Back to Journals » ImmunoTargets and Therapy » Volume 5
The role of adipokines in chronic inflammation
Authors Mancuso P
Received 24 August 2015
Accepted for publication 12 February 2016
Published 23 May 2016 Volume 2016:5 Pages 47—56
DOI https://doi.org/10.2147/ITT.S73223
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Michael Shurin
Peter Mancuso
Department of Nutritional Sciences, School of Public Health, University of Michigan, Ann Arbor, MI, USA
Abstract: Adipose tissue has traditionally been defined as connective tissue that stores excess calories in the form of triacylglycerol. However, the physiologic functions attributed to adipose tissue are expanding, and it is now well established that adipose tissue is an endocrine gland. Among the endocrine factors elaborated by adipose tissue are the adipokines; hormones, similar in structure to cytokines, produced by adipose tissue in response to changes in adipocyte triacylglycerol storage and local and systemic inflammation. They inform the host regarding long-term energy storage and have a profound influence on reproductive function, blood pressure regulation, energy homeostasis, the immune response, and many other physiologic processes. The adipokines possess pro- and anti-inflammatory properties and play a critical role in integrating systemic metabolism with immune function. In calorie restriction and starvation, proinflammatory adipokines decline and anti-inflammatory adipokines increase, which informs the host of energy deficits and contributes to the suppression of immune function. In individuals with normal metabolic status, there is a balance of pro- and anti-inflammatory adipokines. This balance shifts to favor proinflammatory mediators as adipose tissue expands during the development of obesity. As a consequence, the proinflammatory status of adipose tissue contributes to a chronic low-grade state of inflammation and metabolic disorders associated with obesity. These disturbances are associated with an increased risk of metabolic disease, type 2 diabetes, cardiovascular disease, and many other pathological conditions. This review focuses on the impact of energy homeostasis on the adipokines in immune function.
Keywords: calorie restriction, obesity, adipose tissue, type 2 diabetes, macrophage, infection, chronic low-grade inflammation
Introduction
It is now well recognized that adipose serves as a depot for excess energy storage and as an endocrine gland that produces several biological mediators known to regulate blood pressure, reproductive function, appetite, glucose homeostasis, angiogenesis, and immune function.1 Adipose tissue produces both pro- and anti-inflammatory mediators that influence local and systemic inflammation. Among these mediators are the adipokines, proteins produced by cells within white adipose tissue that function as hormones.2 As a family of mediators, the adipokines consist of true adipokines that are predominantly produced by pre- and mature adipocytes and classical cytokines that are produced by adipocytes as well as immune cells found in the stromal vascular fraction (SVF) of adipose tissue and many other cell types outside adipose tissue depots. The balance of pro- and anti-inflammatory adipokines is dictated by many different factors, including the nutritional/metabolic status of the host, the presence of infection or systemic inflammation, oxidative stress, smoking status, age, and sex.3–9 Most importantly, adipokines play a major role in the regulation of the inflammatory response in adipose tissue during the development of obesity and in response to infection or systemic inflammation. This review focuses on the ability of adipokines to regulate the inflammatory response in the setting of chronic calorie restriction and obesity.10–13
Cellular composition of adipose tissue
Adipose tissue is composed of mature adipocytes, preadipocytes, mesenchymal cells, and cells within the SVF that include vascular endothelial and smooth muscle cells, fibroblasts, and several different leukocyte subsets (Figure 1). Interestingly, nearly all immune cells, such as resident macrophages, mast cells, monocytes, dendritic cells, natural killer cells, B-cells, T-cells, neutrophils, and eosinophils, have been found in adipose tissue.14–18 These cells play a critical role in adipose tissue remodeling and repair in lean mice and humans. Although their function in calorie restriction is poorly understood, immune cell populations in general decline during calorie restriction and increase in obesity. Resident and recruited macrophages are the most abundant type of immune cells in adipose tissue. These cells have been characterized as having M1 (classically activated) or M2 (alternatively activated) phenotypes. M1 macrophages appear to be primed for host defense against infection, while M2 macrophages are thought to play an important role in tissue remodeling and repair. Recent evidence suggests that this dichotomous classification may be an oversimplification, since macrophages may exhibit different phenotypes that span a spectrum of activation states.19,20 They also play a critical role in orchestrating the inflammatory response in obesity and type 2 diabetes (T2D).21
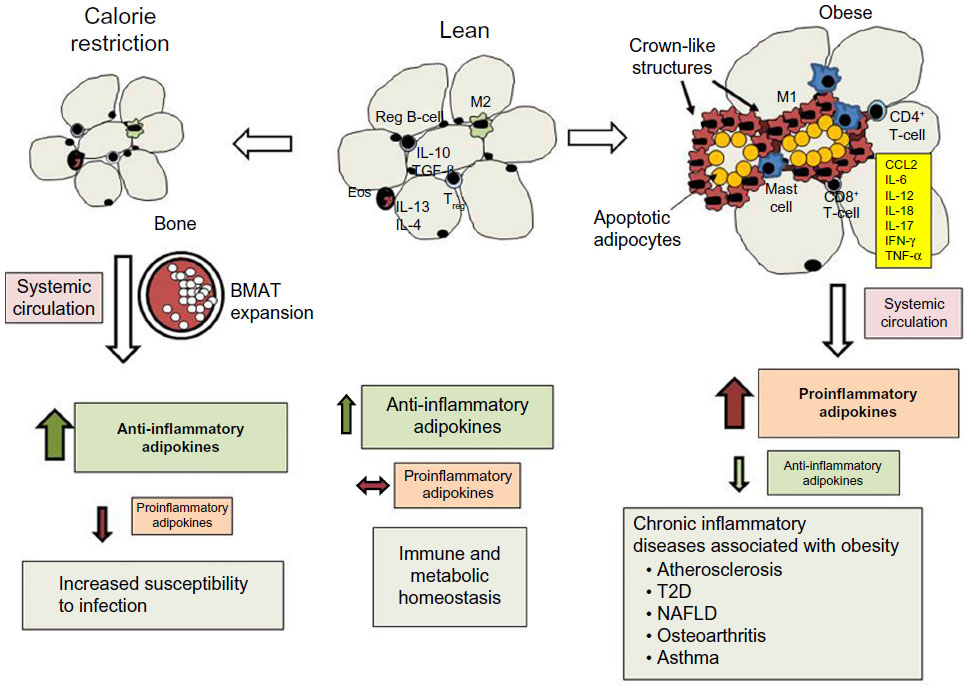 | Figure 1 Effects of calorie restriction and obesity on adipose tissue leukocyte populations, adipokine secretion, and chronic inflammation. |
Mast cells, which are known to mediate acute inflammation in type 1 hypersensitivity responses and host defense against parasitic organisms, are also found in adipose tissue.22 Dendritic cells are professional antigen-presenting cells that recognize foreign antigens and present them to T-cells via major histocompatibility-complex molecules. Adipose tissue dendritic cells have been found in mice and humans and may play an important role in T-helper (TH)-17 cell responses.18,23 The most abundant granulocyte found in blood, the neutrophil, can also be found in adipose tissue. While these cells play a prominent role in host defense against bacterial infections, their function in adipose tissue is not clear. These cells transiently infiltrate murine adipose tissue with the initiation of high-fat-diet feeding and mediate insulin resistance in mice.24,25 Natural killer (NK) cells, best known for their role in the early host response to viral infections and in killing tumor cells, have recently been shown to promote macrophage proliferation and polarization in adipose tissue and insulin resistance in mice fed a high-fat diet.26
B-cells are most commonly studied for their contribution to host defense against infection and autoimmune disease. In adipose tissue, several different B-cell subsets have been identified, and these cells contribute to local and systemic inflammation by secreting cytokines, producing antibodies, and presenting antigens to T-cells.27 Finally, several different T-cell subsets, such as CD4+ TH cells, which include Treg, TH1, and TH2, and CD8+ T-cells, have been identified in adipose tissue, where they regulate local inflammation through the secretion of cytokines that influence the differentiation and polarization of macrophages.28,29 The activation state, differentiation, and proliferation of immune cells in adipose tissue are profoundly influenced by anti- and proinflammatory cytokines, lipid mediators, and adipokines secreted within local fat pads and in circulation.30–34
