Back to Journals » Nature and Science of Sleep » Volume 13
The Effects of Genetic Background for Diurnal Preference on Sleep Development in Early Childhood
Authors Morales-Muñoz I ![]() , Kantojärvi K, Uhre VM, Saarenpää-Heikkilä O
, Kantojärvi K, Uhre VM, Saarenpää-Heikkilä O ![]() , Kylliäinen A, Pölkki P, Himanen SL, Karlsson L, Karlsson H, Paavonen EJ, Paunio T
, Kylliäinen A, Pölkki P, Himanen SL, Karlsson L, Karlsson H, Paavonen EJ, Paunio T
Received 19 October 2020
Accepted for publication 15 January 2021
Published 16 February 2021 Volume 2021:13 Pages 219—228
DOI https://doi.org/10.2147/NSS.S287163
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Dr Steven Shea
Isabel Morales-Muñoz,1,2 Katri Kantojärvi,1,3 Veli-Matti Uhre,1 Outi Saarenpää-Heikkilä,4,5 Anneli Kylliäinen,6 Pirjo Pölkki,7 Sari-Leena Himanen,8,9 Linnea Karlsson,10– 12 Hasse Karlsson,10– 12 E Juulia Paavonen,1,13 Tiina Paunio1,3
1Department of Public Health Solutions, Finnish Institute for Health and Welfare, Helsinki, Finland; 2Institute for Mental Health, School of Psychology, University of Birmingham, Birmingham, UK; 3Department of Psychiatry and SleepWell Research Program, Faculty of Medicine, University of Helsinki and Helsinki University Central Hospital, Helsinki, Finland; 4Pediatric Clinic, Tampere University Hospital, Tampere, Finland; 5Tampere Centre for Child Health Research, University of Tampere and Tampere University Hospital, Tampere, Finland; 6Psychology, Faculty of Social Sciences, Tampere University, Tampere, Finland; 7Department of Social Sciences, University of Eastern Finland, Kuopio, Finland; 8Department of Clinical Neurophysiology, Tampere University Hospital, Medical Imaging Centre and Hospital Pharmacy, Pirkanmaa Hospital District, Tampere, Finland; 9Faculty of Medicine and Health Technology, Tampere University, Tampere, Finland; 10The FinnBrain Birth Cohort Study, Turku Brain and Mind Center, Department of Clinical Medicine, University of Turku, Turku, Finland; 11Centre for Population Health Research, Turku University Hospital and University of Turku, Turku, Finland; 12Department of Psychiatry, University of Turku and Turku University Hospital, Turku, Finland; 13Pediatric Research Center, Child Psychiatry, University of Helsinki and Helsinki University Hospital, Helsinki, Finland
Correspondence: Isabel Morales-Muñoz Tel +358 29 524 6000
Email [email protected]
Tiina Paunio Tel +358 29 410 2941
Email [email protected]
Purpose: No previous research has examined the impact of the genetic background of diurnal preference on children´s sleep. Here, we examined the effects of genetic risk score for the liability of diurnal preference on sleep development in early childhood in two population-based cohorts from Finland.
Participants and Methods: The primary sample (CHILD-SLEEP, CS) comprised 1420 infants (695 girls), and the replication sample (FinnBrain, FB; 962 girls) 2063 infants. Parent-reported sleep duration, sleep-onset latency and bedtime were assessed at three, eight, 18 and 24 months in CS, and at six, 12 and 24 months in FB. Actigraphy-based sleep latency and efficiency were measured in CS in 365 infants at eight months (168 girls), and in 197 infants at 24 months (82 girls). Mean standard scores for each sleep domain were calculated in both samples. Polygenic risk scores (PRS) were used to quantitate the genetic risk for eveningness (PRSBestFit) and morningness (PRS10kBest).
Results: PRSBestFit associated with longer sleep-onset latency and later bedtime, and PRS10kBest related to shorter sleep-onset latency in CS. The link between genetic risk for diurnal preference and sleep-onset latency was replicated in FB, and meta-analysis resulted in associations (P< 0.0005) with both PRS-values (PRSBestFit: Z=3.55; and PRS10kBest: Z=− 3.68). Finally, PRSBestFit was related to actigraphy-based lower sleep efficiency and longer sleep latency at eight months.
