Back to Journals » Journal of Inflammation Research » Volume 19
Utilizing High-Throughput Screening of dHL-60 Inflammatory Behaviors to Evaluate Honey-Derived Molecules’ Potential as Immunomodulatory Biomaterial Additives
Authors Main EN ![]() , Huang JC, Bowlin GL
, Huang JC, Bowlin GL ![]()
Received 22 October 2025
Accepted for publication 14 February 2026
Published 24 February 2026 Volume 2026:19 564685
DOI https://doi.org/10.2147/JIR.S564685
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Shouya Feng
Evan N Main, James C Huang, Gary L Bowlin
Department of Biomedical Engineering, University of Memphis, Memphis, TN, 38152, USA
Correspondence: Gary L Bowlin, Email [email protected]
Introduction: The ability of honey varieties to both prevent infection and prevent a dysregulated immune reaction to an implanted biomaterial provides a strong case for their viability as biomaterial additives. However, the mechanisms, components, and concentration ranges underlying honey’s anti-inflammatory activity remain incompletely understood. The present study sought to screen serial concentrations of the common flavonoid and phenolic components: pinobanksin, pinocembrin, chrysin, and methyl syringate to determine concentrations best to reduce intracellular reactive oxygen species (ROS) activity and neutrophil extracellular trap (NET) release (NETosis) in a differentiated HL60 (dHL60) model.
Methods: HL60s were differentiated into neutrophil-like cells (dHL60s) using a validated protocol, and were cultured with concentrations ranging from 1 nM to 1 mM of each flavonoid and from 10 μM to 2 mM of methyl syringate. NETosis and ROS were measured via Sytox Orange staining and DCFH-DA assay.
Results: ROS activity was moderately inhibited by chrysin but increased by methyl syringate. Flavonoids failed to reduce NET levels. Methyl syringate significantly reduced NETosis in a dose-dependent manner, but increased ROS.
Discussion: The present study provides proof of concept for the honey-derived phenolic compound methyl syringate as a therapeutic candidate to reduce neutrophil-mediated inflammation, specifically NETosis, in response to implanted biomaterials. Methyl syringate is highlighted because NETosis can profoundly affect the progression of downstream inflammatory responses in neutrophils arriving at the site and in other cell types within the microenvironment. Furthermore, it proposes a high-throughput method to screen for potential therapeutic compounds prior to primary neutrophil investigations, thereby addressing the current lack of neutrophil-targeted drug discovery efforts.
Keywords: biomaterial additives, HL60, host-biomaterial response, NETosis, neutrophil, ROS
Introduction
Host-Biomaterial Response
Neutrophils are the most abundant leukocyte and the primary drivers of the acute phase of inflammation.1 While previously thought to be merely short-lived phagocytic effector cells, neutrophils are now recognized to encompass a broad range of phenotypes and to perform specialized functions in both inflammation and tissue remodeling.2 Upon injury, infection, or biomaterial implantation, neutrophils release pro-inflammatory cytokines and chemokines, attracting other immune cells and a surge of additional neutrophils at the implantation site. This pro-inflammatory signaling cascade also induces pro-inflammatory phenotypes in neutrophils arriving at the site, thereby further amplifying pro-inflammatory signaling and perpetuating a vicious cycle that sustains unresolved acute inflammation.3 A unique and specialized cellular process, NETosis, occurs in a subset of neutrophils, in which they extrude their intracellular contents and eject NETs composed of histones and intertwined chromatin fibers. Coated with proteins from neutrophil granules, these chromatin fibers isolate and facilitate the degradation of pathogens and foreign bodies. NETosis induces the release of enzymes, matrix metalloproteinases, and signals, including damage-associated molecular patterns (DAMPs) with degradative and pro-inflammatory qualities.4,5 Free radicals, including ROS, oxidize bacterial membranes and degrade foreign materials, and are also potently generated and released by neutrophils.6,7 The NET-forming and ROS-generating abilities of neutrophils confer high efficacy in protecting the host against invasive pathogens. However, acceptance and integration of an implanted biomaterial can be disrupted when the neutrophil-mediated acute inflammatory response becomes dysregulated, leading to NET release that encapsulates the biomaterial surface and, by forming a chromatin-nanofiber barrier, impedes host-device integration.8 This disproportionate neutrophil response during biomaterial implantation adversely affects the microenvironment at the host tissue-implant interface. It spurs the transition from acute inflammation to pathological chronic inflammation. Thus, before biomaterials or therapeutic additives are considered fully biocompatible, an evaluation of the acute inflammatory response they elicit is warranted. Unless the research is within the niche of neutrophil inflammation, this vital evaluation is often unexplored or overlooked, leading to the production of “biocompatible” biomaterials that are unable to perform their intended functions because the acute inflammatory phase is improperly resolved.9 The present study aimed first to establish a high-throughput, reproducible, and minimally burdensome initial screening procedure for therapeutic agents, and then to screen four promising bioactive molecules to determine which are capable of reducing neutrophil-mediated inflammatory behaviors and at what therapeutic concentrations.
Honey: Special Types and Recent Research
In recent decades, research into the potential of honey as a wound-healing agent has increased. Several types of honey, originating from around the world, have recently been investigated for purported medicinal and wound-healing applications. While the many types of honey found to be beneficial are of great note, for brevity, the authors suggest the review paper and meta-analysis: “Biological properties and therapeutic activities of honey in wound healing: A narrative review and meta-analysis”.10 For example, Manuka honey, derived from the Leptospermum scoparium shrub native to New Zealand, has received special attention. Like other honey types, it contains antioxidant compounds, and its high sugar content confers an osmotic potential that can promote wound debridement. However, unlike other types of honey, it contains, at a high concentration, a potent antibacterial agent, methylglyoxal, which is effective against certain antibiotic-resistant bacteria. Combined, the properties of Manuka honey make it a promising agent for wound healing and infection prevention in the wound microenvironment.11 In recent years, Manuka honey has also exhibited anti-inflammatory properties, reducing neutrophil-mediated inflammation within a therapeutic window. Notably, the anti-inflammatory activity of Manuka honey is retained even when embedded in electrospun biomaterial fibers.12,13 For a comprehensive review of honey as a wound healing or biomaterial additive, and the factors that make Manuka honey a promising honey type, the authors suggest “Honey-Based Templates in Wound Healing and Tissue Engineering” by Benjamin Minden Birkenmeier et. al.11
Previous studies using HL60 models have shown a dose-dependent decrease in ROS generation, chemotaxis, pro-inflammatory cytokine release, and upstream signaling markers in response to co-culture with Manuka honey. Additionally, Manuka honey was incorporated into PDO tissue-engineering templates at concentrations of 0.1, 1, or 10% (v/v). This study found that when incorporated into the PDO template, Manuka honey reduced NET formation by up to 75%. However, concentrations of Manuka honey at 10% or higher increased NET formation, likely due to elevated glucose levels, which stimulate neutrophils.13,14
In vitro studies using an HL60 cell line neutrophil model demonstrate that Manuka honey downregulates the inflammatory NF-κB signaling pathway, chemotaxis, and the release of several key inflammatory cytokines.12,15 Additional in vitro pilot studies of neutrophils within the therapeutic window have indicated reduced MMP-9 release and reduced NETosis in the presence of Manuka honey. Although the pathways through which whole Manuka honey can inhibit the pro-inflammatory characteristics of neutrophils remain to be fully elucidated, the presence of flavonoids with antioxidant properties provides a clue to these inner workings. These flavonoids include pinobanksin, pinocembrin, chrysin, and methyl syringate, which have been found to reduce the activity of pivotal upstream NETosis activators and pro-inflammatory signals, myeloperoxidase (MPO) and ROS.16,17 To elucidate how best to use honeys, such as Manuka honey (or its isolated active components), for biomaterial applications, it is first necessary to determine which components are most responsible for antioxidant and anti-inflammatory activity and at which concentration ranges.
HL60 Cell Line
HL60s are promyelocytic progenitor cells that can be differentiated via coincubation with DMSO to a neutrophil-like phenotype, validated to mimic neutrophil ROS production, NET formation (although significantly diminished compared to primary neutrophils), and inflammatory cytokine release.5,18–22 Unlike primary neutrophils, dHL60s are an immortalized cell line derived from a single leukemia patient and thus do not undergo spontaneous apoptosis during culture, and circumvent patient-to-patient variability. However, these cells are not entirely identical in their behavior to primary human neutrophils and often exhibit diminished effect sizes and responses, particularly in NET formation. Therefore, dHL60 models were used in this study to investigate general trends and to serve as a proof of concept for subsequent primary human neutrophil studies.
