Back to Journals » ImmunoTargets and Therapy » Volume 14
Tumor Immunotherapy Targeting B7-H3: From Mechanisms to Clinical Applications
Authors Guo Y, Wang X, Zhang C, Chen W, Fu Y, Yu Y, Chen Y, Shao T, Zhang J ![]() , Ding G
, Ding G ![]()
Received 20 November 2024
Accepted for publication 16 March 2025
Published 27 March 2025 Volume 2025:14 Pages 291—320
DOI https://doi.org/10.2147/ITT.S507522
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Flavio Salazar-Onfray
Yining Guo,1,2 Xudong Wang,2 Chen Zhang,2 Weiwu Chen,2 Yutian Fu,2 Yanlan Yu,2 Yicheng Chen,2 Tiejuan Shao,1 Jie Zhang,2 Guoqing Ding2,3
1College of Basic Medical Sciences, Zhejiang Chinese Medical University, Hangzhou, 310053, People’s Republic of China; 2Department of Urology, Sir Run Run Shaw Hospital, School of Medicine, Zhejiang University, Hangzhou, 310016, People’s Republic of China; 3National Engineering Research Center of Innovation and Application of Minimally Invasive Instruments, Sir Run Run Shaw Hospital, School of Medicine, Zhejiang University, Hangzhou, 310028, People’s Republic of China
Correspondence: Jie Zhang, Department of Urology, Sir Run Run Shaw Hospital, School of Medicine, Zhejiang University, Hangzhou, 310016, People’s Republic of China, Email [email protected] Guoqing Ding, Department of Urology, Sir Run Run Shaw Hospital, School of Medicine, Zhejiang University, Hangzhou, 310016, People’s Republic of China, Email [email protected]
Abstract: B7-H3 (CD276) is an immune checkpoint from the B7 family of molecules and is abnormally expressed in tumor cells as a co-inhibitory molecule to promote tumor progression. Within the tumor microenvironment (TME), B7-H3 promotes tumor progression by impairing the T cell response, driving the polarization of tumor-associated macrophages (TAMs) to M2 phenotype, and inhibiting the function of other immune cells. In addition, B7-H3 promotes tumor cell proliferation, migration, invasion, metabolism disorder, angiogenesis, and resistance to treatment to promote tumor progression through its non-immunological functions. Immunotherapy targeting B7-H3, as well as combinations with other immune checkpoint therapies, have shown certain efficacy. In this review, we synthesizes the expression of B7-H3 and its mechanism to promote tumor progression through inducing immunomodulation and non-immunological functions, as well as its role of B7-H3 in tumor therapy, aiming to provide a reference for the clinical treatment of tumors.
Keywords: tumor, B7-H3 (CD276), immune checkpoint, tumor immunotherapy, tumor microenvironment
Graphical Abstract:

Introduction
The immune system has an intrinsic ability to recognize and eliminate spontaneous malignant tumors, a phenomenon that is significantly influenced by the tumor microenvironment (TME)1. T cells, as central executors of the immune system, are regulated by antigen-specific signals and also receive co-regulatory signals provided by B7 and other molecules.2 The B7 family is recognized as a group of immunomodulatory ligands. Currently known members of the B7 family include B7-1 (CD80), B7-2 (CD86), B7-DC (PD-L2), B7-H1 (PD-L1), B7-H2 (CD275), B7-H3 (CD276), B7-H4, B7-H5, B7-H6, and B7-H7 (HHLA2).3 Some B7 family members, like B7-1, B7-2, and B7-H2, bind to CD28 to provide co-stimulation, promoting T cell activation, proliferation, and differentiation.4 Others, like B7-H4 and PD-L1, bind to inhibitory receptors such as CTLA-4 and PD-1, delivering co-inhibitory signals to suppress excessive T cell activation and maintain immune balance.4 Most B7 family proteins are, widely expressed on immune cells and various cancer tissues, facilitate tumor immune evasion through their co-inhibitory effects.5
Among the B7 family members, B7-H3 has garnered significant attention due to its role in cancer progression. B7-H3, also known as CD276, was first identified as a member of the B7 family of immune checkpoint proteins in 2001.6 B7-H3 is a type I transmembrane protein featuring extracellular Ig-like domains, a transmembrane region, and an intracellular domain.7 In normal physiology, B7-H3 maintains immune balance by regulating the activity of immune cells.8 B7-H3 can also act as a T cell co-stimulatory protein in vitro and in autoimmune disease models.9 However, numerous scholars have found that B7-H3 is closely associated with many malignant tumors, and its overexpression correlates with poorer patient prognosis, such as pancreatic cancer, cutaneous melanoma, and breast cancer, among others.10–20 In recent years, new insights into the functions of B7-H3 and its roles in cancer development have been gained. As a result, immunotherapy targeting B7-H3 has emerged as a promising strategy. In this review, we summarize the advances regarding the potentially key role of B7-H3 in tumor pathogenesis and progression. The findings from these studies may provide new insights and lead to breakthroughs for tumor immunotherapy.
B7-H3 Expression and Regulation in Tumors
B7-H3 mRNA is widely expressed in both lymphoid and non-lymphoid organs.6 B7-H3 mRNA shows robust expression in many prevalent cancer types, including prostate, pancreatic, breast, colorectal, ovarian, and lung cancers.20 However, at the protein level, B7-H3 is expressed at very low levels or is virtually non-existent in normal tissues, and is more restricted to the cell surface, for instance, on activated T cells, B cells, natural killer (NK) cells, and antigen-presenting cells (APCs).8,21 B7-H3 is commonly overexpressed in human malignant tumors and is often associated with poor clinical outcomes in patients.8,22–25 Notably, high levels of B7-H3 was detected in tumors with known driver mutations (eg EGFR, ESR1, KRAS, SPOP, ERG, AR, TP53, MYC, PTEN) or undruggable therapeutic targets (TP53, APC).20,26
Despite B7-H3 ubiquitous mRNA expression, protein levels are highly regulated, indicating strict post-transcriptional and post-translational control mechanisms (Figure 1). There exists a close and complex relationship between signaling pathways and protein transcription. Aberrant activation of mTOR pathway is one of the most common dysregulations in tumors, and it is closely related to the occurrence and progression of various cancers. ILT4 and BRD4 both upregulate the expression of B7-H3 via the PI3K/AKT/mTOR signaling pathway.27,28 Recent research have provided more detailed insights, that hyperactivated mTORC1 in tumors upregulates B7-H3 expression via direct phosphorylation of the transcription factor YY2 by p70 S6 kinase.29 Non-coding RNAs also affect the transcription and expression regulation of proteins. The expression of B7-H3 protein was inversely correlated with microRNA 29 (miR-29) and miR-1253 levels in both cell lines and tumor tissues tested.30–32 The long non-coding RNA (lncRNA) TUG1 Inhibits miR-29c, leading to upregulation of B7-H3 expression in macrophages.33 In addition, the lncRNA NEAT1 sponged miR-214, thereby enhancing the expression of B7-H3 and promoting the polarization of M2 TAMs, which in turn accelerates the progression of multiple myeloma.34 LINC01123 promotes immune escape by sponging miR-214-3p to regulate B7–H3 in head and neck squamous-cell carcinoma (HNSCC).35 In addition, modifications of mRNA can affect the expression levels of proteins. In colorectal cancer, the downregulation of B7-H3 m6A modification affects its binding to specific RNA-binding proteins, stabilizes B7-H3 mRNA, and promotes its translation, ultimately facilitating immune evasion.36 Together, these various post-transcriptional mechanisms reveal the different ways in which B7-H3 expression is regulated, affecting the occurrence and development of tumors. Furthermore, post-translational modifications of B7-H3 also play a crucial role. B7-H3 is a highly glycosylated protein.37 Current evidence shows that the abnormal glycosylation of B7-H3 in tumors is of great significance for protein expression and ligand-receptor binding.38 Aberrant glycosylation of B7-H3 stabilizes the protein itself, resulting in the accumulation of B7-H3 in tumors, which can be used as an important marker of oral cancer.39 Aberrant B7-H3 glycosylation is highly expressed in triple negative breast cancer (TNBC), and N-glycosylation of B7-H3 at NXT motif sites is responsible for its protein stability and immunosuppression in TNBC tumors.40 In addition, in vivo ubiquitination and TUBE assays have revealed that B7-H3 in non-small cell lung cancer (NSCLC) cells is regulated by the ubiquitin-proteasome system, with ubiquitination occurring at both K48 and K63 sites.41 Moreover, B7-H3 protein may also be controlled by other post-translational regulatory mechanisms, which still need to be further explored, and may provide new insights for tumor immunotherapy.
 |
Figure 1 B7-H3 post-transcriptional and post-translational regulatory mechanisms. B7-H3 mRNA is regulated by the transcription factor YY2 and m6A modification, with various microRNAs and lncRNAs also impacting its expression. Glycosylation and ubiquitination of B7-H3 are also important for the stability of protein expression. Created in BioRender. Yining, G. (2025) https://BioRender.com/2cvopy1. |
The Role of B7-H3 in Tumors
The Immunomodulatory Function of B7-H3 in the TME
The TME includes a rich diversity of immune cells (such as APCs, T cells, NK cells, B cells, etc), tumor cells, the tumor stroma, and other cell types.42 Given the current limited knowledge about the ligands of B7-H3, a comprehensive explanation of its immunomodulatory mechanisms is not yet feasible.43 However, recent studies have indicated that the upregulation of B7-H3 in the TME is highly correlated with tumor immune evasion44 (Figure 2).
 |
Figure 2 Immunomodulatory function B7-H3 in the TME. B7-H3 inhibits the proliferation and function of anti-tumor T cells, including CD8+ T cells, CD4+ T cells, Th1 cells, and Th2 cells. It also promotes the differentiation of T cells into tumor-promoting Tregs, thereby facilitating tumor progression. B7-H3 induces macrophages to shift from the anti-tumor M1 phenotype to the tumor-promoting M2 phenotype. Additionally, B7-H3 inhibits the activation of NK cells. TAMs and tumor-resident DCs also express B7-H3, contributing to their immunosuppressive functions. Created in BioRender. Yining, G. (2025) https://BioRender.com/0cj2xez. |
B7-H3 Facilitates Multidimensional T Cells Suppression
The immune response mediated by T cells is of paramount importance in anti-tumor immunity.45 A variety of co-stimulatory and co-inhibitory receptors are expressed to regulate and direct T cell function.46 B7-H3 was initially characterized as a co-stimulatory molecule with antitumor capabilities. In a mouse lymphoma model, B7-H3 was shown to activate CD8+ T cells and NK cells, leading to tumor regression.47
However, accumulating evidence now highlights its predominant role as a co-inhibitory molecule in facilitating tumor progression. In melanoma, the requirement of NK and CD8+ T cells in B7-H3-mediated tumor immunity has been established. Inhibition of B7-H3 has been shown to potentially augment both the quantity and cytolytic function of tumor antigen-specific CD8+ T cells.48 Similarly, intestinal epithelial B7-H3 deletion elevates tumor-infiltrating CD8+ T cells and their interferon-gamma (IFN-γ) production in colorectal cancer.49 Furthermore, tumor-intrinsic B7-H3 suppresses CD8+ T cell proliferation and cytotoxicity within the TME, correlating inversely with CD8+ T cell infiltration and co-stimulatory molecule expression.50,51 Recent research has indicated that B7-H3 is markedly overexpressed in the vasculature system associated with brain metastases (BrM) in both mice and humans. Therapeutic intervention using B7-H3 antibodies has led to a significant increase in CD8+ T cell infiltration in BrM, thereby demonstrating the potential of B7-H3 as a therapeutic target for BrM.52 Therapeutically, tumor-expressed B7-H3 has been observed to inhibit the function of CD8+ T cells in ovarian cancer that is insensitive to PD-1 blockade therapy. This suggests that B7-H3 could represent a viable target for patients unresponsive to PD-L1/PD-1 inhibitory interventions.50 Collectively, these findings suggest that B7-H3, as expressed by tumors, may suppress CD8+ T cell function. Additionally, studies have shown that tumor cells can inhibit the activity of CD4+ T cells, particularly cytotoxic CD4+ T cells (such as CD38+CD39+CD4+ T cells), by upregulating B7-H3 expression.29 As such, B7-H3 is deemed to have considerable potential as a target for immunotherapy.
