Back to Journals » Drug Design, Development and Therapy » Volume 20
Ursodeoxycholic Acid Alleviates DSS/AOM-Induced Colorectal Cancer in Mice by Inhibiting PI3K/Akt/mTOR Signaling Pathway
Authors Huang Z, Hong H, Yang H, He C
Received 16 August 2025
Accepted for publication 11 January 2026
Published 23 January 2026 Volume 2026:20 556850
DOI https://doi.org/10.2147/DDDT.S556850
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 4
Editor who approved publication: Dr Solomon Tadesse Zeleke
Zuoxi Huang,1 Huazhang Hong,1 Hongbao Yang,1 Chong He2
1Department of Gastrointestinal Surgery, Jiangxi Provincial People’s Hospital, The First Affiliated Hospital of Nanchang Medical College, Nanchang, Jiangxi, People’s Republic of China; 2Jiangxi Trauma Clinical Medical Research Center, Department of Emergency Surgery, The First Affiliated Hospital, Jiangxi Medical College, Nanchang University, Nanchang, Jiangxi, People’s Republic of China
Correspondence: Chong He, Email [email protected]
Introduction: Ursodeoxycholic acid (UDCA) demonstrates potential therapeutic effects against colorectal cancer (CRC) due to its anti-inflammatory and immunomodulatory properties; however, its precise molecular mechanisms remain incompletely understood.
Methods: This study employed an integrative approach combining network pharmacology, molecular docking, and in vivo validation in an AOM/DSS-induced mouse model to investigate the specific molecular targets of UDCA and its associated effects on the gut microenvironment.
Results: UDCA significantly alleviated colitis-associated tumorigenesis and reduced tumor burden, which was associated with the inhibition of the PI3K/Akt/mTOR signaling pathway. Molecular docking and experimental verification identified EGFR as a key upstream target directly engaged by UDCA to suppress this oncogenic axis. Furthermore, UDCA treatment improved the tumor microenvironment, characterized by suppressed pro-inflammatory cytokines, regulated metabolic gene expression (including CYP19A1 and HMGCR), and a shift toward gut microbiota homeostasis through the enrichment of beneficial taxa and short-chain fatty acids.
Conclusion: These findings suggest that UDCA exerts its anti-tumor effects primarily through direct inhibition of the EGFR-mediated PI3K/Akt/mTOR pathway, accompanied by partial restoration of the intestinal immune-metabolic microenvironment. This study provides new mechanistic insights supporting the therapeutic application of UDCA in CRC.
Keywords: ursodeoxycholic acid, colorectal cancer, network pharmacology, molecular docking, intestinal flora, PI3K/Akt/mTOR signaling pathway
Introduction
Colorectal cancer is one of the most prevalent malignancies worldwide, characterized by high morbidity and mortality.1 The incidence and progression of Colorectal cancer are influenced by multiple factors, including genetic predisposition, lifestyle, dietary habits, and the intestinal microenvironment.2,3 In recent years, extensive research into the pathogenesis of Colorectal cancer has provided increasing evidence that dysbiosis of the intestinal microbiota and chronic inflammation are significant risk factors for the disease.4 Consequently, the identification of therapeutic agents capable of modulating the intestinal microenvironment, suppressing inflammatory responses, and exhibiting anticancer activity has emerged as a crucial focus in Colorectal cancer research.
Ursodeoxycholic acid (UDCA), a naturally occurring bile acid, is widely recognized for its ability to regulate bile acid metabolism and exhibit anti-inflammatory, antioxidant, and immunomodulatory properties. Previous studies have demonstrated that UDCA exerts biochemical effects primarily by regulating bile acid synthesis and improving bile acid composition.5,6 Furthermore, UDCA has been shown to improve the intestinal microenvironment by enhancing the composition of beneficial bacteria while reducing harmful bacterial populations.7 For instance, Fathima et al found that UDCA modulated pro-inflammatory cytokine levels in activated macrophages, mitigating chronic enteritis. Similarly, Biagioli et al reported that specific bile acids, including UDCA derivatives, could reverse disease progression and intestinal microbiota dysregulation in colitis models.8–10
However, while the general anti-inflammatory and microbiota-regulating effects of UDCA are well-documented, the precise molecular mechanisms governing its direct anti-tumor activity remain unclear. Specifically, it is not yet fully understood whether UDCA targets specific oncogenic receptors to disrupt intracellular proliferation signals. The PI3K/Akt/mTOR signaling pathway is a central regulator of cell proliferation and survival, and its hyperactivation—often driven by upstream receptors such as the Epidermal Growth Factor Receptor (EGFR)is a critical driver of CRC tumorigenesis. Despite the broad clinical interest in UDCA, whether it can mitigate CRC by directly targeting the EGFR-mediated PI3K/Akt/mTOR axis has not been systematically investigated. This represents a significant gap in understanding the full therapeutic potential of UDCA beyond its immunomodulatory roles.
Therefore, this study aims to identify novel therapeutic targets of UDCA in CRC using an integrative approach. By combining network pharmacology and molecular docking with in vivo experimental validation, we sought to determine whether UDCA inhibits the PI3K/Akt/mTOR pathway via direct binding to EGFR, and how this mechanism acts synergistically with the remodeling of the gut microenvironment to suppress CRC progression.
Materials and Methods
Database and Materials
Databases utilized in this study included PubChem (https://pubchem.ncbi.nlm.nih.gov), UniProt (https://www.uniprot.org), GeneCards (https://www.genecards.org), BATMAN-TCM (http://bionet.ncpsb.org/batman-tcm/), and DAVID (https://david.ncifcrf.gov/). Network analysis was conducted using the STRING online platform (https://string-db.org/), while Cytoscape software was employed for visualization.
UDCA was procured from Jiangyin Tianjiang Pharmaceutical Co., Ltd. (China), while azoxymethane (AOM, analytical grade) was obtained from Sigma-Aldrich (USA). Dextran sulfate sodium (DSS) was obtained from Shanghai Yisheng Biotechnology Co., Ltd., and TNF-α, IL-6, IL-17, and IFN-γ assay kits were acquired from Shanghai Enzyme-Linked Biotechnology Co., Ltd. Hematoxylin-eosin (HE) staining kit were obtained from Shanghai Biyuntian Biotechnology Co., Ltd. PCR primers were supplied by Suzhou Jinweizhi Biotechnology Co., Ltd., and 10% formalin fixative was sourced from Solebaol (China). Rabbit polyclonal antibodies against PI3K, p-PI3K, p-AKT, AKT, mTOR, and p-mTOR were purchased from Bioworld Technology, Inc (USA); BCA protein concentration assay reagent (minibioTM) was purchased from Bio Biotechnology Co., LTD (China).
Acquisition of UDCA Targets, Colorectal Cancer Targets and Their Common Targets
UDCA was searched in the PubChem database, and its SMILES notation was retrieved for target prediction using the SwissTargetPrediction platform. The identified targets and gene information were then standardized using the UniProt database. The bile acid InChIKey was input into the BATMAN-TCM database (http://bionet.ncpsb.org.cn/batman-tcm/index.php) to predict potential targets using “Colorectal cancer” as the keyword. The GeneCards database (https://www.genecards.org/) was queried for Colorectal cancer-related targets, though the search yielded an extensive dataset. The Venny tool (https://bioinfogp.cnb.csic.es/tools/venny/) was used to identify the intersection of bile acid-associated targets and Colorectal cancer-related targets, thereby determining potential therapeutic targets for UDCA in Colorectal cancer treatment.11 Thereby determining potential therapeutic targets for UDCA in Colorectal cancer treatment. Specifically, a probability threshold of > 0.1 was applied in SwissTargetPrediction to ensure target reliability.
Construction of Potential Target PPI Network and Screening of Core Targets
The common target genes of UDCA and Colorectal cancer were uploaded to the STRING online analysis platform (https://string-db.org/) to construct the protein-protein interaction (PPI) network. The species parameter was set to “Homo sapiens,” and the minimum interaction score was adjusted to 0.9 to ensure data reliability, while all other settings remained at their default values. The results were saved in TSV format, imported into Cytoscape 3.9.2, and subsequently analyzed. Cytoscape software was utilized to visualize node degree values using variations in size and color.12
GO and KEGG Pathway Enrichment Analysis of Core Target Genes
The intersecting genes were input into the DAVID database for Gene Ontology (GO) functional enrichment analysis and Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway enrichment analysis. Pathways and GO terms with a p-value < 0.01 were filtered from the enriched items. The top 30 pathways and top 10 GO terms, ranked by count values, were selected and visualized.13
Molecular Docking
The 2D structure of UDCA was retrieved from the PubChem database (http://pubchem.ncbi.nlm.nih.gov/) and converted into a 3D structure using ChemOffice. The resulting structure was saved in MOL2 format. The RCSB PDB database (http://www.rcsb.org/) was used to screen protein targets, selecting high-resolution crystal structures as molecular docking receptors. PyMOL software was used to remove water molecules and phosphate groups from the protein structure, which was then saved as a PDB file. The Molecular Operating Environment (MOE) 2019 software was employed for energy minimization of the compounds, preprocessing of target proteins, and identification of active binding pockets. Finally, molecular docking was performed using MOE 2019. The selection of MOE 2019 was based on its advanced AMBER14:EHT force field and refined scoring algorithms, which provide superior accuracy in identifying active binding pockets and estimating binding energies compared to standard docking tools. Binding affinity was assessed based on binding energy, and the results were visualized using PyMOL and Discovery Studio software.14
Animal Experiment Verification
Experimental Animals and Grouping
This study complies with the ARRIVE guidelines. The experimental design was approved by the Experimental Animal Ethics Committee of Nanchang Medical College (NYLLSC20241206). A total of 50 male Balb/c mice were randomly assigned to five groups: normal group, model group, low-dose UDCA group (60 mg/kg), medium-dose UDCA group (90 mg/kg), and high-dose UDCA group (120 mg/kg), with 10 mice per group. Randomization was performed based on body weight to ensure baseline physiological consistency across all groups. From the 8th day onward, UDCA was administered once daily via oral gavage at doses of 60, 90, and 120 mg/kg, respectively. Doses were determined based on preliminary screening and conversion of human therapeutic doses. Male mice were chosen to minimize variability, as female mice are known to exhibit resistance to AOM/DSS-induced tumorigenesis due to hormonal protection. Following a one-week acclimatization period, all groups except the normal group received an intraperitoneal injection of 10 mg/kg AOM on the first day. To induce Colorectal cancer, a 3% DSS solution was administered during the 2nd, 5th, and 8th weeks. From the 8th day onward, based on preliminary experimental results, the low-dose (60 mg/kg), medium-dose (90 mg/kg), and high-dose (120 mg/kg) UDCA groups were administered UDCA via oral gavage. Body weight was monitored throughout the experiment. At the end of the experiment, the mice were euthanized, and abdominal aortic blood and intact colorectal tissues were collected. Four intestinal tissue samples were randomly selected from each group for paraffin embedding and sectioning. A portion of the remaining six tissues was used to prepare a single-cell suspension, while the rest was stored at −80°C for further analysis.