Unmet Need in Immunomodulatory Biomaterials and Biomaterial Additives
Rampant neutrophil inflammatory activity and recruitment to a biomaterial implant site shift the host biomaterial continuum towards repair and away from regeneration. Instead of replenishing functional tissue, the body isolates the foreign material from host tissue with a fibrous capsule.23,24 Recent interest has focused on the design of immunomodulatory biomaterials to mitigate foreign body reactions. Yet few studies have focused on modulating the acute phase of inflammation to achieve higher device success rates, and even fewer have targeted neutrophil inflammatory behavior.25–28 An inappropriate neutrophil response to an implant can have deleterious cascading effects, ultimately leading to device failure. However, little investigation has thus far focused on therapeutics and additives that can be incorporated into biomaterial designs to reduce excessive neutrophil inflammation and enable tissue regeneration.
Need for Assay Techniques to Quickly Screen Wide Selections of Biomaterial Additive Candidates to Determine Promise
A primary reason for the paucity of research on biomaterial additives and bioactive compounds that target neutrophil inflammatory behavior is that neutrophils are particularly difficult to work with in vitro. This difficulty is primarily due to neutrophilic behaviors. First, they are terminally differentiated and cannot be cultured or expanded in vitro, necessitating the isolation of primary neutrophils, typically from peripheral blood.29 Even then, the time window for initiating neutrophil isolation after blood draw is very narrow, as phagocytic capacity, bacterial clearance, chemotaxis, and oxidative activity are lost before then.30 Additionally, even when using animal blood draws, the costs of animal models and the time required to obtain IACUC approval can be prohibitive. IRB approval is required for isolating human blood from donors, and substantial donor-to-donor variability in immune activity has been observed.31
Additionally, neutrophil behaviors have been proven to be dependent on a myriad of factors, including blood glucose concentrations (even in non-diabetic donors), smoking history, underlying health conditions, medications, alcohol use, BMI, and other factors.32–36 Neutrophils are also heavily sensitive to shear stress. They can therefore be challenging to use for high-throughput testing, which requires extensive pipetting, as multichannel pipetting is not recommended due to shear stress, resulting in slow processing times.37,38
Thus, the goals of this study were two-fold. The first objective was to develop high-throughput methods to assay a broad range of concentrations of multiple potential immunomodulatory biomaterial additives. These methods would ideally be reproducible, rapid microplate models that mimic neutrophil activation upon biomaterial implantation. This would enable screening of compounds and therapeutic ranges for potential effects on neutrophil behavior, thereby enabling further, more detailed, and focused mechanistic assays on primary human neutrophils. The second objective was to use these high-throughput methods to screen for potential biomaterial additives commonly found in various Honey varieties that have been shown to have anti-inflammatory and pro-wound-healing effects. This study hypothesized that, if pinobanksin, pinocembrin, chrysin, and methyl syringate were co-cultured with pro-inflammatory-stimulated dHL60 cells, intracellular ROS and NETosis would be reduced.
Materials and Methods
The following materials and methods describe the sample, cell, and bioactive preparation; the experimental design and differentiation process; and the analytical approaches used to evaluate the effects of isolated honey bioactive components on dHL60 NET formation and intracellular ROS activity. All experiments were conducted under controlled conditions to ensure reproducibility and minimize variability, with appropriate controls included throughout. Detailed descriptions of reagents, experimental procedures, and data analysis methods are provided to enable replication of the study and facilitate interpretation of the results.
Cell Culture
HL60 cells were purchased from the American Type Culture Collection (ATCC, CCL 240, Manassas, VA, USA) and cultured at a cell density of 1×105 to 1×106 using complete cell culture media. Cell culture media consisted of RPMI-1640 Medium with 2.05 mM L-Glutamine (Hyclone, Logan, UT, USA) with 10% v/v heat-inactivated fetal bovine serum (FBS) (Hyclone), 1% penicillin-streptomycin (Corning, NY, USA). Cells were maintained in T-75 tissue culture flasks (Thermo Scientific, Rochester, NY, USA) in a sterile incubator (Fisher Scientific) at 100% humidity, 5% CO2, and 37°C. Cells were passaged every four days and resuspended in media to a cell concentration of 1×105. Cells were not allowed to exceed 30 passages due to potential genetic drift.
HL60 Differentiation into dHL60s
HL60 cells were differentiated to a neutrophil-like morphology and phenotype by the inclusion of 1.25% dimethyl sulfoxide (DMSO) (Fisher Scientific, Hampton, NH, USA) in the culture medium for six days, with replenishment of DMSO and cell culture medium on the third day. This HL60 differentiation method has been validated in previous studies.12,15,39–41
Confirmation of dHL60 Differentiation
Differentiation was confirmed by morphological examination via fluorescence microscopy. Cells were transferred to 96-well plates (Fisher Scientific), permeabilized with 0.17 mM Triton X-100 (Fisher Scientific) for 5 minutes, and then fixed in 10% buffered formalin (Fisher Scientific). Fixed and permeabilized cells were stained with 4′,6-diamidino-2-phenylindole (DAPI) (NucBlue Fixed Cell Stain ReadyProbes reagent) (Invitrogen, Carlsbad, CA, USA) for 5 minutes at stock concentration. Cells were stained with phalloidin-conjugated Alexa Fluor 488 (ActinGreen 488 ReadyProbes reagent) (Invitrogen) for 30 minutes at stock concentration according to the manufacturer’s protocols to co-localize cells with their nuclei. Stained cells were resuspended in PBS (Gibco), smeared onto glass slides, and the PBS allowed to evaporate at room temperature, protected from light. Slides were then treated with ProLong Gold antifade reagent (Invitrogen) overnight at room temperature to homogenize the focal plane and to prevent photobleaching. Imaging of fluorescent samples was performed with a Nikon A1 confocal scanning laser fluorescence microscope (Nikon, Tokyo, Japan). Differentiated cells (those with neutrophil-characteristic kidney-shaped nuclei) were counted in three randomly selected regions on four separate slides, compared with the number of undifferentiated cells, and expressed as a percentage of total differentiated cells.
Bioactive Selection
Chrysin (Thermo Scientific, cat. no. 110320050), pinocembrin (Sigma Aldrich product no. P5239), pinobanksin (Sigma Aldrich product no. 68530), and methyl syringate (Sigma Aldrich, SKU no. S409448) were obtained in powder form and stock solutions dissolved in DMSO (Fisher). Pinobanksin, pinocembrin, and chrysin concentrations in honey are on the order of 100–500 nM, and previous findings have determined the therapeutic window of honey, specifically, Manuka honey, to be between 0.1–1% v/v, corresponding to 0.1–5 nM of each flavonoid.15,42 To determine if these flavonoids are effective even at ranges outside what is found in honey, serial concentrations between 1 nM and 1 mM of each were assayed. Reports of methyl syringate concentrations in Manuka honey exhibit substantial variability.43,44 Thus, a wide concentration range was selected for this study, from 10 μM to 2 mM.
IntraROS
According to manufacturer instructions, dHL60 cells were dyed with 2′,7′ –dichlorofluorescein diacetate (DCFDA, Abcam, product no. ab113851). Cells were then seeded at 100,000 per well, 150 µL total well volume in HBSS (Gibco) with 0.2% FBS, and 10 mM N-2-hydroxyethylpiperazine-N-2-ethane sulfonic acid (HEPES, Corning, Corning, NY, USA, ref. no. 25–060-CI) with 100 nM Phorbol 12-myristate, 13-acetate (PMA) (Sigma Aldrich, St. Louis, MO, USA) to stimulate intracellular ROS activity and the respective flavonoid and methyl syringate concentrations were added (along with negative controls of unstimulated cells and positive controls of stimulated cells without flavonoids or methyl syringate).45,46 Intracellular ROS activity was quantified via SpectraMax i3 spectrophotometer (Molecular Devices, San Jose, CA, USA) at an Ex/Em of 485/535 at 3 and 6 hours. Sample groups were assayed in quadruplicates, and assays were run with three biological replicates. The results were expressed as relative ROS activity (percent of positive control).