Conclusion: Genetic liability to diurnal preference for eveningness relates to longer sleep-onset during the first two years of life, and to objectively measured lowered sleep efficiency. These findings enhance our understanding on the biological factors affecting sleep development, and contribute to clarify the physiological sleep architecture in early childhood.
Keywords: genetic risk, chronotype, sleep, early childhood, cohorts
Introduction
Circadian rhythms are cyclical changes in cellular, molecular, and biological processes repeating approximately every 24 hours.1 They are driven by an internal “master clock”, and play a central role in the regulation of many physiological processes, including sleep–wake cycles.2 Chronotype, or diurnal preference, refers to the individual timing of the endogenous circadian system affecting the preferred behavioral patterns including timing of sleep and wake.3 Chronotype falls on a continuum with individuals in one extreme who are referred to as morning types, and individuals in the other extreme, being referred to as evening types.
Circadian rhythmicity has a genetic basis4 and the genetic mechanisms of the core circadian clock has been also established.5 Consistently, inter-individual differences in chronotype are also determined by genetic factors,1 although age, gender and environmental light exposure may also explain this variation. For example, twin and family studies have estimated the heritability of chronotype to be up to 50%.6–8 Furthermore, recent genome-wide association studies (GWAS) have identified specific genetic variations associated with self-reported chronotype.9–12 Based on these studies, one can calculate quantitative polygenic risk scores (PRS), which can be used for estimation of individual genetic proneness for diurnal preference.
In adolescents and adults, evening chronotype associates with negative social and health consequences.13,14 Furthermore, chronotype influences sleep quality.15–17 However, little is known about its effect on sleep-wake patterns in early childhood. The scarce existing studies indicate that parent-reported morningness in toddlers associates with earlier bedtimes, sleep onset times, and wake times,18 while evening types show bedtime resistance, and wake in a negative mood.19 Further, we recently reported that self-reported maternal eveningness associated with several sleep problems in children from three to 24 months of age.20
Examining chronotype-sleep association is relevant in early childhood because this stage is characterized by substantial inter-individual differences in sleep patterns.21,22 Furthermore, a high percentage of parents report their children experiencing sleep difficulties during the first two years of life.22,23 Therefore, a better understanding of the factors affecting sleep development is needed. However, all the previous studies in children have only used parent-reported measures of chronotype, while none have utilized data about the genetic factors of chronotype, such as the polygenic risk estimates.
The main objective of this study was to examine whether PRS for eveningness (http://www.nealelab.is/blog/2017/7/19/rapid-gwas-of-thousands-of-phenotypes-for-337000-samples-in-the-uk-biobank) and morningness,12 are related to parent-reported nighttime and daytime sleep duration, sleep-onset latency and bedtime during the first two years of life, in two population-based birth cohorts. We selected these sleep variables because our main interest was to investigate those sleep variables that assumingly would be related to circadian preference, ie bedtime, sleep onset latency and sleep duration.20 They present a high variability among infants in early childhood,22 and this high variability is likely to be related to several influencing variables, such as genetic factors, and in this case, polygenic risk scores for chronotype. Further, we also examined whether differences between boys and girls exist, and how the genetic risk for diurnal preference manifests in sleep at different stages of early childhood. Finally, we studied the relation of the genetic risk scores on actigraphy-based sleep measures. We hypothesized that PRS for eveningness would relate to shorter sleep duration, longer sleep-onset latency and later bedtime,20 while PRS for morningness would associate with longer sleep duration, shorter sleep-onset latency and earlier bedtime.18
Participants and Methods
Participants
The primary sample of this study, CHILD-SLEEP (CS), is a birth cohort in Pirkanmaa, southern Finland.24 For this study, we used parental questionnaires during pregnancy (32nd week), umbilical cord blood samples at birth, questionnaire-based sleep measures of the infants at three, eight, 18 and 24 months, and actigraphy-based sleep measures at eight and 24 months. The initial dataset comprised 1679 families. Further, 14 children with chronic illnesses were excluded. From this initial sample, 1420 umbilical cord blood samples were available. Of these, we had questionnaire data from 99.2% (N=1409) infants at three months, 90.3% (N=1282) at eight months, 80.6% (N=1145) at 18 months, and 65.8% (N=935) at 24 months. While all the questionnaires and umbilical blood samples were obtained from the whole sample, actigraphy measures were only conducted in 365 cases at eight months, and 197 at 24 months.
