Back to Journals » Cancer Management and Research » Volume 17

Noncoding RNA-Mediated Regulation of Myeloid-Derived Suppressor Cells in Cancer
Authors Luo K ![]() , Xu Y, Chen J, Song JJ, Zhang R, Zhang W, Jiang P
, Xu Y, Chen J, Song JJ, Zhang R, Zhang W, Jiang P
Received 1 July 2025
Accepted for publication 16 October 2025
Published 30 October 2025 Volume 2025:17 Pages 2567—2587
DOI https://doi.org/10.2147/CMAR.S550896
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 4
Editor who approved publication: Professor Seema Singh
Kengjun Luo,1 Ying Xu,2,3 Jiahao Chen,1 Jingyang JY Song,1 Rui Zhang,1 Wenbo Zhang,1 Pengcheng Jiang1
1Department of General Surgery, The Affiliated People’s Hospital of Jiangsu University, Zhenjiang, Jiangsu, 212000, People’s Republic of China; 2Department of Laboratory Center, The Affiliated People’s Hospital of Jiangsu University, Zhenjiang, Jiangsu, 212000, People’s Republic of China; 3Jiangsu Key Laboratory of Medical Science and Laboratory Medicine, School of Medicine, Jiangsu university, Zhenjiang, Jiangsu, 212013, People’s Republic of China
Correspondence: Wenbo Zhang, Department of General Surgery, The Affiliated People’s Hospital of Jiangsu University, Zhenjiang, 212000, People’s Republic of China, Email [email protected] Pengcheng Jiang, Department of General Surgery, The Affiliated People’s Hospital of Jiangsu University, Zhenjiang, 212000, People’s Republic of China, Email [email protected]
Abstract: Myeloid-derived suppressor cells (MDSCs) arise from myeloid progenitors in the bone marrow and, under the influence of tumor- and immune-cell-derived cytokines, chemokines, and growth factors, enhance immunosuppressive activity within the tumor microenvironment (TME). Noncoding RNAs (ncRNAs)—including microRNAs (miRNAs), long noncoding RNAs (lncRNAs), and circular RNAs (circRNAs)—have emerged as critical regulators of MDSCs biology. Recent evidence has shown that ncRNAs are intimately involved in MDSCs recruitment, differentiation, and suppressive function by modulating key signaling pathways, including STAT3, NF-κB, and PI3K/AKT. Mechanistically, ncRNAs act through epigenetic control (eg, histone modifications and chromatin remodeling), post-transcriptional regulation (eg, miRNA sponging), and fine-tuning of gene networks. These insights highlight RNA-based strategies that target ncRNAs to disrupt MDSCs-mediated immune suppression and potentiate antitumor immunity, while acknowledging ongoing challenges such as delivery specificity, stability, and off-target effects. This review synthesizes current understanding of how ncRNAs regulate MDSCs via major signaling axes and discusses implications for cancer progression and therapeutic development.
Keywords: ncRNAs, MDSCs, tumor, tumor microenvironment, tumor immunity
Introduction
Cancer remains a leading cause of mortality worldwide. According to GLOBOCAN 2022, nearly 20 million new cases and 9.7 million deaths were reported in that year alone.1 Despite advances in serological testing, imaging, and endoscopy, the limited specificity of current diagnostic methods often results in late-stage detection. Likewise, standard therapeutic approaches such as neoadjuvant radiotherapy, adjuvant chemotherapy, and surgery have achieved only modest improvements in patient outcomes. These limitations underscore the urgent need for innovative strategies that target the biological mechanisms underpinning tumor progression and immune evasion.
A primary mechanism driving these clinical challenges is the tumor’s ability to create a profoundly immunosuppressive tumor microenvironment (TME). This complex ecosystem allows cancer cells to evade immune surveillance and resist immunotherapy. Among the key architects of this immunosuppressive network are myeloid-derived suppressor cells (MDSCs). While their foundational biology was defined in a seminal review,2 our current understanding continues to evolve, particularly regarding their complex roles in the human tumor microenvironment.3 Initially described in tumor-bearing hosts as immature myeloid cells with potent suppressive activity, MDSCs have been progressively consolidated under consensus criteria, with refinements to their markers, developmental routes, and functional boundaries.4,5 Driven by tumor- and immune-derived signals, MDSCs exert profound immunosuppressive effects, thereby facilitating tumor immune escape and resistance to conventional therapies.5,6 Their central role has made MDSCs attractive targets for next-generation precision oncology. At the same time, unresolved complexity remains in their markers, developmental routes, and functional states.4 In parallel with advances in tumor immunology, a deeper understanding of gene regulation has revealed noncoding RNAs (ncRNAs) as essential regulators of virtually all cellular processes.7 These molecules—including long noncoding RNAs (lncRNAs), circular RNAs (circRNAs), microRNAs (miRNAs), and PIWI-interacting RNAs (piRNAs)—are now recognized to constitute the majority of the human transcriptome.7,8 Beyond their regulation of tumor-intrinsic processes such as proliferation, migration, and apoptosis, ncRNAs also fine-tune immune-cell function and fate decisions. Through these roles, ncRNAs help shape the tumor microenvironment, influencing immune escape and therapeutic sensitivity.9,10 Recent studies have highlighted a direct intersection between ncRNAs and MDSCs.11 For example, miR-155 has been implicated in modulating MDSC function and has been explored in combination delivery platforms,12 whereas lncRNA HOTAIR recruits EZH2 to drive histone modifications that affect MDSC differentiation and function.13 Other ncRNAs regulate tumor–immune interactions via chromatin remodeling, miRNA sponging, or exosome-mediated communication.14–16 Notably, several regulatory effects appear context dependent, and contradictory findings have been reported across tumor types; these limitations are summarized in the sections below. Collectively, these findings position ncRNAs not only as biomarkers for cancer diagnosis but also as potential therapeutic targets for modulating MDSC activity.17 Beyond ncRNA-mediated regulation, MDSC programs are shaped by cytokine/chemokine cues and pharmacologic or metabolic interventions. Tumor-derived GM-CSF/G-CSF/IL-6–STAT3 signaling, arginase-1/iNOS-driven metabolic rewiring, and nanoparticle-based drug delivery systems have each demonstrated MDSC-suppressive activity in preclinical models.2,3,15 These strategies provide a complementary framework against which ncRNA-centered approaches can be interpreted. This review therefore summarizes the molecular mechanisms by which ncRNAs regulate MDSCs across diverse tumor contexts, highlighting their implications for cancer progression and therapy.
MDSCs: Key Roles in Tumor Biology
The Origin and Development of MDSCs
MDSCs are pivotal immunosuppressive cells originating from myeloid progenitors in the bone marrow. Under normal physiological conditions, these progenitor cells differentiate into essential immune effectors, including monocytes, macrophages, dendritic cells, and granulocytes, forming the frontline defense of the host.18 However, pathological states such as chronic inflammation or cancer disrupt this orderly differentiation, leading instead to the abnormal accumulation of immunosuppressive MDSCs.19 This disrupted maturation significantly facilitates immune evasion by tumors and is involved in various other pathological scenarios, as illustrated in Figure 1.
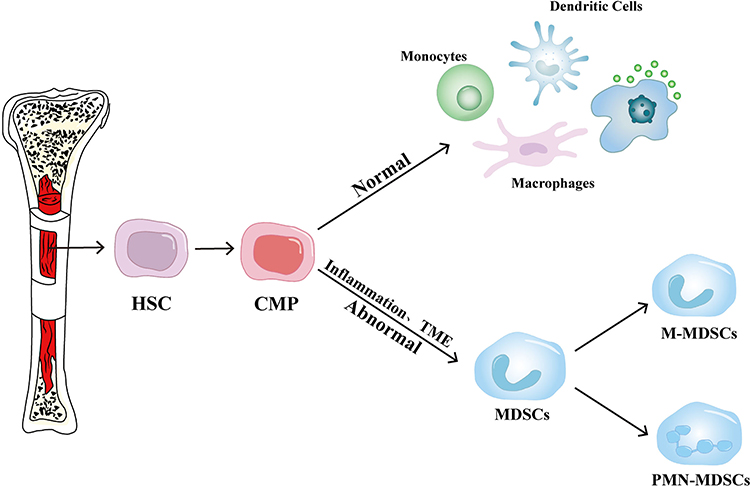 |
Figure 1 Origin and development of MDSCs. Under normal physiological conditions, hematopoietic stem cells (HSCs) and common myeloid progenitors (CMPs) differentiate into monocytes, dendritic cells, and macrophages. However, in the presence of inflammation or abnormalities in the tumor microenvironment (TME), CMPs give rise to immature MDSCs. These MDSCs further differentiate into monocytic MDSCs (M-MDSCs) or polymorphonuclear MDSCs (PMN-MDSCs), both of which play a key role in mediating immunosuppression within the TME. |
The differentiation of MDSCs is intricately governed by diverse inflammatory signals, notably granulocyte-macrophage colony-stimulating factor (GM-CSF) and granulocyte colony-stimulating factor (G-CSF), which play key roles in their regulation.20 These cytokines often accumulate excessively within the tumor microenvironment, inducing myeloid progenitors to adopt immunosuppressive characteristics.21 Additionally, other cytokines like interleukins (IL-6, IL-1β), chemokines, and tumor necrosis factor-alpha (TNF-α) significantly contribute to this differentiation process.22–24 The combined action of these factors promotes aberrant differentiation of myeloid progenitors, ultimately leading to the enhanced production of immunosuppressive MDSCs under pathological conditions.
Subtypes of MDSCs
MDSCs represent a pathological state of activated monocytes and relatively immature neutrophils. In murine models, MDSCs are generally divided into two main subsets, distinguished by the hallmark surface markers GR-1 and CD11b: monocytic MDSCs (M-MDSCs) and granulocytic MDSCs (G-MDSCs), also referred to as polymorphonuclear MDSCs (PMN-MDSCs). M-MDSCs are characterized by the surface markers CD11b+Ly6ChighLy6Glow/−, while PMN-MDSCs are defined by CD11b+Ly6Clow/−Ly6G+.25 Unlike murine MDSCs, human MDSCs lack the expression of GR-1, resulting in distinct classification criteria. In human peripheral blood mononuclear cells, MDSCs are typically identified by the phenotype CD33+, CD11b+, and HLA-DRlow/−. Based on the expression of CD15 and CD14 markers, human MDSCs are classified into granulocytic MDSCs (G-MDSCs) and monocytic MDSCs (M-MDSCs).26 Additionally, a distinct subset of bone marrow progenitor cells with MDSC-like characteristics, termed “early-stage myeloid-derived suppressor cells” (eMDSCs), has been identified exclusively in humans. This population primarily comprises bone marrow progenitor and precursor cells, accounting for less than 5% of the total MDSC population. eMDSCs are defined by the surface markers Lin−HLA-DR−CD33+ CD11b+, where “Lin” refers to a lineage-negative panel that includes CD3, CD14, CD15, CD19, and CD5627–29 (Figure 2).
