.1016/j.drudis.2025.104305
80. Xuan J, Feng W, Wang J, et al. Antimicrobial peptides for combating drug-resistant bacterial infections. Drug Resistance Updates. 2023;68:100954. doi:10.1016/j.drup.2023.100954
81. Neghabi Hajigha M, Hajikhani B, Vaezjalali M, et al. Antiviral and antibacterial peptides: mechanisms of action. Heliyon. 2024;10(22):e40121. doi:10.1016/j.heliyon.2024.e40121
82. Grönberg A, Mahlapuu M, Ståhle M, et al. Treatment with LL −37 is safe and effective in enhancing healing of hard-to-heal venous leg ulcers: a randomized, placebo-controlled clinical trial. Wound Repair Regen. 2014;22(5):613–621. doi:10.1111/wrr.12211
83. Shin JM, Gwak JW, Kamarajan P, et al. Biomedical applications of nisin. J Appl Microbiol. 2016;120(6):1449–1465. doi:10.1111/jam.13033
84. Sierra JM, Fusté E, Rabanal F, et al. An overview of antimicrobial peptides and the latest advances in their development. Expert Opin Biol Ther. 2017;17(6):663–676. doi:10.1080/14712598.2017.1315402
85. Mensa B, Howell GL, Scott R, et al. Comparative mechanistic studies of brilacidin, daptomycin, and the antimicrobial peptide LL16. Antimicrob Agents Chemother. 2014;58(9):6324–6334. doi:10.1128/JB.183.21.6324-6334.2001
86. Falagas ME, Kyriakidou M, Voulgaris GL, et al. Clinical use of intravenous polymyxin B for the treatment of patients with multidrug-resistant Gram-negative bacterial infections: an evaluation of the current evidence. J Glob Antimicrob Resist. 2021;24:342–359. doi:10.1016/j.jgar.2020.12.026
87. Wang SZ, Hu JT, Zhang C, et al. The safety and efficacy of daptomycin versus other antibiotics for skin and soft-tissue infections: a meta-analysis of randomised controlled trials. BMJ Open. 2014;4(6):e004744. doi:10.1136/bmjopen-2013-004744
88. Díez-Aguilar M, Hernández-García M, Morosini MI, et al. Murepavadin antimicrobial activity against and resistance development in cystic fibrosis Pseudomonas aeruginosa isolates. J Antimicrob Chemother. 2021;76:984–992. doi:10.1093/jac/dkaa529
89. Yang X, Yousef AE. Antimicrobial peptides produced by Brevibacillus spp.: structure, classification and bioactivity: a mini review. World J Microbiol Biotechnol. 2018;34(4):57. doi:10.1007/s11274-018-2437-4
90. Sinha M, Kaushik S, Kaur P, et al. Antimicrobial lactoferrin peptides: the hidden players in the protective function of a multifunctional protein. Int J Pept. 2013;2013:390230. doi:10.1155/2013/390230
91. Douglas EJA, Wulandari SW, Lovell SD, et al. Novel antimicrobial strategies to treat multi-drug resistant Staphylococcus aureus infections. Microb Biotechnol. 2023;16:1456–1474. doi:10.1111/1751-7915.14268
92. Rubinchik E, Dugourd D, Algara T, et al. Antimicrobial and antifungal activities of a novel cationic antimicrobial peptide, omiganan, in experimental skin colonisation models. Int J Antimicrob Agents. 2009;34(5):457–461. doi:10.1016/j.ijantimicag.2009.05.003
93. Sultana A, Luo H, Ramakrishna S. Antimicrobial peptides and their applications in biomedical sector. Antibiotics. 2021;10:1094.
94. Willcox MD, Hume EB, Aliwarga Y, et al. A novel cationic-peptide coating for the prevention of microbial colonization on contact lenses. J Appl Microbiol. 2008;105(6):1817–1825. doi:10.1111/j.1365-2672.2008.03942.x
95. Hasan M, Ruksznis C, Wang Y, et al. Antimicrobial peptides inhibit polyinosinic-polycytidylic acid-induced immune responses. J Immunol. 2011;187(11):5653–5659. doi:10.4049/jimmunol.1102144
96. Buda De Cesare G, Cristy SA, Garsin DA, et al. Antimicrobial peptides: a new frontier in antifungal therapy. mBio. 2020:11. doi:10.1128/mBio.02123-20.
97. Shah P, Hsiao FS-H, Ho Y-H, et al. The proteome targets of intracellular targeting antimicrobial peptides. Proteomics. 2016;16(8):1225–1237. doi:10.1002/pmic.201500380
98. da Costa de Souza G, Roque-Borda CA, Pavan FR. Beta-lactam resistance and the effectiveness of antimicrobial peptides against KPC-producing bacteria. Drug Dev Res. 2022;83:1534–1554. doi:10.1002/ddr.21990
99. Piers KL, Brown MH, Hancock RE. Recombinant DNA procedures for producing small antimicrobial cationic peptides in bacteria. Gene. 1993;134:7–13. doi:10.1016/0378-1119(93)90168-3
100. Martínez B, Rodríguez A, Suárez E. Antimicrobial peptides produced by bacteria: the bacteriocins. In: Villa TG, Vinas M, editors. New Weapons to Control Bacterial Growth. Cham: Springer International Publishing; 2016:15–38.
101. Stumpe S, Schmid R, Stephens DL, et al. Identification of OmpT as the protease that hydrolyzes the antimicrobial peptide protamine before it enters growing cells of Escherichia coli. J Bacteriol. 1998;180(15):4002–4006. doi:10.1128/jb.180.15.4002-4006.1998
102. Wang J, Ma W, Fang Y, et al. Core oligosaccharide portion of lipopolysaccharide plays important roles in multiple antibiotic resistance in Escherichia coli. Antimicrob Agents Chemother. 2021;65(10):e0034121. doi:10.1128/aac.00341-21
103. Smith AE, Kim S-H, Liu F, et al. PagP activation in the outer membrane triggers R3 core oligosaccharide truncation in the cytoplasm of Escherichia coli O157:H7. J Biol Chem. 2008;283(7):4332–4343. doi:10.1074/jbc.M708163200
104. Salazar J, Alarcón M, Huerta J, et al. Phosphoethanolamine addition to the Heptose I of the Lipopolysaccharide modifies the inner core structure and has an impact on the binding of Polymyxin B to the Escherichia coli outer membrane. Arch Biochem Biophys. 2017;620:28–34. doi:10.1016/j.abb.2017.03.008
105. Necula G, Bacalum M, Radu M. Interaction of tryptophan- and arginine-rich antimicrobial peptide with E. coli outer membrane-a molecular simulation approach. Int J Mol Sci. 2023;24. doi:10.3390/ijms24032005
106. Weatherspoon-Griffin N, Zhao G, Kong W, et al. The CpxR/CpxA two-component system up-regulates two Tat-dependent peptidoglycan amidases to confer bacterial resistance to antimicrobial peptide. J Biol Chem. 2011;286(7):5529–5539. doi:10.1074/jbc.M110.200352
107. Weatherspoon-Griffin N, Yang D, Kong W, et al. The CpxR/CpxA two-component regulatory system up-regulates the multidrug resistance cascade to facilitate Escherichia coli resistance to a model antimicrobial peptide. J Biol Chem. 2014;289(47):32571–32582. doi:10.1074/jbc.M114.565762
108. Chakraborty K, Ghosh S, Koley H, et al. Bacterial exotoxins downregulate cathelicidin (hCAP-18/LL-37) and human beta-defensin 1 (HBD-1) expression in the intestinal epithelial cells. Cell Microbiol. 2008;10:2520–2537. doi:10.1111/j.1462-5822.2008.01227.x
109. Campos MA, Vargas MA, Regueiro V, et al. Capsule polysaccharide mediates bacterial resistance to antimicrobial peptides. Infect Immun. 2004;72(12):7107–7114. doi:10.1128/iai.72.12.7107-7114.2004
110. Chaput C, Spindler E, Gill RT, et al. O-antigen protects gram-negative bacteria from histone killing. PLoS One. 2013;8(8):e71097. doi:10.1371/journal.pone.0071097
111. Sun L, Zhang Y, Cai T, et al. CrrAB regulates PagP-mediated glycerophosphoglycerol palmitoylation in the outer membrane of Klebsiella pneumoniae. J Lipid Res. 2022;63(9):100251. doi:10.1016/j.jlr.2022.100251
112. Llobet E, Martínez-Moliner V, Moranta D, et al. Deciphering tissue-induced Klebsiella pneumoniae lipid A structure. Proc Natl Acad Sci U S A. 2015;112(46):E6369–78. doi:10.1073/pnas.1508820112
113. Mirshekar M, Zadeh RG, Moghadam MT, et al. Upregulation of pmrA, pmrB, pmrC, phoQ, phoP, and arnT genes contributing to resistance to colistin in superbug Klebsiella pneumoniae isolates from human clinical samples in Tehran, Iran. New Microbes New Infect. 2024;59:101275. doi:10.1016/j.nmni.2024.101275
114. Singh S, Pathak A, Kumar A, et al. Emergence of chromosome-borne colistin resistance gene mcr-1 in clinical isolates of Klebsiella pneumoniae from India. Antimicrob Agents Chemother. 2018:62. doi:10.1128/aac.01885-17.