Detection of Cytokines in Serum of Mice
The expression levels of tumor necrosis factor-α (TNF-α), interleukin-6 (IL-6), interleukin-4 (IL-4), and interferon-α (IFN-α) were measured using ELISA kits following the manufacturer’s instructions.
Pathological Morphology of Intestinal Tissue
Colon tissue samples were excised, fixed in 4% paraformaldehyde, embedded in paraffin, stained, and examined under a microscope. Pathological changes were assessed.
The mRNA Expression Levels of CYP19A1, HMGCR, RARA, NR0B2 and EGFR in Colon Tissues Were Detected by qRT-PCR
A total of 20 mg of colon tissue was homogenized in an ice bath, followed by the addition of 500 μL of isothiocyanate solution and 10 μL of mercaptoethanol. Following homogenization, the mixture was centrifuged at 13,000×g for 10 minutes at 4°C, and the supernatant was collected. The supernatant was transferred to a DNA enrichment tube and centrifuged at 4°C for 2 minutes to remove DNA. The supernatant was discarded, and an appropriate volume of absolute ethanol was added, mixed, and transferred to an RNA enrichment tube. RNA was then separated from proteins by centrifugation at 4°C for 2 minutes. The RNA precipitate was resuspended in a small volume of deionized water or buffer solution, followed by two rounds of purification using an RNA cleaning and concentration buffer. Subsequently, 30 μL of 0.1% DEPC-treated water was added to the purified RNA and centrifuged at 13,000×g for 5 minutes at 4°C. The final purified RNA was collected for downstream analysis. The primers used in this study are listed in Table 1, with β-actin selected as the reference gene for normalization. Relative quantification of target gene mRNA expression was performed using the 2−ΔΔCt method, with all experiments conducted in triplicate.
 |
Table 1 Information for Primer Sequences |
Detection of Diversity and Genus Level Changes of Intestinal Flora in Various Mice
Total genomic DNA was extracted from fecal samples using the GeneJET Genomic DNA Purification Kit. Due to sample storage constraints during the initial collection phase, a representative subset of n=4 mice per group was randomly selected for 16S rRNA high-throughput sequencing. The V3-V4 hypervariable regions of the 16S rRNA gene were amplified using primers 338F and 806R. PCR amplicons were purified using the AxyPrep DNA Gel Extraction Kit, quantified via the QuantiFluor™-ST system, and paired-end sequenced on an Illumina MiSeq PE300 platform (Shanghai Meiji Biomedical Technology Co., Ltd).Bioinformatics analysis was performed using the QIIME 2 platform (version 2022.2). Raw sequence data were demultiplexed, quality-filtered using fastp, and merged using FLASH. Amplicon sequence variants (ASVs) were generated using the DADA2 plugin for denoising. Taxonomic assignment was performed using the Naïve Bayes classifier trained on the SILVA 138 reference database. Alpha diversity indices (Chao1, Shannon) and Beta diversity metrics (Bray-Curtis distance) were calculated using QIIME 2 scripts.
Detection of Short-Chain Fatty Acids
For short-chain fatty acid (SCFA) analysis, 0.1 g of intestinal contents from each group of mice was precisely weighed and homogenized with 1 mL of PBS solution via vortex mixing to obtain a 10% (w/v) fecal dilution. Subsequently, 500 μL of the fecal dilution was combined with 100 μL of a crotonic acid-meta-phosphoric acid mixture and vortexed. The samples were then stored at −20 °C for 24 h. Following incubation, samples were thawed at 4 °C and centrifuged. The supernatant was collected, filtered through a 0.45-μm aqueous phase membrane, and subjected to further analysis. Using crotonic acid as an internal standard, SCFA concentrations in the sample solutions were quantified via gas chromatography (GC) on a system equipped with a DB-FFAP column (Agilent; 0.32 mm × 30 m × 0.5 μm).
Western Blot Was Used to Detect the Expression of Related Proteins in the PI3K/Akt/mTOR Signaling Pathway
The expression levels of PI3K, Akt, and mTOR signaling proteins in tumor tissues were analyzed by Western blot. Tumor tissues were homogenized and centrifuged at 12,000 rpm for 15 min, and the supernatant was collected to determine total protein concentration using the bicinchoninic acid (BCA) assay. Protein samples were separated by 10% SDS-PAGE and transferred onto PVDF membranes. The membranes were washed three times with TBST buffer (5 min per wash) and blocked with 5% skim milk powder at room temperature for 2 h. After three additional TBST washes (10 min each), the membranes were incubated overnight at 4°C with the following primary antibodies (all purchased from Abmart, Shanghai, China): rabbit anti-p-PI3K (1:1,000), PI3K (1:500), p-Akt (1:1,000), Akt (1:1,000), p-mTOR (1:500), mTOR (1:500), Foxp3 (1:500), and ROR-γt (1:1,000). Following TBST washes, HRP-conjugated goat anti-rabbit IgG (1:5,000) was added, and the membranes were incubated for 1 h at room temperature. After final TBST washes, protein bands were visualized using a chemiluminescence reagent. Gray values were quantified, and relative protein expression levels were normalized to controls.
Data Analysis
Statistical analyses were performed using SPSS 19.0 (IBM) and GraphPad Prism 8.02. All experiments utilized n = 10 biologically independent mice per group, with the exception of microbiome profiling (n = 4/group), and results are expressed as mean ± SD. Prior to hypothesis testing, data distributions were assessed for normality using the Shapiro–Wilk test and for homogeneity of variance using Levene’s test. Data satisfying normal distribution assumptions (eg, body weight, colon length, cytokine levels) were analyzed using One-way ANOVA followed by Tukey’s post-hoc test, while non-normally distributed data were evaluated using the Kruskal–Wallis test with Dunn’s multiple comparisons. To rigorously address the limited sample size (n=4) and compositional nature of the 16S rRNA data, a non-parametric resampling framework was employed; specifically, group differences were tested via permutation-based ANOVA (10,000 iterations), with pairwise comparisons conducted using post-hoc permutation t-tests with Bonferroni adjustment to control for false discovery rates. Additionally, to quantify biological relevance beyond p-values, effect sizes were estimated using bootstrap-derived 95% confidence intervals (10,000 resamples). Statistical significance was defined as P < 0.05.
Experimental Results
Results of Network Pharmacology Experiments
Figure 1 shows the results of network pharmacology. Figure 1A presents a Venn diagram depicting the common target genes of UDCA and Colorectal cancer. As illustrated in Figure 1A, UDCA has 148 potential target genes. A total of 10,060 potential target genes were identified for Colorectal cancer. The intersection of UDCA target genes and Colorectal cancer-related target genes was identified using a bioinformatics visualization platform, resulting in 129 common target genes (Figure 1A). The topological properties of the PPI network were analyzed using the CytoNCA plugin, and core targets were identified based on degree centrality. Figure 1B illustrates the PPI network of the common targets between UDCA and colorectal cancer. As depicted in Figure 1B, the nine core target genes of UDCA in Colorectal cancer treatment are SRC, ESR1, EGFR, PPARA, RXRA, NR0B2, RARA, HMGCR, and CYP19A1.Figure 1C presents the Gene Ontology (GO) functional enrichment analysis, while Figure 1D illustrates the Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway enrichment analysis. To further clarify the primary pharmacological mechanisms of UDCA against CRC, the top 5 most significantly enriched GO terms and KEGG pathways were identified and summarized in Table 2 As shown in Figure 1C, GO analysis is categorized into three domains: biological process (BP), molecular function (MF), and cellular component (CC). GO functional enrichment analysis indicated that UDCA plays multiple roles in modulating Colorectal cancer progression. UDCA may influence Colorectal cancer cell proliferation and apoptosis by modulating intracellular receptor signaling pathways and gene transcription. These effects may involve steroid metabolism and hormone-mediated signaling pathways, suggesting a novel mechanism for cancer treatment. Regarding cellular components, the impact of UDCA on cell membrane structure and membrane microdomains is significant, potentially enhancing intercellular signaling and cell adhesion, thereby inhibiting tumor metastasis. Additionally, UDCA may regulate gene expression by binding to nuclear receptors and activating transcription factors, thereby inhibiting cancer cell growth. Furthermore, UDCA may exert anticancer effects by interacting with steroid and prostaglandin receptors, reinforcing its potential in Colorectal cancer treatment. Figure 1D indicates that UDCA may contribute to Colorectal cancer treatment through multiple signaling pathways, encompassing metabolic, immune, endocrine, and hormonal mechanisms. Specifically, UDCA may influence Colorectal cancer initiation and progression by modulating receptor activation mechanisms in chemical carcinogenesis, thereby affecting cell proliferation, apoptosis, and the expression of tumor-related genes. Additionally, UDCA may activate the PPAR signaling pathway, thereby regulating lipid metabolism and anti-inflammatory responses, ultimately influencing Colorectal cancer cell growth and migration. UDCA may exert anticancer effects by modulating the interaction of neuroactive ligands with their receptors, thereby influencing the crosstalk between the nervous system and the gut. Furthermore, UDCA may influence Colorectal cancer development by modulating the estrogen signaling pathway, highlighting its potential role in hormone-related cancers. UDCA may also modulate the immune microenvironment of Colorectal cancer by regulating Th17 cell differentiation, thereby influencing the intestinal immune response. Additionally, UDCA may influence Colorectal cancer cell proliferation and apoptosis by modulating steroid hormone biosynthesis, thereby impacting cancer progression. UDCA may also enhance the efficacy of endocrine therapy in Colorectal cancer by modulating endocrine resistance mechanisms. UDCA modulates the intestinal environment and metabolism in colorectal cancer via bile secretion and adipokine signaling pathways. Additionally, its association with bladder and non-small cell lung cancer mechanisms suggests broad anticancer potential. These findings underscore UDCA’s therapeutic value across various cancer types through diverse molecular pathways.
