Back to Journals » Clinical Pharmacology: Advances and Applications » Volume 18
Transcriptomic Modulation of Inflammasome-Related Genes Following Metformin Administration in Breast Cancer Patients with Paclitaxel-Induced Neuropathy
Authors Malayeri A, Etemad Nezhad M, Ramesh F, Karimpourian H, Mahmoudian-Sani MR ![]()
Received 18 November 2025
Accepted for publication 26 May 2026
Published 4 June 2026 Volume 2026:18 582439
DOI https://doi.org/10.2147/CPAA.S582439
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 4
Editor who approved publication: Dr Khaled Deeb
Alireza Malayeri,1 Mehdi Etemad Nezhad,2 Farrokh Ramesh,3 Hossein Karimpourian,4 Mohammad-Reza Mahmoudian-Sani4
1Medicinal Plant Research Center, Ahvaz Jundishapur University of Medical Sciences, Ahvaz, Iran; 2Student Research Committee, Faculty of Pharmacy, Ahvaz Jundishapur University of Medical Sciences, Ahvaz, Iran; 3Department of Pharmacology, School of Pharmacy, Ahvaz Jundishapur University of Medical Sciences, Ahvaz, Iran; 4Thalassemia and Hemoglobinopathy Research Center, Health Research Institute, Ahvaz Jundishapur University of Medical Sciences, Ahvaz, Iran
Mohammad-Reza Mahmoudian-Sani
Thalassemia and Hemoglobinopathy Research Center, Health Research Institute, Ahvaz Jundishapur University of Medical Sciences, Ahvaz, Iran
, Tel +98 061-33750410
, Fax +98 061-33750427
, Email [email protected]
Introduction: Chemotherapy-induced peripheral neuropathy (CIPN) is a common adverse effect of paclitaxel therapy. The inflammasome complex, including apoptosis-associated speck-like protein containing a CARD (ASC), Caspase-1, and NOD-like receptor family pyrin domain-containing 3 (NLRP3), is implicated in inflammatory signaling pathways related to neuropathy. This study aimed to evaluate transcriptomic changes in genes involved in the inflammasome pathway in breast cancer patients, with a specific focus on metformin treatment initiated after the onset of neuropathy.
Materials and Methods: A total of 51 breast cancer patients receiving paclitaxel were included (26 controls, 25 metformin). Metformin was initiated following the clinical onset of neuropathy. Plasma samples were collected at baseline, cycle 6, and cycle 12. Expression levels of ASC, Caspase-1, and NLRP3 were quantified as fold-change. Statistical analyses included normality testing (Shapiro–Wilk), temporal comparisons (Friedman or repeated-measures ANOVA), group comparisons (Mann–Whitney U-test), correlation (Spearman), and Receiver Operating Characteristic (ROC) analysis.
Results: No significant baseline differences were observed between groups. In the control group, ASC expression increased over time (p = 0.0005), while Caspase-1 and NLRP3 showed no significant temporal changes. In neuropathic patients, ASC (p < 0.0001), Caspase-1 (p = 0.0007), and NLRP3 (p = 0.04) expression levels were higher in the metformin group. ROC analysis demonstrated moderate discriminatory ability for ASC (AUC = 0.74) and Caspase-1 (AUC = 0.70), whereas NLRP3 showed weaker performance (AUC = 0.62). Correlation analysis revealed positive associations between ASC and Caspase-1, suggesting coordinated gene expression. These findings reflect transcriptomic modulation rather than functional inflammasome activation.
Conclusion: Metformin administration after neuropathy onset was associated with transcriptomic changes in genes involved in the inflammasome pathway. However, these findings should be interpreted cautiously, as plasma RNA may not reflect protein activity or neural tissue processes, and further validation studies are required.
Keywords: breast cancer, paclitaxel, metformin, inflammasome, ASC, caspase-1, NLRP3, chemotherapy-induced neuropathy
Introduction
Breast cancer is one of the most commonly diagnosed malignancies worldwide and remains a leading cause of cancer-related mortality among women.1,2 Chemotherapy is a cornerstone of treatment; however, it is frequently associated with adverse effects, among which chemotherapy-induced peripheral neuropathy (CIPN) is particularly significant. CIPN is commonly observed with taxane-based regimens such as paclitaxel and manifests clinically as numbness, tingling, and pain in the extremities.3–5 Approximately 60% of patients receiving paclitaxel develop varying degrees of neuropathy.6 This condition is often dose-dependent and is associated with structural and functional alterations in peripheral nerves, particularly in the dorsal root ganglion (DRG), as well as activation of inflammatory pathways.7 Metformin, a widely prescribed antidiabetic agent, has attracted considerable attention due to its pleiotropic effects beyond glycemic control.8 Previous studies have shown that metformin can modulate multiple signaling pathways, including AMPK and MEK/ERK, and suppress inflammatory responses by inhibiting NOD-like receptor family pyrin domain containing 3 (NLRP3) activation and downregulating pro-inflammatory mediators.9,10 Furthermore, metformin has garnered substantial interest for its anti-inflammatory and neuroprotective properties.11 In experimental models, metformin has demonstrated neuroprotective effects by reducing NLRP3, Apoptosis-associated speck-like protein containing a CARD (ASC), and Caspase-1 expression, inhibiting microglial activation, and modulating pain-related ion channels such as TRPV1 and ASIC3.10,12–14 These findings suggest a potential role for metformin in mitigating neuropathic processes. Neurotoxicity induced by anticancer agents involves multiple mechanisms, including axonal degeneration, DRG neuronal injury, and disruption of neuroimmune homeostasis. Emerging evidence indicates that neuroinflammation, involving both innate and adaptive immune responses, plays a role in the development of CIPN. Glial cells, including satellite glial cells, Schwann cells, microglia, and astrocytes, contribute to the maintenance and amplification of inflammatory signaling in the peripheral and central nervous systems.15 The inflammasome signaling pathway, particularly the NLRP3 inflammasome, has been recognized as an important mediator in this process, regulating the activation of Caspase-1 and the maturation of pro-inflammatory cytokines such as IL-1β and IL-18.16–18 The NLRP3 inflammasome is a multiprotein complex consisting of the NLRP3 sensor, the adaptor protein ASC, and the effector Caspase-1, and its activation is triggered by cellular stress signals such as mitochondrial dysfunction, reactive oxygen species production, ionic imbalance, and lysosomal damage.19–23 In breast cancer, the expression of inflammasome-related components has been associated with tumor characteristics and clinical outcomes.24–26 Additionally, metformin has been reported to exert anticancer and anti-inflammatory effects by modulating pathways such as NF-κB/IL-6 and reducing oxidative stress.12,19,27 Unlike previous studies that primarily focused on metformin’s prophylactic effects in preventing CIPN, the distinction between prophylactic and therapeutic approaches is critical. Prophylactic strategies aim to prevent the onset of neuropathy, whereas therapeutic interventions are initiated after the development of clinical symptoms and may involve different molecular responses. In the present study, metformin was administered only after the onset of neuropathy, representing a therapeutic approach. Circulating plasma RNA has emerged as a minimally invasive approach for evaluating systemic molecular alterations. Although plasma RNA does not directly represent gene expression within neural tissues, it reflects contributions from multiple sources, including circulating immune cells, endothelial cells, and extracellular vesicles. Therefore, it may provide indirect insight into systemic inflammatory and stress-related responses associated with CIPN. Given these findings, it may be hypothesized that metformin could influence inflammasome-related pathways in patients experiencing CIPN. However, clinical evidence regarding its role in this context remains limited. Importantly, the effect of metformin when administered after the onset of neuropathy has not been adequately investigated. Therefore, the present study was designed to evaluate the transcriptomic profile of key inflammasome-related genes, including ASC, Caspase-1, and NLRP3, in breast cancer patients receiving paclitaxel chemotherapy.