115. Llobet E, March C, Giménez P, et al. Klebsiella pneumoniae OmpA confers resistance to antimicrobial peptides. Antimicrob Agents Chemother. 2009;53(1):298–302. doi:10.1128/aac.00657-08
116. Hsu CR, Chang IW, Hsieh PF, et al. A novel role for the Klebsiella pneumoniae sap (sensitivity to antimicrobial peptides) transporter in intestinal cell interactions, innate immune responses, liver abscess, and virulence. J Infect Dis. 2019;219:1294–1306. doi:10.1093/infdis/jiy615
117. Padilla E, Llobet E, Doménech-Sánchez A, et al. Klebsiella pneumoniae acrab efflux pump contributes to antimicrobial resistance and virulence. Antimicrob Agents Chemother. 2010;54(1):177–183. doi:10.1128/aac.00715-09
118. Guina T, Yi EC, Wang H, et al. A phop-regulated outer membrane protease of Salmonella enterica serovar typhimurium promotes resistance to alpha-helical antimicrobial peptides. J Bacteriol. 2000;182(14):4077–4086. doi:10.1128/jb.182.14.4077-4086.2000
119. Guo L, Lim KB, Gunn JS, et al. Regulation of lipid a modifications by salmonella typhimurium virulence genes phoP-phoQ. Science. 1997;276(5310):250–253. doi:10.1126/science.276.5310.250
120. Kawasaki K, China K, Nishijima M. Release of the lipopolysaccharide deacylase pagl from latency compensates for a lack of lipopolysaccharide aminoarabinose modification-dependent resistance to the antimicrobial peptide polymyxin b in Salmonella enterica. J Bacteriol. 2007;189(13):4911–4919. doi:10.1128/jb.00451-07
121. Strandberg KL, Richards SM, Gunn JS, Bäumler AJ. Cathelicidin antimicrobial peptide expression is not induced or required for bacterial clearance during Salmonella enterica infection of human monocyte-derived macrophages. Infect Immun. 2012;80(11):3930–3938. doi:10.1128/iai.00672-12
122. Tamayo R, Choudhury B, Septer A, et al. Identification of cptA, a pmra-regulated locus required for phosphoethanolamine modification of the Salmonella enterica serovar typhimurium lipopolysaccharide core. J Bacteriol. 2005;187(10):3391–3399. doi:10.1128/jb.187.10.3391-3399.2005
123. Honeycutt JD, Wenner N, Li Y, et al. Genetic variation in the MacAB-TolC efflux pump influences pathogenesis of invasive Salmonella isolates from Africa. PLoS Pathog. 2020;16(8):e1008763. doi:10.1371/journal.ppat.1008763
124. Parra-Lopez C, Baer MT, Groisman EA. Molecular genetic analysis of a locus required for resistance to antimicrobial peptides in Salmonella typhimurium. EMBO j. 1993;12:4053–4062. doi:10.1002/j.1460-2075.1993.tb06089.x
125. Humphreys S, Stevenson A, Bacon A, et al. The alternative sigma factor, sigmaE, is critically important for the virulence of Salmonella typhimurium. Infect Immun. 1999;67:1560–1568. doi:10.1128/iai.67.4.1560-1568.1999
126. Islam D, Bandholtz L, Nilsson J, et al. Downregulation of bactericidal peptides in enteric infections: a novel immune escape mechanism with bacterial DNA as a potential regulator. Nat Med. 2001;7:180–185. doi:10.1038/84627
127. West NP, Sansonetti P, Mounier J, et al. Optimization of virulence functions through glucosylation of Shigella LPS. Science. 2005;307(5713):1313–1317. doi:10.1126/science.1108472
128. Arbibe L, Kim DW, Batsche E, et al. An injected bacterial effector targets chromatin access for transcription factor NF-kappaB to alter transcription of host genes involved in immune responses. Nat Immunol. 2007;8:47–56. doi:10.1038/ni1423
129. Devine DA, Marsh PD, Percival RS, et al. Modulation of antibacterial peptide activity by products of Porphyromonas gingivalis and Prevotella spp. Microbiology. 1999;145(4):965–971. doi:10.1099/13500872-145-4-965
130. Belas R, Manos J, Suvanasuthi R. Proteus mirabilis zapa metalloprotease degrades a broad spectrum of substrates, including antimicrobial peptides. Infect Immun. 2004;72(9):5159–5167. doi:10.1128/iai.72.9.5159-5167.2004
131. McCoy AJ, Liu H, Falla TJ, et al. Identification of Proteus mirabilis mutants with increased sensitivity to antimicrobial peptides. Antimicrob Agents Chemother. 2001;45(7):2030–2037. doi:10.1128/aac.45.7.2030-2037.2001
132. Kooi C, Sokol PA. Burkholderia cenocepacia zinc metalloproteases influence resistance to antimicrobial peptides. Microbiology. 2009;155(9):2818–2825. doi:10.1099/mic.0.028969-0
133. Foschiatti M, Cescutti P, Tossi A, et al. Inhibition of cathelicidin activity by bacterial exopolysaccharides. Mol Microbiol. 2009;72(5):1137–1146. doi:10.1111/j.1365-2958.2009.06707.x
134. Loutet SA, Valvano MA. Extreme antimicrobial Peptide and polymyxin B resistance in the genus burkholderia. Front Microbiol. 2011;2:159. doi:10.3389/fmicb.2011.00159
135. Fehlner-Gardiner CC, Valvano MA. Cloning and characterization of the Burkholderia vietnamiensis norM gene encoding a multi-drug efflux protein. FEMS Microbiol Lett. 2002;215(2):279–283. doi:10.1111/j.1574-6968.2002.tb11403.x
136. Pérez A, Poza M, Fernández A, et al. Involvement of the AcrAB-TolC efflux pump in the resistance, fitness, and virulence of Enterobacter cloacae. Antimicrob Agents Chemother. 2012;56(4):2084–2090. doi:10.1128/aac.05509-11
137. Huang L, Feng Y, Zong Z. Heterogeneous resistance to colistin in Enterobacter cloacae complex due to a new small transmembrane protein. J Antimicrob Chemother. 2019;74(9):2551–2558. doi:10.1093/jac/dkz236
138. Boll JM, Tucker AT, Klein DR, et al. Reinforcing lipid a acylation on the cell surface of Acinetobacter baumannii promotes cationic antimicrobial peptide resistance and desiccation survival. mBio. 2015;6(3):e00478–15. doi:10.1128/mBio.00478-15
139. Chin C-Y, Gregg KA, Napier BA, et al. A pmrb-regulated deacetylase required for lipid a modification and polymyxin resistance in Acinetobacter baumannii. Antimicrob Agents Chemother. 2015;59(12):7911–7914. doi:10.1128/aac.00515-15
140. Adams MD, Nickel GC, Bajaksouzian S, et al. Resistance to colistin in Acinetobacter baumannii associated with mutations in the pmrab two-component system. Antimicrob Agents Chemother. 2009;53(9):3628–3634. doi:10.1128/aac.00284-09
141. Uppalapati SR, Sett A, Pathania R. The outer membrane proteins ompa, caro, and oprd of Acinetobacter baumannii confer a two-pronged defense in facilitating its success as a potent human pathogen. Front Microbiol. 2020;11:589234. doi:10.3389/fmicb.2020.589234
142. Lin MF, Lin YY, Lan CY. Contribution of EmrAB efflux pumps to colistin resistance in Acinetobacter baumannii. J Microbiol. 2017;55:130–136. doi:10.1007/s12275-017-6408-5
143. Chen X, Su S, Yan Y, et al. Anti-Pseudomonas aeruginosa activity of natural antimicrobial peptides when used alone or in combination with antibiotics. Frontiers in Microbiology. 2023;14:
144. Tzeng Y-L, Ambrose KD, Zughaier S, et al. Cationic antimicrobial peptide resistance in neisseria meningitidis. J Bacteriol. 2005;187(15):5387–5396. doi:10.1128/jb.187.15.5387-5396.2005
145. Lei J, Sun L, Huang S, et al. The antimicrobial peptides and their potential clinical applications. Am J Transl Res. 2019;11:3919–3931.