Anti-inflammatory adipokines
Adiponectin, C1q/TNF-related proteins (CTRPs), omentin, and secreted frizzled-related protein 5 (SFRP5) are anti-inflammatory adipokines produced by adipose tissue.35–38 Adiponectin is the best-known and most abundant adipokine found in human serum, with concentrations typically in the μg/mL range.39 Unlike other adipokines, which are found in greatest quantities in visceral and subcutaneous adipose tissue (SCAT), it is predominantly produced by bone marrow adipose tissue (BMAT).40 Calorie restriction, aging, estrogen deficiency, T1D, and treatment with thiazolidinediones increase, while obesity, T2D, oxidative stress, and cigarette-smoke exposure decrease serum adiponectin levels.41–46 Adiponectin is a complex molecule that forms low-, intermediate-, and high molecular weight complexes in circulation. Its effects are mediated through the AdipoR1 and AdipoR2 receptors, which activate AMPK in immune cells and tissues.47 In particular, the high-molecular-weight complex has anti-inflammatory properties known to inhibit inflammation by blocking NF-κB activation and reducing such cytokines as TNFα, IL-6, and IL-18.2,48–50
Adiponectin-knockout mice exhibit enhanced inflammatory responses, suggesting a major role for adiponectin in suppressing systemic and tissue inflammation.31,51–53 For example, adiponectin attenuates vascular inflammation, which may play a protective role against aortic aneurism.51 The protective effects of adiponectin in the vasculature may be mediated by locally produced perivascular adipose tissue.54 Moreover, the lungs of adiponectin-knockout animals exhibit an emphysematous phenotype, with enlarged air spaces and activated alveolar macrophages capable of producing higher levels of TNFα and MMP12.31 These animals also develop pulmonary arterial hypertension characterized by perivascular inflammation.53 Adiponectin plays a protective role in acute lung injury and myocardial ischemia, where it reduces cellular infiltration.52,55–57 In contrast to these studies showing a protective anti-inflammatory effect, adiponectin may play a proinflammatory role in arthritic joints by promoting COX2 expression and the synthesis of PGE2, which increases inflammation and pain.58
Much less is known regarding other adipokines with anti-inflammatory properties, such as CTRP, omentin, and SFRP5. The CTRPs are structurally similar to adiponectin, and at least 15 isoforms have been described.59 CTRP3 has been shown to reduce cytokine production in human monocytes and adipocytes stimulated with lipopolysaccharide and free fatty acids by inhibiting TLR4 activation.59 In addition, CTRP13 inhibits inflammation in lipid-loaded hepatocytes and improves insulin sensitivity.60 Omentin is a novel adipokine that inhibits TNFα-induced endothelial cell COX2 expression and induces endothelial nitric oxide synthase.61 SFRP5 has anti-inflammatory effects in adipose tissue and in macrophages that are mediated through the suppression of noncanonical Wnt5a/JNK signaling, which ultimately inhibits macrophage TNFα, IL-1β, and CCL2–MCP1 synthesis.62 Clearly, there is a need to increase our understanding of the biology of these novel anti-inflammatory adipokines and their role in opposing the effects of proinflammatory mediators and adipokines.
Proinflammatory adipokines
Leptin is the best known proinflammatory adipokine that increases in proportion to white adipose tissue mass, and was first described as a satiety hormone.11 The long form of the leptin receptor, which is expressed by nearly all immune cells, initiates intracellular signaling to activate the tyrosine kinase JAK2, the latent transcription factor STAT3, MAPK, ERK1/2, and PI3K pathways that activate the innate immune response.63 Leptin can directly enhance the production of several proinflammatory cytokines, such as IL-6, IL-12, IL-18, and TNFα, the chemokines IL-8 and CCL2/MCP-1, and the lipid mediators PGE2, cysteinyl leukotrienes (cysLTs), and leukotriene B4 (LTB4) in peripheral blood monocytes and resident tissue macrophages in mice and humans.8,64–67 Leptin can also induce the production of reactive oxygen intermediates in macrophages, neutrophils, and endothelial cells and potentiate IFNγ-induced expression of nitric oxide synthase.68–70 Leptin enhances platelet aggregation and promotes leukocyte–endothelial cell interactions by increasing the expression of adhesion molecules on myeloid cells and vascular endothelial cells.68,71–73 TH1 and TH17 responses are enhanced with leptin, which can also prevent T-cell apoptosis.74
Resistin and resistin-like molecules were first characterized in mice as mediators of insulin resistance, T2D, and metabolic syndrome.75 While adipose tissue is the primary source of this adipokine in mice, monocytes and macrophages are the most important sources of resistin in humans.76 In addition, there are substantial differences between mouse and human resistin amino acid sequence homology, indicating that the physiological actions of this adipokine may differ in mice and humans.77 To address these controversial issues, Qatanani et al created a humanized mouse that expressed human resistin in macrophages but not adipose tissue that has provided a robust system to substantiate the importance of human resistin in T2D and obesity.76 The proinflammatory effects of resistin are mediated through CAP1, recently identified as a receptor for resistin. It initiates cAMP-mediated PKA activation and NF-κB-related transcription of inflammatory cytokines in human monocytes.78
There are several other less studied proinflammatory adipokines that have been implicated in the promotion of inflammation associated with obesity, including chemerin, retinol binding protein 4 (RBP4), and lipocalin 2 (LCN2).2 Chemerin is an adipocyte-derived chemoattractant for monocytes and dendritic cells that is produced by mature adipocytes. It is secreted as a preprohormone that requires enzymatic cleavage by extracellular proteases.79 RBP4 is a member of the lipocalin family of proteins that transports retinol from the liver to the peripheral tissues.80 RBP4 is produced by the liver, adipose tissue, and macrophages. RBP4 may contribute to inflammation by activating adipose tissue antigen-presenting cells that promote TH1-cell polarization.81 LCN2 is produced by adipocytes and is induced by inflammatory stimuli that activate NF-κB in adipose tissue.82 LCN2, also referred to as neutrophil gelatinase-associated lipocalin, binds and transports hydrophobic molecules, such as retinoids, arachidonic acid, LTB4, platelet-activating factor, and steroids.82,83 LCN2 also plays an important role in host defense against bacterial infections by sequestering iron.84 Through the elaboration of anti- and proinflammatory adipokines that spill over into the systemic circulation, adipose tissue plays a critical role in regulating the inflammatory response in the setting of calorie restriction and obesity.