The replication sample of this study consists of another population-based cohort, the FinnBrain (FB), in southwest Finland (www.finnbrain.fi).25 In all, 5790 out of 8895 women visiting the recruitment sites during pregnancy (12th week) were contacted. A total of 3808 (66%) mothers decided to participate. Umbilical blood sample were obtained from 2063 infants, and 10 cases were excluded due to reported chronic illnesses. We finally included only participants with parent-reported sleep data available, ie 1491 infants at six months, 1214at 12 months and 994 at 24 months. In this case, actigraphy-based sleep measures were not available.
Ethics Statement
The CS cohort was reviewed and approved by the Pirkanmaa Hospital District Ethical Committee (9/3/2011, Ethical Research Permission Code R11032), and the FB cohort by the Ethical Committee of the Southwestern Finland Hospital District (number 57/180/2011). Permission for the recruitment procedure in both cohorts was received from the physicians at the target health centers. Parental informed consent was obtained for all participants, in accordance with the Declaration of Helsinki.
Measures
Genetic Estimates for Chronotype
In both cohorts, DNA samples were extracted according to standard procedures at the Finnish Institute for Health and Welfare. DNA samples for CS were genotyped with Illumina Infinium PsychArray and for FB with Illumina Infinium PsychArray and Illumina Infinium Global Screening Array at Estonian Genome Centre. Quality control was performed with PLINK 1.9 (www.cog-genomics.org/plink/1.9/).26 Markers were removed for missingness (>5%) and Hardy–Weinberg equilibrium (p-value < 1 x 10–6). Individuals were checked for missing genotypes (>5%), relatedness (identical by descent calculation, PI_HAT>0.2) and population stratification (multidimensional scaling). Principal component analysis (PCA) for population stratification was calculated with PLINK 1.9. Genotyped data in CS was imputed with SHAPEITv227 and IMPUTEv228 against Finnish WGS and 1000 genomes reference panels. Genotyped data of FB was pre-phased with Eagle v2.429 and imputed with Beagle 4.1.30 SISU-project whole-genome sequencing data was used as imputation reference panel.
PRSBestFit was derived from UK Biobank sample consisting of 301,143 individuals (UK Biobank GWAS Manifest 20,170,915, phenotype code: 1180; http://www.nealelab.is/blog/2017/7/19/rapid-gwas-of-thousands-of-phenotypes-for-337000-samples-in-the-uk-biobank). In UK Biobank sample phenotype for eveningness was estimated with the question “Do you consider yourself to be?” “Definitely a ‘morning’ person, More a ‘morning’ than an ‘evening’ person, More an ‘evening’ than a ‘morning’ person or Definitely an ‘evening’ person”. We tested the best fit p-value threshold for eveningness with a national sample of FINRISK where information about the chronotype was collected with the same question as in UK Biobank study. Best fit p-value threshold was 0.007. This was used as a measure of eveningness. We used PRSice program31 to estimate PRS for studied individuals. PRS10kBest was derived from genome-wide association studies (GWAS) from 697,828 UK Biobank and 23andMe,12 which was treated as a measure of morningness. In 23andMe cohort, phenotype for morningness was estimated with two identical questions (“Are you naturally a night person or a morning person?”). For the first instance, the possible answers were “Night owl”, “Early bird” and “Neither”, and for the second “Night person”, “Morning person”, “Neither”, “It depends” and “I’m not sure”. Individuals with discordant or neutral responses to both were excluded. 10,000 most significant SNPs were included in PRS10kBest. Detailed description of this procedure has been described elsewhere.12 Correlations between PRSBestFit and PRS10kBest showed moderate associations in CS (r=−0.361, p<0.001) and FB (r=−0.314, p<0.001).
Subjective Sleep Measures
The Brief Infant Sleep Questionnaire (BISQ) was used to evaluate infant sleep quality.32 We selected the following items: (i) nocturnal sleep hours; (ii) daytime sleep hours; (iii) sleep-onset latency; and (iv) bedtime. BISQ was used in CS at all time points. Furthermore, sleep-onset latency and bedtime were also obtained in FB, at six and 12 months. The four sleep items were based on open-ended items.