NETosis
dHL60 cells were seeded at 100,000 per well, 150 µL total well volume in HBSS with 0.2% FBS and 10 mM HEPES (Corning), with 4 µM Calcium Ionophore A23187 (Sigma Aldrich) to stimulate NETosis. Flavonoid and methyl syringate concentrations were added (along with negative controls of unstimulated cells and positive controls of stimulated cells without flavonoids or methyl syringate). At 3- and 6-hour timepoints, dHL60s were stained with 0.25 µM Sytox orange (Invitrogen) and incubated in the dark at room temperature to stain extracellular NET-associated DNA. The plates were centrifuged at 1750 rpm for 5 minutes using a Sorvall Legend XTR Centrifuge (Rotor ID: 75003624, Thermo Scientific). After centrifugation, 100 µL supernatant was removed to reduce background fluorescence. Sample groups were assayed in quadruplicate, and assays were run with three biological replicates. These methods were adapted from and validated in previous studies that employed high-throughput quantitative evaluation of NETosis in primary neutrophils.47,48
Cell Viability Assessment
Based on the data collected from NETosis and intracellular ROS assays, cytotoxicity of methyl syringate was assessed at and above the concentration range used for NETosis and intracellular ROS. dHL60 cells were seeded at 100,000 per well, 150 µL total well volume in HBSS with 0.2% FBS and 10 mM HEPES (Corning), with concentrations of methyl syringate ranging from 10 µM to 4 mM. Stimulated controls for both 4µM calcium ionophore and 100 nM PMA, as well as unstimulated controls, were included for comparison. Viability was assayed using a CyQUANT™ MTT Cell Viability Assay Kit (Invitrogen, Cat. No. V13154). Briefly, cells were treated with 10 µL of 12 mM 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT), and incubated at 37°C, 5% CO2, 100% humidity for 2 hours. After 2 hours of incubation, the plates were centrifuged at 1750 rpm for 5 minutes using a Sorvall Legend XTR Centrifuge (Rotor ID: 75003624, Thermo Scientific). After centrifugation, 125 µL supernatant was removed, and insoluble formazan was solubilized by the addition of 50 µL DMSO into each well. Absorbance was measured at 540 nm using a SpectraMax i3 spectrophotometer (Molecular Devices).
Statistics
All graphing and statistical analyses for NETosis and ROS data were performed using R (Version 4.3.0, R Foundation, Indianapolis, IN, USA). All data were pooled, normalized to the mean of the positive control, and expressed as a percentage of the positive control. Due to the non-normality of the data determined by the Shapiro–Wilk test, non-parametric Wilcoxon rank sum tests were utilized to compare sample groups to the positive control data at an alpha value of 0.05. Subsequent statistical analyses and data visualization for cellular viability were performed in GraphPad Prism (Version 8.4.3; GraphPad Software, San Diego, CA, USA). Due to the non-normality of the data, as also determined by the Shapiro–Wilk test, a non-parametric Kruskal–Wallis test was performed with post-hoc Dunn’s multiple comparisons. P-values were adjusted for multiple comparisons and the alpha value was also 0.05.
Results
The following section presents the experimental findings from the analyses described above, focusing on the effects of isolated honey-derived bioactive components on neutrophil NETosis and intracellular ROS activity. Results are reported in a sequence aligned with the study objectives, with quantitative data summarized and statistically evaluated to support interpretation. Figures are referenced as appropriate to illustrate key trends and outcomes.
Confirmation of HL60 Differentiation
Based on the counting of lobed or kidney-shaped nuclei versus round nuclei in a high visual field, the percentage of differentiated cells was 68.4%. These results were within the range reported previously.15,49,50 Morphological characteristics of differentiated and undifferentiated cells were consistent with those reported by Wang et al for nuclear morphology and actin reorganization under the same differentiation protocol (Figures 1 and 2). Although differentiation of 100% of cells was not achieved, corroborating prior investigations indicated that 68.4% was sufficient to proceed with NETosis and ROS activity assays. It was also determined that, due to potential replicate-to-replicate variability in total differentiation levels, all data would be normalized to the positive control, and negative controls of unstimulated cells would be assayed alongside the sample groups to confirm that cells could be stimulated and thus differentiated. Following the reported differentiation protocol, statistical analyses of all experimental datasets yielded p-values <0.0001 for stimulated versus unstimulated cells, with large effect sizes, indicating that cells were reliably induced to exhibit neutrophil-like behaviors.
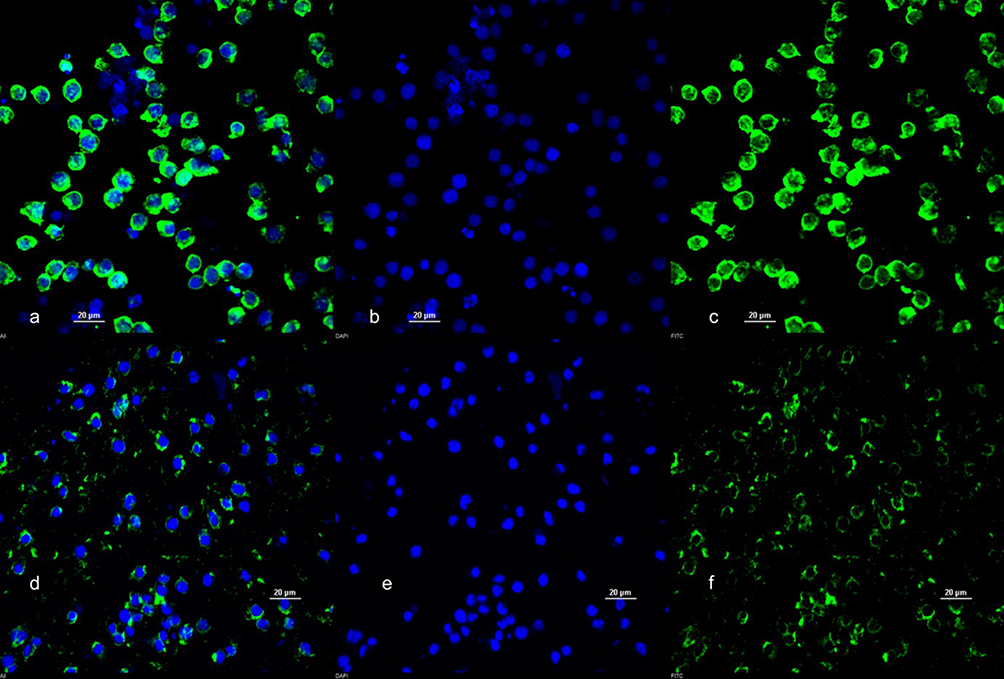 |
Figure 1 DAPI and Actin Staining of undifferentiated HL-60 Cells (a–c) and dHL-60 cells (d–f). Images taken at 60X magnification. Scale bar = 20 µM. |
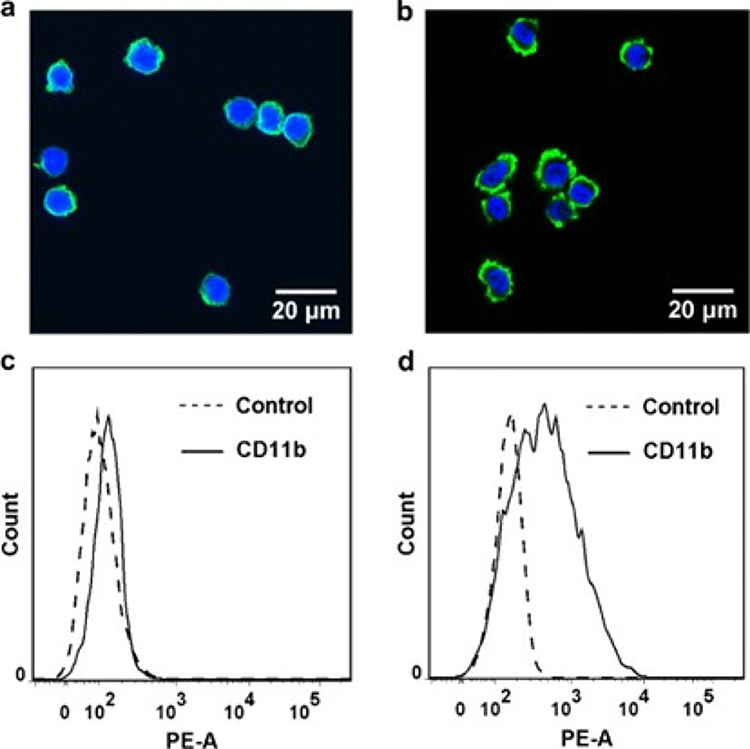 |
Figure 2 DMSO-treated HL-60 differentiation. (a and b) Confocal images of intact (a) and DMSO-differentiated (b) HL-60 cells. (c and d) Mac-1 expression on intact (c) and DMSO-differentiated (d) HL-60 cells. Adapted from Wang, Chengzhi & Li, Ning & Zhang, Chen & Sun, Shujin & Gao, Yuxin & Long, Mian. (2015). Effects of Simulated Microgravity on Functions of Neutrophil-like HL-60 Cells. Microgravity Science and Technology. 27. 10.1007/s12217-015-9473-6.51 |
Chrysin
The data presented by Figure 3 indicate that chrysin fails to reduce dHL60 NET release with a lack of statistically significant changes at the three-hour timepoint (Figure 3a) and even demonstrating mild statistically significant (less than 25% increase in median values) increases in the 5–50 nM range, at 200 nM, and at 400 nM at the six-hour timepoint (Figure 3b). However, an interesting trend was observed when individual experiment replicates were analyzed. Chrysin solutions were prepared in sufficient quantities for two experiments and then remade for the third experiment.
