Back to Journals » Diabetes, Metabolic Syndrome and Obesity » Volume 18
The Proteomic Analysis of Intermittent Fasting Alone or with GLP-1RA in NAFLD Rats
Authors Shao Y ![]() , Xu S
, Xu S ![]() , Ma Y
, Ma Y ![]() , Zhang J, Guo X, Zhang T
, Zhang J, Guo X, Zhang T ![]() , Yuan G
, Yuan G ![]()
Received 17 July 2025
Accepted for publication 29 October 2025
Published 5 November 2025 Volume 2025:18 Pages 4073—4083
DOI https://doi.org/10.2147/DMSO.S550262
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Jae Woong Sull
Yimin Shao,1 Shengya Xu,1 Yuanyuan Ma,2 Junqing Zhang,1 Xiaohui Guo,1 Tingting Zhang,1 Geheng Yuan1
1Department of Endocrinology, Peking University First Hospital, Beijing, People’s Republic of China; 2Laboratory Animal Center, Peking University First Hospital, Beijing, People’s Republic of China
Correspondence: Tingting Zhang, Department of Endocrinology, Peking University First Hospital, No. 8 Xishiku Street, Xicheng District, Beijing, 100034, People’s Republic of China, Tel +86 10 83575103, Fax +86 10 66552395, Email [email protected] Geheng Yuan, Department of Endocrinology, Peking University First Hospital, No. 8 Xishiku Street, Xicheng District, Beijing, 100034, People’s Republic of China, Tel +86 10 83575103, Fax +86 10 66552395, Email [email protected]
Background: Intermittent fasting (IF) and glucagon-like peptide-1 receptor agonists (GLP-1RA) offer effective therapeutic options for nonalcoholic fatty liver disease (NAFLD). This study aimed to examine the effects of alternate-day fasting (ADF) alone and with liraglutide, and to explore the mechanisms behind each treatment.
Methods: To establish the model for NAFLD, Sprague-Dawley rats were given a high-fat diet for 25 weeks. Subsequently, the rats were assigned to the ADF, ADF combined with liraglutide (A+L), and control groups for an additional 5 weeks. Evaluations were performed on liver morphology, body weight, serum lipid profiles, insulin sensitivity, and liver proteomics.
Results: Compared to the control group, ADF alone demonstrated a reduction in body weight (37.1± 15.56 g vs − 20.68± 15.58 g, p< 0.05), food intake (0.30± 0.002 g/d/rat vs 0.18± 6.21 g/d/rat), blood lipid levels, and ALT concentration (139.0± 15.57 U/L vs 91.25± 41.9 U/L, p< 0.05), while also improving the NAFLD score. Furthermore, ADF in conjunction with liraglutide exhibited superior effects in reducing body weight (− 96.15± 15.78 g), food intake (− 10.25± 0.01 g/d/rat), triglyceride (17.53± 0.25 nmol/L), and LDL concentrations (6.93± 0.35mmol/L), as well as ameliorating insulin resistance and lowering the NAFLD score relative to the ADF group (p< 0.05). The proteomic analysis indicates that G protein-coupled receptor 39 (GPR39) and transmembrane protein 41b (Tmem41b) were significantly upregulated in the A+L group compared to the other two groups. Additionally, hydroxysteroid 17β-dehydrogenase 2 (HSD17B2) was significantly diminished in both the ADF and A+L groups relative to the control group.
Conclusion: Intermittent fasting, in conjunction with GLP-1RA, further enhances metabolic health in individuals with NAFLD as a result of obesity. This study provides support for forthcoming clinical trials and aims to establish new therapeutic targets in the context of NAFLD.
Keywords: intermittent fasting, non-alcoholic fatty liver disease, glucagon-like peptide-1, proteomic changes, obesity
Introduction
Obesity, particularly abdominal obesity, is a risk factor for metabolic disorders such as hyperinsulinemia, diabetes, and dyslipidemia.1–3 Non-alcoholic fatty liver disease (NAFLD), which is associated with obesity, has been witnessing a rising prevalence and has emerged as a global public health issue,4,5 the incidence of which is predicted to rise by as much as 56% globally in the next ten years.6 At present, only resmetirom has been approved by the Food and Drug Administration (FDA) for the treatment of NAFLD and lifestyle interventions, such as weight loss and exercise, continue to be the cornerstones for treatment.7
Intermittent fasting (IF) is a widely recognized and debated dietary practice that includes a range of fasting protocols. Research has demonstrated that intermittent fasting is ultimately effective in decreasing body weight, adiposity, and insulin resistance.8,9 Several studies have emphasized IF’s therapeutic potential in NAFLD with the potential of metabolism improvement,10,11 but the specific mechanisms remain unclear, particularly in the liver.12 Nevertheless, some contradictory results remain. Studies have found that alternate-day fasting (ADF) increases visceral fat content.13 There are numerous challenges with IF when it comes to achieving and maintaining sustainable weight loss.14
Glucagon-like peptide-1 receptor agonists (GLP-1RAs) are medications originally developed to treat type 2 diabetes, which can inhibit appetite, increase satiety, and facilitate weight loss.15,16 Our previous study also found that GLP-1 treatment can significantly reduce visceral fat while activating AMPK.15 Additionally, several studies show that GLP-1RAs can reduce liver enzymes and oxidative stress, as well as improve liver histology in murine models of nonalcoholic steatohepatitis (NASH).17,18 Liraglutide efficacy and action in NASH (LEAN study) showed that liraglutide was safe, well-tolerated, and resulted in improvements in histological steatosis and hepatocyte ballooning.19 Long-acting GLP-1RA semaglutide treatment demonstrates a significantly higher percentage of patients achieving NASH resolution compared to placebo.20
We hypothesized that GLP-1RAs could help curb appetite, resulting in better adherence to long-term intermittent fasting. This, in turn, can lead to more sustainable weight loss and improvements in NAFLD. Therefore, we sought to explore the effects of ADF and its potential mechanisms in the treatment of NAFLD. Furthermore, we examined the impacts of ADF both alone and in combination with liraglutide, as well as the related mechanisms.