Materials and Methods
Sample Collection and Ethical Considerations
The biological samples used in this study were obtained from the existing biobank of the Thalassemia and Hemoglobinopathy Research Center, Health Research Institute, Ahvaz Jundishapur University of Medical Sciences. These samples had been collected previously under an approved research protocol. The present study was designed and executed with new objectives and hypotheses, and it was assigned an independent ethical approval code separate from that of the prior project. All procedures were performed in accordance with the moral principles outlined in the Declaration of Helsinki, and the study protocol was approved by the Ethics Committee of Ahvaz Jundishapur University of Medical Sciences (Ethics Code: IR.AJUMS.REC.1403.539). Only basic demographic data (age and sex) were shared across datasets; all subsequent analyses and research objectives were entirely independent.
Study Design and Patient Population
This prospective cohort study included 51 breast cancer patients undergoing paclitaxel chemotherapy. Patients were divided into a control group (n = 26) and a metformin-treated group (n = 25). Importantly, metformin was initiated only after the clinical onset of neuropathy, representing a therapeutic rather than prophylactic approach. The study included breast cancer patients undergoing paclitaxel-based chemotherapy. Eligible participants were adults (≥18 years) with a confirmed histopathological diagnosis of breast cancer and scheduled to receive paclitaxel chemotherapy. Patients with uncontrolled diabetes, pre-existing neuropathy, renal or hepatic impairment, significant psychiatric illness, pregnancy, known metformin intolerance, or use of medications affecting neuropathy or inflammatory pathways were excluded. All participants provided written informed consent after receiving a complete explanation of the study aims and procedures, and enrollment began only after ethical approval was obtained. Neuropathy was assessed using the National Cancer Institute Common Terminology Criteria for Adverse Events (NCI-CTCAE v5.0) at baseline and during each chemotherapy cycle. Metformin treatment was initiated in patients who developed Grade ≥2 neuropathy, and the cumulative paclitaxel dose at neuropathy onset was recorded.
Patient Selection and Group Allocation
During paclitaxel treatment, all patients were regularly evaluated for signs of CIPN. Following the development of neuropathy after several cycles of chemotherapy, eligible patients were recruited into the study. A total of 51 patients met the inclusion criteria:25 patients were assigned to the metformin group and received 500 mg metformin three times daily from the time neuropathy was diagnosed until completion of chemotherapy. Twenty-six patients continued routine chemotherapy without metformin and were designated as the control group.
Sample Collection
Peripheral blood samples were collected at baseline, cycle 6, and cycle 12. Plasma was separated by centrifugation and stored at −80°C until further analysis.
Treatment Protocol and Sampling Timeline
All patients received standard paclitaxel chemotherapy at a dose of 75 mg/m2, administered intravenously once weekly for a total of 12 cycles. Peripheral blood samples (5 mL) were collected at three designated time points: baseline (before initiation of chemotherapy), after the 6th cycle, and after the 12th cycle.
Sample Processing and Plasma Storage
Immediately after collection, blood samples were transported on ice to the laboratory. Plasma separation was performed by centrifugation at 3000 rpm for 7 minutes at 4°C. The isolated plasma was aliquoted into RNase-free microtubes and stored at −80°C until molecular analyses were performed.
RNA Extraction
Total RNA was extracted from 400 µL of plasma using a commercial RNA extraction kit (Cat. No. FAWBR050, Favorgen Biotech Corp., Ping-Tung, Taiwan) according to the manufacturer’s instructions. RNA concentration and purity were assessed by spectrophotometry, and samples with an OD260/280 ratio of 1.8–2.0 were considered acceptable In brief, blood cells were lysed using a guanidinium thiocyanate-containing buffer, and Proteinase K was added to ensure complete protein degradation. After ethanol addition, RNA was bound to a silica membrane column and subjected to sequential washes with the provided buffers. Purified RNA was eluted in RNase-free water with a final volume of 30–50 μL. All extraction steps were performed under RNase-free conditions, and samples were stored at −80°C until downstream processing.
cDNA Synthesis
cDNA was synthesized using a reverse transcription kit (Cat. No. YT4500, Favorgen Biotech Corp., Ping-Tung, Taiwan) following the manufacturer’s protocol. The reaction mixture contained M-MuLV Reverse Transcriptase, 5× reaction buffer, dNTP mix, RNase inhibitor, random hexamer primers, and Oligo(dT)18 primers. For Oligo(dT)18-primed reactions, reverse transcription was performed at 42°C for 60 minutes, whereas reactions primed with random hexamers were conducted at 37°C. Enzyme inactivation was performed at 70°C for 5 minutes, and the resulting cDNA was stored at −20°C until further quantitative PCR (qPCR) analysis.
Real-Time PCR
Quantitative real-time PCR was performed using SYBR Green Master Mix (Cat. No. YT2551, Favorgen Biotech Corp., Ping-Tung, Taiwan) in a final reaction volume of 20 µL. All reactions were performed in triplicate, and a no-template control (NTC) was included to assess contamination. Amplification efficiency was within the acceptable range (90–110%). Quantitative PCR was performed on an Applied Biosystems StepOne Real-Time PCR System. Each 20-µL amplification reaction consisted of 10 µL SYBR Green Master Mix, 0.4 µL of each primer (10 µM), template cDNA, and nuclease-free water. The thermal cycling conditions were as follows: initial denaturation at 95°C for 3 minutes, followed by 40 cycles of denaturation at 95°C for 10 seconds, annealing at 60°C for 10 seconds, and extension at 72°C for 20 seconds. Melting curve analysis was performed to verify the specificity of the PCR products. GAPDH was used as the internal control, and relative gene expression levels were calculated using the 2^−ΔΔCt method. GAPDH was used as the reference gene due to its stable expression across samples. ASC, Caspase-1, and NLRP3 were selected as key components of the canonical inflammasome pathway.
Quality Control and Technical Considerations
All laboratory procedures were conducted in a blinded manner with respect to group allocation. Samples that did not meet RNA quality thresholds or internal quality control parameters were excluded from analysis. Each qPCR run included no-template controls (NTC) and an internal housekeeping control to ensure assay accuracy and reliability.
Statistical Analysis
Gene expression levels were calculated using the 2^-ΔΔCt method. Normality of distribution for fold-change values of NLRP3, ASC, and Caspase-1 was assessed using the Shapiro–Wilk test. Because most variables were non-normally distributed, nonparametric statistical methods were employed for subsequent analyses. Temporal within-group changes across baseline, cycle 6, and cycle 12 were evaluated using the Friedman test; significant outcomes were further examined using Dunn’s multiple-comparison test. For variables exhibiting normal distributions, such as NLRP3, a Repeated-Measures ANOVA followed by Tukey’s post hoc test was applied. Between-group comparisons (metformin versus control) among neuropathic patients were performed using the Mann–Whitney U-test due to non-normality. Results are presented as median (interquartile range, IQR), as appropriate. The diagnostic performance of each gene was assessed using receiver operating characteristic (ROC) analysis, with area under the curve (AUC), standard error, and 95% confidence intervals (CI) reported. Associations among gene expression levels were evaluated using Spearman correlation coefficient. Statistical analysis was performed using IBM SPSS Statistics for Windows, version 21 (IBM Corp., Armonk, NY, USA), GraphPad Prism, version 10 (GraphPad Software, San Diego, CA, USA), and GenEx, with p-values < 0.05 considered statistically significant.
