")
Back to Journals » Biologics: Targets and Therapy » Volume 15
The Impact of NK Cell-Based Therapeutics for the Treatment of Lung Cancer for Biologics: Targets and Therapy
Authors Hamilton G, Plangger A
Received 9 April 2021
Accepted for publication 15 June 2021
Published 7 July 2021 Volume 2021:15 Pages 265—277
DOI https://doi.org/10.2147/BTT.S290305
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Dr Doris Benbrook
Gerhard Hamilton, Adelina Plangger
Department of Vascular Surgery, Medical University of Vienna, Vienna, Austria
Correspondence: Gerhard Hamilton
Department of Vascular Surgery, Medical University of Vienna, Spitalgasse 23, Vienna, A-1090, Austria
Tel +43-1-40400
Email [email protected]
Abstract: Lung cancer has a dismal prognosis and novel targeted therapies leave still room for major improvements and better outcomes. Immunotherapy targeting immune checkpoint (IC) proteins, either as single agents or in combination with chemotherapy, is active but responders constitute only approximately 10– 15% of non-small cell lung cancer (NSCLC) patients. Other effector immune cells such as CAR-T cells or NK cells may help to overcome the limitations of the IC inhibitor therapies for lung cancer. NK cells can kill tumor cells without previous priming and are present in the circulatory system and lymphoid organs. Tissue-residing NK cells differ from peripheral effector cells and, in case of the lung, comprise CD56bright CD16-negative populations showing high cytokine release but low cytotoxicity in contrast to the circulating CD56dim CD16-positive NK cells exhibiting high cytotoxic efficacy. This local attenuation of NK cell killing potency seems due to a specific stage of NK differentiation, immunosuppressive factors as well as presence of myeloid-derived suppressor cells (MDSCs) and regulatory T cells (TREGs). Improved NK cell-based immunotherapies involve IL-2-stimulated effector cells, NK cells expanded with the help of cytokines, permanent NK cell lines, induced pluripotent stem cell-derived NK cells and NK cells armed with chimeric antigen receptors. Compared to CAR T cell therapy, NK cells administration is devoid of graft-versus-host disease (GvHD) and cytokine-release syndrome. Although NK cells are clearly active against lung cancer cells, the low-cytotoxicity differentiation state in lung tumors, the presence of immunosuppressive leucocyte populations, limited infiltration and adverse conditions of the microenvironment need to be overcome. This goal may be achieved in the future using large numbers of activated and armed NK cells as provided by novel methods in NK cell isolation, expansion and stimulation of cytotoxic activity, including combinations with monoclonal antibodies in antibody-dependent cytotoxicity (ADCC). This review discusses the basic characteristics of NK cells and the potential of NK cell preparations in cancer therapy.
Keywords: natural killer cells, immunotherapy, chimeric antigen receptor, antibody-dependent cytotoxicity, lung cancer, interleukins
Background
Lymphoid non-T cells that can kill virally infected and tumor cells were described more than four decades ago and termed natural killer (NK) cells.1 NK cells can attack tumor cells without priming and their activity depends on a range of stimulatory and inhibitory receptors.2,3 NK cells comprise about 5–15% of the human peripheral blood mononuclear cells (PBMCs) and are part of the native immune system that screen cell membranes of autologous cells for a reduced expression of MHC class I molecules and increased expression of cell stress markers.4,5 NK cells mediate the direct and rapid killing of freshly isolated human cancer cells from hematopoietic and solid tumors.6,7 (Figure 1) NK cells in human peripheral blood, bone marrow and various tissues are characterized by the absence of T cell receptors (TCR) and the corresponding CD3 molecules as well as by the expression of neural cell adhesion molecule (NCAM/CD56).8 Human NK cells are generated from multilineage CD34+ hematopoietic progenitors in the bone marrow and their maturation occurs at this site of origin as well as in the lymphoid organs but not in thymus.9 In blood, NK cells show a turnover time of approximately 2 weeks with a doubling within 13.5 days in vivo and in vitro cytokine stimulation of peripheral blood NK cells can result in expansion with a median of 16 (range 11–30) population doublings.10
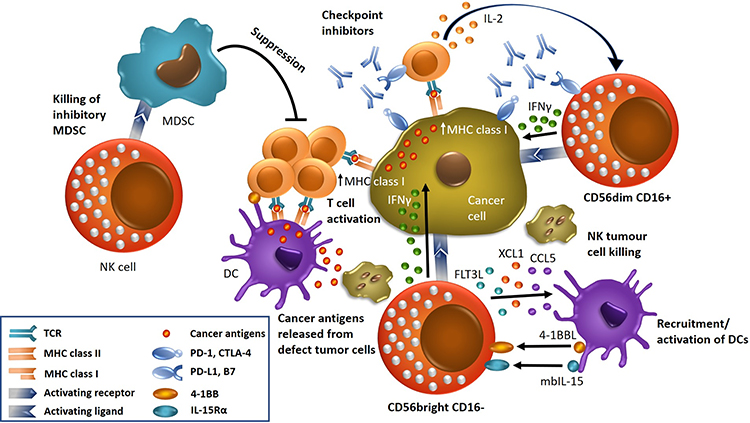 |
Figure 1 NK cells and other immune cells in the tumor microenvironment. NK cells of the CD56dim CD16+ phenotype secrete interferon-γ (IFN-γ), which increases the expression of MHC class I of tumor cells, enhancing the presentation of tumor antigens to T cells. Inhibitory checkpoint molecules expressed by NK cells can be blocked using specific monoclonal antibodies (ICIs). NK cells of the CD56bright CD16- phenotype recruit dendritic cells (DCs) to the tumor microenvironment (TME) and drive their maturation via chemokine ligands CCL5, XCL1 and FMS-related tyrosine kinase 3 ligand (FLT3L). DCs in turn stimulate NK and T cells via membrane-bound IL-15 (mbIL-15) and 4–1BBL secretion. Eventually, NK cells lyse tumor cells resulting in release of cancer antigens, which are then presented by DCs, to provoke specific T cell activation in relation with MHC class I molecules. The immunotherapeutic effect of NK cells includes the removal of immunosuppressive MDSCs. |
NK cells are not only present in peripheral blood, lymph nodes, spleen, and bone marrow but they can also migrate to sites of inflammation in response to distinct chemoattractants. The majority of CD56dim subpopulation of the whole NK cells in peripheral blood (approximately 90%) exhibits high expression of the Fc receptor FcRIII (CD16), killer cell immunoglobulin-like receptors (KIRs) and perforin-mediated cytotoxicity whereas a minor population of CD56bright CD16- KIR- CD94/NKG2A+ (approximately 5–15%) of NK cells is primarily producing cytokines, including IFN-γ and TNF-α11–13 These two NK cell populations have been termed conventional NK cells in contrast to distinct tissue-resident NK cell populations localizing to liver, lymphoid tissue, bone, lung, kidney, gut and uterine tissue as well as distinct adaptive NK cell populations.14 However, CD56 and CD16 are not specific for NK cells and, furthermore, the heterogeneous tissue-resident populations show expression of adhesion molecules and CD69 and may represent an immature NK cell type. Adaptive NK cells are observed in connection with viral infections and exhibit memory cell-like properties. Overall, a wide diversity of receptor expressions of NK cells has been observed and, so far, the function of many of these subpopulations has not been fully characterized.
NK cells can eliminate target cells controlled by signals derived from activating (eg, NCRs or NKG2D) and inhibitory receptors (eg, KIRS or NKG2A).15–17 Normal host cells are protected from NK cells attacks through inhibitory KIRs, that identify the self-MHC class I molecules.15 In particular, the germline-encoded NK receptors include the activating receptors NKG2D, DNAM-1, the natural killing receptors NKp30, NKp44, NKp46, and NKp80, the SLAM-family (Signaling Lymphocyte Activating Molecule) receptors for the elimination of hematopoietic tumor cells and the inhibitory KIRs.18 The activating signaling molecules promote tumor cell killing, cytokine production, immune cell activation, and proliferation and the NKpXX receptors, when engaged, all trigger alterations of the cellular calcium flux and NK cell-mediated killing and secretion of IFN-γ (Figure 1).
