")
Back to Journals » Journal of Inflammation Research » Volume 16
The Role of Inflammation-Associated Factors in Head and Neck Squamous Cell Carcinoma
Authors Li K, Zeng X, Liu P, Zeng X, Lv J, Qiu S, Zhang P
Received 4 July 2023
Accepted for publication 16 September 2023
Published 27 September 2023 Volume 2023:16 Pages 4301—4315
DOI https://doi.org/10.2147/JIR.S428358
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Adam D Bachstetter
Kang Li,1,2,* Xianhai Zeng,1,2,* Peng Liu,1,2 Xiaoxia Zeng,2 Jie Lv,3 Shuqi Qiu,1,2 Peng Zhang1,2
1Department of Graduate and Scientific Research, Zunyi Medical University Zhuhai Campus, Zhuhai, Guangdong, People’s Republic of China; 2Department of Otorhinolaryngology, Longgang Otorhinolaryngology Hospital & Shenzhen Key Laboratory of Otorhinolaryngology, Shenzhen Institute of Otorhinolaryngology, Shenzhen, Guangdong, People’s Republic of China; 3School of Computer Science and Engineering, Yulin Normal University, Yulin, Guangxi, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Shuqi Qiu; Peng Zhang, Longgang Otorhinolaryngology Hospital & Shenzhen Key Laboratory of Otorhinolaryngology, Shenzhen Institute of Otorhinolaryngology, No. 3004 Longgang Avenue, Shenzhen, Guangdong, People’s Republic of China, Tel +86-755-28989999, Fax +86-755-28980505, Email [email protected]; [email protected]
Abstract: Head and neck squamous cell carcinoma (HNSCC), which originates in the head or neck tissues, is characterized by high rates of recurrence and metastasis. Inflammation is important in HNSCC prognosis. Inflammatory cells and their secreted factors contribute to the various stages of HNSCC development through multiple mechanisms. In this review, the mechanisms through which inflammatory factors, signaling pathways, and cells contribute to the initiation and progression of HNSCC have been discussed in detail. Furthermore, the diagnostic and therapeutic potential of targeting inflammation in HNSCC has been discussed to gain new insights into improving patient prognosis.
Keywords: inflammation, head and neck squamous cell carcinoma, HNSCC, mediators, progression, therapy
Introduction
Head and neck tumors originate from the epithelial tissues of the paranasal sinuses, nasal cavity, oral cavity, pharynx, and larynx. Head and neck squamous cell carcinoma (HNSCC) is the predominant malignancy affecting these regions, and is the sixth most common cancer worldwide, with an annual diagnosis of over 500,000 new cases.1 The established risk factors of HNSCC include tobacco and alcohol consumption; betel nut chewing is also an established risk factor of oral cancer.2,3 HNSCC is characterized by aggressive tumor progression, high recurrence and metastasis rates, and poor prognosis.4 The primary treatment modalities for HNSCC are radiation therapy and chemotherapy. Despite advances in these therapies, the 5-year survival rate of Patients with HNSCC remains <50%, except for early-stage tumors.5 The frequent diagnosis of HNSCC in the advanced stage poses significant treatment challenges. Therefore, the early diagnosis and treatment of HNSCC are critical for improving patient prognosis.
In recent years, immunotherapy has emerged as a promising treatment approach for HNSCC, with beneficial outcomes in selected patients. This warrants an in-depth study of the tumor microenvironment (TME) and mechanisms of immune evasion in HNSCC. TME comprises inflammatory cells and their secreted factors that may promote tumor progression. Prolonged inflammatory stimulation and immune activation lead to chronic inflammation, which results in repetitive tissue damage and regeneration.6 In fact, inflammation is a hallmark of cancer and an initiator of malignant transformation. More and more studies have shown that inflammatory cells, mediators, and pathways contribute to the development and immune evasion of various cancer types, including HNSCC. Therefore, exploring the molecular mechanisms through which inflammation affects HNSCC progression and metastasis is crucial to develop novel therapeutic strategies. In this review, the mechanisms underlying tumor-associated inflammation, clinical significance of inflammation-related biomarkers, and potential therapeutic approaches for HNSCC have been discussed.7
Role of Inflammation-Related Mediators in the Progression of HNSCC
Chronic inflammation triggers sustained tissue damage and significantly contributes to the onset and progression of head and neck tumors. Several inflammation-related mediators have been implicated in HNSCC, which are discussed in detail in the following sections (Figure 1).
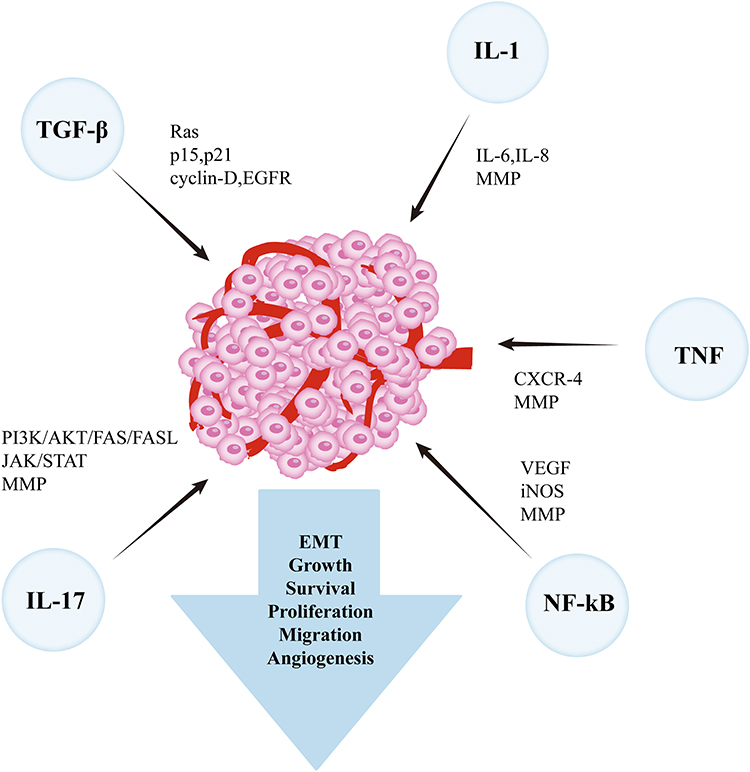 |
Figure 1 Inflammation-related mediators involved in promoting tumor progression. |
Interleukin (IL)-1
Interleukin (IL)-1 is a proinflammatory cytokine that instigates cellular and organ inflammation. The IL-1 family comprises 11 members, of which IL-1α, IL-1β, and the IL-1 receptor antagonist (IL-1RA) are most widely studied. IL-1 receptor family members include both the activators and inhibitors of inflammation, thereby exerting diverse effects on inflammatory processes.8 IL-1 drives tumor progression, invasion, and metastasis, primarily through chronic inflammation and immune-suppression.
IL-1α is known to upregulate IL-8 and IL-6 via the autocrine activation of NF-κB and AP-1.Tumor cells produce high levels of IL-1α, which induce the proliferation of cancer-associated fibroblasts (CAFs) that produce C-C motif ligand (CCL) 7, C-X-C motif ligand (CXCL) 1, IL-8, and CCL2 to accelerate tumor progression.9 The N-terminal domain of IL-1α helps transform cells in the bone marrow or vascular perivascular regions into a malignant phenotype.10 In addition, IL-1α enhances the migration of tumor cells within the endothelium, and the high expression of IL-1α in tumors is related to a higher risk of distant metastasis and worse overall survival.11 Thus, the up-regulation of IL-1α is a prognostic biomarker of cancer.12 IL-1β is a critical mediator of chronic inflammation and is related to various cancers. Upon stimulation by IL-1β, dysfunctional oral keratinocytes (DOKs) and oral squamous cell carcinoma (OSCC) cells secrete high levels of IL-6 and IL-8. IL-1β also facilitates tumor progression by promoting angiogenesis and epithelial-mesenchymal transition (EMT).13 By contrast, IL-1β derived from tumor-infiltrating inflammatory cells can recruit neutrophils and inhibit tumor growth.14 The upregulation of IL-1β in tongue squamous cell carcinoma (TSCC)cells increases the expression of CXCR4, a chemokine receptor involved in tumor growth and metastasis. IL-1β also upregulates Notch expression, and Notch signaling may be involved in cancer progression through the upregulation of CXCR4.15 The high expression of IL-1β in tumor cells is also associated with lymph node metastasis. Overall, IL-1β has been identified as a key nodal gene in oral carcinogenesis and is a promising diagnostic marker and therapeutic target.16 IL-1RA is a naturally occurring receptor antagonist with two structural variants: the intracellular subtype (ICIL1RA) and the secretory subtype (SIL1RA). Both variants are upregulated in acute or chronic inflammatory responses. IL-1RA upregulation protects epithelial cells from environmental stimuli by competing with IL-1α.17 Furthermore, IL-1RA can inhibit tumor angiogenesis and metabolism by blocking IL-1β. Considering its anti-inflammatory and antioncogenic effects,17 enhancing the expression of IL-1RA can be a potential new strategy for cancer treatment.