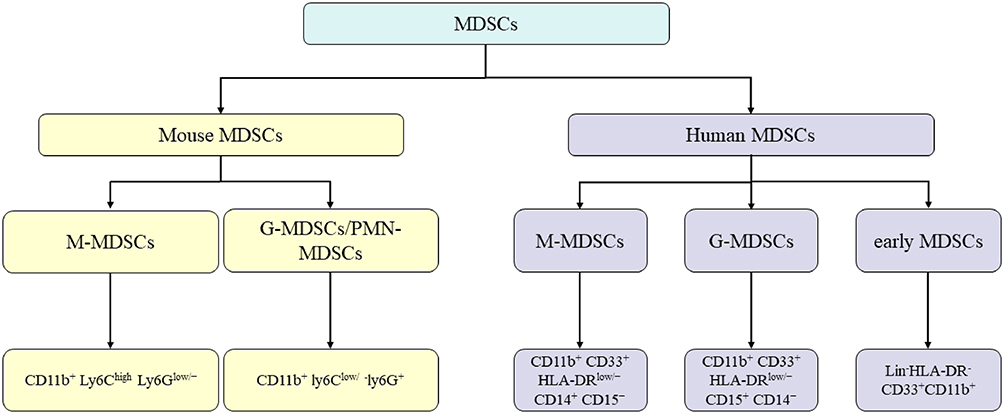 |
Figure 2 The Classification and Characteristics of MDSCs. MDSCs are broadly classified into monocytic MDSCs (M-MDSCs) and granulocytic/polymorphonuclear MDSCs (G-MDSCs/PMN-MDSCs) in both mice and humans. In mice, M-MDSCs are identified by the surface markers CD11b+ Ly6Chigh Ly6G low/−, while G-MDSCs/PMN-MDSCs are characterized as CD11b+ Ly6Clow/− Ly6G+. In humans, M-MDSCs are CD11b+ CD33+ HLA-DRlow/− CD14+ CD15−.and G-MDSCs are CD11b+ CD33+ HLA-DRlow/− CD15+ CD14−. An additional subset, early-stage MDSCs (eMDSCs), is identified in humans by the markers Lin− HLA-DR− CD33+ CD11b+. |
Mechanisms of MDSCs Formation
MDSCs are pivotal in establishing the TME and play an essential role in immune evasion and tumor progression. Key signaling pathways such as STAT3, NF-κB, and PI3K/AKT play crucial roles in regulating MDSCs recruitment, differentiation, and expansion. These pathways are activated by cytokines and growth factors within the TME and work synergistically to promote MDSCs function. STAT3, for example, promotes MDSCs proliferation and survival, contributing to their expansion within the TME.30,31 NF-κB signaling enhances MDSCs differentiation and facilitates their immune suppressive function.32 PI3K/AKT signaling is involved in metabolic reprogramming, supporting MDSCs survival and immune suppression.33 Together, these signaling pathways orchestrate the formation of an immune-suppressive niche that not only facilitates tumor growth but also promotes resistance to immune checkpoint inhibitors and other cancer therapies.
The recruitment, differentiation, and expansion of MDSCs are regulated by a sophisticated interplay of cytokines, chemokines, and growth factors secreted by tumor and immune cells.3 Tumor-derived chemokines such as CXCL8 (in humans) and CXCL5 (in mice) are key regulators of MDSCs recruitment, directing these cells to the tumor site, where they suppress the infiltration and function of effector immune cells, including T cells and natural killer (NK) cells.34 Furthermore, signaling through B7H3 and CXCR4 is integral to MDSCs recruitment, with tumor-derived exosomes activating the TLR2/NF-κB pathway, thereby enhancing MDSCs infiltration and fostering immune suppression within the TME.35,36
Upon entering the TME, MDSCs differentiate into various immunosuppressive cell types, such as regulatory T cells (Tregs), tumor-associated macrophages (TAMs), and cancer-associated fibroblasts (CAFs), all of which contribute to the tumor’s ability to escape immune detection.37 The differentiation of MDSCs is primarily driven by cytokines such as G-CSF, IL-6, and GM-CSF, which facilitate the conversion of MDSCs into Tregs and TAMs, thereby promoting the establishment of an immunosuppressive microenvironment conducive to tumor growth.38,39 In addition, netrin-1, secreted by tumor cells, significantly facilitates the differentiation of MDSCs via its receptor A2BR, thereby amplifying immune escape and accelerating tumor progression.40
The expansion of MDSCs is primarily driven by persistent inflammation and the continuous secretion of growth factors such as G-CSF, GM-CSF, and IL-6, alongside activation of the CXCR2/CXCL1 signaling axis. This not only facilitates MDSCs migration into the tumor site but also enhances their proliferative capacity, resulting in a significant accumulation of MDSCs within the TME.41 The increased presence of MDSCs exacerbates immune suppression, further promoting tumor metastasis and resistance to both conventional and immunotherapies.42 Moreover, MDSCs maintain an immunosuppressive niche in the TME through the secretion of cytokines such as IL-10 and TGF-β, which dampen effector immune responses and allow tumor cells to evade immune detection and resist therapeutic interventions.43,44
Importantly, targeting MDSCs recruitment, differentiation, and expansion holds significant therapeutic potential, as it may enhance the efficacy of immune-based therapies and improve patient outcomes in cancer treatment.
Mechanisms of Action of MDSCs
MDSCs are a population of myeloid-derived cells with potent immunosuppressive properties, and they are commonly found in the majority of cancer types.45 The key mechanisms underlying their activity can be outlined as follows:
Initially, MDSCs suppress immune responses through a variety of mechanisms, exerting multiple effects on T cell function. By secreting immunosuppressive molecules, including arginase and nitric oxide synthase, MDSCs directly interfere with T cell activity. These molecules deplete critical amino acids, such as L-arginine, in the local microenvironment, thereby restricting T cell proliferation and activation.46 Moreover, MDSCs indirectly suppress T cell activation and function by secreting immunosuppressive cytokines, notably TGF-β and IL-10.47 These factors collectively dampen T cell responsiveness, significantly weakening the overall immune defense.
In addition to direct suppression of T cells, MDSCs enhance immune suppression indirectly by stimulating the expansion and function of Tregs. The increased number and activity of Tregs restrict the activation of other immune populations, further suppressing immune responses.48 Direct cell-to-cell interactions between MDSCs and T cells, involving inhibitory surface molecules such as PD-1 ligands, have also been shown to suppress T cell functions, thus intensifying the immune-suppressive environment.49
Furthermore, MDSCs actively support tumor progression and metastasis by modulating the TME. They release multiple cytokines and chemokines, such as VEGF, IL-6, and IL-1β, exacerbating local inflammation and creating conditions favorable for tumor growth. Additionally, MDSCs facilitate tumor angiogenesis, enhancing blood vessel formation to supply nutrients, further promoting tumor expansion and dissemination.50
Lastly, MDSCs boost tumor invasiveness by supporting metastatic processes. They secrete enzymes like matrix metalloproteinases (MMPs), which degrade and remodel the extracellular matrix (ECM), facilitating tumor cell invasion and spread.51 MDSCs also migrate to distant pre-metastatic sites, where they produce cytokines such as CXCL12, establishing an immunosuppressive microenvironment conducive to metastatic tumor cell colonization and growth, thus significantly aiding tumor metastasis and progression.52
Together, these mechanisms position MDSCs as critical factors in tumor growth, immune suppression, and metastasis (Figure 3).
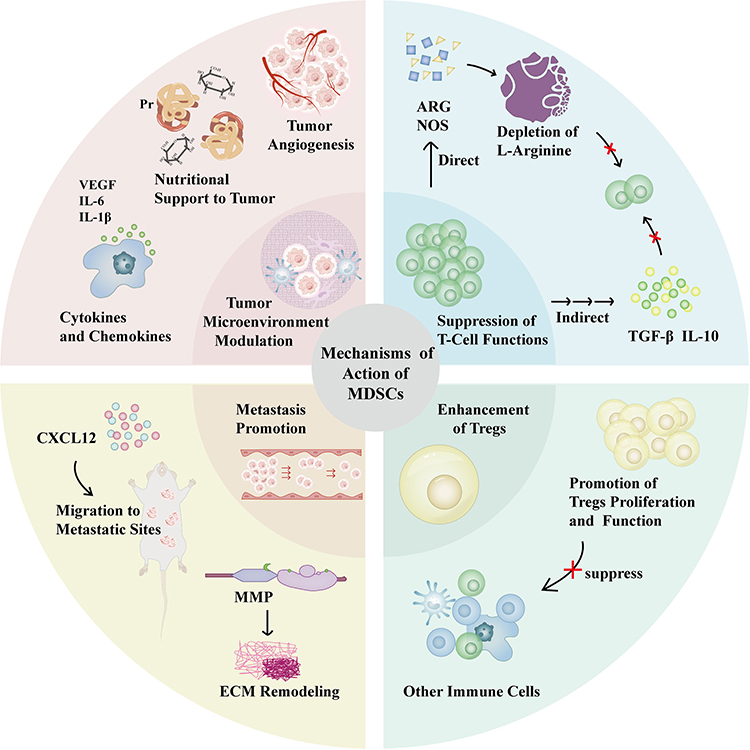 |
Figure 3 Schematic representation of the functional roles of MDSCs in the TME. MDSCs utilize multiple mechanisms to suppress immune responses and facilitate cancer progression. They secrete immunosuppressive molecules (eg, ARG, NOS, TGF-β, IL-10), which suppress T-cell function both directly and indirectly. MDSCs also promote the proliferation and activity of Tregs and release cytokines (eg, VEGF, IL-6, IL-1β) to remodel the TME and drive inflammation. In addition, MDSCs contribute to tumor angiogenesis by supporting vascular development, facilitate ECM remodeling through the secretion of MMPs, and migrate to pre-metastatic sites, where they secrete chemokines such as CXCL12 to establish an immunosuppressive niche. Collectively, these mechanisms enhance tumor growth, immune evasion, and metastasis. |
Regulation of MDSCs by miRNAs in Tumors
Digestive System Tumor
In digestive system tumors, miRNAs play a pivotal role in modulating the immunosuppressive functions of MDSCs through various signaling pathways, including STAT3, PI3K/AKT, TGF-β/SMAD, NF-κB, and PD-L1-mediated immune evasion. These pathways regulate MDSC recruitment, differentiation, and activation, thus facilitating tumor progression and immune escape.
The STAT3 pathway is a central regulator of MDSC activity. It is crucial for MDSC proliferation, survival, and immune suppression. miR-155 enhances MDSC immunosuppressive functions in colorectal cancer by targeting SOCS1, which amplifies STAT3 activation and accelerates tumor progression.53 In esophageal cancer, CAFs secrete IL-6 and exosomal miR-21, which activate STAT3 signaling, inducing M-MDSC generation and promoting the secretion of immunosuppressive cytokines like IL-10 and TGF-β, which inhibit CD8+ T cell activity and contribute to chemotherapy resistance.54
The PI3K/AKT pathway is another critical signaling mechanism in MDSCs regulation. miR-494 in gastric cancer is regulated by tumor-secreted TGF-β, which upregulates miR-494 expression, leading to PTEN downregulation and PI3K/AKT activation. PTEN is a tumor suppressor gene that negatively regulates the PI3K/AKT pathway. In its absence or reduced expression, this pathway becomes hyperactivated, which promotes MDSCs-mediated inhibition of T cell proliferation and interferon-γ production, further promoting immune suppression.55 Likewise, miR-26b-5p promotes differentiation and immunosuppressive activities of MDSCs in esophageal cancer by targeting PTEN, facilitating immune evasion and metastatic progression.56
The TGF-β/SMAD signaling pathway significantly influences MDSC function, driving their differentiation, improving survival, and enhancing their immunosuppressive capacity. In hepatocellular carcinoma, decreased expression of miR-101-3p and miR-490-3p elevates TGFBR1 levels, resulting in increased infiltration and immunosuppressive potential of MDSCs.57 Moreover, in gastric cancer, CAF-derived TGF-β further amplifies MDSC-mediated immune suppression via miR-494, forming a self-reinforcing loop that strengthens TGF-β/SMAD signaling.55
The NF-κB pathway represents another critical route through which MDSCs are regulated, particularly under inflammatory and immunosuppressive conditions. For instance, in gastric cancer, miR-130b enhances NF-κB activity by targeting CYLD, boosting the immunosuppressive functions of SLFN4+ MDSCs. This activation promotes gastric epithelial cell proliferation and the initiation of premalignant lesions.58 NF-κB signaling further amplifies the release of pro-inflammatory cytokines and immunosuppressive mediators, thus supporting tumor immune escape.