The impact of adipokines on immune function during calorie restriction
Prolonged calorie restriction reduces the amount of energy in the form of triacylglycerol (TAG) stored in adipose tissue. The size of individual adipocytes within visceral adipose tissue (VAT) declines. If caloric restriction is extended, SCAT declines as well, but this response requires more time.85 In contrast, BMAT expands during calorie restriction, replacing active hematopoietic cells.40 Calorie restriction also has a profound impact on the adipokines and mediators of inflammation produced within adipose tissue.
The production of anti-inflammatory adipokines, such as adiponectin, CTRP, omentin, and SFRP5, increases.40,86–88 Unlike other adipokines, more adiponectin is produced by BMAT than by SCAT or VAT.40 Adiponectin plays an important role in adipogenesis and insulin sensitivity via AMPK signaling, which promotes fatty acid oxidation and glucose uptake in skeletal muscle and liver.89,90 The increase in omentin that accompanies calorie restriction may be protective against cardiovascular disease.37 Omentin may reduce vascular inflammation associated with atherosclerosis, since it reduces macrophage adhesion to endothelial cells in vitro by inhibiting ICAM1 and VCAM1 expression.37 SFRP5 may also play a protective role in cardiovascular disease.62 The increased secretion of SFRP5 following calorie restriction in obese patients may contribute to improvements in atherosclerosis and protect against myocardial ischemia–reperfusion injury.62 IL-10 has also been shown to increase during calorie restriction. It is produced by regulatory B-cells within adipose tissue, where it suppresses inflammatory cytokine production by resident CD8+ T-cells.91 Tregs elaborate TGFβ and IL-10, which helps maintain an anti-inflammatory environment.28 In contrast, the production of proinflammatory adipokines (leptin, resistin, RBP4), cytokines (IL-1β, IL-6. IL-17, TNFα), chemokines (CCL2 and MIP2), and lipid mediators (cysLTs and LTB4) decline.3,10,33,76,92,93 While calorie restriction has been shown to suppress inflammation, protect against cardiovascular disease, improve glucose homeostasis in T2D, and increase the life span of mice, it also has negative consequences on host defense against infection.3,94–96
Studies by Gardner et al evaluated the impact of a calorie-restricted diet on host defense against a mouse adaptive influenza virus, PR8, in young and aged mice.94,95 In these studies, young and aged mice that consumed 40% fewer calories than their ad libitum-fed counterparts exhibited greater weight loss, mortality, lung viral titers, and impaired NK-cell cytotoxic function following influenza virus infection.94,95 Reduced IFNα and IFNβ production was also associated with increased viral burdens of young calorie-restricted mice.95 In a similar study, refeeding calorie-restricted mice improved survival and nearly restored NK-cell numbers and cytotoxic function at all time points following influenza infection. While refeeding calorie-restricted mice reconstituted adipose tissue, it did not restore leptin to levels observed in ad libitum-fed animals, and this may have accounted for the differences in NK-cell numbers and function.96
Acute calorie restriction, a common occurrence in critically ill patients and starvation, rapidly mobilizes energy stores from adipose tissue, shrinking the size of VAT and SCAT adipose tissue. In general, both anti- and proinflammatory adipokines decline with prolonged fasting. For example, serum leptin levels decline rapidly and are disproportionately lower than would be expected for a given fat mass in humans and mice.4,97 Unlike leptin, serum adiponectin levels remain stable after 3 days of fasting and decline slightly during a prolonged fast.98,99 The decline of leptin in adaptation to starvation has been examined in murine models to determine the impact of leptin in thymic atrophy and pneumococcal pneumonia.3,10,92,100 As leptin levels decline during fasting, glucocorticoid levels rise, and this contributes to peripheral blood T- and B-lymphocyte apoptosis and diminished thymic and bone marrow cellularity. These events are prevented if leptin levels are maintained during fasting.92,101
Fasting and chronic energy malnutrition are known to suppress the immune response to infection, and leptin levels may predict survival in severe acute childhood malnutrition.102 In mice infected with Streptococcus pneumoniae, 48 hours of fasting reduced pulmonary bacterial clearance, and this was associated with reduced neutrophil counts in peripheral blood and bronchoalveolar lavage. Lung-homogenate cytokine (IL-6), chemokine (MIP2), and LTB4 synthesis in alveolar macrophages were also reduced by fasting. Alveolar macrophages obtained from mice after fasting exhibited defective phagocytosis and killing of S. pneumoniae. Interestingly, all of these responses were restored when exogenous leptin was administered during fasting.3 Using a similar model of leptin depletion by starvation and lipopolysaccharide-induced sepsis, Faggioni et al demonstrated that leptin improved survival, and this improvement was associated with lower levels of systemic TNFα.100 These studies demonstrate the physiologic importance of the adipokines generated in calorie restriction and starvation, which play a crucial role in the host response to infection and sepsis. Since infectious disease remains a leading cause of morbidity and mortality in undernourished patients and in severely malnourished children in low-income nations, more research is needed to understand the role of adipokines and immunosuppression associated with energy malnutrition.103
Hyperplasia and hypertrophy of adipocytes in VAT and SCAT during obesity
In contrast to calorie restriction and starvation, obesity is characterized by energy excess and the expansion of white-adipose tissue that contributes to a chronic state of low-grade inflammation and increased risk of chronic disease (Figure 1). Adipose tissue expansion occurs as excess energy is stored in the form of TAG. Under conditions of normal metabolic homeostasis, the expansion of adipose tissue can buffer excess dietary lipids.85 As excess energy accumulates, the adipose tissue must expand to store excess TAG through the process of adipocyte hyperplasia and hypertrophy.104,105 Hyperplasia occurs through the differentiation of preadipocytes into mature adipocytes, and this process occurs in all adipose-tissue depots, but is more prominent in SCAT.106 The enlargement of SCAT around the femoral gluteal area, also known as the gynoid pattern of obesity, is well suited for lipid buffering, and this helps maintain homeostasis during fluctuation of dietary lipids.106,107 With regard to the role of adipokines in this process, adiponectin promotes adipogenesis and hyperplasia through the activation of PPARγ. SFRP5 expression increases during the expansion of adipose tissue, where it seems to play an important role in suppressing oxidative metabolism and adipocyte growth during obesity.108 The expansion of SCAT via adipocyte hyperplasia is associated with a lower risk of metabolic disease, higher levels of adiponectin, and lower levels of proinflammatory adipokines.109–111 In contrast, the expansion of adipose tissue through the process of hypertrophy is associated with adipose tissue inflammation.2 While adipocyte hypertrophy occurs in all adipose tissue depots, its appearance in VAT is highly correlated with the accumulation of immune cells, proinflammatory adipokines, and metabolic dysfunction.2,112
Changes in immune cell composition and phenotype in adipose tissue during obesity
As adipose tissue expands, the number of eosinophils and levels of IL-13 and IL-4 decline in mice and humans.113 The population of T-cells changes as well, with a decline Treg cells and increases in CD4+ and CD8+ T-cells.28,29,114 In addition to a decline in anti-inflammatory cytokine and adipokine levels, the number of CD4+ T-cells that secrete the TH1 cytokine IFNγ and CD8+ T-cells increases, promoting the differentiation of resident macrophages into the classically activated or M1-macrophage phenotype.21,29 A substantial increase in adipose tissue macrophages occurs as a consequence of the recruitment of peripheral blood monocytes that respond to chemokines (CCL2, CXCL5).14 Neutrophils are also recruited to adipose tissue following dietary high-fat feeding in mice, and the chemokines for their recruitment have not been identified.115 Finally, mast cells also increase in the SVF of VAT in obese mice and human subjects, and these cells contribute to the proinflammatory state by releasing TNFα, IL-6, and lipid mediators, such as cysLTs.16,116
Upon further expansion, the proinflammatory cytokines IL-6, TNFα, and IL-18 and adipokines leptin and resistin promote a proinflammatory environment and contribute to metabolic dysfunction.11,117–121 The expansion of individual adipocytes is limited by the extracellular matrix and hypoxia resulting from rarefaction.122 As a consequence, these cells undergo apoptosis and eventually necrosis.104 Crown-like structures are frequently found in adipose tissue sections from obese mice and humans, and these are necrotic adipocytes surrounded by M1 macrophages.13,104 There are distinct differences in the type and number of inflammatory cells within adipose tissue depots in VAT and SCAT of lean and obese human subjects, and these differences are correlated with metabolic disease risk.123 Compared with VAT, there are fewer macrophages, mast cells, and other immune cells in SCAT, suggesting less adipose tissue inflammation in SCAT compared with VAT.16 Crown-like structures in obese mice are also more prevalent in VAT versus SCAT.16 In total, the location and quality of excess adipose tissue may profoundly influence the inflammatory state of obese individuals.