T helper (Th) cells play an important role in tumor immunity by secreting cytokines to activate and enhance the function of other immune cells. Th1 cells express IFN-γ and IL-2 as their effector cytokines to mediate cellular immunity, while Th2 cells express IL-4, IL-5, and IL-10 to mediate humoral immunity. Microarray analysis of B7-H3 peptide, B7-H3 negative and overexpressed tumor cells showed that B7-H3 inhibits the T cell proliferation, the Th1 cell response and the production of cytokines IL-2 and IFN-γ.53 Similarly, B7-H3 on breast cancer cells binds to its receptor on T cells and inhibits the release of IFN-γ and the proliferation of Th1 cells by suppressing the activation of the PI3K/AKT/mTOR signaling pathway.54 However, another study found that both Th1-type, IFN-γ and IL-2, and Th2-type, IL-4, cytokines were increased upon B7-H3 depletion.55 At present, there are few studies on how B7-H3 acts on Th cells, and the mechanism remains unclear, so it is necessary to elucidate the mechanism of B7-H3 on Th cell polarization.
B7-H3 plays a critical role in immunosuppression via interactions with regulatory T cells (Tregs). Tumor cells may promote the differentiation of T cells into Tregs by increasing the expression of B7-H3.56 There is the evidence that B7-H3 expression is positively associated with the density of tumor-infiltrating Foxp3+ Tregs, which assist tumor cells in evading immunosurveillance.57 Likewise, in the intermediate-risk/high-risk subcohort of prostate cancer, there was a significant positive correlation between Tregs density and B7-H3 mRNA expression.58 However, inhibiting B7-H3 alone caused compensatory immunosuppressive effects, including up-regulation of PD-L1 expression in myeloid cells and increased Treg infiltration into tumors.55 It emphasizes the need for combinatorial strategies to overcome Treg-mediated suppression. These findings suggest that further investigation into the relationship between B7-H3 and Tregs could lay the groundwork for developing clinical treatment strategies in order to enhancing anti-tumor immunity.
In summary, B7-H3 orchestrates a complex immunosuppressive network through interactions with CD8+ T cells, CD4+ T cells, Th cells, and Tregs (Figure 2). Although existing research has given us a preliminary understanding of the potential functions of B7-H3, further up-to-date studies and in-depth exploration of its mechanisms are still needed to enhance our comprehension.
B7-H3 Modulates Tumor-Associated Macrophage (TAMs) Polarization and Function
Macrophages play a dominant role in the TME, where M1-type macrophages exhibit antitumor properties, while M2-type macrophages exert immunosuppressive effects and promote cancer cell growth.59 In progressive tumors, M2 TAMs are a major component of the TME, and their presence within tumor tissues significantly contributes to the formation of an immunosuppressive TME.60 B7-H3 enhances the polarization of TAMs toward the pro-tumor M2 phenotype across multiple malignancies.61 In HNSCC, B7-H3 is positively correlated with immature myeloid cells and the proportion of pro-M2 TAMs.62 Studies have shown that B7-H3 induces the polarization of macrophages from the antitumor M1 phenotype to the pro-tumor M2 phenotype, leading to an increase in the proportion of pro-tumor macrophages.63 Mechanistically, B7-H3 in tumor cells drives M2 polarization through pathways such as the CCL2-CCR2 axis in ovarian cancer,64 while its genetic ablation reduces M2-TAM infiltration and enhances IFN-γ+ CD8+ T cell recruitment.64 In addition, TAMs can also lead to increased expression of B7-H3 in tumors. Studies have shown that there is a subset of cells that express both macrophage and tumor signatures simultaneously in the tumor, and this group of double-positive TAMs is formed by phagocytosis of tumor cells and mediates immunosuppression through the transformation of M1 macrophages into M2 macrophages, as well as the up-regulation of the immune checkpoint protein B7-H3.65 This suggests that high expression of B7-H3 influences the polarization of TAMs (Figure 2), but a detailed elucidation of the molecular mechanisms and signaling pathways involved still requires further investigation.
The immunomodulatory role of B7-H3 extends to TAMs functional reprogramming. Research has indicated that both human and murine tumor-associated macrophages in urothelial bladder cancer (BLCA) exhibit high B7-H3, which activates lysosomal signaling pathways and the transcription factor JUN, thereby affecting the efferocytosis of TAMs and promoting tumor immune escape.66 Similarly, in pancreatic neuroendocrine tumors (PNEN), B7-H3-expressing TAMs suppress T cell proliferation and IFN-γ production, suggesting that B7-H3 could be a potential target for immunotherapy in PNEN.67 These findings collectively position B7-H3 as a central regulator of TAM-mediated immunosuppression, though detailed molecular mechanisms warrant further elucidation.
B7-H3 Modulates Other Immune Cells
NK cells are able to recognize tumor cells through their surface receptors and directly kill them by releasing cytotoxins. B7-H3 inhibits the activation of both mouse and human NK cells48 (Figure 2). In prostate cancer models, treatment with B7-H3 inhibitors restores NK cell cytolytic function and enhances tumor infiltration.55 In like manner, the latest camelid B7-H3 tri-specific killer delivers an NK cell specific IL-15 signal to expand NK cells, facilitating the treatment of prostate cancer.68 Dendritic cells (DCs) are pivotal antigen-presenting cells bridging innate and adaptive immunity. B7-H3 is significantly upregulated in tumor-residing DCs in NSCLC, causing DCs to have immunosuppressive effects. This results in increased levels of the inhibitory cytokine IL-10 and decreased secretion of the immunostimulating cytokine IL-12, ultimately weakening the ability of DCs to stimulate T cells.69 This suggests that high expression of B7-H3 can significantly inhibit the immunostimulatory potential of DCs (Figure 2). However, the precise mechanisms through which B7-H3 exerts its effects on other immune cells remain to be fully elucidated.
The Non-Immunomodulatory Roles of B7-H3 in Tumors
Beyond its impact on the immune system, B7-H3 also exerts pro-tumorigenic effects through a variety of non-immunomodulatory mechanisms. It promotes tumor progression by stimulating tumor cell proliferation, disrupting metabolism, enhancing migration and invasion, facilitating angiogenesis, and conferring resistance to tumor treatments. B7-H3’s immunosuppressive functions weaken anti-tumor immune responses, thereby creating favorable conditions for tumor cell proliferation and metastasis. Meanwhile, its non-immune regulatory functions enhance the biological properties of tumor cells, further shaping the TME that supports tumor progression. This dual mechanism makes B7-H3 a key driver of tumor immune evasion and progression. B7-H3’s functional plasticity allows it to dynamically shift between immune regulation and tumor-intrinsic adaptation based on microenvironmental cues.
B7-H3 Drives Tumor Proliferation
Tumor cell proliferation drives tumor growth and spread, influenced by both the TME and the intrinsic properties of tumor cells. B7-H3 has been identified as a promoter of cell proliferation in various cancer types, including glioma,70 breast cancer,71 colorectal cancer,72 and many others.23 Functional studies demonstrate that silencing B7-H3 can significantly inhibit the proliferation, invasion, and migration of breast cancer, melanoma and the lung adenocarcinoma models.71,73 Mechanistically, B7-H3 activates oncogenic signaling pathways within cancer cells, such as ERK, PI3K, and STAT3, potentially driving accelerated proliferation and tumor growth.74,75 Concurrently, B7-H3 leads to the suppression of apoptotic pathways, a significant reduction in cell cycle markers CDK4, CDK6, and cyclin-D1, and increased proliferation marker phosphorylated AKT, resulting in cell cycle arrest.32 Notably, B7-H3 expression on circulating epithelial tumor cells (CETCs) is positively correlated with the expression of the proliferative marker Ki-67 in breast cancer.13 Additionally, the stemness of tumor stem cells significantly drives their proliferation. Compared to regular cancer cells, B7-H3 is overexpressed in the stem cell population. B7-H3 enhances the interaction between B-RAF and MEK by increasing Major Vault Protein (MVP), thereby activating MEK and inducing stem cell proliferation.76 These findings suggest that B7-H3 plays a multifaceted role in tumor progression, not only through its immunosuppressive functions but also by directly promoting tumor cell proliferation, highlighting its potential as a target for a broad range of cancer therapies (Figure 3A).