Methods
Animals and Procedures
All experiments were conducted under permits from the Experimental Animal Ethics Committee of Peking University First Hospital, China (Permit number: J202005). All animal experiments followed the National Institutes of Health Guidelines for the Care and Use of Laboratory Animals and complied with ARRIVE guidelines. Six-week-old female Sprague-Dawley (SD) rats were purchased from Beijing HFK Bioscience Company. All rats were housed (4 rats per cage) in an air-conditioned room at 22 ± 2 °C with controlled ambient conditions and a 12-hour light–12-hour dark cycle (lights on at 8:00 AM). Drinking water and a high-fat diet (HFD) containing 60% fat (HFK Bioscience, Beijing, China) were provided freely.
All the rats were fed with HFD for 25 weeks to establish obese NAFLD models.21 Then the rats were randomly assigned into three groups: (1) the alternate-day fasting group (ADF, n=4) had total deprivation of food and ad libitum access to water from 08:00–08:00 on alternate days with ad libitum food and water access, and received a subcutaneous injection of 0.9% saline of equal volume with liraglutide once a day; (2) the ADF and liraglutide A+L group (A+L, n=4) had total deprivation of food and ad libitum access to water from 08:00–08:00 on alternate days with ad libitum food and water access, and received a subcutaneous injection of liraglutide (300 μg/kg) once a day; (3) the control group fed ad libitum to HFD (Control, n=4) and received a subcutaneous injection of 0.9% saline of equal volume with liraglutide once a day. Fasting blood glucose level, food intake and body weight were measured every week. After 5 weeks of intervention, the rats were deeply anesthetized with pentobarbital 80mg/kg before they were sacrificed. The liver was dissected and weighed.
Oral Glucose Tolerance Test (OGTT)
For the glucose tolerance test, after 16 hours of fasting, the rats were intragastrically administered 50% glucose (2 mg/g of body weight). Glucose levels were then measured at 0, 30, 60, and 120 minutes using a Glucometer Elite (Roche Corp). Subsequently, the area under the curve (AUC) was analyzed with GraphPad Prism 9 for macOS (La Jolla, CA, USA).
For measuring serum insulin levels, blood samples (50 μL) were collected from the caudal vein at 0, 30, 60, and 120 minutes after glucose loading into a heparinized microhematocrit tube. Insulin levels were tested using an Ultra-Sensitive rat Insulin Enzyme-Linked Immunosorbent Assay (ELISA) Kit (Feimobio, Beijing, China).
Serum Collection and Measurement of Blood Parameters
Blood samples were collected via angular vein puncturing before and after diet intervention. The serum levels of insulin, triglyceride (TG), low-density lipoprotein (LDL), and free fatty acid (FFA) were determined by corresponding commercial ELISA kits (Feimobio, Beijing, China). Alanine aminotransferase (ALT) and Aspartate aminotransferase (AST) were detected by Olympus AU640 automatic analyzer and Fujifilm DRI-CHEM 7000 dry biochemical analyzer.
Histological Analysis of Liver Tissue
The liver was fixed with 4% paraformaldehyde, embedded in paraffin, cut into 4μm thick sections, and stained with hematoxylin-eosin. All images were captured by an Olympus microscope.
NAFLD was confirmed histologically using the NAFLD Activity Score (NAS) system, which measures changes in NAFLD according to several histologic structural characteristics. The scoring system comprised 14 histological features, four of which were assessed using a semi-quantitative methodology: steatosis (0–3), lobular inflammation (0–2), hepatocellular ballooning (0–2), and fibrosis (0–4). The remaining nine features were documented as either present or absent.22
Proteomic Bioinformatics Analysis
Proteomics was performed by Biotree Tech (Shanghai, China). For nano–liquid chromatography-tandem mass spectrometry (LC-MS/MS) analysis, 2 μL of total peptides were separated and analyzed with a nano-Ultra Performance Liquid Chromatography (UPLC) coupled to a Q Exactive HFX Orbitrap instrument (Thermo Fisher Scientific) with a nano-electrospray ion source.
Data-dependent acquisition (DDA) was performed in profile and positive mode with an Orbitrap analyzer at a resolution of 120,000 (@200 m/z) and m/z range of 350–1600 for MS1; for MS2, the resolution was set to 4,500 with a fixed first mass of 110 m/z. The dynamic exclusion time window was 45 seconds. Single-charged peaks and peaks with charge exceeding six were excluded from the DDA procedure.
The classification of differentially expressed proteins was conducted in accordance with the annotations obtained from the UniProt Knowledge Base (https://www.uniprot.org/), the terms of gene ontology, and the Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway (https://www.genome.jp/kegg/pathway.html). Ingenuity Pathway Analysis (IPA, Ingenuity Systems, Redwood City, CA, USA) was utilized to conduct disease and functional analyses, as well as to establish networks with the lists of differentially expressed proteins. IPA employs a right-tailed Fisher’s test to compute a p-value for each network, known as the network score, and to determine which pathways are significantly associated with the input data that are mapped to proteins in the comprehensive Ingenuity Pathways Knowledge Base (Qiagen, Germantown, MD, USA).