Ethical Considerations
This study was conducted in accordance with the Declaration of Helsinki. Written informed consent was obtained from all participants prior to enrollment. Renal function was monitored during the study period, and adverse effects of metformin, including gastrointestinal symptoms, were recorded. No serious adverse events were observed.
Results
A total of 51 breast cancer patients receiving paclitaxel chemotherapy were included in this study, comprising 26 patients in the control group and 25 patients in the metformin-treated group. No statistically significant baseline demographic differences were observed between the groups at study entry. Normality assessment of fold-change data using the Shapiro–Wilk test indicated that ASC expression showed non-normal distributions at all evaluated time points, whereas Caspase-1 showed mixed distributions, and NLRP3 generally showed normal distributions across treatment cycles (Table 1). Based on these findings, non-parametric statistical analyses were primarily applied. Longitudinal analysis within the control group demonstrated a significant time-dependent increase in ASC expression across chemotherapy cycles (Table 2). Post hoc analysis revealed significant differences between baseline and Cycle 6 (p = 0.0135) and between baseline and Cycle 12 (p = 0.0005), whereas no significant difference was observed between Cycle 6 and Cycle 12. In contrast, Caspase-1 and NLRP3 expression levels did not demonstrate significant temporal variation during treatment. Figure 1 illustrates the longitudinal fold-change patterns of ASC, Caspase-1, and NLRP3 across baseline, Cycle 6, and Cycle 12. ASC showed a progressive increase over time, whereas Caspase-1 and NLRP3 remained relatively stable Between-group comparisons showed higher median fold-changes in ASC, Caspase-1, and NLRP3 expression in metformin-treated patients compared with controls (Tables 3 and 4). ASC showed the greatest difference, with median expression levels increasing from 1.21 in controls to 2.73 in the metformin group (p < 0.0001). Caspase-1 expression also showed a moderate increase (1.18 vs. 1.76, p = 0.0007). Although NLRP3 reached nominal statistical significance (0.81 vs. 10.67, p = 0.04), substantial inter-individual variability and broad confidence intervals were observed, suggesting considerable biological heterogeneity. Therefore, the NLRP3 findings should be interpreted cautiously. Hodges–Lehmann estimation further supported intergroup differences, particularly for ASC and Caspase-1. Figure 2 demonstrates the distribution of fold-change values for all three genes in both study groups. ROC curve analysis was performed to evaluate the discriminatory performance of inflammasome-related transcripts between metformin-treated and control patients (Table 5). ASC demonstrated the strongest discriminatory performance among the evaluated markers (AUC = 0.74, 95% CI: 0.6339–0.8464), followed by Caspase-1 (AUC = 0.70, 95% CI: 0.5928–0.8167). In contrast, NLRP3 showed relatively weak discriminatory ability (AUC = 0.62, 95% CI: 0.5082–0.7413), with performance only marginally above random classification. These findings suggest that ASC and Caspase-1 may possess moderate transcript-level discriminatory potential, whereas the biomarker relevance of NLRP3 remains uncertain and requires validation in larger cohorts. Figure 3 presents the ROC curves for ASC, Caspase-1, and NLRP3. Correlation analysis