Tumor Necrosis Factor (TNF)-α
Tumor necrosis factor (TNF)-α participates in inflammation, immune stress, and different stages of tumor progression, such as initiation, proliferation, angiogenesis, invasion, and metastasis. TNF-α exerts its effects through two distinct receptors: TNFR1 and TNFR2.TNFR1 mediates inflammatory responses and activates apoptosis signaling; TNFR2regulates cell adhesion, migration, proliferation, and survival.18
Studies have shown that TNF-α is closely related to inflammation, angiogenesis, and lymphangiogenesis in HNSCC. It upregulates vascular endothelial growth factor (VEGF) via the NF-κB pathway, and then promotes the formation of blood vessels and lymphatic vessels.19 TNF-α also facilitates EMT and tumor metastasis through transforming growth factor (TGF)-β-dependent mechanisms.20 Increased TNF-α expression in oral cancer cells has a relation to enhanced proliferation, and its downregulation can weaken this proliferative potency.21 Sandra et al showed that TNF-α inhibits the apoptosis of HNSCC cells by activating the phosphatidylinositol-3-OH kinase (PI3K) and NF-κB pathways.22 Moreover, TNF-α also mediates tumor cell proliferation and invasion by upregulating CXCR4.23 The upregulation of matrix metalloproteinases (MMPs), particularly MMP-9, is characteristic of HNSCC. Increased MMP-9 protein expression has been detected in the saliva protein database of oral cancer. Ruokolainen et al demonstrated that MMP-9 is a prognostic marker of HNSCC.24,25 Additionally, MMP-9 promotes TGF-β1-induced EMT by upregulating vimentin and downregulating E-cadherin, and aberrantly high MMP-9 expression can disintegrate the basement membrane, thereby facilitating tumor invasion and metastasis.26 MMP-9 expression in laryngeal tumors is connected to higher vascular density, and increased MMP-9 expression in HNSCC is closely related with invasion, metastasis, and angiogenesis.27 TNF-α binds to the MMP-9 promoter and enhances its transcription. TNF-α-induced EMT may also be associated with the induction of cancer stem cells (CSCs).28,29 CD44, a marker of CSCs, is correlated to EMT, drug resistance, and apoptosis inhibition in HNSCC and other tumors. Multiple studies have shown a positive correlation between CD44 expression and the mesenchymal phenotype and metastasis in various cancers.30 Moreover, TNF-α is a potential biomarker for predicting HNSCC patient survival and personalized treatment based on clinical TNM staging.31
NF-κB
NF-κB is a transcriptional factor regulating the expression of inflammation- and survival-related cytokines and other factors. The NF-κB family consists of five members: NF-κB1(p50), NF-κB2 (p52), RelA (p65), RelB, and c-Rel.32 NF-κB can be activated by the classical(canonical) or alternative (noncanonical) pathways. The classical pathway is stimulated by TNF-α, IL-1, and lipopolysaccharide (LPS), whereas the alternative pathway is initiated by specific subgroups of TNF family cytokines and other factors, such as CD40L, BAFF, and lymphotoxin-beta (LTβ).33 Following activation, NF-κB dimers translocate to the nucleus and bind to the promoter or enhancer regions of target genes, and thus accelerate tumor proliferation, metastasis, and drug resistance. NF-κB-regulated target genes include cell cycle regulators (eg, cyclin D1, c-MYC, and others) and apoptosis-related factors that either interfere with caspase activation and cytochrome c release (eg, BCL-XL, A1/Bfl-1) induced by anticancer drugs or inhibit the pro-apoptotic JNK pathway.34 NF-κB promotes tumor progression by upregulating MMP-935 and induces EMT and metastasis following activation by IL-8 and EGF.36
NF-κB participates in the development of HNSCC associated with viral pathogens and carcinogens. NF-κB protein is typically activated in nasopharyngeal carcinoma (NPC), which can be triggered by Epstein-Barr virus (EBV) infection, and its overexpression has a relation to poor prognosis. Carcinogen-related HNSCC is linked to tobacco use and betel nut chewing. Cigarette smoke contains polycyclic aromatic hydrocarbons and reactive oxygen species (ROS) that can damage DNA and stimulate the secretion of proinflammatory cytokines such as TNF-α and IL-1 from the epithelial cells. The binding of damaged DNA molecules, TNF-α, and IL-1 to their cognate receptors activates the IKK-NF-κB signaling pathway.37 Tobacco and betel nut consumption have a relation to decreased IκBα expression and NF-κB pathway activation in head and neck tumors.38,39 NF-κB activation in HNSCC cells contributes to blood and lymphatic metastasis via the activation of the pro-angiogenic VEGF and inducible nitric oxide (NO) synthase (iNOS). The iNOS-generated NO is related to increased tumor vascular density, growth, invasion, and metastasis.40,41 In line with these findings, NF-κB inhibitors have been shown to inhibit the invasion and metastasis of HNSCC cells in vitro, warranting further clinical investigation. Furthermore, given its involvement in tumor radio resistance, NF-κB is a potential marker of post radiotherapy prognosis.42
Transforming Growth Factor (TGF)-β
TGF-β is a pleiotropic cytokine that regulates various physiological functions, including cell growth, differentiation, apoptosis, migration, inflammation, and angiogenesis. The TGF-βsuperfamily includes TGFβ1, TGF-β2, and TGF-β3.43 Depending on the anatomical location and cellular environment, TGF-β signaling exerts both tumor-suppressive and tumor-promoting functions via the Smad family proteins.44,45 Smad2, TGF-βRII, and Smad4 primarily function as tumor suppressors but may promote tumor progression through TGF-βsignaling.46
Smad4 and TGF-βRII are frequently deleted or downregulated in squamous cell carcinoma.47,48 The loss of Smad4 disrupts TGF-β-mediated growth inhibition, leading to excessive cell proliferation, reduced apoptosis, and increased Smad3-driven inflammation. In addition, the loss of Smad4 can contribute to cetuximab resistance via the JNK and MAPK pathways. While Smad4 is necessary for TGF-β-induced EMT, its loss may facilitate EMT through alternative pathways.49–51 By contrast, the loss of TGF-βRII promotes Ras-driven malignant progression, resulting in the decreased expression of cyclin-dependent kinase inhibitors p15 and p21 and increased expression of cyclin-D and EGFR. Furthermore, the loss of TGF-βRII enhances the expression of endogenous TGF-β1, thus promoting inflammation and angiogenesis to facilitate tumor progression in HNSCC. TGF-βRII loss also augments cell migration and invasion through integrin-FAK-Src signaling.52 Tumor cells lacking Smad2, TGF-βRII, or Smad4 are resistant to the inhibitory effects of TGF-β signaling and undergo EMT.53 Additionally, TGF-β can induce the transformation of normal fibroblasts into CAFs by regulating fibronectin and MET, thereby promoting tumor progression.54 Consequently, TGF-β is a potential biomarker of HNSCC, and the selective blockade of the TGF-β/Smad signaling is a novel therapeutic strategy.55
Il-17
IL-17 is a pro-inflammatory cytokine that maintains mucosal immunity and barrier integrity, and therefore fosters inflammation in TME and facilitates tumor progression.56 Numerous studies have reported that IL-17 levels are elevated in HNSCC and that targeting IL-17 production can impede tumor cell proliferation.57,58 IL-17 promotes tumor cell proliferation by activating the JAK/STAT3 pathway59 and inhibits apoptosis by blocking FAS-associated death domain protein through the PI3K/AKT pathway. Consistent with this, silencing IL-17 expression in laryngeal cancer cells triggered apoptosis through the PI3K/AKT/FAS/FASL pathway.60 Furthermore, IL-17 can accelerate the invasion and metastasis of thyroid cancer cells in a dose-dependent manner by activating ERK1/2 and upregulating MMP-9.61 IL-17 is also a potential prognostic marker of HNSCC, as high levels of IL-17 have been significantly associated with reduced 5-year survival rates, and IL-17 expression was inversely correlated with overall survival in HNSCC.62,63
Role of Inflammation-Related Pathways in the Progression of HNSCC
Inflammatory pathways are of utmost importance in the pathogenesis of HNSCC and can offer novel therapeutic targets. These pathways and their roles in the development and progression of HNSCC have been discussed in the following sections (Figure 2).