All proteomics data were analyzed with R (version 3.6.3) and visualized using Tableau (version 2019.1.2). Outliers surpassing 1.5 times the interquartile range were removed from specific plots to enhance clarity. Fold changes were computed using median values for each group. For statistical analysis, imputation was performed by determining the mean and standard deviation for each protein based on the three lowest values to fill in missing data. A protein was subject to imputation if it was present in 1 or 2 out of 5 samples within a condition and detected in more than 3 samples across all other conditions.
Western Blot Analysis
Total protein was extracted and subsequently quantified utilizing a BCA protein quantification kit (Beyotime, Shanghai, People’s Republic of China). A total of 50 μg of protein from each sample was separated via sodium dodecyl sulfate polyacrylamide gel electrophoresis (12%) and subsequently transferred to a nitrocellulose membrane (0.45 μm pore size; PALL Corporation, Port Washington, NY, USA). After blocking, the membranes underwent incubation with primary antibodies against G protein-coupled receptor 39 (GPR 39, 1:1000, Bioss, Beijing, China), hydroxysteroid 17β-dehydrogenase 2 (HSD17B2, 1:1000, Bioss, Beijing, China), Nuclear factor E2 related factor 2 (Nrf2, 1:1000, Bioss, Beijing, China), and β-actin (1:5000, Abcam, Cambridge, MA, United States) respectively. Following incubation of HRP-labeled secondary anti-rabbit (KeyGen Biotech, Nanjing, China), the bands were visualized by using enhanced chemiluminescence reagents (EMD Millipore, Billerica, MA, USA).
Statistical Analysis
Data were expressed as mean ± S.E.M. Significant differences among groups were evaluated by one-way analysis of variance and Turkey’s multiple comparisons test or by unpaired two-tailed Student’s t-test by using Graph-Pad Prism version 9.0 software. Data were considered to be significantly different if p< 0.05.
Results
ADF Combination with Liraglutide Better Ameliorated Obesity and Insulin Resistance of HFD-Induced Obesity Rats
In comparison to the control group, body weight significantly decreased in the ADF and A+L groups (Figure 1A). The average relative change in body weight was determined by subtracting the pre-intervention weight from the weekly measured weights. The body weight change in the control and ADF groups was 37.1±15.56 g and −20.68±15.58 g, respectively (Table 1, p<0.05). In contrast, the A+L group experienced a change of −96.15±15.78 g (p<0.05). No differences in food intake were observed between the ADF and control groups, but the combined intervention significantly reduced it compared to the ADF group (Figure 1B).
 |
Table 1 Characteristics After Intervention of Each Group |
 |
Figure 1 Effects of ADF alone or in combination with liraglutide on the body weight, glucose and insulin resistance. (A) Body weight and body weight change. (B) Food intake and food intake change. (C) Glucose tolerance test and the area under the curve of the glucose tolerance test. (D) Insulin release test and the area under the curve of the insulin release test. & represents the statistical difference between the control and ADF group (and p < 0.05, andand p < 0.01, andandandand p <0.0001). # represents the statistical difference between the ADF and A+L group (# p < 0.05, ## p < 0.01, ### p < 0.001, #### p < 0.0001). * represents the statistical difference between the control and A+L group (* p < 0.05, ** p < 0.01, *** p < 0.001, **** p < 0.0001). Data are expressed as mean ± SEM. N = 4 for each group, ns for non-significant. |
ADF alone did not enhance glucose tolerance and insulin resistance, showing no significant difference in the AUC of glucose and insulin compared to the control group (Figure 1C and D, p>0.05). However, the AUC of glucose and insulin in the A+L group was significantly lower than that in the control group (Figure 1C and D, p<0.05).
ADF Combination with Liraglutide Showed Better Effects in Lowering Blood Lipid Levels
Serum FFA, triglyceride, and LDL concentrations decreased significantly in the two intervention groups compared with the control group (Figure 2A–C). In comparison to the ADF group, the combination intervention resulted in further reductions in triglyceride and LDL levels (Figure 2B and C, p<0.05). The administration of ADF alone resulted in a significant reduction of ALT concentrations in comparison to the control group; however, the combination intervention did not yield any further reduction (Figure 2D). Additionally, the concentrations of AST did not demonstrate significant changes in both the ADF and A+L groups (Figure 2E).
 |
Figure 2 Effects of ADF alone or in combination with liraglutide on serum lipid parameters. (A) Free fatty acid; (B) Triglyceride; (C) Low-density lipoprotein changes in each group of rats. (D) Alanine aminotransferase; (E) Aspartate aminotransferase changes in each group of rats after intervention. Data are expressed as mean ± SEM. N=4 for each group, * p < 0.05, ** p < 0.01, *** p < 0.001, **** p < 0.0001, ns for non-significant. |
ADF Alone and in Combination with Liraglutide Improved Hepatic Steatosis in HFD-Induced NAFLD Rat Model
HE staining revealed that HFD led to a marked hepatocellular ballooning and mild lobular inflammation in the liver (Figure 3A). At the same time, these changes were reversed by either ADF alone (Figure 3B) or the combination intervention (Figure 3C). The A+L group demonstrated a greater reduction in NAFLD activity score than the ADF group, indicating that the combination intervention could further enhance hepatic steatosis compared to ADF alone (Figure 3D). The liver weight did not decrease significantly with ADF alone, but it decreased significantly in the A+L group (Figure 3E).