The interaction between KIRs and self-MHC molecules governs the maturation of NK cell, a process termed “licensing”.11,19,20 As alternative of MHC downregulation, cancer cells may be recognized by the overexpression of binding molecules for activating NK cell receptors. Ligands for the activating NKG2D receptor, such as MHC class I polypeptide-related sequence A (MICA), MICB and others are presented by cancer cells preferentially in response to cellular stress.21 A separate mechanism known as antibody-dependent cell cytotoxicity (ADCC) results in elimination of antibody-coated cell via the CD16 FcRIII receptor.22
NK cell-mediated lysis of target cells is mainly achieved through the release of the cytotoxic effector perforin and granzymes A and B but NK cells also produce a range of cytokines, both proinflammatory and immunosuppressive, such as IFN-γ, TNF-α and IL10, respectively, as well as growth factors such as granulocyte macrophage colony-stimulating factor (GM-CSF), granulocyte colony-stimulating factor (G-CSF) and IL-3 (Figure 1). CD56dim NK cells can produce very rapidly IFN-γ within 2 to 4 hours after triggering through NKp46 and NKp30 activating receptors (ARs).12,13 NK cell–derived cytokine production impacts dendritic cells, macrophages and neutrophils and empower NK cells to regulate subsequent antigen-specific T and B cell responses. Activated NK cells lose CD16 (FcRIII) and CD62 ligand through the disintegrin and metalloprotease 17 (ADAM17), and inhibition of this protease enhances CD16-mediated NK cell function. Cytokine stimulation also downregulates CD16 and upregulates CD56 expression. Moreover, certain cytokines can greatly enhance the cytotoxicity and cytokine production of the CD162 CD56bright and CD161 CD56dim NK cell subsets, respectively.23,24
In cancer patients, NK cells target cells low/deficient of MHC-class I or bearing “altered-self” stress-inducible proteins.17,25 Besides tumor cell killing through release of perforin and granzyme and secretion of immunoregulatory mediators such as nitric oxide (NO) effects cell death mediated by TNF-family members such as Fas-L or TRAIL. The degree of tumor infiltration of NK cells seems to have prognostic value in gastric carcinoma, colorectal carcinoma and lung carcinomas, thus indicating a protective role of the NK cell infiltrate.26,27 NK cell infiltration of tumors depends on their expression of heparinase.28 NK cells may further attract T cells to the tumor region and elevate inflammatory responses through secretion of cytokines and chemokines.29 Furthermore, NK cells have been suggested to suppress metastasis through elimination of circulating tumor cells (CTCs).30
NK cells seem well suited for anticancer immunotherapy and cells for clinical administration can be isolated from peripheral or umbilical cord blood. Peripheral blood NK cells are prepared by leukapheresis and further enriched by density gradient centrifugation (Figure 2). Subsequently, the combination of T cell depletion with CD56 cell enrichment yields highly purified NK cell populations.31 NK cells gained from peripheral blood of healthy persons are typically in a resting state and can be activated by exposure to IL-2. However, supplementation with IL-2 and infusion to cancer patients has resulted in severe side effects, such as vascular leak syndrome and liver toxicity.32 Studies with native autologous NK cells have yielded disappointing results. The most efficient NK cell expansion was observed with K562 NK target cells co-expressing membrane-bound IL-15 (mbIL-15) and 4–1BBL.31 This technique yields enough NK to provide cells for at least four infusions at 50 million cells/per kg from one leukapheresis product observing GMP conditions.31 However, many mechanisms mediate NK cell suppression in the tumor microenvironment (TME), several of which also impair T cell responses.33,34 In case of NK cells, NKG2D ligand release can occur by shedding and these soluble ligands prevent NK cell-tumor cell interaction and the cytotoxic response.35,36
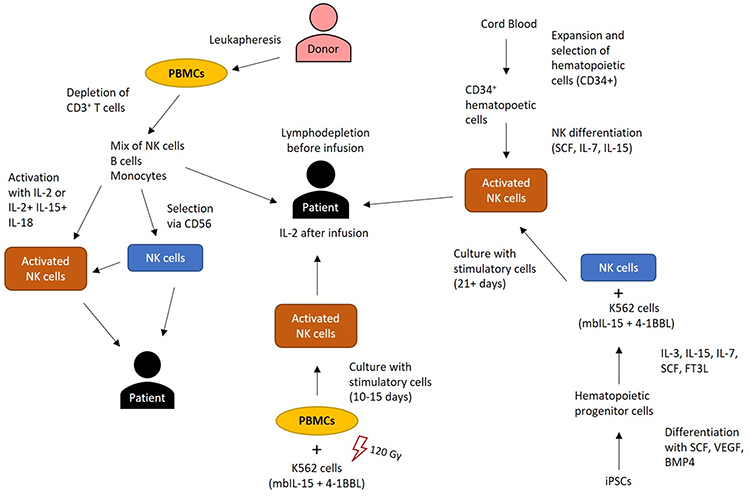 |
Figure 2 Isolation, activation and propagation of allogeneic NK cells. Peripheral blood mononuclear cells (PBMCs) are prepared from healthy donors by leukapheresis. PBMC depletion of CD3+ T cells, prevents GvHD after infusion and further purification is achieved by positive CD56+ cell selection. These cell preparations are infused or activated with IL-2 or a mixture of IL-12, IL-15 and IL-18. Another method for NK cell stimulation involves ex vivo coculture with the K562 cell line expressing membrane-bound IL-15 (mbIL-15) and 4–1BBL that is irradiated to abolish expansion. Umbilical cord blood NK cells can be used similar to peripheral blood NK cells or enriched for CD34+ hematopoietic progenitors, followed by differentiation to NK cells. NK cells can be gained from induced pluripotent stem cells (iPSCs) via successive hematopoietic and NK cell differentiation, followed by stimulation with cells expressing mbIL-21. Before infusion of allogeneic NK cells, patients receive lymphodepleting chemotherapy to facilitate temporary engraftment of the infused NK cells. |
In summary, NK cells are functional in tumor surveillance and can be manipulated by artificial activation techniques to present a highly effective anticancer tool against hematopoietic malignancies and, dependent on successful further rearming and mobilization, against solid tumors in the future.
NK Cells of the Lung
The lungs are frequently challenged by pathogens, environmental damages and tumors and contain a large population of innate immune cells.37,38 Involvement of NK cells in lung diseases, such as cancer, chronic obstructive pulmonary disease (COPD), asthma and infections, has been amply reported.39 Chronic inflammation drives the irreversible obstruction of the lung function in COPD and local NK cells show hyperresponsiveness in COPD and kill autologous lung CD326+ epithelial cells.40 Therefore, targeting NK cells may represent a novel strategy for treating COPD. Furthermore, NK cells from cigarette smoke-exposed mice produce higher levels of IFN-γ upon stimulation with cytokines or toll-like receptor (TLR) ligands.41
Lung NK cells account for approximately 10–20% of local lymphocytes and have migrated to the lungs from bone marrow.42 These cells exhibit the phenotype of the CD56dim CD16+ subset and are located in the parenchyma.43 Lung NK cells show major differences in phenotype and function to those from other tissues and, for example, KIR-positive NK cells and differentiated CD57+ NKG2A− cells are found in higher numbers in the lungs compared to matched peripheral blood.37,38 In vivo, human lung NK cells respond poorly to activation by target cells in comparison to peripheral blood NK cells, most likely due to suppressive effects of alveolar macrophages and soluble factors in the fluid of the lower respiratory tract.44 The presence of hypofunctional NK cells seems to regulate the pulmonary homeostasis in the presence of constantly irritation by environmental and autologous antigens.
Unlike other tissues, the lung NK cell diversity and its acquisition have been very little studied, especially regarding the resident lung populations. Although the majority of lung NK cells are of a non-tissue-resident phenotype, a small CD56bright CD49a+ lung NK cell subset has been found.45 NK cell diversity occurs for the main resident population within the lung, namely CD49a+CD56bright CD16− NK cells that can be split into four different resident subpopulations according to the residency markers CD69 and CD103.47 The CD69+CD103+ subset is the most important as compared to single positive or double negative subsets. The respective significance of these subsets in terms of ontogeny, differentiation, or functionality remains to be characterized.
The CD16− NK cells in the human lung comprises a heterogeneous cell population and the CD69+CD49a+CD103− and CD69+CD49a+CD103+ tissue-resident NK cells are clearly distinct from other NK cell subsets in the lung and other tissues, whereas CD69spCD16− NK cells (lacking expression of CD49a and/or CD103) largely represent conventional CD69−CD16− NK cells.47 Furthermore, lung tissue-resident NK cells are functionally competent and constitute a first line of defense in the human lung. Protein and gene expression signatures of CD16− NK cell subsets correlated with distinct patterns of expression of CD69, CD49a, and CD103 and corroborated the CD69+CD49a+CD103− and CD69+CD49a+CD103+ NK cells as tissue-resident NK cells.48 In contrast, CD69spCD16− NK cells are more similar to CD69−CD16− NK cells and showed lower expression of genes associated with tissue-residency.
On the course of NK cell differentiation less differentiated NK cells are hypofunctional but respond stronger to cytokine stimulation and more differentiated NK cells exert more potent ADCC-dependent cell killing.46,49 The early activation antigen CD69 is expressed on a wide range of tissue-resident lymphocytes, including T cells and NK cells, and promotes retention of the cells in the tissue.38,50 Highly differentiated and hypofunctional CD69+ CD56dim CD161+ NK cells constitute the dominant NK cell population in the human lung. In summary, these results indicate that the human lung is mainly populated by NK cells migrating between lung and blood, rather than by CD69-positive tissue-resident cells. The mechanisms controlling this distribution of the lymphocyte populations is not known but may comprise changes in the homing of NK cells, increased apoptosis of NK cells and increased expansion or recruitment of tissue-resident T cells.