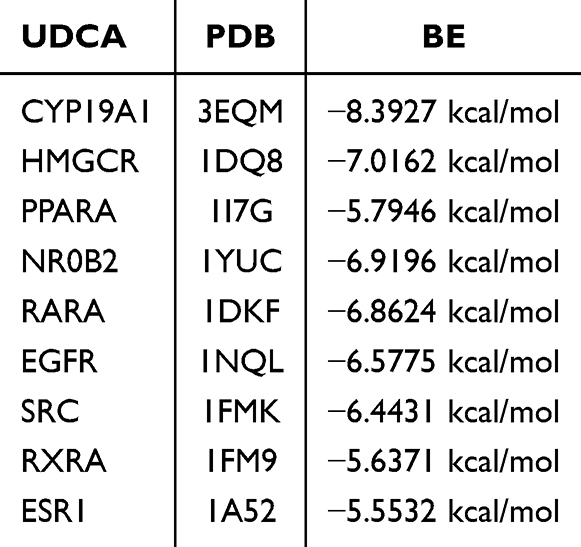 |
Table 2 KEGG Pathway Enrichment Analysis of Potential Targets for UDCA in Colorectal Cancer |
 |
Figure 1 Figure 1. Network pharmacology analysis of UDCA in the treatment of CRC. (A) Venn diagram showing the 129 shared targets between UDCA and CRC. (B) PPI network of shared targets. Node size and color intensity (yellow to red) represent the degree centrality, reflecting the relative importance of the protein in the network. (C) GO enrichment analysis. The bars are categorized by Biological Process (BP, Orange), Cellular Component (CC, green), and Molecular Function (MF, navy). (D) KEGG pathway enrichment analysis. The color gradient indicates the statistical significance, and the bar length represents the gene count. |
Molecular Docking Results
To investigate the interaction between UDCA and its protein targets, molecular docking experiments were conducted using MOE 2019 software. The docking results revealed significant binding activity between UDCA and multiple protein targets, as illustrated in Figure 2. The binding energies between UDCA and these core targets were quantitatively analyzed to evaluate their interaction strength, with the detailed results summarized in Table 3. The molecular docking energy values for UDCA and its protein targets ranged from −5.5532 to −8.3927 kcal/mol. Based on the docking energy criteria, two docking interactions exhibited strong binding activity, while seven demonstrated good binding activity (Figure 2A–J). Notably, UDCA exhibited strong binding affinity with CYP19A1 (−8.3927 kcal/mol) and HMGCR (−7.0162 kcal/mol) (Figure 2A and B). Specifically, in the CYP19A1 protein receptor (Figure 2A), UDCA forms hydrogen bonds with Met374, Arg115, and Ala306, as well as hydrophobic interactions with Leu477, Val370, Cys437, Ile133, and Trp224. In the HMGCR protein receptor (Figure 2B), UDCA forms a hydrogen bond with Leu862 and hydrophobic interactions with His861, Ala856, Leu853, and Leu562. For the PPARA protein receptor (Figure 2C), UDCA forms hydrogen bonds with Thr279 and hydrophobic interactions with Cys276, Val332, Met330, Leu321, Val324, and Met220. In the NR0B2 protein receptor (Figure 2D), UDCA forms hydrogen bonds with Arg393, His390, and Val406, along with hydrophobic interactions with Ile387, Met345, Ile416, Leu424, Met428, and His390.
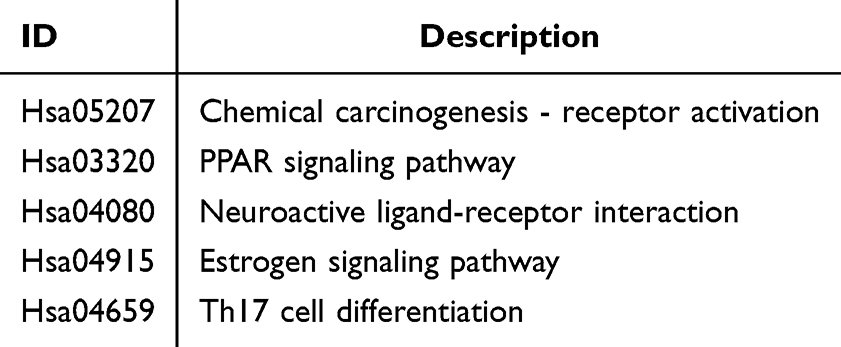 |
Table 3 Molecular Docking Results of UDCA with Potential Target Proteins |
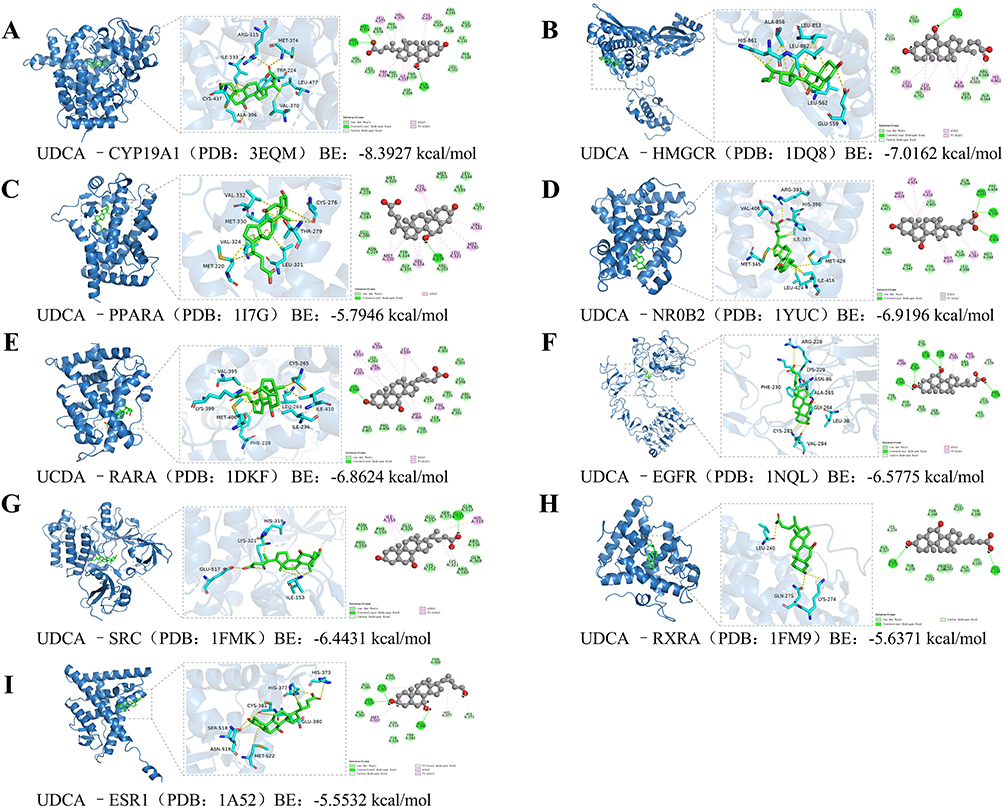 |
Figure 2 Molecular docking simulation revealing the binding modes and affinity between UDCA and core target proteins. Representative 3D and 2D docking poses of UDCA with nine key therapeutic targets are shown. For each panel, the left view displays the global 3D complex structure; the middle view highlights the zoomed-in binding pocket showing critical amino acid residues and hydrogen bonds (indicated by yellow dashed lines); and the right view presents the 2D ligand-receptor interaction diagram. (A) SRC (PDB: 1FMK), (B) ESR1 (PDB: 1A52), (C) EGFR (PDB: 1NQL), (D) PPARA (PDB: 1I7G), (E) RXRA (PDB: 1FM9), (F) NR0B2 (PDB: 1YUC), (G) RARA (PDB: 1DKF), (H) HMGCR (PDB: 1DQ8), and (I) CYP19A1 (PDB: 3EQM). Note: The calculated Binding Energy (BE) is provided below each panel. Lower BE values (≤ −5.0 kcal/mol) indicate higher binding affinity and stability between UDCA and the target protein. |
In the RARA protein receptor (Figure 2E), UDCA forms hydrogen bonds with Lys399 and hydrophobic interactions with Cys365, Ile410, Val395, Ile236, Leu269, Phe228, and Met406. Similarly, in the EGFR protein receptor (Figure 2F), UDCA forms hydrogen bonds with Cys283, Gly364, Leu38, Asn86, and Arg228, along with hydrophobic interactions with Val284, Ala265, Phe230, and Lys229, and hydrocarbon interactions with Lys229 and Cys283. In the SRC protein receptor (Figure 2G), UDCA forms a hydrogen bond with Glu517, a hydrophobic interaction with His319, Lys321, and Ile153, and an electrostatic interaction with Lys321.