Adipose tissue inflammation
Inflammation most often occurs in response to infection, irritation, allergic and autoimmune responses, or tissue trauma and is classically characterized by the cardinal signs of inflammation, which include heat, redness, swelling, pain, and loss of function.124 These responses develop as a consequence of the production of proinflammatory mediators, such as lipid mediators (prostaglandins and leukotrienes), cytokines, chemokines, and cellular debris resulting from the release of intracellular constituents or “danger” signals.124 These mediators can induce vasodilation, increase microvascular permeability, activate local immune cells, increase the production of leukocytes from the bone marrow, recruit peripheral blood leukocytes to the site of inflammation, and trigger nociceptors to induce the sensation of pain. Unlike the classic inflammatory response that is activated by pattern-recognition receptors during an infection, the inflammatory response in obesity is initiated by intrinsic signals, such as nutrient sensing, the unfolded protein response, and endoplasmic reticulum stress, and is often referred to as meta-inflammation.125 These intrinsic signals promote an inflammatory response through the activation of the NF-κB-signaling pathway, the production of proinflammatory cytokines, generation of reactivate oxygen species, and proinflammatory adipokine synthesis by adipocytes and immune cells within adipose tissue.125 Conversely, anti-inflammatory adipokine synthesis, eg, adiponectin, declines as a consequence of the unfolded protein response and endoplasmic reticulum stress.126 Under these circumstances, resident and recruited leukocytes within adipose tissue elaborate proinflammatory mediators that not only contribute to local inflammation but spill over into the systemic circulation, causing a chronic state of low-grade inflammation.
Contribution of adipokines to inflammation associated with obesity
Proinflammatory adipokines, which increase in obese individuals, contribute to systemic inflammation and diseases associated with obesity. For example, leptin levels are correlated with the severity of illness in several diseases, such as osteoarthritis, multiple sclerosis, nonalcoholic fatty liver disease, hepatic fibrosis, renal disease, atherosclerosis, and thrombosis.34,127–135 Resistin levels are elevated in obese humans and are associated with a greater risk of cardiovascular disease.106 Resistin may promote chronic inflammation and insulin resistance by enhancing monocyte recruitment through the induction of CCL2 in inflamed VAT in humans and atherosclerosis by promoting monocyte foam cell formation and endothelial cell interactions with the vascular endothelium.136,137 Likewise, chemerin inhibits the maturation of preadipocytes and alters the metabolic functions of mature adipocyte cell lines in vitro.138 While chemerin levels in human subjects are correlated with obesity and metabolic syndrome, a causal role for chemerin in adipose tissue inflammation and obesity-associated disease has not been reported.139 In addition, RBP4 is produced by the liver, adipose tissue, and macrophages, and its expression increases with increasing body mass index, waist circumference, and visceral adiposity in human subjects.140,141 It inhibits intracellular signaling events induced by insulin, such as phosphorylation of IRS1, and contributes to insulin resistance in T2D.142,143 RBP4 may contribute to inflammation by activating adipose tissue antigen-presenting cells that promote TH1-cell polarization.81 Finally, LCN2 may play a critical role in cardiovascular disease associated with obesity, since it is markedly elevated in atherosclerotic plaques.144 However, a causal role for LCN2 in vascular tissue remodeling in atherosclerotic lesions associated with obesity has not been established, and additional research is needed to understand the role of LCN2 in diseases associated with obesity.
Conclusion
Adipose tissue produces several endocrine factors, cytokines, and chemokines that regulate physiologic processes and immune function. Among these mediators are the adipokines, hormones produced by adipocytes, and cells within the SVF of adipose tissue. There are pro- and anti-adipokines that provide a means of communication between energy stored in adipose tissue and several organ systems to integrate metabolism with several physiologic functions. As adipose tissue shrinks during calorie restriction, anti-inflammatory adipokines rise and proinflammatory adipokines decline, resulting in increased insulin sensitivity and suppressed immune function. As adipose tissue expands during obesity, there is an increase in proinflammatory and a reduction in anti-inflammatory adipokines, which contributes to local and systemic inflammation and disturbances in glucose homeostasis. Future therapeutic interventions may target adipokines and their intracellular signaling cascades to enhance immune function in calorie restriction or ameliorate chronic inflammation and T2D in obese patients.
Disclosure
The author reports no conflicts of interest in this work.