Additionally, miRNAs regulate immune evasion mechanisms involving PD-L1. For example, in colorectal cancer, miR-570 expression is suppressed by ZnC, thereby enhancing the immunosuppressive role of MDSCs and accelerating tumor progression.59 miR-93 and miR-106b also influence the bone marrow microenvironment by repressing CXCL12 expression, reducing stem cell migration and diminishing MDSC-mediated suppression of T cells via the inhibition of PD-L1.60
Gynecological Tumor
miRNAs significantly influence MDSCs activity in gynecological tumors through various signaling cascades, including SOCS3-JAK/STAT, NF-κB, HIF-1α, and oxidative stress pathways. These pathways collectively regulate MDSCs recruitment, differentiation, and immunosuppressive functions, facilitating immune evasion and tumor progression.
The SOCS3/JAK/STAT pathway represents a critical regulatory mechanism in gynecological cancers mediated by miRNAs. miR-9 targets SOCS3, a known suppressor of JAK/STAT signaling, promoting MDSC recruitment and increasing their suppressive capacity within the breast cancer microenvironment, thus supporting tumor growth and immune evasion.61 Exosomal miR-9 and miR-181a secreted by breast cancer cells also intensify JAK/STAT activation by targeting SOCS3 and PIAS3, significantly expanding eMDSCs and enhancing their immunosuppressive effects, further enabling tumor progression and immune escape.62 Similarly, miR-155 aids the recruitment and activity of MDSCs in tumors, contributing to immune suppression. Delivery systems using miR-155 inhibitors have demonstrated potential in reducing MDSC populations, impairing their suppressive functions, and boosting antitumor CD8+ T cell responses.12
NF-κB signaling is another essential miRNA target in gynecological cancers. miR-146a suppresses NF-κB signaling in breast cancer, reducing immunosuppressive factor secretion by MDSCs and promoting their polarization toward an M1 phenotype. This shift diminishes MDSC numbers and enhances CD8+ T cell-mediated antitumor immunity.63 Furthermore, miR-146a and miR-155 regulate MDSC function via the NF-κB pathway, modulating the activities of effector T cells and Tregs.64 In ovarian cancer, miR-211 inhibits both NF-κB and STAT3 signaling through targeting CHOP, thus disrupting MDSC differentiation and immunosuppressive functions. CHOP, an ER stress-related transcription factor, regulates cellular stress responses and apoptosis pathways. Through downregulating CHOP, miR-211 impairs MDSC immunosuppression, enhancing CD4+ and CD8+ T cell antitumor responses.65,66
The HIF-1α pathway also critically modulates MDSC functions in gynecological malignancies via miRNAs. Specifically, miR-210 enhances MDSC immunosuppression by influencing HIF-1α expression, suppressing effector T cell activity, and increasing IL-10 and TGF-β production, thereby supporting immune evasion and tumor progression.67
Finally, miRNAs regulate oxidative stress pathways affecting MDSC activity. In ovarian cancer, miR-17-5p and miR-20a downregulate STAT3 and NADPH oxidase subunits (p47phox, gp91phox), decreasing ROS and H2O2 production. This reduction mitigates MDSC-mediated suppression of antigen-specific CD4+ and CD8+ T cells, impeding tumor advancement.68
Hematologic Tumor
In lymphoma, the upregulated expression of miR-30a enhances the immunosuppressive capabilities of MDSCs and inhibits T cell function by increasing the secretion of immunosuppressive molecules, including Arg-1 and IL-10, thereby facilitating tumor immune evasion.69 Similarly, miR-21 and miR-155 intensify MDSCs functions through the activation of multiple signaling pathways such as TGF-β, IL-6, and JUN, further promoting immune suppression in the tumor microenvironment. Conversely, miR-130b influences MDSCs activity via the IGF1 signaling pathway, aiding in immune suppression and tumor advancement. Additionally, decreased miR-28 expression is linked to reduced MDSCs numbers and diminished immunosuppressive function, suggesting its critical role in modulating immune responses.70
Lung Cancer
Within lung cancer, miR-300 facilitates tumor immune evasion and progression by regulating the KLF9/GADD34 signaling axis, thereby promoting the expansion and immunosuppressive phenotype of MDSCs in the TME.71 Additionally, the receptor complex LILRB4/gp49B strengthens MDSCs immunosuppressive functions by suppressing miR-1 family expression, thus favoring MDSC differentiation into the immunosuppressive M2 subtype.72 MDSC-derived exosomal miR-126a critically modulates the TME, significantly enhancing MDSC-mediated immunosuppression and promoting tumor growth.73 Furthermore, miR-143-3p boosts MDSCs immunosuppressive activity by targeting the tumor suppressor gene ITM2B, resulting in the activation of PI3K/Akt signaling.74 miR-21 regulates MDSCs stability and survival by downregulating SORBS1 expression, promoting MDSC accumulation and their immunosuppressive capacity in the TME.75 Finally, miR-21a enhances MDSC-driven immune suppression through suppression of PDCD4, increasing autocrine IL-6 production and subsequent activation of STAT3 phosphorylation, further consolidating immune evasion in lung cancer.76
Glioma
In glioma, MSC-derived exosomes containing miR-21 significantly enhance MDSC immunosuppressive functions through activation of the PTEN/PI3K/AKT/HIF-1α signaling pathway.77,78 Tumor-derived exosomes enriched with miR-1246 induce differentiation and activation of monocytic MDSCs (M-MDSCs) via the DUSP3/ERK pathway, thus increasing their suppressive capacity.79 Under hypoxic conditions, glioma-derived exosomes (H-GDEs) carrying miR-10a and miR-21 notably enhance MDSC expansion and their immunosuppressive function. Specifically, miR-10a targets the RORA gene to activate IκBα/NF-κB signaling, while miR-21 inhibits PTEN to enhance PI3K/AKT pathway activity.78 Exosomal miR-1298-5p derived from glioma cells significantly promotes MDSC immunosuppressive functions by targeting MSH2 and activating NF-κB signaling, leading to increased NOS2 and TGF-β expression. Further experimental evidence indicates these miR-1298-5p-loaded exosomes markedly augment MDSC suppression of T cell proliferation.80 miR-29a enhances MDSCs proliferation by targeting the Hbp1 gene, whereas miR-92a augments their immunosuppressive functions by repressing Prkar1a, activating the PKA/p-STAT3 signaling cascade.81
Melanoma
In melanoma, tumor-derived exosomes carry miRNAs (including miR-146a, miR-155, miR-125b, miR-100, etc.) that target and regulate key signaling pathways such as STAT3, NF-κB, and PI3K/AKT. These miRNAs promote MDSCs expansion and activation, while suppressing the proliferation and function of effector T cells.82 In CD133+ melanoma stem cells, downregulation of miR-92 leads to upregulation of integrins αV and α5, enhancing TGF-β activation. This, in turn, increases the immunosuppressive effects of MDSCs via the SMAD2 signaling pathway.83 Under tumor conditions, activation of the CXCR2 signaling pathway significantly upregulates the expression of miR-449c, which inhibits STAT6 translation. This shift alters the differentiation balance of bone marrow progenitor cells, promoting the expansion of M-MDSCs.84
Other Tumors