NK Cells and NSCLC
Although the incidence of lung cancer is declining, the survival rates remain poor due to a lack of early detection and only recent progress in targeted cancer therapies that are still only feasible for a limited subpopulation of patients.51,52 The host of immune cells involved in lung cancer include CD4+ and CD8+ T lymphocytes, neutrophils, monocytes, macrophages, innate lymphoid cells (ILCs), dendritic cells and NK cells. In lung cancer patients, peripheral NK cell cytotoxicity and INF-γ production was reported to be reduced.53–56 Especially, a lower cytotoxic activity in NK cells was observed in smokers due to the suppression of the induction of IL-15 and IL-15-mediated NK cell functions in human PBMCs.57 Furthermore, the granzyme B release by NK cells from lung cancer tissue is lower compared to adjacent normal tissue.58 Additionally, peripheral NK cells of NSCLC patients are present in lower cell numbers and display a distinctive receptor expression with downregulation of NKp30, NKp80, CD16, DNAM1, KIR2DL1, and KIR2DL2, but upregulation of NKp44, NKG2A, CD69, and HLA-DR. Furthermore, low levels of IFN-γ and CD107a result in impaired cytotoxicity and promotion of tumor growth.54,59,60 The CD56bright CD16-NK cell subset is highly enriched in the tumor infiltrate and show activation markers, including NKp44, CD69, and HLA-DR.59–61 However, the release of soluble factors by NSCLC tumor cells inhibit the activity of granzyme B and perforin and the induction of IFN-γ in intratumoral NK cells and suggest a local inhibition of NK cells by the NSCLC TME.62 T cell immune checkpoint molecules programmed cell death 1 (PD-1), cytotoxic T lymphocyte antigen 4 (CTLA4), lymphocyte activation gene 3 protein (LAG3) and TIM3 are expressed by subpopulations of NK cells and might reduce NK antitumor responses. In solid tumors, vascular supply may be ineffective causing hypoxia and low nutrient levels in the TME that may impair NK cell metabolism and antitumor cytotoxicity as demonstrated in lung experimental animal models.63,64 Additionally, the CD56bright CD16- NK cells enhance protumor neoangiogenesis through secretion of VEGF, placental growth factor and IL-8/CXCL8.65
NK Cells and SCLC
Small cell lung cancer (SCLC) is a pulmonary neuroendocrine cancer linked to smoking that has a dismal prognosis and invariably develops resistance to chemotherapy within a short time.66 Despite a high tumor mutational burden, immune checkpoint inhibitors show minor prolongation of survival in SCLC patients.66,67 In particular, Nivolumab (anti-PD1 antibody) was approved for third-line treatment and the combination of atezolizumab (anti-PDL1 antibody) with carboplatin and etoposide was approved for first-line treatment of disseminated SCLC, resulting in minor survival gains.68,69 NK cells are critical in suppressing lung tumor growth and while low MHC expression would make SCLC resistant to adaptive immunity, this should make SCLCs susceptible to NK cell killing.64,70 In comparison to the peripheral blood NK cells of healthy individuals, the NK cells of SCLC patients are present in equal cell counts but exhibit lower cytotoxic activity, downregulation of NKp46 and perforin expression.55 Lack of effective NK surveillance seems to contribute to SCLC progress, primarily through the reduction of NK-activating ligands (NKG2DL). SCLC primary tumors possess very low levels of NKG2DL mRNA and SCLC lines largely fail to express NKG2DL at the protein level.66,71 Accordingly, restoring NKG2DL in experimental models suppressed tumor growth and metastasis in a NK cell-dependent manner. Furthermore, histone deacetylase (HDAC) inhibitors induced NKG2DL re-expression and resulted in tumor suppression by NK and T cells. Actually, SCLC and neuroblastoma are the two tumor types with lowest NKG2DL-expression. In conclusion, epigenetic silencing of NKG2DL results in a defect of NK cell activation and immune escape of SCLC and neuroblastoma. Poor immune infiltrates in SCLC tumors combined with reduced NK and T cell recognition of the tumor cells seem to contribute to immune resistance of SCLCs.72
Suppression of the Activity of Lung NK Cells
A majority of NSCLC patients do not benefit from the current IC-directed immunotherapy. CD56dim CD16+ NK cells comprise the majority of NK cells in human lungs and express KIRs and a more differentiated phenotype compared with NK cells in the peripheral blood.38,73 However, human lung NK cells were hyporesponsive toward target cell stimulation, irrespective of priming with IFN-α. NK cells are activated by MICA and MICB expressed by stressed tumor cells and are recognized by NK cell receptors NKG2D.74 Preclinical studies show that NKG2A or TIGIT blockade enhances antitumor immunity mediated by NK cells.2 However, the poor infiltration of NK cells into solid tumors, alterations in activating/inhibitory signals and adverse TME conditions decrease the NK-mediated killing. NK cells can be inactivated by different cells such as Tregs and MDSCs but also by soluble mediators such as adenosine.75,76 Adenosine represents one of the most potent immunosuppressive factors in solid tumors that is produced in the tumor stroma by degradation of extracellular ATP.77–79 ATP and ADP are degraded by membrane-expressed ectonucleotidases such as CD39 and enhance the influx and the suppressive capacity of Tregs and MDSCs in solid tumors. NK cells are strongly involved in eliminating circulating tumor cells (CTCs), but their activity can be inhibited by soluble factors, such as TGF-β derived from M2 macrophages.80,81 One approach uses cytokines to selectively boost both the number as well as the efficacy of anti-tumor functions of peripheral NK cells.82 The gene signature of NK cell dysfunction in human NSCLC revealed an altered migratory behavior with downregulation of the sphingosine-1-phosphate receptor 1 (S1PR1) and CX3C chemokine receptor 1 (CX3CR1).83 Additionally, the expression of the immune inhibitory molecules CTLA-4 and killer cell lectin like receptor (KLRC1) were elevated in intratumoral NK cells and CTLA-4 blockade could partially restore the impaired MHC class II expression on dendritic cell (DC). In summary, the intratumoral NK dysfunction can be attributed to direct crosstalk between tumor and NK cells, activated platelets and soluble factors, such as TGF-β, prostaglandin E2, indoleamine-2,3-dioxygenase, adenosine and IL-10.19,26,54,83 In addition, a specific migratory signature could explain the exclusion of NK cells from the tumor interior. NK cells in NSCLC distribute to the intratumoral fibrous septa and to the borders between tumor cells and surrounding stroma.54,59 It has been suggested that a barrier of extracellular matrix proteins may be responsible for the restriction of NK cells primarily to the tumor stroma, such preventing direct NK cell–tumor cell interactions.84,85 In contradiction, ultrastructural investigations demonstrated NK cells are rather flexible and capable of extravasation and intratumoral migration.59 CD56bright CD162+ NK cells express CCR5 that is known to mediate the chemoattraction of specific leukocyte subtypes and explain their accumulation in tumor tissues.13 Infiltration of the tumors by NK cells was reported to be linked with a favorable prognosis in lung cancer.26,86 However, Platonova et al reported that NK cell infiltration lacks any correlation with clinical outcomes in NSCLC.47,54 The poor prognostic significance of NK cells in NSCLC seems to be associated with the intratumoral NK cell dysfunction in patients with intermediate or advanced-stage tumors.
Stimulation of NK Cell Migration and Cytotoxic Activity
It would be of great importance to target chemokine receptors on NK cells to enable them to enter tumor tissues. NK cells acquire inhibitory functions within the TME, the reversion of which will enable NK cells to activate other immune cells and exert antitumor cytotoxic functions.87 In addition, several clinical trials based on NK cell checkpoints are ongoing, targeting KIR, TIGIT, lymphocyte-activation gene 3, TIM3 and KLRC1.88 NK cell dysfunction favors tumor progress and restoring NK cell functions would represent an important potential strategy to inhibit lung cancer. These approaches include the activation of NK cells by exposing to interleukins such as IL-2, IL-12, IL-15, IL-18, the blockade of inhibitory receptors of NK cells by targeting NKG2A, KIR2DL1 and KIR2DL2 as well as the enhancement of NK cell glycolysis by inhibition of fructose-1,6-bisphosphatase 1 and altering the immunosuppressive TME by neutralization of TGF-β.37,53 Pilot clinical trials of NK cell-based therapies such as administration of cytokines, NK-92 cell lines and allogenic NK cell immunotherapy showed promising outcomes on the lung cancer survival with less adverse effects. However, due to the lack of larger clinical trials, the NK cell targeting strategy has not been approved for lung cancer treatment so far.