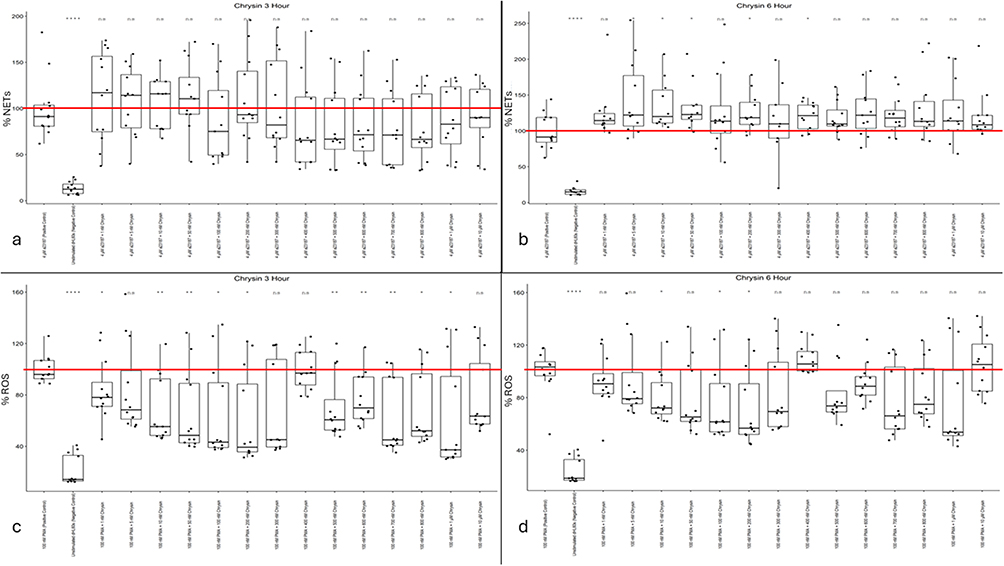 |
Figure 3 (a) Chrysin NET levels at three hours post-Calcium Ionophore A23187 stimulus. (b) Chrysin NET levels at six hours post-Calcium Ionophore A23187 stimulus. (c) Chrysin ROS activity levels at three hours post-PMA stimulus. (d) Chrysin ROS activity levels at six hours post-PMA stimulus. Mean positive control value (100%) is indicated by a red line. Single asterisk indicates p<0.05. Double asterisk indicates p<0.01. Quadruple asterisk indicates p<0.0001. |
In contrast to the NET data, chrysin effectively reduced dHL60 ROS activity. Across a broad therapeutic range between 1 nM and 1 µM, chrysin provided statistically significant (except for the 5 nM, 300 nM, and 400 nM data sets) reductions in intracellular ROS activity by up to 60% (Figure 3c). At the six-hour time point, chrysin shows some promise in its ability to downregulate dHL60 intracellular ROS activity, with mild statistical significance being demonstrated throughout the 10–200 nM range. Median values at 10 nM, 100 nM, and 200 nM indicated decreases in intracellular ROS activity by nearly 40% (Figure 3d).
Pinocembrin
The data presented in Figure 4a and b indicate that pinocembrin may, in fact, be an agonist for dHL60 NET formation, specifically within the 100–300 nM range. Median NET levels increased by 50% in response to incubation with dHL60 and 200 nM pinocembrin (Figure 4a). Levels of NET activation were more variable in the six-hour data than in the three-hour data, with median values increasing relative to the positive control but not reaching statistical significance (Figure 4b). Thus, it can be reasonably concluded that pinocembrin is not responsible for the anti-NET-forming effects demonstrated by honey varieties such as Manuka honey.
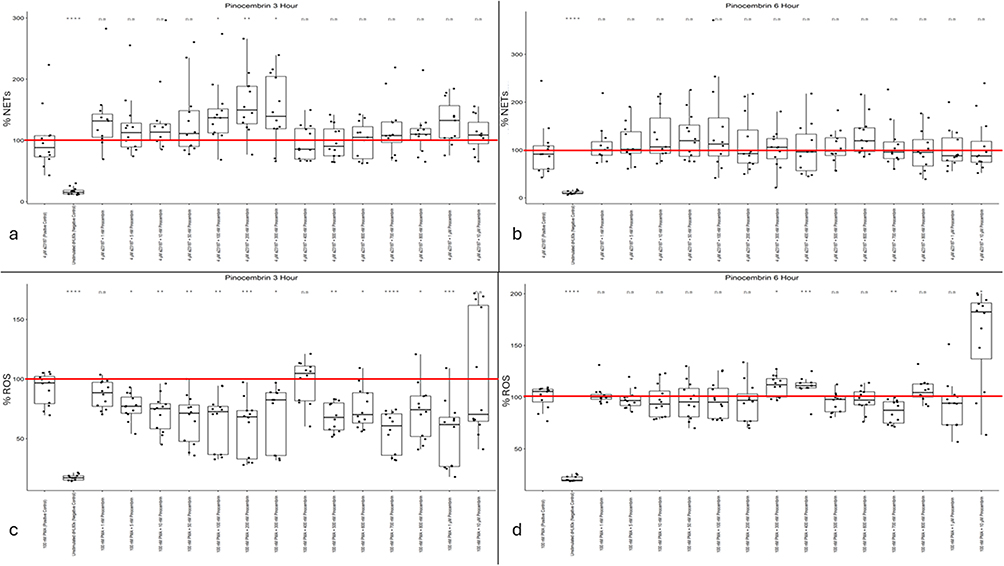 |
Figure 4 (a) Pinocembrin NET levels at three hours post-Calcium Ionophore A23187 stimulus. (b) Pinocembrin NET levels at six hours post-Calcium Ionophore A23187 stimulus. (c) Pinocembrin ROS activity levels at three hours post-PMA stimulus. (d) Pinocembrin ROS activity levels at six hours post-PMA stimulus. Mean positive control value (100%) is indicated by a red line. Single asterisk indicates p<0.05. Double asterisk indicates p<0.01. Triple asterisk indicates p<0.001. Quadruple asterisk indicates p<0.0001. |
As with the chrysin dataset, pinocembrin yields neutral to agonistic effects on NET formation but strong promise in lowering intracellular ROS activity in dHL60. However, in contrast to the chrysin data, this effect is observed only acutely (3 hours after PMA stimulation). The three-hour dataset (Figure 4c) shows a wide therapeutic range from 5 nM to 1 μM, with consistent data, with strong statistical significance, clustered between 10% and 40% reductions in overall intracellular ROS activity (except for 400 nM pinocembrin). These effects subsided by the six-hour time point (Figure 4d). They resulted in a substantial increase in intracellular ROS activity at the highest pinocembrin concentration (10 μM) and a mild yet statistically significant increase at the 400 nM concentration. Overall, these data suggest a pattern similar to that observed for chrysin: pinocembrin acts as an antioxidant but not as a NETosis inhibitor.
Pinobanksin
The data presented by Figure 5a and b indicate that, similar to the other flavonoids selected for this study, this flavonoid has a neutral to agonistic effect on dHL60 NET release. At 3 hours post calcium ionophore treatment, pinobanksin concentrations show no statistically significant differences relative to cells treated with A23187 alone (positive control) (Figure 5a). At six hours post calcium ionophore stimulus, pinobanksin treatment consistently results in higher levels of NETosis across the range of 5 nM to 800 nM (Figure 5b). This dataset likely rules out pinobanksin as a potential anti-NET-forming biomaterial additive.
