Back to Journals » International Journal of Nanomedicine » Volume 21
Therapeutic Potential of Mesenchymal Stem Cell-Derived Extracellular Vesicles in Digestive System Tumors: Modulating the Chronic Inflammatory Microenvironment
Authors Kang H ![]() , Ding T, Lin H, Wang F, Zhu Y
, Ding T, Lin H, Wang F, Zhu Y ![]() , Chen P, Chen H, Huang J, Wang X
, Chen P, Chen H, Huang J, Wang X
Received 2 February 2026
Accepted for publication 10 May 2026
Published 18 May 2026 Volume 2026:21 600593
DOI https://doi.org/10.2147/IJN.S600593
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Prof. Dr. Anderson Oliveira Lobo
Huapei Kang,1,2,* Tao Ding,1– 3,* Haihong Lin,2,3 Fangsheng Wang,1– 3 Yifan Zhu,2,3 Puwen Chen,1,2 Hanlu Chen,2,3 Junyun Huang,1– 3 Xiaoling Wang1– 3
1The First School of Clinical Medicine, Gannan Medical University, Ganzhou, People’s Republic of China; 2Laboratory Medicine, First Affiliated Hospital of Gannan Medical University, Ganzhou, People’s Republic of China; 3College of Medical Technology, Gannan Medical University, Ganzhou, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Xiaoling Wang, The First School of Clinical Medicine, Gannan Medical University, Ganzhou, People’s Republic of China, Email [email protected]
Abstract: Digestive system tumors (including colorectal cancer, hepatocellular carcinoma, and gastric cancer) have become one of the most serious global cancer burdens, and their pathological evolution and chronic inflammatory microenvironments are closely related. Chronic inflammation moves into cancer by angling immune cell infiltration, pro-inflammatory stuff, and aberrant signaling pathways, which advance tumor cell growth factor and burst, hitching a ride. In this context, mesenchymal stem cell-derived extracellular vesicles (MSC-EVs)—naturally occurring nan vesicles enveloped by a bilipidic layer—are loaded with bioactive substances like proteins, miRNAs, which present considerable promise in remodeling the chronic inflammatory microenvironments and arresting tumor development. MSC - EVs have the benefits of low risk of being immunogenic, good biological compatibility, and targeted homing capabilities; however, the main aspect is that they adjust immune cells and interfere with key signaling systems using the bioactive materials that they carry. This can be carried out to modulate the chronic inflammatory environment, and it is also carriers to carry specific loads (miRNAs, chemotherapeutic drugs, etc.) to inhibit the tumor cell biological behavior. However, the potential pro-tumorigenic effects of MSC-EVs under specific conditions cannot be overlooked, as they may also transport cargos that accelerate tumor progression. Similarly, EVs derived from non-mesenchymal sources have been shown to drive malignancy through various signaling mechanisms. Although MSC-EVs have great clinical value, there are still problems such as the standardization of preparation, the standardization of heterogeneity control, and the standardization of safety evaluation. Current engineering modification strategies can enhance the effectiveness of engineering. In the future, innovative combinations of technologies and approaches may even lead to more rapid progress toward translation from the bench to the bedside. This paper mainly discusses MSC-EVs regulation mechanism towards chronic inflammation, and the potential for it to inhibit or promote tumor growth, as well as conducting a systematic review of current research on MSC-EVs as a treatment agent for digestive system tumors. Finally, It outlines challenges in clinical application and future prospects.
Keywords: chronic inflammation, digestive system tumors, mesenchymal stem cells, extracellular vesicles, immunomodulation, targeted therapy
Introduction
In recent years, digestive system tumors have become an important part of the world’s cancer burden. From the view of epidemiology, digestive system cancers are the main sources in terms of incidence and death from these kinds of cancers: colorectal cancer (CRC), hepatocellular carcinoma (HCC), and gastric cancer (GC).1 In spite of the various causes for tumorigenesis and development, chronic inflammation plays a bigger part in raising cancer threats. Its role is very important in the beginning of a tumor. It helps a tumor get started and makes it grow worse. Inflammatory cells exert their influence on the surrounding tumor cells by continuously emitting chemokines, reactive nitrogen species (RNS), and reactive oxygen species (ROS).2 In a certain pathologic evolution of digestive system tumors, chronic inflammation has become a major factor in tumor initiation and progression. Inflammatory bowel disease (IBD) is a major player in initiating and promoting CRC. Persistent chronic inflammation in IBD triggers oxidative stress, which cranks out massive amounts of ROS and RNS via NADPH oxidase and NOS, directly causing oxidative DNA damage. This inflammatory microenvironment does more than just interfere with DNA repair and cause mutations in key tumor suppressors like p53; it also activates oncogenic pathways. By driving abnormal cell growth and clonal expansion, these processes ultimately fuel the progression toward colorectal cancer.3 In HCC, hepatic inflammation causes HCC to start and grow through various different ways. It promotes the survival and spread of HCC stem cells through pathways such as the IL-6/Signal Transducer and Activator of Transcription 3(STAT3) pathway, and also causes DNA damage and genetic disorders by producing ROS and RNS, which leads to the creation of an environment where genetic changes linked with HCC development are favorable.4 It is the same link between chronic gastritis caused by Helicobacter pylori (H. pylori) and GC. The cause is that H. pylori activates the NOD1/nuclear factor κB (NF-κB) pathway, driving the release of pro-inflammatory factors such as IL-8 and creating a chronic inflammatory environment. This chronic inflammatory state leads to severe damage to the gastric mucosa, which in turn induces gastric atrophy and ultimately progresses to gastric cancer.5 Currently, surgical resection remains the primary curative approach for digestive system tumors. However, clinical practice has evolved toward a multimodal integrated management model, which systematically incorporates chemotherapy and targeted therapy based on pathological staging alongside early intervention.6 In addition to the current treatment approaches, preventing and treating digestive system cancers by controlling prolonged inflammation has become a major prevention and cure method.7
Due to the significant effects on the immune system, the therapy using mesenchymal stem cells (MSCs) has received extensive research. A lot of preclinical research points out that the good effects MSCs bring about are mostly because of their paracrine mechanisms. As the major paracrine mediators, extracellular vesicles derived from mesenchymal stem cells (MSC-EVs) function by transporting biomolecules such as nucleic acids and proteins to participate in the regulation of inflammation.8 In terms of existing studies, it’s well established that MSC-EVs inhibit tumor proliferation and metastasis through immunomodulation and antitumor responses as well as the potential to become a good drug carrier for targeted drug delivery.9 In short, MSC-EVs have excellent clinical application potential to reduce tumor growth and to prevent tumorigenesis in the chronic inflammatory microenvironment and to prevent and treat the tumor directly. The focus should be placed on exploring the specific mechanisms by which MSC-EVs regulate chronic inflammation, analyzing their dual roles in tumor progression, and, based on current research advances, thoroughly examining the challenges and engineering optimization strategies encountered during their transition from basic research to clinical application.
Molecular Composition and Biological Functions of EVs
Intercellular signaling is crucial for multicellular organisms to maintain order, ensure normal functioning, and respond to pathological changes. Extracellular vesicles (EVs) are the main carriers of intercellular signaling system. According to their origin and size, extracellular vesicles can be classified into: exosomes, microvesicles, and apoptotic bodies.10 However, given that most current isolation techniques struggle to physically distinguish exosomes from other sources of nanovesicles, the latest guidelines published by the International Society for Extracellular Vesicles (ISEV) recommend using “extracellular vesicles” as a general umbrella term.11 Therefore, although many original studies cited in this review employ the term “exosomes,” to ensure terminological rigor, this paper will consistently use “mesenchymal stem cell-derived extracellular vesicles (MSC-EVs)” hereafter. Extensive research has fairly thoroughly explained the biological traits and functions of EVs. The secretion of EVs by parental cells is a complex physiological process, intricately regulated through mechanisms involving endocytosis and intracellular sorting. EVs are formed during biogenesis via the multivesicular body (MVB) pathway, Early endosomes are created by endocytic invagination or fusion with vesicles originating from the Golgi apparatus. They then develop into late endosomes, producing intraluminal vesicles (ILVs). And what’s more, MVB fuse with the plasma membrane to release ILVs as EVs. This process relies on the coordinated regulation of the endosomal sorting complex required for transport (ESCRT) machinery and non-ESCRT-dependent pathways.12 The outer membrane of EVs consists of a lipid bilayer. This membrane is composed of various lipids, including cholesterol, sphingomyelin, and phosphatidylethanolamine. These lipid components not only maintain membrane rigidity but also actively participate in cargo sorting and intercellular signaling processes.13 Bioactive molecules are a large part of what can be found inside EVs. Proteins can be classified into many kinds, such as the transmembrane protein family (eg, CD9, CD63, CD53, CD81), bioactive enzymes, and signaling proteins Nucleic acids are of different kinds, including mRNA, miRNA, and circular RNA.14 The main biological function of EVs is to serve as a medium of intercellular communication, carrying bioactive molecules contained within the EVs inside the compartments of the EVs and targeted towards and recognized by the cell, so as to exert control over the regulation of physiological or pathological processes in such cells. After EVs recognize receptor cells, there are three ways that the target cells could be activated by the EVs. The first pathway involves ligands on the surface of EVs binding to receptors on the surface of target cells, thereby activating downstream signaling pathways within the target cells.15 The second pathway involves EVs directly releasing their cargo into the cytoplasm of target cells through membrane fusion.16 The third pathway involves target cells directly engulfing entire EVs through endocytosis, after which the cargo of the EVs is released directly into the cytoplasm.17 Endocytosis is one of the primary pathways for EVs to enter cells (Figure 1). The pathways by which EVs enter cells are influenced by surface molecules on EVs, target cell characteristics, and the microenvironment where EVs are present.18 After entering cells, EVs deliver their bioactive molecules to target cells. These bioactive molecules influence the physiological activities of target cells by regulating gene expression, altering signaling pathways, and inducing immune responses. In the field of medical research, based on the core biological functions of MSC-EVs and their advantages of excellent biocompatibility and low immunogenicity, MSC-EVs are employed as natural carriers for therapeutic drugs. They achieve specific therapeutic effects by targeting and delivering specific therapeutic drugs or engineered bioactive molecules into cells.