146. Wang Y, Song M, Chang W. Antimicrobial peptides and proteins against drug-resistant pathogens. The Cell Surface. 2024;12:100135. doi:10.1016/j.tcsw.2024.100135
147. Garvey M. Antimicrobial peptides demonstrate activity against resistant bacterial pathogens. Infect Dis Rep. 2023;15:454–469. doi:10.3390/idr15040046
148. Islam T, Tamanna NT, Sagor MS, et al. Antimicrobial peptides: a promising solution to the rising threat of antibiotic resistance. Pharmaceutics. 2024;16(12):1542. doi:10.3390/pharmaceutics16121542
149. Wang Z, Wang X, Wang J. Recent advances in antibacterial and antiendotoxic peptides or proteins from marine resources. Mar Drugs. 2018;16:57. doi:10.3390/md16020057
150. Bhunia A, Mohanram H, Domadia PN, et al. Designed beta-boomerang antiendotoxic and antimicrobial peptides: structures and activities in lipopolysaccharide. J Biol Chem. 2009;284:21991–22004. doi:10.1074/jbc.M109.013573
151. Diamond G, Beckloff N, Weinberg A, et al. The roles of antimicrobial peptides in innate host defense. Curr Pharm Des. 2009;15(21):2377–2392. doi:10.2174/138161209788682325
152. Duarte-Mata DI, Salinas-Carmona MC. Antimicrobial peptides´ immune modulation role in intracellular bacterial infection. Front Immunol. 2023;14. doi:10.3389/fimmu.2023.1119574.
153. Bechinger B, Gorr SU. Antimicrobial Peptides: mechanisms of Action and Resistance. J Dent Res. 2017;96:254–260. doi:10.1177/0022034516679973
154. Hillerich B, Westpheling J. A new GntR family transcriptional regulator in streptomyces coelicolor is required for morphogenesis and antibiotic production and controls transcription of an abc transporter in response to carbon source. J Bacteriol. 2006;188(21):7477–7487. doi:10.1128/jb.00898-06
155. Kraus D, Peschel A. Molecular mechanisms of bacterial resistance to antimicrobial peptides. In: Shafer WM, editor. Antimicrobial Peptides and Human Disease. Berlin Heidelberg: Berlin, Heidelberg: Springer; 2006:231–250.
156. Rivera KG, Tanaka KJ, Buechel ER, et al. Antimicrobial peptide recognition motif of the substrate binding protein SapA from nontypeable haemophilus influenzae. Biochemistry. 2024;63(3):294–311. doi:10.1021/acs.biochem.3c00562
157. Omar ED, Mat H, Abd Karim AZ, et al. Comparative analysis of logistic regression, gradient boosted trees, svm, and random forest algorithms for prediction of acute kidney injury requiring dialysis after cardiac surgery. Int J Nephrol Renovasc Dis. 2024;17:197–204. doi:10.2147/ijnrd.S461028
158. G K, K PI, Hasin AJ, et al. Support vector machines: a literature review on their application in analyzing mass data for public health. Cureus. 2025;17(1):e77169. doi:10.7759/cureus.77169
159. Bajiya N, Choudhury S, Dhall A, et al. AntiBP3: a method for predicting antibacterial peptides against gram-positive/negative/variable bacteria. Antibiotics. 2024;13(2):168. doi:10.3390/antibiotics13020168
160. Aguilera-Puga MDC, Cancelarich NL, Marani MM, et al. Accelerating the discovery and design of antimicrobial peptides with artificial intelligence. Methods Mol Biol. 2024;2714:329–352. doi:10.1007/978-1-0716-3441-7_18
161. Schonlau M, Zou RY. The random forest algorithm for statistical learning. Stata J. 2020;20(1):3–29. doi:10.1177/1536867x20909688
162. Peng S, Rajjou L. Unifying antimicrobial peptide datasets for robust deep learning-based classification. Data Brief. 2024;56:110822. doi:10.1016/j.dib.2024.110822
163. Han B, Ren C, Wang W, et al. Computational prediction of protein intrinsically disordered region related interactions and functions. Genes. 2023:14. doi:10.3390/genes14020432.
164. Chen T, Guestrin C. XGBoost: a scalable tree boosting system. In
165. Chung CR, Chien CY, Tang Y, et al. An ensemble deep learning model for predicting minimum inhibitory concentrations of antimicrobial peptides against pathogenic bacteria. iScience. 2024;27:110718. doi:10.1016/j.isci.2024.110718
166. Zhao Y, Li Q, Sun M, et al. EBAMP: an efficient de novo broad-spectrum antimicrobial peptide discovery framework. Cell Rep. 2025;44:116215. doi:10.1016/j.celrep.2025.116215
167. Weissenow K, Rost B. Are protein language models the new universal key? Curr Opin Struct Biol. 2025;91:102997. doi:10.1016/j.sbi.2025.102997
168. Ching T, Himmelstein DS, Beaulieu-Jones BK, et al. Opportunities and obstacles for deep learning in biology and medicine. J R Soc Interface. 2018:15. doi:10.1098/rsif.2017.0387.