In thyroid cancer, miR-486-3p has been seen as a target of the NF-κB2 gene. Downregulation of miR-486-3p contributes to the excessive activation of the NF-κB2 signaling pathway, which in turn promotes cancer cell metastasis and invasion. Restoration of miR-486-3p expression significantly inhibits PMN-MDSC-mediated activation of NF-κB2, thereby reducing thyroid cancer cell migration and invasion.85 Additionally, miR-142-3p regulates the immunosuppressive function of MDSCs by targeting the C/EBPβ pathway. Downregulation of C/EBPβ reduces the number of TAMs in fibrosarcoma, while simultaneously promoting the accumulation of M-MDSCs, thereby enhancing immune suppression.86 Under hypoxic conditions, exosomes secreted by oral squamous cell carcinoma cells exhibit significantly upregulated expression of miR-21, which promotes the expansion of MDSCs and further enhances their immunosuppressive function.87
In summary, miRNAs play a critical role in regulating the recruitment, differentiation, and immunosuppressive functions of tumor-associated MDSCs across various cancer types (Figure 4) (Table 1).
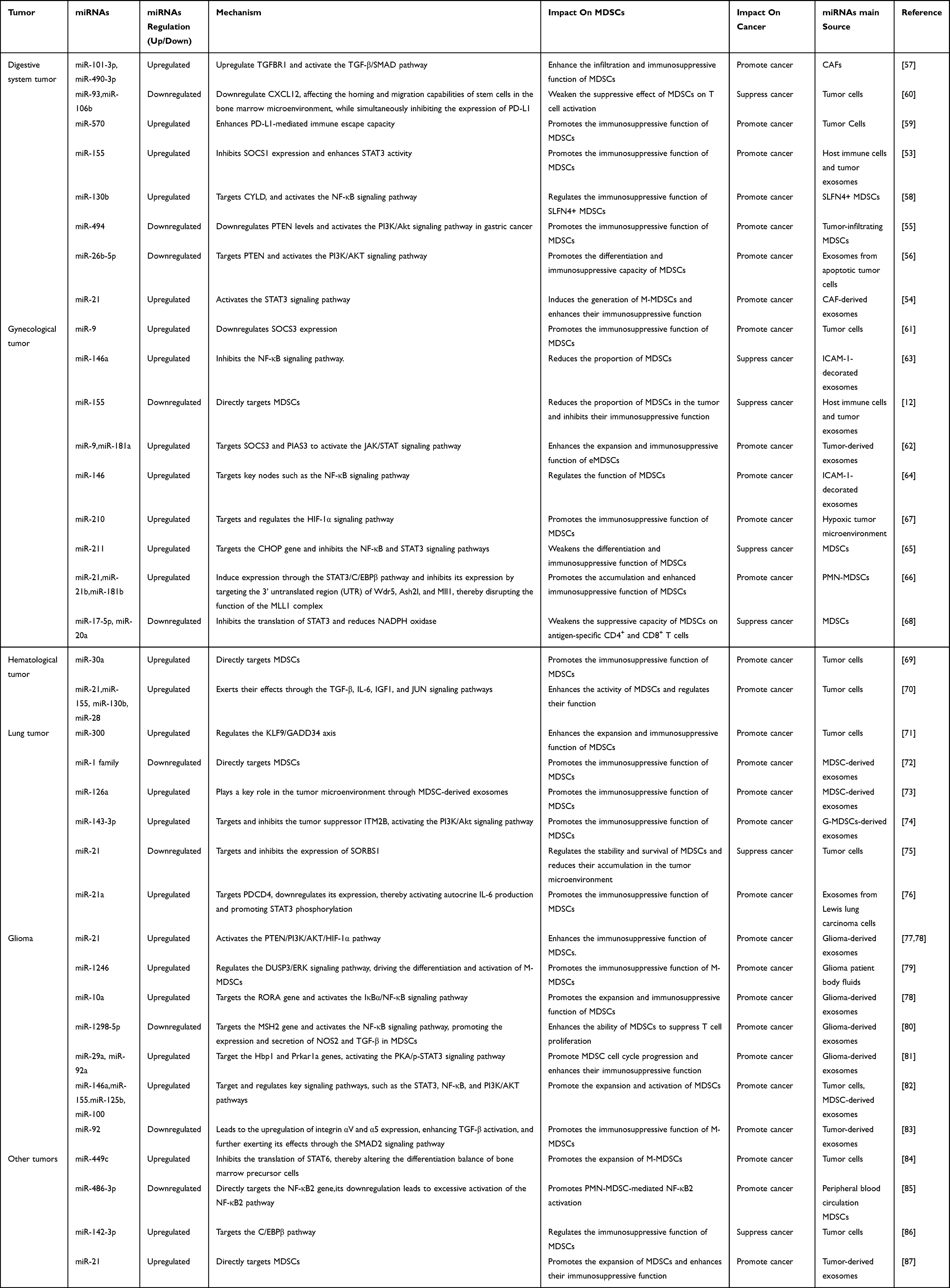 |
Table 1 Mechanisms of miRNAs Regulation of Tumor MDSCs |
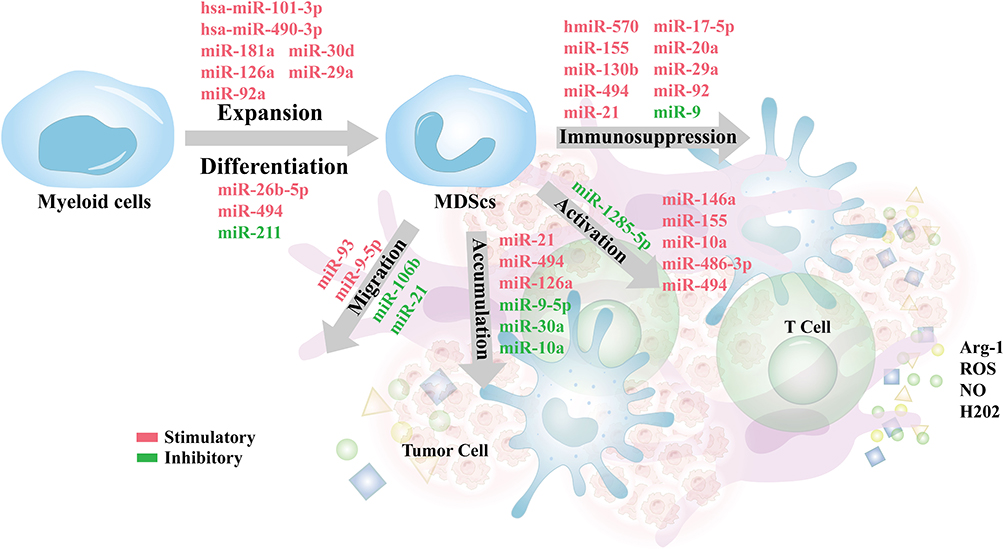 |
Figure 4 Regulatory roles of miRNAs in MDSCs expansion, differentiation, accumulation, activation, and immunosuppressive functions within the tumor microenvironment. miRNAs such as miR-101-3p, miR-490-3p, and miR-30d promote MDSCs expansion, while miR-494 and miR-211 regulate their differentiation. The accumulation of MDSCs is influenced by miRNAs including miR-21, miR-494, and miR-126a. Activation of MDSCs is driven by miRNAs such as miR-155, miR-9, and miR-146a, which enhance their immunosuppressive capacities through the production of Arg-1, ROS, NO, and H2O2, ultimately suppressing effector T cell function. |
Regulation of MDSCs by lncRNAs in Tumors
Digestive System Tumor
Recent study has shown that lncRNA Lnc-17 Rik enhances the immunosuppressive functions of MDSCs by increasing Arg-1 activity in M-MDSCs and boosting H2O2 production inPMN-MDSCs.88 Zheng et al demonstrated that Pvt1 is upregulated by HIF-1α under hypoxic conditions, thereby modulating the function and differentiation of G-MDSCs.89 MIR4435-2HG, a long non-coding RNA previously identified as oncogenic, was reported by Zhang et al to facilitate tumor initiation and progression in models of colitis-associated cancer and spontaneous intestinal adenomatous polyposis. This pro-tumorigenic effect is associated with the loss of MIR4435-2HG, which results in an increased infiltration of PMN-MDSCs into the tumor microenvironment and augments their immunosuppressive functions, rather than directly affecting cancer cell proliferation, migration, or invasion.90 Furthermore, Lnc-C/EBPβ modulates the expression of C/EBPβ LIP and WDR5, leading to the downregulation of IL4i1, which in turn impacts MDSC differentiation. The reduced IL4i1 levels may impair the immunoregulatory function of MDSCs, thereby influencing tumor immune evasion.91 Overexpression of lncRNA 57Rik not only upregulates key immunosuppressive genes, including Arg-1 and NOS2, thereby enhancing the suppressive function of MDSCs, but also promotes the release of metabolic byproducts such as Arg-1, NO, and H2O2, which inhibit T cell proliferation and function.92 Recent studies have shown that Olfr29-ps1, a lncRNA, is upregulated in MDSCs in response to the tumor-associated factor IL6. This upregulation results in the suppression of miR-214-3p expression, which leads to increased MyD88 levels. Consequently, elevated MyD88 expression enhances both the immunosuppressive function and differentiation of MDSCs.93 Studies have shown that lncRNA HOTAIR (Hox antisense intergenic RNA), a member of the HOXC gene cluster, promotes CCL2 secretion through its overexpression, which in turn induces the proliferation of monocytes and MDSCs.13 CRNDE (Colorectal Neoplasia Differentially Expressed), a long non-coding RNA implicated in tumor promotion, has been detected across multiple cancer types. Li et al revealed that CRNDE upregulates CXCL3 expression, leading to the recruitment of G-MDSCs into the TME, which subsequently suppresses CD8+ T cell infiltration and function.94
Lung Cancer
Studies have shown that lncRNA RUNXOR is upregulated in the peripheral blood of lung cancer patients, while its target gene RUNX1 is downregulated in these patients. Knockdown of RUNXOR reduces Arg1 expression in MDSCs.95 Another Study has shown that lncRNA Snhg6 promotes the differentiation of CD11b+Ly6G−Ly6Chigh M-MDSCs, while having no effect on the differentiation of CD11b+Ly6G+Ly6Clow PMN-MDSCs.96 In the tumor microenvironment, lncRNA AK036396 is highly expressed in PMN-MDSCs. Further investigation reveals that silencing lncRNA AK036396 significantly reduces the immunosuppressive functions of PMN-MDSCs, including decreased Arg1 activity and ROS production, while promoting their maturation. Additionally, lncRNA AK036396 directly interacts with the Fcnb protein, and its silencing destabilizes Fcnb, thereby further reducing the immunosuppressive capacity of PMN-MDSCs. Furthermore, silencing Fcnb also attenuates the immunosuppressive function of PMN-MDSCs and delays tumor progression.97 HOTAIRM1 (HOXA Transcript Antisense RNA, Myeloid-Specific 1) is a myeloid-specific lncRNA located between the HOXA1 and HOXA2 genes in humans. Research by Tian et al demonstrated that HOTAIRM1 expression is significantly downregulated in the tumor tissues of lung cancer patients. Overexpression of HOTAIRM1 inhibits the immunosuppressive functions of MDSCs. Furthermore, HOTAIRM1 regulates MDSC function by modulating HOXA1 expression, thereby affecting the immune microenvironment and tumor progression in lung cancer.98
Other Tumors
Beyond the aforementioned cancers, lncRNAs have also been shown to regulate MDSCs in other tumors. Notably, inhibition of Malat1 in the primary TME of triple-negative breast cancer (TNBC) has been reported to promote a more immune-stimulatory environment. This is achieved by reducing the immunosuppressive effects of TAMs and MDSCs on T cells, thereby enhancing T cell responses.99 Another study emphasized that lnc-chop promotes the expression of key enzymes associated with the immunosuppressive function of MDSCs, including ARG-1, NOS2, NOX2, and COX-2, thereby playing a pivotal role in tumor and inflammatory microenvironments.100 The expression of PROM1, CCAT1, HOTAIR, and MUC19 has been reported to be closely associated with the recruitment of MDSCs in HPV-positive head and neck squamous cell carcinoma (HNSCC).101 Recent studies have identified a novel lncRNA, LncOVM, which forms a complex with the protein PPIP5K2. This complex promotes the proliferation and metastasis of ovarian cancer cells. Notably, PPIP5K2 plays a role in regulating the secretion of complement C5, which in turn influences tumor microenvironment remodeling and facilitates the recruitment of neutrophil- MDSCs.102 Research has shown that Lnc-H19, via its encoded protein H19-IRP, facilitates the formation of an immunosuppressive microenvironment, thereby promoting the proliferation of GBM cells and their immune evasion. Additionally, H19-IRP enhances the accumulation of MDSCs by regulating the transcription of CCL2 and Galectin-9, which exacerbates tumor-induced immune suppression and drives tumor metastasis and growth.103 Studies have shown that Lnc-AGAP2-AS1 exacerbates the immunosuppressive effects within the TME by regulating the secretion of TGF-β1 from MDSCs. This, in turn, inhibits effector T cell function and promotes tumor metastasis.104 In summary, lncRNAs play a pivotal role in regulating tumor-associated MDSCs across various cancers, and are closely linked to tumor initiation, progression, and prognosis (Figure 5). This section reviews the current research on the involvement of lncRNAs in modulating MDSCs in tumors, as summarized in Table 2.