revealed positive associations among several inflammasome-related transcripts (Table 6). The strongest correlation was observed between ASC and Caspase-1 within the metformin group (r = 0.62, p < 0.001), whereas a slightly weaker but still significant association was identified in the control group (r = 0.55, p < 0.01). A weaker correlation was also observed between ASC and NLRP3 in the metformin group (r = 0.32, p = 0.04). Other correlations were either weak or non-significant. These findings indicate coordinated transcriptomic expression patterns among selected inflammasome-related genes. Figure 4 illustrates the correlation patterns among ASC, Caspase-1, and NLRP3 expression levels in both study groups. Overall, the observed findings indicate transcriptomic modulation of inflammasome-related genes following metformin administration after neuropathy onset. However, because the present study was limited to plasma RNA analysis without protein-level or functional validation, these findings should not be interpreted as direct evidence of functional inflammasome activation. In addition, the relatively small sample size and substantial biological heterogeneity, particularly for NLRP3, warrant cautious interpretation of the results.
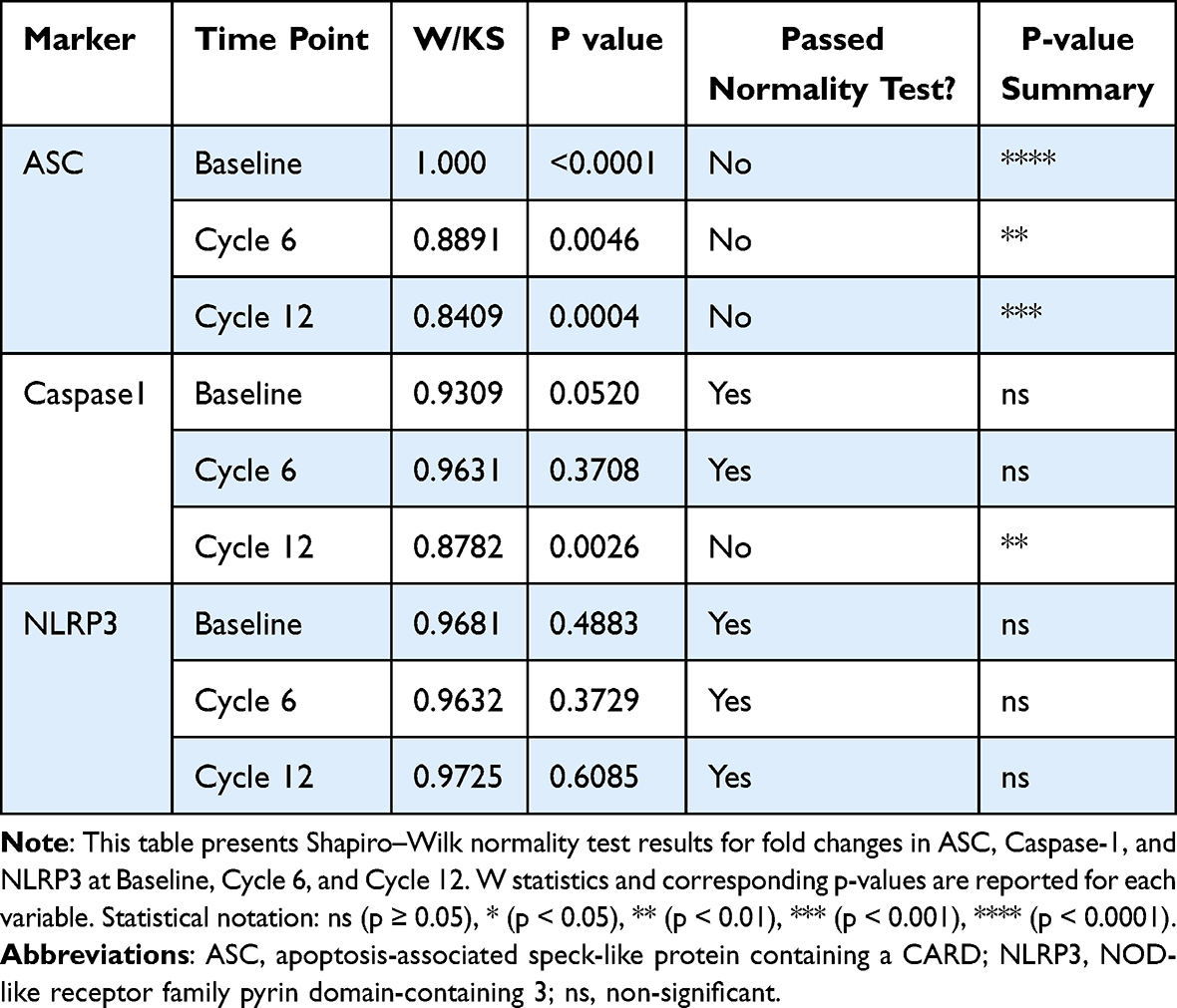 |
Table 1 Shapiro–Wilk Test for Normality of Fold-Change Data for ASC, Caspase-1, and NLRP3 at Baseline, Cycle 6, and Cycle 12 |
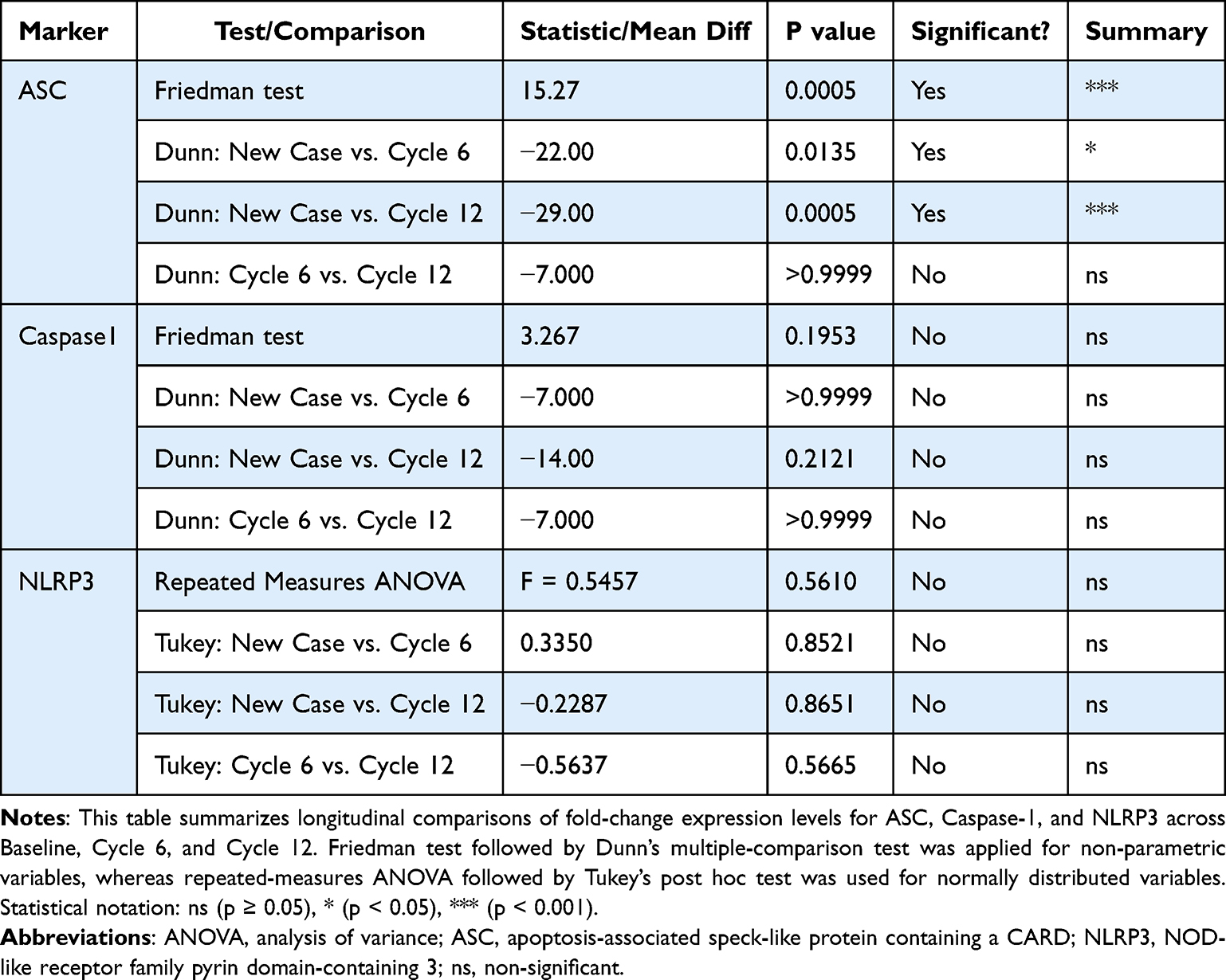 |
Table 2 Comparative Analysis of Longitudinal Fold-Change Expression Across Treatment Cycles |
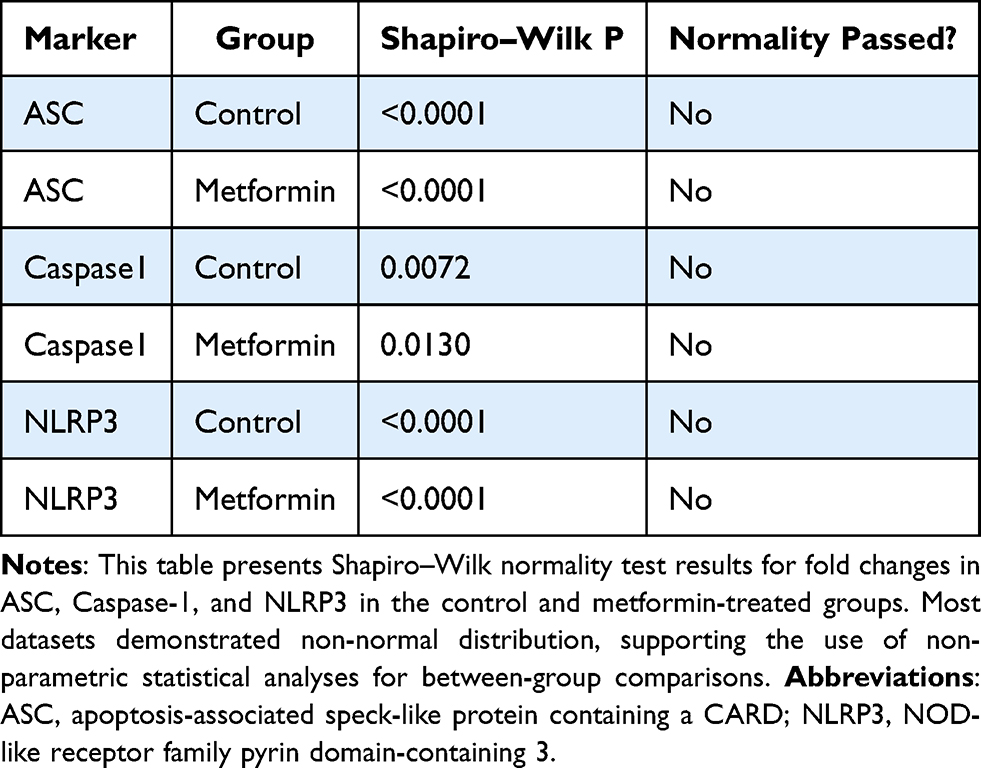 |
Table 3 Shapiro–Wilk Test for Fold-Change Data Normality in Control and Metformin Groups |
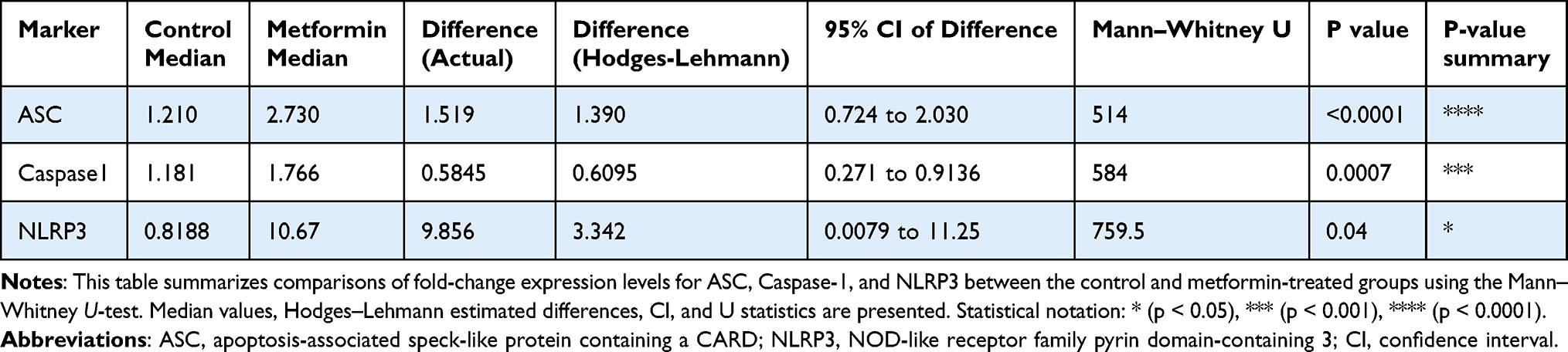 |
Table 4 Comparison of Fold-Change Expression Levels Between Control and Metformin Groups |
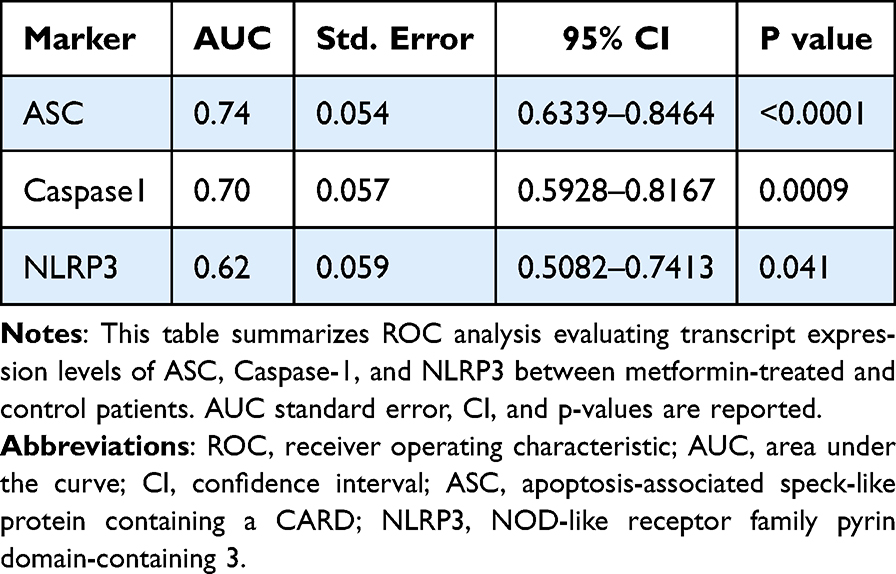 |
Table 5 ROC Analysis of Inflammasome-Related Transcripts |
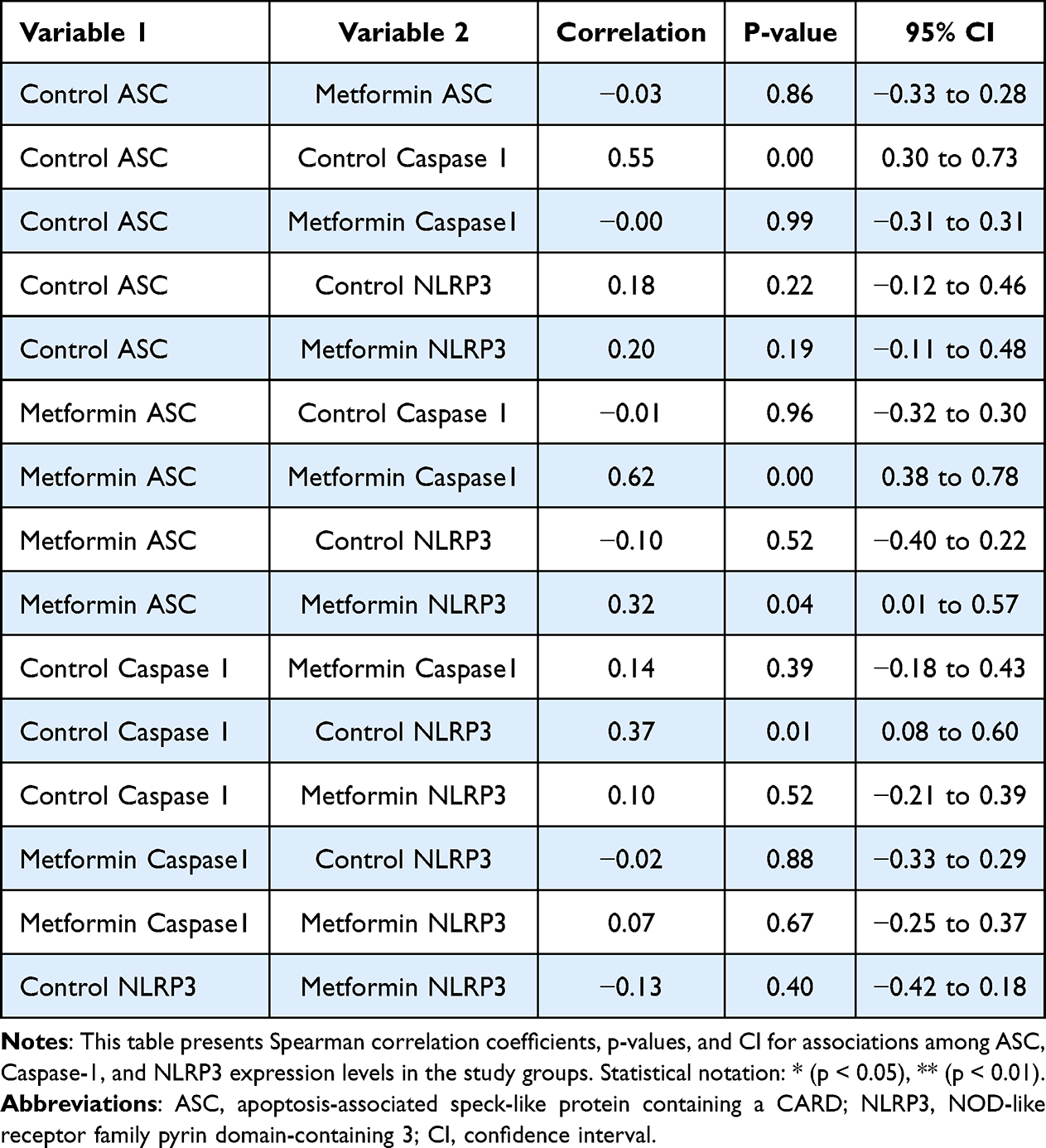 |
Table 6 Spearman Correlation Analysis Among Inflammasome-Related Transcripts |
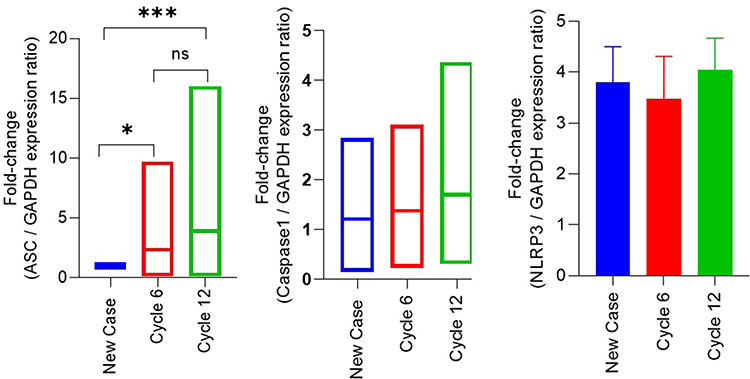 |
Figure 1 Longitudinal Fold-Change Expression Patterns of ASC, Caspase-1, and NLRP3 During Paclitaxel Chemotherapy. Abbreviations: ASC, apoptosis-associated speck-like protein containing a CARD; NLRP3, NOD-like receptor family pyrin domain-containing 3; IQR, interquartile range; ANOVA, analysis of variance; ns, non-significant. Notes: This figure illustrates fold-change expression levels of ASC, Caspase-1, and NLRP3 at Baseline, Cycle 6, and Cycle 12 in breast cancer patients receiving paclitaxel chemotherapy. Data are presented as median and IQR. Statistical analyses were performed using the Friedman test or a repeated-measures ANOVA, depending on the data distribution. Statistical notation: ns (p ≥ 0.05), * (p < 0.05), *** (p < 0.001). |
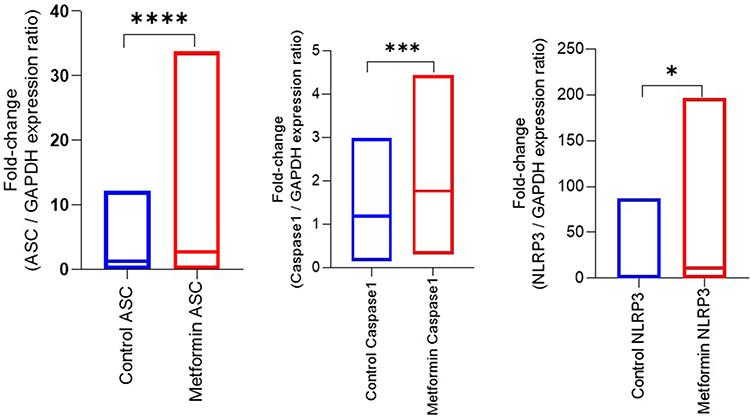 |
Figure 2 Comparison of Fold-Change Expression Levels Between Control and Metformin Groups. Abbreviations: ASC, apoptosis-associated speck-like protein containing a CARD; NLRP3, NOD-like receptor family pyrin domain-containing 3; IQR, interquartile range. Notes: This figure compares fold changes in ASC, Caspase-1, and NLRP3 expression between control and metformin-treated patients. Data are presented as median and IQR. Statistical comparisons were performed using the Mann–Whitney U-test. Statistical notation: * (p < 0.05), *** (p < 0.001), **** (p < 0.0001). |