169. Mohsenzadegan K, Tavakkoli V, Kyamakya K. A deep-learning based visual sensing concept for a robust classification of document images under real-world hard conditions. Sensors. 2021;21. doi:10.3390/s21206763
170. Gillani M, Pollastri G. Protein subcellular localization prediction tools. Comput Struct Biotechnol J. 2024;23:1796–1807. doi:10.1016/j.csbj.2024.04.032
171. López-Siles M, Tajuelo A, Caravaca-Fuentes P, et al. Identification and characterization of amphipathic antimicrobial peptides with broad spectrum activity against multi-drug resistant bacteria. Curr Res Microbial Sci. 2025;8:100363. doi:10.1016/j.crmicr.2025.100363
172. Chatzimiltis S, Agathocleous M, Promponas VJ, et al. Post-processing enhances protein secondary structure prediction with second order deep learning and embeddings. Comput Struct Biotechnol J. 2025;27:243–251. doi:10.1016/j.csbj.2024.12.022
173. Lindemann B, Müller T, Vietz H, et al. A survey on long short-term memory networks for time series prediction. Procedia CIRP. 2021;99:650–655. doi:10.1016/j.procir.2021.03.088
174. Liu Q, Jiang Z, Hasan MZ, et al. Bidirectional linear recurrent models for sequence-level multisource fusion. Information Fusion. 2026;126:103601. doi:10.1016/j.inffus.2025.103601
175. Wang G, L M. Machine learning prediction of antimicrobial peptides. Methods Mol Biol. 2022;2405:1–37. doi:10.1007/978-1-0716-1855-4_1
176. Cho K, Merrienboer B, Bahdanau D, et al. Learning Phrase Representations Using RNN Encoder-Decoder for Statistical Machine Translation. 2014. doi:10.3115/v1/D14-1179
177. Devlin J, Chang M-W, Lee K, et al. BERT: pre-training of deep bidirectional transformers for language understanding. 2018.
178. Yu X, Weber IT, Harrison RW. Sparse representation for prediction of hiv-1 protease drug resistance. Proc SIAM Int Conf Data Min. 2013;2013:342–349. doi:10.1137/1.9781611972832.38
179. Mou M, Pan Z, Zhou Z, et al. A transformer-based ensemble framework for the prediction of protein-protein interaction sites. Research. 2023;6:0240. doi:10.34133/research.0240
180. Zhao M, Zhang Y, Wang M, et al. dsAMP and dsAMPGAN: deep learning networks for antimicrobial peptides recognition and generation. Antibiotics. 2024;13(10):948. doi:10.3390/antibiotics13100948
181. Wang Y, Wang L, Li C, et al. AMP-EBiLSTM: employing novel deep learning strategies for the accurate prediction of antimicrobial peptides. Front Genet. 2023;14:1232117. doi:10.3389/fgene.2023.1232117
182. Ferruz N, Schmidt S, Höcker B. ProtGPT2 is a deep unsupervised language model for protein design. Nat Commun. 2022;13(1):4348. doi:10.1038/s41467-022-32007-7
183. Madani A, Krause B, Greene ER, et al. Large language models generate functional protein sequences across diverse families. Nat Biotechnol. 2023;41(8):1099–1106. doi:10.1038/s41587-022-01618-2
184. Strodthoff N, Wagner P, Wenzel M, et al. UDSMProt: universal deep sequence models for protein classification. Bioinformatics. 2020;36(8):2401–2409. doi:10.1093/bioinformatics/btaa003
185. Saleem N, Kumar N, El-Omar E, et al. Harnessing machine learning approaches for the identification, characterization, and optimization of novel antimicrobial peptides. Antibiotics. 2025:14. doi:10.3390/antibiotics14121263.
186. Lin Z, Akin H, Rao R, et al. Evolutionary-scale prediction of atomic-level protein structure with a language model. Science. 2023;379(6637):1123–1130. doi:10.1126/science.ade2574
187. Meier J, Rao R, Verkuil R, et al. Language models enable zero-shot prediction of the effects of mutations on protein function. bioRxiv. 2021;2021. doi:10.1101/2021.07.09.450648
188. Notin P, Dias M, Frazer J, et al. Tranception: protein fitness prediction with autoregressive transformers and inference-time retrieval. Int. Conf. Mach. Learn. 2022.
189. Raffel C, Shazeer N, Roberts A, et al. Exploring the limits of transfer learning with a unified text-to-text transformer. J. Mach. Learn. Res. 2019.
190. Tan C, Sun F, Kong T, et al. A survey on deep transfer learning:
191. Wei CH, Kao HY, Lu Z. GNormPlus: an integrative approach for tagging genes, gene families, and protein domains. Biomed Res Int. 2015;2015:918710. doi:10.1155/2015/918710
192. Chawla N, Bowyer K, Hall L, et al. SMOTE: synthetic minority over-sampling technique. J artif intell res. 2002;16:321–357. doi:10.1613/jair.953
193. Bronstein M, Bruna J, Cohen T, et al. Geometric deep learning: grids, groups, graphs, geodesics, and gauges. arXiv preprint arXiv. 2021.
194. Sun J, Kulandaisamy A, Liu J, et al. Machine learning in computational modelling of membrane protein sequences and structures: from methodologies to applications. Comput Struct Biotechnol J. 2023;21:1205–1226. doi:10.1016/j.csbj.2023.01.036
195. Gligorijević V, Renfrew PD, Kosciolek T, et al. Structure-based protein function prediction using graph convolutional networks. Nat Commun. 2021;12(1):3168. doi:10.1038/s41467-021-23303-9
196. Kipf T, Welling M. Semi-Supervised Classification with Graph Convolutional Networks; 2016. doi:10.48550/arXiv.1609.02907
197. Hao Y, Liu X, Fu H, et al. PGAT-ABPp: harnessing protein language models and graph attention networks for antibacterial peptide identification with remarkable accuracy. Bioinformatics. 2024:40. doi:10.1093/bioinformatics/btae497.
198. Yan K, Lv H, Guo Y, et al. sAMPpred-GAT: prediction of antimicrobial peptide by graph attention network and predicted peptide structure. Bioinformatics. 2023:39. doi:10.1093/bioinformatics/btac715.
199. Chiu YP, Yao L, Tang Y, et al. ANIA: an inception-attention network for predicting minimum inhibitory concentration of antimicrobial peptides. Brief Bioinform. 2026:27. doi:10.1093/bib/bbag023.
200. Sun T-J, Bu H-L, Yan X, et al. LABAMPsGCN: a framework for identifying lactic acid bacteria antimicrobial peptides based on graph convolutional neural network. Front Genetics. 2022;13:2022. doi:10.3389/fgene.2022.1062576
201. Jumper J, Evans R, Pritzel A, et al. Highly accurate protein structure prediction with AlphaFold. Nature. 2021;596(7873):583–589. doi:10.1038/s41586-021-03819-2
202. Varadi M, Anyango S, Deshpande M, et al. AlphaFold protein structure database: massively expanding the structural coverage of protein-sequence space with high-accuracy models. Nucleic Acids Res. 2022;50(D1):D439–d444. doi:10.1093/nar/gkab1061
203. Fu Y. Advances in the application of deep learning for antimicrobial peptide screening. Agri Sci Food Process. 2026;3(2):49–72. doi:10.62762/ASFP.2026.121905
204. Goodfellow I, Warde-Farley D, Pouget-Abadie J, et al. Generative Adversarial Nets. ArXiv. 2014.
205. Zervou MA, Doutsi E, Pantazis Y, et al. De novo antimicrobial peptide design with feedback generative adversarial networks. Int J Mol Sci. 2024:25. doi:10.3390/ijms25105506.
206. Vivekananthan S. Comparative analysis of generative models: enhancing image synthesis with vaes, gans, and stable diffusion. arXiv preprint arXiv. 2024.