References
Scherer PE. Adipose tissue: from lipid storage compartment to endocrine organ. Diabetes. 2006;55(6):1537–1545. | |
Ouchi N, Parker JL, Lugus JJ, Walsh K. Adipokines in inflammation and metabolic disease. Nat Rev Immunol. 2011;11(2):85–97. | |
Mancuso P, Huffnagle GB, Olszewski MA, Phipps J, Peters-Golden M. Leptin corrects host defense defects following acute starvation in murine pneumococcal pneumonia. Am J Respir Crit Care Med. 2006; 173(2):212–218. | |
Ahima RS, Prabakaran D, Mantzoros C, et al. Role of leptin in the neuroendocrine response to fasting. Nature. 1996;382(6588):250–252. | |
Sull JW, Kim HJ, Yun JE, Park EJ, Kim G, Jee SH. Serum adiponectin is associated with smoking status in healthy Korean men. Endocr J. 2009;56(1):73–78. | |
Isidori AM, Strollo F, Morè M, et al. Leptin and aging: correlation with endocrine changes in male and female healthy adult populations of different body weights. J Clin Endocrinol Metab. 2000;85(5):1954–1962. | |
Somech R, Reif S, Golander A, Spirer Z. Leptin and C-reactive protein levels correlate during minor infection in children. Isr Med Assoc J. 2007;9(2):76–78. | |
Mancuso P, Gottschalk A, Phare SM, Peters-Golden M, Lukacs NW, Huffnagle GB. Leptin-deficient mice exhibit impaired host defense in Gram-negative pneumonia. J Immunol. 2002;168(8):4018–4024. | |
Nakanishi S, Yamane K, Kamei N, Nojima H, Okubo M, Kohno N. A protective effect of adiponectin against oxidative stress in Japanese Americans: the association between adiponectin or leptin and urinary isoprostane. Metabolism. 2005;54(2):194–199. | |
Lord GM, Matarese G, Howard JK, Baker RJ, Bloom SR, Lechler RI. Leptin modulates the T-cell immune response and reverses starvation-induced immunosuppression. Nature. 1998;394(6696):897–901. | |
Friedman JM, Halaas JL. Leptin and the regulation of body weight in mammals. Nature. 1998;395(6704):763–770. | |
Takabatake N, Nakamura H, Abe S, et al. Circulating leptin in patients with chronic obstructive pulmonary disease. Am J Respir Crit Care Med. 1999;159(4 Pt 1):1215–1219. | |
Lumeng CN, Bodzin JL, Saltiel AR. Obesity induces a phenotypic switch in adipose tissue macrophage polarization. J Clin Invest. 2007;117(1):175–184. | |
Weisberg SP, McCann D, Desai M, Rosenbaum M, Leibel RL, Ferrante AW Jr. Obesity is associated with macrophage accumulation in adipose tissue. J Clin Invest. 2003;112(12):1796–1808. | |
Deiuliis J, Shah Z, Shah N, et al. Visceral adipose inflammation in obesity is associated with critical alterations in T regulatory cell numbers. PLoS One. 2011;6(1):e16376. | |
Altintas MM, Azad A, Nayer B, et al. Mast cells, macrophages, and crown-like structures distinguish subcutaneous from visceral fat in mice. J Lipid Res. 2011;52(3):480–488. | |
Ohmura K, Ishimori N, Ohmura Y, et al. Natural killer T cells are involved in adipose tissues inflammation and glucose intolerance in diet-induced obese mice. Arterioscler Thromb Vasc Biol. 2010;30(2):193–199. | |
Bedford PA, Todorovic V, Westcott ED, et al. Adipose tissue of human omentum is a major source of dendritic cells, which lose MHC class II and stimulatory function in Crohn’s disease. J Leukoc Biol. 2006;80(3):546–554. | |
Cildir G, Akincilar SC, Tergaonkar V. Chronic adipose tissue inflammation: all immune cells on the stage. Trends Mol Med. 2013;19(8):487–500. | |
Ginhoux F, Schultze JL, Murray PJ, Ochando J, Biswas SK. New insights into the multidimensional concept of macrophage ontogeny, activation and function. Nat Immunol. 2015;17(1):34–40. | |
Lumeng CN, Deyoung SM, Saltiel AR. Macrophages block insulin action in adipocytes by altering expression of signaling and glucose transport proteins. Am J Physiol Endocrinol Metab. 2007;292(1):E166–E174. | |
Poglio S, De Toni-Costes F, Arnaud E, et al. Adipose tissue as a dedicated reservoir of functional mast cell progenitors. Stem Cells. 2010;28(11):2065–2072. | |
Bertola A, Ciucci T, Rousseau D, et al. Identification of adipose tissue dendritic cells correlated with obesity-associated insulin-resistance and inducing Th17 responses in mice and patients. Diabetes. 2012;61(9):2238–2247. | |
Talukdar S, Oh DY, Bandyopadhyay G, et al. Neutrophils mediate insulin resistance in mice fed a high-fat diet through secreted elastase. Nat Med. 2012;18(9):1407–1412. | |
Meakin PJ, Morrison VL, Sneddon CC, et al. Mice lacking β2-integrin function remain glucose tolerant in spite of insulin resistance, neutrophil infiltration and inflammation. PLoS One. 2015;10(9):e0138872. | |
Wensveen FM, Jelenčić V, Valentić S, et al. NK cells link obesity-induced adipose stress to inflammation and insulin resistance. Nat Immunol. 2015;16(4):376–385. | |
Winer DA, Winer S, Chng MH, Shen L, Engleman EG. B lymphocytes in obesity-related adipose tissue inflammation and insulin resistance. Cell Mol Life Sci. 2013;71(6):1033–1043. | |
Feuerer M, Herrero L, Cipolletta D, et al. Lean, but not obese, fat is enriched for a unique population of regulatory T cells that affect metabolic parameters. Nat Med. 2009;15(8):930–939. | |
Nishimura S, Manabe I, Nagasaki M, et al. CD8+ effector T cells contribute to macrophage recruitment and adipose tissue inflammation in obesity. Nat Med. 2009;15(8):914–920. | |
De Rosa V, Procaccini C, Calì G, et al. A key role of leptin in the control of regulatory T cell proliferation. Immunity. 2007;26(2):241–255. | |
Summer R, Little FF, Ouchi N, et al. Alveolar macrophage activation and an emphysema-like phenotype in adiponectin-deficient mice. Am J Physiol Lung Cell Mol Physiol. 2008;294(6): L1035–L1042. | |
Farooqi IS, Matarese G, Lord GM, et al. Beneficial effects of leptin on obesity, T cell hyporesponsiveness, and neuroendocrine/metabolic dysfunction of human congenital leptin deficiency. J Clin Invest. 2002;110(8):1093–1103. | |
Rodriguez L, Graniel J, Ortiz R. Effect of leptin on activation and cytokine synthesis in peripheral blood lymphocytes of malnourished infected children. Clin Exp Immunol. 2007;148(3):478–485. | |
Matarese G, Carrieri PB, La Cava A, et al. Leptin increase in multiple sclerosis associates with reduced number of CD4+CD25+ regulatory T cells. Proc Natl Acad Sci U S A. 2005;102(14):5150–5155. | |
Gutiérrez-Vidal R, Vega-Badillo J, Reyes-Fermín LM, et al. SFRP5 hepatic expression is associated with non-alcoholic liver disease in morbidly obese women. Ann Hepatol. 2015;14(5):666–674. | |
Ohashi K, Shibata R, Murohara T, Ouchi N. Role of anti-inflammatory adipokines in obesity-related diseases. Trends Endocrinol Metab. 2014;25(7):348–355. | |
Tan YL, Zheng XL, Tang CK. The protective functions of omentin in cardiovascular diseases. Clin Chim Acta. 2015;448:98–106. | |