The Infant Sleep Questionnaire (ISQ) assesses infant sleeping habits on an interval scale with seven to nine response alternatives.33 For this study, sleep-onset latency at all time points in CS cohort was selected, as follows: How long does it usually take to settle your baby off to sleep on average?: 1=<10 minutes; 2=10–20 minutes; 3=20–30 minutes; 4=30–40 minutes; 5=40–50 minutes; 6=50–60 minutes; or 7=≥1 hour. We additionally included the ISQ sleep-onset latency item in this study to control for potential subjective bias associated with open-ended items (ie BISQ items), which can result in data with differential quality.34
The Sleep Disturbance Scale for Children (SDSC) assesses the frequency of a variety of sleep-related behaviors in preschoolers,35 children and adolescents,36 as follows: “How long after going to bed does your child usually fall asleep: 1=<15 minutes; 2=15–30 minutes; 3=30–45 minutes; 4=45–60 minutes; or 5≤60 minutes.” This scale was used to measure sleep-onset latency at 24 months in FB cohort.
The World Health Organization (WHO)-coordinated survey sleep items are derived from the school children´s health behavior study.37 In FB, parents were asked to report about their child bedtime at the age of 24 months, as follows: “When does your child usually go to bed?” There were alternatives ranging from 21:00 to 02:00 hours or later, in half hour intervals.
Objective Sleep Measures
For this study, Actiwatch 7 was used (CamNtech Ltd, Cambridge, United Kingdom). Parents were asked to place the actigraphy on the thigh of their infant for three consecutive days (ie two nights) and to complete the sleep-log. Nighttime activity data were scored using the sleep analysis program provided by the manufacturer. This device uses the Oakley algorithm,38 which has been previously validated in infants.39 Further, we used a medium sensitivity (wake threshold), following previous studies using similar devices in pediatric population.40 All cases with only one night recorded (n = 10) were excluded from the final analyses. Our sleep variables of interest were sleep efficiency and sleep latency.
Covariates
The following covariates were considered: age (days), sex, three top principal components to adjust the effect estimates for population stratification and season of assessment. Seasons were defined in terms of the amount of daily light at the date of assessment: (i) IL= increasing light: Feb 5-May 5; (ii) L= light: May 6-Aug 5; (iii) ID= increasing darkness: Aug 6-Nov 6; and (iv) D= dark: Nov 7-Feb 4.
Sociodemographic variables and description of the variables of interest appear in Table 1.
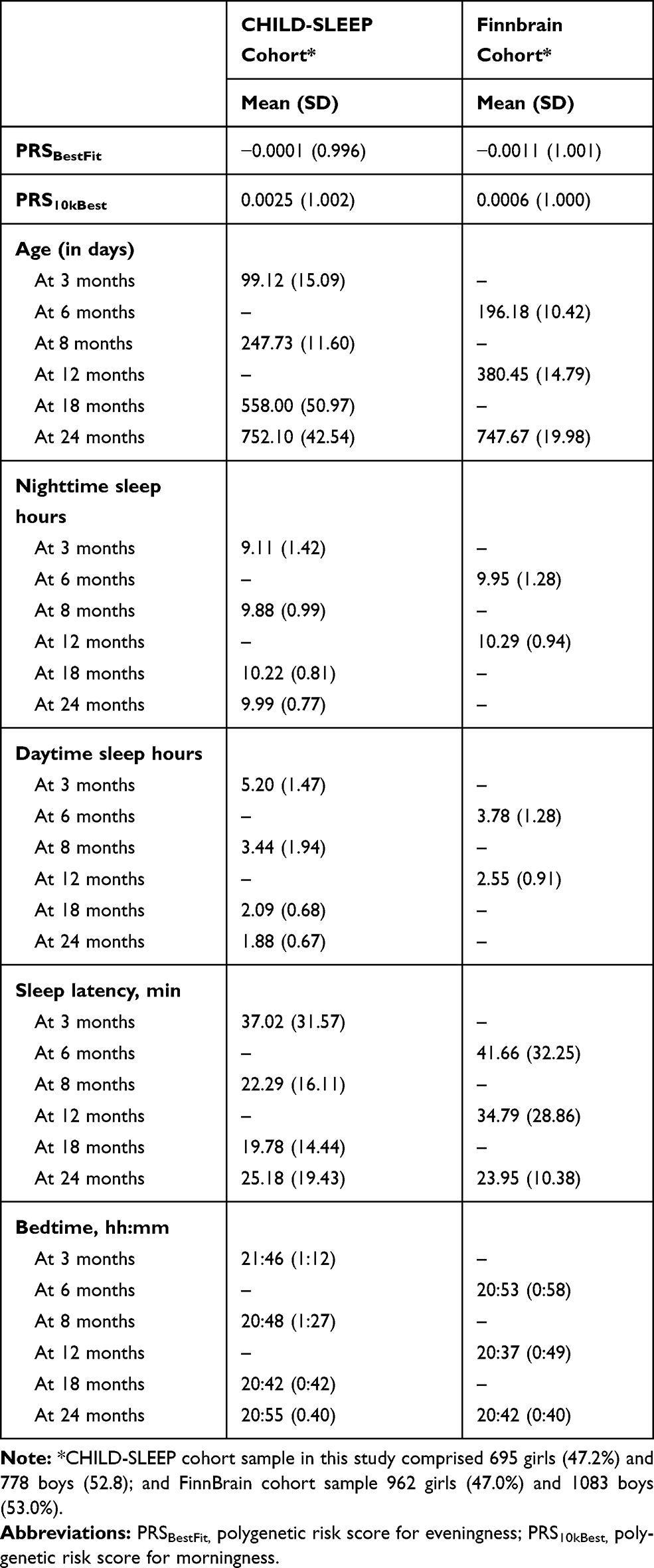 |