 |
Figure 3 ADF and ADF in combination with liraglutide improved hepatic steatosis. (A) HE staining of liver in the control group. (B) HE staining of liver in the ADF group. (C) HE staining of liver in the A+F group. The black arrow indicates the balloon cells. Scale bars: 100 μm. (D) NAFLD activity score. (E) Liver weight of each group. Data are expressed as mean ± SEM. N=4 for each group, * p < 0.05, ** p < 0.01, *** p < 0.001, **** p < 0.0001, ns for non-significant. |
Proteomic Analysis of Livers
To gain a better understanding of potential mechanisms, we employed quantitative proteomics to explore the ADF and A+L-triggered proteomic analysis. After a 5-week intervention, the rat livers from different groups were harvested and processed for preparing proteomic samples. Proteins were tagged using tandem mass tags (TMT), which are isobaric chemical labels that enable multiplexing for relative quantitative proteomic analysis.23 The labeled proteins underwent mass spectrometry analysis. Finally, proteins with at least one unique peptide were identified, and their corresponding false discovery rate was below 0.05%. The principal component analysis demonstrated that the control, ADF and A+L group were clearly separated from each other (Figure 4A). Of the 5608 proteins quantified in the liver, 4926 proteins were quantified in all three groups (Figure 4B). The screening conditions for differential proteins were unique peptide ≥ 1, fold change≥ 1.2 or ≤ 0.83, and p-value < 0.05. Analysis of volcano data revealed 214 significantly different proteins in the livers of the control and ADF groups, with 145 proteins down-regulated and 69 proteins up-regulated (Figure 4C). In the A+L group, 315 proteins were down-regulated and 153 proteins were up-regulated compared to the control group (Figure 4D), while 106 proteins were down-regulated and 100 proteins were up-regulated in the A+L group relative to the ADF group (Figure 4E). In addition, through the pathway enrichment of differential proteins in the KEGG database,24 we found that pathways related to metabolic processes, lipid metabolism, and bile secretion were most significantly influenced among each group (Figure 4F and G).
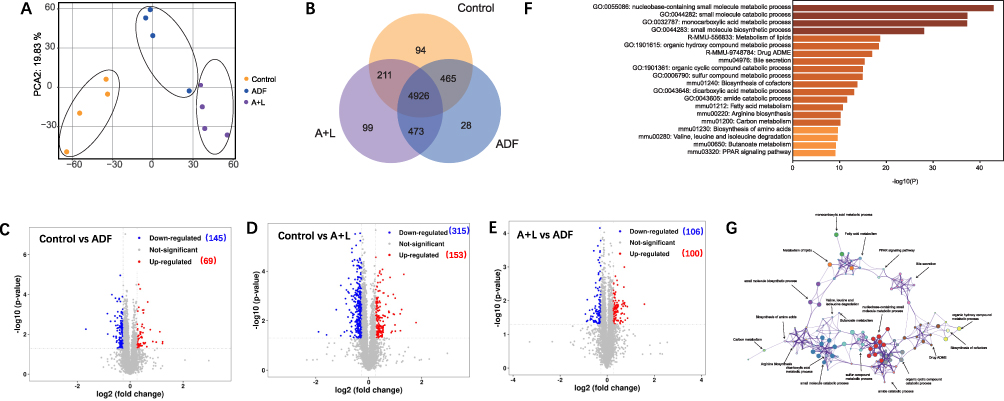 |
Figure 4 Analysis of ADF and combination of ADF and liraglutide on the proteomics of livers from NAFLD rat models. (A) Principal component analysis among each group. Each dot represents the technique replicas in the plot. (B) Venn diagram of coexpressed proteins in each group. (C–E) Volcano plot of the P-values vs the log 2protein abundance differences between control and ADF group (C), control and A+L group (D), and A+L and ADF group (E). Colors: red, upregulation; blue, down-regulation; gray, non-significant differences. (F) KEGG enrichment analysis differential protein among each group. (G) Network for the significantly changed proteins. N=4 for each group. |
Then we conducted a hierarchical cluster analysis of differentially expressed proteins and visualized the results using heat maps (Figure 5A). For lipid metabolism-related pathways, after the intervention, the proteins were downregulated in the bile secretion and amino acid biosynthesis pathways. In contrast, most proteins in the PPAR signaling and fatty acid metabolism pathways were upregulated compared to the control group. The expression of proteins associated with oxidative stress and metabolic pathways exhibited inconsistency between the ADF and A+L groups (Figure 5A). In the most markedly affected metabolic pathways, we found that GPR39, Tmem41b, Inosine-5’-monophosphate dehydrogenase (Impdh2), UDP-glucose 6-dehydrogenase (Ugdh), and α-1,6-mannosyl-glycoprotein2-β-N-acetylglucosaminyltransferase (Mgat2) were increased in the ADF and A+L groups compared with the control group. Still, significant changes between the ADF and A+L groups were observed only for GPR39 and Tmem41b (Figure 5B). Compared to the control group, the expressions of Glycogen debranching enzyme (Agl), 1,4-α-glucan branching enzyme (Gbe1), and HSD17B2 were reduced in both the ADF and A+L groups. However, the combination treatment did not further decrease Agl, Gbe1, and HSD17B2 (Figure 5B).