Furthermore, in the RXRA protein receptor (Figure 2H), UDCA forms hydrogen bonds with Leu240 and Gln275 and a C-H interaction with Lys274. Finally, in the ESR1 protein receptor (Figure 2I), UDCA forms hydrogen bonds with Cys381, Ser518, and Glu380, hydrophobic interactions with Met522, His377, Cys381, and C-H interactions with His373, His377, and Asn519. These findings suggest that UDCA exhibits significant binding activity with multiple protein targets, primarily through hydrogen bonding and hydrophobic interactions. UDCA may regulate its protein targets through multiple pathways.
Animal Experiment Results
Effects of UDCA on Body Weight, Colon Length and Colon Tissue Pathology in Mice with Colorectal Cancer
Figure 3 illustrates the protective effects of UDCA against AOM/DSS-induced colorectal cancer (CRC) in mice. As shown in Figure 3A, AOM/DSS induction caused significant body weight loss compared to the control group (P < 0.01). However, UDCA administration dose-dependently alleviated this weight loss, with the high-dose group exhibiting the most robust recovery (P < 0.01). Consistently, the model group showed significant colon shortening—a hallmark of inflammation (P < 0.01; Figure 3B). This shortening was significantly mitigated by UDCA treatment in a dose-dependent manner; notably, colon length in the high-dose group was restored to near-control levels. Histological analysis (Figure 3C) revealed that the model group underwent characteristic pathological changes, including inflammatory cell infiltration, crypt distortion (loss of polarity, branching, and abscess formation), goblet cell depletion, and metaplasia (Paneth cell and villous). These dysplastic and inflammatory lesions presented primarily in an intermittent and multifocal pattern. Conversely, UDCA treatment markedly improved colonic mucosal architecture. While the low-dose group showed mild inflammation, the high-dose group demonstrated significant mucosal recovery with minimal inflammatory infiltration. Collectively, these findings indicate that UDCA exerts a potent therapeutic effect against colitis-associated CRC by preserving colonic integrity and mitigating pathological damage.
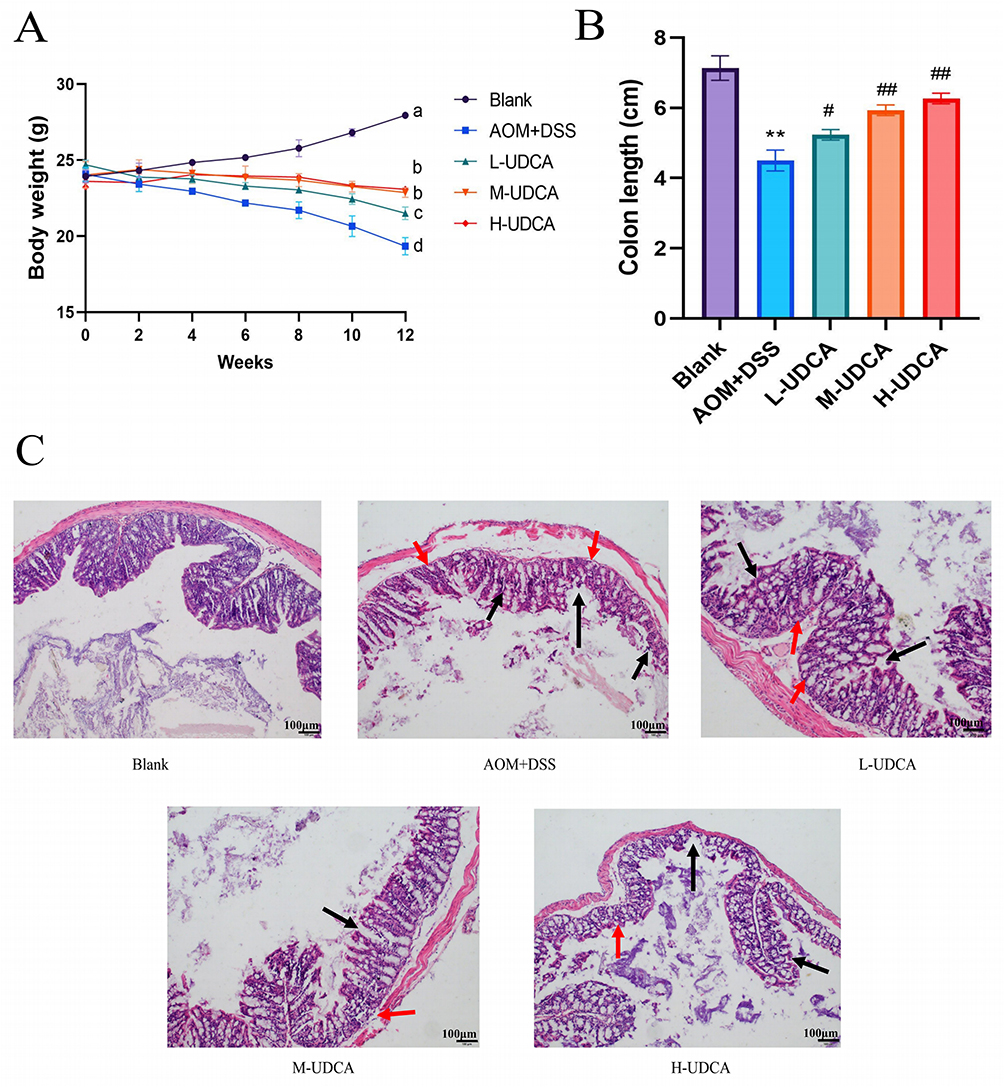 |
Figure 3 Protective effects of UDCA against AOM/DSS-induced colorectal cancer in mice. (A) Changes in body weight during the experimental period. Notes: Different letters indicate statistically significant differences between groups (P < 0.05). (B) Statistical analysis of colon length. (C) Representative H&E staining results of colonic tissues (magnification, 100×). Scale bar = 100 μm. Black arrows indicate inflammatory cell infiltration, and red arrows indicate crypt distortion and dysplasia. UDCA was administered at low (60 mg/kg), medium (90 mg/kg), and high (120 mg/kg) doses. Data are presented as mean ± SD (n = 10). ** p < 0.01 vs the Blank group; # p < 0.05 and ## p < 0.01 vs the Model group. |
Regulatory Effect of UDCA on Serum Inflammatory Factors in Mice with Colorectal Cancer
Figure 4 illustrates the regulatory effects of UDCA on serum cytokine levels. Compared to the control group, the AOM/DSS group exhibited a systemic inflammatory state characterized by significantly upregulated levels of TNF-α, IL-6, and IL-4 (P < 0.01; Figure 4A–C). UDCA administration effectively attenuated these pro-inflammatory cytokines in a dose-dependent manner. Notably, the medium- and high-dose UDCA groups showed the most profound suppression of inflammatory secretion (P < 0.01). Conversely, serum levels of IFN-α—a critical anti-tumor cytokine—were significantly suppressed in the model group (P < 0.01; Figure 4D). UDCA treatment reversed this suppression, with high-dose administration restoring IFN-α levels to near-normal values. Collectively, these data suggest that UDCA mitigates CRC progression by dampening systemic inflammation and reprogramming the immune microenvironment, particularly at medium and high doses.
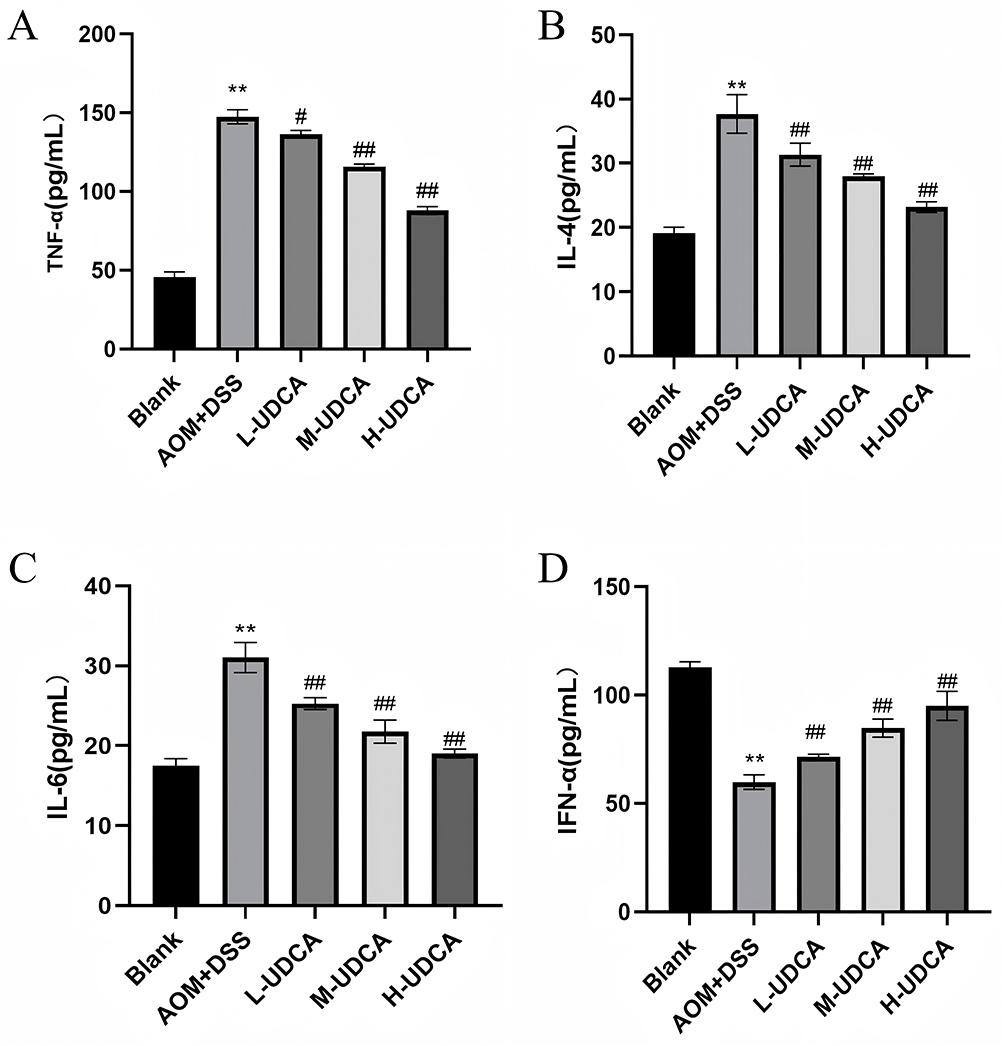 |
Figure 4 Regulatory effects of UDCA on serum inflammatory cytokine levels in AOM/DSS-induced CRC mice. (A) TNF-α concentration. (B) IL-4 concentration. (C) IL-6 concentration. (D) IFN-α concentration. Note: The levels of cytokines were detected by ELISA. Data are presented as mean ± SD. UDCA dosages: low (60 mg/kg), medium (90 mg/kg), and high (120 mg/kg). ** P < 0.01 vs the Blank group; # P < 0.05, ## P < 0.01 vs the Model group. |
Effect of UDCA on the mRNA Expression Levels of CYP19A1, HMGCR, RARA, NR0B2 and EGFR in Colon Tissue of Mice with Colorectal Cancer
Figure 5 illustrates the effect of UDCA on the mRNA expression of CYP19A1, HMGCR, RAR-α, NR0B2, and EGFR in a mouse model of AOM+DSS-induced Colorectal cancer. The results indicated that, compared with the blank control group, CYP19A1 mRNA expression was significantly downregulated (Figure 5A, P < 0.01), whereas HMGCR (Figure 5B), NR0B2 (Figure 5D), and EGFR (Figure 5E) were significantly upregulated (P < 0.01) in the AOM+DSS-induced model group. However, RAR-α expression did not differ significantly between the model and blank control groups (Figure 5C).