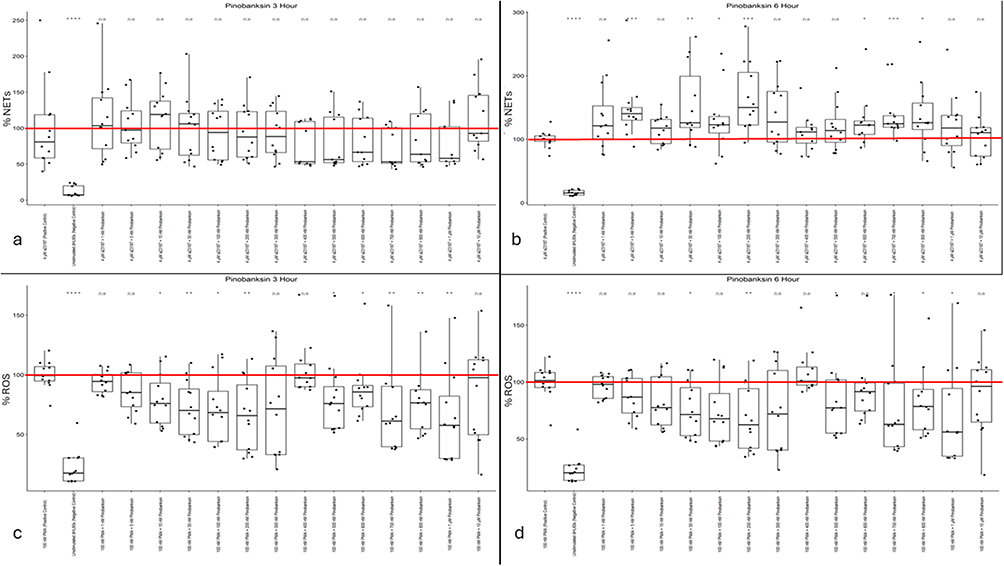 |
Figure 5 (a) Pinobanksin NET levels at three hours post-Calcium Ionophore A23187 stimulus. (b) Pinobanksin NET levels at six hours post-Calcium Ionophore A23187 stimulus. (c) Pinobanksin ROS activity levels at six hours post-PMA stimulus. (d) Pinobanksin ROS activity levels at six hours post-PMA stimulus. Mean positive control value (100%) is indicated by a red line. Single asterisk indicates p<0.05. Single asterisk indicates p<0.05. Double asterisk indicates p<0.01. Triple asterisk indicates p<0.001. Quadruple asterisk indicates p<0.0001. |
While the data for calcium ionophore stimulated dHL60 NET formation indicates that pinobanksin is not promising as a biomaterial additive, the ROS-inhibiting behavior of pinobanksin shown in Figure 5c and d contradict this conclusion. Rather, in concentrations ranging from 10 nM to 1 µM showed consistent, substantial reductions in dHL60 intracellular ROS activity in the three-hour data set (Figure 5c). These ROS activity reductions lasted till the six-hour data set in the 50 nM, 200 nM, 500 nM, and 700–1000 nM concentrations of pinobanksin (Figure 5d). The concentration of pinobanksin that showed the most consistent and significant effect on ROS activity was 1 μM, which remained below 60% of the positive-control level in both the three-hour and six-hour data sets.
Methyl Syringate
The data presented in Figure 6a and b demonstrate a high magnitude of NET inhibition from Calcium Ionophore A23187-stimulated dHL60s treated with methyl syringate lasting until the six-hour timepoint. The data in both datasets showed high within-well consistency and between-biological-replicate consistency. At 3 hours post-stimulus, a therapeutic range of 600 µM to 2 mM methyl syringate reduced dHL60 NET release by more than 50%, except for the 800 µM dataset, which did not reach statistical significance (Figure 6a). This trend persisted through the six-hour time point, with similar reductions in NET formation observed at the same concentrations as in the three-hour data. The data from the six-hour time point indicate that the substantial, tightly controlled reduction in dHL60 NET formation in response to calcium ionophore treatment is well conserved over time (Figure 6b). In contrast to all previously reported trends in this study, methyl syringate potently inhibits NET release from stimulated dHL60s. The findings suggest that among the prevalent compounds in honey selected for this study, methyl syringate is the only one that reliably reduces NETosis, implying that this phenolic component is responsible for previously reported anti-inflammatory and pro-wound-healing effects of various honey types.
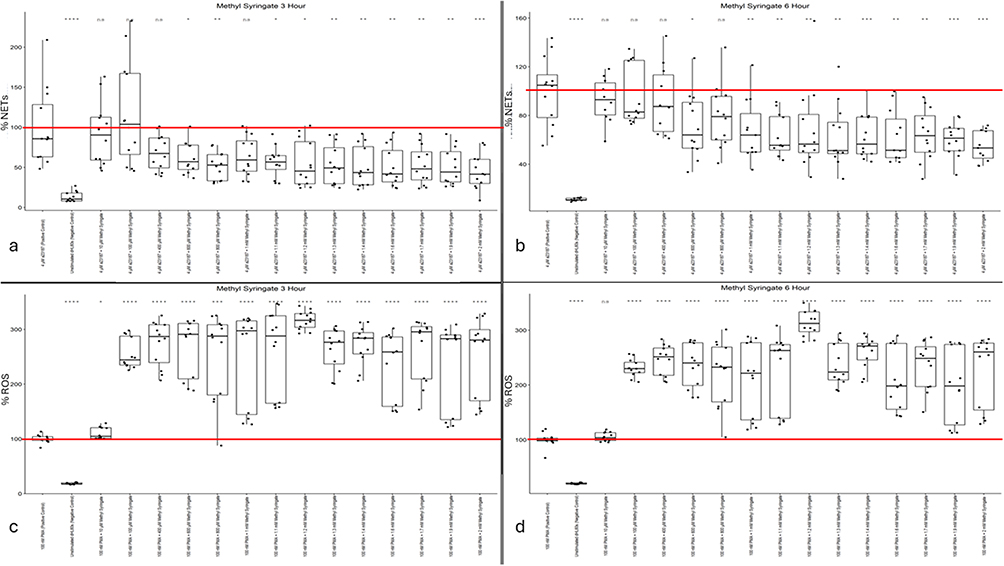 |
Figure 6 (a) Methyl Syringate NET levels at three hours post-Calcium Ionophore A23187 stimulus. (b) Methyl Syringate NET levels at six hours post-Calcium Ionophore A23187 stimulus. (c) Methyl Syringate ROS activity levels at three hours post-PMA stimulus. (d) Methyl Syringate ROS activity levels at three hours post-PMA stimulus. Mean positive control value (100%) is indicated by a red line. Single asterisk indicates p<0.05. Double asterisk indicates p<0.01. Triple asterisk indicates p<0.001. Quadruple asterisk indicates p<0.0001. |
Although the methyl syringate NETosis data showed potent inhibitory effects, the three-hour and six-hour intracellular ROS data showed a pronounced increase in intracellular ROS activity. When stimulated with PMA, methyl syringate exacerbated ROS accumulation in the cytoplasm of dHL60s, resulting in levels up to threefold higher than those observed with PMA alone. In the three-hour data, the lowest dose of methyl syringate did not affect ROS activity, whereas at higher concentrations a strong agonistic effect was observed (Figure 6c). In the six-hour data, however, all doses of methyl syringate, even 10 µM, increased intracellular ROS activity (Figure 6d). These trends were consistent throughout all biological repeats of the assays. ROS activity appears to peak at the 1.2 mM concentration of methyl syringate, resulting in an above 310% increase in cytoplasmic ROS activity.
Cellular Viability
The data from the MTT assay demonstrates that even at twice the upper range of methyl syringate concentrations assayed, there is no observed cytotoxicity after 6 hour incubation with methyl syringate when compared to A23187 or PMA alone. The only statistical significance was found between 100 µM and 4 mM methyl syringate groups (p=0.2) (Figure 7). Statistical analysis found most p-values to be greater than 0.9 between groups, indicating a high confidence level that there is no distinguishable difference between groups.
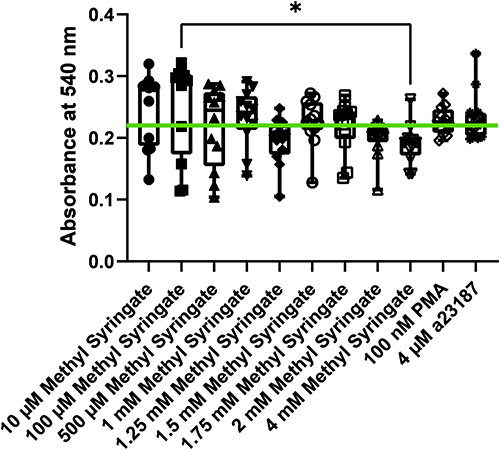 |
Figure 7 Cellular viability as measured by MTT assay at six hours. Mean absorbance of control cells is indicated as a green line. Single asterisk indicates p<0.05. |
Discussion
Despite ROS activity being an essential precursor to NET release, methyl syringate increased ROS without a corresponding increase in NETosis.5 In fact, the opposite effect was observed, where an increase in intracellular ROS corresponded to an inverse effect on NET release. This phenomenon suggests that inhibition of NETosis by methyl syringate occurs either downstream of, or independently of, intracellular ROS accumulation. This theory is consistent with the potent inhibitory effect of methyl syringate on MPO. MPO is an enzyme that drives neutrophil inflammatory behavior, specifically NETosis.52 What is surprising, however, is that this MPO inhibition did not also reduce intracellular ROS levels. Perhaps inhibition or deactivation of downstream effects leads to the accumulation of intracellular ROS. Further mechanistic and signaling studies are required to elucidate the interplay between ROS and NETosis upon treatment with methyl syringate and to explain it with certainty and in detail.