Auguet T, Quintero Y, Riesco D, et al. New adipokines vaspin and omentin: circulating levels and gene expression in adipose tissue from morbidly obese women. BMC Med Genet. 2011;12:60. | |
Scherer PE, Williams S, Fogliano M, Baldini G, Lodish HF. A novel serum protein similar to C1q, produced exclusively in adipocytes. J Biol Chem. 1995;270(45):26746–26749. | |
Cawthorn WP, Scheller EL, Learman BS, et al. Bone marrow adipose tissue is an endocrine organ that contributes to increased circulating adiponectin during caloric restriction. Cell Metab. 2014;20(2): 368–375. | |
Combs TP, Berg AH, Rajala MW, et al. Sexual differentiation, pregnancy, calorie restriction, and aging affect the adipocyte-specific secretory protein adiponectin. Diabetes. 2003;52(2):268–276. | |
Fazeli PK, Horowitz MC, MacDougald OA, et al. Marrow fat and bone – new perspectives. J Clin Endocrinol Metab. 2013;98(3): 935–945. | |
Hong SC, Yoo SW, Cho GJ, et al. Correlation between estrogens and serum adipocytokines in premenopausal and postmenopausal women. Menopause. 2007;14(5):835–840. | |
Imagawa A, Funahashi T, Nakamura T, et al. Elevated serum concentration of adipose-derived factor, adiponectin, in patients with type 1 diabetes. Diabetes Care. 2002;25(9):1665–1666. | |
Yu JG, Javorschi S, Hevener AL, et al. The effect of thiazolidinediones on plasma adiponectin levels in normal, obese, and type 2 diabetic subjects. Diabetes. 2002;51(10):2968–2974. | |
Yuan H, Wong LS, Bhattacharya M, et al. The effects of second-hand smoke on biological processes important in atherogenesis. BMC Cardiovasc Disord. 2007;7:1. | |
Yamauchi T, Kamon J, Ito Y, et al. Cloning of adiponectin receptors that mediate antidiabetic metabolic effects. Nature. 2003; 423(6941):762–769. | |
Yokota T, Oritani K, Takahashi I, et al. Adiponectin, a new member of the family of soluble defense collagens, negatively regulates the growth of myelomonocytic progenitors and the functions of macrophages. Blood. 2000;96(5):1723–1732. | |
Yamaguchi N, Argueta JG, Masuhiro Y, et al. Adiponectin inhibits Toll-like receptor family-induced signaling. FEBS Lett. 2005;579(30): 6821–6826. | |
Chandrasekar B, Boylston WH, Venkatachalam K, Webster NJ, Prabhu SD, Valente AJ. Adiponectin blocks interleukin-18-mediated endothelial cell death via APPL1-dependent AMP-activated protein kinase (AMPK) activation and IKK/NF-κB/PTEN suppression. J Biol Chem. 2008;283(36):24889–24898. | |
Yoshida S, Fuster JJ, Walsh K. Adiponectin attenuates abdominal aortic aneurysm formation in hyperlipidemic mice. Atherosclerosis. 2014;235(2):339–346. | |
Konter JM, Parker JL, Baez E, et al. Adiponectin attenuates lipopolysaccharide-induced acute lung injury through suppression of endothelial cell activation. J Immunol. 2012;188(2):854–863. | |
Medoff BD, Okamoto Y, Leyton P, et al. Adiponectin-deficiency increases allergic airway inflammation and pulmonary vascular remodeling. Am J Respir Cell Mol Biol. 2009;41(4):397–406. | |
Ruan CC, Ge Q, Li Y, et al. Complement-mediated macrophage polarization in perivascular adipose tissue contributes to vascular injury in deoxycorticosterone acetate-salt mice. Arterioscler Thromb Vasc Biol. 2015;35(3):598–606. | |
Shore SA, Terry RD, Flynt L, Xu A, Hug C. Adiponectin attenuates allergen-induced airway inflammation and hyperresponsiveness in mice. J Allergy Clin Immunol. 2006;118(2):389–395. | |
Verbout NG, Benedito L, Williams AS, et al. Impact of adiponectin overexpression on allergic airways responses in mice. J Allergy (Cairo). 2013;2013:349520. | |
Shibata R, Sato K, Pimentel DR, et al. Adiponectin protects against myocardial ischemia-reperfusion injury through AMPK- and COX-2-dependent mechanisms. Nat Med. 2005;11(10):1096–1103. | |
Bas S, Finckh A, Puskas GJ, et al. Adipokines correlate with pain in lower limb osteoarthritis: different associations in hip and knee. Int Orthop. 2014;38(12):2577–2583. | |
Kopp A, Bala M, Buechler C, et al. C1q/TNF-related protein-3 represents a novel and endogenous lipopolysaccharide antagonist of the adipose tissue. Endocrinology. 2010;151(11):5267–5278. | |
Wei Z, Peterson JM, Wong GW. Metabolic regulation by C1q/TNF-related Protein-13 (CTRP13): activation of AMP-activated protein kinase and suppression of fatty acid-induced JNK signaling. J Biol Chem. 2011;286(18):15652–15665. | |
Yamawaki H, Kuramoto J, Kameshima S, Usui T, Okada M, Hara Y. Omentin, a novel adipocytokine inhibits TNF-induced vascular inflammation in human endothelial cells. Biochem Biophys Res Commun. 2011;408(2):339–343. | |
Nakamura K, Sano S, Fuster JJ, et al. Secreted fizzled-related protein 5 diminishes cardiac inflammation and protects the heart from ischemia-reperfusion injury. J Biol Chem. 2016;291(6):2566–2575. | |
La Cava A, Matarese G. The weight of leptin in immunity. Nat Rev Immunol. 2004;4(5):371–379. | |
Loffreda S, Yang S, Lin H, et al. Leptin regulates proinflammatory immune responses. FASEB J. 1998;12(1):57–65. | |
Gainsford T, Willson TA, Metcalf D, et al. Leptin can induce proliferation, differentiation, and functional activation of hemopoietic cells. Proc Natl Acad Sci U S A. 1996;93(25):14564–14568. | |
Mancuso P, Canetti C, Gottschalk A, Tithof PK, Peters-Golden M. Leptin augments alveolar macrophage leukotriene synthesis by increasing phospholipase activity and enhancing group IVC iPLA2 (cPLA2γ) protein expression. Am J Physiol Lung Cell Mol Physiol. 2004;287(3):L497–L502. | |
Yamagishi SI, Edelstein D, Du XL, Kaneda Y, Guzmán M, Brownlee M. Leptin induces mitochondrial superoxide production and monocyte chemoattractant protein-1 expression in aortic endothelial cells by increasing fatty acid oxidation via protein kinase A. J Biol Chem. 2001;276(27):25096–25100. | |
Raso GM, Pacilio M, Esposito E, Coppola A, Di Carlo R, Meli R. Leptin potentiates IFN-γ-induced expression of nitric oxide synthase and cyclo-oxygenase-2 in murine macrophage J774A.1. Br J Pharmacol. 2002;137(6):799–804. | |
Caldefie-Chezet F, Poulin A, Vasson MP. Leptin regulates functional capacities of polymorphonuclear neutrophils. Free Radic Res. 2003;37(8):809–814. | |
Bouloumie A, Marumo T, Lafontan M, Busse R. Leptin induces oxidative stress in human endothelial cells. FASEB J. 1999;13(10):1231–1238. | |
Konstantinides S, Schäfer K, Koschnick S, Loskutoff DJ. Leptin-dependent platelet aggregation and arterial thrombosis suggests a mechanism for atherothrombotic disease in obesity. J Clin Invest. 2001;108(10):1533–1540. | |
Santos-Alvarez J, Goberna R, Sánchez-Margalet V. Human leptin stimulates proliferation and activation of human circulating monocytes. Cell Immunol. 1999;194(1):6–11. | |
Zarkesh-Esfahani H, Pockley G, Metcalfe RA, et al. High-dose leptin activates human leukocytes via receptor expression on monocytes. J Immunol. 2001;167(8):4593–4599. | |