 |
Figure 5 Effect of UDCA on the mRNA expression of CYP19A1, HMGCR, RARA, NR0B2, and EGFR in colonic tissues. (A) Relative mRNA expression of CYP19A1. (B) Relative mRNA expression of HMGCR. (C) Relative mRNA expression of RARA (RAR-α). (D) Relative mRNA expression of NR0B2. (E) Relative mRNA expression of EGFR. The relative mRNA expression levels were determined by qRT-PCR. UDCA was administered at low (60 mg/kg), medium (90 mg/kg), and high (120 mg/kg) doses. Data are expressed as mean ± SD (n = 10). ** P < 0.01 vs the Blank group; # P < 0.05, ## P < 0.01 vs the AOM+DSS group (Model). |
In the UDCA-treated groups, the expression levels of these genes exhibited varying degrees of regulation. Specifically, CYP19A1 expression was significantly restored in the M-UDCA and H-UDCA groups (Figure 5A, P < 0.01), approaching control levels. HMGCR expression was reduced across all UDCA-treated groups, with the most pronounced decrease observed in the M-UDCA and H-UDCA groups (Figure 5B, P < 0.01). Additionally, NR0B2 and EGFR expression was significantly reduced in the M-UDCA and H-UDCA groups (Figure 5D and E, P < 0.01), approaching levels observed in the blank control group. However, L-UDCA exhibited a comparatively weaker downregulating effect on EGFR (Figure 5E, P < 0.05). In contrast, RAR-α expression remained unchanged across all UDCA-treated groups (Figure 5C), suggesting that this gene may not be a primary target of UDCA.
In conclusion, UDCA treatment significantly modulated the expression of CYP19A1, HMGCR, NR0B2, and EGFR, particularly at medium and high doses, where their expression levels closely resembled those of the blank control group. These findings suggest that UDCA may exert anti-Colorectal cancer effects. However, RAR-α expression remained unaltered among the groups, suggesting that it may not be involved in UDCA-mediated anti-tumor effects.
Effect of UDCA on the Expression of Phosphorylated Proteins of PI3K/AKT/mTOR Signaling Pathway in Colon Cancer Tissues of Mice
The Western blot results are presented in Figure 6. The phosphorylation levels of PI3K, Akt, and mTOR in colon cancer tissues of mice were markedly elevated compared to the control group (P < 0.01). Following UDCA treatment, phosphorylated PI3K, Akt, and mTOR protein levels were significantly reduced in a dose-dependent manner (P < 0.01) (Figure 6A–H). In contrast, total PI3K, Akt, and mTOR protein expression remained unchanged across all groups.
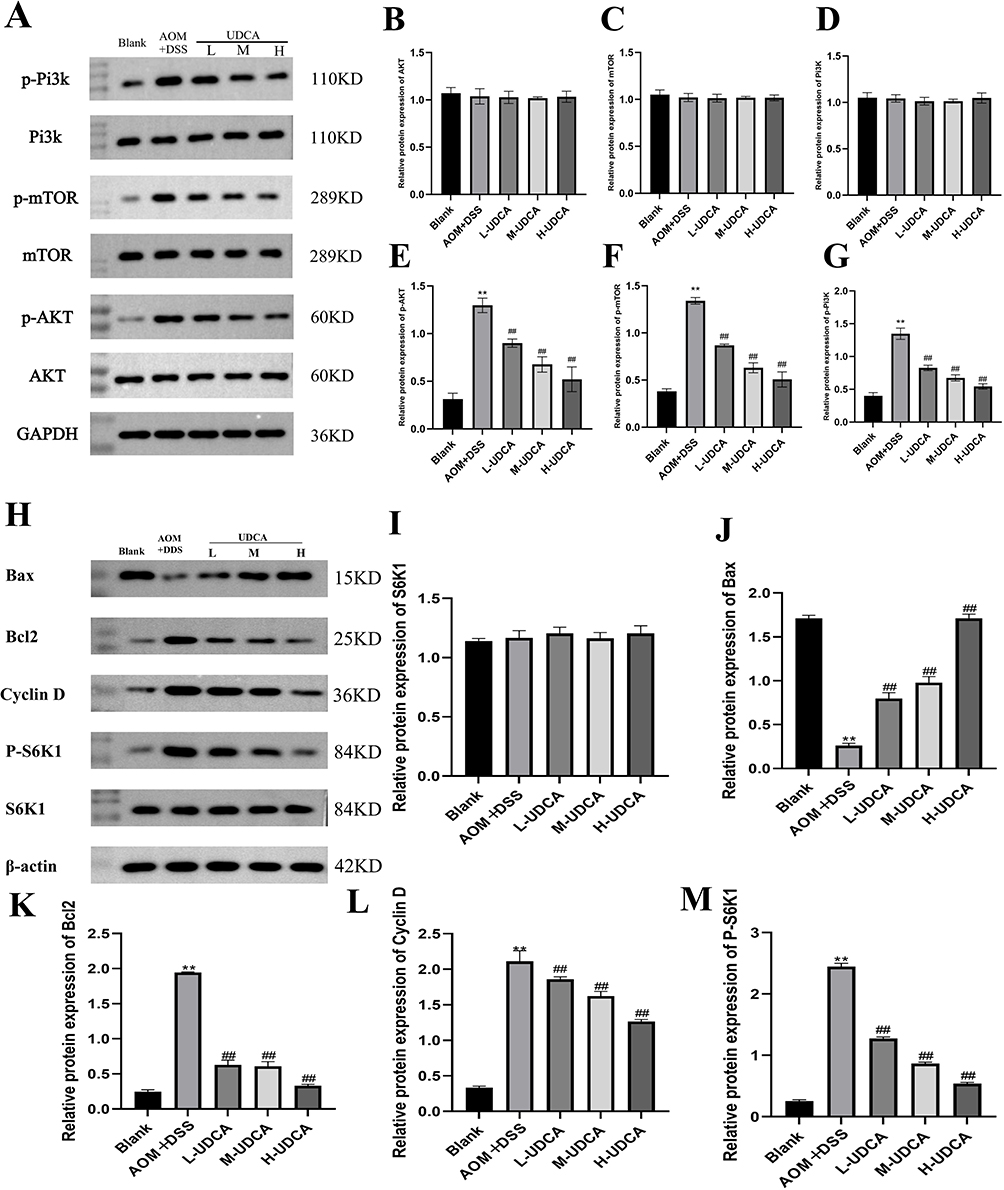 |
Figure 6 Effect of UDCA on the expression of phosphorylated proteins and downstream functional effectors of the PI3K/Akt/mTOR signaling pathway in colon cancer tissues. (A) Representative Western blot bands showing the expression of p-PI3K, PI3K, p-mTOR, mTOR, p-AKT, and AKT. GAPDH was used as a loading control (B–G) Quantitative analysis of the relative protein expression levels for the core PI3K/Akt/mTOR signaling components. (H) Representative Western blot bands showing the expression of downstream functional markers, including the pro-apoptotic protein Bax, anti-apoptotic protein Bcl2, cell cycle regulator Cyclin D, and the phosphorylated and total forms of the mTOR substrate S6K. β-actin was used as the internal loading control for this specific set. (I–M) Quantitative analysis of the relative protein expression levels of S6K1, Bax, Bcl2, Cyclin D, and P-S6K1, respectively. Data are presented as mean ± SD. ** P < 0.01 vs the Blank group; ## P < 0.01 vs the Model group. |
To further evaluate the downstream functional consequences of this signaling inhibition, we examined the expression of key effectors related to apoptosis, cell cycle, and protein synthesis (Figure 6H–M). Compared to the blank group, the model group showed a significant downregulation of the pro-apoptotic protein Bax (Figure 6J, P < 0.01) and a marked upregulation of the anti-apoptotic protein Bcl2 (Figure 6K, P < 0.01). UDCA treatment effectively reversed this trend, significantly increasing Bax expression and decreasing Bcl2 levels, thereby restoring the Bax/Bcl2 balance. Additionally, the elevated levels of Cyclin D, a critical cell cycle regulator, were dose-dependently suppressed by UDCA (Figure 6L, P < 0.01). Furthermore, UDCA treatment led to a significant reduction in the phosphorylation of S6K1 (P-S6K1), the primary downstream substrate of mTOR, while total S6K1 remained constant (Figure 6I and M, P < 0.01). Collectively, these findings demonstrate that UDCA ameliorates colon cancer progression not only by inhibiting the core PI3K/Akt/mTOR pathway activation but also by subsequently modulating downstream markers to promote apoptosis and arrest the cell cycle.