Most of studies regarding NK cell-based immunotherapy have been performed in hematologic malignancies. However, there are increasingly data available that show that NK cells can selectively recognize and kill cancer stem cells in solid tumors.89 Furthermore, Kim et al showed the essential role of NK cells in prevention of lung metastasis.90 Additionally, Zhang et al studied the efficacy of adaptive transfer of NK and cytotoxic T-lymphocytes mixed effector cells in NSCLC patients.91 A prolonged overall survival was detectable in patients after administration of NK cell-based immunotherapy. In a trial of Lin et al, the clinical outcomes of cryosurgery combined with allogenic NK cell immunotherapy for the treatment of advanced NSCLC were improved with elevated immune functions and quality of life.92
The efficacy of NK cell-based adoptive immunotherapy was also investigated in SCLC patients. Ding et al studied the efficacy and safety of cellular immunotherapy with autologous NK, γδT cells and cytokine-induced killer cells as maintenance therapy for 29 SCLC patients and demonstrated an increased survival of the patients.93 Importantly, lung cancer-infiltrating NK cells can mainly function as producers of relevant cytokines, either beneficial or detrimental for the antitumor immune response, and activation can transform CD56bright CD162+ KIR2+ NK cells into CD56dim CD161+ KIR1+ NK cells with higher cytotoxic activity.94 The switch from a CD56bright phenotype to a CD56dim NK cell signature can take place in lymph nodes during inflammation and these cells circulate into peripheral blood as KIR+CD16+ NK cells with low cytotoxic ability. However, the secondary lymphoid organ (SLO) NK cells acquire cytotoxic activity upon stimulation with IL-2. Malignant NSCLC tumor areas show high presence of Tregs and minor NK cell infiltration, whereas non-malignant regions were oppositely populated, containing NK cells with marked cytotoxicity ex vivo.95 IL-2 activation of PMBCs exhibit increased cytotoxic activity against primary lung cancer cells, that is further elevated by IL-12 treatment.96 The adoptive transfer of NK cells is a therapeutic strategy currently being investigated in various cancer types. For example, Krause et al treated a NSCLC patient and 11 colorectal cancer patients with autologous transfer of NK cells activated ex vivo by a peptide derived from heat shock protein 70 (Hsp70) plus low-dose IL-2.97 The NK cell reinfusion revealed minor adverse effects and yielded promising immunological alterations.
Adaptive-like CD56dim CD16+ NK cells that were found in studies in mice and humans in peripheral blood have a distinctive phenotypic and functional profile compared to conventional NK cells.31,98 These cells have a high target cell responsiveness, as well as a longer life time and a recall potential comparable to that of memory T cells.99 Whereas adoptive NK cell transfer showed promising activities in the treatment of hematological malignancies, elimination of solid tumor cells failed due to insufficient migration and tumor infiltration.100 Furthermore, a CD49a+ KIR+ NKG2C+ CD56bright CD16− adaptive NK cell population with features of residency exists in human lung, that is distinct from adaptive-like CD56dim CD16+ peripheral blood NK cells.43 NK cells with an adaptive-like CD49a+ NK cell expansion in the lung proved to be hyperresponsive toward cancer cells. Despite their in vivo priming, the presence of adaptive-like CD49a+ NK cells in the lung did not correlate with any clinical parameters.
Immune Checkpoints and NK Cells
At the time of diagnosis, the majority (80%) of lung cancer patients present with locally advanced or metastatic disease that continues to progress despite chemotherapy.101 Lung cancer remains the leading cause of cancer death worldwide despite the responses found for immune checkpoint inhibitors (ICIs), including programmed death receptor-1 (PD1) or PD ligand 1 (PDL1)-blockade therapy.102 These ICIs has achieved marked tumor regression in some patients with advanced PD1/PDL1-positive lung cancer; however, lasting responses were limited to a 15% subpopulation of patients.103 IFN-γ, released by cytotoxic NK and T cells, is a critical enhancer of PDL1 expression on tumors and a predictor of response to immunotherapies.104 The high failure rate of immunotherapy seems to be a consequence of low tumor PDL1 expression and the action of further immunosuppressive mechanisms in the TME.105
NK cells expanded from induced-pluripotent stem cells (iPSCs) increased PDL1 expression of tumor cell lines, sensitized non-responding tumors from patients with lung cancer to PD1-targeted immunotherapy and killed PDL1- patient tumors (Figure 2).102 In contrast, native NK cells, that are susceptible to immunosuppression in the TME, had no effect on tumor PDL1 expression. Accordingly, only combined treatment of expanded NK cells and PD1-directed inhibitors resulted in synergistic tumor cell kill of initially non-responding patient tumors. A randomized control trial in patients with PDL1+ NSCLC found that the combination treatment of NK cells with the PD1 inhibitor pembrolizumab was well-tolerated and improved overall and progression-free survival in patients compared single agent pembrolizumab treatment.106 Importantly, during this clinical study no adverse events associated with the administration of NK cells were detected.
Future NK Cell Immunotherapies
Early trials of autologous NK cell therapy from leukapheresis have demonstrated potency against several metastatic cancers but patients developed vascular leak syndrome due to a high level of IL-2.32,107 In contrast, other studies reported that these autologous NK cells failed to demonstrate clinical responses or efficacy at large.108,109 Adoptive transfer of ex vivo IL-2 activated NK cells showing better outcomes than the systemic administration of IL-2.107,110 The development of novel NK cell-mediated immunotherapies presumes a rich source of suitable NK cells for adoptive transfer and an enhancement of the NK cell cytotoxicity and durability in vivo. Potential sources comprise haploidentical NK cells, umbilical cord blood NK cells, stem cell-derived NK cells, permanent NK cell lines, adaptive NK cells, cytokine-induced memory-like NK cells and chimeric antigen receptor (CAR) NK cells (Figure 2). Augmentation of the cytotoxicity and persistence of NK cells under clinical investigation is promoted by cytokine-based agents, NK cell engager molecules and ICIs.111,112 Despite some successes, most patients failed to respond to unmodified NK cell-based immunotherapy.113
Clonal NK cell lines, such as NK-92, KHYG-1 and YT cells, are an alternative source of allogeneic NK cells, and the NK-92 cell line has been extensively tested in clinical trials.114–116 NK-92 cells are easily expanded with doubling times between 24 and 36 hours.115 NK-92 has received FDA approval for trials in patients with solid tumors.116 These cells are genetically unstable, which requires them to be irradiated prior to infusion. Irradiated NK-92 cells have been observed to kill tumor cells in patients with cancer, although irradiation limits the in vivo persistence of these cells to a maximum of 48 hours.117 The results are still short of a significant clinical benefit.118 An NK-92- derived product (haNK) has been engineered to express a high-affinity variant of CD16 as well as endogenous IL-2 in order to enhance effector function (Figure 2).119–121 For example, Dinutuximab is a product of human-mouse chimeric mAb (ch14.18 mAb), which has demonstrated high efficacy against GD2-positive neuroblastoma cells in vitro and melanoma cells in vivo.122 In MHC-I expressing tumor cells, the effector functions of autologous NK cells are often inhibited by KIR that can be blocked with the help of anti-KIR (IPH2101).123 Stem cell-derived NK cell products from multiple sources are currently being tested clinically, including those originating from umbilical cord blood stem cells or iPSCs.124,125 NK cells account for ~5–15% of all lymphocytes in peripheral blood, whereas they constitute up to 30% of the lymphocytes in umbilical cord blood.126 iPSC-derived NK cells were triple gene- modified to express cleavage-resistant CD16, a chimeric antigen receptor (CAR) targeting CD19 and a membrane-bound IL-15 receptor signaling complex in order to promote their persistence.127 Thus, investigations to provide highly active modified NK cells in numbers sufficient for clinical application are actively pursued.
CAR NK Cells
CAR T cells are derived from autologous T cells and genetically engineered to express an antibody single-chain variable fragment (scFv) targeting a tumor-associated antigen.128 CAR T cell therapies achieved objective response rates of >80% in patients with acute lymphocytic leukemia (ALL) and B cell non-Hodgkin lymphoma.129–131 However, the drawbacks of CAR T therapy include severe adverse events such as GvHD, cytokine-release syndrome and neurological toxicities, besides inefficiencies of T cell isolation, modification and expansion as well as exorbitant costs.132 CAR NK therapy is expected to circumvent some of these problems, including the high toxicities. Primary NK cells are not ideal sources for the generation of CAR cell products, due to difficulties in cell isolation, transduction and expansion. However, NK cell expansion could be greatly improved by involvement of a K562 leukemia cell line feeder modified to express membrane-bound IL-15 (mbIL-15; Figure 2).133 Denman et al improved this method adding membrane-bound 4–1BBL to the K562 cell line resulting in a high expansion of NK cells within a short time.134,135 Nevertheless, current clinical trials of CAR NK cells rely mainly on processing of stem cell-derived or progenitor NK cells.136 Genetic engineering of NK cells has been performed by viral transduction or electroporation of mRNA.3 Many clinical trials of CAR NK-92 cells are ongoing, but the requirement for irradiation and resulting short persistence are limitations to the clinical efficacy of these products. NK92-CD16 cells preferentially killed tyrosine kinase inhibitor (TKI)-resistant NSCLC cells when compared with their parental NSCLC cells.137 Moreover, NK92-CD16 cell-induced cytotoxicity against TKI-resistant NSCLC cells was increased in the presence of cetuximab, an EGFR-targeting monoclonal antibody. A number of Phase I trials of CAR NK cells from various sources, including autologous peripheral blood NK cells, umbilical cord blood NK cells, NK-92 cells and iPSCs were designed to target diverse cancers, such as ALL, B cell malignancies, NSCLC, ovarian cancer or glioblastoma, and are currently active.