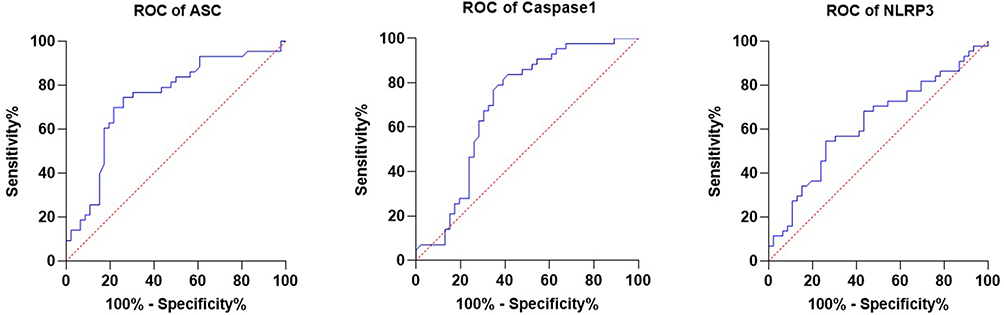 |
Figure 3 ROC Curves for ASC, Caspase-1, and NLRP3. Abbreviations: ROC, receiver operating characteristic; AUC, area under the curve; ASC, apoptosis-associated speck-like protein containing a CARD; NLRP3, NOD-like receptor family pyrin domain-containing 3. Notes: This figure presents ROC curves evaluating the discriminatory performance of ASC, Caspase-1, and NLRP3 transcript expression levels between metformin-treated and control patients. AUC values were calculated for each marker. |
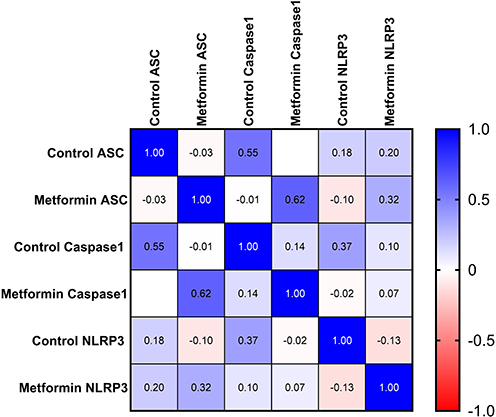 |
Figure 4 Correlation Patterns Among ASC, Caspase-1, and NLRP3 Expression Levels. Abbreviations: ASC, apoptosis-associated speck-like protein containing a CARD; NLRP3, NOD-like receptor family pyrin domain-containing 3. Notes: This figure illustrates Spearman correlation analysis among ASC, Caspase-1, and NLRP3 transcript expression levels in control and metformin-treated patients. Correlation coefficients and corresponding p-values are presented within the figure. |
Discussion
In this exploratory cohort study of 51 paclitaxel-treated breast cancer patients (25 receiving metformin, 26 controls), gene expression of three principal inflammasome components, ASC, Caspase-1, and NLRP3, was examined in plasma samples. Median fold-change expression of all three genes was significantly higher in the metformin group than in controls, with notable increases in ASC (p < 0.0001), Caspase-1 (p = 0.0007), and NLRP3 (p = 0.04). Temporal intra-group analysis revealed that only ASC showed a significant time-dependent increase from Baseline to Cycle 12 (p = 0.0005), while Caspase-1 and NLRP3 did not. ROC analysis indicated that ASC (AUC = 0.74) and Caspase-1 (AUC = 0.70) provided moderate to good discrimination between the metformin and control groups, whereas NLRP3 exhibited lower discrimination (AUC = 0.62). Combined biomarker models may improve discriminatory performance and should be evaluated in future studies. Correlation analysis revealed a significant increase in ASC–Caspase-1 correlation in the metformin group (r ≈ 0.62, p < 0.001) compared with controls (r ≈ 0.55, p < 0.01), suggesting coordinated transcriptomic regulation rather than direct evidence of increased inflammasome activity. Importantly, these findings should be interpreted as transcriptomic changes rather than evidence of enhanced inflammatory activity. Increased mRNA expression does not necessarily correspond to increased protein levels or functional activation of the inflammasome pathway. This distinction is critical when interpreting plasma-based gene expression data. These findings are partially consistent with previous studies reporting NLRP3 involvement in paclitaxel-induced neuropathy (PIN), such as Jia et al (2017), who demonstrated increased inflammasome activity in neural tissues of paclitaxel-treated animals.28 Conversely, other studies (Ha et al29 and Bakry et al11 reported anti-inflammatory and protective effects of metformin on the NLRP3 pathway. Ha et al demonstrated that metformin can inhibit NLRP3 assembly through AMPK activation in microglia, and Bakry et al reported a reduction in paclitaxel-induced peripheral neuropathy (PIPN) in patients treated with metformin. The apparent discrepancy with the present findings may be explained by several key factors. First, the sample source differs: previous studies mainly examined target tissues such as dorsal root ganglia or neural