207. Dean SN, Alvarez JAE, Zabetakis D, et al. PepVAE: variational autoencoder framework for antimicrobial peptide generation and activity prediction. Front. Microbiol. 2021;12:
208. Lee J-S. Evaluating generative patent language models. World Patent Inf. 2023;72:102173. doi:10.1016/j.wpi.2023.102173
209. Ho J, Jain A, Abbeel P. Denoising diffusion probabilistic models. Adv. Neural Inf. Process. Syst. 2020.
210. Wang Y, Song M, Liu F, et al. Artificial intelligence using a latent diffusion model enables the generation of diverse and potent antimicrobial peptides. Sci Adv. 2025;11(6):eadp7171. doi:10.1126/sciadv.adp7171
211. Popova M, Isayev O, Tropsha A. Deep reinforcement learning for de novo drug design. Sci Adv. 2018;4(7):eaap7885. doi:10.1126/sciadv.aap7885
212. Sampa MB, Aziz NHA. Application of reinforcement learning techniques in de novo drug design: a systematic literature review. Health Sci Rep. 2026;9(3):e72132. doi:10.1002/hsr2.72132
213. Eckhart L, Lenhof K, Herrmann L, et al. How to predict effective drug combinations – moving beyond synergy scores. iScience. 2025;28(6):112622. doi:10.1016/j.isci.2025.112622
214. Viruly L, Suhartono MT, Nurilmala M, et al. Identification and characterization of antimicrobial peptide (AMP) candidate from Gonggong Sea Snail (Leavistrombus turturella) extract. J Food Sci Technol. 2023;60(1):44–52. doi:10.1007/s13197-022-05585-z
215. Powers D. Evaluation: from Precision, Recall and F-Factor to ROC, Informedness, Markedness & Correlation. Mach Learn Technol. 2008;2.
216. Chicco D, Jurman G. The advantages of the Matthews correlation coefficient (MCC) over F1 score and accuracy in binary classification evaluation. BMC Genomics. 2020;21(1):6. doi:10.1186/s12864-019-6413-7
217. Bajiya N, Kumar N, Raghava GPS. Prediction of inhibitory peptides against E.coli with desired MIC value. Sci Rep. 2025;15(1):4672. doi:10.1038/s41598-025-86638-z
218. Ibisanmi TA, Jiang X, Willcox M, et al. Recent advances in computational antimicrobial peptide discovery through big data, modeling, and artificial intelligence and their interplay in ushering the next golden era of drug development. Front Bioinform. 2026;6. doi:10.3389/fbinf.2026.1749404
219. Yao L, Pang Y, Wan J, et al. ABPCaps: a novel capsule network-based method for the prediction of antibacterial peptides. Appl Sci. 2023;13(12):6965. doi:10.3390/app13126965
220. Bhadra P, Yan J, Li J, et al. AmPEP: sequence-based prediction of antimicrobial peptides using distribution patterns of amino acid properties and random forest. Sci Rep. 2018;8(1):1697. doi:10.1038/s41598-018-19752-w
221. Yan J, Bhadra P, Li A, et al. Deep-AmPEP30: improve short antimicrobial peptides prediction with deep learning. Mol Ther Nucleic Acids. 2020;20:882–894. doi:10.1016/j.omtn.2020.05.006
222. Xiao X, Shao YT, Cheng X, et al. iAMP-CA2L: a new CNN-BiLSTM-SVM classifier based on cellular automata image for identifying antimicrobial peptides and their functional types. Brief Bioinform. 2021:22. doi:10.1093/bib/bbab209.
223. Veltri D, Kamath U, Shehu A, Hancock J. Deep learning improves antimicrobial peptide recognition. Bioinformatics. 2018;34(16):2740–2747. doi:10.1093/bioinformatics/bty179
224. Yan J, Zhang B, Zhou M, et al. A deep learning method for predicting the minimum inhibitory concentration of antimicrobial peptides against Escherichia coli using Multi-Branch-CNN and Attention. mSystems. 2023;8:e0034523. doi:10.1128/msystems.00345-23
225. Van Oort CM, Ferrell JB, Remington JM, et al. AMPGAN v2: machine Learning-Guided Design of Antimicrobial Peptides. J Chem Inf Model. 2021;61(5):2198–2207. doi:10.1021/acs.jcim.0c01441
226. Das P, Zhao P, Perera D, et al. Molecular dynamics-guided material model for the simulation of shock-induced pore collapse in β-octahydro-1,3,5,7-tetranitro-1,3,5,7-tetrazocine (β-HMX). J Appl Phys. 2021:130. doi:10.1063/5.0056560.
227. Wang X-F, Tang J-Y, Sun J, et al. ProT-diff: a modularized and efficient strategy for de novo generation of antimicrobial peptide sequences by integrating protein language and diffusion models. Advanced Science. 2024;11(43):2406305. doi:10.1002/advs.202406305
228. Porto WF, Fensterseifer ICM, Ribeiro SM, et al. Joker: an algorithm to insert patterns into sequences for designing antimicrobial peptides. Biochim Biophys Acta Gen Subj. 2018;1862:2043–2052. doi:10.1016/j.bbagen.2018.06.011
229. Yoshida M, Hinkley T, Tsuda S, et al. Using evolutionary algorithms and machine learning to explore sequence space for the discovery of antimicrobial peptides. Chem. 2018:4. doi:10.1016/j.chempr.2018.01.005.
230. Hossain H, Ahmed S. Hospital wastewater microbiome & resistome: an overlooked one health crisis driving superbugs in low- and middle-income countries (LMICs). New Microbes New Infect. 2025;68:101653. doi:10.1016/j.nmni.2025.101653
231. Cai J, Yan J, Un C, et al. BERT-AmPEP60: a BERT-based transfer learning approach to predict the minimum inhibitory concentrations of antimicrobial peptides for Escherichia coli and Staphylococcus aureus. J Chem Inf Model. 2025;65(7):3186–3202. doi:10.1021/acs.jcim.4c01749
232. Li W, Huang B, Guo M, et al. Unveiling the evolution of antimicrobial peptides in gut microbes via foundation-model-powered framework. Cell Reports. 2025:44. doi:10.1016/j.celrep.2025.115773.
233. King AM, Zhang Z, Glassey E, et al. Systematic mining of the human microbiome identifies antimicrobial peptides with diverse activity spectra. Nature Microbiology. 2023;8(12):2420–2434. doi:10.1038/s41564-023-01524-6
234. Haryini S, Manku KK, Deshkar A, et al. Chapter One - Antimicrobial peptides: bioinformatic advances and translational therapeutics to combat antibiotic resistance. In: Donev R, editor. Advances in Protein Chemistry and Structural Biology. Academic Press; 2026. 1–59.
235. Ali W, Elsahn A, Ting DSJ, et al. Host DEFENCE PEPTIDES: A POTENT ALTERNATIVE TO COMBAT ANTIMICROBIAL RESISTANCE IN THE ERA of the COVID-19 Pandemic. Antibiotics. 2022:11. doi:10.3390/antibiotics11040475.