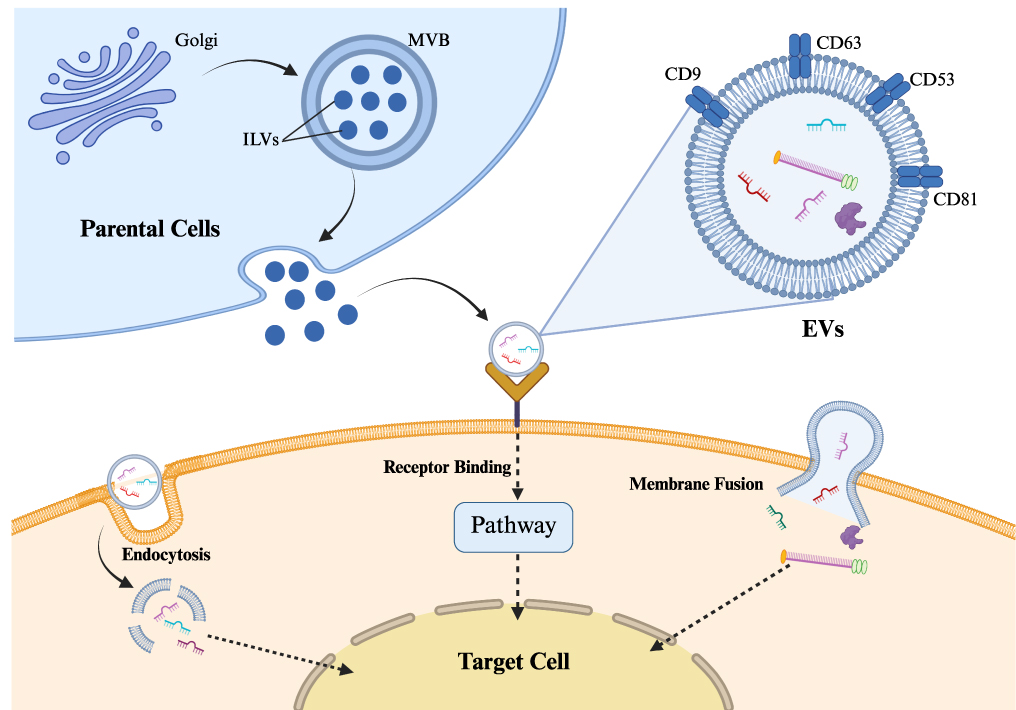 |
Figure 1 Biogenesis, molecular composition, and cellular uptake pathways of EVs.The schematic illustrates the release of EVs from donor cells via the multivesicular body (MVB) pathway, their structure containing bioactive cargo (proteins, nucleic acids) and surface markers (eg, CD9,CD53, CD63, CD81), and the three primary mechanisms for interacting with target cells: endocytosis, receptor binding, and membrane fusion. |
Inflammatory Diseases of the Digestive System: A Microenvironment Suitable for Digestive System Tumors
Major Mechanisms of the Chronic Inflammatory Microenvironment
Inflammation is a protective response that occurs when the body encounters various exogenous and endogenous injury factors that cause damage to cells, tissues, and organs. The inflammatory response is characterised by a series of complex reactions, including the elimination of injurious factors, the absorption of necrotic tissue and cells, and the repair of damage.19 Inflammation can be classified into two distinct types: acute inflammation and chronic inflammation. When the mechanisms responsible for resolving inflammation become imbalanced, the inflammatory response fails to subside, leading to a persistently activated inflammatory state. This type of inflammatory response is termed chronic inflammation.20 Chronic inflammation has been found in multiple systems of the human body, and there is strong evidence indicating that it is a clear risk factor for the development of various tumors of the digestive system. The inflammatory microenvironment, which is shaped by chronic inflammation, contains a variety of immune cells, including T cells, B cells, and macrophages. These immune cells activate signaling pathways such as NF-κB/JAK-STAT by secreting cytokines, including IL-6 and TNF-α. This process consequently drives the biological behaviors associated with tumor initiation, proliferation, migration, and invasion.21 The NF-κB and STAT3 signaling pathways play a major role in the transition from chronic inflammation to tumors. Extracellular inflammatory signals are converted into intracellular pro-tumor programs through these pathways, specifically by synergistically regulating the expression of tumor-promoting genes, including angiogenesis-related genes, chemokines, and immunosuppressive cytokines.22 During the process of tumour development, the NF-κB signaling pathway confers anti-apoptotic properties to tumour cells. This is achieved through the direct regulation of the expression of anti-apoptotic genes, including c-IAPs, c-FLIP, A1/Bfl1, TRAF1, and TRAF2. This enables tumour cells to evade programmed cell death, thereby facilitating their massive proliferation.23 It has been shown that the NF-κB signaling pathway can also induce the expression of vascular endothelial growth factor (VEGF) and matrix metalloproteinases, thereby promoting tumor angiogenesis.24 The STAT3 signaling pathway and the NF-κB signaling pathway form a synergistic relationship through a positive feedback loop. Specifically, NF-κB activates STAT3 by inducing IL-6, while STAT3 continuously enhances NF-κB activity by upregulating the S1PR1 receptor, thereby establishing a positive feedback cycle.25 In addition, abnormally activated immune cells in the chronic inflammatory microenvironment can secrete large amounts of ROS and RNS, which can directly damage DNA, leading to base modifications, DNA single-strand or double-strand breaks, and chromosomal aberrations.26 Chronic inflammation can also reprogram cellular epigenetics, primarily manifested in two ways: On one hand, pro-inflammatory cytokines such as IL-1β, TNF-α and ROS induce hypermethylation of CpG islands in tumor suppressor gene promoters by recruiting DNA methyltransferase and histone methyltransferase complexes, thereby leading to transcriptional silencing. On the other hand, chronic inflammation also induces genome-wide hypomethylation and abnormal histone modifications, further promoting the aberrant activation of oncogenes.27 The inflammatory response is a series of reactions designed to protect cells, tissues, and organs from exogenous or endogenous damage. However, the inflammatory microenvironment shaped by prolonged chronic inflammation promotes tumor initiation and progression. Selected digestive system tumors will be discussed in detail in the following sections.
Chronic Inflammatory Diseases and CRC
IBD is a classic example of chronic inflammation in the pathological evolution of colorectal diseases. This condition causes recurrent damage to the intestinal mucosa, leading to persistent inflammatory responses. Its progression involves a complex pathological process encompassing three main factors: dysregulation of immune responses, disruption of the gut microbiota, and genetic susceptibility.28 Immune dysregulation represents the primary mechanism underlying the pathological progression of chronic inflammation. This mechanism manifests primarily as dysfunction of immune cells, characterized by excessive activation of Th17 cells and insufficient function of regulatory T cells (Tregs).29 Dysregulation of immune cells results in ongoing immune attacks on the intestinal mucosa. Factors that promote inflammation, such as TNF-α, IL-6, and IFN-γ, are highly expressed in chronic inflammation. These pro-inflammatory factors enhance vascular permeability, enabling immune cells to migrate to the site of inflammation and intensify the inflammatory response.30 In addition, increased levels of IL-17 secreted by Th17 cells can induce the production of proinflammatory cytokines such as IL-6 and TNF-α by activating the mitogen-activated protein kinase (MAPK) pathway.31,32 Long-term exposure to an inflammatory microenvironment significantly increases the risk of carcinogenesis in colorectal tissues.33 From the perspective of pathological evolution, IBD-associated CRC follows the sequence of “inflammation—dysplasia—carcinogenesis.” Chronic inflammation drives the accumulation of gene mutations, ultimately leading to cancer development.34 The inflammatory response subjects intestinal mucosal cells to persistent immune attacks, leading to abnormal proliferation of mucosal cells.35 Mucosal cell dysplasia is classified into low-grade dysplasia and high-grade dysplasia. Both types represent well-defined precancerous lesions.36 High-grade dysplasia may progress to invasive adenocarcinoma.37 Furthermore, at the genetic level, the release of large amounts of ROS or RNS by immune cells can directly damage DNA, thereby affecting the function of key genes.38 ROS directly causes point mutations or deletions in the tumor suppressor gene p53, causing it to lose its DNA repair and apoptosis regulation functions.39 In addition, studies have shown that mutations in the Wingless-type MMTV integration site family (Wnt) pathway negative regulator gene RNF43 lead to sustained activation of Wnt signaling, thereby upregulating the expression of oncogenes such as c-Myc.40 In an animal model study, mice with mutations in the Wnt pathway regulatory gene APC accelerated tumor differentiation within an inflammatory microenvironment.41
Chronic Inflammatory Diseases and HCC