tissue, whereas this study assessed plasma gene expression. Plasma-derived RNA may originate from circulating immune cells, endothelial cells, or extracellular vesicles, and therefore may reflect systemic responses rather than direct neuronal inflammasome activity. Emerging evidence suggests that innate immunity plays a crucial role in CIPN, with systemic inflammatory responses contributing to neuropathic processes.30 Second, differences between transcriptional and functional regulation must be considered. Metformin may inhibit inflammasome activation at the protein or enzymatic level without reducing mRNA expression. Therefore, increased transcript levels may represent compensatory or adaptive responses rather than enhanced pathway activation. Third, the timing of metformin administration is a critical factor. Bakry et al initiated metformin before chemotherapy as a prophylactic intervention, whereas in the present study, metformin was administered after the onset of neuropathy. This distinction between prophylactic and therapeutic use may result in fundamentally different molecular responses. Additional factors, including dosage, metabolic status, and comorbid conditions, may also influence gene expression patterns. Consistent with the present findings, Cristiano et al31 demonstrated that taxanes activate glial cells, leading to the release of pro-inflammatory cytokines and modulation of inflammasome-related pathways. Multi-omics analyses in paclitaxel-treated patients further indicate that CIPN involves complex and interconnected molecular networks.32 Chen et al33 also confirmed the role of NLRP3 signaling in inflammatory pain pathways. However, the observed increase in Caspase-1 and ASC transcripts in the present study should not be interpreted as evidence of increased inflammasome-mediated inflammation, but rather as potential indicators of systemic or compensatory gene regulation. The increased ASC–Caspase-1 correlation in the metformin group likely reflects coordinated pathway regulation rather than causative inflammatory activation. ASC showed the most pronounced temporal change, which may reflect upstream signaling dynamics, cellular stress responses, or release from circulating sources, such as extracellular vesicles. Nevertheless, without protein-level validation, these interpretations remain speculative. Analysis indicated moderate diagnostic performance for ASC and Caspase-1; however, given the limited sample size and lack of external validation, these findings should be interpreted cautiously. The present study has several strengths, including a longitudinal design with multiple sampling time points and comprehensive statistical analyses. However, several limitations should be acknowledged. The present study was limited to transcriptomic data without corresponding protein-level or functional assessments, such as IL-1β, IL-18, or Caspase-1 activity measurements, thereby restricting mechanistic interpretation. In addition, increased transcript abundance should not be interpreted as direct evidence of functional inflammasome activation. The relatively small sample size, substantial inter-individual biological variability, and absence of an independent external validation cohort may further limit statistical power and generalizability. Multiple-comparison correction was not applied; therefore, borderline findings, particularly for NLRP3, should be interpreted cautiously. Because treatment allocation