This work has been further validated by subsequent primary neutrophil studies from this lab, which, while showing an inverse effect on ROS activity, corroborate the high effect size in NET inhibition by methyl syringate. Interestingly, investigations based on the results of this study found no increase in intracellular ROS activity in response to methyl syringate in primary human neutrophils, but rather a significant inhibition. Additionally, methyl syringate was more effective than whole Manuka honey at reducing ROS but was somewhat less effective at reducing NETosis. Although no spike in ROS was observed at methyl syringate concentrations in primary neutrophils, a trend similar to the methyl syringate ROS data presented in this study was observed when a combination of chrysin, pinocembrin, pinobanksin, and methyl syringate was co-cultured with neutrophils. These findings prompted the use of the CyQUANT MTT Cell Viability Assay (Invitrogen) on the combination of flavonoid and methyl syringate concentrations in differentiated HL60s. The findings suggested a mild reduction in viability (~20%) at the highest combined concentration of all flavonoids and methyl syringate. These reductions in viability, however, do not scale proportionally with the reductions in NETosis observed in both the present study and subsequent primary neutrophil studies.53 Additionally, the methods used for detecting intracellular ROS are viability-dependent, and a loss of cell viability would reduce total fluorescence. NET-associated extracellular DNA is also defined by pronounced chromatin decondensation and its diffusion beyond the cellular boundary. It is detected by Sytox Orange staining in the absence of fixation and permeabilization. In contrast, necrosis is identified by the presence of intact yet condensed nuclei. In contrast, apoptosis is characterized by preserved cellular membrane integrity, which precludes Sytox dye entry and limits nucleic acid staining. These staining patterns provide a reliable basis for distinguishing an observed reduction in NET formation from necrotic and apoptotic cell death.48 This previous data was corroborated in the present study, with no cytotoxicity observed even at twice the levels assayed for NETosis and ROS. Therefore, although methyl syringate increased intracellular ROS levels, cellular viability was maintained even at the six-hour time point. The present study validates these findings, employing a high-throughput, high-reproducibility screening method to mitigate donor-to-donor variability, sourcing difficulties, and the labor-intensive methods required for primary neutrophil work. The validated methods in this study enable researchers to reliably and effectively screen for neutrophil-targeted therapeutics, facilitating further testing of acute immune responses to bioactive compounds. Thus, by removing the obstacles that neutrophil investigations entail, more research will be encouraged to target acute-phase inflammation without the risk of heavy time and resource investment for no return.
The increase in intracellular ROS activity is consistent with previous studies showing increased ROS generation and the presence of H2O2 in the extracellular medium in response to whole Manuka honey.54,55 Notably, even as intracellular ROS activity increased, antioxidant activity and cytoprotection against oxidative stress were maintained. Previous publications report promising data indicating that Manuka honey protects mitochondrial function, activates the AMPK/Nrf2/ARE signaling pathway, and induces the expression of antioxidant enzymes, such as superoxide dismutase (SOD) and catalase (CAT), in fibroblasts.16 Perhaps the protection that specific honey types afford from oxidative stress does not occur intracellularly or does not occur in a way that corresponds to a decrease in intracellular ROS activity. It is possible that these two events can coincide: ROS activity is indeed increased within the cell, but honey also induces an extracellular or cytoprotective factor. Intracellular ROS activity is only one of myriad mechanisms for assessing oxidative protection, and honey may protect cells in other ways. Further study of this matter should include assays of extracellular ROS scavenging and lipid peroxidation. Now that a substantial effect has been observed, specifically that in differentiated and stimulated HL60s, methyl syringate induces intracellular ROS build-up, but lowers NET release, further mechanistic studies involving primary neutrophils can be performed. Neutrophil signaling is complex, context-dependent, and not yet fully understood; consequently, broad-ranging, comprehensive investigative techniques are also limited. Therefore, having a starting point, such as a potential step or series of steps, within a signaling cascade is critical. The findings of this investigation, notably the discrepancy between increased ROS activity and decreased NET release, provide an opportunity to investigate the signaling events linking superoxide generation by NADPH oxidase to NET formation and subsequent release.
While this study provides proof of concept for further research into honey as a biomaterial additive and a therapeutic agent to reduce neutrophil-mediated inflammation, it has significant limitations. The first and perhaps most prominent limitation is the fact that HL60 cells are not primary human neutrophils. While HL60 Cells are a valuable model for mimicking the physiological responses of primary human neutrophils, there is no one-for-one correspondence. HL60s were chosen for this exploratory pilot study to determine if the selected bioactive molecules held further promise in terms of modulating neutrophil inflammatory responses, and, if so, at what concentration ranges, as the range that these compounds exist in honey is widely variable, even in honeys currently in use as wound healing therapeutics. The rationale for using an HL60 model is the inability to culture and expand neutrophils in vitro; the need to obtain human blood from a moderate to large number of donors; the associated institutional review board approvals; and the high donor-to-donor variability of neutrophils, which necessitates a large donor pool. Thus, it is reasonable to use promising preliminary data as justification before proceeding with primary human neutrophil studies. A second limitation of the current study is that it demonstrates that methyl syringate reduces NETosis but does not explain how or why this occurs. Now that a substantial effect has been observed with methyl syringate, and other compounds that did not show as strong a benefit have been eliminated, more in-depth, narrowly focused investigations may be conducted. The mechanisms by which methyl syringate reduces NET formation must now be elucidated. Future mechanistic studies should be conducted involving primary human neutrophils to investigate likely pathways that could be interrupted by methyl syringate, including NADPH oxidase (NOX) inhibition, Peptidyl Arginine Deiminase 4 (PAD4) inhibition, which are essential components in the ROS-NETosis pathway, as well as NF-κB phosphorylation, which is an essential upstream signaling event for neutrophil inflammatory activation.56,57 Additionally, an interesting trend was observed when individual experiment replicates were analyzed. For example, chrysin solutions were prepared in sufficient quantities for two experiments and then remade for the third experiment. While experiment two did not meet our exclusion criteria to discard a biological replicate (p-value of greater than 0.001 between positive and negative controls, indicating either a failure to induce an inflammatory response or some form of contamination causing negative control wells to become stimulated), the data does suggest that a loss of efficacy may be the result of storage in aqueous solutions for longer than two days (Supplementary Figure 1). These trends were notable, as no changes were made with regard to cell culture conditions, treatment times, and cell and passage numbers, indicating that the reduced effect likely resulted from chrysin itself. However, these trends were outside the current scope of the study and emphasize the need for in-depth investigations into the chemical stability of these compounds outside the honey milieu.
Another essential obstacle in translating honey-derived therapeutics to clinical applications is the presence of confounding physicochemical properties inherent to whole honey, emphasizing the necessity of first evaluating isolated bioactive components. Honey exhibits high osmolarity and low pH, both of which are well-established contributors to the antibacterial activity of many honey types. These factors can independently modulate neutrophil behavior, specifically, thereby complicating attribution of observed anti-inflammatory effects to specific molecular constituents. Consequently, assessment of isolated compounds, such as methyl syringate, is critical for delineating direct immunomodulatory activity from secondary effects driven by nonspecific environmental changes. The pH of the local microenvironment is a central regulator of neutrophil activation and inflammatory outcomes. Most honey exhibits a moderately acidic pH (3.5–4.5), a condition that enhances neutrophil antimicrobial function and promotes cytokine and chemokine production, potentially amplifying both the magnitude and duration of inflammation. However, despite inducing localized acidification, previous findings demonstrate that whole Manuka honey treatment resulted in limited cytokine and chemokine release. This dissociation suggests that phenolic and antioxidant constituents exert dominant anti-inflammatory effects that may counterbalance pH-driven neutrophil activation. Nonetheless, without isolating individual components, the relative contributions of molecular bioactivity versus physicochemical conditions cannot be definitively resolved. In addition to pH, neutrophil function is susceptible to osmotic stress within the inflammatory microenvironment. Elevated osmolarity can initiate intracellular signaling pathways, including altered calcium flux and downstream activation cascades, that promote the release of pro-inflammatory mediators and influence chemotactic responsiveness. Because whole honey simultaneously alters osmotic conditions and delivers multiple bioactive molecules, isolating individual components is essential to disentangle these overlapping stimuli. These findings support a stepwise experimental strategy in which isolated honey constituents are first evaluated under controlled conditions to establish causal mechanisms, followed by reintegration into whole-honey or biomaterial-based systems. Such an approach is critical for the rational design, optimization, and translational application of honey-derived immunomodulatory therapies. By determining which components contribute to the dual-purpose (antibacterial and anti-inflammatory properties) that many honey strains have demonstrated, the first step in this experimental strategy can be resolved.57–61
The findings of this study elucidate the components of honey that inhibit neutrophil-mediated inflammation. Methyl syringate emerges as a strong candidate for reducing NETosis observed in previous Manuka honey studies in both dHL60 models and primary human neutrophil studies.13,15,28 Methyl syringate could thus be used as an anti-NET drug to mitigate the adverse downstream effects of NETosis or as an additive to Manuka honey to enhance its anti-inflammatory properties. These capacities can be further extended into the field of biomaterials, allowing implants to pass through the acute phase of inflammation without extensive NET build-up and the resultant pro-inflammatory signaling and enzymatic degradation. Honey, specifically Manuka honey, has already shown potential as a biomaterial additive; adding methyl syringate could further enhance its efficacy in reducing NETosis, thereby enabling the development of biomaterial implants that effectively target acute-phase inflammation and improve device outcomes.