236. Rathore AS, Kumar N, Choudhury S, et al. Prediction of hemolytic peptides and their hemolytic concentration. Commun Biol. 2025;8(1):176. doi:10.1038/s42003-025-07615-w
237. Afzal S, Bajwa M, Haroon MS, et al. Unequal budgeting and limited WHO AWaRe Access antibiotics in Punjab: procurement policy gaps in Pakistan’s National Action Plan against antimicrobial resistance. Arch Public Health. 2025;83(1):259. doi:10.1186/s13690-025-01720-2
238. Ngashangva N, Huidrom S, Devi IS. Antimicrobial peptides: natural templates for next-generation therapeutics against antimicrobial resistance. Front Cell Infect Microbiol. 2026;15:
239. Seal S, Mahale M, García-Ortegón M, et al. Machine learning for toxicity prediction using chemical structures: pillars for success in the real world. Chem Res Toxicol. 2025;38(5):759–807. doi:10.1021/acs.chemrestox.5c00033
240. Nilsson JB, Greenbaum J, Peters B, et al. NetMHCpan-4.2: improved prediction of CD8+ epitopes by use of transfer learning and structural features. Front Immunol. 2025;16:
241. Wang H, Chen M, Zhang X, et al. Peptide-based PROTACs: current Challenges and Future Perspectives. Curr Med Chem. 2024;31(2):208–222. doi:10.2174/0929867330666230130121822
242. Adrio J-L, Demain AL. Recombinant organisms for production of industrial products. Bioeng Bugs. 2010;1(2):116–131. doi:10.4161/bbug.1.2.10484
243. Newkirk SE, Kelly JJ, Hourn N, et al. Mapping the modification landscape of mhc-i epitopes: a framework for immunogenic peptidomimetic antigen design. bioRxiv. 2026. doi:10.64898/2026.03.06.710184
244. Waibel I, Schneider TN, Fischer FJ, et al. Bayesian optimization for efficient multiobjective formulation development of biologics. Mol Pharm. 2025;22(11):6636–6645. doi:10.1021/acs.molpharmaceut.5c00591
245. Franco P, Mullot R, Owczarek V. Pareto multi-objective optimization for high locality-preserving space-filling curve identification. Swarm Evol Comput. 2025;92:101797. doi:10.1016/j.swevo.2024.101797
246. Wan F, Kontogiorgos-Heintz D, de la Fuente-Nunez C. Deep generative models for peptide design. Digital Discovery. 2022;1(3):195–208. doi:10.1039/d1dd00024a
247. Wagner FL, Sagmeister P, Tampone TG, et al. Self-optimizing flow reactions for sustainability: an experimental bayesian optimization study. ACS Sustainable Chem. Eng. 2024;12(26):10002–10010. doi:10.1021/acssuschemeng.4c03253
248. Alzain M, Daghistani H, Shamrani T, et al. Antimicrobial peptides: mechanisms, applications, and therapeutic potential. Infect Drug Resist. 2025;18:4385–4426. doi:10.2147/idr.S514825
249. Enninful GN, Kuppusamy R, Tiburu E, et al. Assessing the effect of substitution of arginine with homoarginine in two antimicrobial peptides. Int. J. Pept. Res. Ther. 2026;32(2):24. doi:10.1007/s10989-026-10804-8
250. Johnson TS, Bourdine AA, Deber CM. Hydrophobic moment drives penetration of bacterial membranes by transmembrane peptides. J Biol Chem. 2023;299. doi:10.1016/j.jbc.2023.105266
251. Ye X, Cui N, Ou W, et al. Explainable optimized 3D-MoRSE descriptors for the power conversion efficiency prediction of molecular passivated perovskite solar cells through machine learning. J Mater Chem A. 2024;12(38):26224–26233. doi:10.1039/D4TA03547J
252. Zhang Q, Elkins CA. Antimicrobial peptides: from discovery to developmental applications. Applied and Environmental Microbiology. 2025;91(4):e02115–24. doi:10.1128/aem.02115-24
253. Stavrogiannis C, Sofos F, Karakasidis TE. Machine learning-enhanced molecular dynamics: current state, challenges and perspectives. Arch Comput Methods Eng. 2026;33(4):6125–6147. doi:10.1007/s11831-026-10505-x
254. Huo X, Xu J, Xu M, et al. An improved 3D quantitative structure-activity relationships (QSAR) of molecules with CNN-based partial least squares model. Artificial Intelligence in the Life Sciences. 2023;3:100065. doi:10.1016/j.ailsci.2023.100065
255. Klebe G, Abraham U, Mietzner T. Molecular similarity indices in a comparative analysis (CoMSIA) of drug molecules to correlate and predict their biological activity. J Med Chem. 1994;37(24):4130–4146. doi:10.1021/jm00050a010
256. Stokes JM, Yang K, Swanson K, et al. A deep learning approach to antibiotic discovery. Cell. 2020;180(4):688–702.e13. doi:10.1016/j.cell.2020.01.021
257. Allec SI, Ziatdinov M. Active and transfer learning with partially Bayesian neural networks for materials and chemicals††Electronic supplementary information (ESI) available. See Digital Discovery. 2025;4(5):1284–1297. doi:10.1039/D5DD00027K
258. Zhang H, Shi Y, Wang Y, et al. Biological sequence representation methods and recent advances: a review. Biology. 2025:14. doi:10.3390/biology14091137
259. Zhou Q, Sun L, Ou Y, et al. Artificial intelligence drives the identification and screening of novel antibiotics and antimicrobial peptides. Briefings Bioinf. 2026:27. doi:10.1093/bib/bbag107.
260. Schneider G. Automating drug discovery. Nat Rev Drug Discov. 2018;17(2):97–113. doi:10.1038/nrd.2017.232
261. Siqueira-Neto JL, Wicht KJ, Chibale K, et al. Antimalarial drug discovery: progress and approaches. Nat Rev Drug Discov. 2023;22(10):807–826. doi:10.1038/s41573-023-00772-9
262. Wongchai M, Wongkaewkhiaw S, Kanthawong S, et al. Dual-function antimicrobial-antibiofilm peptide hybrid to tackle biofilm-forming Staphylococcus epidermidis. Ann Clinic Microbiol Antimicrob. 2024;23(1):44. doi:10.1186/s12941-024-00701-7
263. Almatroudi A. Biofilm resilience: molecular mechanisms driving antibiotic resistance in clinical contexts. Biology. 2025;14:165. doi:10.3390/biology14020165
264. Park K-H, Jung J, Shin SU, et al. Preclinical and clinical evaluation of vancomycin plus delpazolid combination therapy for MRSA bacteremia: a multicenter, double-blinded, randomized, parallel design, phase IIa clinical trial. Microbiol Spectr. 2026;14(4):e0336125. doi:10.1128/spectrum.03361-25
265. Zhang H, Lv J, Ma Z, et al. Advances in antimicrobial peptides: mechanisms, design innovations, and biomedical potential. Molecules. 2025;30(7):1529. doi:10.3390/molecules30071529
266. Moini Jazani A, Ayati MH, Nadiri AA, et al. Efficacy of hydrotherapy, spa therapy, and balneotherapy for psoriasis and atopic dermatitis: a systematic review. Int J Dermatol. 2023;62(2):177–189. doi:10.1111/ijd.16080
267. Wang B, Lin P, Zhong Y, et al. Explainable deep learning and virtual evolution identifies antimicrobial peptides with activity against multidrug-resistant human pathogens. Nat Microbiol. 2025;10(2):332–347. doi:10.1038/s41564-024-01907-3
268. Giacometti A, Cirioni O, Gov Y, et al. RNA III inhibiting peptide inhibits in vivo biofilm formation by drug-resistant Staphylococcus aureus. Antimicrob Agents Chemother. 2003;47(6):1979–1983. doi:10.1128/aac.47.6.1979-1983.2003
269. François P, Schrenzel J, Götz F. Biology and regulation of staphylococcal biofilm. Int J Mol Sci. 2023;24(6):5218. doi:10.3390/ijms24065218
270. Gao H, Guan F, Luo B, et al. DLFea4AMPGen de novo design of antimicrobial peptides by integrating features learned from deep learning models. Nat Commun. 2025;16(1):9134. doi:10.1038/s41467-025-64378-y
271. Bonifacio-Velez de Villa EI, Montoya-Alfaro ME, Negrón-Ballarte LP, et al. Predicting antimicrobial peptide activity: a machine learning-based quantitative structure-activity relationship approach. Pharmaceutics. 2025:17. doi:10.3390/pharmaceutics17080993.