Chronic inflammation manifests as chronic hepatitis in the liver. The aetiology of chronic hepatitis is multifarious and intricate. Common contributory factors encompass persistent infection with hepatitis B virus (HBV) or hepatitis C virus (HCV), long-term alcoholic liver disease, and non-alcoholic fatty liver disease.42 The chronic inflammatory microenvironment that is established due to chronic hepatic inflammation has been shown to increase the risk of HCC occurrence and progression. This chronic hepatic inflammatory microenvironment is characterized by persistent hepatocyte injury and death alongside compensatory regeneration, resulting in complex genetic alterations.43 From the perspective of pathological progression, hepatocellular carcinoma associated with chronic hepatitis follows a typical sequence of “inflammation — liver fibrosis — cirrhosis — carcinogenesis”.44 Immune dysregulation has been identified as the primary mechanism underlying the evolutionary sequence of chronic inflammation that contributes to liver carcinogenesis. Hepatocellular stellate cells, which act as key mediators in chronic inflammation, have been shown to actively recruit myeloid-derived suppressor cells (MDSCs) via the SDF-1/CXCR4 axis. This process has been found to result in immune cell functional exhaustion and immune evasion.45 In the context of chronic inflammation, a variety of immunosuppressive cell subsets, such as Tregs and MDSCs, are present. These immunosuppressive cells secrete cytokines, including IL-10 and TGF-β, which have been shown to suppress antitumor immune responses.46,47 In addition, TNF-α and IL-6, which are secreted by immune cells or hepatic parenchymal cells, are persistently overexpressed in the chronic inflammatory microenvironment. This enhances inflammatory responses and activates key signaling pathways, thereby promoting malignant proliferation of hepatocytes.48 The NF-κB and MAPK signaling pathways are crucial signaling pathways in the progression of inflammation to hepatocellular carcinoma. Cytokines such as TNF-α and IL-6 suppress the normal apoptotic program of hepatocytes by activating the NF-κB signaling pathway, thereby upregulating the expression of anti-apoptotic genes like Bcl-2.49 By activating the MAPK signaling pathway, particularly the c-Jun N-terminal kinase (JNK) signaling pathway. Research indicates that the JNK1 signaling pathway promotes the development of hepatocellular carcinoma by inducing malignant proliferation and selective cell death in hepatocytes, thereby facilitating tumor formation. It also enhances cancer cell growth by upregulating pro-proliferative factors such as cyclin D1.50 In addition, another study indicates that the TOX3 protein promotes the initiation, progression, and metastasis of hepatocellular carcinoma by targeting key molecules in the MAPK signaling pathway, such as p-ERK1/2.51 In addition, immune cells and damaged liver cells produce large amounts of ROS and RNS, which can directly damage DNA, leading to DNA strand breaks, base modifications, and other injuries, thereby triggering gene mutations and genomic instability.52
Chronic Inflammatory Diseases and GC
The development of GC is also highly correlated with long-term chronic inflammation. The progression from chronic inflammation to GC follows a stepwise pathological process: non-atrophic gastritis—atrophic gastritis—intestinal metaplasia—dysplasia—invasive carcinoma.53 H. pylori infection promotes DNA damage and malignant proliferation in gastric mucosal epithelial cells through pathways such as upregulating spermidine oxidase and increasing the production of acrolein, a bacterial metabolite, thereby accelerating the malignant transformation of gastric mucosal epithelial cells.54 In the progression from gastritis to GC, persistent dysregulation of immune function serves as a key factor, with the imbalance in immune mechanisms acting as the initiating step in this evolutionary process. In chronic gastritis induced by factors such as H. pylori infection, the gastric mucosal epithelium recruits and activates a large number of immune cells (eg, neutrophils, macrophages, T cells). These immune cells release substantial amounts of pro-inflammatory cytokines (eg, IL-8, IL-1β, TNF-α), chemokines, and growth factors.55 Cytokines such as IL-8, IL-1β, and TNF-α enhance vascular permeability and also attract more immune cells to migrate to the site of inflammation, creating a vicious cycle.56 A study found that the IL-1β signaling pathway activates CREB and C/EBPβ transcription factors by regulating ERK1/2-dependent mechanisms. This process consequently induces gastric cancer cell proliferation and enhances their survival rate.57 In addition, H. pylori infection represents a pivotal external factor. Lipopolysaccharide (LPS) constitutes a component of the outer membrane of H. pylori. It is recognized by Toll-like receptors (TLRs) situated on the surface of gastric epithelial cells and immune cells. Following binding, it activates downstream immune signaling pathways. A study has shown that the binding of LPS to Toll-like receptor 4 (TLR4) on the surface of gastric epithelial cells and immune cells activates the NF-κB pathway, thereby enhancing the inflammatory response.58 In addition, the virulence factor Cytotoxin-associated gene A (CagA) carried by H. pylori can exert immunosuppressive effects by influencing multiple signaling pathways, ultimately leading to malignant proliferation of tumor cells, promoting tumor angiogenesis, and enhancing cellular invasiveness. CagA promotes β-catenin release and transcriptional activation by activating the phosphoinositide 3-kinase/protein kinase B pathway (PI3K/AKT) and the Wnt/β-catenin pathway, thereby driving cell proliferation, morphological changes, and migration.59 Under the combined influence of endogenous and exogenous factors, the genomes of gastric epithelial cells persistently exposed to an inflammatory microenvironment sustain damage. Neutrophils and macrophages, among other immune cells, secrete large amounts of ROS and RNS, which can directly induce DNA damage. This leads to the inactivation or abnormal activation of key genes, thereby facilitating the accumulation of genetic mutations that contribute to carcinogenesis.60 (Table 1 and Figure 2)
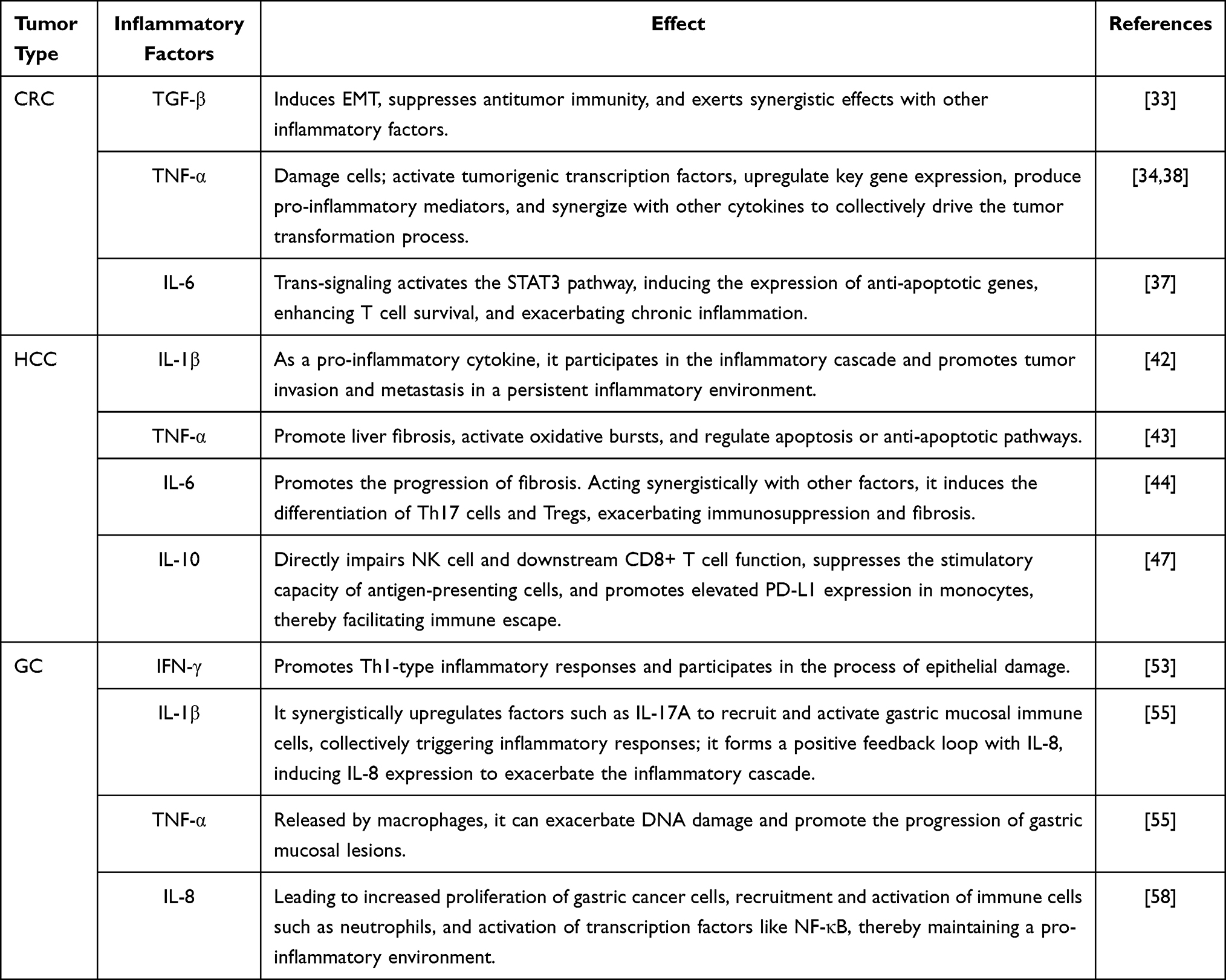 |
Table 1 Roles of Key Inflammatory Factors in the Microenvironment of Digestive System Tumors |
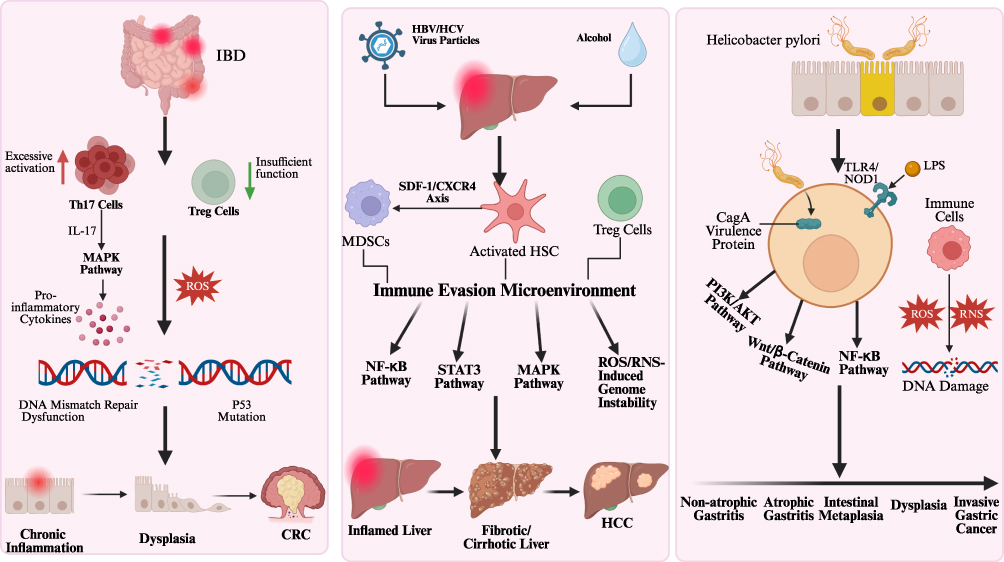 |
Figure 2 Schematic representation of the transition from chronic inflammation to tumorigenesis in digestive system tumors.The diagram illustrates the distinct pathological progressions for CRC, HCC, and GC, highlighting key drivers such as immune dysregulation, activation of oncogenic signaling pathways and genetic instability induced by oxidative stress and pathogens.Upward and downward arrows indicate as labeled; arrows without text labels represent the direction of signaling transduction/flow. |
Mechanism of Action of MSC-EVs in Treating Chronic Inflammation
MSC-EVs’ Role in Regulating Immune Cells
MSC-EVs can modulate the inflammatory microenvironment by carrying various bioactive molecules (miRNA, proteins, lipids, etc), thereby exerting anti-inflammatory, anti-fibrotic, and tissue-repairing effects.61 MSC-EVs can regulate immune cells (such as T cells, B cells, and macrophages) within the inflammatory microenvironment through multiple mechanisms. Macrophages are one of the primary immune cell types in the inflammatory microenvironment. Based on their functional properties, they can be classified into pro-inflammatory M1 and anti-inflammatory M2 subtypes. MSC-EVs improve the chronic inflammatory microenvironment by regulating the polarization of macrophages from the M1 to the M2 phenotype.62 Studies have shown that in a mouse model of colitis, MSC-EVs can alleviate colitis symptoms by inducing M2b-type polarization in macrophages and enhancing their ability to secrete the anti-inflammatory factor IL-10, thereby suppressing the production of pro-inflammatory factors (IL-1β, IL-6, TNF-α).63 MSC-EVs also promote macrophage M2 polarization by delivering miR-182 to target and suppress TLR4 expression.64 In an in vitro experimental study, MSC-EVs carrying miR-148a significantly reduced the expression levels of M1 macrophage markers (iNOS, TNF-α, IL-6) while simultaneously upregulating the expression levels of M2 markers (IL-10, CD206, Arg-1).65 Tregs are a key T cell subset that maintains immune homeostasis and suppresses autoimmune responses. In inflammatory microenvironments, Tregs exhibit reduced numbers or functional inactivation. Research indicates that MSC-EVs can induce Treg generation. MSC-EVs contain TGF-β1, which promotes the expression of the Foxp3 transcription factor, thereby inducing Treg production.66 Furthermore, in another study, bone marrow-derived MSC-EVs effectively promoted Treg cell differentiation and functional recovery by delivering JKAP protein through the AKT/Extracellular Signal-Regulated Kinase (ERK) signaling pathway.67 Dendritic cells initiate immune responses through antigen presentation. MSC-EVs block dendritic cells maturation and attenuate their ability to activate T cells by upregulating IL-10 and TGF-β while simultaneously suppressing the expression of co-stimulatory molecules such as CD40, thereby inducing immune tolerance.68 Overall, MSC-EVs systematically regulate immune cell populations within the inflammatory microenvironment through their bioactive molecules, redirecting the originally imbalanced immune response back to a balanced state.