Conclusion
In summary, methyl syringate was the most effective honey component tested in this study. The increase in intracellular ROS activity in response to methyl syringate concentrations was surprising; however, the scientific literature corroborates that this phenomenon can coexist with cytoprotection and anti-inflammatory wound resolution. The present study demonstrates that isolated Manuka honey-derived components modulate dHL60 inflammatory responses to varying degrees of efficacy, with methyl syringate emerging as a particularly potent regulator of NET formation. These findings advance the understanding of how specific phenolic constituents contribute to the immunomodulatory properties traditionally attributed to whole honey. By establishing a foundation for component-level evaluation, this work supports a stepwise approach to developing honey-derived anti-inflammatory strategies. Thus, future studies incorporating primary human neutrophils, especially in mechanistic and signaling studies, complex co-culture systems, and biomaterial interfaces, will be essential to translate these insights into clinically relevant applications. Additionally, these validated, high-throughput assays enable screening of neutrophil-targeting therapeutics and biomaterial additives, thereby facilitating more accessible, reproducible, and efficient research in this vital area of reducing dysregulated innate immune responses to biomaterials, implants, and infections.
Data Sharing Statement
Any and all raw data supporting the conclusions of this article will be made available by the authors upon request.
Acknowledgment
The authors thank Dr. Benjamin Minden-Birkenmaier for his training, advice, and counsel regarding HL-60 work and laboratory protocols. The authors thank Lauren Thompson and Dr. Omar Skalli for their assistance in confocal microscopy and fluorescence imaging preparation. The authors also thank Kian Ziai for his hard work as an undergraduate research trainee.
Author Contributions
Evan N. Main: Data curation, formal analysis, investigation, validation, visualization, preparation, conceptualization, methodology, writing – original draft, writing – review and editing. James C. Huang: Investigation, validation, data curation, writing – review and editing. Gary L. Bowlin: Funding acquisition, resources, project administration, supervision, conceptualization, methodology, writing, review, and editing. All authors gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This research was funded by the National Institutes of Health, Department of Health and Human Services, National Institute of Biomedical Imaging and Bioengineering, award number 1R15EB033752-01A1.
Disclosure
The authors declare no conflict of interest.
References
1. Loh JT, Lam K-P. Neutrophils in the pathogenesis of rheumatic diseases. Rheumatol Immunol Res. 2022;3:120–17. doi:10.2478/rir-2022-0020
2. Cassatella MA. The Neutrophil: An Emerging Regulator of inflammatory and Immune Response. Karger Medical and Scientific Publishers; 2003.
3. Keshari RS, Jyoti A, Dubey M, et al. Cytokines induced neutrophil extracellular traps formation: implication for the inflammatory disease condition. PLoS One. 2012;7:e48111.
4. Ling S, Xu J-W. NETosis as a pathogenic factor for heart failure. Oxid Med Cell Longev. 2021;2021. doi:10.1155/2021/6687096
5. Remijsen Q, Kuijpers T, Wirawan E, Lippens S, Vandenabeele P, Vanden Berghe T. Dying for a cause: NETosis, mechanisms behind an antimicrobial cell death modality. Cell Death Differ. 2011;18:581–588. doi:10.1038/cdd.2011.1
6. Main EN, Bowlin GL. Potential for Manuka honey‐inspired therapeutics to improve the host–biomaterial response. Medcomm–Biomater Appl. 2022;1:e18.
7. Selders GS, Fetz AE, Radic MZ, Bowlin GL. An overview of the role of neutrophils in innate immunity, inflammation, and host-biomaterial integration. Regen Biomater. 2017;4:55–68. doi:10.1093/rb/rbw041
8. Fetz AE, Bowlin GL. Neutrophil extracellular traps: inflammation and biomaterial preconditioning for tissue engineering. Tissue Eng B. 2022;28:437–450. doi:10.1089/ten.teb.2021.0013
9. Ratner B. Vascular grafts: technology success/technology failure. BME Frontiers. 2023;4:0003. doi:10.34133/bmef.0003
10. Oryan A, Alemzadeh E, Moshiri A. Biological properties and therapeutic activities of honey in wound healing: a narrative review and meta-analysis. J Tissue Viability. 2016;25(2):98–118. doi:10.1016/j.jtv.2015.12.002
11. Minden-Birkenmaier BA, Bowlin GL. Honey-based templates in wound healing and tissue engineering. Bioengineering. 2018;5:46. doi:10.3390/bioengineering5020046
12. Minden-Birkenmaier BA, Meadows MB, Cherukuri K, et al. The effect of manuka honey on dHL-60 cytokine, chemokine, and matrix-degrading enzyme release under inflammatory conditions. Med One. 2019;4:e190005.
13. Minden-Birkenmaier BA, Smith RA, Radic MZ, van der Merwe M, Bowlin GL. Manuka honey reduces NETosis on an electrospun template within a therapeutic window. Polymers. 2020;12:1430. doi:10.3390/polym12061430
14. Menegazzo L, Ciciliot S, Poncina N, et al. NETosis is induced by high glucose and associated with type 2 diabetes. Acta Diabetologica. 2015;52:497–503. doi:10.1007/s00592-014-0676-x
15. Minden-Birkenmaier BA, Cherukuri K, Smith RA, Radic MZ, Bowlin GL. Manuka honey modulates the inflammatory behavior of a dHL-60 neutrophil model under the cytotoxic limit. Int J Biomater. 2019;2019:1–12. doi:10.1155/2019/6132581
16. Alvarez-Suarez JM, Giampieri F, Cordero M, et al. Activation of AMPK/Nrf2 signalling by Manuka honey protects human dermal fibroblasts against oxidative damage by improving antioxidant response and mitochondrial function promoting wound healing. J Funct Foods. 2016;25:38–49. doi:10.1016/j.jff.2016.05.008
17. Alvarez-Suarez J, Gasparrini M, Forbes-Hernández T, Mazzoni L, Giampieri F. The composition and biological activity of honey: a focus on Manuka honey. Foods. 2014;3:420–432. doi:10.3390/foods3030420
18. Moghanloo E, Ghorbani E, Beikverdi MS, et al. The netosis formation of HL-60 cell differentiated to neutrophil-like cells by LPS. J Hum Environ Health Promot. 2018;4:138. doi:10.29252/jhehp.4.3.8
19. Manda‐Handzlik A, Bystrzycka W, Wachowska M, et al. The influence of agents differentiating HL‐60 cells toward granulocyte‐like cells on their ability to release neutrophil extracellular traps. Immunol Cell Biol. 2018;96:413–425. doi:10.1111/imcb.12015
20. Verdon R, Gillies SL, Brown DM, et al. Neutrophil activation by nanomaterials in vitro: comparing strengths and limitations of primary human cells with those of an immortalized (HL-60) cell line. Nanotoxicology. 2021;15:1–20. doi:10.1080/17435390.2020.1834635
21. Sontag W. Modulation of cytokine production by interferential current in differentiated HL‐60 cells. Bioelectromagnetics. 2000;21:238–244. doi:10.1002/(SICI)1521-186X(200004)21:3<238::AID-BEM10>3.0.CO;2-Y
22. Lin K-T, Xue J-Y, Sun FF, Wong PY. Reactive oxygen species participate in peroxynitrite-induced apoptosis in HL-60 cells. Biochem Biophys Res Commun. 1997;230:115–119. doi:10.1006/bbrc.1996.5897
23. Anderson JM, Rodriguez A, Chang DT. Foreign body reaction to biomaterials. Semin Immunol. 2008;20:86–100. doi:10.1016/j.smim.2007.11.004
24. Anderson JM. Biological responses to materials. Annu Rev Mater Res. 2001;31:81–110. doi:10.1146/annurev.matsci.31.1.81
25. Fetz AE, Neeli I, Buddington KK, et al. Localized delivery of Cl-amidine from electrospun polydioxanone templates to regulate acute neutrophil NETosis: a preliminary evaluation of the PAD4 inhibitor for tissue engineering. Front Pharmacol. 2018;9:289. doi:10.3389/fphar.2018.00289
26. Fetz AE, Neeli I, Rodriguez IA, Radic MZ, Bowlin GL. Electrospun template architecture and composition regulate neutrophil NETosis in vitro and in vivo. Tissue Eng Part A. 2017;23:1054–1063. doi:10.1089/ten.tea.2016.0452
27. Fetz AE. Electrospun Biomaterial-Induced Neutrophil Extracellular Traps: Characterization and Regulation for Biomaterial-Guided Tissue Regeneration. The University of Memphis; 2021.