was not randomized, potential selection bias and differences in neuropathy severity between groups cannot be excluded. Furthermore, clinical improvement in neuropathy symptoms following metformin treatment was not systematically quantified using longitudinal symptom-based outcome measures. Another important limitation is the uncertain cellular origin of circulating plasma RNA, which may derive from immune cells, endothelial cells, platelets, or extracellular vesicles rather than directly from neural tissues. Consequently, RNA-based findings cannot be directly equated with neural tissue pathology or functional inflammasome activity and may instead reflect systemic, compensatory, or non-neuronal transcriptomic responses. Therefore, the present findings should be considered exploratory and hypothesis-generating rather than definitive evidence of functional regulation of the inflammasome. Future studies with larger cohorts, randomized designs, and integrated multi-level analyses incorporating protein, enzymatic, and tissue-specific assessments are required. Comparative studies evaluating prophylactic versus post-onset metformin administration are particularly important to clarify potential context-dependent effects.
Conclusion
This study showed that metformin administration after the onset of PIN was associated with altered expression of inflammasome-related genes, particularly ASC and Caspase-1, in breast cancer patients. ROC and correlation analyses suggested moderate transcript-level discriminatory ability for ASC and Caspase-1, whereas NLRP3 demonstrated weaker performance. However, these findings were limited to plasma RNA expression and should not be interpreted as direct evidence of functional inflammasome activation or inflammatory activity. The observed changes may reflect systemic or compensatory transcriptomic responses rather than direct neuropathic mechanisms. Therefore, the biological and clinical significance of these findings remains uncertain. Future studies incorporating protein validation, Caspase-1 activity assays, tissue-specific and tumor microenvironment analyses, cytokine and immune-cell profiling, RNA sequencing, and direct comparisons between prophylactic and post-onset metformin administration are required to clarify the biological relevance of these observations.
Ethical Approval
The research ethics committee at the Ahvaz Jundishapur University of Medical Sciences adheres to the IR code of ethics.AJUMS.REC.1403.539 approved our work. All participants provided written informed consent, and the study was conducted in accordance with the Declaration of Helsinki.
Acknowledgment
The authors sincerely thank their esteemed colleagues at the Clinical Research Development Unit, Golestan Hospital, Ahvaz Jundishapur University of Medical Sciences, Ahvaz, Iran, for their invaluable collaboration and support.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Disclosure of AI Tool Use
This manuscript has utilized Generative AI tools in its preparation. Specifically, OpenAI’s ChatGPT was employed to refine language and generate summaries. This use aimed to enhance the clarity and coherence of the text while preserving the research’s integrity and originality.
Funding
The Vice Chancellor of Research Affairs at Ahvaz Jundishapur University of Medical Sciences provides financial support (Grant: CMRC-0324).
Disclosure
The authors declare that they have no conflicts of interest in this work.