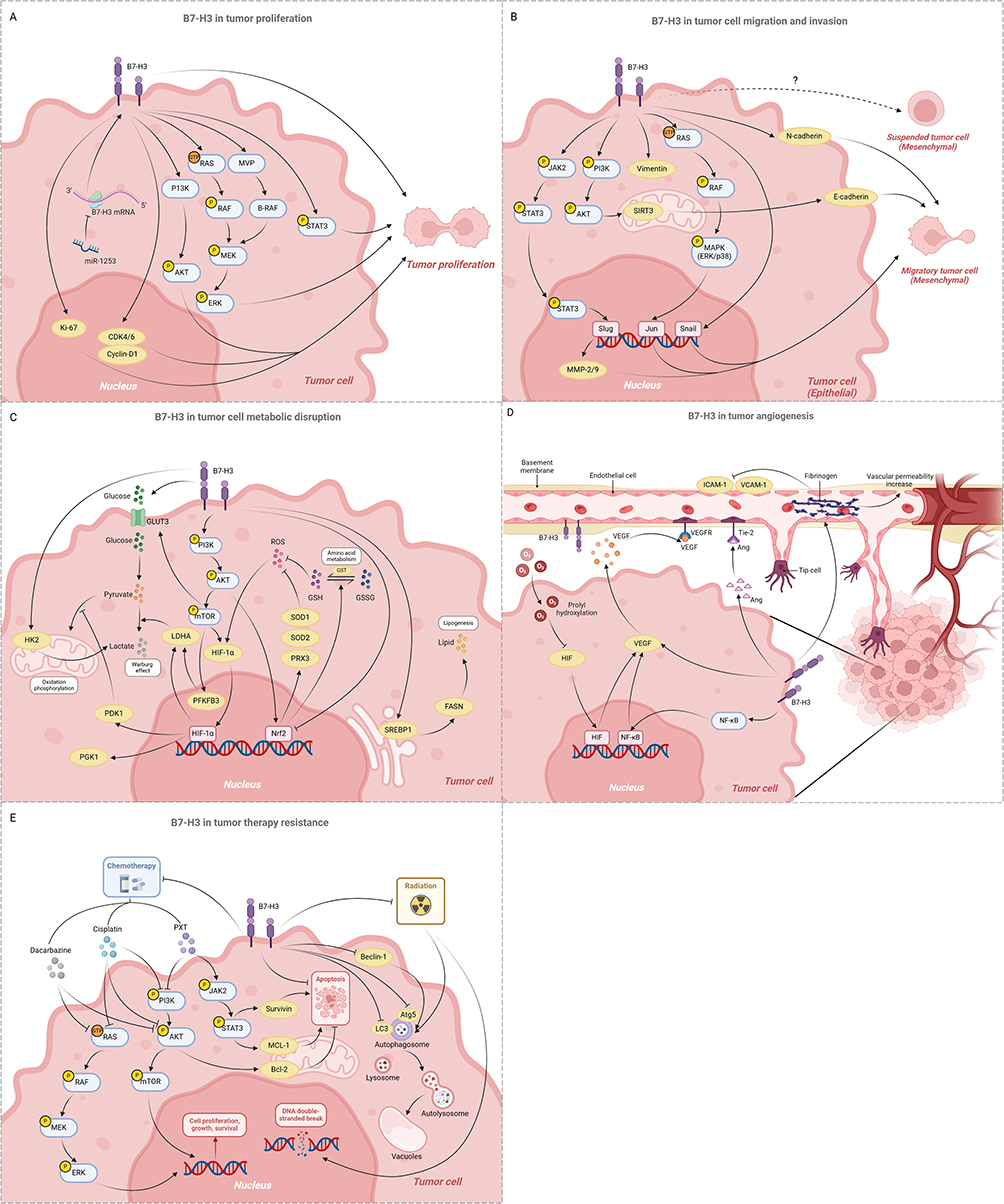 |
Figure 3 The non-immunomodulatory roles of B7-H3 in tumors. (A) B7-H3 drives tumor proliferation by activating oncogenic signaling pathways within tumor cells (eg, ERK, PI3K, and STAT3), inhibiting apoptosis pathways (eg, reducing cell cycle markers such as CDK4, CDK6, and cyclin-D1), and enhancing the stemness of tumor stem cells (B7-H3 upregulates MVP, activates MEK, and induces stem cell proliferation) Created in BioRender. Yining, G. (2025) https://BioRender.com/lps362n. (B) B7-H3 promotes epithelial-mesenchymal transition (EMT) by activating various signaling pathways (including JAK2/STAT3, PI3K/AKT, p38/ERK signaling pathways, and downstream EMT-related proteins), thereby facilitating tumor cell migration and invasion Created in BioRender. Yining, G. (2025) https://BioRender.com/ qqmks36. (C) B7-H3 causes metabolic disruption of tumor cells by driving aberrant glycolysis, lipid metabolism, and amino acid metabolismCreated in BioRender. Yining, G. (2025) https://BioRender.com/s9axlvi. (D) B7-H3 stimulates tumor angiogenesis by stimulating the secretion of VEGF, activating the Ang/Tie-2 pathway, and altering vascular structure at the molecular level Created in BioRender. Yining, G. (2025) https://BioRender.com/x16n3aq. (E) B7-H3 enhances tumor resistance to chemotherapy and radiotherapy by upregulating anti-apoptotic proteins, downregulating autophagy-related proteins, and inhibiting the repair of DNA double-strand breaks. Created in BioRender. Yining, G. (2025) https://BioRender.com/95tk9tm. |
B7-H3 Promotes Tumor Cell Migration and Invasion
The migration and invasion of tumor cells are significant indicators of cancer progression, often associated with poor prognosis and high mortality rates. Clinically, B7-H3 has been confirmed to promote tumor cell migration and invasion across various cancer types, commonly linked to adverse outcomes.72,77 Epithelial-mesenchymal transition (EMT) endows cells with the ability to invade and metastasize, representing a crucial step in tumor metastasis. In glioma cells with overexpression of B7-H3, the JAK2/STAT3 signaling pathway is activated, inducing glioma invasion and participating in EMT through the JAK2/STAT3/Slug/MMP-2/-9 pathway.78 Similarly, in hepatocellular carcinoma, B7-H3 has been shown to promote EMT via the JAK2/STAT3/Slug signaling axis.79 Evidence suggests that silencing B7-H3 can downregulate the expression of EMT marker proteins, such as MMP-2, vimentin, Snail, and N-cadherin, attenuating the migratory and invasive capabilities of lung adenocarcinoma cells.73 Furthermore, B7-H3 can upregulate the expression of Sirtuins protein 3 (SIRT3) in NSCLC cell lines through the PI3K/AKT pathway, thereby enhancing the expression of E-cadherin and endowing tumor cells with stronger migratory and invasive abilities, thus facilitating EMT.78 In clear cell renal cell carcinoma (CCRCC), B7-H3 promotes the EMT process by activating the fibronectin-mediated PI3K/AKT and p38/ERK mitogen-activated protein kinase (MAPK) signaling pathways.80 Despite differences in the mechanisms, these studies collectively highlight the significant impact of B7-H3 on tumor invasion and metastasis, demonstrating its potential as a therapeutic target for disrupting these processes in cancer (Figure 3B). Additionally, a novel phenotype has been identified in the migration and invasion of tumor cells: the adherent-to-suspension transition (AST). This process involves the reprogramming of adherent cells into suspension cells by specific hematopoietic transcription factors, thereby endowing tumor cells with enhanced invasiveness and metastatic potential.81 To date, research focusing on AST is quite limited, and studies exploring the relationship between B7-H3 and AST have not yet been reported. This represents a promising avenue for future investigation and may offer new perspectives for the treatment of metastatic tumors.
B7-H3 Causes Metabolic Disruption of Tumor Cells
Abnormal tumor metabolism, including aberrations in glycolysis and lipid metabolism, plays a critical role in tumor occurrence, development, and metastasis.82 Both in vivo and in vitro studies have confirmed that overexpression of B7-H3 promote glucose intake and lactate production, driving aberrant glycolysis in tumors.83 B7-H3 enhances the Warburg effect by upregulating the protein levels of hypoxia-inducible factor 1α (HIF-1α) and its downstream glycolytic enzymes, including lactate dehydrogenase (LDHA), phosphoglycerate kinase 1 (PGK1), and Pyruvate dehydrogenase kinase 1 (PDK1).83,84 Mechanistically, B7-H3 promotes the reactive oxygen activity-dependent stability of HIF-1α by inhibiting the activity of stress-activated transcription factor Nrf2 and its target genes, including antioxidant enzymes such as SOD1, SOD2, and PRX3, further amplifying glycolysis and tumor progression.83 In oral squamous carcinoma, B7-H3 activates the PI3K/AKT/mTOR to upregulate the expression of downstream HIF-1α, glucose transporter 3 (GLUT3), and phosphofructokinase-2/fructose-2,6-bisphosphatase 3 (PFKFB3), allowing tumor cells to generate energy through glycolysis even in an oxygen-rich environment.85 Additionally, hexokinase 2 (HK2) is a key mediator in the regulation of glucose metabolism. B7-H3 increases glucose consumption and lactate production by promoting HK2 expression in colorectal cancer cells, thereby enhancing cell migration, invasion, and apoptotic resistance.86 HK2 inhibition reverses B7-H3-driven aerobic glycolysis, confirming its functional role.87 Furthermore, B7-H3 regulates lipid metabolism in tumor cells, which is one of the key metabolic alterations in tumor progression. B7-H3 increases lipogenesis through the SREBP1/FASN signaling pathway, thereby promoting the occurrence and development of lung cancer.88 Additionally, B7-H3 enhances the stemness of gastric cancer (GC) cells and promotes tumorigenesis and progression by activating the expression of Nrf2 through the AKT pathway, thereby modulating glutathione (GSH) metabolism in GC cells.89 However, the relationship between B7-H3 and other amino acid metabolism disorders in tumors remains unexplored, highlighting a critical research gap. In summary, these studies consistently demonstrate that B7-H3 may favor tumor growth by manipulating tumor immunosurveillance through alterations in the metabolism of glucose, lipids, and amino acids in tumor and immune cells (Figure 3C). Further studies should explore its crosstalk with immunometabolic pathways.
B7-H3 Stimulates Tumor Angiogenesis
Angiogenesis, the process of generating new blood vessels from pre-existing ones, is one of the hallmarks of cancer and plays a complex role in the development of solid tumors.90 Hypoxia in the TME triggers HIF activation, which drives angiogenesis through transcriptional upregulation of vascular endothelial growth factor (VEGF).91,92 Studies have shown that B7-H3 can promote angiogenesis by stimulating the secretion of VEGF.93 In colorectal cancer, B7-H3 upregulates VEGF expression through the activation of the NF-κB pathway, thereby promoting tumor angiogenesis; conversely, downregulation of B7-H3 significantly inhibits the in vitro tube formation of human umbilical vein endothelial cells (HUVECs).92 Angiogenin (Ang)/Tie-2 (Tek) represents another pathway in tumor angiogenesis, and the expression levels of B7-H3 and Tie-2 are positively correlated, with immunofluorescence staining confirming the co-expression of B7-H3 and Tie-2 in the vascular endothelium of CCRCC.94 Therefore, B7-H3 may promote the angiogenesis of CCRCC through the Tie-2 pathway, offering a potential therapeutic strategy for targeting angiogenesis in the context of cancer. Among various primary BrM, the expression of B7-H3 on tumor vasculature is significantly increased.52 Blocking B7-H3 leads to a reduction in extravascular fibrinogen, thereby reducing the permeability of the blood-brain barrier, as well as an increase in cellular adhesion molecules ICAM-1 and VCAM-1, which alters the structure of the blood vessels at the molecular level, ultimately suppressing BrM growth.52 Targeting of B7-H3 could inhibit neoangiogenesis and tumor growth95 (Figure 3D). Furthermore, considering the complexity of angiogenesis, future research should take into account the interactions between B7-H3 and other angiogenic factors to gain a more comprehensive perspective.