CAR NK cells derived from iPSCs, such as the triple-gene-modified constructions are described as a promising alternative. For example, a tri-specific killer engager (TriKE) consists of two scFvs, one targeting CD16 on NK cells and the other targeting CD33 on AML cells, linked by an IL-15 domain that promotes NK cell survival and proliferation.138 Controlled clinical trials with larger patient cohorts are required to validate these early results. Immunosuppressive factors of the TME, such as low glucose, hypoxia and MDSCs, Treg cells and tumor associated macrophages (TAMs) still suppress the antitumor functions of CAR-NK cells. Low efficiency of CAR-transduction, limited cell expansion and the scarcity of suitable targets impede the use of CAR-NK therapy despite of reports of therapeutic efficacy and safety.139
The cytokine gene transfer approaches, including interleukins and stem cell factor (SCF), have been shown to induce NK cell proliferation and increases survival capacity in vivo.140 The use of primary CAR-NK and CAR-NK lines in hematological tumors showed high specificity and cytotoxicity toward the target cells.141,142 So far, only a few clinical trial studies of CAR-NK have been registered on ClinicalTrials.gov.143 The combination of blocking ICIs on CAR-NK cells can lead to a highly efficient cancer-redirected cytotoxic activity.144,145 However, hematological cancers are responsible for only 6% of all cancer deaths and solid tumor are much more difficult to target by NK/CAR NK-based immunotherapy.146
Conclusion
Both the unmodified and the engineered forms of NK cell treatment are showing promise in pilot clinical trials in patients with cancer.147 This kind of immunotherapy seems to combine efficacy, safety, and relative ease of effector cell supply. The lung is populated by NK cells at a specific differentiation stage releasing cytokines but exhibiting low cytotoxicity. Poor tumor infiltration, immunosuppressive factors and cell types as well as hypoxic conditions in the TME limit the activity of NK cells. Therefore, larger numbers of activated, cytotoxic competent and armed NK cells will be required for successful therapy.
Acknowledgments
We wish to thank B. Rath for help in the preparation of the manuscript and T. Hohenheim for enduring endorsement.
Disclosure
The authors report no conflicts of interest in this work.
References
1. Herberman RB, Nunn ME, Lavrin DH. Natural cytotoxic reactivity of mouse lymphoid cells against syngeneic acid allogeneic tumors. I. Distribution of reactivity and specificity. Int J Cancer. 1975;16:216–229. doi:10.1002/ijc.2910160204
2. Chiossone L, Dumas PY, Vienne M, Vivier E. Natural killer cells and other innate lymphoid cells in cancer. Nat Rev Immunol. 2018;18:671–688. doi:10.1038/s41577-018-0061-z
3. Shimasaki N, Coustan-Smith E, Kamiya T, Campana D. Expanded and armed natural killer cells for cancer treatment. Cytotherapy. 2016;18(11):1422–1434. doi:10.1016/j.jcyt.2016.06.013
4. Caligiuri MA. Human natural killer cells. Blood. 2008;112(3):461–469. doi:10.1182/blood-2007-09-077438
5. Waldhauer I, Steinle A. NK cells and cancer immunosurveillance. Oncogene. 2008;27(45):5932–5943. doi:10.1038/onc.2008.267
6. Carlsten M, Malmberg KJ, Ljunggren HG. Natural killer cell-mediated lysis of freshly isolated human tumor cells. Int J Cancer. 2009;124(4):757–762. doi:10.1002/ijc.24082
7. Li L, Li W, Wang C, et al. Adoptive transfer of natural killer cells in combination with chemotherapy improves outcomes of patients with locally advanced colon carcinoma. Cytotherapy. 2018;20(1):134–148. doi:10.1016/j.jcyt.2017.09.009
8. Lanier LL, Testi R, Bindl J, Phillips JH. Identity of Leu-19 (CD56) leukocyte differentiation antigen and neural cell adhesion molecule. J Exp Med. 1989;169:2233–2238. doi:10.1084/jem.169.6.2233
9. Yu J, Freud AG, Caligiuri MA. Location and cellular stages of natural killer cell development. Trends Immunol. 2013;34(12):573–582. doi:10.1016/j.it.2013.07.005
10. Fujisaki H, Kakuda H, Shimasaki N, et al. Expansion of highly cytotoxic human natural killer cells for cancer cell therapy. Cancer Res. 2009;69(9):4010–4017. doi:10.1158/0008-5472.CAN-08-3712
11. Raulet DH, Guerra N. Oncogenic stress sensed by the immune system: role of natural killer cell receptors. Nat Rev Immunol. 2009;9(8):568–580. doi:10.1038/nri2604
12. Poli A, Michel T, Theresine M, Andres E, Hentges F, Zimmer J. CD56bright natural killer (NK) cells: an important NK cell subset. Immunology. 2009;126(4):458–465. doi:10.1111/j.1365-2567.2008.03027.x
13. Campbell JJ, Qin S, Unutmaz D, et al. Unique subpopulations of CD561 NK and NK-T peripheral blood lymphocytes identified by chemokine receptor expression repertoire. J Immunol. 2001;166(11):6477–6482. doi:10.4049/jimmunol.166.11.6477
14. Freud AG, Mundy-Bosse BL, Yu J, Caligiuri MA. The broad spectrum of human natural killer cell diversity. Immunity. 2017;47(5):820–833. doi:10.1016/j.immuni.2017.10.008
15. Campbell KS, Hasegawa J. Natural killer cell biology: an update and future directions. J Allergy Clin Immunol. 2013;132(3):536–544. doi:10.1016/j.jaci.2013.07.006
16. Kärre K, Ljunggren HG, Piontek G, Kiessling R. Selective rejection of H-2-deficient lymphoma variants suggests alternative immune defence strategy. Nature. 1986;319(6055):675–678. doi:10.1038/319675a0
17. Kärre K. NK cells, MHC class I molecules and the missing self. Scand J Immunol. 2002;55(3):221–228. doi:10.1046/j.1365-3083.2002.01053.x
18. Marcus A, Gowen BG, Thompson TW, et al. Recognition of tumors by the innate immune system and natural killer cells. Adv Immunol. 2014;122:91–128. doi:10.1016/B978-0-12-800267-4.00003-1
19. Morvan MG, Lanier LL. NK cells and cancer: you can teach innate cells new tricks. Nat Rev Cancer. 2016;16(1):7–19. doi:10.1038/nrc.2015.5
20. Elliott JM, Yokoyama WM. Unifying concepts of MHC-dependent natural killer cell education. Trends Immunol. 2011;32(8):364–372. doi:10.1016/j.it.2011.06.001
21. Bauer S, Groh V, Wu J, et al. Activation of NK cells and T cells by NKG2D, a receptor for stress-inducible MICA. Science. 1999;285(5428):727–729. doi:10.1126/science.285.5428.727
22. Ferris RL, Jaffee EM, Ferrone S. Tumor antigen-targeted, monoclonal antibody-based immunotherapy: clinical response, cellular immunity, and immunoescape. J Clin Oncol. 2010;28:4390–4399. doi:10.1200/JCO.2009.27.6360
23. Vivier E, Raulet DH, Moretta A, et al. Innate or adaptive immunity? The example of natural killer cells. Science. 2011;331(6013):44–49. doi:10.1126/science.1198687
24. Deniz G, van de Veen W, Akdis M. Natural killer cells in patients with allergic diseases. J Allergy Clin Immunol. 2013;132(3):527–535. doi:10.1016/j.jaci.2013.07.030
25. Ingram Z, Madan S, Merchant J, et al. Targeting natural killer T cells in solid malignancies. Cells. 2021;10(6):1329. doi:10.3390/cells10061329
26. Villegas FR, Coca S, Villarrubia VG, et al. Prognostic significance of tumor infiltrating natural killer cells subset CD57 in patients with squamous cell lung cancer. Lung Cancer. 2002;35(1):23–28. doi:10.1016/s0169-5002(01)00292-6
27. Takanami I, Takeuchi K, Giga M. The prognostic value of natural killer cell infiltration in resected pulmonary adenocarcinoma. J Thorac Cardiovasc Surg. 2001;121(6):1058–1063. doi:10.1067/mtc.2001.113026
28. Putz EM, Mayfosh AJ, Kos K, et al. NK cell heparanase controls tumor invasion and immune surveillance. J Clin Invest. 2017;127(7):2777–2788. doi:10.1172/JCI92958
29. Malmberg KJ, Carlsten M, Björklund A, Sohlberg E, Bryceson YT, Ljunggren HG. Natural killer cell-mediated immunosurveillance of human cancer. Semin Immunol. 2017;31:20–29. doi:10.1016/j.smim.2017.08.002
30. López-Soto A, Gonzalez S, Smyth MJ, Galluzzi L. Control of metastasis by NK cells. Cancer Cell. 2017;32(2):135–154. doi:10.1016/j.ccell.2017.06.009
31. Lee DA, Verneris MR, Campana D. Acquisition, preparation, and functional assessment of human NK cells for adoptive immunotherapy. Methods Mol Biol. 2010;651:61–77. doi:10.1007/978-1-60761-786-0_4