272. Rodriguez-Leal CM, González-Corralejo C, Candel FJ, et al. Towards a holistic approach to pulmonary infections.
273. Franza M, Varricchio R, Alloisio G, et al. Zebrafish (Danio rerio) as a Model System to Investigate the Role of the Innate Immune Response in Human Infectious Diseases. Int J Mol Sci. 2024;25(22):12008. doi:10.3390/ijms252212008
274. Lavertu A, Corbeil J, Germain P. QMAP: a benchmark for standardized evaluation of antimicrobial peptide mic and hemolytic activity regression. bioRxiv. 2026;2026. doi:10.64898/2026.02.03.703041.
275. Wan F, Wong F, Collins JJ, et al. Machine learning for antimicrobial peptide identification and design. Nat Rev Bioeng. 2024;2(5):392–407. doi:10.1038/s44222-024-00152-x
276. Torrent Rodríguez A, Font IBA, Barrantes González M, et al. Clinically important pharmacokinetic drug-drug interactions with antibacterial agents. Rev Esp Quimioter. 2024;37(4):299–322. doi:10.37201/req/037.2024
277. Rowland M, Thomas NT. Clinical Pharmacokinetics: Concepts and Applications, 4e. Lippincott Williams & Wilkins; 2010.
278. Jang YE, Kwon M, Kwon CW, et al. Integrative peptide drug development: chemical engineering, ai-driven design, and cell-penetrating peptides. Pharmaceutics. 2026;18(5):537. doi:10.3390/pharmaceutics18050537
279. Das P, Sercu T, Wadhawan K, et al. Accelerated antimicrobial discovery via deep generative models and molecular dynamics simulations. Nat Biomed Eng. 2021;5(6):613–623. doi:10.1038/s41551-021-00689-x
280. Torres MDT, Wan F, de la Fuente-Nunez C. Deep learning reveals antibiotics in the archaeal proteome. Nat Microbiol. 2025;10(9):2153–2167. doi:10.1038/s41564-025-02061-0
281. Mookherjee N, Anderson MA, Haagsman HP, et al. Antimicrobial host defence peptides: functions and clinical potential. Nat Rev Drug Discov. 2020;19(5):311–332. doi:10.1038/s41573-019-0058-8
282. Greber KE, Dawgul M. Antimicrobial Peptides Under Clinical Trials. Curr Top Med Chem. 2016;17(5):620–628. doi:10.2174/1568026616666160713143331
283. Ali W, Elsahn A, Ting DSJ, et al. Host defence peptides: a potent alternative to combat antimicrobial resistance in the era of the COVID-19 Pandemic. Antibiotics. 2022;11(4):475. doi:10.3390/antibiotics11040475
284. De Groot AS, Scott DW. Immunogenicity of protein therapeutics. Trends Immunol. 2007;28(11):482–490. doi:10.1016/j.it.2007.07.011
285. Perron GG, Zasloff M, Bell G. Experimental evolution of resistance to an antimicrobial peptide. Proc Biol Sci. 2006;273(1583):251–256. doi:10.1098/rspb.2005.3301
286. Andersson DI, Hughes D, Kubicek-Sutherland JZ. Mechanisms and consequences of bacterial resistance to antimicrobial peptides. Drug Resist Updat. 2016;26:43–57. doi:10.1016/j.drup.2016.04.002
287. Lazzaro BP, Zasloff M, Rolff J. Antimicrobial peptides: application informed by evolution. Science. 2020;368. doi:10.1126/science.aau5480
288. Butler MS, Henderson IR, Capon RJ, et al. Antibiotics in the clinical pipeline as of December 2022. J Antibiot. 2023;76:431–473. doi:10.1038/s41429-023-00629-8
289. Budhwani Z, Buragina JT, Lang J, et al. Characterization of the novel leaderless bacteriocin, bawcin, from bacillus wiedmannii. Int J Mol Sci. 2023;24(23):16965. doi:10.3390/ijms242316965
290. Ceballos Benavides M, Castillo Muñoz J, Marcillo Villota K, et al. Enhancing the antitumor efficacy of nisin through advanced nanosystems: a systematic review of in vitro studies. Pharmaceuticals. 2026;19(4):611. doi:10.3390/ph19040611
291. Fodeke SD, Musa EA, Ishaya I. Assessment of antimicrobial stewardship practices using process measures in a nigerian tertiary hospital: a retrospective study. Antimicrobial Stewardship Healthcare Epidemiol. 2026;6(1):e75. doi:10.1017/ash.2026.10343
292. Hancock RE, Sahl HG. Antimicrobial and host-defense peptides as new anti-infective therapeutic strategies. Nat Biotechnol. 2006;24:1551–1557. doi:10.1038/nbt1267
293. Zimmer J, Hobkirk J, Mohamed F, et al. On the functional overlap between complement and anti-microbial peptides. Front Immunol. 2015;5. doi:10.3389/fimmu.2014.00689
294. Ju Y, Weng L, Lin T, et al. Advances of antimicrobial peptides in the treatment of multidrug-resistant bacteria. BioDrugs. 2025;39(6):899–922. doi:10.1007/s40259-025-00740-2
295. Goldberg K, Lobov A, Antonello P, et al. Cell-autonomous innate immunity by proteasome-derived defence peptides. Nature. 2025;639(8056):1032–1041. doi:10.1038/s41586-025-08615-w
296. Ren J, Yan G, Yang L, et al. Cancer chemoprevention: signaling pathways and strategic approaches. Signal Transduct Target Ther. 2025;10:113. doi:10.1038/s41392-025-02167-1
297. Namburu JR, Rajendra Sanosh AB, Poosarla CS, et al. Streptococcus M utans-Specific Antimicrobial Peptide C16G2-Mediated Caries Prevention: a Review. Front Dent. 2022;19:17. doi:10.18502/fid.v19i17.9963
298. Kaplan CW, Sim JH, Shah KR, et al. Selective membrane disruption: mode of action of C16G2, a specifically targeted antimicrobial peptide. Antimicrob Agents Chemother. 2011;55(7):3446–3452. doi:10.1128/aac.00342-11
299. Mercer DK, Robertson JC, Miller L, et al. NP213 (Novexatin®): a unique therapy candidate for onychomycosis with a differentiated safety and efficacy profile. Med Mycol. 2020;58(8):1064–1072. doi:10.1093/mmy/myaa015
300. Fu J, Zong X, Jin M, et al. Mechanisms and regulation of defensins in host defense. Signal Transduct Target Ther. 2023;8(1):300. doi:10.1038/s41392-023-01553-x
301. Naylor NR, Hasso-Agopsowicz M, Kim C, et al. The global economic burden of antibiotic-resistant infections and the potential impact of bacterial vaccines: a modelling study. BMJ Glob Health. 2025;10(6):e016249. doi:10.1136/bmjgh-2024-016249
302. Perfilyev A, Gæde A, Hooton S, et al. Large-scale analysis of bacterial genomes reveals thousands of lytic phages. Nat Microbiol. 2026;11:42–52. doi:10.1038/s41564-025-02203-4
303. Jülke EM, Beck-Sickinger AG. Peptide therapeutics: current status and future opportunity with focus on nose-to-brain delivery☆. Peptides. 2025;188:171404. doi:10.1016/j.peptides.2025.171404
304. Morowitz MJ, Babrowski T, Carlisle EM, et al. The human microbiome and surgical disease. Ann Surg. 2011;253(6):1094–1101. doi:10.1097/SLA.0b013e31821175d7
305. Zheng S, Tu Y, Li B, et al. Antimicrobial peptide biological activity, delivery systems and clinical translation status and challenges. J Transl Med. 2025;23(1):292. doi:10.1186/s12967-025-06321-9