Effect of UDCA on Intestinal Flora in Mice with Colorectal Cancer
Permutation-based ANOVA revealed significant overall group differences in microbial metrics (P < 0.01). Specifically, DSS-induced colorectal cancer (CRC) modeling profoundly reduced alpha diversity versus normal controls (Shannon/Chao1 indices: P < 0.01), while UDCA intervention significantly restored diversity relative to model group (P < 0.01), indicating attenuation of CRC-associated richness depletion (Figure 7A and B). Permutational multivariate analysis (PERMANOVA, Bray-Curtis) confirmed compositional divergence: normal vs model (P < 0.01) and model vs UDCA (P < 0.01), with UDCA-treated microbiota exhibiting closer PCoA proximity to controls, suggesting structural normalization (Figure 7C). Critically, genus-level permutation tests identified CRC-associated depletion of beneficial Bacteroides (P < 0.01) and Lactobacillus (P < 0.01), alongside enrichment of pathobionts Pseudomonasand Helicobacter (P < 0.01 for both); these dysbiotic patterns were significantly reversed by UDCA (P < 0.01), correlating with a probiotic-enriched profile (Figure 7D–H). Interpretation note: Statistical inferences are constrained by n=4/group and observed microbiota remodeling represents associative relationships requiring functional validation.
 |
Figure 7 Analysis of intestinal microbiota diversity in each group of mice (A) Chao1 index, (B) Shannon index, and (C) principal component analysis, (D) bar chart of relative abundance of species at genus level, (E) relative abundance of Bacteroides, (F) relative abundance of Pseudomonas, (G) relative abundance of Lactobacillus and (H) relative abundance of Helicobacter (low dose = 60 mg/kg, medium dose = 90mg/kg and high dose = 120 mg/kg). Note: TData represent median ± interquartile range (IQR) derived from n = 4 biologically independent mice per group. Statistical analysis: Permutation-based ANOVA (10,000 iterations) with post-hoc permutation t-tests (Bonferroni-adjusted). Effect sizes shown with bootstrap 95% CIs (10,000 resamples). **P < 0.01 vs the blank group; #P < 0.05, ##P < 0.01 vs the model group. |
UDCA Restores Intestinal Metabolic Homeostasis by Upregulating Short-Chain Fatty Acids
Figure 8 illustrates the impact of UDCA on the metabolic profile of short-chain fatty acids (SCFAs) in colonic contents. Given the pivotal role of SCFAs in maintaining gut homeostasis, we quantified their levels across different groups. Compared to the control group, the colon cancer model mice exhibited a marked depletion of total SCFAs (Figure 8F), including significant reductions in acetate (Figure 8A), propionate (Figure 8B), butyrate (Figure 8C), isobutyrate (Figure 8D), and valerate (Figure 8E) (P < 0.01). Strikingly, UDCA treatment effectively reversed this metabolic dysregulation. The administration of UDCA significantly upregulated these SCFAs (Figure 8A–F, P < 0.01), restoring their concentrations to levels comparable to those of the healthy controls. These findings suggest that UDCA promotes the production of beneficial metabolites, which may contribute to the improvement of mucosal barrier function and anti-tumor immunity.
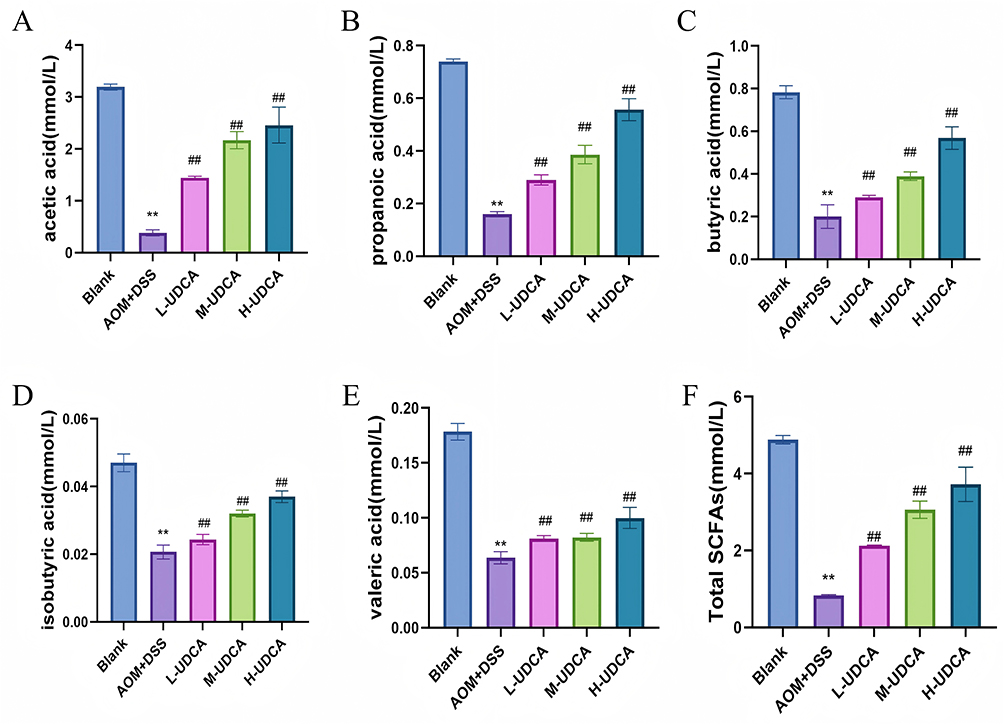 |
Figure 8 Effect of UDCA on the levels of Short-Chain Fatty Acids (SCFAs) in intestinal contents. The concentrations of SCFAs were determined by GC. (A) Acetic acid. (B) Propionic acid. (C) Butyric acid. (D) Isobutyric acid. (E) Valeric acid. (F) Total SCFAs. Note: UDCA dosages: low (60 mg/kg), medium (90 mg/kg), and high (120 mg/kg). Data are expressed as mean ± SD (n = 10). ** P < 0.01 vs the Blank group; ## P < 0.01 vs the Model group. |
Discussion
By integrating network pharmacology, molecular docking, and in vivo experiments, this study investigated the specific molecular targets governing the therapeutic effects of UDCA in Colorectal cancer (CRC). Network pharmacological predictions identified common targets of UDCA and CRC, including genes such as SRC, ESR1, EGFR, PPARA, RXRA, NR0B2, RARA, HMGCR, and CYP19A1, which are extensively involved in cell proliferation, apoptosis, metabolism, and immune regulation. KEGG enrichment analysis further suggested that UDCA may exert anti-tumor effects through the PPAR signaling pathway, Th17 cell differentiation, and bile secretion pathways. Molecular docking results revealed that UDCA exhibited strong binding affinity with core targets (binding energy ≤ −7.0 kcal/mol), implying a capacity for direct interaction, particularly with the upstream receptor EGFR. Consequently, CYP19A1, HMGCR, RARA, NR0B2, and EGFR were identified as potential therapeutic targets. Further qRT-PCR results confirmed that UDCA treatment significantly mitigated the aberrant expression profiles of these genes, specifically reversing the downregulation of CYP19A1 mRNA and the upregulation of HMGCR, EGFR, and NR0B2 mRNA in the model group.
The modulation of CYP19A1 is particularly intriguing. The cytochrome P450 superfamily plays a crucial role in physiological processes. While CYP1A1 is a well-known downstream target of the AhR pathway associated with colitis remission,15–17 CYP19A1 (aromatase) shares significant structural homology with CYP1A1, including the heme-binding domain and substrate recognition sites.18 Although no direct association between CYP19A1 and CRC has been definitively established, Chen et al found that proinflammatory cytokines (IL-1β, TNF-α) in the tumor microenvironment may suppress CYP19A1 expression via JAK/STAT signaling.19 Our findings indicate that UDCA treatment restores CYP19A1 levels. While we cannot yet conclude this is a driver of efficacy, the restoration of CYP19A1 accompanies the anti-tumor effects, suggesting it may serve as a biomarker of resolving inflammation. While CYP19A1 (aromatase) is traditionally associated with tumor promotion in estrogen-dependent tissues like the breast, its role in the colon appears to be protective. Local estrogen production in the colonic mucosa primarily signals through Estrogen Receptor beta (ERβ) to exert anti-inflammatory and pro-apoptotic effects. Our results show that UDCA restores CYP19A1 mRNA levels, which correlates with the suppression of pro-inflammatory cytokines and the inhibition of the PI3K/Akt/mTOR oncogenic axis. We hypothesize that UDCA-mediated CYP19A1 upregulation may facilitate the restoration of a protective hormonal microenvironment, thereby synergistically contributing to the overall anti-tumor efficacy. However, further studies utilizing specific ERβ antagonists or direct measurements of local estrogen levels are required to definitively confirm this hierarchical relationship. Additionally, hyperactivation of HMGCR, NR0B2, and EGFR is strongly associated with cell proliferation. Jiang et al20 found that HMGCR promotes tumor growth, we observed reduced HMGCR expression following UDCA treatment. Similarly, while NR0B2 (SHP) loss is often linked to metabolic abnormalities,21 our data in this specific AOM/DSS model showed elevated NR0B2 in the model group, which was suppressed by UDCA. Most critically, our findings confirm that UDCA ameliorates colitis-associated tumorigenesis effectively associated with the suppression of aberrant EGFR activation. Conversely, RAR-α expression remained unchanged, suggesting UDCA exerts its effects independently of this receptor.