B7-H3 Contributes to Tumor Therapy Resistance
Increased expression of B7-H3 in tumors is associated with the development of therapeutic resistance.71 B7-H3 knockdown sensitizes melanoma cells to MAPK and AKT/mTOR pathways-targeted agents (dacarbazine, cisplatin), reversing chemoresistance.96,97 Paclitaxel (PXT) is a commonly used chemotherapeutic drug in clinical practice. However, studies have found that in breast cancer, B7-H3 overexpression drives PXT resistance via Jak2/Stat3 signaling pathway activation, upregulating anti-apoptotic proteins.98 Targeting B7-H3 can enhance the sensitivity of breast cancer to paclitaxel, a finding of significant importance for developing treatment strategies for paclitaxel-refractory cancers. Similarly, B7-H3 activates the PI3K/AKT signaling pathway and upregulates the protein levels of B-cell lymphoma 2 (BCL-2) in ovarian cancer cell lines, resulting in chemoresistance and sustained tumor growth.99 Radiotherapy, which damages the DNA structure of cancer cells leading to apoptosis, is another standard treatment modality. B7-H3 downregulates the expression of autophagy-related proteins such as light chain 3 (LC3), autophagy-related protein 5 (Atg5), and Beclin-1, thereby enhancing the resistance of cancer cells to radiotherapy through the suppression of cellular autophagy.100 Moreover, B7-H3 can also increase the resistance of gastric cancer cells to radiotherapy by inhibiting cellular apoptosis and DNA double-strand break repair.100 However, no studies have yet elucidated the specific mechanisms by which B7-H3 affects these autophagy-related proteins. Targeted approaches, such as CRISPR screening or co-immunoprecipitation assays, are urgently needed to verify these potential interactions. Overall, these findings suggest that B7-H3 decreases the sensitivity of tumor cells to a range of chemotherapeutic drugs and radiotherapy, making it a novel target for enhancing the effectiveness of traditional cancer treatments (Figure 3E). To fully understand the impact of B7-H3 and optimize its application in cancer treatment, it is necessary to further elucidate its mechanisms of action and expand research on the role of B7-H3 in different treatment modalities.
Current Status of Immunotherapy Strategies Targeting B7-H3
B7-H3’s multifaceted role in tumor biology has propelled it as a prime therapeutic target. Current immunotherapeutic agents targeting B7-H3 encompass monoclonal antibodies (mAbs), bispecific antibodies (BsAbs), antibody-drug conjugates (ADCs), chimeric antigen receptor T cells (CAR-T), and other modalities. A number of anti-B7-H3 approaches have been studied in preclinical or clinical trials, with details of the clinical trials presented in Table 1.
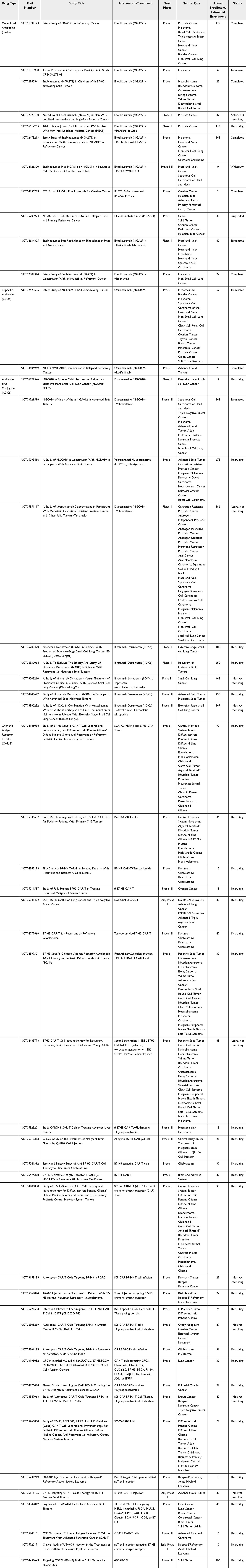 |
Table 1 Trials of Immunotherapeutic Agents Targeting B7-H3 |
Enoblituzumab (MGA271), a humanized mAb with engineered Fc domains to enhance its antitumor function by increasing binding to the activating receptor, CD16A, and reducing that to the inhibitory receptor, CD32B, was the first mAb tested against B7-H3-expressing tumors.101 It is used to treat B7-H3-expressing refractory tumors, such as melanoma (NCT01391143), and other B7-H3-expressing tumors, including osteosarcoma and Ewing’s sarcoma (NCT02982941).102 In localized prostate cancer (NCT02923180), treatment with enoblituzumab was found to be relatively safe, exhibiting no off-target antibodies or antigen spread. Importantly, after treatment with enoblituzumab, significant activation of the immune system was observed in the TME, involving both T cells and myeloid cells.103 Another study evaluated the preclinical in vitro efficacy of vobramitamab duocarmazine, an investigational ADC featuring a humanized B7-H3 mAb, demonstrating good in vitro activity in the treatment of acute myeloid leukemia.19 A study on the Fc optimization (SDIE modification) of the mAb of B7-H3 for acute myeloid leukemia has shown it can significantly induce NK cell activation and effectively inhibit leukemia growth in vivo, without causing off-target immune activation.104 Unfortunately, patient heterogeneity limits universal efficacy. A recent study found that ITGB6 is a key gene mediating differential response to anti-B7-H3 treatment and can be used as a predictive biomarker of anti-B7-H3 immunotherapy response.105 The role of B7-H3 monoclonal antibodies in targeting advanced metastatic or recurrent diseases remains to be explored.
BsAbs, also known as bispecific T cell engagers (BiTEs), are artificially synthesized proteins that can recognize two different antigens, and simultaneously bind to antigens on tumour cells and the CD3 subunits on T cells.106 Obrindatamab (MGD009) is generated by linking the anti-CD3 mAb scFv to the anti-B7-H3 mAb single-chain variable fragment (scFv), enabling dual binding to CD3 and B7-H3 and redirecting T cells to kill B7-H3-expressing tumor cells.107 In preclinical models, obrindatamab enhances T cell cytotoxicity by promoting tumor-dependent secretion of IFN-γ, TNF-α, and IL-2; in immunodeficient mice, tumor growth is significantly reduced, and survival rates are increased.108 These promising results have led to a Phase I open-label dose-escalation study for refractory B7-H3-expressing tumors (NCT03406949).102 Studies have fabricated MMP-2-sensitive anti-B7-H3×CD3 bispecific antibody (biAb)/dEGCG-based nanoparticles to improve glioblastoma therapy by enhancing the ferroptotic effect and reversing immune inhibition.109 Additionally, numerous clinical trials are currently evaluating the therapeutic efficacy of B7-H3 inhibitors in combination with other checkpoint inhibitors.110,111
ADCs combine the specificity of monoclonal antibodies with the potency of highly cytotoxic agents, selectively and stably delivering potent anticancer agents to tumor cells.112,113 Duocarmazine (MGC018) is a duocarmycin-based ADC targeting B7-H3.114 MGC018 specifically binds to B7-H3, delivering the potent DNA alkylating agent duocarmycin to tumor cells, inducing DNA damage and cell death, thereby providing a more potent antitumor effect than traditional mAbs.114 Importantly, the “bystander effect”—whereby released payloads diffuse to neighboring B7-H3-negative cells—may enhance efficacy in heterogeneous tumors.114 A Phase I dose expansion study is currently underway for NSCLC, triple-negative breast cancer (TNBC), and metastatic castration-resistant prostate cancer (mCRPC), as indicated by the clinical trial registration NCT03729596.114 MGC018 has shown preliminary clinical activity in B7-H3-expressing preclinical models of neuroblastoma.115 Moreover, the clinical trials of Ifinatamab deruxtecan (I-DXd, DS-7300a) have also had a broad impact. This ADC targets B7-H3 and is Non-coding RNAs also affect composed of a humanized anti-B7-H3 monoclonal antibody linked to a topoisomerase I inhibitor payload (DXd). The ADC has demonstrated significant antitumor activity and good safety in clinical trials116 and has been granted orphan drug designation by the US FDA and the European Commission for the treatment of SCLC. The ongoing Phase III IDeate-Lung02 trial will further validate its potential in first-line treatment.
Traditional therapies often lack specificity for tumor cells, potentially causing severe side effects. In contrast, B7-H3-targeted CAR-T cells can specifically recognize and kill tumor cells expressing B7-H3, reducing damage to normal cells.117 B7-H3-targeted CAR-T therapy has demonstrated significant potential in a series of preclinical model studies across various cancer types,118–121 and clinical trials have confirmed its effectiveness. A Phase I clinical study of B7-H3 CAR-T therapy for diffuse intrinsic pontine glioma (NCT04185038) showed sustained improvement in clinical and radiographic symptoms in some patients, with repeated intracranial injections of CAR-T cells inducing local immune activation.122 In vitro co-culture models and mouse xenograft models of acute myeloid leukemia have shown that B7-H3 CAR-T therapy is effective, exhibiting antigen-dependent toxicity with acceptable side effects.123 Recently, iCas9.B7-H3 CAR-T have effectively killed uveal melanoma cells in vitro and eradicated liver metastasis of uveal melanoma in mouse model, outperforming B7-H3 mAbs.124 This method embeds a caspase-9 (iCas9) suicide gene, regulated by the chemical inducer AP1903, into B7-H3 CAR-T cells.124 This allows for rapid and conditional elimination of iC9.B7-H3 CAR-T cells via AP1903 administration in case of adverse effects, functioning as a “safety on/off switch”. The findings support a Phase I clinical trial of iCas9.B7-H3 CAR-T cells to treat patients with metastatic uveal melanoma. However, CAR-T monotherapy may not achieve high cure rates for patients with solid tumors that are difficult to treat. This is possibly due to insufficient target antigen expression on the tumor surface, which impairs CAR-T cell recognition and attack, and the complex TME, where inhibitory cells and factors suppress CAR-T cell function and reduce their efficacy in solid tumors. Consequently, some scholars have used a novel high-content and high-throughput screening method to identify that ingenol-3-angelate enhances the activity of B7-H3 CAR-T cells by activating PKCα to upregulate B7-H3 on the target cell surface.125 In addition, an increasing number of studies have focused on modifying B7-H3 CAR-T cells to improve their therapeutic efficacy, fostering the development of novel combination strategies for CAR-T immunotherapy.126–131 Collectively, these studies demonstrate that B7-H3 CAR-T represents a promising therapeutic strategy for targeting human tumors.
In recent years, the number of trials evaluating B7-H3 targeted therapy has increased significantly with the accumulation of experimental evidence. These studies not only enhance our understanding of B7-H3’s role in tumorigenesis but also provide insights into the potential therapeutic benefits of targeting this molecule, offering new alternatives and clinical benefits for tumor patients. However, the majority of B7-H3 targeted clinical trials are currently in Phase I/II, and the efficacy data that have been published are limited. Early results have primarily focused on safety and maximum tolerated dose, while biomarker-driven mechanistic data, such as the dynamics of immune cell changes, are rarely reported. Therefore, the unique expression patterns of B7-H3 in tumors make it a potential breakthrough target. Future research needs to conduct more in-depth mechanistic analyses. Targeting B7-H3 represents a new and feasible option for tumor treatment in the future.