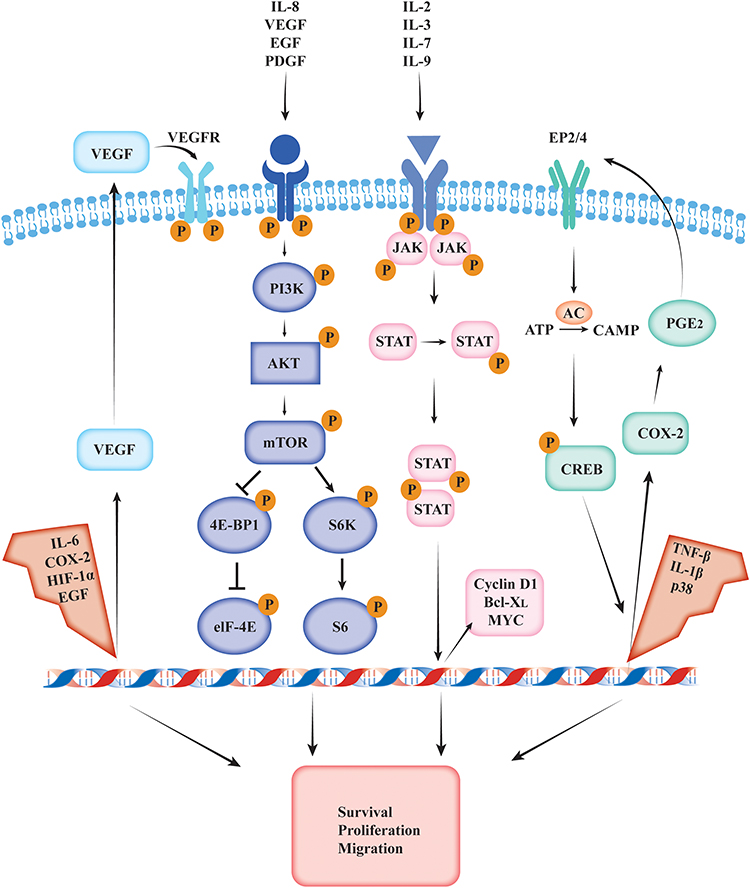 |
Figure 2 VEGF, mTOR, STAT, and COX-2 inflammation-related signaling pathways involved in head and neck squamous cell carcinoma progression. |
VEGF Pathway
The VEGF family comprises seven human-origin subtypes: VEGF-A, -B, -C, -D, -E, -F, and PLGF.64 VEGF ligands signal through cell surface receptor tyrosine kinases, notably VEGFR- 1 (Flt-1), VEGFR-2 (Flk-1/FDR), and VEGFR-3 (Flt-4). VEGFR-1 is primarily expressed in hematopoietic stem cells, macrophages, monocytes, and vascular endothelial cells, and its function is contingent on the tissue or cell type during development. VEGFR-2 primarily regulates angiogenesis, endothelial cell mitosis and chemotaxis, and vascular permeability, whereas VEGFR-3 promotes lymphangiogenesis.65
VEGF accelerates tumor progression by enhancing vascular permeability, which leads to the extravasation of high molecular weight molecules and development of edema, and consequently facilitates neovascularization.66 It is regulated by hypoxia-inducible factor (HIF)- 1α in the low oxygen conditions of TME.67,68 Moreover, VEGF inhibits endothelial cell apoptosis by activating the anti-apoptotic kinase Akt/PKB and upregulating Bcl-2 and A1.69 Wu et al demonstrated that VEGF-A blockade delayed EMT and angiogenesis in NPC.70 The thyroid-stimulating hormone (TSH) regulates the vascularization and growth of thyroid tumors by stimulating VEGF secretion from cancer cells.71 Sun et al showed that the inhibition of the VEGF/VEGFR2 pathway using proanthocyanidin B2 (PB2) suppressed angiogenesis and proliferation in OSCC.72
The interaction between VEGF-C/D and their receptor VEGFR3 stimulates the proliferation and migration of lymphatic endothelial cells and thus promotes lymphatic vessel formation and potentially contributes to lymph node metastasis.73 Li et al showed that EBV-infected NPC cells facilitated LN metastasis by inducing cancer-associated lymphangiogenesis and that the administration of anti-VEGF-C antibodies inhibited this process.74 Furthermore, VEGF is a valuable prognostic indicator of various tumors. Ceric et al identified VEGF-C as a risk factor of papillary thyroid carcinoma (PTC) and found that VEGF-C expression correlated significantly with tumor size and recurrence.75 Huang et al further demonstrated that VEGF-C is a noninvasive diagnostic biomarker of laryngeal squamous cell carcinoma (LSCC).76
Mechanistic Target of Rapamycin (mTOR) Pathway
The mechanistic target of rapamycin (mTOR) exists in two distinct protein complexes (mTORC1 and mTORC2). mTORC1 is involved in protein synthesis, autophagy, and metabolic regulation, and mTORC2 contributes to actin cytoskeleton polarization.77 The PI3K/AKT/mTOR pathway is involved in cell survival, migration, proliferation, and differentiation as well as in physiological processes such as angiogenesis and protein synthesis.78 The downstream targets of mTOR signaling regulate multiple inflammatory factors, eg, cytokines, chemokines, proliferation, invasion, extracellular matrix (ECM) remodeling, and fibrosis.77 Thus, mTOR is a potential therapeutic target for acute and chronic inflammatory conditions. The PI3K/AKT/mTOR pathway is frequently dysregulated in tumor cells.
The mTOR pathway controls tumorigenesis by regulating protein synthesis, translation of oncogenic proteins, autophagy, and apoptosis. The downstream effectors of mTORC1, including ribosomal protein S6 kinase 1 (S6K1) and eukaryotic translation initiation factor 4E- binding protein 1 (4EBP1), promote angiogenesis by regulating HIF-1α at the translational level.79 The mTOR pathway is activated in HNSCC cells,80 and inhibiting this pathway can impede their growth and metastasis.81,82
The PI3K/AKT/mTOR pathway is also related to the proliferation and invasion of OSCC cells.83 The inactivation of the AKT/mTOR pathway using glioma-associated oncogene (Gli) antagonist-61 (GANT61) inhibited the survival, invasion, and EMT of anaplastic thyroid cancer (ATC) cells, whereas its activation in thyroid cancer cells promoted stemness and metastasis.84,85 Huang et al demonstrated that knocking out placenta specific gene 8 (PLAC8) in NPC tissue induced autophagy and apoptosis in tumor cells, and inhibited proliferation and EMT via the inhibition of the mTOR pathway.86 Besides, Yu et al found a correlation between the mTOR pathway and the T stage, N stage, clinical stage, recurrence, and distant metastasis in HNSCC; the activation of the mTOR pathway was shown to be related to shorter survival and lower survival rates in NPC. Therefore, mTOR is a promising prognostic biomarker of head and neck cancer.87
Signal Transducers and Activators of Transcription (STAT) Pathway
The mammalian signal transducers and activators of transcription (STAT) family consists of seven members: STAT1, STAT2, STAT3, STAT4, STAT5a, STAT5b, and STAT6. The STAT signaling pathway engages in cellular proliferation, differentiation, and apoptosis, along with the regulation of cytokine-dependent inflammation and immunity.88 Mutations in STAT family proteins are essential for the induction and maintenance of pro-cancerous inflammatory microenvironments. In addition, the JAK/STAT pathway is aberrantly activated by autocrine/paracrine cytokines as well as by mutations in STAT and upstream oncogenes in various cancers. STAT3 is known to upregulate anti-apoptotic proteins (such as BCL-XL and C-MYC) and facilitate cell survival and mitosis induced by Src-mediated signaling.89,90 STATs are also connected to the progression of HNSCC. Multiple studies have indicated that this pathway promotes the proliferation and survival of HNSCC cells by regulating factors involved in cell cycle progression (eg, cyclin D1) and apoptosis (BclxL, Bcl-2, Mcl-1, Fas, etc.).91,92 Furthermore, STATs can enhance tumor invasion and metastasis by downregulating the mucin-like glycoprotein episialin (MUC1) and reducing cell adhesion.93 STAT3 expression is negatively correlated to that of the epithelial factor E-cadherin in squamous cell carcinoma, and the inhibition of E-cadherin leads to tumor cell invasion and metastasis.94 STAT3 also facilitates tumor invasion by activating the MMPs (MMP-2, MMP-1, MMP-9, and MMP-10), and STATs are known to enhance tumor angiogenesis by upregulating VEGF via HIF-1α.91,95 The activation of the JAK2/STAT3 pathway in follicular thyroid carcinoma (FTC) cells enhanced their proliferation, EMT, and invasion, whereas its inactivation suppressed tumor cell proliferation and promoted apoptosis.96 The persistent activation of the JAK/STAT signaling pathway accelerated the proliferation and invasion of NPC cells, and the targeting of the key genes of this pathway had an inhibitory effect on tumor cell proliferation, invasion, and metastasis in vitro and in vivo.97,98
The JAK/STAT pathway is associated with the chemoresistance of tumor cells. Cisplatin resistance in HNSCC cells is induced by activated STAT through AKR1C1, a member of the aldo-keto reductase 1 family.99 Although BRAFV600E is a common mutation in thyroid cancer, the cells routinely develop resistance to BRAFV600E inhibitors through the JAK/STAT pathway.100 Therefore, the STAT pathway is a promising therapeutic target for recalcitrant HNSCC.101
Cyclooxygenase (COX)-2 Pathway
Two subtypes of cyclooxygenase (COX)—the constitutive COX-1 and inducible COX-2—have been identified in mammals.102 The COX-2 pathway is involved in the conversion of arachidonic acid to prostaglandins, which regulate physiological and pathological processes such as vascular constriction, vasodilation, and inflammation.103
While COX-2 is expressed at low levels in normal tissues, it is induced in the early stages of cancer. High COX-2 expression contributes to apoptosis resistance, proliferation, transformation, angiogenesis, inflammation, invasion, metastasis, chemoresistance, and poor prognosis in several cancers. The inhibition of the COX-2 pathway can inhibit tumor progression.104,105 Zhu et al found that the high expression of COX-2 in the CAFs was related with poor survival rates and distant metastasis in NPC patients and that COX-2 was overexpressed in CAFs at the site of distant metastasis.106 Epidermal growth factor receptor (EGFR) overexpression induces the generation of the COX-2 metabolite PGE2 via angiopoietin-like 4 (ANGPTL4), which promotes angiogenesis, EMT, and tumor metastasis.107 COX-2 can also accelerate tumor invasion by regulating the expression of VEGF to facilitate lymphangiogenesis.108 The carcinogenic potential of COX-2 is likely driven by its downstream proinflammatory factors such as PGE2. The latter suppresses T and B lymphocyte proliferation and natural killer cell function and mediates chronic inflammation by promoting vasodilation and angiogenesis.109,110
COX-2 expression is significantly correlated to lymph node metastasis in NPC patients.111 In fact, NPC has the highest metastasis rate among head and neck cancers, and one of the main reasons for treatment failure is distant metastasis.112 Therefore, COX-2 is a prognostic biomarker of NPC and other HNSCCs, and the decrease in its expression before and after chemotherapy has been shown to be connected with the survival duration and survival rate of NPC patients.113 Finally, high COX-2 expression is an independent predictor of adverse long-term outcomes in non-metastatic NPC, regardless of the clinical stage.114
Inflammation-Associated Cells Mediate the Progression of HNSCC
Inflammatory cells are a critical component of TME that promotes tumor development and immune responses. The functions of different inflammation-related cells in HNSCC progression have been discussed below.