Reis BS, Lee K, Fanok MH, et al. Leptin receptor signaling in T cells is required for Th17 differentiation. J Immunol. 2015;194(11): 5253–5260. | |
Steppan CM, Bailey ST, Bhat S, et al. The hormone resistin links obesity to diabetes. Nature. 2001;409(6818):307–312. | |
Qatanani M, Szwergold NR, Greaves DR, Ahima RS, Lazar MA. Macrophage-derived human resistin exacerbates adipose tissue inflammation and insulin resistance in mice. J Clin Invest. 2009;119(3): 531–539. | |
Yang RZ, Huang Q, Xu A, et al. Comparative studies of resistin expression and phylogenomics in human and mouse. Biochem Biophys Res Commun. 2003;310(3):927–935. | |
Lee S, Lee HC, Kwon YW, et al. Adenylyl cyclase-associated protein 1 is a receptor for human resistin and mediates inflammatory actions of human monocytes. Cell Metab. 2014;19(3):484–497. | |
Rourke JL, Dranse HJ, Sinal CJ. Towards an integrative approach to understanding the role of chemerin in human health and disease. Obes Rev. 2013;14(3):245–262. | |
Newcomer ME. Retinoid-binding proteins: structural determinants important for function. FASEB J. 1995;9(2):229–239. | |
Moraes-Vieira PM, Yore MM, Dwyer PM, Syed I, Aryal P, Kahn BB. RBP4 activates antigen-presenting cells, leading to adipose tissue inflammation and systemic insulin resistance. Cell Metab. 2014;19(3):512–526. | |
Wang HH, Wu MM, Chan MW, Pu YS, Chen CJ, Lee TC. Long-term low-dose exposure of human urothelial cells to sodium arsenite activates lipocalin-2 via promoter hypomethylation. Arch Toxicol. 2014;88(8):1549–1559. | |
Bratt T, Ohlson S, Borregaard N. Interactions between neutrophil gelatinase-associated lipocalin and natural lipophilic ligands. Biochim Biophys Acta. 1999;1472(1–2):262–269. | |
Flo TH, Smith KD, Sato S, et al. Lipocalin 2 mediates an innate immune response to bacterial infection by sequestrating iron. Nature. 2004;432(7019):917–921. | |
Goossens GH. The role of adipose tissue dysfunction in the pathogenesis of obesity-related insulin resistance. Physiol Behav. 2008;94(2):206–218. | |
Urbanová M, Dostálová I, Trachta P, et al. Serum concentrations and subcutaneous adipose tissue mRNA expression of omentin in morbid obesity and type 2 diabetes mellitus: the effect of very-low-calorie diet, physical activity and laparoscopic sleeve gastrectomy. Physiol Res. 2014;63(2):207–218. | |
Schulte DM, Müller N, Neumann K, et al. Pro-inflammatory Wnt5a and anti-inflammatory sFRP5 are differentially regulated by nutritional factors in obese human subjects. PLoS One. 2012;7(2):e32437. | |
Clément K, Viguerie N, Poitou C, et al. Weight loss regulates inflammation-related genes in white adipose tissue of obese subjects. FASEB J. 2004;18(14):1657–1669. | |
Ye RR, Scherer PE. Adiponectin, driver or passenger on the road to insulin sensitivity? Mol Metab. 2013;2(3):133–141. | |
Yamauchi T1, Kamon J, Minokoshi Y, et al. Adiponectin stimulates glucose utilization and fatty-acid oxidation by activating AMP-activated protein kinase. Nat Med. 2002;8(11):1288–1295. | |
Nishimura S, Manabe I, Takaki S, et al. Adipose natural regulatory B cells negatively control adipose tissue inflammation. Cell Metab. 2013;18(5):759–766. | |
Howard J, Lord G, Matarese G, et al. Leptin protects mice from starvation-induced lymphoid atrophy and increases thymic cellularity in ob/ob mice. J Clin Invest. 1999;104(8):1051–1059. | |
Harvey AE, Lashinger LM, Hays D, et al. Calorie restriction decreases murine and human pancreatic tumor cell growth, nuclear factor-κB activation, and inflammation-related gene expression in an insulin-like growth factor-1−dependent manner. PLoS One. 2014;9(5): e94151. | |
Gardner EM. Caloric restriction decreases survival of aged mice in response to primary influenza infection. J Gerontol A Biol Sci Med Sci. 2005;60(6):688–694. | |
Ritz BW, Aktan I, Nogusa S, Gardner EM. Energy restriction impairs natural killer cell function and increases the severity of influenza infection in young adult male C57BL/6 mice. J Nutr. 2008;138(11):2269–2275. | |
Clinthorne JF, Adams DJ, Fenton JI, Ritz BW, Gardner EM. Short-term re-feeding of previously energy-restricted C57BL/6 male mice restores body weight and body fat and attenuates the decline in natural killer cell function after primary influenza infection. J Nutr. 2010;140(8):1495–1501. | |
Boden G, Chen X, Mozzoli M, Ryan I. Effect of fasting on serum leptin in normal human subjects. J Clin Endocrinol Metab. 1996;81(9):3419–3423. | |
Merl V, Peters A, Oltmanns KM, et al. Serum adiponectin concentrations during a 72-hour fast in over- and normal-weight humans. Int J Obes Relat Metab Disord. 2005;29(8):998–1001. | |
Fazeli PK, Lun M, Kim SM, et al. FGF21 and the late adaptive response to starvation in humans. J Clin Invest. 2015;125(12):4601–4611. | |
Faggioni R, Moser A, Feingold KR, Grunfeld C. Reduced leptin levels in starvation increase susceptibility to endotoxic shock. Am J Pathol. 2000;156(5):1781–1787. | |
Fujita Y, Yanagida H, Mimori T, et al. Prevention of fasting-mediated bone marrow atrophy by leptin administration. Cell Immunol. 2012;273(1):52–58. | |
Bartz S, Mody A, Hornik C, et al. Severe acute malnutrition in childhood: hormonal and metabolic status at presentation, response to treatment, and predictors of mortality. J Clin Endocrinol Metab. 2014;99(6):2128–2137. | |
Katona P, Katona-Apte J. Clinical practice: the interaction between nutrition and infection. Clin Infect Dis. 2008;46(10):1582–1588. | |
Cinti S, Mitchell G, Barbatelli G, et al. Adipocyte death defines macrophage localization and function in adipose tissue of obese mice and humans. J Lipid Res. 2005;46(11):2347–2355. | |
Berger E, Héraud S, Mojallal A, et al. Pathways commonly dysregulated in mouse and human obese adipose tissue: FAT/CD36 modulates differentiation and lipogenesis. Adipocyte. 2015;4(3):161–180. | |
Drolet R, Richard C, Sniderman AD, et al. Hypertrophy and hyperplasia of abdominal adipose tissues in women. Int J Obes (Lond). 2008;32(2):283–291. | |
Frayn KN. Adipose tissue as a buffer for daily lipid flux. Diabetologia. 2002;45(9):1201–1210. | |
Mori H, Prestwich TC, Reid MA, et al. Secreted Frizzled-related protein 5 suppresses adipocyte mitochondrial metabolism through WNT inhibition. J Clin Invest. 2012;122(7):2405–2416. | |
Seidell JC, Pérusse L, Després JP, Bouchard C. Waist and hip circumferences have independent and opposite effects on cardiovascular disease risk factors: the Quebec Family Study. Am J Clin Nutr. 2001;74(3):315–321. | |
Milewicz A, Jdrzejuk D, Dunajska K, Lwow F. Waist circumference and serum adiponectin levels in obese and non-obese postmenopausal women. Maturitas. 2010;65(3):272–275. | |