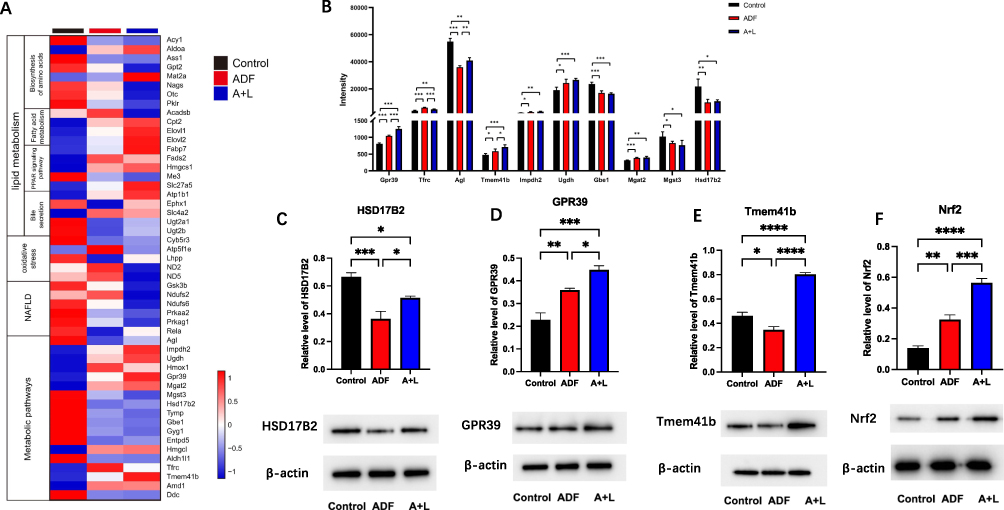 |
Figure 5 Possible genes involved in the most obviously changed pathways after ADF and ADF combined with liraglutide intervention. (A) Heatmap of genes involved in lipid metabolism, oxidative stress, NAFLD, and metabolic pathways in liver. (B) Genes involved in the metabolic pathways of the liver. Protein expression of GPR39 (C), HSD17B2 (D), Tmem41b (E) and Nrf2 (F) in the liver by Western blot. Data are expressed as mean ± SEM. N=4 for each group, * p < 0.05, ** p < 0.01, *** p < 0.001, ****p < 0.0001. |
Potential Mechanisms of ADF Alone and in Combination with Liraglutide Improving NAFLD
After both interventions, HSD17B2 was decreased compared with the control group and there was no difference between the ADF and A+L groups. Therefore, HSD17B2 may be influenced by ADF alone. By Western blot, we verified that the HSD17B2 protein expression was reduced both after ADF alone and in combination with liraglutide (Figure 5C).
In the obviously changed proteins, only changes of GRP39 and Tmem41b were consistent and statistical significance existed between the ADF and A+L group. We verified the expression of GPR39 and Tmem41b proteins in the liver using Western blot analysis. In accordance with the proteomic results, GPR39 protein expression increased significantly after ADF alone and in combination with liraglutide. Moreover, the protein expression of GPR39 in the A+L group was much higher than in the ADF group (Figure 5D). In contrast, Tmem41b protein expression decreased in the ADF group compared to the control group, which contradicted the proteomics results. However, it increased in the A+L group compared to the ADF group (Figure 5E). As GPR39 has been reported to reduce lipid accumulation in a cellular model of NAFLD by activating SIRT1/Nrf2 signaling,25 we further tested the protein expression of Nrf2 in the livers of each group. We found a similar trend of Nrf2 protein expression as GPR39 in each group (Figure 5F), which was most activated in the A+L group.
Discussion
The present study clarified the effects of ADF alone and ADF in combination with liraglutide on the improvement of HFD-induced NAFLD rats. Compared with ADF alone, the A+L group showed better effects in ameliorating obesity, insulin resistance, lowering blood lipid levels and improving NAFLD activity score. To our knowledge, it is the first time to report the proteomics results after ADF alone and in combination with liraglutide.
IF is a beneficial dietary treatment for obesity. However, a recent study using proteomics technology found that after ADF, visceral fat accumulated in the body, resulting in reduced lipolysis and increased fatty acid synthesis in visceral fat.13 Therefore, the long-term effectiveness of ADF in alleviating NAFLD remains controversial. In our study, we first clarified the effects of ADF alone on NAFLD and found ADF mitigated metabolic disorders and hepatic steatosis in the livers of HFD rats. Although body weight decreased in both the ADF and A+L groups, with the most significant reduction in the A+L group, glucose tolerance and insulin levels improved only with the combination intervention. Then we compared the effects of ADF alone and in combination with liraglutide in alleviating NAFLD. As anticipated, the combination treatment demonstrated a more significant enhancement in metabolic disorders and hepatic steatosis among obese rats.
The proteomics results revealed that the involvement of metabolic pathways was most markedly affected in the A+L group. Among the significantly altered liver proteins, two are of particular interest. The first one is GPR39, a seven-transmembrane protein composed of 462 amino acids, which plays a vital role in glucose-induced insulin secretion and the maturation of pancreatic β-cell function.26 Data in the GEO database (GSE63067) indicated that GPR39 was down-regulated in tissues from NAFLD patients. GPR39 deficiency was associated with increased fat accumulation in a high-fat diet, possibly due to reduced energy expenditure and adipocyte lipolytic activity.26 Recently, studies have proved that GPR39 activation of Nrf2 inhibits lipid accumulation both in NAFLD model rats and patients.27,28 The effects caused by overexpression of GPR39 concentrate on its activation of SIRT1 and downstream Nrf2 signaling pathway, as well as the positive regulation of mitochondrial function.25 The disruption of mitochondrial structure and function in hepatocytes, along with the ensuing oxidative stress response, represents a crucial pathogenic mechanism of NAFLD.29 Herein, we found that liver GPR39 and Nrf2 expression were significantly enhanced only in the A+L group. The GPR39 may serve as a potential target for NAFLD. However, this study is a preliminary exploration, and more animal experiments are required to demonstrate.