Intestinal dysbiosis is a critical driver of the inflammation-carcinoma sequence.22–24 To investigate the effects of UDCA on this ecological network, we analyzed microbial composition. The AOM/DSS model exhibited reduced α-diversity and an increase in pathogenic genera (Pseudomonas, Helicobacter) with a concomitant decrease in beneficial genera (Bacteroides, Lactobacillus). UDCA intervention was associated with a shift toward normalization. Pseudomonas, an opportunistic pathogen known to activate NF-κB and promote inflammation,25–28 and Helicobacter, a known carcinogen,29,30 were significantly reduced. Conversely, Lactobacillus, which strengthens the intestinal barrier via the CLA-PPAR-γ axis,31 was enriched, aligning with observations of its depletion in CRC patients.32 These microbial shifts correlate with the reduced inflammatory burden observed, suggesting that gut microbiota remodeling may be a contributing factor to the therapeutic milieu created by UDCA. The EGFR-mediated PI3K/Akt/mTOR signaling pathway constitutes a central regulatory network governing cell proliferation and survival. Dysregulation of this axis, driven by PIK3CA mutations or PTEN loss, is a hallmark of CRC pathogenesis.33 EGFR activates this cascade via intrinsic tyrosine kinase activity and Src kinase activation, fueling tumor growth and angiogenesis. A key finding of our study is the identification of EGFR as a direct molecular target, suggesting that UDCA functions partially as a natural EGFR inhibitor. Our data demonstrate that UDCA suppresses EGFR transcript levels and inhibits downstream PI3K/Akt/mTOR phosphorylation. To further substantiate the direct nature of this molecular intervention, we conducted a deeper exploration of the downstream functional effectors within the PI3K/Akt/mTOR cascade. The observed shift in the Bax/Bcl2 ratio and the dose-dependent suppression of Cyclin D and P-S6K1—occurring in the absence of changes to total protein expression—collectively underscore that UDCA acts as a specific modulator that directly disrupts the phosphorylation relay of this oncogenic axis. This distinction between specific phosphorylation inhibition and stable total protein stability effectively differentiates UDCA’s direct signaling interference from broader, indirect consequences of an improved microenvironment. This provides a precise molecular mechanism distinct from broad anti-inflammatory effects. These mechanistic insights provide a rationale for repurposing UDCA. By targeting the EGFR-PI3K axis, UDCA offers a potential adjuvant strategy, particularly in combination with chemotherapies like 5-fluorouracil. Unlike conventional targeted therapies that often induce toxicity, UDCA possesses a favorable safety profile. In our study, high-dose UDCA (120 mg/kg) was well-tolerated and even improved body weight recovery compared to the model group, indicating an absence of overt systemic toxicity. We acknowledge that the AOM/DSS model primarily mimics CAC, which follows an “inflammation-dysplasia-carcinoma” sequence, whereas sporadic CRC typically arises from the “adenoma-carcinoma” sequence initiated by APC mutations. However, the findings of this study may still have broader implications for sporadic CRC for several reasons. First, the EGFR/PI3K/Akt/mTOR signaling axis, which we identified as the primary target of UDCA, is a central oncogenic driver frequently hyperactivated in both CAC and sporadic CRC. Second, although sporadic CRC lacks a history of overt clinical colitis, “smoldering” or subclinical inflammation within the tumor microenvironment is known to promote its progression. Given that UDCA demonstrates multi-faceted roles in regulating bile acid metabolism and restoring gut microbiota homeostasis—factors that are critically involved in the etiology of both types of CRC—it is plausible that UDCA could exert similar anti-tumor effects in sporadic cases. Nonetheless, further validation using models such as ApcMin/+ mice or patient-derived organoids is needed to confirm these findings in the context of sporadic tumorigenesis.”
However, several limitations of this study must be acknowledged. First, regarding the microbiome analysis, the limited sample size (n=4 per group) is a notable constraint. Although we employed rigorous permutation-based statistical methods (10,000 iterations) and bootstrap validation to mitigate false discoveries, the observed taxonomic alterations should be interpreted as exploratory associations rather than definitive evidence of microbiota-driven efficacy. Future studies incorporating larger cohorts and fecal microbiota transplantation (FMT) are essential to establish a definitive causal link. Second, while our study focused on systemic cytokines and intracellular signaling, we identified that UDCA significantly and dose-dependently reduced serum IL-6 levels. Given that IL-6 is an indispensable cytokine for the differentiation of naïve T cells into Th17 cells, its suppression by UDCA likely interrupts the initial stages of Th17 lineage commitment. This aligns with our KEGG enrichment analysis showing the involvement of the Th17 cell differentiation pathway. Although direct quantification of T-cell subsets via flow cytometry was not performed in this study, the robust reduction in IL-6 provides strong circumstantial evidence for an attenuated Th17-mediated inflammatory response. Third, the AOM/DSS model primarily recapitulates the transition from chronic inflammation to dysplasia and early tumorigenesis rather than advanced metastatic disease. Regarding the experimental design, UDCA administration was initiated on day 8, following the AOM injection and the first cycle of DSS induction. This timing represents an “interventional prevention” model targeting the early lesion stage, rather than a purely prophylactic approach or a treatment for established macroscopic tumors. At this juncture, the colonic tissue has undergone initial genetic insult and is experiencing peak acute inflammation, which constitutes the “critical window” for the transition from inflammation to dysplasia. By intervening at this stage, we demonstrate UDCA’s capacity to interrupt the malignant transformation process by suppressing early molecular dysregulation and reshaping the immune-metabolic microenvironment. It is plausible that the antitumor efficacy of UDCA might be attenuated if treatment were delayed until the formation of visible tumors, as advanced lesions often develop significant autonomy and a more consolidated, immunosuppressive microenvironment. Therefore, our findings emphasize UDCA’s potential as a potent agent for interrupting the “inflammation-to-carcinoma” progression, offering particular relevance for high-risk populations, such as patients with chronic inflammatory bowel disease (IBD). Finally, while we did not observe overt toxicity, comprehensive toxicological evaluations, including liver enzymes and lipid profiles, were not performed in this efficacy-focused study and will be required in future pre-clinical assessments. Furthermore, while our study establishes a robust correlation between UDCA, EGFR inhibition, and microbiota restoration, future genetic knockdown or knockout studies are warranted to definitively confirm the hierarchical relationship between these molecular and microbial changes.
Conclusions
In summary, this study utilized an integrated network pharmacology and in vivo approach to investigate the molecular mechanisms of UDCA in colitis-associated tumorigenesis. Our findings suggest that UDCA exerts its therapeutic effects primarily through direct inhibition of the EGFR-mediated PI3K/Akt/mTOR signaling axis, accompanied by the partial restoration of intestinal immune-metabolic homeostasis. These mechanistic insights provide a molecular rationale for the clinical repurposing of UDCA. Future research should focus on exploring the synergistic effects of UDCA with existing chemotherapy regimens and validating the causal contributions of the observed microbiota and metabolic alterations.
Data Sharing Statement
The data that support the findings of this study are available from the corresponding author upon reasonable request.
Ethics Approval and Consent to Participate
The experimental protocols were approved by the Experimental Animal Ethics Committee of Nanchang Medical College (Approval No. NYLLSC20241206). All animal procedures were performed in strict accordance with the “Guide for the Care and Use of Laboratory Animals” (National Research Council, 8th Edition) to ensure the welfare of the laboratory animals. Regarding the human data utilized in the network pharmacology analysis, all data were obtained from legally public and open databases (including TCMSP, GeneCards, and GEO). No recruitment of human subjects or collection of clinical samples was performed in this study. In accordance with Article 32, Items 1 and 2 of the “Measures for Ethical Review of Life Science and Medical Research Involving Human Subjects” (issued on February 18, 2023, China), this research is exempt from Institutional Review Board (IRB) approval as it involves the secondary use of de-identified, publicly available data and does not involve privacy or safety risks to human subjects.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This work was supported by Jiangxi Trauma Clinical Medical Research Center (No. 20223BCG74007) Jiangxi Provincial Science and technology program of traditional Chinese Medicine (No.2022A017).
Disclosure
All authors disclosed no conflicts of interest in this work.