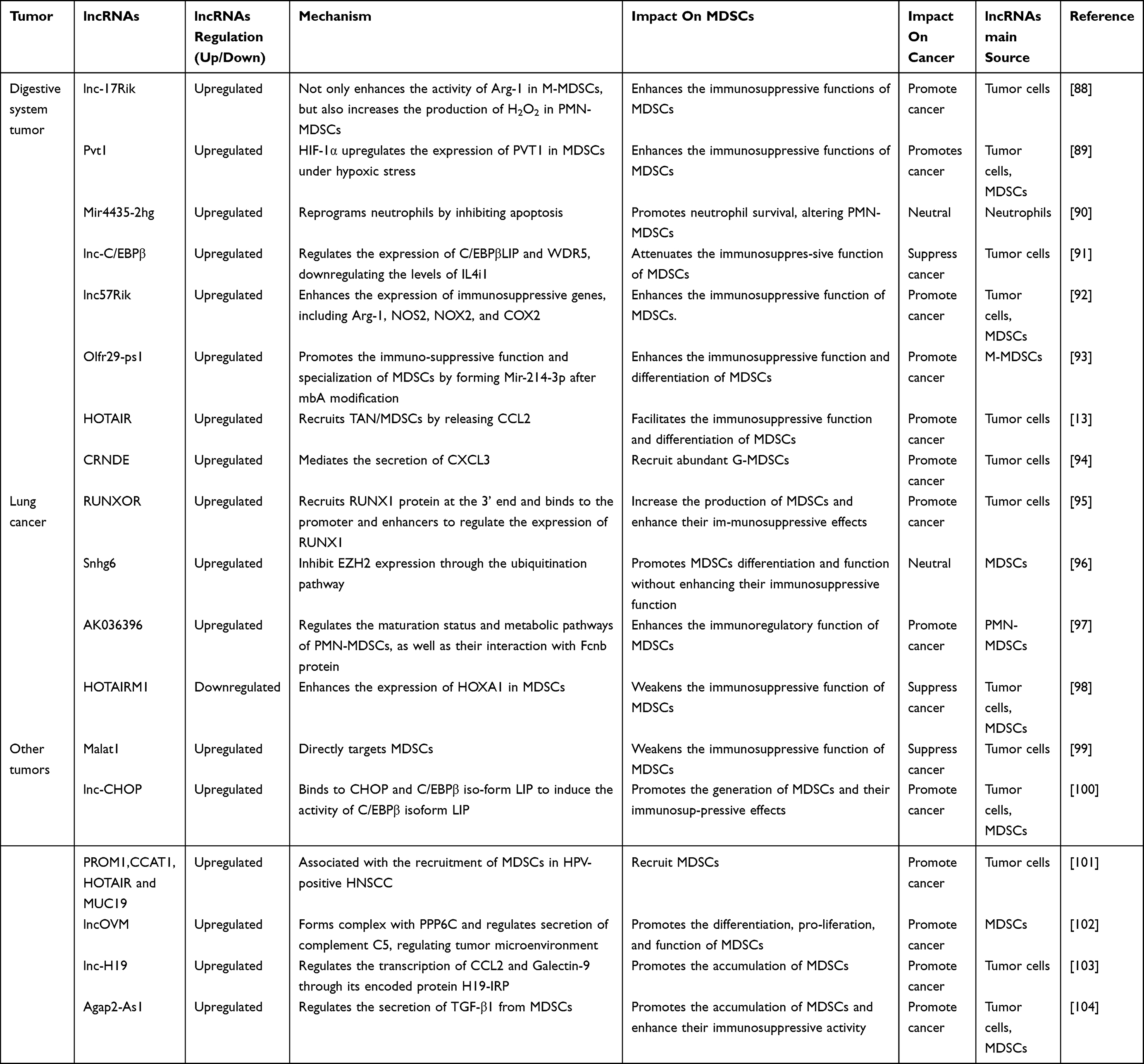 |
Table 2 Mechanisms of lncRNAs Regulation of Tumor MDSCs |
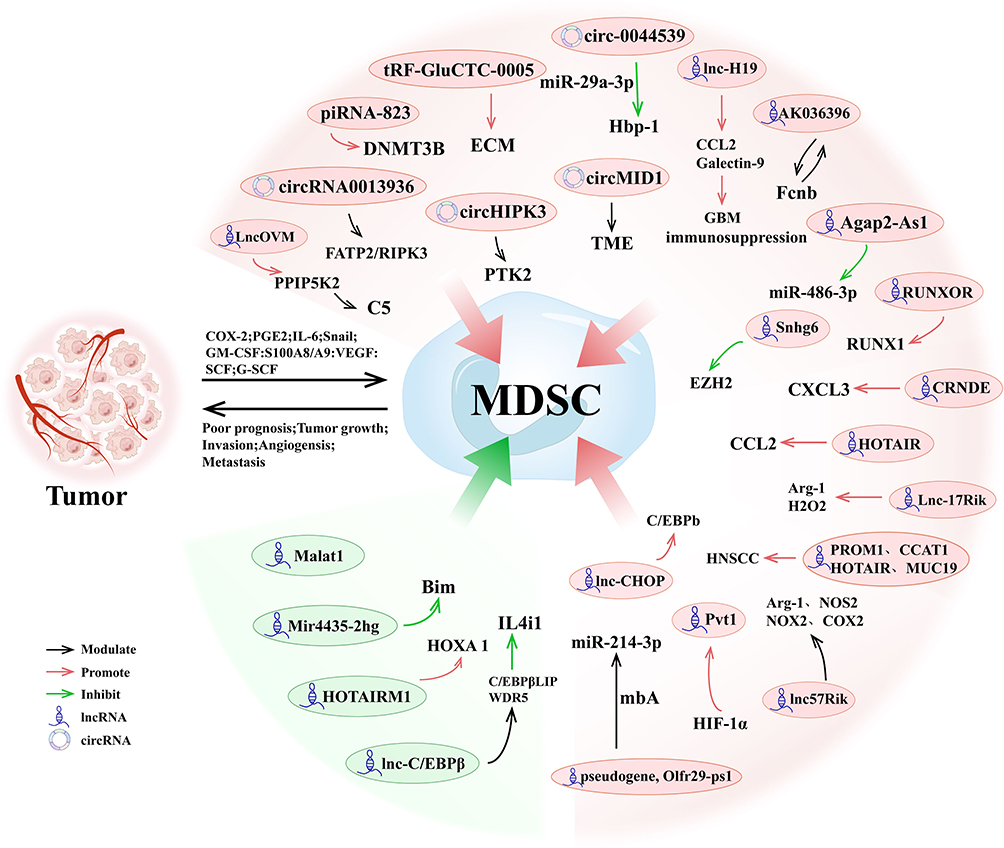 |
Figure 5 Regulation of MDSCs by ncRNAs in Tumors. This figure illustrates the roles of various ncRNAs in regulating MDSCs and promoting tumor immune evasion within the tumor microenvironment. Key ncRNAs, including lncRNAs, circRNAs, miRNAs, and piRNAs, modulate MDSC activation, differentiation, and immunosuppressive functions via pathways such as FAT2/RIPK3 (circHIPK3), CXCL3/CCL2 (HOTAIR), and Hbp-1 inhibition (miR-29a-3p). |
Regulation of MDSCs by Other ncRNAs in Tumors
Digestive System Tumor
Circ-0044539 modulates the immunosuppressive activity of MDSCs through exosomal signaling and plays a pivotal role in lymph node metastasis of hepatocellular carcinoma. It exerts its effects by targeting miR-29a-3p to inhibit Hbp-1, which enhances the levels of Arg1 and NO in PMN-MDSCs and suppresses T cell proliferation, thereby establishing an immunosuppressive pre-metastatic lymph node microenvironment.105 tRF-GluCTC-0005 activates hepatic stellate cells via exosomes, upregulating WDR1 expression and promoting the production of ECM components and the secretion of factors such as IL-6. This process recruits MDSCs and establishes an immunosuppressive pre-metastatic niche, thereby facilitating liver metastasis of pancreatic cancer.106
Urinary System Tumor
Exosomes derived from bladder cancer containing circRNA_0013936 have been shown to enhance the immunosuppressive activity of PMN-MDSCs by modulating the expression of fatty acid transporter protein 2 (FATP2) and receptor-interacting protein kinase 3 (RIPK3).107 MDSCs transfer the S100A9 protein via exosomes, promoting the generation of circMID1 in prostate cancer. Through the circMID1/miR-506-3p/MID1 axis, MDSCs regulate the tumor microenvironment, enhancing their immunosuppressive activity and accelerating tumor progression.108
Other Tumors
The circHIPK3/PTK2 signaling axis facilitates the conversion of M-MDSCs into M2 macrophages in lung cancer, thereby strengthening the immunosuppressive characteristics of the tumor microenvironment.109 In the multiple myeloma tumor microenvironment, G-MDSCs amplify the immunosuppressive function of MDSCs by inducing the expression of piRNA-823. This piRNA activates DNMT3B and triggers widespread DNA methylation, indirectly maintaining MDSC-mediated suppression and facilitating immune evasion.110 This section summarizes current research on the roles of other ncRNAs in modulating MDSCs in tumors, as presented in Table 3.
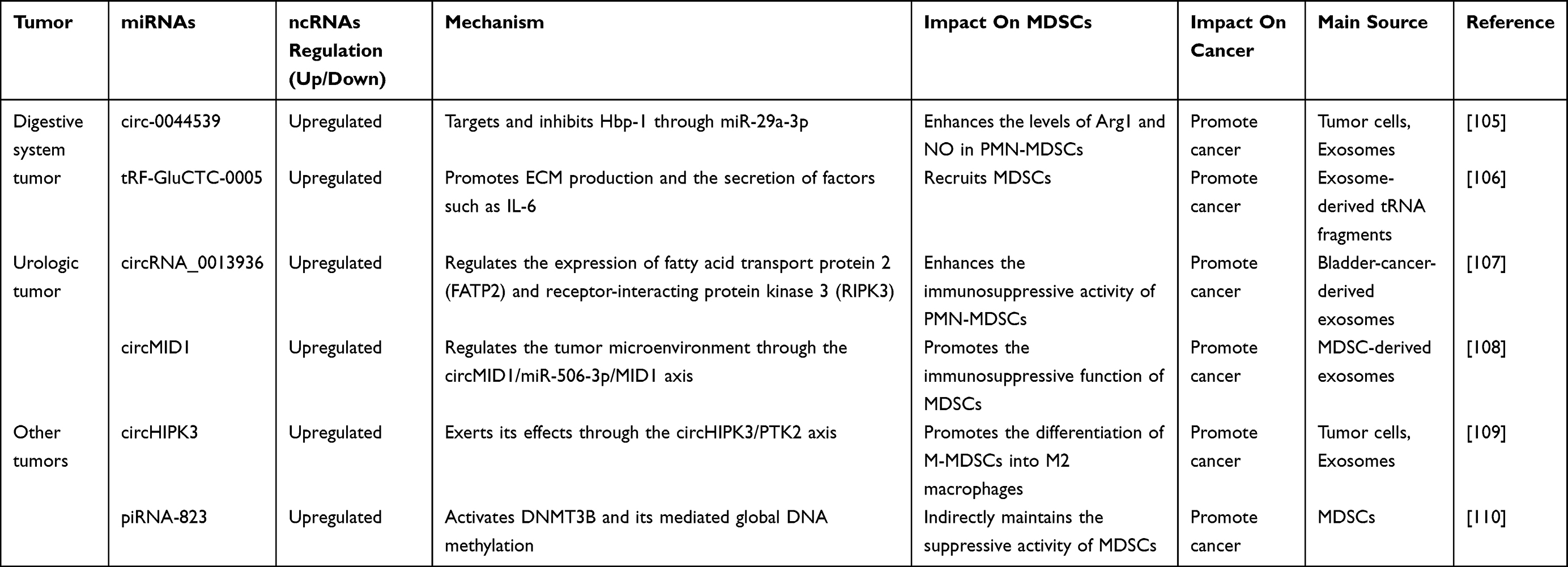 |
Table 3 The Mechanisms of Other ncRNAs Involvement in Regulating Tumor MDSCs |
Therapeutic Strategies Targeting ncRNAs in MDSCs
Targeting oncogenic ncRNAs with antisense oligonucleotides (ASOs) or small molecule inhibitors represents a promising therapeutic strategy.111,112 For instance, targeting HOTAIR or circHIPK3 can diminish MDSC-driven immunosuppression and bolster antitumor immune responses.109,113
In addition to suppressing oncogenic ncRNAs, delivering synthetic mimics of tumor-suppressive ncRNAs holds potential for reprogramming MDSCs into a less suppressive phenotype. This approach can complement existing therapies by reducing the immunosuppressive barrier created by MDSCs.111,114
Integrating ncRNA-targeted strategies with immune checkpoint blockade, such as anti-PD-1/PD-L1 therapies, can potentiate antitumor immune responses.Additionally, integrating these strategies with chemotherapies or targeted therapies, such as CSF1R inhibitors, can further disrupt MDSC activity and improve treatment efficacy.112,115
To achieve effective delivery, novel platforms such as nanoparticle-based systems have been developed to specifically target ncRNAs in MDSCs. For instance, dual-responsive PEG-lipid polyester nanoparticles enable precise siRNA delivery to the TME, reducing MDSC-mediated immunosuppression while enhancing T cell activation.114 These advances highlight the potential of ncRNA-targeting strategies as a transformative approach to modulate MDSCs activity and improve the outcomes of cancer immunotherapy.
Conclusion
Recent studies have notably enhanced the comprehension of how ncRNAs regulate MDSCs within the tumor microenvironment (TME). Various ncRNAs, such as miRNAs, lncRNAs, and circRNAs, influence MDSC recruitment, differentiation, and immunosuppressive activities by modulating essential signaling pathways, including STAT3, NF-κB, and PI3K/AKT, thus promoting immune escape and tumor progression. Moreover, ncRNAs impact tumor-immune interactions via processes such as histone modification, chromatin remodeling, and modulation of miRNA functions, thereby intensifying immune suppression and facilitating tumor growth.
MDSC regulation involves intricate networks comprising cytokines, chemokines, and growth factors secreted by tumor cells as well as immune cells. Collectively, these molecules strengthen the immunosuppressive environment, accelerating tumor advancement. Additionally, Tregs interact with MDSCs, further consolidating immune suppression.
Targeting oncogenic ncRNAs through antisense oligonucleotides (ASOs) or small molecule inhibitors represents a promising therapeutic approach to disrupting MDSC-induced immune suppression. Additionally, strategies aiming to reprogram MDSCs into less immunosuppressive phenotypes using synthetic mimics of tumor-suppressive ncRNAs could complement existing cancer treatments. Integrating these interventions with immune checkpoint blockade therapies, such as anti-PD-1/PD-L1 antibodies, or targeted agents like CSF1R inhibitors, might enhance anti-tumor immune responses and clinical efficacy. Collectively, ncRNAs converge on a limited set of myeloid programs-prominently STAT3, NF-κB, and PI3K/AKT-to influence MDSC specification, maintenance, and suppressive function. Mechanistically, epigenetic writers/readers (eg, EZH2), post-transcriptional control (miRNA sponging), and exosome-mediated transfer act in combination and are context dependent across tumor types. Therapeutically, ncRNA-directed strategies (sense/antisense, siRNA, miRNA mimics, and delivery via nanoparticles or exosomes) are promising adjuncts to immunotherapy but face challenges in cell-type–specific delivery, durability, and off-target effects. We outline testable priorities: (i) pathway-centric triage, comparing JAK/STAT pathway-dominant tumors, which are often associated with immune dysregulation and tumor progression, with TGF-β pathway-dominant tumors, which are frequently linked to immune suppression and tumor metastasis; (ii) cell-selective delivery platforms for MDSCs; (iii) paired biomarker panels (circulating ncRNAs and exosomal ncRNAs associated with myeloid states); and (iv) rigorous cross-species standards for MDSC definition. In line with the synthesis above, future work should prioritize pathway-centric triage, MDSC-selective delivery platforms for ncRNA agents, paired biomarker panels integrating circulating/exosomal ncRNAs with myeloid states, and cross-species standards for MDSC definition.