Effects of MSC-EVs on Inflammation-Related Signaling Pathways
In the chronic inflammatory microenvironment, a complex network of cellular regulation is formed by a series of intricate signaling pathways. MSC-EVs can regulate inflammatory signaling by intervening in key steps of these pathways. A Study has shown that MSC-EVs regulate the NF-κB signaling pathway by delivering miR-146a, which can target and suppress the expression of TRAF6 and IRAK1. This process results in the inhibition of NF-κB activation and the alleviation of inflammatory responses.69 Another study has found that MSC-EVs exert a direct influence on TLR4 expression by delivering miR-145-5p, thereby inducing inhibition of NF-κB activation.70 In addition, MSC-EVs have been shown to inhibit the phosphorylation of p38 mitogen-activated protein kinase (p38 MAPK) within the MAPK family through the delivery of specific miRNAs, thereby slowing the progression of chronic inflammation. For example, in the chronic inflammatory process associated with intervertebral disc degeneration, there is overexpression of the LRG1 protein, which leads to the activation of the p38 MAPK signaling pathway, as evidenced by elevated levels of phosphorylated p38 protein, miR-129-5p is a key cargo of MSC-EVs that directly targets and suppresses LRG1 protein expression, thereby inhibiting phosphorylation within the p38 MAPK signaling pathway.71 MSC-EVs can also affect signaling pathways related to anti-inflammation and tissue repair. Adipose tissue-derived MSC-EVs promote PI3K/AKT signaling pathway activation by carrying miR-126, which enhances phosphorylation of the core protein Akt.72 This mechanism inhibits endothelial cell apoptosis and enhances cellular survival capacity. In another study, adipose tissue-derived MSC-EVs stimulated osteoblast proliferation and differentiation by carrying miR-136-5p, which suppressed the expression of LDL receptor-related protein 4 (LRP4)—a downstream target gene of the Wnt/β-catenin pathway—thereby promoting fracture healing.73 MSC-EVs simultaneously regulate inflammation-related signaling pathways through a multi-level, multi-target approach, thereby achieving the suppression of pro-inflammatory signals, activation of anti-inflammatory signals, and restoration of repair and regeneration-related signaling pathways. (Figure 3 and Table 2)
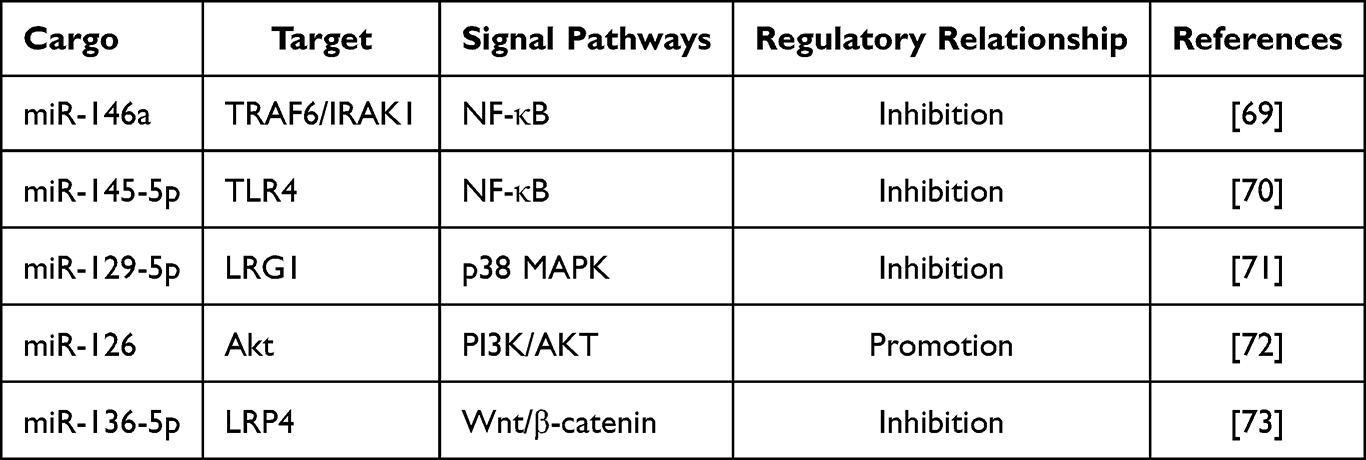 |
Table 2 Regulatory Effects of MSC-EV Cargo on Signaling Pathways |
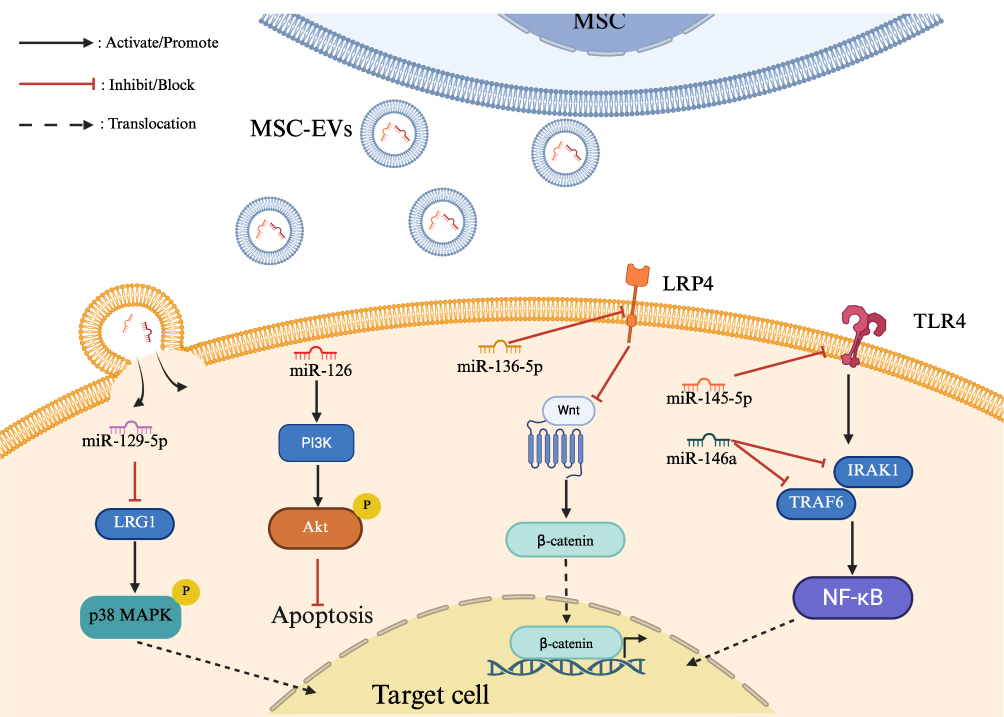 |
Figure 3 Molecular mechanisms of MSC-EV-mediated regulation of inflammation-related signaling pathways.The diagram illustrates how specific miRNAs delivered by MSC-EVs target key molecules (eg, LRG1, TLR4, TRAF6) to modulate downstream signaling cascades, including the inhibition of pro-inflammatory NF-κB and p38 MAPK pathways, and the activation of cell survival and repair pathways such as PI3K/Akt and Wnt/β-catenin. |
Progress in Research on MSC-EVs for Treating Digestive System Tumors
Advantages of MSC-EVs in Tumor Therapy
MSC-EVs demonstrate tremendous potential as natural nanoscale delivery vehicles. MSC-EVs derived from human sources exhibit low immunogenicity due to their biomimetic membrane structure and reduced membrane-bound protein content, significantly reducing the risk of triggering severe immune rejection reactions and thereby substantially enhancing treatment safety.74 The intrinsic low immunogenicity of MSC-EVs renders it superior to most synthetic carriers in numerous treatment regimens. In addition, the structural integrity of EVs is maintained by the lipid bilayer that envelops them, thus safeguarding the integrity of their internal cargo during circulation.75 It is also noteworthy that MSC-EVs demonstrate superior inherent capabilities for crossing biological barriers. Given their natural bilayer lipid structure and abundant surface adhesion proteins, MSC-EVs possess the potential to penetrate the blood-brain barrier.76 Moreover, MSC-EVs have been shown to be capable of encapsulating a variety of therapeutic molecules. Utilising engineering approaches allows MSC-EVs to carry specific mRNA or modified siRNA, thus enabling functional delivery within target cells.77 This phenomenon can be attributed to the homing properties exhibited by MSC-EVs, which facilitate their migration to tumor sites and areas of inflammation. Consequently, they are well-suited to serve as carriers for the precise delivery of encapsulated, specific miRNAs to tumor tissues.78 In addition to their role as carriers, the bioactive molecules transported by MSC-EVs have been shown to possess the capacity to directly suppress tumors. These effects are achieved by the regulation of complex signaling networks, thereby inhibiting tumor biology. Angiogenesis plays a key role in tumor growth and metastasis. Tumor blood vessels supply oxygen and nutrients to tumor cells while aiding in the removal of metabolic waste products.79 A study shows that MSC-EVs can downregulate the expression of key pro-angiogenic factors by targeting VEGF through the miR-16 they carry. This process subsequently inhibits the formation of new blood vessels in tumor tissues.80 Epithelial-mesenchymal transition (EMT) is a pivotal dynamic process in cell biology. This process entails the loss of polarity and intercellular junctions by epithelial cells through specific signaling pathways, resulting in their transformation into migratory mesenchymal cells. EMT is reactivated during tumorigenesis, thereby conferring metastatic properties to cancer cells. These properties include enhanced motility, invasiveness, and resistance to apoptotic stimuli.81 MSC-EVs can suppress tumor cell migration and invasion capabilities by adjusting the EMT process, thereby effectively curbing distant metastasis of tumors.82 Immunosuppression represents a primary mechanism through which tumors are able to evade the immune system. MSC-EVs have been shown to regulate immune cell activation and functional states by delivering specific miRNAs to immune cells, including T cells and natural killer cells.83 MSC-EVs exhibit significant therapeutic potential across diverse tumor subtypes. Although they excel at delivering therapeutic payloads, evidence also indicates suggests the potential of MSC-EVs to promote cancer progression. Within specific tumor microenvironments(TME), MSC-EVs facilitate tumor proliferation, EMT and angiogenesis. By modulating various signaling pathways, these vesicles significantly accelerate oncogenic progression.84 Particularly within the digestive system, evidence indicates that MSC-EVs exert dual regulatory effects on tumor progression, contingent upon specific microenvironmental conditions or their distinct molecular cargo.