Another significantly enhanced protein was Tmem41b. Recent studies have found that hepatic Tmem41b, as an endoplasmic reticulum scramblase, participates in lipoprotein generation and the maintenance of lipid homeostasis30 and is associated with autophagy, while the deletion of Tmem41b results in the accumulation of cytoplasmic lipid droplets.31–33 Downregulation of Tmem41b is prevalent in obese mice, rhesus monkeys, and fatty liver patients, while phospholipid scramblase supplementation can alleviate the symptoms of fatty liver.30 We found that Tmem41b expression was significantly increased in the A+L group. However, the exact mechanisms behind the upregulation of GRP39 and Tmem41b after combination therapy remain unclear.
Compared with the control group, HSD17B2 was downregulated in both ADF and the A+L group, and there was no difference between ADF alone and the A+L group. HSD17B2 is a member of the HSD17B family. Recently, HSD17B13 has been demonstrated to play a significant role in the pathogenesis of chronic liver diseases, particularly NAFLD.34–36 To the best of our knowledge, this research represents the first investigation into the role of HSD17B2 in NAFLD. As we know, HSD17B2 catalyzes the interconversion of estradiol and estrone, mediates the oxidation of estradiol (E2) back to estrone, and then loss activity.37 Estrogen is a significant regulator of lipid homeostasis, improving hepatic lipid metabolism.38 Estradiol reduces both liver lipid production by improving β oxidation and very low-density lipoprotein secretion, which may reduce lipotoxicity and thus mitigate the progression of NASH.39 There is no direct evidence of liver estradiol enhancement in the present study, which needs further study. Selective hepatic HSD17B2 inhibitors may be a future treatment for NAFLD.
The present study has some limitations. This study uses the term NAFLD, consistent with the diagnostic criteria and era of data collection. We acknowledge the recent transition to the terminology of metabolic dysfunction-associated steatotic liver disease (MASLD) and will adopt this framework in our subsequent research. Another limitation is the lack of a separate liraglutide group. A previous study compared the effects of liraglutide alone and caloric restriction in adults with obesity, demonstrating that caloric restriction is more effective than liraglutide alone in reducing visceral fat and improving insulin resistance.40 We focused on evaluating the synergistic effects of liraglutide in combination with ADF, demonstrating that their combined intervention outperformed ADF alone in improving NAFLD. Furthermore, our findings indicate that ADF, both independently and alongside liraglutide, holds therapeutic promise for diet-induced NAFLD. Nonetheless, certain groups, such as individuals with diabetes, may require tailored protocols. Therefore, conducting safety studies is essential prior to wider use. Future studies should explore the efficacy of combining GLP-1RAs with other intermittent fasting paradigms, such as time-restricted feeding (TRF).
Conclusion
In conclusion, combining ADF with liraglutide offers a more effective strategy, significantly enhancing the management of metabolic dysregulation and hepatic steatosis in NAFLD. This synergistic effect is partly due to increased expression of GPR39 and TMEM41B—two molecular targets linked to NAFLD development. Notably, ADF alone may boost hepatic estrogen signaling by reducing HSD17B2 levels, providing a new mechanistic perspective on NAFLD remission. Overall, our research highlights the therapeutic promise of integrating intermittent fasting with liraglutide and encourages further clinical studies on this combined approach to improve metabolic health in obesity-related NAFLD.
Data Sharing Statement
The original data for this study can be obtained from the corresponding author upon reasonable request.
Author Contributions
YMS contributed to the conceptualization, data curation, formal analysis, investigation, methodology, project administration, and writing-original draft. SYX contributed to the conceptualization, data curation, methodology, and project administration. YYM contributed to the investigation and project administration. JQZ contributed to the conceptualization and supervision. XHG contributed to the conceptualization and supervision. TTZ contributed to the conceptualization, funding acquisition, resources, supervision, writing- review and editing. GHY contributed to the conceptualization, data curation, formal analysis, resources, supervision, writing- review and editing. All authors took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This work was supported by grants from China Diabetes Young Scientific Talent Research Project (2018‑N‑01) and National High Level Hospital Clinical Research Funding (Interdepartmental Clinical Research Project of Peking University First Hospital, 2022CR04).
Disclosure
The authors have no relevant financial or non-financial interests to disclose.