32. Rosenberg SA, Lotze MT, Muul LM, et al. Observations on the systemic administration of autologous lymphokine-activated killer cells and recombinant interleukin-2 to patients with metastatic cancer. N Engl J Med. 1985;313(23):1485–1492. doi:10.1056/NEJM198512053132327
33. Shimasaki N, Coustan-Smith E, Kamiya T, Campana D. Mechanisms of resistance to NK cell immunotherapy. Cancers. 2020;12(4):893. doi:10.3390/cancers12040893
34. Guillerey C, Huntington ND, Smyth MJ. Targeting natural killer cells in cancer immunotherapy. Nat Immunol. 2016;17(9):1025–1036. doi:10.1038/ni.3518
35. Chitadze G, Bhat J, Lettau M, Janssen O, Kabelitz D. Generation of soluble NKG2D ligands: proteolytic cleavage, exosome secretion and functional implications. Scand J Immunol. 2013;78(2):120–129. doi:10.1111/sji.12072
36. Zingoni A, Vulpis E, Nardone I, et al. Targeting NKG2D and NKp30 ligands shedding to improve NK cell-based immunotherapy. Crit Rev Immunol. 2016;36(6):445–460. doi:10.1615/CritRevImmunol.2017020166
37. Cong J, Wei H. Natural killer cells in the lungs. Front Immunol. 2019;10:1416. doi:10.3389/fimmu.2019.01416
38. Marquardt N, Kekäläinen E, Chen P, et al. Human lung natural killer cells are predominantly comprised of highly differentiated hypofunctional CD69−CD56dim cells. J Allergy Clin Immunol. 2017;139(4):1321–1330.e4. doi:10.1016/j.jaci.2016.07.043
39. Culley FJ. Natural killer cells in infection and inflammation of the lung. Immunology. 2009;128(2):151–163. doi:10.1111/j.1365-2567.2009.03167.x
40. Barnes PJ. Immunology of asthma and chronic obstructive pulmonary disease. Nat Rev Immunol. 2008;8:183–192. doi:10.1038/nri2254
41. Freeman CM, Stolberg VR, Crudgington S, et al. Human CD56+ cytotoxic lung lymphocytes kill autologous lung cells in chronic obstructive pulmonary disease. PLoS One. 2014;9(7):e103840. doi:10.1371/journal.pone.0103840
42. Trinchieri G. Biology of natural killer cells. Adv Immunol. 1989;47:187–376. doi:10.1016/S0065-2776(08)60664-1
43. Marquardt N, Beziat V, Nyström S, et al. Cutting edge: identification and characterization of human intrahepatic CD49a1 NK cells. J Immunol. 2015;194(6):2467–2471. doi:10.4049/jimmunol.1402756
44. Robinson BW, Pinkston P, Crystal RG. Natural killer cells are present in the normal human lung but are functionally impotent. J Clin Invest. 1984;74:942–950. doi:10.1172/JCI111513
45. Cooper GE, Ostridge K, Khakoo SI, Wilkinson TMA, Staples KJ. Human CD49a+ lung natural killer cell cytotoxicity in response to influenza A virus. Front Immunol. 2018;9:1671. doi:10.3389/fimmu.2018.01671
46. Björkström N, Riese P, Heuts F, et al. Expression patterns of NKG2A, KIR, and CD57 define a process of CD56dim NK-cell differentiation uncoupled from NK-cell education. Blood. 2010;116(19):3853–3864. doi:10.1182/blood-2010-04-281675
47. Hervier B, Russick J, Cremer I, Vieillard V. NK cells in the human lungs. Front Immunol. 2019;10:1263. doi:10.3389/fimmu.2019.01263
48. Marquardt N, Kekäläinen E, Chen P, et al. Unique transcriptional and protein-expression signature in human lung tissue-resident NK cells. Nat Commun. 2019;10(1):3841. doi:10.1038/s41467-019-11632-9
49. Lopez-Verges S, Milush J, Pandey S, et al. CD57 defines a functionally distinct population of mature NK cells in the human CD56dimCD161 NK-cell subset. Blood. 2010;116:3865–3874. doi:10.1182/blood-2010-04-282301
50. Sathaliyawala T, Kubota M, Yudanin N, et al. Distribution and compartmentalization of human circulating and tissue-resident memory T cell subsets. Immunity. 2013;38:187–197. doi:10.1016/j.immuni.2012.09.020
51. Nasim F, Sabath BF, Eapen GA. Lung cancer. Med Clin North Am. 2019;103(3):463–473. doi:10.1016/j.mcna.2018.12.006
52. Nguyen AH, Berim IG, Agrawal DK. Cellular and molecular immunology of lung cancer: therapeutic implications. Expert Rev Clin Immunol. 2014;10(12):1711–1730. doi:10.1586/1744666X.2014.975692
53. Aktaş ON, Öztürk AB, Erman B, Erus S, Tanju S, Dilege Ş. Role of natural killer cells in lung cancer. J Cancer Res Clin Oncol. 2018;144(6):997–1003. doi:10.1007/s00432-018-2635-3
54. Platonova S, Cherfils-Vicini J, Damotte D, et al. Profound coordinated alterations of intratumoral NK cell phenotype and function in lung carcinoma. Cancer Res. 2011;71(16):5412–5422. doi:10.1158/0008-5472.CAN-10-4179
55. Al Omar SY, Marshall E, Middleton D, Christmas SE. Increased killer immunoglobulin-like receptor expression and functional defects in natural killer cells in lung cancer. Immunology. 2011;133(1):94–104. doi:10.1111/j.1365-2567.2011.03415.x
56. Yang L, Wang L, Zhang Y. Immunotherapy for lung cancer: advances and prospects. Am J Clin Exp Immunol. 2016;5(1):1–20.
57. Mian MF, Lauzon NM, Stampfli MR, Mossman KL, Ashkar AA. Impairment of human NK cell cytotoxic activity and cytokine release by cigarette smoke. J Leukoc Biol. 2008;83(3):774–784. doi:10.1189/jlb.0707481
58. Castriconi R, Cantoni C, Della Chiesa M, et al. Transforming growth factor beta 1 inhibits expression of NKp30 and NKG2D receptors: consequences for the NK-mediated killing of dendritic cells. Proc Natl Acad Sci USA. 2003;100(7):4120–4125. doi:10.1073/pnas.0730640100
59. Carrega P, Morandi B, Costa R, et al. Natural killer cells infiltrating human non-small-cell lung cancer are enriched in CD56 bright CD16(−) cells and display an impaired capability to kill tumor cells. Cancer. 2008;112(4):863–875. doi:10.1002/cncr.23239
60. Lavin Y, Kobayashi S, Leader A, et al. Innate immune landscape in early lung adenocarcinoma by paired single-cell analyses. Cell. 2017;169:750–765.e17. doi:10.1016/j.cell.2017.04.014
61. Björkström NK, Ljunggren HG, Michaëlsson J. Emerging insights into natural killer cells in human peripheral tissues. Nat Rev Immunol. 2016;16(5):310–320. doi:10.1038/nri.2016.34
62. Hodge G, Barnawi J, Jurisevic C, et al. Lung cancer is associated with decreased expression of perforin, granzyme B and IFNγ by infiltrating lung tissue T cells, NKT-like and NK cells. Clin Exp Immunol. 2014;178(1):79–85. doi:10.1111/cei.12392
63. O’Brien KL, Finlay DK. Immunometabolism and natural killer cell responses. Nat Rev Immunol. 2019;19(5):282–290. doi:10.1038/s41577-019-0139-2
64. Cong J, Wang X, Zheng X, et al. Dysfunction of natural killer cells by FBP1-induced inhibition of glycolysis during lung cancer progression. Cell Metab. 2018;28(2):243–255.e5. doi:10.1016/j.cmet.2018.06.021
65. Bruno A, Focaccetti C, Pagani A, et al. The proangiogenic phenotype of natural killer cells in patients with non – small cell lung cancer. Neoplasia. 2013;15:133–142. doi:10.1593/neo.121758
66. Hamilton G, Rath B. Immunotherapy for small cell lung cancer: mechanisms of resistance. Expert Opin Biol Ther. 2019;19(5):423–432. doi:10.1080/14712598.2019.1592155
67. Zhu M, Huang Y, Bender ME, et al. Evasion of innate immunity contributes to small cell lung cancer progression and metastasis. Cancer Res. 2021;81(7):1813–1826. doi:10.1158/0008-5472.can-20-2808
68. Regzedmaa O, Zhang H, Liu H, Chen J. Immune checkpoint inhibitors for small cell lung cancer: opportunities and challenges. Onco Targets Ther. 2019;12:4605–4620. doi:10.2147/OTT.S204577
69. Pakkala S, Owonikoko TK. Immune checkpoint inhibitors in small cell lung cancer. J Thorac Dis. 2018;10(Suppl 3):S460–S467. doi:10.21037/jtd.2017.12.51
70. Vivier E, Ugolini S. Natural killer cells: from basic research to treatments. Front Immunol. 2011;2:18. doi:10.3389/fimmu.2011.00018
71. Doyle A, Martin WJ, Funa K, et al. Markedly decreased expression of class I histocompatibility antigens, protein, and mRNA in human small-cell lung cancer. J Exp Med. 1985;161(5):1135–1151. doi:10.1084/jem.161.5.1135
72. Busch S, Hanke M, Kargl J, Metz H, MacPherson D, Houghton A. Lung cancer subtypes generate unique immune responses. J Immunol. 2016;197(11):4493–4503. doi:10.4049/jimmunol.1600576
73. Hofman P. New insights into the interaction of the immune system with non-small cell lung carcinomas. Transl Lung Cancer Res. 2020;9(5):2199–2213. doi:10.21037/tlcr-20-178
74. Schmidt L, Eskiocak B, Kohn R, et al. Enhanced adaptive immune responses in lung adenocarcinoma through natural killer cell stimulation. Proc Natl Acad Sci USA. 2019;116:17460–17469. doi:10.1073/pnas.1904253116