Tumor-Associated Macrophages (TAMs)
Monocytes derived from the bone marrow differentiate into macrophages at the site of inflammation under the influence of growth factors. Macrophages are versatile immune cells that regulate tissue homeostasis, phagocytose pathogens, and promote wound healing.115 TAMs infiltrate into the tumor tissues or reside within TME and regulate tumor growth, tumor angiogenesis, immune modulation, metastasis, and chemoresistance. Activated macrophages can be classified into the classically activated M1 and alternatively activated M2 phenotypes.116 The M1 macrophages promote antitumor Th1 responses, whereas M2 macrophages facilitate tissue repair and Th2 responses as well as exhibit a tumor-promoting phenotype.117 Although M1 and M2 polarization is finely balanced in many tumors, TAMs typically exhibit functions similar to M2 macrophages, then accelerate tumor progression.118 Macrophage polarization in TME is driven by multiple cytokines and growth factors, such asIL-4 produced by the CD4+ T cells and/or tumor cells as well tumor cell-derived CSF1 andGM-CSF.119
TAMs facilitate the growth of HNSCC tumors through epidermal growth factor (EGF)signaling. Furthermore, the infiltrating TAMs enhance lymphatic vessel density and promote angiogenesis in HNSCC tissues, thereby facilitating tumor metastasis. TAMs are highly concentrated in the hypoxic and avascular regions of tumors, in which they upregulate VEGF, HIF-regulated genes, and MMPs, resulting in vascular regeneration and allowing the cancer cells to survive in the adverse TME.120,121 Pirilä et al have shown that TAMs regulate the adhesion, migration, and invasion of human TSCC cells.122 Macrophages promote the adhesion of tumor cells to the ECM by secreting bone sialoprotein (also known as SPARC) and facilitate tumor EMT, invasion, and metastasis through TGF-β.123,124 Additionally, macrophages express immune checkpoints such as programmed cell death protein 1 (PD-1) and cytotoxic T lymphocyte antigen 4 (CTLA-4), which suppress the activity of T cells, B cells, and other immune cells by inhibiting TCR and BCR signaling.119 There is experimental evidence suggesting that the quantity of TAMs in the tumor tissues or periphery is predictive of disease progression and 5-year survival in oral cancer patients.125
Myeloid-Derived Suppressor Cells (MDSCs)
Myeloid-derived suppressor cells (MDSCs) are a heterogeneous population consisting of immature dendritic cells, granulocytes, macrophages, and bone marrow progenitor cells that infiltrate TME and enable the tumor cells to evade immune responses. An immunosuppressive TME prevents excessive tissue damage caused by unresolved inflammation and fosters tumor progression.126 MDSCs identified in human malignancies are typically characterized by the expression of CD33 and CD11b. They inhibit T-cell immune responses through the following mechanisms: (1) upregulation of arginase and subsequent depletion of arginine, which is crucial for T-cell proliferation; (2) production of ROS and iNOSto impair cytotoxic T-cell activity; and (3) suppression of T cell function by producing peroxynitrite (ONOO-).127 Recent studies have shown that MDSCs can accelerate tumor angiogenesis by differentiating into endothelial-like cells (ECs) that produce high levels of VEGF. Besides, MDSCs also facilitate metastasis by stimulating MMP-9 production and promoting EMT.128,129
MDSCs have been implicated in the progression of HNSCC. Pang et al reported an increased abundance of MDSCs in OSCC tissues; these cells facilitated tumor cell proliferation, invasion, metastasis, and angiogenesis in OSCC. Conversely, the microenvironment of OSCC is conducive to the infiltration and function of MDSCs.130 Furthermore, inhibiting the accumulation and function of MDSCs can slow the development of oral cancer, and then highlight the potential of targeting MDSCs as a therapeutic strategy for head and neck tumors.131
Intratumoral MDSCs can serve as potential marker of prognosis and tumor stage. For instance, Angell et al demonstrated that MDSC evaluation is capable of predicting the risk and stage of thyroid cancer cases. Among patients who underwent surgery for a solitary thyroid nodule, malignant nodules exhibited a significantly higher average proportion of granulocytic CD11bHLA-DR + lowHIF1a MDSCs than benign nodules. In addition, measuring the phenotype of these MDSCs may be useful for predicting various cancer types.132 Furthermore, blocking MDSC infiltration into HNSCC tumors can restore T cell function and antitumor responses, therefore potentially improving prognosis.133
Tumor-Infiltrating Lymphocytes (TILs)
Tumor-infiltrating lymphocytes (TILs) encompass a diverse range of immune cells and constitute a crucial component of TME. While T cells are significant in cancer, the presence and functional significance of other cell types remains to be elucidated. T lymphocytes can be categorized into the αβ and γδ types based on their surface receptors. The αβ T cells are the predominant type and further differentiate into CD3+ CD4+ helper T cells and CD3+ CD8+cytotoxic T cells, which consist of Th1, Th2, Th9, Th17, Th22, Tfh, and regulatory T (Treg) cell subsets.134 The different T cell subsets exert varying effects on TME. For instance, the CD8+T cells and Th1 cells exhibit anti-inflammatory effects and restrain tumor growth, whereasTh22 cells promote tumor growth by secreting IL-22.135,136
With recent advances in immunotherapy, numerous studies have explored the prognostic value of TILs in cancer. The TIL load is an independent prognostic factor of HNSCC, and the high infiltration of CD8+ TILs is associated with improved survival outcomes. The infiltration of functional effector T cells induces the expression of various immune checkpoint molecules, such as indoleamine 2,3-dioxygenase (IDO) and programmed cell death ligand 1 (PD-L1),137 which are related to a favorable prognosis. PD-L1 promotes the infiltration of CD8+ TILs and improves the accuracy of TNM staging for HNSCC.138 Furthermore, TILs can predict tumor sensitivity to radiotherapy and chemotherapy. For example, the high density of CD8+ TILs is an effective biomarker of the sensitivity to chemoradiotherapy in nasopharyngeal and oropharyngeal cancer patients.139
Clinical Use of Inflammation-Targeted Drugs in HNSCC
The high incidence, poor prognosis, and therapeutic recalcitrance of HNSCC necessitate the identification of novel molecular therapeutic targets. Given the critical role of inflammation in the progression of HNSCC, immunotherapy with a specific focus on TME is a promising intervention. The current progress in the immunotherapeutic strategies against HNSCC is summarized in this section (Table 1).
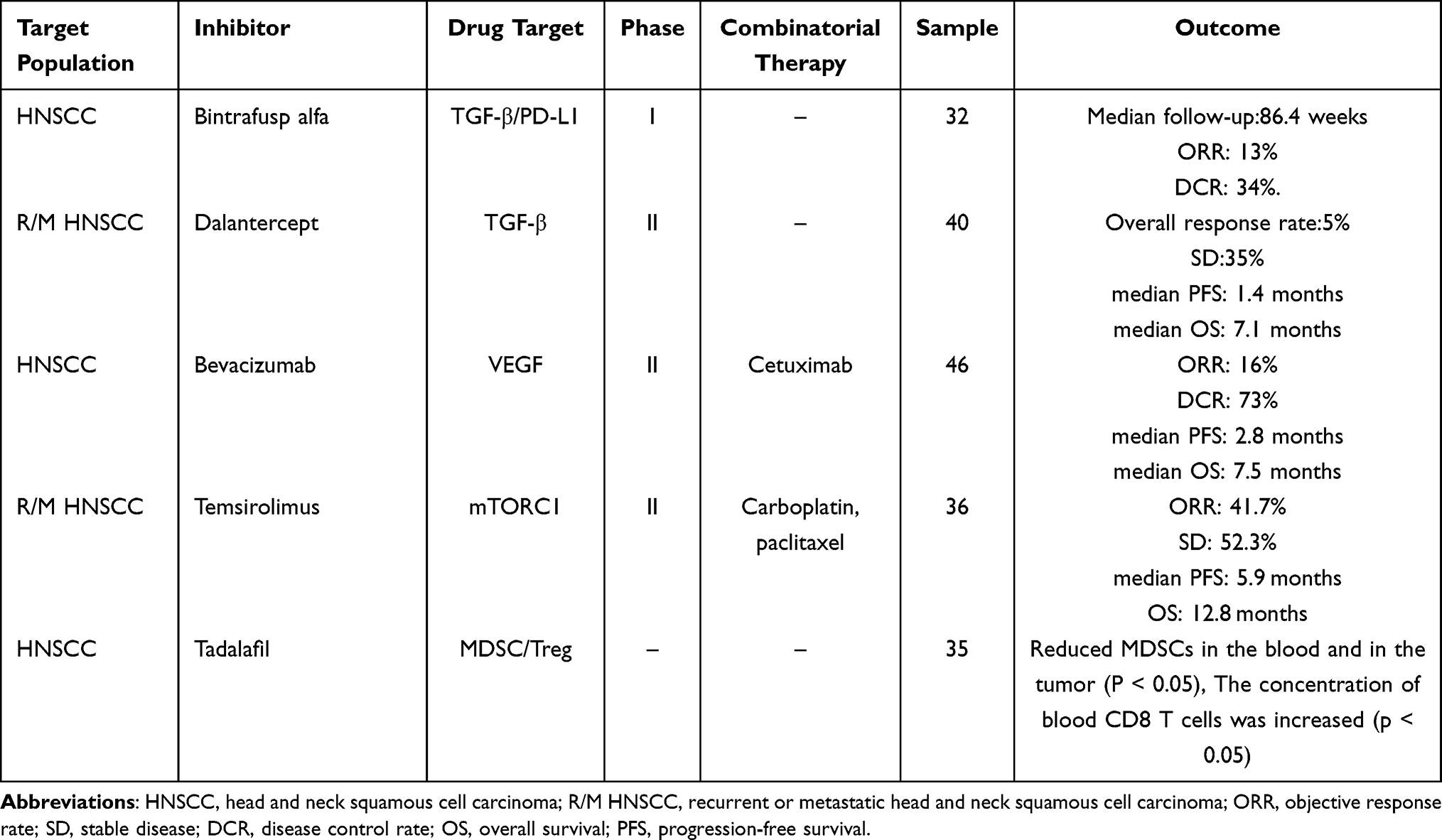 |
Table 1 Evaluation of Inflammation as a Therapeutic Target in Head and Neck Squamous Cell Carcinoma |
Bintrafusp alfa is a bifunctional fusion protein that targets both TGF-β and PD-L1. It has demonstrated superior efficacy in suppressing tumor progression than using TGF-β trap or anti-PD-L1 antibodies alone, making it a promising treatment option for patients with advanced HNSCC. Bintrafusp alfa was found to have a manageable safety profile in clinica ltrials, with potential TGF-β-related adverse events being observed and managed in a timely manner.140
Dalantercept is a fusion protein that blocks activin receptor-like kinase 1 (ALK1), a type I receptor of the TGF-β superfamily, and inhibits tumor angiogenesis. It has implied a dose-dependent antitumor effect with acceptable toxicity against highly vascularized recurrent or metastatic HNSCC (R/M HNSCC). Further research is warranted to explore the potential of dalantercept in combination with radiotherapy and chemotherapy.141
Bevacizumab is an anti-VEGF monoclonal antibody that has been used in combination with anti-EGFR antibody cetuximab in Phase 2 trials for HNSCC. The simultaneous blockade of VEGF and EGFR has achieved significant antitumor effects in vivo and in vitro by inhibiting neo-angiogenesis. In addition, the combination therapy has demonstrated good tolerability and activity in previously treated patients.142
Temsirolimus is an mTOR inhibitor that acts synergistically with cisplatin and paclitaxel. The combination of temsirolimus with low-dose cisplatin and paclitaxel is associated with a lower rate of treatment discontinuation due to adverse events than the current first-line standard EXTREME regimen, indicating acceptable safety. In phase 2 trials of HNSCC, temsirolimus-based treatment showed higher response rate than the standard treatment. Everolimus, also an mTOR inhibitor, was ineffective against R/M HNSCC in a phase 2 trial when used as a monotherapy. However, the combination of everolimus with cetuximab and cisplatin as a first-line treatment for R/M HNSCC achieved 60% objective response rate. The enhanced response rate may be attributed to the synergistic effect of mTOR inhibition and cytotoxic chemotherapy.143
The infiltration of MDSCs and regulatory T cells (Tregs) into TME suppresses the immune response and is often associated with poor prognosis. Tadalafil has been shown to significantly reduce the number of circulating and intratumoral MDSCs and Tregs in Patients with HNSCC through phosphodiesterase-5 inhibition, resulting in the increased proportion of anti-tumor CD8+ T cells. Tadalafil exhibits the greatest immunomodulatory activity at moderate doses, whereas high doses may have negative effects on tumor immunity.144
Conclusion
Inflammation is a key factor in the initiation and progression of HNSCC and offers novel therapeutic targets and prognostic indicators for effective tumor management. Inflammatory factors fuel tumor growth and angiogenesis, and should be investigated as potential therapeutic targets against cancer. While several inflammation-related factors have been identified as the diagnostic and prognostic biomarkers of HNSCC, further prospective research is warranted before clinical translation. The continued exploration of the complex interplay between inflammation and HNSCC may shed light on innovative therapeutic approaches and improve patient outcomes.