306. Alo B, Lamers C. Crossing barriers: advancements in macromolecular therapeutics for neurodegenerative diseases and strategies to overcome the blood-brain barrier. ACS Pharmacol Transl Sci. 2025;8(8):2353–2383. doi:10.1021/acsptsci.5c00153
307. Faltus T. The Medicinal Phage-Regulatory Roadmap For Phage Therapy Under Eu Pharmaceutical Legislation. Viruses. 2024;16(3):443. doi:10.3390/v16030443
308. Hashim NT, Babiker R, Rahman MM, et al. Targeting bacterial infections in periodontal disease: from conventional antibiotics to next-generation therapeutics. Antibiotics. 2026;15(4):397. doi:10.3390/antibiotics15040397
309. Sharma D, Dhiman I, Das S, et al. Recent advances in therapeutic peptides: innovations and applications in treating infections and diseases. ACS Omega. 2025;10(17):17087–17107. doi:10.1021/acsomega.5c02077
310. Price R. O’Neill report on antimicrobial resistance: funding for antimicrobial specialists should be improved. Eur J Hosp Pharm. 2016;23(4):245–247. doi:10.1136/ejhpharm-2016-001013
311. Joseph J, Boby S, Mooyottu S, et al. Antibiotic potentiators as a promising strategy for combating antibiotic resistance. Npj Antimicrobials and Resistance. 2025;3(1):53. doi:10.1038/s44259-025-00112-4
312. Ferreira PM, Silva FFMD, Santos JSD, et al. The therapeutic potential of antimicrobial peptides isolated from the skin secretions of anurans of the genus boana in the face of the global antimicrobial resistance crisis. Toxins. 2025;17:312. doi:10.3390/toxins17070312
313. Peng H, Chen IA, Qimron U. Engineering phages to fight multidrug-resistant bacteria. Chemical Reviews. 2025;125:933–971. doi:10.1021/acs.chemrev.4c00681
314. Santos-Júnior CD, Torres MDT, Duan Y, et al. Discovery of antimicrobial peptides in the global microbiome with machine learning. Cell. 2024;187:3761–3778.e16. doi:10.1016/j.cell.2024.05.013
315. Veiga A, Ribeiro V, Ramírez-Jiménez RA, et al. Biofunctional silk sericin hydrogels: a versatile platform with potential for tissue healing and regeneration. Colloids Surf B. 2025;255:114916. doi:10.1016/j.colsurfb.2025.114916
316. Lin C, Xiong S, Cui F, et al. Deep learning in antimicrobial peptide prediction. J Chem Inf Model. 2025;65(14):7373–7392. doi:10.1021/acs.jcim.5c00530
317. Hegde M, Nebel J-C, Rahman F. Language modelling techniques for analysing the impact of human genetic variation. Bioinf Biol Insights. 2025;19:11779322251358314. doi:10.1177/11779322251358314
318. Wu W, Chen H, Liu Z, et al. Machine learning-based QSPR modeling for predicting the n-octanol/air partition coefficient of polybrominated diphenyl ethers. iScience. 2026:29. doi:10.1016/j.isci.2026.115393.
319. Miao H, Wang Z, Chen S, et al. Application of machine learning in the discovery of antimicrobial peptides: exploring their potential for ulcerative colitis therapy. bioRxiv. 2025;2025. doi:10.1101/2025.06.17.660148
320. Lundberg SM, Erion G, Chen H, et al. From local explanations to global understanding with explainable ai for trees. Nat Mach Intell. 2020;2(1):56–67. doi:10.1038/s42256-019-0138-9
321. Tanifuji R, Oguri H. Chemo-enzymatic total synthesis: current approaches toward the integration of chemical and enzymatic transformations. Beilstein J Org Chem. 2024;20:1693–1712. doi:10.3762/bjoc.20.151
322. Chumpol W, Lunha K, Jiemsup S, et al. In vitro synergistic effects of antibiotic combinations against multidrug-resistant Streptococcus suis from diseased pigs. Antibiotics. 2026:15. doi:10.3390/antibiotics15020136.
323. Talapko J, Meštrović T, Juzbašić M, et al. Antimicrobial peptides-mechanisms of action, antimicrobial effects and clinical applications. Antibiotic). 2022:11. doi:10.3390/antibiotics11101417.
324. Riemer A, Freund V. Generative artificial intelligence in pharmaceutical drug development: a systematic review of time and cost efficiency across discovery, preclinical, and clinical phases. Intelligent Pharmacy. 2026;4(2):145–158. doi:10.1016/j.ipha.2025.12.006
325. Krones F, Marikkar U, Parsons G, et al. Review of multimodal machine learning approaches in healthcare. Inf Fusion. 2025;114:102690. doi:10.1016/j.inffus.2024.102690
326. Roder M, Pereira CR, Papa JP, et al. Vision transformers as SOTA models for lithological classification of Brazilian pre-salt rocks. Appl Comput Geosci. 2025;28:100301. doi:10.1016/j.acags.2025.100301
327. Wei J, Tay Y, Bommasani R, et al. Emergent abilities of large language models. arXiv preprint arXiv:2206.07682. 2022.
328. Qin L, Chen Q, Zhou Y, et al. A survey of multilingual large language models. Patterns (N Y). Patterns (New York, N.Y.). 2025;6(1):101118. doi:10.1016/j.patter.2024.101118
329. Hanna CC, Hermant YO, Harris PWR, et al. Discovery, synthesis, and optimization of peptide-based antibiotics. Acc Chem Res. 2021;54:1878–1890. doi:10.1021/acs.accounts.0c00841
330. Dharmasivam M, Kaya B, Akinware A, et al. Leading artificial intelligence–driven drug discovery platforms: 2025 landscape and global outlook. Pharmacol Rev. 2026;78:100102. doi:10.1016/j.pharmr.2025.100102
331. Tristancho-Baró A, Franco-Fobe LE, Ariza MP, et al. Genomic characterization of carbapenemase-producing enterobacteriaceae from clinical and epidemiological human samples. Antibiotics. 2025;14:42. doi:10.3390/antibiotics14010042
332. Singla R, Khan S, Silsarma A, et al. Bedaquiline resistance and treatment outcomes among patients with tuberculosis previously exposed to bedaquiline in India: a multicentric retrospective cohort study. Clin Infect Dis. 2025;81:846–852. doi:10.1093/cid/ciaf068
333. Punetha A, Kotiya D. Advancements in oncoproteomics technologies: treading toward translation into clinical practice. Proteomes. 2023;11:2. doi:10.3390/proteomes11010002
334. Kryshtafovych A, Schwede T, Topf M, et al. Critical assessment of methods of protein structure prediction (CASP)—Round XIV. Proteins. 2021;89(12):1607–1617. doi:10.1002/prot.26237
335. Rieke N, Hancox J, Li W, et al. The future of digital health with federated learning. NPJ Digit Med. 2020;3(1):119. doi:10.1038/s41746-020-00323-1
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Predicting Antibiotic Resistance in ICUs Patients by Applying Machine Learning in Vietnam
Tran Quoc V, Nguyen Thi Ngoc D, Nguyen Hoang T, Vu Thi H, Tong Duc M, Do Pham Nguyet T, Nguyen Van T, Ho Ngoc D, Vu Son G, Bui Duc T
Infection and Drug Resistance 2023, 16:5535-5546
Published Date: 22 August 2023
Application of Antimicrobial Peptides in Wound Dressings
Zhu A, Chen B, Ma J, Wang J, Tang R, Liu L, Sun W, Zheng X, Pan G
Drug Design, Development and Therapy 2025, 19:8523-8539
Published Date: 19 September 2025