75. Sun JC, Lanier LL. NK cell development, homeostasis and function: parallels with CD8+ T cells. Nat Rev Immunol. 2011;11(10):645–657. doi:10.1038/nri3044
76. Sek K, Mølck C, Stewart GD, et al. Targeting adenosine receptor signaling in cancer immunotherapy. Int J Mol Sci. 2018;19:3837. doi:10.3390/ijms19123837
77. Inoue Y, Yoshimura K, Kurabe N, et al. Prognostic impact of CD73 and A2A adenosine receptor expression in non-small-cell lung cancer. Oncotarget. 2017;8:8738–8751. doi:10.18632/oncotarget.14434
78. Mediavilla-Varela M, Castro J, Chiappori A, et al. A novel antagonist of the immune checkpoint protein adenosine A2a receptor restores tumor-infiltrating lymphocyte activity in the context of the tumor microenvironment. Neoplasia. 2017;19:530–536. doi:10.1016/j.neo.2017.02.004
79. Vigano S, Alatzoglou D, Irving M, et al. Targeting adenosine in cancer immunotherapy to enhance T-cell function. Front Immunol. 2019;10:925. doi:10.3389/fimmu.2019.00925
80. Heeke S, Mograbi B, Alix-Panabières C, et al. Never travel alone: the crosstalk of circulating tumor cells and the blood microenvironment. Cells. 2019;8(7):714. doi:10.3390/cells8070714
81. Wang WC, Zhang XF, Peng J, et al. Survival mechanisms and influence factors of circulating tumor cells. BiomedRes Int. 2018;2018:6304701. doi:10.1155/2018/6304701
82. De Vito C, Mikulak J, Zaghi E, et al. NK cells to cure cancer. Semin Immunol. 2019;41:101272. doi:10.1016/j.smim.2019.03.004
83. Russick J, Joubert PE, Gillard-Bocquet M, et al. Natural killer cells in the human lung tumor microenvironment display immune inhibitory functions. J Immunother Cancer. 2020;8(2):e001054. doi:10.1136/jitc-2020-001054
84. Kuppen PJ, Van Der Eb MM, Jonges LE, et al. Tumor structure and extracellular matrix as a possible barrier for therapeutic approaches using immune cells or adenoviruses in colorectal cancer. Histochem Cell Biol. 2001;115(1):67–72. doi:10.1007/s004180000224
85. Clift R, Souratha J, Garrovillo SA, Zimmerman S, Blouw B. Remodeling the tumor microenvironment sensitizes breast tumors to anti-programmed death-ligand 1 immunotherapy. Cancer Res. 2019;79(16):4149–4159. doi:10.1158/0008-5472.CAN-18-3060
86. Jin S, Deng Y, Hao JW, et al. NK cell phenotypic modulation in lung cancer environment. PLoS One. 2014;9(10):e109976. doi:10.1371/journal.pone.0109976
87. Topalian SL, Hodi FS, Brahmer JR, et al. Safety, activity, and immune correlates of Anti–PD-1 antibody in cancer. N Engl J Med. 2012;366(26):2443–2454. doi:10.1056/NEJMoa1200690
88. Sun H, Sun C. The rise of NK cell checkpoints as promising therapeutic targets in cancer immunotherapy. Front Immunol. 2019;10:2354. doi:10.3389/fimmu.2019.02354
89. Luna JI, Grossenbacher SK, Murphy WJ, Canter RJ. Targeting cancer stem cells with natural killer cell immunotherapy. Expert Opin Biol Ther. 2017;17(3):313–324. doi:10.1080/14712598.2017.1271874
90. Kim S, Iizuka K, Aguila HL, et al. In vivo natural killer cell activities revealed by natural killer cell-deficient mice. Proc Natl Acad Sci USA. 2000;97(6):2731–2736. doi:10.1073/pnas.050588297
91. Zhang G, Zhao H, Wu J, et al. Adoptive immunotherapy for non-small cell lung cancer by NK and cytotoxic T lymphocytes mixed effector cells: retrospective clinical observation. Int Immunopharmacol. 2014;21(2):396–405. doi:10.1016/j.intimp.2014.04.026
92. Lin M, Liang SZ, Wang XH, et al. Clinical efficacy of percutaneous cryoablation combined with allogenic NK cell immunotherapy for advanced non-small cell lung cancer. Immunol Res. 2017;65(4):880–887. doi:10.1007/s12026-017-8927-x
93. Ding X, Cao H, Chen X, et al. Cellular immunotherapy as maintenance therapy prolongs the survival of the patients with small cell lung cancer. J Transl Med. 2015;13:158. doi:10.1186/s12967-015-0514-0
94. Romagnani C, Juelke K, Falco M, et al. CD56bright CD162 KIR2 NK cells display longer telomeres and acquire features of CD56dim NK cells upon activation. J Immunol. 2007;178:4947–4955. doi:10.4049/jimmunol.178.8.4947
95. Esendagli G, Bruderek K, Goldmann T, et al. Malignant and nonmalignant lung tissue areas are differentially populated by natural killer cells and regulatory T cells in non-small cell lung cancer. Lung Cancer. 2008;59(1):32–40. doi:10.1016/j.lungcan.2007.07.022
96. Hiraki A, Kiura K, Yamane H, et al. Interleukin-12 augments cytolytic activity of peripheral blood mononuclear cells against autologous lung cancer cells in combination with IL-2. Lung Cancer. 2002;35:329–333. doi:10.1016/s0169-5002(01)00424-x
97. Krause SW, Gastpar R, Andreesen R, et al. Treatment of colon and lung cancer patients with ex vivo heat shock protein 70-peptide-activated, autologous natural killer cells: a clinical phase I trial. Clin Cancer Res. 2004;10:3699–3707. doi:10.1158/1078-0432.CCR-03-0683
98. Brownlie D, Scharenberg M, Mold J, et al. Expansions of adaptive-like NK cells with a tissue-resident phenotype in human lung and blood. Proc Natl Acad Sci U S A. 2021;118(11):e2016580118. doi:10.1073/pnas.2016580118
99. Cerwenka A, Lanier LL. Natural killer cell memory in infection, inflammation and cancer. Nat Rev Immunol. 2016;16(2):112–123. doi:10.1038/nri.2015.9
100. Chen Z, Yang Y, Liu LL, Lundqvist A. Strategies to augment natural killer (NK) cell activity against solid tumors. Cancers. 2019;11(7):1040. doi:10.3390/cancers11071040
101. Howlader N, Noone AM, Krapcho M. SEER Cancer Statistic Review. Bethesda, MD: National Cancer Institute; 2019.
102. Poznanski SM, Ritchie TM, Fan IY, et al. Expanded human NK cells from lung cancer patients sensitize patients’ PDL1-negative tumors to PD1-blockade therapy. J Immunother Cancer. 2021;9(1):e001933. doi:10.1136/jitc-2020-001933
103. Yu Y, Zeng D, Ou Q, et al. Association of survival and immune-related biomarkers with immunotherapy in patients with non-small cell lung cancer: a meta-analysis and individual patient-level analysis. JAMA Netw Open. 2019;2(7):e196879. doi:10.1001/jamanetworkopen.2019.6879
104. Chen S, Crabill GA, Pritchard TS, et al. Mechanisms regulating PD-L1 expression on tumor and immune cells. J Immunother Cancer. 2019;7(1):305. doi:10.1186/s40425-019-0770-2
105. Chang CH, Qiu J, O’Sullivan D, et al. Metabolic competition in the tumor microenvironment is a driver of cancer progression. Cell. 2015;162(6):1229–1241. doi:10.1016/j.cell.2015.08.016
106. Lin M, Luo H, Liang S, et al. Pembrolizumab plus allogeneic NK cells in advanced non-small cell lung cancer patients. J Clin Invest. 2020;130:2560–2569. doi:10.1172/JCI132712
107. Barkholt L, Alici E, Conrad R, et al. Safety analysis of ex vivo-expanded NK and NK-like T cells administered to cancer patients: a phase I clinical study. Immunotherapy. 2009;1(5):753–764. doi:10.2217/imt.09.47
108. Parkhurst MR, Riley JP, Dudley ME, Rosenberg SA. Adoptive transfer of autologous natural killer cells leads to high levels of circulating natural killer cells but does not mediate tumor regression. Clin Cancer Res. 2011;17(19):6287–6297. doi:10.1158/1078-0432.CCR-11-1347
109. Mantovani S, Oliviero B, Varchetta S, Mele D, Mondelli MU. Natural killer cell responses in hepatocellular carcinoma: implications for novel immunotherapeutic approaches. Cancers. 2020;12(4):926. doi:10.3390/cancers12040926
110. Ruggeri L, Capanni M, Urbani E, et al. Effectiveness of donor natural killer cell alloreactivity in mismatched hematopoietic transplants. Science. 2002;295(5562):2097–2100. doi:10.1126/science.1068440
111. Kang S, Gao X, Zhang L, Yang E, Li Y, Yu L. The advances and challenges of NK cell-based cancer immunotherapy. Curr Oncol. 2021;28(2):1077–1093. doi:10.3390/curroncol28020105
112. Hermanson DL, Kaufman DS. Utilizing chimeric antigen receptors to direct natural killer cell activity. Front Immunol. 2015;6:195. doi:10.3389/fimmu.2015.00195
113. Myers JA, Miller JS. Exploring the NK cell platform for cancer immunotherapy. Nat Rev Clin Oncol. 2021;18(2):85–100. doi:10.1038/s41571-020-0426-7
114. Gong JH, Maki G, Klingemann HG. Characterization of a human cell line (NK-92) with phenotypical and functional characteristics of activated natural killer cells. Leukemia. 1994;8(4):652–658.