References
1. Roesel R, Strati F, Basso C, et al. Combined tumor-associated microbiome and immune gene expression profiling predict response to neoadjuvant chemo-radiotherapy in locally advanced rectal cancer. Oncoimmunology. 2025;14:2465015. doi:10.1080/2162402X.2025.2465015
2. Chen Y, Fang H, Chen H, et al. Bifidobacterium inhibits the progression of colorectal tumorigenesis in mice through fatty acid isomerization and gut microbiota modulation. Gut Microbes. 2025;17:2464945. doi:10.1080/19490976.2025.2464945
3. Katona BW, Shukla A, Hu W, et al. Microbiota and metabolite-based prediction tool for colonic polyposis with and without a known genetic driver. Gut Microbes. 2025;17:2474141. doi:10.1080/19490976.2025.2474141
4. Zheng J, Zhou C, Li Z, et al. Alcaligenes faecalis promotes colitis to colorectal cancer transition through IgA+ B cell suppression and vinculin acetylation. Gut Microbes. 2025;17:2473511. doi:10.1080/19490976.2025.2473511
5. Basu S, Običan SG, Bertaggia E, et al. Unresolved alterations in bile acid composition and dyslipidemia in maternal and cord blood after UDCA treatment for intrahepatic cholestasis of pregnancy. Am J Physiol Gastrointest Liver Physiol. 2025;328:G364–19. doi:10.1152/ajpgi.00266.2024
6. Bessone F, Hillotte GL, Tamagnone N, Arnedillo D, Roma MG. Ursodeoxycholic acid for the management of drug-induced liver injury: role of hepatoprotective and anti-cholestatic mechanisms. J. Clin. Transl. Hepatol. 2025;13:162–168. doi:10.14218/JCTH.2024.00325
7. Francini E, Pazmay GVB, Fumarola S, Procopio AD, Olivieri F, Marchegiani F. Bi-directional relationship between bile acids (BAs) and Gut Microbiota (GM): UDCA/TUDCA, probiotics, and dietary interventions in elderly people. Int J Mol Sci. 2025;26:1759. doi:10.3390/ijms26041759
8. Liu S, Pang Y, Wang X, Lin R, Tunala S. Application Value of novel inflammatory indicators in response to ursodeoxycholic acid therapy in patients with primary biliary cholangitis. Int J Gene Med. 2025;18:897–905. doi:10.2147/IJGM.S493132
9. Fathima A, Jamma T. UDCA ameliorates inflammation driven EMT by inducing TGR5 dependent SOCS1 expression in mouse macrophages. Sci Rep. 2024;14:24285. doi:10.1038/s41598-024-75516-9
10. Biagioli M, Di Giorgio C, Massa C, et al. Microbial-derived bile acid reverses inflammation in IBD via GPBAR1 agonism and RORγt inverse agonism. Biomed Pharmacothe. 2024;181:117731. doi:10.1016/j.biopha.2024.117731
11. Wang Y, Peng X, Qian B, Wang L, Wang J. The integration of metabolites from Forsythia suspensa and gut microbiota ameliorates drug-induced liver injury: network pharmacology and molecular docking studies. Artif. Cells Nanomed. Biotechnol. 2025;53:105–121. doi:10.1080/21691401.2025.2475088
12. Liang H, Chen Z, Zhu M, et al. Efficacy and potential pharmacological mechanism of Astragalus-Salvia miltiorrhiza combination in diabetic nephropathy: integrating meta-analysis, network pharmacology, molecular docking, and experimental validation. Renal Failure. 2025;47:2466116. doi:10.1080/0886022X.2025.2466116
13. Kanehisa M, Furumichi M, Sato Y, Matsuura Y, Ishiguro-Watanabe M. KEGG: biological systems database as a model of the real world. Nucleic Acids Res. 2025;53:D672–d677. doi:10.1093/nar/gkae909
14. Jiang J, Deng X, Xu C, Wu Y, Huang J. Naringenin inhibits ferroptosis to reduce radiation-induced lung injury: insights from network Pharmacology and molecular docking. Pharm Biol. 2025;63:1–10. doi:10.1080/13880209.2025.2465312
15. Griffin D, Carson R, Moss D, et al. Ruthenium drug BOLD-100 Regulates BRAFMT colorectal cancer cell apoptosis through AhR/ROS/ATR signaling axis modulation. Mol Cancer Res. 2024;22:1088–1101. doi:10.1158/1541-7786.MCR-24-0151
16. Yu A, Yoon D, An HB, et al. The regulatory role of ACP5 in the diesel exhaust particle-induced AHR inflammatory signaling pathway in a human bronchial epithelial cell line. Sci Rep. 2025;15:8826. doi:10.1038/s41598-024-84280-9
17. Wang X, Xie X, Li Y, et al. Quercetin ameliorates ulcerative colitis by activating aryl hydrocarbon receptor to improve intestinal barrier integrity. Phytother Res. 2024;38:253–264. doi:10.1002/ptr.8027
18. Ye W, Tang Q, Wang L, et al. Contribution of CYP19A1, CYP1A1, and CYP1A2 polymorphisms in coronary heart disease risk among the Chinese Han population. Funct Integrat Genomics. 2022;22:515–524. doi:10.1007/s10142-022-00850-y
19. Chen Y, Luo X, Li Y, et al. PTX3 impairs granulosa cell function by promoting the secretion of inflammatory cytokines in M1 macrophages via the JAK pathway. Steroids. 2025;214:109541. doi:10.1016/j.steroids.2024.109541
20. Jiang X, Wang M, Cui G, et al. Tetramethylpyrazine attenuates the cancer stem cell like-properties and doxorubicin resistance by targeting HMGCR in breast cancer. Phytomedicine. 2025;136:156344. doi:10.1016/j.phymed.2024.156344
21. Lam KK, Sethi R, Tan G, et al. The orphan nuclear receptor NR0B2 could be a novel susceptibility locus associated with microsatellite-stable, APC mutation-negative early-onset colorectal carcinomas with metabolic manifestation. Genes Chromosomes Cancer. 2021;60:61–72. doi:10.1002/gcc.22904
22. Ruiz-Malagón AJ, Rodríguez-Sojo MJ, Redondo E, Rodríguez-Cabezas ME, Gálvez J, Rodríguez-Nogales A. Systematic review: the gut microbiota as a link between colorectal cancer and obesity. Obesity Rev. 2025;26:e13872. doi:10.1111/obr.13872
23. Pan L, Wang X, Yang B, Liu Y, Tang D. Importance of intestinal microflora: dried toad skin-radix clematidis plasma component analysis and anti-CRC core target study. J. Pharm. Biomed. Anal. 2025;260:116802. doi:10.1016/j.jpba.2025.116802
24. Li Z, Deng L, Cheng M, et al. Emerging role of bile acids in colorectal liver metastasis: from molecular mechanism to clinical significance (Review. Int J Oncol. 2025;66:1–18. doi:10.3892/ijo.2025.5730
25. Swart AL, Laventie BJ, Sütterlin R, et al. Pseudomonas aeruginosa breaches respiratory epithelia through goblet cell invasion in a microtissue model. Nature Microbiology. 2024;9:1725–1737. doi:10.1038/s41564-024-01718-6
26. Zelikman S, Dudkevich R, Korenfeld-Tzemach H, et al. PemB, a type III secretion effector in Pseudomonas aeruginosa, affects Caenorhabditis elegans life span. Heliyon. 2024;10:e29751. doi:10.1016/j.heliyon.2024.e29751
27. Lee DH, Seol M, Lee YR, et al. Difference of colorectal cancer microbial community by metagenomics and culture-based methods. 2019;37:489.
28. Affhan S, Dachang W, Xin Y, Shang D. Lactic acid bacteria protect human intestinal epithelial cells from Staphylococcus aureus and Pseudomonas aeruginosa infections. Genet. Mol. Res. 2015;14:17044–17058. doi:10.4238/2015.December.16.5
29. Frías-Ordoñez JS, Riquelme A, Marulanda-Fernandez H, et al. Helicobacter pylori and colorectal cancer: meeting sir austin bradford hill’s causality criteria. Helicobacter. 2025;30:e70024. doi:10.1111/hel.70024
30. Kuo YC, Ko HJ, Yu LY, et al. Kill two birds with one stone? the effect of helicobacter pylori eradication in decreased prevalence of gastric cancer and colorectal cancer. Cancers. 2024;16:3881. doi:10.3390/cancers16223881
31. Chen Y, Ma W, Zhao J, et al. Lactobacillus plantarum ameliorates colorectal cancer by ameliorating the intestinal barrier through the CLA-PPAR-γ Axis. J. Agric. Food Chem. 2024;72:19766–19785. doi:10.1021/acs.jafc.4c02824
32. Toumazi D, Constantinou C. A fragile balance: the important role of the intestinal microbiota in the prevention and management of colorectal cancer. Oncology. 2020;98:593–602. doi:10.1159/000507959
33. Yu L, Wei J, Liu P. Attacking the PI3K/Akt/mTOR signaling pathway for targeted therapeutic treatment in human cancer. Semi Cancer Biol. 2022;85:69–94. doi:10.1016/j.semcancer.2021.06.019
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Anti-Inflammatory Effects and Molecular Mechanisms of Shenmai Injection in Treating Acute Pancreatitis: Network Pharmacology Analysis and Experimental Verification
He Y, Hu C, Liu S, Xu M, Liang G, Du D, Liu T, Cai F, Chen Z, Tan Q, Deng L, Xia Q
Drug Design, Development and Therapy 2022, 16:2479-2495
Published Date: 2 August 2022
A Novel Approach Based on Gut Microbiota Analysis and Network Pharmacology to Explain the Mechanisms of Action of Cichorium intybus L. Formula in the Improvement of Hyperuricemic Nephropathy in Rats

Amatjan M, Li N, He P, Zhang B, Mai X, Jiang Q, Xie H, Shao X
Drug Design, Development and Therapy 2023, 17:107-128
Published Date: 20 January 2023
Network Pharmacology and Experimental Validation to Explore That Celastrol Targeting PTEN is the Potential Mechanism of Tripterygium wilfordii (Lév.) Hutch Against IgA Nephropathy

Zhao J, Liu H, Xia M, Chen Q, Wan L, Leng B, Tang C, Chen G, Liu Y, Zhang L, Liu H
Drug Design, Development and Therapy 2023, 17:887-900
Published Date: 23 March 2023
Investigating the Mechanism of Action of Schisandra chinensis Combined with Coenzyme Q10 in the Treatment of Heart Failure Based on PI3K-AKT Pathway

Wen S, Yang K, Bai Y, Wu Y, Liu D, Wu X, Zhang X, Sun J
Drug Design, Development and Therapy 2023, 17:939-957
Published Date: 27 March 2023
Network Pharmacology Analysis of the Mechanisms Underlying the Therapeutic Effects of Yangjing Zhongyu Tang on Thin Endometrium
Zhang L, Li H, Zhang L, Zu Z, Xu D, Zhang J
Drug Design, Development and Therapy 2023, 17:1805-1818
Published Date: 17 June 2023