Progress in MSC-EVs Treatment Research for CRC
Current research on MSC-EVs for the treatment of CRC reveals a concentration on two distinct aspects: the regulation of tumor cell behavior and the utilization of EVs as vehicles for drug delivery. On the one hand, the miRNAs carried by MSC-EVs can directly regulate tumor cell behavior. In a study, human bone marrow-derived MSC-EVs were found to be rich in miR-22-3p. This microRNA directly targets and suppresses the expression of the RAP2B protein, thereby downregulating the activation level of the downstream key PI3K/AKT signaling pathway.85 Ultimately, this inhibits the proliferation, migration, and invasive capabilities of CRC cells. In another study, integrin alpha 2 (ITGA2), a key molecule mediating cell-extracellular matrix adhesion, was found to be highly expressed and closely associated with tumor invasion and metastasis. Bone marrow-derived MSC-EVs overexpressing miR-16-5p restricted CRC progression by downregulating ITGA2.86 In addition, studies have indicated that MSC-EVs derived from human umbilical cords can directly target and bind to ITGA6 by carrying miR-3940-5p, thereby inhibiting the TGF-β1 signaling pathway and ultimately suppressing the invasion, metastasis, and EMT of CRC cells.87 On the other hand, MSC-EVs can serve as natural drug delivery carriers. Through engineered modifications, they can carry specific miRNAs or chemotherapy drugs and deliver them directly to CRC cells for targeted therapeutic effects. A study has demonstrated that the fusion of MSC-EVs with folate-modified liposomes to form hybrid extracellular vesicles and subsequent loading with paclitaxel significantly enhances drug delivery efficiency. The hybrid system under investigation exhibited folate-enhanced tumor targeting in colorectal cancer models. The system achieved this enhanced targeting by activating CD4+/CD8+ T cells, promoting M1 macrophage polarization, and suppressing Tregs cells. These mechanisms acted in a synergistic manner, resulting in the significant enhancement of antitumor efficacy.88 In contrast to the anti-tumor effects of MSC-EVs, certain studies have highlighted the pro-tumorigenic roles of CRC cell-derived EVs in disease progression. This dichotomy underscores the inherent potential of EVs to facilitate CRC development, depending significantly on their parental cell origin. Hypoxia represents a hallmark of the CRC tumor microenvironment. Hypoxia significantly boosts hypoxia-inducible factor-1α (HIF-1α) expression in CRC cells, leading them to release more EVs packed with miR-210-3p. These EVs are then taken up by nearby normoxic tumor cells. Once inside, the exosomal miR-210-3p directly targets and shuts down the tumor suppressor CELF2. This loss of CELF2 drives the G1-to-S phase transition and ramps up cell proliferation, while at the same time blocking apoptosis.89 Beyond localized effects, CRC-derived EVs play a pivotal role in orchestrating distant metastasis by priming the pre-metastatic niche. Evidence shows that EVs released by CRC cells, packed with miR-221 and miR-222, can home to the liver. Once there, they shut down SPINT1 expression in hepatic stellate cells and trigger a massive secretion of hepatocyte growth factor. This remodeling of the hepatic landscape promotes a pro-tumorigenic microenvironment that helps recruit circulating tumor cells and facilitates the establishment of metastatic lesions.90
Progress in MSC-EVs Treatment Research for HCC
A large body of preclinical research evidence indicates that MSC-EVs possess the ability to suppress HCC through a variety of mechanisms. For example, bone marrow-derived MSC-EVs significantly inhibit hepatocellular carcinoma cell proliferation and induce apoptosis by regulating cell cycle-related genes.91 In another study, bone marrow-derived MSC-EVs carry the lncRNA C5orf66-AS1, competitively bind to miR-127-3p to attenuate its inhibitory effect on DUSP1, thereby suppressing ERK pathway activation.92 This ultimately inhibits the proliferation, migration, and invasive capabilities of HCC stem cells. MSC-EVs can regulate the EMT of hepatocellular carcinoma cells by delivering specific miRNAs. Research has demonstrated that MSC-EVs derived from human umbilical cord inhibit the expression of ADAM10 by delivering miR-451a, thereby suppressing the EMT process and ultimately reducing the migration and invasion capabilities of hepatocellular carcinoma cells.93 Research has also demonstrated that MSC-EVs can exert therapeutic effects by targeting hepatocellular carcinoma cells through engineered modifications such as targeted peptide conjugation or miRNA transfection. Adipose tissue-derived MSC-EVs modified with miR-122 induce apoptosis and cell cycle arrest in hepatocellular carcinoma cells by downregulating the expression of ADAM10, IGF1R, and CCNG1 genes.94 In contrast to the inhibitory effects of MSC-EVs, EVs derived from highly metastatic hepatocellular carcinoma cells promote tumor progression. These cells exhibit significant oncogenic potential by releasing EVs that transport the calcium-binding protein S100A4 to low-metastatic recipient cells. S100A4 within the EVs triggers the phosphorylation and activation of the STAT3 signaling pathway; activated STAT3 drives the expression of osteopontin and its downstream stemness-related genes. This signaling axis significantly enhances the invasiveness and metastatic capacity of recipient cells, thereby promoting an increase in the overall malignancy of the tumor by enhancing their stemness.95
Progress in MSC-EVs Treatment Research for GC
It is noteworthy that research on MSC-EVs has primarily focused on their pro-tumorigenic effects, though some studies also indicate their potential for anti-cancer activity. MSC-EVs promote tumor progression through multiple oncogenic mechanisms during the evolution of GC. MSC-EVs activate the AKT signaling axis and initiate EMT, a process that reinforces stem-like properties while augmenting the proliferative, migratory, and invasive potential of GC cells.96 In another study, human bone marrow-derived MSC-EVs activated the Hedgehog pathway through upregulation of key molecules (Shh, Gli1, Ptch1, Smo), thereby promoting the proliferation and metastasis of gastric cancer cells.97 Bone marrow-derived MSC-EVs also have been demonstrated to possess the capacity to modulate the process of angiogenesis in GC. Specifically, these EVs have been found to activate the ERK1/2 and p38 MAPK pathways, which in turn results in the overexpression of VEGF and chemokine receptor.98 This series of events has been shown to promote tumor angiogenesis. Notably, regarding their role in drug resistance, MSC-EVs can activate the CaM-Ks/Raf/MEK/ERK pathway, upregulate multidrug resistance protein expression, and induce GC cells to develop resistance to 5-fluorouracil (5-FU).99 In addition, MSC-EVs can induce multidrug resistance in GC cells to cisplatin and vincristine by delivering miR-301b-3p to target and suppress the expression of thioredoxin-interacting protein (TXNIP).100 Despite these tumorigenic findings, conflicting evidence suggests that MSC-EVs exert an inhibitory effect in GC. Recent studies have revealed that small extracellular vesicles derived from human umbilical cord-derived mesenchymal stem cells can significantly suppress GC proliferation and metastasis by delivering miR-13896 to silence the ATG2A-mediated autophagy pathway, thereby inducing apoptosis.101
Challenges, Engineering Strategies, and Combination Therapy for MSC-EVs
Challenges in Standardization in Clinical Translation and Frameworks for Quality Control
MSC-EVs have strong capability of controlling the inflammatory microenvironment, and its transpose has great prospect for application during clinical translation in regard of curing digestive system tumors. A substantial body of research has been conducted on MSC-EVs to date, most of it remaining at the preclinical stage is still at the preclinical level. Although a great deal of progress has been made in the preclinical domain, MSC-EVs in clinical settings still encounter various challenges. First, the absence of standardized isolation methodologies constitutes a significant hindrance. Presently, there is an absence of consolidated standards for EVs isolation. There are substantial variations in yield among the various preparation methods, which complicates the fulfillment of clinical application requirements.102 The sources of MSCs used include bone marrow, adipose tissue, and umbilical cord. Given the variety of passages, culture conditions, and isolation and purification methods—such as ultracentrifugation, exclusion chromatography, and immunocapture—there will be substantial disparities in the final MSC-EV products, including size, amount, and content.103 The reproducibility of the research is low, and the clinical efficacy is unstable In addition, the stability of EVs is highly dependent on the conditions under which they are stored. EVs from different sources may require distinct preservation conditions, but the optimal conditions for long-term storage remain to be elucidated.104 During the clinical translation of MSC-EVs, establishing a rigorous and reproducible quality control (QC) system is a fundamental prerequisite for ensuring both safety and efficacy. In alignment with the latest MISEV2023 recommendations and the broader consensus in the field, it is essential that the QC framework covers three core dimensions: physical properties, biochemical composition, and biological potency.11 The physical characterization of EVs mainly relies on Nanoparticle Tracking Analysis (NTA) or Dynamic Light Scattering (DLS). NTA provides high-resolution particle size distribution curves and accurate concentration data, while DLS focuses more on analyzing the overall size distribution of the sample. Additionally, Transmission Electron Microscopy (TEM) is widely regarded as the gold standard for observing vesicle morphology and double-checking the accuracy of NTA measurements.105 Regarding biochemical composition, it is necessary to simultaneously evaluate multiple categories of markers to verify the presence and integrity of EVs, including surface proteins, cytosolic proteins, non-EV structures, intracellular organelle markers, and tissue-specific markers.106 Immunomodulatory activity is a key criterion in the evaluation of biological potency. Recent studies indicate that the levels of TGF-β1 within MSC-EVs can function as a molecular marker to predict their immunomodulatory activity.107 Such marker-based quantitative assays offer superior precision and reproducibility compared to traditional cell-based co-culture experiments.
Risks of the Double Effect and Engineering Strategies
Although MSC-EVs demonstrate reduced immunogenicity, caution is warranted regarding their dual roles in tumor therapy. The MSC-EVs exhibit both tumor-suppressing and tumor-promoting activity. To illustrate, under hypoxic conditions, pretreated MSCs exhibit a significant enrichment of miR-21-5p within their secreted EVs. This specific miRNA has been shown to facilitate tumor progression by downregulating the expression of PTEN, PDCD4, and RECK, thereby promoting cancer cell survival, invasion, and EMT.108 Therefore, In order to use MSC-EVs for the treatment of inflammation-related digestive system tumors, it is essential to ensure that they do not promote the growth of tumor cells. Overall, the cargo delivery efficiency of MSC-EVs may remains below the desired threshold. To overcome these challenges, a series of studies on engineering modification techniques for EVs have been conducted within the research field. The engineering of EVs involves two main modifications: surface engineering and cargo engineering. In the field of surface engineering, the predominant strategies for achieving surface modification can be classified into three main categories: genetic engineering, chemical conjugation, and physical targeting approaches. Prior to EVs secretion, the expression of fusion proteins capable of targeting specific cell types was induced in MSCs via genetic engineering. These proteins naturally appear on the membrane surface of EVs secreted by MSCs, enabling specific targeting of designated cells.109 The genetic engineering modification technology enhances therapeutic specificity and efficiency by enabling targeted peptides on the surface of EVs to bind specifically to target cells. In the area of chemical coupling strategies, common approaches involve the utilization of click chemistry, covalent bonding, or electrostatic/hydrophobic interactions for the direct attachment of targeting molecules—such as antibodies or antigens—to the surface of EVs.110 The advantage is that there is more freedom and it can be adjusted for different diseases. However, it should be acknowledged that a series of biochemical reactions are necessary, which may potentially compromise the functionality of proteins on the EVs’ membrane. For the part on physical targeting approaches, One potential strategy entails the co-incubation of superparamagnetic iron oxide nanoparticles with human mesenchymal stem cells, thereby facilitating the formation of nanoparticle-loaded magnetic EVs through the process of endocytosis. The employment of an external magnetic field serves to guide these particles, thus significantly enhancing their targeted delivery efficiency within a living organism.111 The engineering of MSC-EV cargo entails the efficient loading of specific therapeutic molecules into MSC-EVs through methods such as electroporation, ultrasound, incubation, or freeze-thaw cycles. This process encompasses the loading of small-molecule chemotherapeutic drugs, anti-inflammatory drugs, and nucleic acid molecules that are engineered to perform specific functions.112 MSC-EVs show significant potential for clinical translation in the treatment of digestive system tumors; however, achieving this goal requires overcoming several challenges, including standardization, heterogeneity, safety, and delivery efficiency.
Combination Therapy with EVs
Although immune checkpoint inhibitors (ICIs) have achieved remarkable progress in cancer therapy, their clinical response rates are often constrained by the heterogeneity of the TME. Evidence indicates that tumor cells not only express PD-L1 on their surface but also secrete EVs carrying PD-L1. These PD-L1-carrying EVs can reach distant sites to bind with PD-1 on CD8+ T cells, inducing CD8+ T cell apoptosis or functional exhaustion.113 However, these same transport mechanisms can be harnessed for therapeutic gain through bioengineering. In triple-negative breast cancer (TNBC) models, donor cells such as MDA-MB-231 can be genetically engineered to secrete EVs that highly express a high-affinity PD-1 variant (havPD-1). These havPD-1-EVs act by competitively binding to membrane-bound PD-L1 on tumor surfaces and neutralizing tumor-derived circulating PD-L1 EVs, effectively shutting down the PD-1/PD-L1 signaling pathway. This blockade not only restores T-cell activation and cytotoxicity but also directly induces apoptosis in tumor cells, thereby producing exceptional antitumor activity.114 Ultimately, while the exact mechanisms of MSC-EV-based combination therapies remain to be fully clarified, their unique transport machinery and immunomodulatory features are hard to overlook. By leveraging bioengineering techniques, MSC-EVs can be further optimized to act as a powerful synergistic platform, holding massive potential to boost the efficacy of ICIs in cancer treatment.