A compelling example of the challenges that lie ahead is the paradoxical nature of key regulators like miR-155. While widely explored as a therapeutic agent due to its immunostimulatory properties—such as enhancing dendritic cell maturation—its net effect in the tumor microenvironment can be profoundly pro-tumorigenic. Seminal studies have revealed that this is because the dominant role of miR-155 in supporting MDSCs and Tregs can outweigh its beneficial anti-tumor functions. This functional dichotomy, where a single ncRNA can be both friend and foe, underscores the complexity of targeting these pathways.
Therefore, despite the promising developments outlined above, several critical challenges remain. These include not only further clarifying the context-dependent regulatory mechanisms of ncRNAs in MDSCs, but also improving delivery specificity and overcoming barriers for effective clinical translation of RNA-based treatments. Continuous research into ncRNA mechanisms and advances in targeted therapeutic strategies hold considerable promise for enhancing the efficacy of cancer immunotherapy.
Abbreviations
ncRNAs, non-coding RNAs; miRNAs, microRNA; lncRNAs, long non-coding RNAs; circRNAs, circular RNAs; piRNAs, PIWI-interacting RNAs; MDSCs, myeloid-derived suppressor cells; M-MDSCs, monocytic MDSCs; G-MDSCs, granulocytic MDSCs; PMN-MDSCs, polymorphonuclearMDSCs; eMDSCs, early-stage myeloid-derived suppressor cells; TME, tumor microenvironment; Tregs, regulatory T cells; TAMs, tumor-associated macrophages; CAFs, cancer-associated fibroblasts; ceRNA, competitive endogenous RNA; SOCS3, Suppressor of Cytokine Signaling 3; TNF-α, tumor necrosis factor-alpha; ECM, extracellular matrix; MMP, matrix metalloproteinases.
Funding
Jiangsu Provincial Key Laboratory of Clinical Laboratory Medicine 2024 Open Fund Program, Grant/Award Number: JSKLM-Y-2024-010; Jiangsu Provincial Health Commission Medical Research General Program, Grant/Award Numbers: M2022100, M2022117; Jiangsu Provincial Natural Science Foundation General Program, Grant/Award Number: BK20231250; the Social Development Foundation of Zhenjiang (SH2023001).
Disclosure
The authors declare no conflicts of interest in this work.
References
1. Bray F, Laversanne M, Sung H, et al. Global cancer statistics 2022: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J Clin. 2024;74(3):229–263. doi:10.3322/caac.21834
2. Gabrilovich DI, Nagaraj S. Myeloid-derived suppressor cells as regulators of the immune system. Nat Rev Immunol. 2009;9(3):162–174. doi:10.1038/nri2506
3. Veglia F, Sanseviero E, Gabrilovich DI. Myeloid-derived suppressor cells in the era of increasing myeloid cell diversity. Nat Rev Immunol. 2021;21(8):485–498. doi:10.1038/s41577-020-00490-y
4. Hegde S, Leader AM, Merad M. MDSC: markers, development, states, and unaddressed complexity. Immunity. 2021;54(5):875–884. doi:10.1016/j.immuni.2021.04.004
5. van Vlerken-Ysla L, Tyurina YY, Kagan VE, Gabrilovich DI. Functional states of myeloid cells in cancer. Cancer Cell. 2023;41(3):490–504. doi:10.1016/j.ccell.2023.02.009
6. Wang T, Wang J, Jiang H, et al. Targeted regulation of tumor microenvironment through the inhibition of MDSCs by curcumin loaded self-assembled nano-filaments. Mater Today Bio. 2022;15:100304. doi:10.1016/j.mtbio.2022.100304
7. Slack FJ, Chinnaiyan AM. The role of non-coding RNAs in oncology. Cell. 2019;179(5):1033–1055. doi:10.1016/j.cell.2019.10.017
8. Nemeth K, Bayraktar R, Ferracin M, Calin GA. Non-coding RNAs in disease: from mechanisms to therapeutics. Nat Rev Genet. 2024;25(3):211–232. doi:10.1038/s41576-023-00662-1
9. Zhang L, Xu X, Su X. Noncoding RNAs in cancer immunity: functions, regulatory mechanisms, and clinical application. Mol Cancer. 2020;19(1):48. doi:10.1186/s12943-020-01154-0
10. Liu SJ, Dang HX, Lim DA, Feng FY, Maher CA. Long noncoding RNAs in cancer metastasis. Nat Rev Cancer. 2021;21(7):446–460. doi:10.1038/s41568-021-00353-1
11. Liu X, Zhao S, Sui H, et al. MicroRNAs/LncRNAs Modulate MDSCs in Tumor Microenvironment. Front Oncol. 2022;12:772351. doi:10.3389/fonc.2022.772351
12. Li K, Wang J, Xie Y, et al. Reactive oxygen species/glutathione dual sensitive nanoparticles with encapsulation of miR155 and curcumin for synergized cancer immunotherapy. J Nanobiotechnology. 2024;22(1):400. doi:10.1186/s12951-024-02575-5
13. Fujisaka Y, Iwata T, Tamai K, et al. Long non-coding RNA HOTAIR up-regulates chemokine (C-C motif) ligand 2 and promotes proliferation of macrophages and myeloid-derived suppressor cells in hepatocellular carcinoma cell lines. Oncol Lett. 2018;15(1):509–514. doi:10.3892/ol.2017.7322
14. Liu Y, Han Y, Zhang Y, Lv T, Peng X, Huang J. LncRNAs has been identified as regulators of Myeloid-derived suppressor cells in lung cancer. Front Immunol. 2023;14:1067520. doi:10.3389/fimmu.2023.1067520
15. Xia Y, Wang D, Piao Y, et al. Modulation of immunosuppressive cells and noncoding RNAs as immunotherapy in osteosarcoma. Front Immunol. 2022;13:1025532. doi:10.3389/fimmu.2022.1025532
16. Tang XH, Guo T, Gao XY, et al. Exosome-derived noncoding RNAs in gastric cancer: functions and clinical applications. Mol Cancer. 2021;20(1):99. doi:10.1186/s12943-021-01396-6
17. Wang H, Zhou F, Qin W, Yang Y, Li X, Liu R. Metabolic regulation of myeloid-derived suppressor cells in tumor immune microenvironment: targets and therapeutic strategies. Theranostics. 2025;15(6):2159–2184. doi:10.7150/thno.105276
18. Nakamura K, Smyth MJ. Myeloid immunosuppression and immune checkpoints in the tumor microenvironment. Cell Mol Immunol. 2020;17(1):1–12. doi:10.1038/s41423-019-0306-1
19. Wu Y, Yi M, Niu M, Mei Q, Wu K. Myeloid-derived suppressor cells: an emerging target for anticancer immunotherapy. Mol Cancer. 2022;21(1):184. doi:10.1186/s12943-022-01657-y
20. Ruffolo LI, Jackson KM, Kuhlers PC, et al. GM-CSF drives myelopoiesis, recruitment and polarisation of tumour-associated macrophages in cholangiocarcinoma and systemic blockade facilitates antitumour immunity. Gut. 2022;71(7):1386–1398. doi:10.1136/gutjnl-2021-324109
21. Calì B, Agnellini AHR, Cioccarelli C, et al. GM-CSF nitration is a new driver of myeloid suppressor cell activity in tumors. Front Immunol. 2021;12:718098. doi:10.3389/fimmu.2021.718098
22. Wang L, Zhu L, Liang C, et al. Targeting N6-methyladenosine reader YTHDF1 with siRNA boosts antitumor immunity in NASH-HCC by inhibiting EZH2-IL-6 axis. J Hepatol. 2023;79(5):1185–1200. doi:10.1016/j.jhep.2023.06.021
23. Sun R, Sun Y, Wu C, et al. CXCR4-modified CAR-T cells suppresses MDSCs recruitment via STAT3/NF-κB/SDF-1α axis to enhance efficacy against pancreatic cancer. Mol Ther. 2023;31(11):3193–3209. doi:10.1016/j.ymthe.2023.09.010
24. Liu NN, Yi CX, Wei LQ, et al. The intratumor mycobiome promotes lung cancer progression via myeloid-derived suppressor cells. Cancer Cell. 2023;41(11):1927–44.e9. doi:10.1016/j.ccell.2023.08.012
25. Dong P, Yan Y, Fan Y, et al. The role of myeloid-derived suppressor cells in the treatment of pancreatic cancer. Technol Cancer Res Treat. 2022;21:15330338221142472. doi:10.1177/15330338221142472
26. Siemińska I, Baran J. Myeloid-derived suppressor cells as key players and promising therapy targets in prostate cancer. Front Oncol. 2022;12:862416. doi:10.3389/fonc.2022.862416
27. Ostrand-Rosenberg S, Lamb TJ, Pawelec G. Here, there, and everywhere: myeloid-derived suppressor cells in immunology. J Immunol. 2023;210(9):1183–1197. doi:10.4049/jimmunol.2200914
28. Lv J, Zhao Y, Zong H, Ma G, Wei X, Zhao Y. Increased levels of circulating monocytic- and early-stage Myeloid-Derived Suppressor Cells (MDSC) in acute myeloid leukemia. Clin Lab. 2021;67(3). doi:10.7754/Clin.Lab.2020.200719
29. He S, Zheng L, Qi C. Myeloid-derived suppressor cells (MDSCs) in the tumor microenvironment and their targeting in cancer therapy. Mol Cancer. 2025;24(1):5. doi:10.1186/s12943-024-02208-3
30. Wang Y, Zhang C, Liu T, et al. Malat1 regulates PMN-MDSC expansion and immunosuppression through p-STAT3 ubiquitination in sepsis. Int J Biol Sci. 2024;20(4):1529–1546. doi:10.7150/ijbs.92267
31. Zhang K, Zakeri A, Alban T, et al. Vista promotes the metabolism and differentiation of myeloid-derived suppressor cells by STAT3 and polyamine-dependent mechanisms. Cell Rep. 2024;43(1):113661. doi:10.1016/j.celrep.2023.113661
32. Liu X, Tang R, Xu J, et al. CRIP1 fosters MDSC trafficking and resets tumour microenvironment via facilitating NF-κB/p65 nuclear translocation in pancreatic ductal adenocarcinoma. Gut. 2023;72(12):2329–2343. doi:10.1136/gutjnl-2022-329349
33. Zhang M, Wang L, Liu W, et al. Targeting inhibition of accumulation and function of myeloid-derived suppressor cells by artemisinin via PI3K/AKT, mTOR, and MAPK pathways enhances anti-pd-l1 immunotherapy in melanoma and liver tumors. J Immunol Res. 2022;2022:2253436. doi:10.1155/2022/2253436
34. Lim HX, Kim TS, Poh CL. Understanding the differentiation, expansion, recruitment and suppressive activities of myeloid-derived suppressor cells in cancers. Int J Mol Sci. 2020;21(10):3599. doi:10.3390/ijms21103599
35. Li BH, Garstka MA, Li ZF. Chemokines and their receptors promoting the recruitment of myeloid-derived suppressor cells into the tumor. Mol Immunol. 2020;117:201–215. doi:10.1016/j.molimm.2019.11.014
36. Kumar V, Cheng P, Condamine T, et al. CD45 Phosphatase Inhibits STAT3 transcription factor activity in myeloid cells and promotes tumor-associated macrophage differentiation. Immunity. 2016;44(2):303–315. doi:10.1016/j.immuni.2016.01.014