Conclusions and Outlook
B7-H3, an emerging immune checkpoint molecule in the B7 family, plays a multifaceted role in cancer, encompassing a range of immunological and non-immunological functions. B7-H3 has a dual role in regulating adaptive and innate immune cells, underscoring its potential as a therapeutic target (Figure 2). At the same time, it is also involved in several biological processes central to tumor pathogenesis, including proliferation, migration, invasion, angiogenesis, and metabolism (Figure 3). Spanning from immunomodulatory to non-immunomodulatory functions, the pleiotropic characteristics of B7-H3 suggest extensive application potential in the therapeutic strategies against cancer. The expression of B7-H3 is typically associated with a poor prognosis and can be used alone or in combination with other molecules as a prognostic marker. Furthermore, the differential expression of B7-H3 across various tumor cell types suggests its potential significance in tumor heterogeneity. Although B7-H3 is predominantly identified as an inhibitory modulator in tumors, its functions may vary across different types of neoplasms. Therefore, caution should be exercised in interpreting the conclusions when translating the findings to other models.
Despite the expanding research on B7-H3, critical knowledge gaps persist in B7-H3 biology. The foremost issue is the unclear identification of the receptors and intracellular binding partners of B7-H3 in both immune and tumor cells, which hinders more profound investigations in this domain. Utilizing AI-driven protein interaction prediction tools, such as AlphaFold-Multimer, ESM-IF1, and DeepMSA2, can significantly accelerate the identification process of B7-H3 receptors and further enhance the accuracy and reliability of predictions. Secondly, post-transcriptional and post-translational modifications of B7-H3 play an exceedingly important role in its functional mechanisms, particularly in the context of tumorigenesis. Notably, there is a significant gap in research on the post-translational modifications of B7-H3, which warrants further exploration, including studies on phosphorylation, acetylation, methylation, glycosylation, ubiquitination, and palmitoylation. Further study of the crosstalk between post-translational modifications will also be of great value. These investigations aim to broaden our in-depth comprehension of B7-H3, thereby aiding in the refinement of B7-H3-targeted therapeutic strategies and facilitating their clinical deployment. Moreover, several emerging technologies have offered unprecedented opportunities for biological research of B7-H3, such as single-cell multi-omics sequencing,132 spatial functional genomics,133 the combined application of single-cell and spatial genomics,134 CRISPR-based screens (iScreen,135 StAR136), and novel sequencing platforms (NIS-Seq,137 UDA-seq138). Future studies can leverage these new technologies to further explore B7-H3’s upstream regulatory mechanisms, downstream effector pathways, and its functions in normal physiology, thereby achieving a more comprehensive understanding of its mechanisms of action in tumor development and progression. Elucidating these mechanisms is essential for development of innovative immunotherapeutic strategies directed against B7-H3.
Therapeutic innovation is accelerating. Overexpression of B7-H3 in many tumors is associated with regulation by various non-coding RNAs, and targeting these naturally occurring non-coding RNAs shows promise in disrupting B7-H3 signaling. Targeting the pro-tumorigenic miRNAs, the introduction of synthetic anti-miRs that are complementary to the miRNAs into tumor cells may effectively prevent the binding of miRNAs to the mRNA of B7-H3, thereby inhibiting the progression of tumorigenesis.139 Moreover, recent artificial lncRNA technology can precisely target and efficiently degrade proteins, effectively addressing the long-standing “undruggable” target issues that have plagued the scientific community.140 Additionally, given the dual nature of B7-H3’s immune and non-immune functions, simultaneous blocking of its immune checkpoint activity (such as antagonistic antibodies) and intrinsic tumor survival pathways (such as small molecule inhibitors disrupting B7-H3-mTOR interactions) may be required. And, given the complex role of B7-H3 in the TME, future research should explore the interactions between B7-H3 and other immune checkpoint molecules, as well as their potential synergistic effects on tumor immune evasion. Therapeutic strategies targeting B7-H3, including mAbs, BsAbs, ADCs, CAR-T therapies, and combinatorial strategies, are rapidly advancing and achieving significant breakthroughs. The development of monoclonal antibodies targeting the B7-H3 glycosylation site, as has been done with PD-1, may be a promising therapeutic approach. Additionally, inhibiting the glycosylation of B7-H3 may enhance the cytotoxic effects of CAR-T cells, thereby improving their anticancer activity in solid tumors.141 And, local delivery strategies for B7-H3 CAR-T cells combined with oncolytic viruses are also a viable option.142 Furthermore, high-throughput and virtual screening of compound libraries143,144 can be used to explore the combination of B7-H3 with other therapeutic modalities, such as small-molecule inhibitors and chemotherapy, which may provide new treatment strategies for refractory tumors. Treatment strategies with B7-H3 as a target have shown promising efficacy and safety in the treatment of various tumor cells, suggesting that targeting B7-H3 could be a valuable strategy for cancer treatment. In terms of clinical trials, a broader evaluation of the safety and efficacy of B7-H3-targeted therapies is warranted. Notably, the clinical application of B7-H3 ADCs and CAR-T cell therapies across various types of cancer will be a focal point of future research. Ultimately, in-depth research on B7-H3 is poised to yield groundbreaking advancements in oncology. Integrating mechanistic insights with cutting-edge technologies will enable the development of more personalized and effective therapeutic regimens for patients.
Acknowledgment
This work was supported by grants from the National Natural Science Foundation of China (32000799 to J.Z.) and Key Research and Development Program of Zhejiang Province (2025C02058 to G.D.). We apologize to those whose work was not cited because of space limitations.
Author Contributions
All authors contributed to data collecting, drafting or revising the article, gave final approval of the version to be published, and agree to be accountable for all aspects of the work.
Disclosure
The authors declare no competing interests in this work. Graphical abstract Created in BioRender. Yining, G. (2025) https://BioRender.com/cny2i40.
References
1. Huang W, Jiang Y, Xiong W, et al. Noninvasive imaging of the tumor immune microenvironment correlates with response to immunotherapy in gastric cancer. Nat Commun. 2022;13(1):5095. doi:10.1038/s41467-022-32816-w
2. Tan S, Day D, Nicholls SJ, Segelov E. Immune checkpoint inhibitor therapy in oncology: current uses and future directions: JACC: cardioOncology state-of-the-art review. JACC. 2022;4(5):579–597. doi:10.1016/j.jaccao.2022.09.004
3. Edner NM, Carlesso G, Rush JS, Walker LSK. Targeting co-stimulatory molecules in autoimmune disease. Nat Rev Drug Discov. 2020;19(12):860–883. doi:10.1038/s41573-020-0081-9
4. Burke KP, Chaudhri A, Freeman GJ, Sharpe AH. The B7:CD28 family and friends: unraveling coinhibitory interactions. Immunity. 2024;57(2):223–244. doi:10.1016/j.immuni.2024.01.013
5. Andrews LP, Yano H, Vignali DAA. Inhibitory receptors and ligands beyond PD-1, PD-L1 and CTLA-4: breakthroughs or backups. Nat Immunol. 2019;20(11):1425–1434. doi:10.1038/s41590-019-0512-0
6. Chapoval AI, Ni J, Lau JS, et al. B7-H3: a costimulatory molecule for T cell activation and IFN-γ production. Nat Immunol. 2001;2(3):269–274. doi:10.1038/85339
7. Getu AA, Tigabu A, Zhou M, Lu J, Ø F, Tan M. New frontiers in immune checkpoint B7-H3 (CD276) research and drug development. Mol Cancer. 2023;22(1):43. doi:10.1186/s12943-023-01751-9
8. Picarda E, Ohaegbulam KC, Zang X. Molecular pathways: targeting B7-H3 (CD276) for human cancer immunotherapy. Clin Cancer Res. 2016;22(14):3425–3431. doi:10.1158/1078-0432.CCR-15-2428
9. Luo L, Zhu G, Xu H, et al. B7-H3 promotes pathogenesis of autoimmune disease and inflammation by regulating the activity of different T cell subsets. PLoS One. 2015;10(6):e0130126. doi:10.1371/journal.pone.0130126
10. Loos M, Hedderich DM, Ottenhausen M, et al. Expression of the costimulatory molecule B7-H3 is associated with prolonged survival in human pancreatic cancer. BMC Cancer. 2009;9(1):463. doi:10.1186/1471-2407-9-463
11. Yamato I, Sho M, Nomi T, et al. Clinical importance of B7-H3 expression in human pancreatic cancer. Br J Cancer. 2009;101(10):1709–1716. doi:10.1038/sj.bjc.6605375
12. Wang J, Chong KK, Nakamura Y, et al. B7-H3 associated with tumor progression and epigenetic regulatory activity in cutaneous melanoma. J Invest Dermatol. 2013;133(8):2050–2058. doi:10.1038/jid.2013.114
13. Pizon M, Schott D, Pachmann U, Pachmann K. B7-H3 (CD276) on circulating epithelial tumor cells (CETCs) correlates with proliferation marker Ki-67 and may be associated with aggressiveness of tumor in breast cancer patients. Ann Oncol. 2017; 28:v580. doi:10.1093/annonc/mdx390.020
14. Maruki Y, Takashima A, Miyamoto T, et al. Expression of B7-H3 (CD276) in surgically resected esophageal squamous cell carcinoma. JCO. 2018;36(4_suppl):70. doi:10.1200/JCO.2018.36.4_suppl.70
15. Inamura K, Takazawa Y, Inoue Y, et al. Tumor B7-H3 (CD276) expression and survival in pancreatic cancer. J Clin Med. 2018;7(7):172. doi:10.3390/jcm7070172
16. Wang Y, Deng J, Wang L, et al. Expression and clinical significance of PD-L1, B7-H3, B7-H4 and Vista in craniopharyngioma. J Immunother Cancer. 2020;8(2):e000406. doi:10.1136/jitc-2019-000406
17. Wang LC, Wang YL, He B, et al. Expression and clinical significance of Vista, B7-H3, and PD-L1 in glioma. Clin Immunol. 2022;245:109178. doi:10.1016/j.clim.2022.109178