Funding
The present study was supported in part by Guangdong Basic and Applied Basic Research Foundation (2021A1515010970); Shenzhen Innovation of Science and Technology Commission (No. JCYJ20210324132407019, LGKCYLWS2022002, LGKCYLWS2021000027, and LGKCYLWS2020099); and Shenzhen Key Medical Discipline Construction Fund (No. SZXK039).
Disclosure
The authors have no relevant financial or non financial interests to disclose for this work.
References
1. Argiris A, Karamouzis MV, Raben D, Ferris RL. Head and neck cancer. Lancet. 2008;371(9625):1695–1709. doi:10.1016/S0140-6736(08)60728-X
2. Tang YC, Hsiao JR, Jiang SS, et al. c-MYC-directed NRF2 drives malignant progression of head and neck cancer via glucose-6-phosphate dehydrogenase and transketolase activation. Theranostics. 2021;11(11):5232–5247. doi:10.7150/thno.53417
3. Yu MZ, Wu MM, Chien HT, et al. Risk prediction models for patients with head and neck cancer among the Taiwanese population. Cancers (Basel). 2022;14(21):5338. doi:10.3390/cancers14215338
4. Belcher R, Hayes K, Fedewa S, Chen AY. Current treatment of head and neck squamous cell cancer. J Surg Oncol. 2014;110(5):551–574. doi:10.1002/jso.23724
5. Kumar D, New J, Vishwakarma V, et al. Cancer-associated fibroblasts drive glycolysis in a targetable signaling loop implicated in head and neck squamous cell carcinoma progression. Cancer Res. 2018;78(14):3769–3782. doi:10.1158/0008-5472.CAN-17-1076
6. Hanahan D, Weinberg RA. Hallmarks of cancer: the next generation. Cell. 2011;144(5):646–674. doi:10.1016/j.cell.2011.02.013
7. Greten FR, Grivennikov SI. Inflammation and cancer: triggers, mechanisms, and consequences. Immunity. 2019;51(1):27–41. doi:10.1016/j.immuni.2019.06.025
8. Dinarello CA. Overview of the IL-1 family in innate inflammation and acquired immunity. Immunol Rev. 2018;281(1):8–27. doi:10.1111/imr.12621
9. Paish HL, Kalson NS, Smith GR, et al. Fibroblasts promote inflammation and pain via IL-1alpha induction of the Monocyte Chemoattractant Chemokine (C-C Motif) ligand 2. Am J Pathol. 2018;188(3):696–714. doi:10.1016/j.ajpath.2017.11.007
10. Stevenson FT, Turck J, Locksley RM, Lovett DH. The N-terminal propiece of interleukin 1 alpha is a transforming nuclear oncoprotein. Proc Natl Acad Sci U S A. 1997;94(2):508–513. doi:10.1073/pnas.94.2.508
11. Leon X, Bothe C, Garcia J, et al. Expression of IL-1alpha correlates with distant metastasis in patients with head and neck squamous cell carcinoma. Oncotarget. 2015;6(35):37398–37409. doi:10.18632/oncotarget.6054
12. Li T, Cheng D, Guo J, et al. SOX9 and IL1A as the potential gene biomarkers of the oral cancer. Comb Chem High Throughput Screen. 2023;26(8):1461–1479. doi:10.2174/1386207325666220628091041
13. Lee CH, Chang JS, Syu SH, et al. IL-1beta promotes malignant transformation and tumor aggressiveness in oral cancer. J Cell Physiol. 2015;230(4):875–884. doi:10.1002/jcp.24816
14. Chen LC, Wang LJ, Tsang NM, et al. Tumour inflammasome-derived IL-1beta recruits neutrophils and improves local recurrence-free survival in EBV-induced nasopharyngeal carcinoma. EMBO Mol Med. 2012;4(12):1276–1293. doi:10.1002/emmm.201201569
15. Sun Y, Zhu D, Wang G, et al. Pro-inflammatory cytokine IL-1beta up-regulates CXC chemokine receptor 4 via Notch and ERK signaling pathways in tongue squamous cell carcinoma. PLoS One. 2015;10(7):e0132677. doi:10.1371/journal.pone.0132677
16. Singh P, Augustine D, Rao RS, et al. Interleukin-1beta and Caspase-3 expression serve as independent prognostic markers for metastasis and survival in oral squamous cell carcinoma. Cancer Biomark. 2019;26(1):109–122. doi:10.3233/CBM-190149
17. Ding Y, Yi J, Wang J, Sun Z. Interleukin-1 receptor antagonist: a promising cytokine against human squamous cell carcinomas. Heliyon. 2023;9(4):e14960. doi:10.1016/j.heliyon.2023.e14960
18. Li P, Yang Y, Yang X, et al. TNFR2 deficiency impairs the growth of mouse colon 553 cancer. Int J Biol Sci. 2023;19(4):1024–1035. doi:10.7150/ijbs.72606
19. Zhang C, Zhu M, Wang W, et al. TNF-alpha promotes tumor lymph angiogenesis in head and neck squamous cell carcinoma through regulation of ERK3. Transl Cancer Res. 2019;8(6):2439–2448. doi:10.21037/tcr.2019.09.60
20. Yoshimatsu Y, Wakabayashi I, Kimuro S, et al. TNF-alpha enhances TGF-beta-induced endothelial-to-mesenchymal transition via TGF-beta signal augmentation. Cancer Sci. 2020;111(7):2385–2399. doi:10.1111/cas.14455
21. Qiu YF, Wang MX, Meng LN, Zhang R, Wang W. MiR-21 regulates proliferation and apoptosis of oral cancer cells through TNF-alpha. Eur Rev Med Pharmacol Sci. 2018;22(22):7735–7741. doi:10.26355/eurrev_201811_16395
22. Sandra F, Matsuki NA, Takeuchi H, et al. TNF inhibited the apoptosis by activation of Akt serine/threonine kinase in the human head and neck squamous cell carcinoma. Cell Signal. 2002;14(9):771–778. doi:10.1016/S0898-6568(02)00025-6
23. Han N, Li X, Wang Y, et al. Increased tumor-infiltrating plasmacytoid dendritic cells promote cancer cell proliferation and invasion via TNF-alpha/NF-kappaB/CXCR-4 pathway in oral squamous cell carcinoma. J Cancer. 2021;12(10):3045–3056. doi:10.7150/jca.55580
24. Hohberger L, Wuertz BR, Xie H, Griffin T, Ondrey F. TNF-alpha drives matrix metalloproteinase-9 in squamous oral carcinogenesis. Laryngoscope. 2008;118(8):1395–1399. doi:10.1097/MLG.0b013e318174e09b
25. Ruokolainen H, Paakko P, Turpeenniemi-Hujanen T. Serum matrix metalloproteinase-9 in head and neck squamous cell carcinoma is a prognostic marker. Int J Cancer. 2005;116(3):422–427. doi:10.1002/ijc.21092
26. Li Y, He J, Wang F, et al. Role of MMP-9 in epithelial-mesenchymal transition of thyroid cancer. World J Surg Oncol. 2020;18(1):181. doi:10.1186/s12957-020-01958-w
27. Wittekindt C, Jovanovic N, Guntinas-Lichius O. Expression of matrix metalloproteinase-9 (MMP-9) and blood vessel density in laryngeal squamous cell carcinomas. Acta Otolaryngol. 2011;131(1):101–106. doi:10.3109/00016489.2010.506886
28. Zhang L, Jiao M, Wu K, et al. TNF-alpha induced epithelial mesenchymal transition increases stemness properties in renal cell carcinoma cells. Int J Clin Exp Med. 2014;7(12):4951–4958.