Table 1 Socio-Demographic Factors and Variables of Interest in CHILD-SLEEP and FinnBrain |
Statistical Analyses
Statistical analyses were performed with SPSS V25.0. The mean standard scores for the main outcome (ie each sleep domain) were calculated by summing the standard score of each time point from one specific sleep domain and dividing by four, which was the number of time points. By using this approach, instead of other longitudinal analysis approaches, such as Latent Growth Curve Analysis, we assume that we are missing several cases, due to attrition, across the time points. However, and taking into account the high variability inter- and intra-subject in sleep development that is observed in early childhood, the approach presented here is the most suitable one for the purpose of this study.
To test our primary hypothesis that genetic risk for diurnal preference is related to sleep, linear regression models were conducted in CS. To do this, mean standard scores for each sleep domain were used as dependent variables, while PRSBestFit and PRS10kBest were used as independent variables. Each independent and dependent variable were conducted in different models, together with all the covariates. The acquired P values were adjusted to control false discovery rate (FDR) using the Benjamini–Hochberg procedure.41
Similar regression models, using mean standard scores were applied in FB cohort. Meta-analyses using the meta data from the CS and FB cohorts were performed using R 3.6042 with “meta” package using fixed effect model.43
To examine whether differences between boys and girls appeared in CS, the interaction terms between sex and PRS were added in a secondary analysis. Furthermore, to examine how the genetic risk for diurnal preference expresses at different time points, separate linear regression models were applied, at each time point in both cohorts. Finally, linear regression models were applied to examine the links between genetic risk for diurnal preference and actigraphy-based sleep measures in CS; first, using mean standard scores for each actigraphy domain; and second, using separate regression analyses at eight and 24 months.
Results
Genetic Risk for Diurnal Preference and Parent-Reported Sleep in CS
PRSBestFit associated with mean standard scores of longer sleep-onset latency using both BISQ and ISQ (R2=0.019, β=0.081, p=0.004; and R2=0.020, β=0.081, p=0.004, respectively). Additionally, PRSBestFit associated with mean standard scores of later bedtime (R2=0.028, β=0.085, p=0.003). PRS10kBest associated with mean standard scores of shorter ISQ sleep-onset latency (R2=0.016, β=−0.060, p=0.034).