18. Shenderov E, De Marzo AM, Lotan TL, et al. Targeting B7-H3 in prostate cancer: Phase 2 trial in localized prostate cancer using the anti-B7-H3 antibody enoblituzumab, with biomarker correlatives. JCO. 2022;40(16_suppl):5015. doi:10.1200/JCO.2022.40.16_suppl.5015
19. Kirkey DC, Blankenfeld M, Hylkema T, et al. CD276 (B7-H3) is an immunotherapeutic target in acute myeloid leukemia with preclinical efficacy of vobramitamab duocarmazine, an investigational CD276 antibody-drug conjugate. Blood. 2023;142(Supplement 1):5958. doi:10.1182/blood-2023-187450
20. Miller CD, Lozada JR, Zorko NA, et al. Pan-cancer interrogation of B7-H3 (CD276) as an actionable therapeutic target across human malignancies. Canc Res Commun. 2024;4(5):1369–79doi:10.1158/2767-9764.CRC-23-0546
21. Wang L, Kang FB, Shan BE. B7-H3-mediated tumor immunology: friend or foe? Int J Cancer. 2014;134(12):2764–2771. doi:10.1002/ijc.28474
22. Zang X, Sullivan PS, Soslow RA, et al. Tumor associated endothelial expression of B7-H3 predicts survival in ovarian carcinomas. Mod Pathol. 2010;23(8):1104–1112. doi:10.1038/modpathol.2010.95
23. Flem-Karlsen K, Fodstad Ø, Tan M, Nunes-Xavier CE. B7-H3 in cancer - beyond immune regulation. Trends Cancer. 2018;4(6):401–404. doi:10.1016/j.trecan.2018.03.010
24. Dong P, Xiong Y, Yue J, Hanley SJB, Watari H. B7H3 as a promoter of metastasis and promising therapeutic target. Front Oncol. 2018;8:264. doi:10.3389/fonc.2018.00264
25. Zhang H, Zhu M, Zhao A, Shi T, Xi Q. B7-H3 regulates anti-tumor immunity and promotes tumor development in colorectal cancer. Biochimica et Biophysica Acta. 2024;1879(1):189031. doi:10.1016/j.bbcan.2023.189031
26. Guo C, Figueiredo I, Gurel B, et al. B7-H3 as a therapeutic target in advanced prostate cancer. Europ Urol. 2023;83(3):224–238. doi:10.1016/j.eururo.2022.09.004
27. Zhang P, Yu S, Li H, et al. ILT4 drives B7-H3 expression via PI3K/AKT/mTOR signalling and ILT4/B7-H3 co-expression correlates with poor prognosis in non-small cell lung cancer. FEBS Lett. 2015;589(17):2248–2256. doi:10.1016/j.febslet.2015.06.037
28. Zhao J, Meng Z, Xie C, et al. B7-H3 is regulated by BRD4 and promotes TLR4 expression in pancreatic ductal adenocarcinoma. Int J Biochem Cell Biol. 2019;108:84–91. doi:10.1016/j.biocel.2019.01.011
29. Liu HJ, Du H, Khabibullin D, et al. mTORC1 upregulates B7-H3/CD276 to inhibit antitumor T cells and drive tumor immune evasion. Nat Commun. 2023;14(1):1214. doi:10.1038/s41467-023-36881-7
30. Xu H, Cheung IY, Guo HF, Cheung NKV. MicroRNA miR-29 modulates expression of immunoinhibitory molecule B7-H3: potential implications for immune based therapy of human solid tumors. Cancer Res. 2009;69(15):6275–6281. doi:10.1158/0008-5472.CAN-08-4517
31. Nygren MK, Tekle C, Ingebrigtsen VA, et al. Identifying microRNAs regulating B7-H3 in breast cancer: the clinical impact of microRNA-29c. Br J Cancer. 2014;110(8):2072–2080. doi:10.1038/bjc.2014.113
32. Kanchan RK, Perumal N, Atri P, et al. MiR-1253 exerts tumor-suppressive effects in medulloblastoma via inhibition of CDK6 and CD276 (B7-H3). Brain Pathol. 2020;30(4):732–745. doi:10.1111/bpa.12829
33. Sun H, Wang T, Zhang W, et al. LncRNATUG1 facilitates Th2 cell differentiation by targeting the miR-29c/B7-H3 axis on macrophages. Front Immunol. 2021;12:631450. doi:10.3389/fimmu.2021.631450
34. Gao Y, Fang P, Li WJ, et al. LncRNA NEAT1 sponges miR-214 to regulate M2 macrophage polarization by regulation of B7-H3 in multiple myeloma. Mol Immunol. 2020;117:20–28. doi:10.1016/j.molimm.2019.10.026
35. Li H, Yang Z, Yang X, et al. LINC01123 promotes immune escape by sponging miR-214-3p to regulate B7–H3 in head and neck squamous-cell carcinoma. Cell Death Dis. 2022;13(2):1–11. doi:10.1038/s41419-022-04542-0
36. Zhou Y, Zhou H, Shi J, et al. Decreased m6A modification of CD34/CD276(B7-H3) leads to immune escape in colon cancer. Front Cell Dev Biol. 2021;9:715674. doi:10.3389/fcell.2021.715674
37. Xiao L, Guan X, Xiang M, et al. B7 family protein glycosylation: promising novel targets in tumor treatment. Front Immunol. 2022;13:1088560. doi:10.3389/fimmu.2022.1088560
38. Li F, Han D, Chen M, et al. A4GALT mediated the glycosylation of b7-h3 in human colorectal cancer cell lines.
39. Chen JT, Chen CH, Ku KL, et al. Glycoprotein B7-H3 overexpression and aberrant glycosylation in oral cancer and immune response. Proc Natl Acad Sci. 2015;112(42):13057–13062. doi:10.1073/pnas.1516991112
40. Huang Y, Zhang HL, Li ZL, et al. FUT8-mediated aberrant N-glycosylation of B7H3 suppresses the immune response in triple-negative breast cancer. Nat Commun. 2021;12(1):2672. doi:10.1038/s41467-021-22618-x
41. Kurz M, Rieger L, Giansanti P, Kuster B, Bassermann F. P01.05 deciphering the function of the ubiquitin-proteasome-system in regulating the immune checkpoint protein B7-H3 (CD276) in non-small cell lung cancer. J ImmunoTher Cancer. 2021; 9:A5.1–A5. doi:10.1136/jitc-2021-ITOC8.9
42. de VKE, Joyce JA. The evolving tumor microenvironment: from cancer initiation to metastatic outgrowth. Cancer Cell. 2023;41(3):374–403. doi:10.1016/j.ccell.2023.02.016
43. Yang S, Wei W, Zhao Q. B7-H3, a checkpoint molecule, as a target for cancer immunotherapy. Int J Bio Sci. 2020;16(11):1767–1773. doi:10.7150/ijbs.41105
44. Koumprentziotis IA, Theocharopoulos C, Foteinou D, et al. New emerging targets in cancer immunotherapy: the role of B7-H3. Vaccines. 2024;12(1):54. doi:10.3390/vaccines12010054
45. Park J, Hsueh PC, Li Z, Ho PC. Microenvironment-driven metabolic adaptations guiding CD8+ T cell anti-tumor immunity. Immunity. 2023;56(1):32–42. doi:10.1016/j.immuni.2022.12.008
46. Chen L, Flies DB. Molecular mechanisms of T cell co-stimulation and co-inhibition. Nat Rev Immunol. 2013;13(4):227–242. doi:10.1038/nri3405
47. Sun X, Vale M, Leung E, Kanwar JR, Gupta R, Krissansen GW. Mouse B7-H3 induces antitumor immunity. Gene Ther. 2003;10(20):1728–1734. doi:10.1038/sj.gt.3302070
48. Lee YH, Martin-Orozco N, Zheng P, et al. Inhibition of the B7-H3 immune checkpoint limits tumor growth by enhancing cytotoxic lymphocyte function. Cell Res. 2017;27(8):1034–1045. doi:10.1038/cr.2017.90
49. Peuker K, Strigli A, Tauriello DVF, et al. Microbiota-dependent activation of the myeloid calcineurin-NFAT pathway inhibits B7H3- and B7H4-dependent anti-tumor immunity in colorectal cancer. Immunity. 2022;55(4):701–717.e7. doi:10.1016/j.immuni.2022.03.008
50. Cai D, Li J, Liu D, et al. Tumor-expressed B7-H3 mediates the inhibition of antitumor T-cell functions in ovarian cancer insensitive to PD-1 blockade therapy. Cell Mol Immunol. 2020;17(3):227–236. doi:10.1038/s41423-019-0305-2
51. Liu J, Kadier A, Guo Y, et al. Effect of tumor CD276 expression on infiltrating immune cells and clinicopathological features of prostate cancer. Prostate Cancer P D. 2023;27:1–3. doi:10.1038/s41391-023-00690-2
52. Bejarano L, Kauzlaric A, Lamprou E, et al. Interrogation of endothelial and mural cells in brain metastasis reveals key immune-regulatory mechanisms. Cancer Cell. 2024;42(3):378–395.e10. doi:10.1016/j.ccell.2023.12.018
53. Lee JS, Kim ME. B7H3 modulate tumor micro-environment via Th1 subset response. J Immunol. 2020;204(1_Supplement):
54. Shao L, Yu Q, Xia R, et al. B7-H3 on breast cancer cell MCF7 inhibits IFN-γ release from tumour-infiltrating T cells. Pathol Res Pract. 2021;224:153461. doi:10.1016/j.prp.2021.153461
55. Shi W, Wang Y, Zhao Y, et al. Immune checkpoint B7-H3 is a therapeutic vulnerability in prostate cancer harboring PTEN and TP53 deficiencies. Sci Trans med. 2023;15(695):eadf6724. doi:10.1126/scitranslmed.adf6724
56. Iida K, Miyake M, Onishi K, et al. Prognostic impact of tumor-infiltrating CD276/Foxp3-positive lymphocytes and associated circulating cytokines in patients undergoing radical nephrectomy for localized renal cell carcinoma. Oncol Lett. 2019;17(4):4004–4010. doi:10.3892/ol.2019.10057
57. Inamura K, Amori G, Yuasa T, Yamamoto S, Yonese J, Ishikawa Y. Relationship of B7-H3 expression in tumor cells and tumor vasculature with FOXP3+ regulatory T cells in renal cell carcinoma. CMAR. 2019;11:7021–7030. doi:10.2147/CMAR.S209205
58. Mendes AA, Lu J, Kaur HB, et al. Association of B7-H3 expression with racial ancestry, immune cell density, and androgen receptor activation in prostate cancer. Cancer. 2022;128(12):2269–2280. doi:10.1002/cncr.34190
59. Yang Q, Guo N, Zhou Y, Chen J, Wei Q, Han M. The role of tumor-associated macrophages (TAMs) in tumor progression and relevant advance in targeted therapy. Acta Pharm Sin B. 2020;10(11):2156–2170. doi:10.1016/j.apsb.2020.04.004
60. Lu H, Shi T, Wang M, et al. B7-H3 inhibits the IFN-γ-dependent cytotoxicity of Vγ9Vδ2 T cells against colon cancer cells. Oncoimmunology. 2020;9(1):1748991. doi:10.1080/2162402X.2020.1748991
61. Mortezaee K. B7-H3 immunoregulatory roles in cancer. Biomed Pharmacother. 2023;163:114890. doi:10.1016/j.biopha.2023.114890
62. Mao L, Fan TF, Wu L, et al. Selective blockade of B7-H3 enhances antitumour immune activity by reducing immature myeloid cells in head and neck squamous cell carcinoma. J Cell Mol Med. 2017;21(9):2199–2210. doi:10.1111/jcmm.13143
63. Pulido R, López JI, Nunes-Xavier CE. B7-H3: a robust target for immunotherapy in prostate cancer. Trends Cancer. 2024;10(7):584–587. doi:10.1016/j.trecan.2024.05.003
64. Miyamoto T, Murakami R, Hamanishi J, et al. B7-H3 suppresses antitumor immunity via the CCL2–CCR2–M2 macrophage axis and contributes to ovarian cancer progression. Cancer Immunol Res. 2022;10(1):56–69. doi:10.1158/2326-6066.CIR-21-0407
65. Wu M, Wu L, Wu W, et al. Phagocytosis of glioma cells enhances the immunosuppressive phenotype of bone marrow-derived macrophages. Cancer Res. 2023;83(5):771–785. doi:10.1158/0008-5472.CAN-22-1570
66. Cheng M, Chen S, Li K, et al. CD276-dependent efferocytosis by tumor-associated macrophages promotes immune evasion in bladder cancer. Nat Commun. 2024;15:2818. doi:10.1038/s41467-024-46735-5
67. Zhong Y, Tian Y, Wang Y, et al. Small extracellular vesicle piR-hsa-30937 derived from pancreatic neuroendocrine neoplasms upregulates CD276 in macrophages to promote immune evasion. Cancer Immunol Res. 2024:OF1–OF14. doi:10.1158/2326-6066.CIR-23-0825.