29. Gomez KE, Wu F, Keysar SB, et al. Cancer cell CD44 mediates macrophage/monocyte- driven regulation of head and neck cancer stem cells. Cancer Res. 2020;80(19):4185–4198. doi:10.1158/0008-5472.CAN-20-1079
30. Chen C, Zhao S, Karnad A, Freeman JW. The biology and role of CD44 in cancer progression: therapeutic implications. J Hematol Oncol. 2018;11(1):64. doi:10.1186/s13045-018-0605-5
31. Andersson BA, Lewin F, Lundgren J, et al. Plasma tumor necrosis factor-alpha and C-reactive protein as biomarker for survival in head and neck squamous cell carcinoma. J Cancer Res Clin Oncol. 2014;140(3):515–519. doi:10.1007/s00432-014-1592-8
32. Liu T, Zhang L, Joo D, Sun SC. NF-kappaB signaling in inflammation. Signal Transduct Target Ther. 2017;2:17023. doi:10.1038/sigtrans.2017.23
33. Luo JL, Kamata H, Karin M. IKK/NF-kappaB signaling: balancing life and death--a new approach to cancer therapy. J Clin Invest. 2005;115(10):2625–2632. doi:10.1172/JCI26322
34. Pacifico F, Leonardi A. Role of NF-kappaB in thyroid cancer. Mol Cell Endocrinol. 2010;321(1):29–35. doi:10.1016/j.mce.2009.10.010
35. Wang S, Liu Q, Zhang Y, et al. Suppression of growth, migration and invasion of highly- metastatic human breast cancer cells by berbamine and its molecular mechanisms of action. Mol Cancer. 2009;8:81. doi:10.1186/1476-4598-8-81
36. Ma T, Zhao Z, Wang Z, Wang C, Zhang L. MiR-940 inhibits migration and invasion of tongue squamous cell carcinoma via regulatingCXCR2/NF-kappaB system-mediated epithelial-mesenchymal transition. Naunyn Schmiedebergs Arch Pharmacol. 2019;392(11):1359–1369. doi:10.1007/s00210-019-01671-w
37. Churg A, Dai J, Tai H, Xie C, Wright JL. Tumor necrosis factor-alpha is central to acute cigarette smoke-induced inflammation and connective tissue breakdown. Am J Respir Crit Care Med. 2002;166(6):849–854. doi:10.1164/rccm.200202-097OC
38. Anto RJ, Mukhopadhyay A, Shishodia S, Gairola CG, Aggarwal BB. Cigarette smoke condensate activates nuclear transcription factor-kappaB through phosphorylation and degradation of IkappaB(alpha): correlation with induction of cyclooxygenase-2. Carcinogenesis. 2002;23(9):1511–1518. doi:10.1093/carcin/23.9.1511
39. Lin SC, Lu SY, Lee SY, et al. Areca (betel) nut extract activates mitogen-activated protein kinases and NF-kappaB in oral keratinocytes. Int J Cancer. 2005;116(4):526–535. doi:10.1002/ijc.21104
40. Zhang J, Peng B. NF-kappaB promotes iNOS and VEGF expression in salivary gland adenoid cystic carcinoma cells and enhances endothelial cell motility in vitro. Cell Prolif. 2009;42(2):150–161. doi:10.1111/j.1365-2184.2009.00588.x
41. Fahey JM, Korytowski W, Girotti AW. Upstream signaling events leading to elevated production of pro-survival nitric oxide in photodynamically-challenged glioblastoma cells. Free Radic Biol Med. 2019;137:37–45. doi:10.1016/j.freeradbiomed.2019.04.013
42. Nariai Y, Mishima K, Yoshimura Y, Sekine J. FAP-1 and NF-kappaB expressions in oral squamous cell carcinoma as potential markers for chemo-radio sensitivity and prognosis. Int J Oral Maxillofac Surg. 2011;40(4):419–426. doi:10.1016/j.ijom.2010.10.020
43. Bertolino P, Deckers M, Lebrin F, Ten DP. Transforming growth factor-beta signal transduction in angiogenesis and vascular disorders. Chest. 2005;128(6 Suppl):585S–590S. doi:10.1378/chest.128.6_suppl.585S
44. Wu F, Weigel KJ, Zhou H, Wang XJ. Paradoxical roles of TGF-beta signaling in suppressing and promoting squamous cell carcinoma. Acta Biochim Biophys Sin (Shanghai). 2018;50(1):98–105. doi:10.1093/abbs/gmx127
45. Massague J. TGFbeta signalling in context. Nat Rev Mol Cell Biol. 2012;13(10):616–630. doi:10.1038/nrm3434
46. Shi Y, Massague J. Mechanisms of TGF-beta signaling from cell membrane to the 628 nucleus. Cell. 2003;113(6):685–700. doi:10.1016/S0092-8674(03)00432-X
47. Hernandez AL, Wang Y, Somerset HL, et al. Inter- and intra-tumor heterogeneity of SMAD4 loss in head and neck squamous cell carcinomas. Mol Carcinog. 2019;58(5):666–673. doi:10.1002/mc.22958
48. Muro-Cacho CA, Anderson M, Cordero J, Munoz-Antonia T. Expression of transforming growth factor beta type II receptors in head and neck squamous cell carcinoma. Clin Cancer Res. 1999;5(6):1243–1248.
49. Natsugoe S, Xiangming C, Matsumoto M, et al. Smad4 and transforming growth factor beta1 expression in patients with squamous cell carcinoma of the esophagus. Clin Cancer Res. 2002;8(6):1838–1842.
50. Ozawa H, Ranaweera RS, Izumchenko E, et al. SMAD4 loss is associated with cetuximab resistance and induction of MAPK/JNK activation in head and neck cancer cells. Clin Cancer Res. 2017;23(17):5162–5175. doi:10.1158/1078-0432.CCR-16-1686
51. David CJ, Huang YH, Chen M, et al. TGF-beta tumor suppression through a lethal 641 EMT. Cell. 2016;164(5):1015–1030. doi:10.1016/j.cell.2016.01.009
52. Lu SL, Herrington H, Reh D, et al. Loss of transforming growth factor-beta type II receptor promotes metastatic head-and-neck squamous cell carcinoma. Genes Dev. 2006;20(10):1331–1342. doi:10.1101/gad.1413306
53. Pang X, Tang YL, Liang XH. Transforming growth factor-beta signaling in head and neck squamous cell carcinoma: insights into cellular responses. Oncol Lett. 2018;16(4):4799–4806. doi:10.3892/ol.2018.9319
54. Huang Q, Hsueh CY, Shen YJ, et al. Small extracellular vesicle-packaged TGFbeta1 promotes the reprogramming of normal fibroblasts into cancer-associated fibroblasts by regulating fibronectin in head and neck squamous cell carcinoma. Cancer Lett. 2021;517:1–13. doi:10.1016/j.canlet.2021.05.017
55. Zheng L, Guan Z, Xue M. TGF-beta signaling pathway-based model to predict the subtype and prognosis of head and neck squamous cell carcinoma. Front Genet. 2022;13:862860. doi:10.3389/fgene.2022.862860
56. Li X, Bechara R, Zhao J, McGeachy MJ, Gaffen SL. IL-17 receptor-based signaling and implications for disease. Nat Immunol. 2019;20(12):1594–1602. doi:10.1038/s41590-019-0514-y
57. Li FJ, Cai ZJ, Yang F, Zhang SD, Chen M. Th17 expression and IL-17 levels in laryngeal squamous cell carcinoma patients. Acta Otolaryngol. 2016;136(5):484–490. doi:10.3109/00016489.2015.1126857
58. Avadhani AV, Parachuru VP, Milne T, Seymour GJ, Rich AM. Multiple cells express interleukin 17 in oral squamous cell carcinoma. J Oral Pathol Med. 2017;46(1):39–45. doi:10.1111/jop.12465
59. Liu T, Zong S, Jiang Y, et al. Neutrophils promote larynx squamous cell carcinoma progression via activating the IL-17/JAK/STAT3 pathway. J Immunol Res. 2021;2021:8078646. doi:10.1155/2021/8078646
60. Song Y, Yang M, Zhang H, et al. IL-17 affects the progression, metastasis, and recurrence of laryngeal cancer via the inhibition of apoptosis through activation of the PI3K/AKT/FAS/FASL pathways. J Immunol Res. 2020;2020:2953191. doi:10.1155/2020/2953191
61. Ren L, Xu Y, Liu C, Wang S, Qin G. IL-17RB enhances thyroid cancer cell invasion and metastasis via ERK1/2 pathway-mediated MMP-9 expression. Mol Immunol. 2017;90:126–135. doi:10.1016/j.molimm.2017.06.034
62. Carvalho D, Zanetti BR, Miranda L, et al. High IL-17 expression is associated with an unfavorable prognosis in thyroid cancer. Oncol Lett. 2017;13(3):1925–1931. doi:10.3892/ol.2017.5638
63. Lee MH, Tung-Chieh CJ, Liao CT, et al. Interleukin 17 and peripheral IL-17-expressing T cells are negatively correlated with the overall survival of head and neck cancer patients. Oncotarget. 2018;9(11):9825–9837. doi:10.18632/oncotarget.23934
64. Dvorak HF. Vascular permeability factor/vascular endothelial growth factor: a critical cytokine in tumor angiogenesis and a potential target for diagnosis and therapy. J Clin Oncol. 2002;20(21):4368–4380. doi:10.1200/JCO.2002.10.088
65. Ferrara N, Gerber HP, LeCouter J. The biology of VEGF and its receptors. Nat Med. 2003;9(6):669–676. doi:10.1038/nm0603-669
66. Christopoulos A, Ahn SM, Klein JD, Kim S. Biology of vascular endothelial growth factor and its receptors in head and neck cancer: beyond angiogenesis. Head Neck. 2011;33(8):1220–1229. doi:10.1002/hed.21588