Garaulet M, Perex-Llamas F, Fuente T, Zamora S, Tebar FJ. Anthropometric, computed tomography and fat cell data in an obese population: relationship with insulin, leptin, tumor necrosis factor-alpha, sex hormone-binding globulin and sex hormones. Eur J Endocrinol. 2000;143(5):657–666. | |
Yusuf S, Hawken S, Ounpuu S, et al. Obesity and the risk of myocardial infarction in 27 000 participants from 52 countries: a case-control study. Lancet. 2005;366(9497):1640–1649. | |
Wu D, Molofsky AB, Liang HE, et al. Eosinophils sustain adipose alternatively activated macrophages associated with glucose homeostasis. Science. 2011;332(6026):243–247. | |
Pacifico L, Di Renzo L, Anania C, et al. Increased T-helper interferon-γ-secreting cells in obese children. Eur J Endocrinol. 2006;154(5):691–697. | |
Talukdar S, Oh DY, Bandyopadhyay G, et al. Neutrophils mediate insulin resistance in mice fed a high-fat diet through secreted elastase. Nat Med. 2012;18(9):1407–1412. | |
Theoharides TC, Sismanopoulos N, Delivanis DA, Zhang B, Hatziagelaki EE, Kalogeromitros D. Mast cells squeeze the heart and stretch the gird: their role in atherosclerosis and obesity. Trends Pharmacol Sci. 2011;32(9):534–542. | |
Fried SK, Bunkin DA, Greenberg AS. Omental and subcutaneous adipose tissues of obese subjects release interleukin-6: depot difference and regulation by glucocorticoid. J Clin Endocrinol Metab. 1998;83(3):847–850. | |
Hotamisligil GS, Shargill NS, Spiegelman BM. Adipose expression of tumor necrosis factor-α: direct role in obesity-linked insulin resistance. Science. 1993;259(5091):87–91. | |
Wood IS, Wang B, Jenkins JR, Trayhurn P. The pro-inflammatory cytokine IL-18 is expressed in human adipose tissue and strongly upregulated by TNFα in human adipocytes. Biochem Biophys Res Commun. 2005;337(2):422–429. | |
Netea MG, Joosten LA, Lewis E, et al. Deficiency of interleukin-18 in mice leads to hyperphagia, obesity and insulin resistance. Nat Med. 2006;12(6):650–656. | |
Huang F, Del-Río-Navarro BE, Pérez-Ontiveros JA, et al. Effect of six-month lifestyle intervention on adiponectin, resistin and soluble tumor necrosis factor-α receptors in obese adolescents. Endocr J. 2014;61(9):921–931. | |
Pasarica M, Sereda OR, Redman LM, et al. Reduced adipose tissue oxygenation in human obesity: evidence for rarefaction, macrophage chemotaxis, and inflammation without an angiogenic response. Diabetes. 2009;58(3):718–725. | |
Harlev AA. Macrophage infiltration and stress-signaling in omental and subcutaneous adipose tissue in diabetic pregnancies. J Matern Fetal Neonatal Med. 2014;27(12):1189–1194. | |
Murphy K. Basic concepts of immunology. In: Janeway’s Immunobiology. 8th ed. London: Taylor and Francis; 2012:1–36. | |
Hotamisligil GS, Erbay E. Nutrient sensing and inflammation in metabolic diseases. Nat Rev Immunol. 2008;8(12):923–934. | |
Hosogai N, Fukuhara A, Oshima K, et al. Adipose tissue hypoxia in obesity and its impact on adipocytokine dysregulation. Diabetes. 2007;56(4):901–911. | |
Karvonen-Gutierrez CA, Harlow SD, Mancuso P, Jacobson J, de Leon CF, Nan B. Leptin levels are associated with radiographic knee osteoarthritis among a cohort of mid-life women. Arthritis Care Res (Hoboken). 2013;65(6):936–944. | |
Karvonen-Gutierrez CA, Harlow SD, Jacobson J, Mancuso P, Jiang Y. The relationship between longitudinal serum leptin measures and measures of magnetic resonance imaging-assessed knee joint damage in a population of mid-life women. Ann Rheum Dis. 2014;73(5):883–889. | |
Dattaroy D, Pourhoseini S, Das S, et al. Micro-RNA 21 inhibition of SMAD7 enhances fibrogenesis via leptin-mediated NADPH oxidase in experimental and human nonalcoholic steatohepatitis. Am J Physiol Gastrointest Liver Physiol. 2015;308(4):G298–G312. | |
Bodary PF, Westrick RJ, Wickenheiser KJ, Shen Y, Eitzman DT. Effect of leptin on arterial thrombosis following vascular injury in mice. JAMA. 2002;287(13):1706–1709. | |
Lim CC, Teo BW, Tai ES, et al. Elevated serum leptin, adiponectin and leptin to adiponectin ratio is associated with chronic kidney disease in Asian adults. PLoS one. 2015;10(3):e0122009. | |
Hasan-Ali H, Abd El-Mottaleb NA, Hamed HB, Abd-Elsayed A. Serum adiponectin and leptin as predictors of the presence and degree of coronary atherosclerosis. Coron Artery Dis. 2011;22(4):264–269. | |
Shamsuzzaman AS, Winnicki M, Wolk R, et al. Independent association between plasma leptin and C-reactive protein in healthy humans. Circulation. 2004;109(18):2181–2185. | |
Petersen KF, Oral EA, Dufour S, et al. Leptin reverses insulin resistance and hepatic steatosis in patients with severe lipodystrophy. J Clin Invest. 2002;109(10):1345–1350. | |
Cochran E, Young JR, Sebring N, DePaoli A, Oral EA, Gorden P. Efficacy of recombinant methionyl human leptin therapy for the extreme insulin resistance of the Rabson-Mendenhall syndrome. J Clin Endocrinol Metab. 2004;89(4):1548–1554. | |
Hsu WY, Chao YW, Tsai YL, et al. Resistin induces monocyte-endothelial cell adhesion by increasing ICAM-1 and VCAM-1 expression in endothelial cells via p38MAPK-dependent pathway. J Cell Physiol. 2011;226(8):2181–2188. | |
Verma S, Li SH, Wang CH, et al. Resistin promotes endothelial cell activation: further evidence of adipokine-endothelial interaction. Circulation. 2003;108(6):736–740. | |
Goralski KB, McCarthy TC, Hanniman EA, et al. Chemerin, a novel adipokine that regulates adipogenesis and adipocyte metabolism. J Biol Chem. 2007;282(38):28175–28188. | |
Ernst MC, Issa M, Goralski KB, Sinal CJ. Chemerin exacerbates glucose intolerance in mouse models of obesity and diabetes. Endocrinology. 2010;151(5):1998–2007. | |
Klöting N, Graham TE, Berndt J, et al. Serum retinol-binding protein is more highly expressed in visceral than in subcutaneous adipose tissue and is a marker of intra-abdominal fat mass. Cell Metab. 2007;6(1):79–87. | |
Broch M, Ramírez R, Auguet MT, et al. Macrophages are novel sites of expression and regulation of retinol binding protein-4 (RBP4). Physiol Res. 2010;59(2):299–303. | |
Yang Q, Graham TE, Mody N, et al. Serum retinol binding protein 4 contributes to insulin resistance in obesity and type 2 diabetes. Nature. 2005;436(7049):356–362. | |
Samaras K, Botelho NK, Chisholm DJ, Lord RV. Subcutaneous and visceral adipose tissue gene expression of serum adipokines that predict type 2 diabetes. Obesity (Silver Spring). 2010;18(5):884–889. | |
Hemdahl AL, Gabrielsen A, Zhu C, et al. Expression of neutrophil gelatinase-associated lipocalin in atherosclerosis and myocardial infarction. Arterioscler Thromb Vasc Biol. 2006;26(1):136–142. |
 © 2016 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2016 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.