115. Klingemann H, Boissel L, Toneguzzo F. Natural killer cells for immunotherapy - advantages of the NK-92 cell line over blood NK cells. Front Immunol. 2016;7:91. doi:10.3389/fimmu.2016.00091
116. Williams BA, Law AD, Routy B, et al. A phase I trial of NK-92 cells for refractory hematological malignancies relapsing after autologous hematopoietic cell transplantation shows safety and evidence of efficacy. Oncotarget. 2017;8(51):89256–89268. doi:10.18632/oncotarget.19204
117. Suck G, Odendahl M, Nowakowska P, et al. NK-92: an ‘off- the-shelf therapeutic’-for adoptive natural killer cell- based cancer-immunotherapy. Cancer Immunol Immunother. 2016;65(4):485–492. doi:10.1007/s00262-015-1761-x
118. Tonn T, Schwabe D, Klingemann HG, et al. Treatment of patients with advanced cancer with the natural killer cell line NK-92. Cytotherapy. 2013;15(12):1563–1570. doi:10.1016/j.jcyt.2013.06.017
119. Jochems C, Hodge JW, Fantini M, et al. An NK cell line (haNK) expressing-high levels of granzyme and engineered to express the-high affinity CD16 allele. Oncotarget. 2016;7:86359–86373. doi:10.18632/oncotarget.13411
120. Nimmerjahn F, Ravetch JV. Fcgamma receptors as regulators of immune responses. Nat Rev Immunol. 2008;8(1):34–47. doi:10.1038/nri2206
121. Pahl JHW, Koch J, Götz JJ, et al. CD16A activation of NK cells promotes NK cell proliferation and memory-like cytotoxicity against cancer cells. Cancer Immunol Res. 2018;6(5):517–527. doi:10.1158/2326-6066.CIR-17-0550
122. Kendra K, Malkovska V, Allen M, Guzman J, Albertini M. In vivo binding and antitumor activity of Ch14.18. J Immunother. 1999;22(5):423–430. doi:10.1097/00002371-199909000-00006
123. Benson DM, Hofmeister CC, Padmanabhan S, et al. A Phase 1 trial of the anti-KIR antibody IPH2101 in patients with relapsed/refractory multiple myeloma. Blood. 2012;120(22):4324–4433. doi:10.1182/blood-2012-06-438028
124. Spanholtz J, Preijers F, Tordoir M, et al. Clinical- grade generation of active-NK cells from cord blood hematopoietic progenitor-cells for immunotherapy using a closed- system culture-process. PLoS One. 2011;6(6):e20740. doi:10.1371/journal.pone.0020740
125. Knorr DA, Ni Z, Hermanson D, et al. Clinical- scale derivation of natural-killer cells from human pluripotent stem cells for-cancer therapy. Stem Cells Transl Med. 2013;2:274–283. doi:10.5966/sctm.2012-0084
126. Luevano M, Daryouzeh M, Alnabhan R, et al. The unique profile of cord blood natural killer cells balances incomplete maturation and effective killing function upon activation. Hum Immunol. 2012;73(3):248–257. doi:10.1016/j.humimm.2011.12.015
127. Woll PS, Martin CH, Miller JS, Kaufman DS. Human embryonic stem cell-derived NK cells acquire functional receptors and cytolytic activity. J Immunol. 2005;175(8):5095–5103. doi:10.4049/jimmunol.175.8.5095
128. Hong M, Clubb JD, Chen YY. Engineering CAR-T cells for next-generation cancer therapy. Cancer Cell. 2020;38(4):473–488. doi:10.1016/j.ccell.2020.07.005
129. Schuster SJ, Bishop MR, Tam CS, et al.; JULIET Investigators. Tisagenlecleucel in adult relapsed or refractory diffuse large B-cell lymphoma. N Engl J Med. 2019;380(1):45–56. doi:10.1056/NEJMoa1804980.
130. Maude SL, Laetsch TW, Buechner J, et al. Tisagenlecleucel in children and young adults with B-cell lymphoblastic leukemia. N Engl J Med. 2018;378(5):439–448. doi:10.1056/NEJMoa1709866
131. Neelapu SS, Locke FL, Bartlett NL, et al. Axicabtagene ciloleucel CAR T- cell therapy in refractory large B- cell lymphoma. N Engl J Med. 2017;377(26):2531–2544. doi:10.1056/NEJMoa1707447
132. Shah NN, Fry TJ. Mechanisms of resistance to CAR T cell therapy. Nat Rev Clin Oncol. 2019;16(6):372–385. doi:10.1038/s41571-019-0184-6
133. Fujisaki H, Kakuda H, Imai C, Mullighan CG, Campana D. Replicative potential of human natural killer cells. Br J Haematol. 2009;145(5):606–613. doi:10.1111/j.1365-2141.2009.07667.x
134. Denman CJ, Senyukov VV, Somanchi SS, et al. Membrane- bound IL-21 promotes sustained ex vivo proliferation of human natural killer cells. PLoS One. 2012;7(1):e30264. doi:10.1371/journal.pone.0030264
135. Ojo EO, Sharma AA, Liu R, et al. Membrane bound IL-21 based NK cell feeder cells drive robust expansion and metabolic activation of NK cells. Sci Rep. 2019;9(1):14916. doi:10.1038/s41598-019-51287-6
136. Carlsten M, Childs RW. Genetic manipulation of NK cells for cancer immunotherapy: techniques and clinical implications. Front Immunol. 2015;6:266. doi:10.3389/fimmu.2015.00266
137. Park HR, Ahn YO, Kim TM, et al. NK92-CD16 cells are cytotoxic to non-small cell lung cancer cell lines that have acquired resistance to tyrosine kinase inhibitors. Cytotherapy. 2019;21(6):603–611. doi:10.1016/j.jcyt.2019.03.312
138. Vallera DA, Felices M, McElmurry R, et al. IL15 trispecific killer engagers (TriKE) make natural killer cells specific to CD33+ targets while also inducing persistence, in vivo expansion, and enhanced function. Clin Cancer Res. 2016;22(14):3440–3450. doi:10.1158/1078-0432.CCR-15-2710
139. Jiang H, Zhang W, Shang P, et al. Transfection of chimeric anti-CD138 gene enhances natural killer cell activation and killing of multiple myeloma cells. Mol Oncol. 2014;8(2):297–310. doi:10.1016/j.molonc.2013.12.001
140. Nagashima S, Mailliard R, Kashii Y, et al. Stable transduction of the interleukin-2 gene into human natural killer cell lines and their phenotypic and functional characterization in vitro and in vivo. Blood. 1998;91(10):3850–3861. doi:10.1182/blood.V91.10.3850
141. Grote S, Mittelstaet J, Baden C, et al. Adapter chimeric antigen receptor (AdCAR)-engineered NK-92 cells: an off-the-shelf cellular therapeutic for universal tumor targeting. Oncoimmunology. 2020;9(1):1825177. doi:10.1080/2162402X.2020.1825177
142. Wang W, Jiang J, Wu C. CAR-NK for tumor immunotherapy: clinical transformation and future prospects. Cancer Lett. 2020;472:175–180. doi:10.1016/j.canlet.2019.11.033
143. Marofi F, Rahman HS, Thangavelu L, et al. Renaissance of armored immune effector cells, CAR-NK cells, brings the higher hope for successful cancer therapy. Stem Cell Res Ther. 2021;12(1):200. doi:10.1186/s13287-021-02251-7
144. Khan M, Arooj S, Wang H. NK cell-based immune checkpoint inhibition. Front Immunol. 2020;11:167. doi:10.3389/fimmu.2020.00167
145. Hu Y, Tian ZG, Zhang C. Chimeric antigen receptor (CAR)-transduced natural killer cells in tumor immunotherapy. Acta Pharmacol Sin. 2018;39(2):167–176. doi:10.1038/aps.2017.125
146. Tokarew N, Ogonek J, Endres S, von Bergwelt-baildon M, Kobold S. Teaching an old dog new tricks: next-generation CAR T cells. Br J Cancer. 2019;120(1):26–37. doi:10.1038/s41416-018-0325-1
147. Molgora M, Cortez VS, Colonna M. Killing the invaders: NK cell impact in tumors and anti-tumor therapy. Cancers (Basel). 2021;13(4):595. doi:10.3390/cancers13040595
 © 2021 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2021 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.