References
1. Engin A. The definition and prevalence of obesity and metabolic syndrome. Adv Exp Med Biol. 2017;960:1–17. doi:10.1007/978-3-319-48382-5_1
2. Fatade YA, Dave EK, Vatsa N, et al. Obesity and diabetes in heart disease in women. Metabolism Target Organ Damage. 2024;4(3):22. doi:10.20517/mtod.2023.30
3. Litwin M, Kułaga Z. Obesity, metabolic syndrome, and primary hypertension. Pediatr Nephrol. 2021;36(4):825–837. doi:10.1007/s00467-020-04579-3
4. Eslam M, Newsome PN, Sarin SK, et al. A new definition for metabolic dysfunction-associated fatty liver disease: an international expert consensus statement. J Hepatol. 2020;73(1):202–209. doi:10.1016/j.jhep.2020.03.039
5. Wong RJ. Epidemiology of metabolic dysfunction-associated steatotic liver disease (MASLD) and alcohol-related liver disease (ALD). Metabolism Target Organ Damage. 2024;4(4):35. doi:10.20517/mtod.2024.57
6. Huang DQ, El-Serag HB, Loomba R. Global epidemiology of NAFLD-related HCC: trends, predictions, risk factors and prevention. Nat Rev Gastroenterol Hepatol. 2021;18(4):223–238. doi:10.1038/s41575-020-00381-6
7. Younossi ZM, Corey KE, Lim JK. AGA clinical practice update on lifestyle modification using diet and exercise to achieve weight loss in the management of nonalcoholic fatty liver disease: expert review. Gastroenterology. 2021;160(3):912–918. doi:10.1053/j.gastro.2020.11.051
8. Patikorn C, Roubal K, Veettil SK, et al. Intermittent fasting and obesity-related health outcomes: an umbrella review of meta-analyses of randomized clinical trials. JAMA Netw Open. 2021;4(12):e2139558. doi:10.1001/jamanetworkopen.2021.39558
9. Cienfuegos S, Gabel K, Kalam F, et al. Effects of 4- and 6-h Time-Restricted Feeding on Weight and Cardiometabolic Health: a Randomized Controlled Trial in Adults with Obesity. Cell Metab. 2020;32(3):366–378.e363. doi:10.1016/j.cmet.2020.06.018
10. Różański G, Pheby D, newton JL, Murovska M, Zalewski P, Słomko J. Effect of Different Types of Intermittent Fasting on Biochemical and Anthropometric Parameters among Patients with Metabolic-Associated Fatty Liver Disease (MAFLD)-A Systematic Review. Nutrients. 2021;14(1):91. doi:10.3390/nu14010091
11. de Cabo R, Mattson MP. Effects of Intermittent Fasting on Health, Aging, and Disease. N Engl J Med. 2019;381(26):2541–2551. doi:10.1056/NEJMra1905136
12. Lavallee CM, Bruno A, Ma C, Raman M. The Role of Intermittent Fasting in the Management of Nonalcoholic Fatty Liver Disease: a Narrative Review. Nutrients. 2022;14(21):4655. doi:10.3390/nu14214655
13. Harney DJ, Cielesh M, Chu R, et al. Proteomics analysis of adipose depots after intermittent fasting reveals visceral fat preservation mechanisms. Cell Reports. 2021;34(9):108804. doi:10.1016/j.celrep.2021.108804
14. Pantelis AG. Standardizing outcomes in metabolic bariatric surgery - more than meets the eye, less than counts the scale. Metabolism and Target Organ Damage. 2024;4(1):8. doi:10.20517/mtod.2023.55
15. Shao Y, Yuan G, Zhang J, Guo X. Liraglutide reduces lipogenetic signals in visceral adipose of db/db mice with AMPK activation and Akt suppression. Drug Design, Development and Therapy. 2015;9:1177–1184. doi:10.2147/dddt.s79175
16. Nauck MA, Quast DR, Wefers J, Meier JJ. GLP-1 receptor agonists in the treatment of type 2 diabetes - state-of-the-art. Molecular Metabolism. 2021;46:101102. doi:10.1016/j.molmet.2020.101102
17. Lee J, Hong SW, Chae SW, et al. Exendin-4 improves steatohepatitis by increasing Sirt1 expression in high-fat diet-induced obese C57BL/6J mice. PLoS One. 2012;7(2):e31394. doi:10.1371/journal.pone.0031394
18. Mells JE, Fu PP, Sharma S, et al. Glp-1 analog, liraglutide, ameliorates hepatic steatosis and cardiac hypertrophy in C57BL/6J mice fed a Western diet. American Journal of Physiology Gastrointestinal and Liver Physiology. 2012;302(2):G225–235. doi:10.1152/ajpgi.00274.2011
19. Armstrong MJ, Gaunt P, Aithal GP, et al. Liraglutide safety and efficacy in patients with non-alcoholic steatohepatitis (LEAN): a multicentre, double-blind, randomised, placebo-controlled Phase 2 study. Lancet. 2016;387(10019):679–690. doi:10.1016/s0140-6736(15)00803-x
20. Newsome PN, Buchholtz K, Cusi K, et al. A Placebo-Controlled Trial of Subcutaneous Semaglutide in Nonalcoholic Steatohepatitis. N Engl J Med. 2021;384(12):1113–1124. doi:10.1056/NEJMoa2028395
21. Flessa CM, Nasiri-Ansari N, Kyrou I, et al. Genetic and Diet-Induced Animal Models for Non-Alcoholic Fatty Liver Disease (NAFLD) Research. International Journal of Molecular Sciences. 2022;23(24):15791. doi:10.3390/ijms232415791
22. Kleiner DE, Brunt EM, Van Natta M, et al. Design and validation of a histological scoring system for nonalcoholic fatty liver disease. Hepatology. 2005;41(6):1313–1321. doi:10.1002/hep.20701
23. Hung CW, Tholey A. Tandem mass tag protein labeling for top-down identification and quantification. Anal Chem. 2012;84(1):161–170. doi:10.1021/ac202243r