68. Zorko N, Shackelford M, Bozicevich A, et al. Effect of camelid B7-H3 tri-specific killer engagers on natural killer cells in patients with prostate cancer. JCO. 2024;42(4_suppl):155. doi:10.1200/JCO.2024.42.4_suppl.155
69. Schneider T, Hoffmann H, Dienemann H, et al. Non-small cell lung cancer induces an immunosuppressive phenotype of dendritic cells in tumor microenvironment by upregulating B7-H3. J Thorac Oncol. 2011;6(7):1162–1168. doi:10.1097/JTO.0b013e31821c421d
70. Zhong C, Tao B, Chen Y, et al. B7-H3 regulates glioma growth and cell invasion through a JAK2/STAT3/Slug-dependent signaling pathway. Onco Targets Ther. 2020;13:2215–2224. doi:10.2147/OTT.S237841
71. Nunes-Xavier C, Flem-Karlsen K, Tekle C, Ø F. Immunoregulatory protein B7-H3 promotes growth and decreases tumor cell sensitivity to anti-cancer agents. Ann Oncol. 2017;28:vii12. doi:10.1093/annonc/mdx509.001
72. Hu X, Xu M, Hu Y, Li N, Zhou L. B7-H3, negatively regulated by miR-128, promotes colorectal cancer cell proliferation and migration. Cell Biochem Biophys. 2021;79(2):397–405. doi:10.1007/s12013-021-00975-0
73. Yu TT, Zhang T, Lu X, Wang RZ. B7-H3 promotes metastasis, proliferation, and epithelial-mesenchymal transition in lung adenocarcinoma. Onco Targets Ther. 2018;11:4693–4700. doi:10.2147/OTT.S169811
74. Li Y, Guo G, Song J, et al. B7-H3 promotes the migration and invasion of human bladder cancer cells via the PI3K/Akt/STAT3 signaling pathway. J Cancer. 2017;8(5):816–824. doi:10.7150/jca.17759
75. Ding M, Liao H, Zhou N, Yang Y, Guan S, Chen L. B7-H3-induced signaling in lung adenocarcinoma cell lines with divergent epidermal growth factor receptor mutation patterns. Biomed Res Int. 2020;2020:8824805. doi:10.1155/2020/8824805
76. Liu Z, Zhang W, Phillips JB, et al. Immunoregulatory protein B7-H3 regulates cancer stem cell enrichment and drug resistance through MVP-mediated MEK activation. Oncogene. 2019;38(1):88–102. doi:10.1038/s41388-018-0407-9
77. Fan TF, Deng WW, Bu LL, Wu TF, Zhang WF, Sun ZJ. B7-H3 regulates migration and invasion in salivary gland adenoid cystic carcinoma via the JAK2/STAT3 signaling pathway. Am J Transl Res. 2017;9(3):1369–1380.
78. Liao H, Ding M, Zhou N, Yang Y, Chen L. B7‑H3 promotes the epithelial‑mesenchymal transition of NSCLC by targeting SIRT1 through the PI3K/AKT pathway. Mol Med Rep. 2022;25(3):79. doi:10.3892/mmr.2022.12595
79. Kang FB, Wang L, Jia HC, et al. B7-H3 promotes aggression and invasion of hepatocellular carcinoma by targeting epithelial-to-mesenchymal transition via JAK2/STAT3/Slug signaling pathway. Cancer Cell Int. 2015;15:45. doi:10.1186/s12935-015-0195-z
80. Xie J, Sun M, Zhang D, Chen C, Lin S, Zhang G. Fibronectin enhances tumor metastasis through B7-H3 in clear cell renal cell carcinoma. FEBS Open Bio. 2021;11(11):2977–2987. doi:10.1002/2211-5463.13280
81. Huh HD, Sub Y, Oh J, et al. Reprogramming Anchorage dependency by adherent-to-suspension transition promotes metastatic dissemination. Mol Cancer. 2023;22(1):63. doi:10.1186/s12943-023-01753-7
82. Park JH, Pyun WY, Park HW. Cancer metabolism: phenotype, signaling and therapeutic targets. Cells. 2020;9(10):2308. doi:10.3390/cells9102308
83. Lim S, Liu H, Madeira da Silva L, et al. Immunoregulatory protein B7-H3 reprograms glucose metabolism in cancer cells by ROS-mediated stabilization of HIF1α. Cancer Res. 2016;76(8):2231–2242. doi:10.1158/0008-5472.CAN-15-1538
84. Deng M, Wu D, Zhang Y, Jin Z, Miao J. MiR-29c downregulates tumor-expressed B7-H3 to mediate the antitumor NK-cell functions in ovarian cancer. Gynecol Oncol. 2021;162(1):190–199. doi:10.1016/j.ygyno.2021.04.013
85. Li Z, Liu J, Que L, Tang X. The immunoregulatory protein B7-H3 promotes aerobic glycolysis in oral squamous carcinoma via PI3K/Akt/mTOR pathway. J Cancer. 2019;10(23):5770–5784. doi:10.7150/jca.29838
86. Pulanco MC, Madsen AT, Tanwar A, Corrigan DT, Zang X. Recent advancements in the B7/CD28 immune checkpoint families: new biology and clinical therapeutic strategies. Cell Mol Immunol. 2023;20(7):694–713. doi:10.1038/s41423-023-01019-8
87. Shi T, Ma Y, Cao L, et al. B7-H3 promotes aerobic glycolysis and chemoresistance in colorectal cancer cells by regulating HK2. Cell Death Dis. 2019;10(4):308. doi:10.1038/s41419-019-1549-6
88. Luo D, Xiao H, Dong J, et al. B7-H3 regulates lipid metabolism of lung cancer through SREBP1-mediated expression of FASN. Biochem Biophys Res Commun. 2017;482(4):1246–1251. doi:10.1016/j.bbrc.2016.12.021
89. Xia L, Chen Y, Li J, et al. B7-H3 confers stemness characteristics to gastric cancer cells by promoting glutathione metabolism through AKT/pAKT/Nrf2 pathway. Chinese Med J. 2023;136(16):1977. doi:10.1097/CM9.0000000000002772
90. Ramjiawan RR, Griffioen AW, Duda DG. Anti-angiogenesis for cancer revisited: is there a role for combinations with immunotherapy? Angiogenesis. 2017;20(2):185–204. doi:10.1007/s10456-017-9552-y
91. Pouysségur J, Dayan F, Mazure NM. Hypoxia signalling in cancer and approaches to enforce tumour regression. Nature. 2006;441(7092):437–443. doi:10.1038/nature04871
92. Wang R, Ma Y, Zhan S, et al. B7-H3 promotes colorectal cancer angiogenesis through activating the NF-κB pathway to induce VEGFA expression. Cell Death Dis. 2020;11(1):55. doi:10.1038/s41419-020-2252-3
93. Lai H, Sun Z, Yang J, Wu P, Guo Y, Sun J. B7-H3 modulates endothelial cell angiogenesis through the VEGF cytokine. Immunol Res. 2019;67(2–3):202–211. doi:10.1007/s12026-019-09084-w
94. Zhang X, Ji J, Zhang G, et al. Expression and significance of B7-H3 and Tie-2 in the tumor vasculature of clear cell renal carcinoma. Onco Targets Ther. 2017;10:5417–5424. doi:10.2147/OTT.S147041
95. Lei X, Ou Z, Yang Z, et al. A pan-histone deacetylase inhibitor enhances the antitumor activity of B7-H3-specific CAR T cells in solid tumors. Clin Cancer Res. 2021;27(13):3757–3771. doi:10.1158/1078-0432.CCR-20-2487
96. Flem-Karlsen K, Tekle C, Andersson Y, Flatmark K, Ø F, Nunes-Xavier CE. Immunoregulatory protein B7-H3 promotes growth and decreases sensitivity to therapy in metastatic melanoma cells. Pigm Cell Melanoma Res. 2017;30(5):467–476. doi:10.1111/pcmr.12599
97. Flem-Karlsen K, Tekle C, Øyjord T, et al. p38 MAPK activation through B7-H3-mediated DUSP10 repression promotes chemoresistance. Sci Rep. 2019;9(1):5839. doi:10.1038/s41598-019-42303-w
98. Liu H, Tekle C, Chen YW, et al. B7-H3 silencing increases paclitaxel sensitivity by abrogating Jak2/Stat3 phosphorylation. Mol Cancer Ther. 2011;10(6):960–971. doi:10.1158/1535-7163.MCT-11-0072
99. Zhou L, Zhao Y. B7-H3 induces ovarian cancer drugs resistance through an PI3K/AKT/BCL-2 signaling pathway. Cancer Manag Res. 2019;11:10205–10214. doi:10.2147/CMAR.S222224
100. Li Y, Yang X, Yao P, et al. B7-H3 increases the radioresistance of gastric cancer cells through regulating baseline levels of cell autophagy. Am J Transl Res. 2019;11(7):4438–4449.