Discussion
This review is a large discussion on the basis of systematically discussing the mechanisms and therapeutic effects of MSC-EVs modulating the digestive system tumors inflammatory microenvironment. The review indicates that MSC-EVs play an important part in inflammatory regulation in the digestive system tumors. Delivering some miRNAs like miR-146a, miR-145-5p etc. to stop the major inflammatory signaling pathways such as NF-κB, MAPK. It reshapes the inflammatory microenvironment by inducing macrophage polarization toward the M2 type and promoting Treg cell production. This is also a brand new theoretical premise of a comprehensive prevention and treatment strategy, which can prevent and treat inflammation and cancer from the source, and intervene accurately in the development process of cancer. More precisely, the potential of MSC-EVs for therapy has a tissue-specific nature for different subtypes of digestive system tumor. In CRC, MSC-EVs reduce tumor cell malignancy via the PI3K/AKT pathway by delivering miR-22-3p. In HCC, they have an antitumor effect on it through the ERK pathway and EMT. However, this therapeutic potential must be carefully weighed against the pro-tumor effects of tumor-derived EVs. As noted, in CRC, hypoxia-induced tumor-derived EVs boost tumor proliferation by delivering miR-210-3p, or reshape the liver’s pre-metastatic niche via miR-221/222. Similarly, in HCC, EVs loaded with S100A4 protein from highly metastatic cells can flip the switch on the STAT3 pathway to enhance invasiveness. These contrasting effects, driven by different parental cells, further explain why MSC-EVs may carry tissue-specific risks or even act as a double-edged sword across different tumor subtypes, such as GC. In the specific context of particular tumors, potential risks must be meticulously analyzed, and targeted optimization of therapeutic outcomes must be actively pursued through engineered modifications. Even though MSC-EVs exhibit considerable therapeutic potential, there are a number of challenges that must be overcome before their clinical translation can be realised. The main problem is that MSC-EVs from various tissues and preparation methods. Heterogeneity comes from greatly different yields, ingredients and biological activities, which seriously reduces the reproducibility and sturdiness of the results. Therefore, establishing a standardized framework for isolation techniques, characterization criteria, and quality control is of paramount importance, necessitating strict adherence to the latest guidelines and field-wide consensus. It is important to note that under certain pathophysiological conditions, MSC-EVs may carry pro-cancer molecules. This makes it imperative to conduct comprehensive evaluation and optimization of their biosafety through rigorous source screening, optimized pretreatment strategies, or cargo editing techniques before clinical translation. In addition, It is also possible that the expected efficiency of MSC - EVs work as a natural carrier can be achieved after making engineered modification techniques. We can modify EVs using genetics to express targeting peptides on the surface of EVs, or modify the surface using click chemistry, so as to improve tumor targeting and therapy. At the same time, directed loading of therapeutic molecules via physical techniques such as electroporation also serves this purpose. Looking forward, there is going to be more research work done to make EVs that can accurately identify tumor tissues and release their curative payload only in specific microenvironments. Meanwhile, Study using MSC-EVs in combination with current treatment approaches like immune checkpoint inhibitors, chemotherapy, in order to overcome tumor resistance. By establishing rigorous preclinical evaluation models and validation systems, we are driving a substantial leap forward in advancing MSC-EVs from preclinical research to clinical application, ultimately providing safer and more effective innovative treatment options for patients with digestive system tumors.
Abbreviations
CRC, colorectal cancer; HCC, hepatocellular carcinoma; GC, gastric cancer; RNS, reactive nitrogen species; ROS, reactive oxygen species; IBD, inflammatory bowel disease; NF-κB, nuclear factor κB; STAT3, Signal Transducer and Activator of Transcription 3; H. pylori, Helicobacter pylori; MSCs, mesenchymal stem cells; EVs, extracellular vesicles; MVB, multivesicular body; ILVs, intraluminal vesicles; ESCRT, endosomal sorting complex required for transport; VEGF, vascular endothelial growth factor; Tregs, regulatory T cells; MAPK, mitogen-activated protein kinase; Wnt; Wingless-type MMTV integration site family, MDSCs, myeloid-derived suppressor cells; LPS, Lipopolysaccharide; TLRs, Toll-like receptors; CagA, Cytotoxin-associated gene A; PI3K/AKT, phosphoinositide 3-kinase/protein kinase B; ERK, extracellular signal-regulated kinase; LRP4, LDL receptor-related protein 4; EMT, epithelial-mesenchymal transition; ITGA2, integrin alpha 2; QC, quality control; ICIs, Immune checkpoint inhibitors.
Acknowledgments
We thank everyone who contributed to this manuscript.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This work was supported by Jiangxi Provincial Natural Science Foundation (20252BAC200537), Ganzhou Key Research and Development Program Project of the Ganzhou Science and Technology Bureau(GZ2024YLJ144).
Disclosure
The authors declare no conflicts of interest in this work.
References
1. Sung H, Ferlay J, Siegel RL, et al. Global cancer statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA a Cancer J Clinicians. 2021;71(3):209–18. doi:10.3322/caac.21660
2. Wen Y, Zhu Y, Zhang C, et al. Chronic inflammation, cancer development and immunotherapy. Front Pharmacol. 2022;13:1040163. doi:10.3389/fphar.2022.1040163
3. Burgos-Molina AM, Téllez Santana T, Redondo M, Bravo Romero MJ. The crucial role of inflammation and the immune system in colorectal cancer carcinogenesis: a comprehensive perspective. Int J Mol Sci. 2024;25(11):6188. doi:10.3390/ijms25116188
4. Yu LX, Ling Y, Wang HY. Role of nonresolving inflammation in hepatocellular carcinoma development and progression. Npj Precision Onc. 2018;2(1):6. doi:10.1038/s41698-018-0048-z
5. Myrou A. Molecular mechanisms and treatment strategies for Helicobacter pylori-induced gastric carcinogenesis and Mucosa-Associated Lymphoid Tissue (MALT) lymphoma. Cureus. 2024;16(5):e60326. doi:10.7759/cureus.60326
6. Zhan T, Betge J, Schulte N, et al. Digestive cancers: mechanisms, therapeutics and management. Signal Trans Target Ther. 2025;10:24. doi:10.1038/s41392-024-02097-4
7. Zhang YH, Chen XL, Wang YR, Hou YW, Zhang YD, Wang KJ. Prevention of malignant digestive system tumors should focus on the control of chronic inflammation. World J Gastrointest Oncol. 2023;15(3):389–404. doi:10.4251/wjgo.v15.i3.389
8. Wang C, Xu M, Fan Q, Li C, Zhou X. Therapeutic potential of exosome‐based personalized delivery platform in chronic inflammatory diseases. Asian J Pharm Sci. 2023;18(1):100772. doi:10.1016/j.ajps.2022.100772
9. Pirisinu M, Pham TC, Zhang DX, Hong TN, Nguyen LT, Le MT. Extracellular vesicles as natural therapeutic agents and innate drug delivery systems for cancer treatment: recent advances, current obstacles, and challenges for clinical translation. Semin Cancer Biol. 2022;80:340–355. doi:10.1016/j.semcancer.2020.08.007
10. Raposo G, Stoorvogel W. Extracellular vesicles: exosomes, microvesicles, and friends. J Cell Biol. 2013;200(4):373–383. doi:10.1083/jcb.201211138
11. Welsh JA, Goberdhan DC, O’Driscoll L, Théry C, Witwer KW. MISEV2023: an updated guide to EV research and applications. J Extracell Vesicles. 2024;13(2):e12416. doi:10.1002/jev2.12416
12. van Niel G, D’Angelo G, Raposo G. Shedding light on the cell biology of extracellular vesicles. Nat Rev Mol Cell Biol. 2018;19(4):213–228. doi:10.1038/nrm.2017.125
13. Gurung S, Perocheau D, Touramanidou L, Baruteau J. The exosome journey: from biogenesis to uptake and intracellular signalling. Cell Commun Signal. 2021;19:47. doi:10.1186/s12964-021-00730-1
14. Mizenko RR, Brostoff T, Rojalin T, et al. Tetraspanins are unevenly distributed across single extracellular vesicles and bias sensitivity to multiplexed cancer biomarkers. J Nanobiotechnol. 2021;19(1):250. doi:10.1186/s12951-021-00987-1
15. Boere J, Malda J, Van De Lest CHA, Van Weeren PR, Wauben MHM. Extracellular vesicles in joint disease and therapy. Front Immunol. 2018;9:2575. doi:10.3389/fimmu.2018.02575
16. Sun SJ, Wei R, Li F, Liao SY, Tse HF. Mesenchymal stromal cell-derived exosomes in cardiac regeneration and repair. Stem Cell Reports. 2021;16(7):1662–1673. doi:10.1016/j.stemcr.2021.05.003
17. Mulcahy LA, Pink RC, Carter DRF. Routes and mechanisms of extracellular vesicle uptake. J Extracell Vesicles. 2014;3. doi:10.3402/jev.v3.24641
18. Kim HI, Park J, Zhu Y, Wang X, Han Y, Zhang D. Recent advances in extracellular vesicles for therapeutic cargo delivery. Exp Mol Med. 2024;56(4):836–849. doi:10.1038/s12276-024-01201-6
19. Medzhitov R. The spectrum of inflammatory responses. Science. 2021;374(6571):1070–1075. doi:10.1126/science.abi5200
20. Furman D, Campisi J, Verdin E, et al. Chronic inflammation in the etiology of disease across the life span. Nat Med. 2019;25(12):1822–1832. doi:10.1038/s41591-019-0675-0
21. Zhao H, Wu L, Yan G, et al. Inflammation and tumor progression: signaling pathways and targeted intervention. Sig Transduct Target Ther. 2021;6(1):263. doi:10.1038/s41392-021-00658-5
22. Fan Y, Mao R, Yang J. NF-κB and STAT3 signaling pathways collaboratively link inflammation to cancer. Protein Cell. 2013;4(3):176–185. doi:10.1007/s13238-013-2084-3
23. Cao Y, Yi Y, Han C, Shi B. NF-κB signaling pathway in tumor microenvironment. Front Immunol. 2024;15:1476030. doi:10.3389/fimmu.2024.1476030
24. Oh A, Pardo M, Rodriguez A, et al. NF-κB signaling in neoplastic transition from epithelial to mesenchymal phenotype. Cell Commun Signal. 2023;21(1):291. doi:10.1186/s12964-023-01207-z