24. Jia X, Peng Y, Ma X, Liu X, Yu K, Wang C. Analysis of metabolic disturbances attributable to sepsis-induced myocardial dysfunction using metabolomics and transcriptomics techniques. Front Mol Biosci. 2022;9:967397. doi:10.3389/fmolb.2022.967397
25. Chen Q, Lou Y. G protein-coupled receptor 39 alleviates mitochondrial dysfunction and hepatocyte lipid accumulation via SIRT1/Nrf2 signaling. J Bioenerg Biomembr. 2023;55(1):33–42. doi:10.1007/s10863-022-09953-4
26. Petersen PS, Jin C, Madsen AN, et al. Deficiency of the GPR39 receptor is associated with obesity and altered adipocyte metabolism. FASEB J. 2011;25(11):3803–3814. doi:10.1096/fj.11-184531
27. Mohs A, Otto T, Schneider KM, et al. Hepatocyte-specific NRF2 activation controls fibrogenesis and carcinogenesis in steatohepatitis. J Hepatol. 2021;74(3):638–648. doi:10.1016/j.jhep.2020.09.037
28. Liu P, Anandhan A, Chen J, et al. Decreased autophagosome biogenesis, reduced NRF2, and enhanced ferroptotic cell death are underlying molecular mechanisms of non-alcoholic fatty liver disease. Redox Biol. 2023;59:102570. doi:10.1016/j.redox.2022.102570
29. Koliaki C, Szendroedi J, Kaul K, et al. Adaptation of hepatic mitochondrial function in humans with non-alcoholic fatty liver is lost in steatohepatitis. Cell Metab. 2015;21(5):739–746. doi:10.1016/j.cmet.2015.04.004
30. Huang D, Xu B, Liu L, et al. TMEM41B acts as an ER scramblase required for lipoprotein biogenesis and lipid homeostasis. Cell Metab. 2021;33(8):1655–1670.e1658. doi:10.1016/j.cmet.2021.05.006
31. Moretti F, Bergman P, Dodgson S, et al. TMEM41B is a novel regulator of autophagy and lipid mobilization. EMBO Reports. 2018;19(9). doi:10.15252/embr.201845889
32. Morita K, Hama Y, Izume T, et al. Genome-wide CRISPR screen identifies TMEM41B as a gene required for autophagosome formation. J Cell Biol. 2018;217(11):3817–3828. doi:10.1083/jcb.201804132
33. Shoemaker CJ, Huang TQ, Weir NR, Polyakov NJ, Schultz SW, Denic V. CRISPR screening using an expanded toolkit of autophagy reporters identifies TMEM41B as a novel autophagy factor. PLoS Biol. 2019;17(4):e2007044. doi:10.1371/journal.pbio.2007044
34. Su W, Wang Y, Jia X, et al. Comparative proteomic study reveals 17β-HSD13 as a pathogenic protein in nonalcoholic fatty liver disease. Proc Natl Acad Sci U S A. 2014;111(31):11437–11442. doi:10.1073/pnas.1410741111
35. Ma Y, Belyaeva OV, Brown PM, et al. 17-Beta Hydroxysteroid Dehydrogenase 13 Is a Hepatic Retinol Dehydrogenase Associated With Histological Features of Nonalcoholic Fatty Liver Disease. Hepatology. 2019;69(4):1504–1519. doi:10.1002/hep.30350
36. Mitsumoto K, Watanabe R, Nakao K, et al. Time-course microarrays reveal early activation of the immune transcriptome in a choline-deficient mouse model of liver injury. Life Sci. 2017;184:103–111. doi:10.1016/j.lfs.2017.07.009
37. Wang MX, Peng ZG. 17β-hydroxysteroid dehydrogenases in the progression of nonalcoholic fatty liver disease. Pharmacology & Therapeutics. 2023;246:108428. doi:10.1016/j.pharmthera.2023.108428
38. Lee C, Jung Y. Sex disparity in the liver regeneration focusing on sex hormones. Metabolism Target Organ Damage. 2023;3(2):10. doi:10.20517/mtod.2023.04
39. Farruggio S, Cocomazzi G, Marotta P, et al. Genistein and 17β-Estradiol Protect Hepatocytes from Fatty Degeneration by Mechanisms Involving Mitochondria, Inflammasome and Kinases Activation. Cell Physiol Biochem. 2020;54(3):401–416. doi:10.33594/000000227
40. Silver HJ, Olson D, Mayfield D, et al. Effect of the glucagon-like peptide-1 receptor agonist liraglutide, compared to caloric restriction, on appetite, dietary intake, body fat distribution and cardiometabolic biomarkers: a randomized trial in adults with obesity and prediabetes. Diabetes, Obesity & Metabolism. 2023;25:2340–2350. doi:10.1111/dom.15113
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Clinical Impact of Semaglutide, a Glucagon-Like Peptide-1 Receptor Agonist, on Obesity Management: A Review
Alorfi NM, Algarni AS
Clinical Pharmacology: Advances and Applications 2022, 14:61-67
Published Date: 3 August 2022
Is Intermittent Fasting Better Than Continuous Energy Restriction for Adults with Overweight and Obesity?
Ye YF, Zhang MX, Lin Z, Tang L
Diabetes, Metabolic Syndrome and Obesity 2022, 15:2813-2826
Published Date: 15 September 2022
Metabolomics Provides Insights into Renoprotective Effects of Semaglutide in Obese Mice
Chen X, Chen S, Ren Q, Niu S, Pan X, Yue L, Li Z, Zhu R, Jia Z, Chen X, Zhen R, Ban J
Drug Design, Development and Therapy 2022, 16:3893-3913
Published Date: 9 November 2022
Alternate-Day High Fat-Normal Chow Diet Ameliorates HFD-Induced Obesity and Restores Intestinal Immunity
Ao DZ, Xu Y, Sun X, Zhang W, Yuan Y
Diabetes, Metabolic Syndrome and Obesity 2022, 15:3843-3853
Published Date: 12 December 2022
Would Combination Be Better: Swimming Exercise and Intermittent Fasting Improve High-Fat Diet-Induced Nonalcoholic Fatty Liver Disease in Obese Rats via the miR-122-5p/SREBP-1c/CPT1A Pathway
Yang K, Liu C, Shao J, Guo L, Wang Q, Meng Z, Jin X, Chen X
Diabetes, Metabolic Syndrome and Obesity 2024, 17:1675-1686
Published Date: 12 April 2024