101. Loo D, Alderson RF, Chen FZ, et al. Development of an Fc-enhanced Anti–B7-H3 monoclonal antibody with potent antitumor activity. Clin Cancer Res. 2012;18(14):3834–3845. doi:10.1158/1078-0432.CCR-12-0715
102. Yonesaka K, Haratani K, Takamura S, et al. B7-H3 negatively modulates CTL-mediated cancer immunity. Clin Cancer Res. 2018;24(11):2653–2664. doi:10.1158/1078-0432.CCR-17-2852
103. Shenderov E, De Marzo AM, Lotan TL, et al. Neoadjuvant enoblituzumab in localized prostate cancer: a single-arm, phase 2 trial. Nat Med. 2023;29(4):888–897. doi:10.1038/s41591-023-02284-w
104. Stefańczyk SA, Hagelstein I, Lutz MS, et al. Induction of NK cell reactivity against acute myeloid leukemia by Fc-optimized CD276 (B7-H3) antibody. Blood Cancer J. 2024;14(1):1–10. doi:10.1038/s41408-024-01050-6
105. Zhang C, Li K, Zhu H, et al. ITGB6 modulates resistance to anti-CD276 therapy in head and neck cancer by promoting PF4+ macrophage infiltration. Nat Commun. 2024;15(1):7077. doi:10.1038/s41467-024-51096-0
106. van de DNWCJ, Zweegman S. T-cell-engaging bispecific antibodies in cancer. Lancet. 2023;402(10396):142–158. doi:10.1016/S0140-6736(23)00521-4
107. Shankar S, Spira AI, Strauss J, et al. A Phase 1, open label, dose escalation study of MGD009, a humanized B7-H3 x CD3 DART protein, in combination with MGA012, an anti-PD-1 antibody, in patients with relapsed or refractory B7-H3-expressing tumors. JCO. 2018;36(15_suppl):TPS2601. doi:10.1200/JCO.2018.36.15_suppl.TPS2601
108. Ma J, Ma P, Zhao C, et al. B7-H3 as a promising target for cytotoxicity T cell in human cancer therapy. Oncotarget. 2016;7(20):29480–29491. doi:10.18632/oncotarget.8784
109. Fan R, Chen C, Mu M, et al. Engineering MMP-2 activated nanoparticles carrying B7-H3 bispecific antibodies for ferroptosis-enhanced glioblastoma immunotherapy. Acs Nano. 2023;17(10):9126–9139. doi:10.1021/acsnano.2c12217
110. Kontos F, Michelakos T, Kurokawa T, et al. B7-H3: an attractive target for antibody-based immunotherapy. Clin Cancer Res. 2021;27(5):1227–1235. doi:10.1158/1078-0432.CCR-20-2584
111. Zekri L, Lutz M, Prakash N, et al. An optimized IgG-based B7-H3xCD3 bispecific antibody for treatment of gastrointestinal cancers. Mol Ther. 2023;31(4):1033–1045. doi:10.1016/j.ymthe.2023.02.010
112. Tarantino P, Carmagnani Pestana R, Corti C, et al. Antibody–drug conjugates: smart chemotherapy delivery across tumor histologies. Ca A Cancer J Clin. 2022;72(2):165–182. doi:10.3322/caac.21705
113. Dumontet C, Reichert JM, Senter PD, Lambert JM, Beck A. Antibody–drug conjugates come of age in oncology. Nat Rev Drug Discov. 2023;22(8):641–661. doi:10.1038/s41573-023-00709-2
114. Scribner JA, Brown JG, Son T, et al. Preclinical development of MGC018, a duocarmycin-based antibody–drug conjugate targeting B7-H3 for solid cancer. Mol Cancer Ther. 2020;19(11):2235–2244. doi:10.1158/1535-7163.MCT-20-0116
115. Brignole C, Calarco E, Bensa V, et al. Antitumor activity of the investigational B7-H3 antibody-drug conjugate, vobramitamab duocarmazine, in preclinical models of neuroblastoma. J Immunother Cancer. 2023;11(9):e007174. doi:10.1136/jitc-2023-007174
116. Ifinatamab Deruxtecan Continues to Demonstrate Promising Objective Response Rates in Patients with Extensive-Stage Small Cell Lung Cancer in IDeate-Lung01 Phase 2 Trial. Merck.com. Available from: https://www.merck.com/news/ifinatamab-deruxtecan-continues-to-demonstrate-promising-objective-response-rates-in-patients-with-extensive-stage-small-cell-lung-cancer-in-ideate-lung01-phase-2-trial/.
117. Jacobson C, Emmert A, Rosenthal MB. CAR T-cell therapy: a microcosm for the challenges ahead in medicare. JAMA. 2019;322(10):923–924. doi:10.1001/jama.2019.10194
118. Du H, Hirabayashi K, Ahn S, et al. Antitumor responses in the absence of toxicity in solid tumors by targeting B7-H3 via chimeric antigen receptor T cells. Cancer Cell. 2019;35(2):221–237.e8. doi:10.1016/j.ccell.2019.01.002
119. Theruvath J, Sotillo E, Mount CW, et al. Locoregionally administered B7-H3-targeted CAR T cells for treatment of atypical teratoid/rhabdoid tumors. Nat Med. 2020;26(5):712–719. doi:10.1038/s41591-020-0821-8
120. Haydar D, Houke H, Chiang J, et al. Cell-surface antigen profiling of pediatric brain tumors: B7-H3 is consistently expressed and can be targeted via local or systemic CAR T-cell delivery. Neuro Oncol. 2021;23(6):999–1011. doi:10.1093/neuonc/noaa278
121. Jiang L, You F, Wu H, et al. B7-H3–targeted CAR-Vδ1T cells exhibit potent broad-spectrum activity against solid tumors. Cancer Res. 2024;84(23):4066–4080. doi:10.1158/0008-5472.CAN-24-0195
122. Vitanza NA, Wilson AL, Huang W, et al. Intraventricular B7-H3 CAR T cells for diffuse intrinsic pontine glioma: preliminary first-in-human bioactivity and safety. Cancer Discov. 2023;13(1):114–131. doi:10.1158/2159-8290.CD-22-0750
123. Lichtman EI, Du H, Shou P, et al. Preclinical evaluation of B7-H3-specific chimeric antigen receptor T cells for the treatment of acute myeloid leukemia. Clin Cancer Res. 2021;27(11):3141–3153. doi:10.1158/1078-0432.CCR-20-2540
124. Ventin M, Cattaneo G, Arya S, et al. Chimeric antigen receptor T cell with an inducible caspase-9 suicide gene eradicates uveal melanoma liver metastases via B7-H3 targeting. Clin Cancer Res. 2024;30(15):3243–3258. doi:10.1158/1078-0432.CCR-24-0071
125. Lee HW, O’Reilly C, Beckett AN, Currier DG, Chen T, DeRenzo C. A high-content screen of FDA approved drugs to enhance CAR T cell function: ingenol-3-angelate improves B7-H3-CAR T cell activity by upregulating B7-H3 on the target cell surface via PKCα activation. J Exp Clin Canc Res. 2024;43(1):97. doi:10.1186/s13046-024-03022-x
126. Timpanaro A, Piccand C, Dzhumashev D, et al. CD276-CAR T cells and Dual-CAR T cells targeting CD276/FGFR4 promote rhabdomyosarcoma clearance in orthotopic mouse models. J Exp Clin Cancer Res. 2023;42(1):293. doi:10.1186/s13046-023-02838-3
127. Haydar D, Crawford J, Chou CH, et al. Immu-01. Evaluating The Impact of Car Design on the Tumor Immune Microenvironment and Anti-Tumor Response in Syngeneic Glioma Models. Neuro-Oncology. 2023;25(Supplement_1):i49. doi:10.1093/neuonc/noad073.188
128. Vitanza N, Ronsley R, Wilson A, et al. TRLS-12. Intraventricular B7-H3 Car T Cells for Diffuse Intrinsic Pontine Glioma: Interim Analysis of Brainchild-03 Arm C. Neuro-Oncology. 2023;25(Supplement_1):i81–i82. doi:10.1093/neuonc/noad073.315
129. Fanuzzi M, Cole A, Singh MP, et al. IMMU-20. Evaluation of B7-H3 Nanobody Car-T Cells in Pediatric Glioblastoma. Neuro-Oncology. 2023;25(Supplement_1):i53–i54. doi:10.1093/neuonc/noad073.207
130. Li X, Sun S, Zhang W, et al. Identification of genetic modifiers enhancing B7-H3-targeting CAR T cell therapy against glioblastoma through large-scale CRISPRi screening. J Exp Clin Canc Res. 2024;43(1):95. doi:10.1186/s13046-024-03027-6
131. Nishimura CD, Corrigan D, Zheng XY, et al. TOP CAR with TMIGD2 as a safe and effective costimulatory domain in CAR cells treating human solid tumors. Sci Adv. 2024;10(19):eadk1857. doi:10.1126/sciadv.adk1857
132. Zeng S, Chen L, Tian J, et al. Integrative analysis of pan-cancer single-cell data reveals a tumor ecosystem subtype predicting immunotherapy response. Npj Precis Onc. 2024;8(1):1–13. doi:10.1038/s41698-024-00703-w
133. Rausch M, Bartels K, Leibold J. Immunotherapy in ovarian cancer: spatial functional genomics to unravel resistance mechanisms. Sig Transduct Target Ther. 2025;10(1):1–3. doi:10.1038/s41392-024-02110-w
134. Zhang J, Peng Q, Fan J, et al. Single-cell and spatial transcriptomics reveal SPP1-CD44 signaling drives primary resistance to immune checkpoint inhibitors in RCC. J Transl Med. 2024;22(1):1157. doi:10.1186/s12967-024-06018-5
135. Behan FM, Iorio F, Picco G, et al. Prioritization of cancer therapeutic targets using CRISPR–Cas9 screens. Nature. 2019;568(7753):511–516. doi:10.1038/s41586-019-1103-9
136. Uijttewaal ECH, Lee J, Sell AC, et al. CRISPR-StAR enables high-resolution genetic screening in complex in vivo models. Nat Biotechnol. 2024:1–13. doi:10.1038/s41587-024-02512-9.
137. Fandrey CI, Jentzsch M, Konopka P, et al. NIS-Seq enables cell-type-agnostic optical perturbation screening. Nat Biotechnol. 2024:1–11. doi:10.1038/s41587-024-02516-5.
138. Li Y, Huang Z, Xu L, et al. UDA-seq: universal droplet microfluidics-based combinatorial indexing for massive-scale multimodal single-cell sequencing. Nat Methods. 2025:1–14. doi:10.1038/s41592-024-02586-y.
139. Inoue J, Inazawa J. Cancer-associated miRNAs and their therapeutic potential. J Hum Genet. 2021;66(9):937–945. doi:10.1038/s10038-021-00938-6
140. Cao C, Li A, Xu C, Wu B, Yao L, Liu Y. Engineering artificial non-coding RNAs for targeted protein degradation. Nat Chem Biol. 2024;1–9. doi:10.1038/s41589-024-01719-w
141. Shi X, Zhang D, Li F, et al. Targeting glycosylation of PD-1 to enhance CAR-T cell cytotoxicity. J Hematol Oncol. 2019;12(1):127. doi:10.1186/s13045-019-0831-5
142. Tonne J, Thompson J, Pulido J, et al. Efficacy of function-enhanced, re-activatable, dual-specific CAR T cells pre-loaded with oncolytic virus for immunotherapy of high-grade glioma. J Clin Oncol. 2023;2023:2062. doi:10.1200/JCO.2023.41.16_suppl.2062
143. Song F, Dotti G. A cocktail of kinase inhibitors that enhance the antitumor effects of CAR-T cell therapy. Nat Immunol. 2025;1–2. doi:10.1038/s41590-024-02049-8
144. Sokouti B. A review on in silico virtual screening methods in COVID-19 using anticancer drugs and other natural/chemical inhibitors. Explor Target Antitumor Ther. 2023;4(5):994–1026. doi:10.37349/etat.2023.00177
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
RNA Modifications Meet Tumors
Yang Z, Zhang S, Xia T, Fan Y, Shan Y, Zhang K, Xiong J, Gu M, You B
Cancer Management and Research 2022, 14:3223-3243
Published Date: 22 November 2022
Noncoding RNA-Mediated Regulation of Myeloid-Derived Suppressor Cells in Cancer
Luo K, Xu Y, Chen J, Song JJ, Zhang R, Zhang W, Jiang P
Cancer Management and Research 2025, 17:2567-2587
Published Date: 30 October 2025