After Benjamini-Hochberg multiple testing correction, the associations between PRSBestFit and longer BISQ and ISQ sleep-onset latency, as well as PRSBestFit and bedtime remained significant (FDR<0.005; see Table 2). For description of the mean values and standard errors of ISQ sleep-onset latency for each quartile of PRSBestFit and PRS10kBest, see Figure 1.
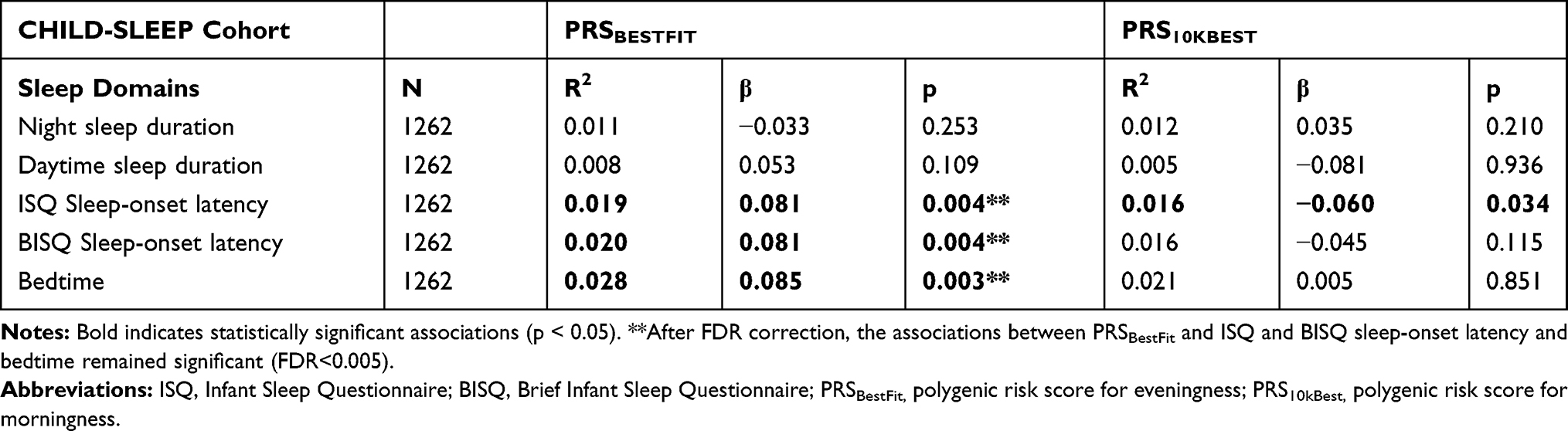 |
Table 2 Associations Between Genetic Risk for Diurnal Preference and Mean Standard Scores for Each Sleep Domain, in CHILD-SLEEP |
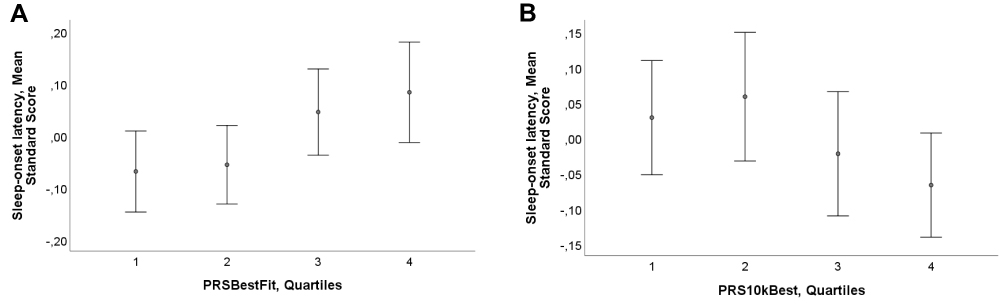 |
Figure 1 Sleep-onset latency and genetic risk for diurnal preference in early childhood. This graph presents the mean and standard error (SE) in parent-reported sleep-onset latency in early childhood, for each quartile of PRSBestFit (A) and PRS10kBest (B), in the CHILD-SLEEP cohort. The measure of parent-reported sleep-onset latency in childhood refers to mean standard scores at three, eight, 18 and 24 months, from the Infant Sleep Questionnaire. Graph A indicates that those subjects situated in the 4th Quartile (ie higher PRS for eveningness) report higher mean standard score values in ISQ sleep-onset latency (ie longer sleep-onset latency). Accordingly, graph B shows that those infants located in the 4th Quartile (ie higher PRS for morningness) obtain lower mean standard values in ISQ sleep-onset latency (ie short sleep-onset latency). Error bars represent 2SE, indicating 95% of values that are less than two standard errors (2SE) away from the mean. |
Replication in FB
We aimed at replication of the significant findings (ie association of PRSBestFit and PRS10kBest with sleep-onset latency and bedtime) in FB. The results only showed association between PRS10kBest and BISQ sleep-onset latency (R2=0.014, β=−0.049, p=0.034). However, no significant associations were observed between any of the PRSes and bedtime (see supplementary Table 1).