67. Dunst J, Stadler P, Becker A, et al. Tumor hypoxia and systemic levels of vascular endothelial growth factor (VEGF) in head and neck cancers. Strahlenther Onkol. 2001;177(9):469–473. doi:10.1007/PL00002428
68. Maxwell PH, Wiesener MS, Chang GW, et al. The tumour suppressor protein VHL targets hypoxia-inducible factors for oxygen-dependent proteolysis. Nature. 1999;399(6733):271–275. doi:10.1038/20459
69. Zachary I, Gliki G. Signaling transduction mechanisms mediating biological actions of the vascular endothelial growth factor family. Cardiovasc Res. 2001;49(3):568–581. doi:10.1016/S0008-6363(00)00268-6
70. Wu A, Luo N, Xu Y, et al. Exosomal LBH inhibits epithelial-mesenchymal transition and angiogenesis in nasopharyngeal carcinoma via downregulating VEGFA signaling. Int J Biol Sci. 2022;18(1):242–260. doi:10.7150/ijbs.66506
71. Song YS, Kim MJ, Sun HJ, et al. Aberrant thyroid-stimulating hormone receptor signaling increases VEGF-A and CXCL8 secretion of thyroid cancer cells, contributing to angiogenesis and tumor growth. Clin Cancer Res. 2019;25(1):414–425. doi:10.1158/1078-0432.CCR-18-0663
72. Sun Q, Zhang T, Xiao Q, Mei B, Zhang X. Procyanidin B2 inhibits angiogenesis and cell growth in oral squamous cell carcinoma cells through the vascular endothelial growth factor (VEGF)/VEGF receptor 2 (VEGFR2) pathway. Bioengineered. 2022;13(3):6500–6508. doi:10.1080/21655979.2022.2033013
73. Yasuoka H, Nakamura Y, Zuo H, et al. VEGF-D expression and lymph vessels play an important role for lymph node metastasis in papillary thyroid carcinoma. Mod Pathol. 2005;18(8):1127–1133. doi:10.1038/modpathol.3800402
74. Li DK, Chen XR, Wang LN, et al. Epstein-Barr virus induces lymphangiogenesis and lympth node metastasis via upregulation of VEGF-C in nasopharyngeal carcinoma. Mol Cancer Res. 2022;20(1):161–175. doi:10.1158/1541-7786.MCR-21-0164
75. Ceric S, Ceric T, Pojskic N, et al. Immunohistochemical expression and prognostic significance of VEGF-C in well-differentiated thyroid cancer. Acta Endocrinol (Buchar). 2020;16(4):409–416. doi:10.4183/aeb.2020.409
76. Huang Q, Ji M, Li F, et al. Diagnostic and prognostic value of plasma cell-free DNA combined with VEGF-C in laryngeal squamous cell carcinoma. Mol Cell Probes. 2023;67:101895. doi:10.1016/j.mcp.2023.101895
77. Suto T, Karonitsch T. The immunobiology of mTOR in autoimmunity. J Autoimmun. 2020;110:102373. doi:10.1016/j.jaut.2019.102373
78. Kaldirim M, Lang A, Pfeiler S, et al. Modulation of mTOR signaling in cardiovascular disease to target acute and chronic inflammation. Front Cardiovasc Med. 2022;(9):907348. doi:10.3389/fcvm.2022.907348
79. Ma J, Han LZ, Liang H, et al. Celastrol inhibits the HIF-1alpha pathway by inhibition of mTOR/p70S6K/eIF4E and ERK1/2 phosphorylation in human hepatoma cells. Oncol Rep. 2014;32(1):235–242. doi:10.3892/or.2014.3211
80. Wang Z, Valera JC, Zhao X, Chen Q, Gutkind JS. mTOR co-targeting strategies for head and neck cancer therapy. Cancer Metastasis Rev. 2017;36(3):491–502. doi:10.1007/s10555-017-9688-7
81. Zheng A, Li F, Chen F, et al. PD-L1 promotes head and neck squamous cell carcinoma cell growth through mTOR signaling. Oncol Rep. 2019;41(5):2833–2843. doi:10.3892/or.2019.7053
82. Xie J, Li Q, Ding X, Gao Y. Targeting mTOR by CZ415 inhibits head and neck squamous cell carcinoma cells. Cell Physiol Biochem. 2018;46(2):676–686. doi:10.1159/000488724
83. Yang Y, Chen D, Liu H, Yang K. Increased expression of lncRNA CASC9 promotes tumor progression by suppressing autophagy-mediated cell apoptosis via the AKT/mTOR pathway in oral squamous cell carcinoma. Cell Death Dis. 2019;10(2):41. doi:10.1038/s41419-018-1280-8
84. Gao H, Wang W, Li Q. GANT61 suppresses cell survival, invasion and epithelial-mesenchymal transition through inactivating AKT/mTOR and JAK/STAT3 pathways in anaplastic thyroid carcinoma. Cancer Biol Ther. 2022;23(1):369–377. doi:10.1080/15384047.2022.2051158
85. Lv J, Liu C, Chen FK, et al. M2-like tumour-associated macrophage-secreted IGF promotes thyroid cancer stemness and metastasis by activating the PI3K/AKT/mTOR pathway. Mol Med Rep. 2021;24(2). doi:10.3892/mmr.2021.12249
86. Huang ML, Qi CL, Zou Y, et al. Plac8-mediated autophagy regulates nasopharyngeal carcinoma cell function via AKT/mTOR pathway. J Cell Mol Med. 2020;24(14):7778–7788. doi:10.1111/jcmm.15409
87. Yu JH, Chen L, Yu JY, Luo HQ, Wang L. PI3K-PKB-mTOR hyperactivation in relation to nasopharyngeal carcinoma progression and prognosis. J Cell Biochem. 2019;120(6):10186–10194.
88. Ni Y, Low JT, Silke J, O’Reilly LA. Digesting the role of JAK-STAT and cytokine signaling in oral and gastric cancers. Front Immunol. 2022;13:835997. doi:10.3389/fimmu.2022.835997
89. Mukthavaram R, Ouyang X, Saklecha R, et al. Effect of the JAK2/STAT3 inhibitor SAR317461 on human glioblastoma tumorspheres. J Transl Med. 2015;13:269. doi:10.1186/s12967-015-0627-5
90. Silva CM. Role of STATs as downstream signal transducers in Src family kinase- mediated tumorigenesis. Oncogene. 2004;23(48):8017–8023. doi:10.1038/sj.onc.1208159
91. Jarnicki A, Putoczki T, Ernst M. Stat3: linking inflammation to epithelial cancer – more than a “gut” feeling? Cell Div. 2010;5:14. doi:10.1186/1747-1028-5-14
92. Siveen KS, Sikka S, Surana R, et al. Targeting the STAT3 signaling pathway in cancer: role of synthetic and natural inhibitors. Biochim Biophys Acta. 2014;1845(2):136–154. doi:10.1016/j.bbcan.2013.12.005
93. Gaemers IC, Vos HL, Volders HH, van der Valk SW, Hilkens J. A stat-responsive element in the promoter of the episialin/MUC1 gene is involved in its overexpression in carcinoma cells. J Biol Chem. 2001;276(9):6191–6199. doi:10.1074/jbc.M009449200
94. Suiqing C, Min Z, Lirong C. Overexpression of phosphorylated-STAT3 correlated with the invasion and metastasis of cutaneous squamous cell carcinoma. J Dermatol. 2005;32(5):354–360. doi:10.1111/j.1346-8138.2005.tb00906.x
95. Lai SY, Johnson FM. Defining the role of the JAK-STAT pathway in head and neck and thoracic malignancies: implications for future therapeutic approaches. Drug Resist Updat. 2010;13(3):67–78. doi:10.1016/j.drup.2010.04.001
96. Li H, Liu J, Fan N, et al. Nav1.6 promotes the progression of human follicular thyroid carcinoma cells via JAK-STAT signaling pathway. Pathol Res Pract. 2022;236:153984. doi:10.1016/j.prp.2022.153984
97. Qing X, Tan GL, Liu HW, et al. LINC00669 insulates the JAK/STAT suppressor SOCS1 to promote nasopharyngeal cancer cell proliferation and invasion. J Exp Clin Cancer Res. 2020;39(1):166. doi:10.1186/s13046-020-01674-z
98. Ling J, Zhang L, Chang A, et al. Overexpression of KITLG predicts unfavorable clinical outcomes and promotes lymph node metastasis via the JAK/STAT pathway in nasopharyngeal carcinoma. Lab Invest. 2022;102(11):1257–1267. doi:10.1038/s41374-022-00817-4
99. Chang WM, Chang YC, Yang YC, et al. AKR1C1 controls cisplatin-resistance in head and neck squamous cell carcinoma through cross-talk with the STAT1/3 signaling pathway. J Exp Clin Cancer Res. 2019;38(1):245. doi:10.1186/s13046-019-1256-2
100. Limberg J, Egan CE, Gray KD, et al. Activation of the JAK/STAT pathway leads to BRAF inhibitor resistance in BRAFV600E positive thyroid carcinoma. Mol Cancer Res. 2023;21(5):397–410. doi:10.1158/1541-7786.MCR-21-0832
101. Geiger JL, Grandis JR, Bauman JE. The STAT3 pathway as a therapeutic target in head and neck cancer: barriers and innovations. Oral Oncol. 2016;56:84–92. doi:10.1016/j.oraloncology.2015.11.022
102. Pan Y, Cao S, Tang J, et al. Cyclooxygenase-2 in adipose tissue macrophages limits adipose tissue dysfunction in obese mice. J Clin Invest. 2022;132(9):e152391. doi:10.1172/JCI152391
103. Dean PT, Hooks SB. Pleiotropic effects of the COX-2/PGE2 axis in the glioblastoma tumor microenvironment. Front Oncol. 2023;12:1116014. doi:10.3389/fonc.2022.1116014