References
1. Trapani D, Murthy SS. Progress in global breast cancer control. Curr Opin Oncol. 2025;37(6):529–13. doi:10.1097/CCO.0000000000001163
2. Chinedu SN, Ogbodo PN, Effiong ME. Disparities in diagnosis, treatment, and outcomes of early-onset breast cancer: a global perspective. Crit Rev Oncol Hematol. 2026;221:105251. doi:10.1016/j.critrevonc.2026.105251
3. Godiveau M, Jahanmohan JP, Abrial C, Durando X. Chemotherapy-induced peripheral neuropathy in patients with breast cancer treated with taxanes (Review). Oncol Lett. 2025;30(5):508. doi:10.3892/ol.2025.15254
4. Dalferth R, Hebbel H, Bauerschlag D, Letsch A, Schmidt T. Effects on chemotherapy-induced peripheral neuropathy by moderate strength training in combination with whole-body vibration in breast cancer patients. Support Care Cancer. 2025;33(11):970. doi:10.1007/s00520-025-09972-y
5. Zhi WI, Chen P, Kwon A, et al. Chemotherapy-induced peripheral neuropathy (CIPN) in breast cancer survivors: a comparison of patient-reported outcomes and quantitative sensory testing. Breast Cancer Res Treat. 2019;178(3):587–595. doi:10.1007/s10549-019-05416-4
6. Mahmoud AMAS, El Said NO, Shash E, Ateyya H. Prevention of paclitaxel-induced peripheral neuropathy: literature review of potential pharmacological interventions. Future J Pharm Sci. 2024;10(1):67. doi:10.1186/s43094-024-00638-w
7. Chen X, Gan Y, NPB A, CHE M. Current understanding of the molecular mechanisms of chemotherapy-induced peripheral neuropathy. Front Mol Neurosci. 2024;17:1345811. doi:10.3389/fnmol.2024.1345811
8. Mallik R, Chowdhury TA. Metformin in cancer. Diabet Res Clin Pract. 2018;143:409–419. doi:10.1016/j.diabres.2018.05.023
9. Xu J, Cai S, Zhao J, et al. Advances in the relationship between pyroptosis and diabetic neuropathy. Front Cell Dev Biol. 2021;9:753660. doi:10.3389/fcell.2021.753660
10. Yang F, Qin Y, Wang Y, et al. Metformin Inhibits the NLRP3 Inflammasome via AMPK/mTOR-dependent Effects in Diabetic Cardiomyopathy. Int J Biol Sci. 2019;15(5):1010–1019. doi:10.7150/ijbs.29680
11. Bakry HM, Mansour NO, ElKhodary TR, Soliman MM. Efficacy of metformin in prevention of paclitaxel-induced peripheral neuropathy in breast cancer patients: a randomized controlled trial. Front Pharmacol. 2023;14:1181312. doi:10.3389/fphar.2023.1181312
12. Ala M, Ala M. Metformin for cardiovascular protection, inflammatory bowel disease, osteoporosis, periodontitis, polycystic ovarian syndrome, neurodegeneration, cancer, inflammation and senescence: what is next? ACS Pharmacol Transl Sci. 2021;4(6):1747–1770. doi:10.1021/acsptsci.1c00167
13. Ge A, Wang S, Miao B, Yan M. Effects of metformin on the expression of AMPK and STAT3 in the spinal dorsal horn of rats with neuropathic pain. Mol Med Rep. 2018;17(4):5229–5237. doi:10.3892/mmr.2018.8541
14. Cao G, Gong T, Du Y, Wang Y, Ge T, Liu J. Mechanism of metformin regulation in central nervous system: progression and future perspectives. Biomed Pharmacother. 2022;156:113686. doi:10.1016/j.biopha.2022.113686
15. Fumagalli G, Monza L, Cavaletti G, Rigolio R, Meregalli C. Neuroinflammatory process involved in different preclinical models of chemotherapy-induced peripheral neuropathy. Front Immunol. 2020;11:626687. doi:10.3389/fimmu.2020.626687
16. Jia L, Zhou Y, Ma L, et al. Inhibition of NLRP3 alleviated chemotherapy-induced cognitive impairment in rats. Neurosci Lett. 2023;793:136975. doi:10.1016/j.neulet.2022.136975
17. Wedel S, Hahnefeld L, Schreiber Y, et al. SAFit2 ameliorates paclitaxel-induced neuropathic pain by reducing spinal gliosis and elevating pro-resolving lipid mediators. J Neuroinflammation. 2023;20(1):149. doi:10.1186/s12974-023-02835-5
18. Starobova H, Nadar EI, Vetter I. The NLRP3 inflammasome: role and therapeutic potential in pain treatment. Front Physiol. 2020;11:1016. doi:10.3389/fphys.2020.01016
19. Du Y, Zhu YJ, Zhou YX, Ding J, Liu JY. Metformin in therapeutic applications in human diseases: its mechanism of action and clinical study. Mol Biomed. 2022;3(1):41. doi:10.1186/s43556-022-00108-w
20. Chen MY, Ye XJ, He XH, Ouyang DY. The signaling pathways regulating nlrp3 inflammasome activation. Inflammation. 2021;44(4):1229–1245. doi:10.1007/s10753-021-01439-6
21. Blevins HM, Xu Y, Biby S, Zhang S. The NLRP3 inflammasome pathway: a review of mechanisms and inhibitors for the treatment of inflammatory diseases. Front Aging Neurosci. 2022;14:879021. doi:10.3389/fnagi.2022.879021
22. Zheng D, Liwinski T, Elinav E. Inflammasome activation and regulation: toward a better understanding of complex mechanisms. Cell Discov. 2020;6:36. doi:10.1038/s41421-020-0167-x
23. Kelley N, Jeltema D, Duan Y, He Y. The NLRP3 inflammasome: an overview of mechanisms of activation and regulation. Int J Mol Sci. 2019;20(13). doi:10.3390/ijms20133328
24. Faria SS, Costantini S, de Lima VCC, et al. NLRP3 inflammasome-mediated cytokine production and pyroptosis cell death in breast cancer. J Biomed Sci. 2021;28(1):26. doi:10.1186/s12929-021-00724-8
25. Deng Z, Lu L, Li B, Shi X, Jin H, Hu W. The roles of inflammasomes in cancer. Front Immunol. 2023;14:1195572. doi:10.3389/fimmu.2023.1195572
26. Wang X, Lin J, Wang Z, Li Z, Wang M. Possible therapeutic targets for NLRP3 inflammasome-induced breast cancer. Discov Oncol. 2023;14(1):93. doi:10.1007/s12672-023-00701-7
27. Roshan MH, Shing YK, Pace NP. Metformin as an adjuvant in breast cancer treatment. SAGE Open Med. 2019;7:2050312119865114. doi:10.1177/2050312119865114
28. Jia M, Wu C, Gao F, et al. Activation of NLRP3 inflammasome in peripheral nerve contributes to paclitaxel-induced neuropathic pain. Mol Pain. 2017;13:1744806917719804. doi:10.1177/1744806917719804
29. Ha J-S, Yeom Y-S, Jang J-H, et al. Anti-inflammatory effects of metformin on neuro-inflammation and NLRP3 Inflammasome activation in BV-2 microglial cells. Biomed Sci Letters. 2019;25(1):92–98. doi:10.15616/BSL.2019.25.1.92
30. Dong N, Lin T. Innate immunity in chemotherapy-induced peripheral neuropathy: recent advances. Front Pain Res. 2025;6:1642306. doi:10.3389/fpain.2025.1642306
31. Cristiano C, Giorgio C, Cocchiaro P, et al. Inhibition of C5aR1 as a promising approach to treat taxane-induced neuropathy. Cytokine. 2023;171:156370. doi:10.1016/j.cyto.2023.156370
32. Sharma A, Johnson KB, Sen A, et al. Longitudinal multi-omics analyses of chemotherapy-induced peripheral neuropathy in response to taxanes. BMC Cancer. 2025;25(1):1591. doi:10.1186/s12885-025-14901-7
33. Chen C, Smith MT. The NLRP3 inflammasome: role in the pathobiology of chronic pain. Inflammopharmacology. 2023;31(4):1589–1603. doi:10.1007/s10787-023-01235-8
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
The NLRP3 Inflammasome as a Novel Therapeutic Target for Cardiac Fibrosis
Fan J, Ren M, Adhikari BK, Wang H, He Y
Journal of Inflammation Research 2022, 15:3847-3858
Published Date: 7 July 2022
Extracellular Histones Activate Endothelial NLRP3 Inflammasome and are Associated with a Severe Sepsis Phenotype

Beltrán-García J, Osca-Verdegal R, Pérez-Cremades D, Novella S, Hermenegildo C, Pallardó FV, García-Giménez JL
Journal of Inflammation Research 2022, 15:4217-4238
Published Date: 25 July 2022
The Role of NLRP3 Inflammasome Signaling on Arrhythmias in Diabetes
Zhang L, Liu HH, Li F, Yang F, Qian LL, Wang RX
Journal of Inflammation Research 2022, 15:6883-6889
Published Date: 29 December 2022
The Mechanism of Pyroptosis and Its Application Prospect in Diabetic Wound Healing
Al Mamun A, Shao C, Geng P, Wang S, Xiao J
Journal of Inflammation Research 2024, 17:1481-1501
Published Date: 6 March 2024
Salmonella-NLRP3 Inflammasome Crosstalk: Host Defense Activation Versus Bacterial Immune Evasion Strategies
Li Y, Xu Y, Jin C, Qiu J, Jiao X, Pan Z, Guo Y
Journal of Inflammation Research 2025, 18:5133-5148
Published Date: 15 April 2025