25. Dibra D, Mishra L, Li S. Molecular mechanisms of oncogene-induced inflammation and inflammation-sustained oncogene activation in gastrointestinal tumors: an underappreciated symbiotic relationship. Biochimica et Biophysica Acta. 2014;1846(1):152–160. doi:10.1016/j.bbcan.2014.05.001
26. Sadrekarimi H, Gardanova ZR, Bakhshesh M, et al. Emerging role of human microbiome in cancer development and response to therapy: special focus on intestinal microflora. J Transl Med. 2022;20(1):301. doi:10.1186/s12967-022-03492-7
27. Chiba T, Marusawa H, Ushijima T. Inflammation-associated cancer development in digestive organs: mechanisms and roles for genetic and epigenetic modulation. Gastroenterology. 2012;143(3):550–563. doi:10.1053/j.gastro.2012.07.009
28. Mehta M, Ahmed S, Dryden G. Immunopathophysiology of inflammatory bowel disease: how genetics link barrier dysfunction and innate immunity to inflammation. Innate Immun. 2017;23(6):497–505. doi:10.1177/1753425917722206
29. Lee SH, Kwon JE, Cho ML. Immunological pathogenesis of inflammatory bowel disease. Intest Res. 2018;16(1):26. doi:10.5217/ir.2018.16.1.26
30. Soufli I, Toumi R, Rafa H, Touil-Boukoffa C. Overview of cytokines and nitric oxide involvement in immuno-pathogenesis of inflammatory bowel diseases. World J Gastrointest Pharmacol Ther. 2016;7(3):353–360. doi:10.4292/wjgpt.v7.i3.353
31. Fujino S, Andoh A, Bamba S, et al. Increased expression of interleukin 17 in inflammatory bowel disease. Gut. 2003;52(1):65–70. doi:10.1136/gut.52.1.65
32. Shen W, Durum SK. Synergy of IL-23 and Th17 cytokines: new light on inflammatory bowel disease. Neurochem Res. 2010;35(6):940–946. doi:10.1007/s11064-009-0091-9
33. Landskron G, De la Fuente M, Thuwajit P, Thuwajit C, Hermoso MA. Chronic inflammation and cytokines in the tumor microenvironment. J Immunol Res. 2014;2014:149185. doi:10.1155/2014/149185
34. Galuppini F, Fassan M, Mastracci L, et al. The histomorphological and molecular landscape of colorectal adenomas and serrated lesions. Pathologica. 2021;113(3):218–229. doi:10.32074/1591-951X-270
35. Frigerio S, Lartey DA, D’Haens GR, Grootjans J. The role of the immune system in IBD-associated colorectal cancer: from pro to anti-tumorigenic mechanisms. Int J Mol Sci. 2021;22(23):12739. doi:10.3390/ijms222312739
36. Stidham R, Higgins P. Colorectal cancer in inflammatory bowel disease. Clin Colon Rectal Surg. 2018;31(03):168–178. doi:10.1055/s-0037-1602237
37. Majumder S, Shivaji UN, Kasturi R, Sigamani A, Ghosh S, Iacucci M. Inflammatory bowel disease-related colorectal cancer: past, present and future perspectives. World J Gastrointest Oncol. 2022;14(3):547–567. doi:10.4251/wjgo.v14.i3.547
38. Westbrook AM, Szakmary A, Schiestl RH. Mechanisms of intestinal inflammation and development of associated cancers: lessons learned from mouse models. Mutat Res. 2010;705(1):40–59. doi:10.1016/j.mrrev.2010.03.001
39. Scarpa M, Castagliuolo I, Castoro C, et al. Inflammatory colonic carcinogenesis: a review on pathogenesis and immunosurveillance mechanisms in ulcerative colitis. World J Gastroenterol. 2014;20(22):6774–6785. doi:10.3748/wjg.v20.i22.6774
40. Nguyen LH, Goel A, Chung DC. Pathways of Colorectal Carcinogenesis. Gastroenterology. 2020;158(2):291–302. doi:10.1053/j.gastro.2019.08.059
41. Ghobashi AH, Lanzloth R, Ladaika CA, Masood A, O’Hagan HM. Single-cell profiling reveals the impact of genetic alterations on the differentiation of inflammation-induced murine colon tumors. Cancers. 2024;16(11):2040. doi:10.3390/cancers16112040
42. Galasso L, Cerrito L, Maccauro V, et al. Inflammatory response in the pathogenesis and treatment of hepatocellular carcinoma: a double-edged weapon. Int J Mol Sci. 2024;25(13):7191. doi:10.3390/ijms25137191
43. Cardin R, Piciocchi M, Bortolami M, et al. Oxidative damage in the progression of chronic liver disease to hepatocellular carcinoma: an intricate pathway. World J Gastroenterol. 2014;20(12):3078–3086. doi:10.3748/wjg.v20.i12.3078
44. Keenan BP, Fong L, Kelley RK. Immunotherapy in hepatocellular carcinoma: the complex interface between inflammation, fibrosis, and the immune response. J Immunother Cancer. 2019;7(1):267. doi:10.1186/s40425-019-0749-z
45. Xu Y, Fang F, Jiao H, et al. Activated hepatic stellate cells regulate MDSC migration through the SDF-1/CXCR4 axis in an orthotopic mouse model of hepatocellular carcinoma. Cancer Immunol Immunother. 2019;68(12):1959–1969. doi:10.1007/s00262-019-02414-9
46. Stewart CA, Metheny H, Iida N, et al. Interferon-dependent IL-10 production by Tregs limits tumor Th17 inflammation. J Clin Invest. 2013;123(11):4859–4874. doi:10.1172/JCI65180
47. Chen Y, Hu H, Yuan X, Fan X, Zhang C. Advances in immune checkpoint inhibitors for advanced hepatocellular carcinoma. Front Immunol. 2022;13:896752. doi:10.3389/fimmu.2022.896752
48. Bergmann J, Müller M, Baumann N, et al. IL-6 trans-signaling is essential for the development of hepatocellular carcinoma in mice. Hepatology. 2017;65(1):89–103. doi:10.1002/hep.28874
49. Guo Q, Jin Y, Chen X, et al. NF-κB in biology and targeted therapy: new insights and translational implications. Signal Transduct Target Ther. 2024;9(1):53. doi:10.1038/s41392-024-01757-9
50. Nakagawa H, Maeda S. Inflammation- and stress-related signaling pathways in hepatocarcinogenesis. World J Gastroenterol. 2012;18(31):4071–4081. doi:10.3748/wjg.v18.i31.4071
51. Peng Y, Yu J, Liu F, et al. Accumulation of TOX high mobility group box family member 3 promotes the oncogenesis and development of hepatocellular carcinoma through the MAPK signaling pathway. MedComm. 2024;5(3):e510. doi:10.1002/mco2.510
52. Fishbein A, Hammock BD, Serhan CN, Panigrahy D. Carcinogenesis: failure of resolution of inflammation? Pharmacol Ther. 2021;218:107670. doi:10.1016/j.pharmthera.2020.107670
53. Correa P, Piazuelo MB. The gastric precancerous cascade. J Dig Dis. 2012;13(1):2–9. doi:10.1111/j.1751-2980.2011.00550.x
54. McNamara KM, Sierra JC, Latour YL, et al. Spermine oxidase promotes Helicobacter pylori-mediated gastric carcinogenesis through acrolein production. Oncogene. 2025;44(5):296–306. doi:10.1038/s41388-024-03218-7
55. Zhang S, Shen Y, Liu H, et al. Inflammatory microenvironment in gastric premalignant lesions: implication and application. Front Immunol. 2023;14:1297101. doi:10.3389/fimmu.2023.1297101
56. D’Souza C, Pediongco T, Wang H, et al. Mucosal-associated invariant T cells augment immunopathology and gastritis in chronic Helicobacter pylori infection. J Immunol. 2018;200(5):1901–1916. doi:10.4049/jimmunol.1701512
57. Resende C, Regalo G, Durães C, et al. Interleukin-1B signalling leads to increased survival of gastric carcinoma cells through a CREB-C/EBPβ-associated mechanism. Gastric Cancer. 2016;19(1):74–84. doi:10.1007/s10120-014-0448-x
58. Ito N, Tsujimoto H, Ueno H, Xie Q, Shinomiya N. Helicobacter pylori-mediated immunity and signaling transduction in gastric cancer. J Clin Med. 2020;9(11):3699. doi:10.3390/jcm9113699
59. Sahan AZ, Hazra TK, Das S. The pivotal role of DNA repair in infection mediated-inflammation and cancer. Front Microbiol. 2018;9:663. doi:10.3389/fmicb.2018.00663
60. Li T, Chen ZJ. The cGAS–cGAMP–STING pathway connects DNA damage to inflammation, senescence, and cancer. J Exp Med. 2018;215(5):1287–1299. doi:10.1084/jem.20180139
61. Asgarpour K, Shojaei Z, Amiri F, et al. Exosomal microRNAs derived from mesenchymal stem cells: cell-to-cell messages. Cell Commun Signal. 2020;18:149. doi:10.1186/s12964-020-00650-6
62. Sica A, Mantovani A. Macrophage plasticity and polarization: in vivo veritas. J Clin Invest. 2012;122(3):787–795. doi:10.1172/JCI59643
63. Liu H, Liang Z, Wang F, et al. Exosomes from mesenchymal stromal cells reduce murine colonic inflammation via a macrophage-dependent mechanism. JCI Insight. 2019;4(24):e131273. doi:10.1172/jci.insight.131273
64. Zhao J, Li X, Hu J, et al. Mesenchymal stromal cell-derived exosomes attenuate myocardial ischaemia-reperfusion injury through miR-182-regulated macrophage polarization. Cardiovasc Res. 2019;115(7):1205–1216. doi:10.1093/cvr/cvz040
65. Tian S, Zhou X, Zhang M, et al. Mesenchymal stem cell-derived exosomes protect against liver fibrosis via delivering miR-148a to target KLF6/STAT3 pathway in macrophages. Stem Cell Res Ther. 2022;13:330. doi:10.1186/s13287-022-03010-y
66. Cosenza S, Toupet K, Maumus M, et al. Mesenchymal stem cells-derived exosomes are more immunosuppressive than microparticles in inflammatory arthritis. Theranostics. 2018;8(5):1399–1410. doi:10.7150/thno.21072
67. Xu FT, Ling Y, Wei HX, et al. BMSC exosomes deliver JKAP to restore Th17/Treg balance via AKT/ERK, alleviating rheumatoid arthritis. iScience. 2025;28(7):112832. doi:10.1016/j.isci.2025.112832
68. Shahir M, Mahmoud Hashemi S, Asadirad A, et al. Effect of mesenchymal stem cell‐derived exosomes on the induction of mouse tolerogenic dendritic cells. J Cell Physiol. 2020;235(10):7043–7055. doi:10.1002/jcp.29601
69. Harrell CR, Jovicic N, Djonov V, Arsenijevic N, Volarevic V. Mesenchymal stem cell-derived exosomes and other extracellular vesicles as new remedies in the therapy of inflammatory diseases. Cells. 2019;8(12):1605. doi:10.3390/cells8121605
70. Jiang Z, Zhang J. Mesenchymal stem cell-derived exosomes containing miR-145-5p reduce inflammation in spinal cord injury by regulating the TLR4/NF-κB signaling pathway. Cell Cycle. 2021;20(10):993–1009. doi:10.1080/15384101.2021.1919825
71. Cui S, Zhang L. microRNA-129-5p shuttled by mesenchymal stem cell-derived extracellular vesicles alleviates intervertebral disc degeneration via blockade of LRG1-mediated p38 MAPK activation. J Tissue Eng. 2021;12:20417314211021679. doi:10.1177/20417314211021679
72. Mizuta Y, Akahoshi T, Guo J, et al. Exosomes from adipose tissue-derived mesenchymal stem cells ameliorate histone-induced acute lung injury by activating the PI3K/Akt pathway in endothelial cells. Stem Cell Res Ther. 2020;11:508. doi:10.1186/s13287-020-02015-9
73. Li D, Li D, Wang Z, et al. Signaling pathways activated and regulated by stem cell-derived exosome therapy. Cell Biosci. 2024;14:105. doi:10.1186/s13578-024-01277-7