28. Minden-Birkenmaier BA. Manuka Honey as a Tissue Engineering Bioactive: Effect on Neutrophil Inflammatory Behavior. The University of Memphis; 2020.
29. Blanter M, Gouwy M, Struyf S. Studying neutrophil function in vitro: cell models and environmental factors. J Inflamm Res. 2021;Volume 14:141–162. doi:10.2147/JIR.S284941
30. Connelly AN, Huijbregts RP, Pal HC, et al. Optimization of methods for the accurate characterization of whole blood neutrophils. Sci Rep. 2022;12:3667. doi:10.1038/s41598-022-07455-2
31. Duarte M, Kuchibhatla M, Khandelwal S, Arepally GM, Lee GM. Heterogeneity in neutrophil responses to immune complexes. Blood Adv. 2019;3:2778–2789. doi:10.1182/bloodadvances.2019000235
32. Brotfain E, Hadad N, Shapira Y, et al. Neutrophil functions in morbidly obese subjects. Clin Exp Immunol. 2015;181:156–163. doi:10.1111/cei.12631
33. Saito Y, Takahashi I, Iwane K, et al. The influence of blood glucose on neutrophil function in individuals without diabetes. Luminescence. 2013;28:569–573. doi:10.1002/bio.2495
34. Sørensen LT, Nielsen HB, Kharazmi A, Gottrup F. Effect of smoking and abstention on oxidative burst and reactivity of neutrophils and monocytes. Surgery. 2004;136:1047–1053. doi:10.1016/j.surg.2004.04.001
35. Sato J, Takahashi I, Umeda T, et al. Effect of alcohol drinking and cigarette smoking on neutrophil functions in adults. Luminescence. 2011;26:557–564. doi:10.1002/bio.1270
36. Beyrau M, Bodkin JV, Nourshargh S. Neutrophil heterogeneity in health and disease: a revitalized avenue in inflammation and immunity. Open Biology. 2012;2:120134. doi:10.1098/rsob.120134
37. Drury JL, Dembo M. Aspiration of human neutrophils: effects of shear thinning and cortical dissipation. Biophys J. 2001;81:3166–3177. doi:10.1016/S0006-3495(01)75953-X
38. Zhelev D, Hochmuth R. Human neutrophils under mechanical stress. In: Cell Mechanics and Cellular Engineering. Springer; 1994:3–21.
39. Jenkins R, Burton N, Cooper R. Effect of manuka honey on the expression of universal stress protein A in meticillin-resistant Staphylococcus aureus. Int J Antimicrob Agents. 2011;37:373–376. doi:10.1016/j.ijantimicag.2010.11.036
40. Wallace A, Eady S, Miles M, et al. Demonstrating the safety of manuka honey UMF® 20+ in a human clinical trial with healthy individuals. Br J Nutr. 2010;103:1023–1028. doi:10.1017/S0007114509992777
41. Minden-Birkenmaier BA, Meadows MB, Cherukuri K, et al. Manuka honey modulates the release profile of a dHL-60 neutrophil model under anti-inflammatory stimulation. J Tissue Viability. 2020;29:91–99. doi:10.1016/j.jtv.2020.03.005
42. Chan CW, Deadman BJ, Manley-Harris M, Wilkins AL, Alber DG, Harry E. Analysis of the flavonoid component of bioactive New Zealand mānuka (Leptospermum scoparium) honey and the isolation, characterisation and synthesis of an unusual pyrrole. Food Chem. 2013;141:1772–1781. doi:10.1016/j.foodchem.2013.04.092
43. El-Senduny FF, Hegazi NM, Abd Elghani GE, Farag MA. Manuka honey is a unique monofloral honey. A comprehensive review of its bioactives, metabolism, action mechanisms, and therapeutic merits. Food Bioscience. 2021;42:101038. doi:10.1016/j.fbio.2021.101038
44. Oelschlaegel S, Gruner M, Wang P-N, Boettcher A, Koelling-Speer I, Speer K. Classification and characterization of manuka honeys based on phenolic compounds and methylglyoxal. J Agric Food Chem. 2012;60:7229–7237. doi:10.1021/jf300888q
45. Grommes J, Vijayan S, Drechsler M, et al. Simvastatin reduces endotoxin-induced acute lung injury by decreasing neutrophil recruitment and radical formation. PLoS One. 2012;7:e38917.
46. Gottlieb E, Vander Heiden MG, Thompson CB. Bcl-xL prevents the initial decrease in mitochondrial membrane potential and subsequent reactive oxygen species production during tumor necrosis factor alpha-induced apoptosis. Mol Cell Biol. 2000;20:5680–5689. doi:10.1128/MCB.20.15.5680-5689.2000
47. Jiang D, Saffarzadeh M, Scharffetter-Kochanek K. In vitro demonstration and quantification of neutrophil extracellular trap formation. Bio-Protocol. 2017;7:e2386–e.
48. Gupta S, Chan DW, Zaal KJ, Kaplan MJ. A high-throughput real-time imaging technique to quantify NETosis and distinguish mechanisms of cell death in human neutrophils. J Immunol. 2018;200:869–879. doi:10.4049/jimmunol.1700905
49. Gallagher R, Collins S, Trujillo J, et al. Characterization of the continuous, differentiating myeloid cell line (HL-60) from a patient with acute promyelocytic leukemia. 1979.
50. Martin S, Bradley J, Cotter T. HL‐60 cells induced to differentiate towards neutrophils subsequently die via apoptosis. Clin Exp Immunol. 1990;79:448–453. doi:10.1111/j.1365-2249.1990.tb08110.x
51. Wang C, Li N, Zhang C, Sun S, Gao Y, Long M. Effects of simulated microgravity on functions of neutrophil-like HL-60 cells. Microgravity Sci Technol. 2015;27:515–527. doi:10.1007/s12217-015-9473-6
52. Kato Y. Neutrophil myeloperoxidase and its substrates: formation of specific markers and reactive compounds during inflammation. J Clin Biochem Nutr. 2016;58:99–104. doi:10.3164/jcbn.15-104
53. Main EN, Huang JC, Bowlin GL. Methyl syringate: a primary driving factor in Manuka honeys ability to ameliorate neutrophil intracellular ROS activity and NETosis. Front Biosci. 2024;29:255. doi:10.31083/j.fbl2907255
54. Afrin S, Forbes-Hernandez TY, Gasparrini M, et al. Strawberry-tree honey induces growth inhibition of human colon cancer cells and increases ROS generation: a comparison with Manuka honey. Int J Mol Sci. 2017;18:613. doi:10.3390/ijms18030613
55. Martinotti S, Pellavio G, Patrone M, Laforenza U, Ranzato E. Manuka honey induces apoptosis of epithelial cancer cells through aquaporin-3 and calcium signaling. Life. 2020;10:256. doi:10.3390/life10110256
56. McDonald PP, Bald A, Cassatella MA. Activation of the NF-κB pathway by inflammatory stimuli in human neutrophils. Blood J Am Soc Hematol. 1997;89:3421–3433.
57. Almasaudi S. The antibacterial activities of honey. Saudi J Biol Sci. 2021;28:2188–2196. doi:10.1016/j.sjbs.2020.10.017
58. Johnston M, McBride M, Dahiya D, Owusu-Apenten R, Nigam PS. Antibacterial activity of Manuka honey and its components: an overview. AIMS Microbiol. 2018;4:655. doi:10.3934/microbiol.2018.4.655
59. Kwakman PH, Te Velde AA, de Boer L, Vandenbroucke-Grauls CM, Zaat SA. Two major medicinal honeys have different mechanisms of bactericidal activity. PLoS One. 2011;6:e17709.
60. Mousa AM, Allemailem KS, Alhumaydhi FA, et al. Cytoprotective antioxidant, anti-inflammatory, and antifibrotic impact of celery seed oil and manuka honey against cyclophosphamide-induced cystitis in rabbits. Evid Based Complement Alternat Med. 2022;2022:1–11. doi:10.1155/2022/2863023
61. Navaei‐Alipour N, Mastali M, Ferns GA, Saberi‐Karimian M, Ghayour‐Mobarhan M. The effects of honey on pro‐and anti‐inflammatory cytokines: a narrative review. Phytother Res. 2021;35:3690–3701. doi:10.1002/ptr.7066
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.