37. Tesi RJ. MDSC; the most important cell you have never heard of. Trends Pharmacol Sci. 2019;40(1):4–7. doi:10.1016/j.tips.2018.10.008
38. Tannenbaum CS, Rayman PA, Pavicic PG, et al. Mediators of inflammation-driven expansion, trafficking, and function of tumor-infiltrating MDSCs. Cancer Immunol Res. 2019;7(10):1687–1699. doi:10.1158/2326-6066.CIR-18-0578
39. Zhang W, Jiang M, Chen J, et al. SOCS3 Suppression Promoted the Recruitment of CD11b(+)Gr-1(-)F4/80(-)MHCII(-) Early-Stage Myeloid-Derived Suppressor Cells and Accelerated Interleukin-6-Related Tumor Invasion via Affecting Myeloid Differentiation in Breast Cancer. Front Immunol. 2018;9:1699. doi:10.3389/fimmu.2018.01699
40. Ducarouge B, Redavid AR, Victoor C, et al. Netrin-1 blockade inhibits tumor associated Myeloid-derived suppressor cells, cancer stemness and alleviates resistance to chemotherapy and immune checkpoint inhibitor. Cell Death Differ. 2023;30(10):2201–2212. doi:10.1038/s41418-023-01209-x
41. Lee YS, Saxena V, Bromberg JS, Scalea JR. G-CSF promotes alloregulatory function of MDSCs through a c-Kit dependent mechanism. Cell Immunol. 2021;364:104346. doi:10.1016/j.cellimm.2021.104346
42. Wang C, Zheng X, Zhang J, et al. CD300ld on neutrophils is required for tumour-driven immune suppression. Nature. 2023;621(7980):830–839. doi:10.1038/s41586-023-06511-9
43. Vetter C, Schieb J, Vedder N, et al. The impact of IL-10 and IL-17 on myeloid-derived suppressor cells in vitro and in vivo in a murine model of asthma. Eur J Immunol. 2024;54(7):e2350785. doi:10.1002/eji.202350785
44. Zeng X, Liao G, Li S, et al. Eliminating METTL1-mediated accumulation of PMN-MDSCs prevents hepatocellular carcinoma recurrence after radiofrequency ablation. Hepatology. 2023;77(4):1122–1138. doi:10.1002/hep.32585
45. Sayyadioskoie SR, Schwacha MG. Myeloid-Derived Suppressor Cells (MDSCs) and the Immunoinflammatory Response to Injury (Mini Review). Shock. 2021;56(5):658–666. doi:10.1097/SHK.0000000000001795
46. Hofer F, Di Sario G, Musiu C, Sartoris S, De Sanctis F, Ugel S. A complex metabolic network confers immunosuppressive functions to Myeloid-Derived Suppressor Cells (MDSCs) within the tumour microenvironment. Cells. 2021;10(10):2700. doi:10.3390/cells10102700
47. Ostrand-Rosenberg S, Sinha P, Beury DW, Clements VK. Cross-talk between myeloid-derived suppressor cells (MDSC), macrophages, and dendritic cells enhances tumor-induced immune suppression. Semin Cancer Biol. 2012;22(4):275–281. doi:10.1016/j.semcancer.2012.01.011
48. Davoodzadeh Gholami M, Kardar GA, Saeedi Y, Heydari S, Garssen J, Falak R. Exhaustion of T lymphocytes in the tumor microenvironment: significance and effective mechanisms. Cell Immunol. 2017;322:1–14. doi:10.1016/j.cellimm.2017.10.002
49. Liu H, Zeng X, Ren X, et al. Targeting tumour-intrinsic N(7)-methylguanosine tRNA modification inhibits MDSC recruitment and improves anti-PD-1 efficacy. Gut. 2023;72(8):1555–1567. doi:10.1136/gutjnl-2022-327230
50. Hack SP, Zhu AX, Wang Y. Augmenting Anticancer Immunity Through Combined Targeting of Angiogenic and PD-1/PD-L1 Pathways: challenges and Opportunities. Front Immunol. 2020;11:598877. doi:10.3389/fimmu.2020.598877
51. Tian D, Yu Y, Zhang L, Sun J, Jiang W. 23-hydroxybetulinic acid reduces tumorigenesis, metastasis and immunosuppression in a mouse model of hepatocellular carcinoma via disruption of the MAPK signaling pathway. Anticancer Drugs. 2022;33(9):815–825. doi:10.1097/CAD.0000000000001325
52. Li J, Shu X, Xu J, et al. S100A9-CXCL12 activation in BRCA1-mutant breast cancer promotes an immunosuppressive microenvironment associated with resistance to immunotherapy. Nat Commun. 2022;13(1):1481. doi:10.1038/s41467-022-29151-5
53. Chen S, Wang L, Fan J, et al. Host miR155 promotes tumor growth through a myeloid-derived suppressor cell-dependent mechanism. Cancer Res. 2015;75(3):519–531. doi:10.1158/0008-5472.CAN-14-2331
54. Zhao Q, Huang L, Qin G, et al. Cancer-associated fibroblasts induce monocytic myeloid-derived suppressor cell generation via IL-6/exosomal miR-21-activated STAT3 signaling to promote cisplatin resistance in esophageal squamous cell carcinoma. Cancer Lett. 2021;518:35–48. doi:10.1016/j.canlet.2021.06.009
55. Moaaz M, Lotfy H, Elsherbini B, Motawea MA, Fadali G. TGF-β enhances the anti-inflammatory effect of tumor- infiltrating CD33+11b+HLA-DR myeloid-derived suppressor cells in gastric cancer: a possible relation to MicroRNA-494. Asian Pac J Cancer Prev. 2020;21(11):3393–3403. doi:10.31557/APJCP.2020.21.11.3393
56. Yin X, Tian M, Zhang J, et al. MiR-26b-5p in small extracellular vesicles derived from dying tumor cells after irradiation enhances the metastasis promoting microenvironment in esophageal squamous cell carcinoma. Cancer Lett. 2022;541:215746. doi:10.1016/j.canlet.2022.215746
57. Eun JW, Ahn HR, Baek GO, et al. Aberrantly expressed micrornas in cancer-associated fibroblasts and their target oncogenic signatures in hepatocellular carcinoma. Int J Mol Sci. 2023;24(5):4272. doi:10.3390/ijms24054272
58. Ding L, Li Q, Chakrabarti J, et al. MiR130b from Schlafen4(+) MDSCs stimulates epithelial proliferation and correlates with preneoplastic changes prior to gastric cancer. Gut. 2020;69(10):1750–1761. doi:10.1136/gutjnl-2019-318817
59. Tang W, Liu H, Li X, et al. Efficacy of zinc carnosine in the treatment of colorectal cancer and its potential in combination with immunotherapy in vivo. Aging. 2022;14(21):8688–8699. doi:10.18632/aging.204380
60. Cioffi M, Trabulo SM, Vallespinos M, et al. The miR-25-93-106b cluster regulates tumor metastasis and immune evasion via modulation of CXCL12 and PD-L1. Oncotarget. 2017;8(13):21609–21625. doi:10.18632/oncotarget.15450
61. Gu X, Wei F, Tong J, et al. MiR-9 promotes G-MDSC recruitment and tumor proliferation by targeting SOCS3 in breast cancer. FASEB j. 2024;38(1):e23388. doi:10.1096/fj.202301764RR
62. Jiang M, Zhang W, Zhang R, et al. Cancer exosome-derived miR-9 and miR-181a promote the development of early-stage MDSCs via interfering with SOCS3 and PIAS3 respectively in breast cancer. Oncogene. 2020;39(24):4681–4694. doi:10.1038/s41388-020-1322-4
63. Duarte-Sanmiguel S, Salazar-Puerta AI, Panic A, et al. ICAM-1-decorated extracellular vesicles loaded with miR-146a and Glut1 drive immunomodulation and hinder tumor progression in a murine model of breast cancer. Biomater Sci. 2023;11(20):6834–6847. doi:10.1039/D3BM00573A
64. Geis-Asteggiante L, Belew AT, Clements VK, et al. Differential Content of Proteins, mRNAs, and miRNAs Suggests that MDSC and their exosomes may mediate distinct immune suppressive functions. J Proteome Res. 2018;17(1):486–498. doi:10.1021/acs.jproteome.7b00646
65. Zheng LE, Huang M, Ye Y, Sun P. MicroRNA-211 regulates proliferation, expansion, and immune inhibitory function of myeloid-derived suppressor cells via mediation of CHOP expression. Immunol Invest. 2023;52(5):616–634. doi:10.1080/08820139.2023.2217843
66. Zhang Z, Huang X, Wang E, Huang Y, Yang R. Suppression of Mll1-Complex by Stat3/Cebpβ-Induced miR-21a/21b/181b maintains the accumulation, homeostasis, and immunosuppressive function of polymorphonuclear myeloid-derived suppressor cells. J Immunol. 2020;204(12):3400–3415. doi:10.4049/jimmunol.2000230
67. Noman MZ, Janji B, Hu S, et al. Tumor-promoting effects of myeloid-derived suppressor cells are potentiated by hypoxia-induced expression of miR-210. Cancer Res. 2015;75(18):3771–3787. doi:10.1158/0008-5472.CAN-15-0405
68. Zhang M, Liu Q, Mi S, et al. Both miR-17-5p and miR-20a alleviate suppressive potential of myeloid-derived suppressor cells by modulating STAT3 expression. J Immunol. 2011;186(8):4716–4724. doi:10.4049/jimmunol.1002989
69. Xu Z, Ji J, Xu J, et al. MiR-30a increases MDSC differentiation and immunosuppressive function by targeting SOCS3 in mice with B-cell lymphoma. FEBS J. 2017;284(15):2410–2424. doi:10.1111/febs.14133
70. Sun R, Zheng Z, Wang L, et al. A novel prognostic model based on four circulating miRNA in diffuse large B-cell lymphoma: implications for the roles of MDSC and Th17 cells in lymphoma progression. Mol Oncol. 2021;15(1):246–261. doi:10.1002/1878-0261.12834
71. Lei Y, Huang Y, Lin J, et al. Mxi1 participates in the progression of lung cancer via the microRNA-300/KLF9/GADD34 Axis. Cell Death Dis. 2022;13(5):425. doi:10.1038/s41419-022-04778-w
72. Su MT, Kumata S, Endo S, Okada Y, Takai T. LILRB4 promotes tumor metastasis by regulating MDSCs and inhibiting miR-1 family miRNAs. Oncoimmunology. 2022;11(1):2060907. doi:10.1080/2162402X.2022.2060907
73. Deng Z, Rong Y, Teng Y, et al. Exosomes miR-126a released from MDSC induced by DOX treatment promotes lung metastasis. Oncogene. 2017;36(5):639–651. doi:10.1038/onc.2016.229
74. Zhou JH, Yao ZX, Zheng Z, et al. G-MDSCs-Derived Exosomal miRNA-143-3p Promotes Proliferation via Targeting of ITM2B in Lung Cancer. Onco Targets Ther. 2020;13:9701–9719. doi:10.2147/OTT.S256378
75. Zhao C, Tang Q, Yang C, et al. Stereotactic body radiation therapy suppresses myeloid-derived suppressor cells by regulating miR-21/Sorbin and SH3 Domain-containing Protein 1 axis. Hum Exp Toxicol. 2024;43:9603271241261307. doi:10.1177/09603271241261307
76. Zhang X, Li F, Tang Y, et al. miR-21a in exosomes from Lewis lung carcinoma cells accelerates tumor growth through targeting PDCD4 to enhance expansion of myeloid-derived suppressor cells. Oncogene. 2020;39(40):6354–6369. doi:10.1038/s41388-020-01406-9