74. Konala VBR, Mamidi MK, Bhonde R, Das AK, Pochampally R, Pal R. The current landscape of the mesenchymal stromal cell secretome: a new paradigm for cell-free regeneration. Cytotherapy. 2016;18(1):13–24. doi:10.1016/j.jcyt.2015.10.008
75. Melzer C, Rehn V, Yang Y, Bähre H, Von Der Ohe J, Hass R. Taxol-loaded MSC-derived exosomes provide a therapeutic vehicle to target metastatic breast cancer and other carcinoma cells. Cancers. 2019;11(6):798. doi:10.3390/cancers11060798
76. Haney MJ, Klyachko NL, Zhao Y, et al. Exosomes as drug delivery vehicles for parkinson’s disease therapy. J Control Release. 2015;207:18–30. doi:10.1016/j.jconrel.2015.03.033
77. Vizoso F, Eiro N, Cid S, Schneider J, Perez-Fernandez R. Mesenchymal stem cell secretome: toward cell-free therapeutic strategies in regenerative medicine. IJMS. 2017;18(9):1852. doi:10.3390/ijms18091852
78. Lin W, Huang L, Li Y, et al. Mesenchymal stem cells and cancer: clinical challenges and opportunities. Biomed Res Int. 2019;2019:1–12. doi:10.1155/2019/2820853
79. Zeng Q, Li S, Chepeha DB, et al. Crosstalk between tumor and endothelial cells promotes tumor angiogenesis by MAPK activation of notch signaling. Cancer Cell. 2005;8(1):13–23. doi:10.1016/j.ccr.2005.06.004
80. Lee JK, Park SR, Jung BK, et al. Exosomes derived from mesenchymal stem cells suppress angiogenesis by down-regulating VEGF expression in breast cancer cells. PLoS One. 2013;8(12):e84256. doi:10.1371/journal.pone.0084256
81. Mittal V. Epithelial mesenchymal transition in tumor metastasis. Annu Rev Pathol. 2018;13:395–412. doi:10.1146/annurev-pathol-020117-043854
82. Wan FZ, Chen KH, Sun YC, et al. Exosomes overexpressing miR-34c inhibit malignant behavior and reverse the radioresistance of nasopharyngeal carcinoma. J Transl Med. 2020;18:12. doi:10.1186/s12967-019-02203-z
83. Luo T, Von Der Ohe J, Hass R. MSC-derived extracellular vesicles in tumors and therapy. Cancers. 2021;13(20):5212. doi:10.3390/cancers13205212
84. Weng Z, Zhang B, Wu C, et al. Therapeutic roles of mesenchymal stem cell-derived extracellular vesicles in cancer. J Hematol Oncol. 2021;14:136. doi:10.1186/s13045-021-01141-y
85. Wang Y, Lin C. Exosomes miR-22-3p derived from mesenchymal stem cells suppress colorectal cancer cell proliferation and invasion by regulating RAP2B and PI3K/AKT pathway. J Oncol. 2021;2021:3874478. doi:10.1155/2021/3874478
86. Xu Y, Shen L, Li F, Yang J, Wan X, Ouyang M. microRNA-16-5p-containing exosomes derived from bone marrow-derived mesenchymal stem cells inhibit proliferation, migration, and invasion, while promoting apoptosis of colorectal cancer cells by downregulating ITGA2. J Cell Physiol. 2019;234(11):21380–21394. doi:10.1002/jcp.28747
87. Li T, Wan Y, Su Z, Li J, Han M, Zhou C. Mesenchymal stem cell-derived exosomal microRNA-3940-5p inhibits colorectal cancer metastasis by targeting integrin α6. Dig Dis Sci. 2021;66(6):1916–1927. doi:10.1007/s10620-020-06458-1
88. Wang X, Li D, Li G, et al. Enhanced therapeutic potential of hybrid exosomes loaded with paclitaxel for cancer therapy. Int J Mol Sci. 2024;25(7):3645. doi:10.3390/ijms25073645
89. Ge L, Zhou F, Nie J, Wang X, Zhao Q. Hypoxic colorectal cancer-secreted exosomes deliver miR-210-3p to normoxic tumor cells to elicit a protumoral effect. Exp Biol Med. 2021;246(17):1895–1906. doi:10.1177/15353702211011576
90. Tian F, Wang P, Lin D, et al. Exosome‐delivered miR‐221/222 exacerbates tumor liver metastasis by targeting SPINT1 in colorectal cancer. Cancer Sci. 2021;112(9):3744–3755. doi:10.1111/cas.15028
91. Yin Z, Jiang K, Li R, Dong C, Wang L. Multipotent mesenchymal stromal cells play critical roles in hepatocellular carcinoma initiation, progression and therapy. Mol Cancer. 2018;17:178. doi:10.1186/s12943-018-0926-6
92. Gu H, Yan C, Wan H, et al. Mesenchymal stem cell-derived exosomes block malignant behaviors of hepatocellular carcinoma stem cells through a lncRNA C5orf66-AS1/microRNA-127-3p/DUSP1/ERK axis. Human Cell. 2021;34(6):1812–1829. doi:10.1007/s13577-021-00599-9
93. Xu Y, Lai Y, Cao L, et al. Human umbilical cord mesenchymal stem cells-derived exosomal microRNA-451a represses epithelial–mesenchymal transition of hepatocellular carcinoma cells by inhibiting ADAM10. RNA Biol. 2021;18(10):1408–1423. doi:10.1080/15476286.2020.1851540
94. Sun Y, Liu G, Zhang K, Cao Q, Liu T, Li J. Mesenchymal stem cells-derived exosomes for drug delivery. Stem Cell Res Ther. 2021;12:561. doi:10.1186/s13287-021-02629-7
95. Sun H, Wang C, Hu B, et al. Exosomal S100A4 derived from highly metastatic hepatocellular carcinoma cells promotes metastasis by activating STAT3. Signal Transduct Target Ther. 2021;6:187. doi:10.1038/s41392-021-00579-3
96. Gu H, Ji R, Zhang X, et al. Exosomes derived from human mesenchymal stem cells promote gastric cancer cell growth and migration via the activation of the Akt pathway. Mol Med Reports. 2016;14(4):3452–3458. doi:10.3892/mmr.2016.5625
97. Qi J, Zhou Y, Jiao Z, et al. Exosomes derived from human bone marrow mesenchymal stem cells promote tumor growth through hedgehog signaling pathway. Cell Physiol Biochem. 2017;42(6):2242–2254. doi:10.1159/000479998
98. Akad F, Mocanu V, Peiu SN, et al. Mesenchymal stem cell-derived exosomes modulate angiogenesis in gastric cancer. Biomedicines. 2023;11(4):1031. doi:10.3390/biomedicines11041031
99. Yang Y, Meng WJ, Wang ZQ. Cancer stem cells and the tumor microenvironment in gastric cancer. Front Oncol. 2022;11:803974. doi:10.3389/fonc.2021.803974
100. Z T, H Z, W Z, et al. microRNA-301b-3p from mesenchymal stem cells-derived extracellular vesicles inhibits TXNIP to promote multidrug resistance of gastric cancer cells. Cell Biol Toxicol. 2023;39(5). doi:10.1007/s10565-021-09675-0
101. Wu P, Wang M, Jin C, et al. Highly efficient delivery of novel MiR-13896 by human umbilical cord mesenchymal stem cell-derived small extracellular vesicles inhibits gastric cancer progression by targeting ATG2A-mediated autophagy. Biomater Res. 2024;28:0119. doi:10.34133/bmr.0119
102. Ludwig N, Whiteside TL, Reichert TE. Challenges in exosome isolation and analysis in health and disease. Int J Mol Sci. 2019;20(19):4684. doi:10.3390/ijms20194684
103. Yuan YG, Wang JL, Zhang YX, Li L, Reza AMMT, Biogenesis GS. Composition and potential therapeutic applications of mesenchymal stem cells derived exosomes in various diseases. IJN. 2023;18:3177–3210. doi:10.2147/IJN.S407029
104. Chang C, Cai RP, Su YM, Wu Q, Su Q. Mesenchymal stem cell-derived exosomal noncoding RNAs as alternative treatments for myocardial ischemia-reperfusion injury: current status and future perspectives. J Cardiovasc Trans Res. 2023;16(5):1085–1098. doi:10.1007/s12265-023-10401-w
105. Kowkabany G, Bao Y. Nanoparticle tracking analysis: an effective tool to characterize extracellular vesicles. Molecules. 2024;29(19):4672. doi:10.3390/molecules29194672
106. Liang Y, Lehrich BM, Zheng S, Lu M. Emerging methods in biomarker identification for extracellular vesicle-based liquid biopsy. J Extracell Vesicles. 2021;10(7):e12090. doi:10.1002/jev2.12090
107. Kaur G, Bae EH, Zhang Y, et al. Biopotency and surrogate assays to validate the immunomodulatory potency of extracellular vesicles derived from mesenchymal stem/stromal cells for the treatment of experimental autoimmune uveitis. J Extracell Vesicles. 2024;13(8):e12497. doi:10.1002/jev2.12497
108. Ren W, Hou J, Yang C, et al. Extracellular vesicles secreted by hypoxia pre-challenged mesenchymal stem cells promote non-small cell lung cancer cell growth and mobility as well as macrophage M2 polarization via miR-21-5p delivery. J Exp Clin Cancer Res. 2019;38(1):62. doi:10.1186/s13046-019-1027-0
109. Yan J, Xiao H, Zhou X, et al. Engineered exosomes reprogram Gli1+ cells in vivo to prevent calcification of vascular grafts and autologous pathological vessels. Sci Adv. 2023;9(29):eadf7858. doi:10.1126/sciadv.adf7858
110. Jung I, Shin S, Baek MC, Yea K. Modification of immune cell-derived exosomes for enhanced cancer immunotherapy: current advances and therapeutic applications. Exp Mol Med. 2024;56(1):19–31. doi:10.1038/s12276-023-01132-8
111. Li X, Wang Y, Shi L, et al. Magnetic targeting enhances the cutaneous wound healing effects of human mesenchymal stem cell-derived iron oxide exosomes. J Nanobiotechnol. 2020;18:113. doi:10.1186/s12951-020-00670-x
112. Tian J, Han Z, Song D, et al. Engineered exosome for drug delivery: recent development and clinical applications. Int J Nanomed. 2023;18:7923–7940. doi:10.2147/IJN.S444582
113. Fang R, Huang B, Li Y, et al. PD‐L1 on tumor‐derived extracellular vesicles induces CD8+ T cell terminal exhaustion and mediates Anti‐PD‐1 resistance in head and neck squamous cell carcinoma. Adv Sci. 2025;13(4):e16348. doi:10.1002/advs.202516348
114. Chen Y, Wang L, Zheng M, et al. Engineered extracellular vesicles for concurrent Anti-PDL1 immunotherapy and chemotherapy. Bioact Mater. 2021;9:251–265. doi:10.1016/j.bioactmat.2021.07.012
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Mesenchymal Stem Cell–Derived Exosomes in Various Chronic Liver Diseases: Hype or Hope?
Zhu L, Wang Q, Guo M, Fang H, Li T, Zhu Y, Jiang H, Xiao P, Hu M
Journal of Inflammation Research 2024, 17:171-189
Published Date: 10 January 2024
Emerging Role and Mechanism of Mesenchymal Stem Cells-Derived Extracellular Vesicles in Rheumatic Disease
Wang Z, Yang C, Yan S, Sun J, Zhang J, Qu Z, Sun W, Zang J, Xu D
Journal of Inflammation Research 2024, 17:6827-6846
Published Date: 30 September 2024
Oral Opportunistic Infection Induced by Stress and Silent Type 2 Diabetes Mellitus in Young Adult Patient: A Case Report
Karina D, Heldayani I, Hidayat W
International Medical Case Reports Journal 2025, 18:59-66
Published Date: 11 January 2025
Extracellular Vesicles in Acute Kidney Injury: Mechanisms, Biomarkers, and Therapeutic Potential
Yan Q, Liu M, Mao J, Zhao Z, Wang B
International Journal of Nanomedicine 2025, 20:6271-6288
Published Date: 17 May 2025
MSC-Derived Extracellular Vesicles: Roles and Molecular Mechanisms for Tissue Repair
Wu T, Liu Y, Wang S, Shi C
International Journal of Nanomedicine 2025, 20:7953-7974
Published Date: 21 June 2025