104. Zhang Y, Tighe S, Zhu YT. COX-2 signaling in the tumor microenvironment. Adv Exp Med Biol. 2020;1277:87–104.
105. Saito S, Ozawa H, Imanishi Y, et al. Cyclooxygenase-2 expression is associated with chemoresistance through cancer stemness property in hypopharyngeal carcinoma. Oncol Lett. 2021;22(1):533. doi:10.3892/ol.2021.12794
106. Zhu Y, Shi C, Zeng L, et al. High COX-2 expression in cancer-associated fibiroblasts contributes to poor survival and promotes migration and invasiveness in nasopharyngeal carcinoma. Mol Carcinog. 2020;59(3):265–280. doi:10.1002/mc.23150
107. Chiang KH, Shieh JM, Shen CJ, et al. Epidermal growth factor-induced COX-2 regulates metastasis of head and neck squamous cell carcinoma through upregulation of angiopoietin- like 4. Cancer Sci. 2020;111(6):2004–2015. doi:10.1111/cas.14400
108. Wu QB, Sun GP. Expression of COX-2 and HER-2 in colorectal cancer and their correlation. World J Gastroenterol. 2015;21(20):6206–6214. doi:10.3748/wjg.v21.i20.6206
109. Nasry W, Rodriguez-Lecompte JC, Martin CK. Role of COX-2/PGE2 mediated inflammation in oral squamous cell carcinoma. Cancers (Basel). 2018;10(10):348. doi:10.3390/cancers10100348
110. Mendes RA, Carvalho JF, Waal I. An overview on the expression of cyclooxygenase-2 in tumors of the head and neck. Oral Oncol. 2009;45(10):e124–8. doi:10.1016/j.oraloncology.2009.03.016
111. Yang G, Deng Q, Fan W, et al. Cyclooxygenase-2 expression is positively associated with lymph node metastasis in nasopharyngeal carcinoma. PLoS One. 2017;12(3):e0173641. doi:10.1371/journal.pone.0173641
112. Cheng C, Yang J, Li SW, et al. HDAC4 promotes nasopharyngeal carcinoma progression and serves as a therapeutic target. Cell Death Dis. 2021;12(2):137. doi:10.1038/s41419-021-03417-0
113. Shui L, Li S, Wang F, et al. Relationship between cyclooxygenase-2 (COX-2) content and prognosis in nasopharyngeal carcinoma before and after radiochemotherapy. J BUON. 2020;25(5):2395–2404.
114. Li YJ, Luo Y, Xie XQ, Li P, Wang F. The prognostic value of COX-2 expression on circulating tumor cells in nasopharyngeal carcinoma: a prospective analysis. Radiother Oncol. 2018;129(2):396–402. doi:10.1016/j.radonc.2018.07.022
115. Wynn TA, Chawla A, Pollard JW. Macrophage biology in development, homeostasis and disease. Nature. 2013;496(7446):445–455. doi:10.1038/nature12034
116. Biswas SK, Mantovani A. Macrophage plasticity and interaction with lymphocyte subsets: cancer as a paradigm. Nat Immunol. 2010;11(10):889–896. doi:10.1038/ni.1937
117. Bao Z, Zeng W, Zhang D, et al. SNAIL induces EMT and lung metastasis of tumours secreting CXCL2 to promote the invasion of M2-type immunosuppressed macrophages in colorectal cancer. Int J Biol Sci. 2022;18(7):2867–2881. doi:10.7150/ijbs.66854
118. Richards DM, Hettinger J, Feuerer M. Monocytes and macrophages in cancer: development and functions. Cancer Microenviron. 2013;6(2):179–191. doi:10.1007/s12307-012-0123-x
119. Noy R, Pollard JW. Tumor-associated macrophages: from mechanisms to therapy. Immunity. 2014;41(1):49–61. doi:10.1016/j.immuni.2014.06.010
120. Vijver SV, Singh A, Mommers-Elshof ETAM, et al. Collagen fragments produced in cancer mediate T cell suppression through leukocyte-associated immunoglobulin-like receptor 1. Front Immunol. 2021;12:733561. doi:10.3389/fimmu.2021.733561
121. Li Y, Hodge J, Liu Q, et al. TFEB is a master regulator of tumor-associated macrophages in breast cancer. J Immunother Cancer. 2020;8(1):e000543. doi:10.1136/jitc-2020-000543
122. Pirila E, Vayrynen O, Sundquist E, et al. Macrophages modulate migration and invasion of human tongue squamous cell carcinoma. PLoS One. 2015;10(3):e0120895. doi:10.1371/journal.pone.0120895
123. Sangaletti S, Di Carlo E, Gariboldi S, et al. Macrophage-derived SPARC bridges tumor cell-extracellular matrix interactions toward metastasis. Cancer Res. 2008;68(21):9050–9059. doi:10.1158/0008-5472.CAN-08-1327
124. Bonde AK, Tischler V, Kumar S, Soltermann A, Schwendener RA. Intratumoral macrophages contribute to epithelial-mesenchymal transition in solid tumors. BMC Cancer. 2012;12:35. doi:10.1186/1471-2407-12-35
125. Kubota K, Moriyama M, Furukawa S, et al. CD163(+)CD204(+) tumor-associated macrophages contribute to T cell regulation via interleukin-10 and PD-L1 production in oral squamous cell carcinoma. Sci Rep. 2017;7(1):1755. doi:10.1038/s41598-017-01661-z
126. Solito S, Marigo I, Pinton L, et al. Myeloid-derived suppressor cell heterogeneity in human cancers. Ann N Y Acad Sci. 2014;1319:47–65. doi:10.1111/nyas.12469
127. Gabrilovich DI, Nagaraj S. Myeloid-derived suppressor cells as regulators of the immune system. Nat Rev Immunol. 2009;9(3):162–174. doi:10.1038/nri2506
128. Kujawski M, Kortylewski M, Lee H, et al. Stat3 mediates myeloid cell-dependent tumor angiogenesis in mice. J Clin Invest. 2008;118(10):3367–3377. doi:10.1172/JCI35213
129. Murdoch C, Muthana M, Coffelt SB, Lewis CE. The role of myeloid cells in the promotion of tumour angiogenesis. Nat Rev Cancer. 2008;8(8):618–631. doi:10.1038/nrc2444
130. Pang X, Fan HY, Tang YL, et al. Myeloid derived suppressor cells contribute to the malignant progression of oral squamous cell carcinoma. PLoS One. 2020;15(2):e0229089. doi:10.1371/journal.pone.0229089
131. Fugle CW, Zhang Y, Hong F, et al. CD24 blunts oral squamous cancer development and dampens the functional expansion of myeloid-derived suppressor cells. Oncoimmunology. 2016;5(10):e1226719. doi:10.1080/2162402X.2016.1226719
132. Angell TE, Lechner MG, Smith AM, et al. Circulating myeloid-derived suppressor cells predict differentiated thyroid cancer diagnosis and extent. Thyroid. 2016;26(3):381–389. doi:10.1089/thy.2015.0289
133. Davis RJ, Van Waes C, Allen CT. Overcoming barriers to effective immunotherapy: mDSCs, TAMs, and Tregs as mediators of the immunosuppressive microenvironment in head and neck cancer. Oral Oncol. 2016;58:59–70. doi:10.1016/j.oraloncology.2016.05.002
134. Paijens ST, Vledder A, de Bruyn M, Nijman HW. Tumor-infiltrating lymphocytes in the immunotherapy era. Cell Mol Immunol. 2021;18(4):842–859. doi:10.1038/s41423-020-00565-9
135. Hibino S, Kawazoe T, Kasahara H, et al. Inflammation-induced tumorigenesis and metastasis. Int J Mol Sci. 2021;22(11):5421. doi:10.3390/ijms22115421
136. Kryczek I, Lin Y, Nagarsheth N, et al. IL-22(+)CD4(+) T cells promote colorectal cancer stemness via STAT3 transcription factor activation and induction of the methyltransferase DOT1L. Immunity. 2014;40(5):772–784. doi:10.1016/j.immuni.2014.03.010
137. Lei Y, Xie Y, Tan YS, et al. Telltale tumor infiltrating lymphocytes (TIL) in oral, head & neck cancer. Oral Oncol. 2016;61:159–165. doi:10.1016/j.oraloncology.2016.08.003
138. Zhou Z, Mu D, Zhang D, et al. PD-L1 in combination with CD8(+)TIL and HIF-1alpha are promising prognosis predictors of head and neck squamous cell carcinoma. Cancer Manag Res. 2020;12:13233–13239. doi:10.2147/CMAR.S285691
139. Kawaguchi T, Ono T, Sato F, et al. CD8+ T cell infiltration predicts chemoradiosensitivity in nasopharyngeal or oropharyngeal cancer. Laryngoscope. 2021;131(4):E1179–E1189. doi:10.1002/lary.29097
140. Cho BC, Daste A, Ravaud A, et al. Bintrafusp alfa, a bifunctional fusion protein targeting TGF-beta and PD-L1, in advanced squamous cell carcinoma of the head and neck: results from a Phase I cohort. J Immunother Cancer. 2020;8(2):e000664. doi:10.1136/jitc-2020-000664
141. Jimeno A, Posner MR, Wirth LJ, et al. A phase 2 study of dalantercept, an activin receptor-like kinase-1 ligand trap, in patients with recurrent or metastatic squamous cell carcinoma of the head and neck. Cancer. 2016;122(23):3641–3649. doi:10.1002/cncr.30317
142. Argiris A, Kotsakis AP, Hoang T, et al. Cetuximab and bevacizumab: preclinical data and Phase II trial in recurrent or metastatic squamous cell carcinoma of the head and neck. Ann Oncol. 2013;24(1):220–225. doi:10.1093/annonc/mds245
143. Dunn LA, Fury MG, Xiao H, et al. A phase II study of temsirolimus added to low-dose weekly carboplatin and paclitaxel for patients with recurrent and/or metastatic (R/M) head and neck squamous cell carcinoma (HNSCC). Ann Oncol. 2017;28(10):2533–2538. doi:10.1093/annonc/mdx346
144. Weed DT, Vella JL, Reis IM, et al. Tadalafil reduces myeloid-derived suppressor cells and regulatory T cells and promotes tumor immunity in patients with head and neck squamous cell carcinoma. Clin Cancer Res. 2015;21(1):39–48 879. doi:10.1158/1078-0432.CCR-14-1711
 © 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.