77. Qiu W, Guo Q, Guo X, et al. Mesenchymal stem cells, as glioma exosomal immunosuppressive signal multipliers, enhance MDSCs immunosuppressive activity through the miR-21/SP1/DNMT1 positive feedback loop. J Nanobiotechnology. 2023;21(1):233. doi:10.1186/s12951-023-01997-x
78. Guo X, Qiu W, Liu Q, et al. Immunosuppressive effects of hypoxia-induced glioma exosomes through myeloid-derived suppressor cells via the miR-10a/Rora and miR-21/Pten Pathways. Oncogene. 2018;37(31):4239–4259. doi:10.1038/s41388-018-0261-9
79. Qiu W, Guo X, Li B, et al. Exosomal miR-1246 from glioma patient body fluids drives the differentiation and activation of myeloid-derived suppressor cells. Mol Ther. 2021;29(12):3449–3464. doi:10.1016/j.ymthe.2021.06.023
80. Qi Y, Jin C, Qiu W, et al. The dual role of glioma exosomal microRNAs: glioma eliminates tumor suppressor miR-1298-5p via exosomes to promote immunosuppressive effects of MDSCs. Cell Death Dis. 2022;13(5):426. doi:10.1038/s41419-022-04872-z
81. Guo X, Qiu W, Wang J, et al. Glioma exosomes mediate the expansion and function of myeloid-derived suppressor cells through microRNA-29a/Hbp1 and microRNA-92a/Prkar1a pathways. Int, J, Cancer. 2019;144(12):3111–3126. doi:10.1002/ijc.32052
82. Huber V, Vallacchi V, Fleming V, et al. Tumor-derived microRNAs induce myeloid suppressor cells and predict immunotherapy resistance in melanoma. J Clin Invest. 2018;128(12):5505–5516. doi:10.1172/JCI98060
83. Shidal C, Singh NP, Nagarkatti P, Nagarkatti M. MicroRNA-92 Expression in CD133(+) Melanoma Stem Cells Regulates Immunosuppression in the Tumor Microenvironment via Integrin-Dependent Activation of TGFβ. Cancer Res. 2019;79(14):3622–3635. doi:10.1158/0008-5472.CAN-18-2659
84. Han X, Luan T, Sun Y, Yan W, Wang D, Zeng X. MicroRNA 449c mediates the generation of monocytic myeloid-derived suppressor cells by targeting STAT6. Mol Cells. 2020;43(9):793–803. doi:10.14348/molcells.2020.2307
85. Chen L, Xiong L, Hong S, et al. Circulating myeloid-derived suppressor cells facilitate invasion of thyroid cancer cells by repressing miR-486-3p. J Clin Endocrinol Metab. 2020;105(8):2704–2718. doi:10.1210/clinem/dgaa344
86. Ledo AM, Sasso MS, Bronte V, et al. Co-delivery of RNAi and chemokine by polyarginine nanocapsules enables the modulation of myeloid-derived suppressor cells. J Control Release. 2019;295:60–73. doi:10.1016/j.jconrel.2018.12.041
87. Li L, Cao B, Liang X, et al. Microenvironmental oxygen pressure orchestrates an anti- and pro-tumoral γδ T cell equilibrium via tumor-derived exosomes. Oncogene. 2019;38(15):2830–2843. doi:10.1038/s41388-018-0627-z
88. Wen J, Xuan B, Gao Y, et al. Lnc-17Rik promotes the immunosuppressive function of Myeloid-Derived suppressive cells in esophageal cancer. Cell Immunol. 2023;385:104676. doi:10.1016/j.cellimm.2023.104676
89. Zheng Y, Tian X, Wang T, et al. Long noncoding RNA Pvt1 regulates the immunosuppression activity of granulocytic myeloid-derived suppressor cells in tumor-bearing mice. Mol Cancer. 2019;18(1):61. doi:10.1186/s12943-019-0978-2
90. Yu H, Chen C, Han F, et al. Long Noncoding RNA MIR4435-2HG suppresses colorectal cancer initiation and progression by reprogramming neutrophils. Cancer Immunol Res. 2022;10(9):1095–1110. doi:10.1158/2326-6066.CIR-21-1011
91. Gao Y, Shang W, Zhang D, et al. Lnc-C/EBPβ Modulates Differentiation of MDSCs Through Downregulating IL4i1 With C/EBPβ LIP and WDR5. Front Immunol. 2019;10:1661. doi:10.3389/fimmu.2019.01661
92. Wang Y, Gao Y, Zhang C, et al. Tumor environment promotes Lnc57Rik-mediated suppressive function of myeloid-derived suppressor cells. J Immunol. 2022;209(7):1401–1413. doi:10.4049/jimmunol.2200195
93. Shang W, Gao Y, Tang Z, Zhang Y, Yang R. The Pseudogene Olfr29-ps1 Promotes the Suppressive Function and Differentiation of Monocytic MDSCs. Cancer Immunol Res. 2019;7(5):813–827. doi:10.1158/2326-6066.CIR-18-0443
94. Li X, Liang S, Fei M, et al. LncRNA CRNDE drives the progression of hepatocellular carcinoma by inducing the immunosuppressive niche. Int J Biol Sci. 2024;20(2):718–732. doi:10.7150/ijbs.85471
95. Tian X, Ma J, Wang T, et al. Long non-coding RNA RUNXOR accelerates MDSC-mediated immunosuppression in lung cancer. BMC Cancer. 2018;18(1):660. doi:10.1186/s12885-018-4564-6
96. Lu W, Cao F, Feng L, et al. LncRNA Snhg6 regulates the differentiation of MDSCs by regulating the ubiquitination of EZH2. J Hematol Oncol. 2021;14(1):196. doi:10.1186/s13045-021-01212-0
97. Tian X, Zheng Y, Yin K, et al. LncRNA AK036396 inhibits maturation and accelerates immunosuppression of polymorphonuclear myeloid-derived suppressor cells by enhancing the stability of ficolin B. Cancer Immunol Res. 2020;8(4):565–577. doi:10.1158/2326-6066.CIR-19-0595
98. Tian X, Ma J, Wang T, et al. Long Non-Coding RNA HOXA Transcript Antisense RNA Myeloid-Specific 1-HOXA1 Axis Downregulates the Immunosuppressive Activity of Myeloid-Derived Suppressor Cells in Lung Cancer. Front Immunol. 2018;9:473. doi:10.3389/fimmu.2018.00473
99. Adewunmi O, Shen Y, Zhang XH, Rosen JM. Targeted Inhibition of lncRNA Malat1 Alters the tumor immune microenvironment in preclinical syngeneic mouse models of triple-negative breast cancer. Cancer Immunol Res. 2023;11(11):1462–1479. doi:10.1158/2326-6066.CIR-23-0045
100. Gao Y, Wang T, Li Y, Zhang Y, Yang R. Lnc-chop promotes immunosuppressive function of myeloid-derived suppressor cells in tumor and inflammatory environments. J Immunol. 2018;200(8):2603–2614. doi:10.4049/jimmunol.1701721
101. Ma X, Sheng S, Wu J, et al. LncRNAs as an intermediate in HPV16 promoting myeloid-derived suppressor cell recruitment of head and neck squamous cell carcinoma. Oncotarget. 2017;8(26):42061–42075. doi:10.18632/oncotarget.14939
102. Li Y, Zhang Q, Wu M, et al. Suppressing MDSC infiltration in tumor microenvironment serves as an option for treating ovarian cancer metastasis. Int J Biol Sci. 2022;18(9):3697–3713. doi:10.7150/ijbs.70013
103. Chen J, Gao Y, Zhong J, et al. Lnc-H19-derived protein shapes the immunosuppressive microenvironment of glioblastoma. Cell Rep Med. 2024;5(11):101806. doi:10.1016/j.xcrm.2024.101806
104. Tian Y, Gao X, Yang X, Chen S, Ren Y. Glioma-derived exosome Lncrna Agap2-As1 promotes glioma proliferation and metastasis by mediating Tgf-β1 secretion of myeloid-derived suppressor cells. Heliyon. 2024;10(9):e29949. doi:10.1016/j.heliyon.2024.e29949
105. Yang Y, Chen XQ, Jia YX, Ma J, Xu D, Xiang ZL. Circ-0044539 promotes lymph node metastasis of hepatocellular carcinoma through exosomal-miR-29a-3p. Cell Death Dis. 2024;15(8):630. doi:10.1038/s41419-024-07004-x
106. Chen W, Peng W, Wang R, et al. Exosome-derived tRNA fragments tRF-GluCTC-0005 promotes pancreatic cancer liver metastasis by activating hepatic stellate cells. Cell Death Dis. 2024;15(1):102. doi:10.1038/s41419-024-06482-3
107. Shi X, Pang S, Zhou J, et al. Bladder-cancer-derived exosomal circRNA_0013936 promotes suppressive immunity by up-regulating fatty acid transporter protein 2 and down-regulating receptor-interacting protein kinase 3 in PMN-MDSCs. Mol Cancer. 2024;23(1):52. doi:10.1186/s12943-024-01968-2
108. Gao F, Xu Q, Tang Z, et al. Exosomes derived from myeloid-derived suppressor cells facilitate castration-resistant prostate cancer progression via S100A9/circMID1/miR-506-3p/MID1. J Transl Med. 2022;20(1):346. doi:10.1186/s12967-022-03494-5
109. Katopodi T, Petanidis S, Domvri K, et al. Kras-driven intratumoral heterogeneity triggers infiltration of M2 polarized macrophages via the circHIPK3/PTK2 immunosuppressive circuit. Sci Rep. 2021;11(1):15455. doi:10.1038/s41598-021-94671-x
110. Ai L, Mu S, Sun C, et al. Myeloid-derived suppressor cells endow stem-like qualities to multiple myeloma cells by inducing piRNA-823 expression and DNMT3B activation. Mol Cancer. 2019;18(1):88. doi:10.1186/s12943-019-1011-5
111. Grillone K, Caridà G, Luciano F, et al. A systematic review of non-coding RNA therapeutics in early clinical trials: a new perspective against cancer. J Transl Med. 2024;22(1):731. doi:10.1186/s12967-024-05554-4
112. Bian J, Shao R, Li J, et al. Mechanism research of non-coding RNA in immune checkpoint inhibitors therapy. Cancer Sci. 2024;115(11):3520–3531. doi:10.1111/cas.16309
113. Yan B, Liu C, Li H, et al. Reversal of HMGA1-mediated immunosuppression synergizes with immunogenic magnetothermodynamic for improved hepatocellular carcinoma therapy. ACS Nano. 2023;17(10):9209–9223. doi:10.1021/acsnano.3c00004
114. Liu Z, Zhao L, Feng Y, et al. Dual-responsive PEG-lipid polyester nanoparticles for siRNA and vaccine delivery elicit anti-cancer immune responses by modulating tumor microenvironment. Biomater Sci. 2023;11(19):6619–6634. doi:10.1039/D3BM01265D
115. Tong X, Qiao S, Dong Z, Zhao X, Du X, Niu W. Targeting CSF1R in myeloid-derived suppressor cells: insights into its immunomodulatory functions in colorectal cancer and therapeutic implications. J Nanobiotechnology. 2024;22(1):409. doi:10.1186/s12951-024-02584-4
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
RNA Modifications Meet Tumors
Yang Z, Zhang S, Xia T, Fan Y, Shan Y, Zhang K, Xiong J, Gu M, You B
Cancer Management and Research 2022, 14:3223-3243
Published Date: 22 November 2022
Tumor Immunotherapy Targeting B7-H3: From Mechanisms to Clinical Applications

Guo Y, Wang X, Zhang C, Chen W, Fu Y, Yu Y, Chen Y, Shao T, Zhang J, Ding G
ImmunoTargets and Therapy 2025, 14:291-320
Published Date: 27 March 2025
Remodeling the Stroma: The Role of ncRNAs in Cancer-Associated Fibroblasts and Therapeutic Implications for Digestive Tumors
Luo K, Hou W, Zhang W
Drug Design, Development and Therapy 2026, 20:600029
Published Date: 23 June 2026