Meta-Analysis
Mean standard score regarding BISQ sleep-onset latency was significantly associated with both genetic scores (PRSBestFit, Z=3.55, p=0.0004; PRS10kBest, Z=−3.68, p=0.0002). Combining BISQ sleep-onset latency from FB-cohort and ISQ sleep-onset latency from CS-cohort strengthened this association (PRSBestFit: Z=3.62, p=0.0003 and PRS10kBest: Z=−4.11, P<0.0001) (see Figure 2).
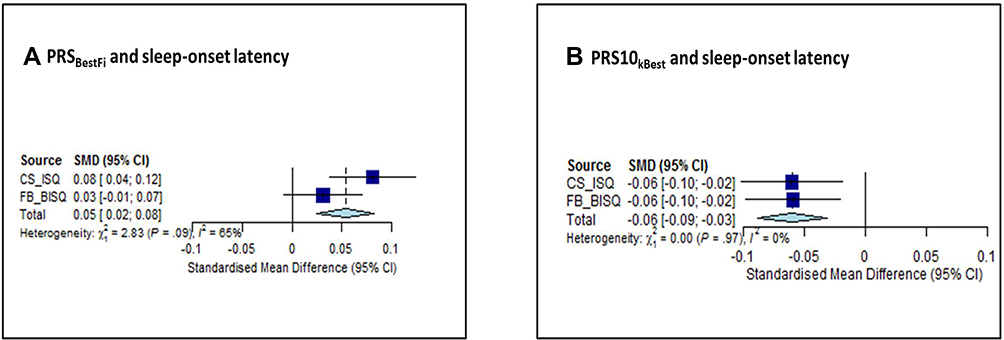 |
Figure 2 Forest plot from the meta-analysis, concerning genetic risk for diurnal preference and sleep-onset latency in early childhood. This graph provides a graphical display of the estimated results when we combined the data from both cohorts (CS and FB) to perform meta-analysis. Graph (A) represents the forest plot from the meta-analysis, concerning the association between PRSBestFit and sleep-onset latency in early childhood. Graph (B) represents the forest plot from the meta-analysis, concerning the association between PRS10kBest and sleep-onset latency in early childhood. First dark blue square on the top, in each graph, represents the standard mean differences (and 95% Confidence Intervals) for sleep-onset latency measured with Infant Sleep Questionnaire in CS, and the dark blue square below, in each graph, refers to the standard mean differences (and 95% Confidence Intervals) for sleep-onset latency measured with Brief Infant Sleep Questionnaire in FB. The light blue diamond on the bottom appearing in each graph represents the overall effect estimate. Abbreviations: SMD, standardized mean difference; CI, confidence interval; CS, CHILD-SLEEP; FB, FinnBrain; ISQ, Infant Sleep Questionnaire; BISQ, Brief Infant Sleep Questionnaire; PRSBestFit, polygenic risk score for eveningness; PRS10kBest, polygenic risk score for morningness. |
Differences Between Boys and Girls in CS
No significant interactions of gender with any of the PRS values were observed in any of the associations.
Associations Between Genetic Risk for Diurnal Preference and Parent-Reported Sleep-Onset Latency at Separate Time Points
To study the temporal pattern of manifestation of genetic risk for diurnal preference on sleep-onset latency, the most consistent result in both cohorts, we analyzed this sleep variable separately at each time point (see supplementary Table 2). As sleep development is highly variable at different stages of early childhood,22 we wanted to investigate whether the effect of PRS for chronotype in sleep-onset latency was more relevant at a specific stage of the child´s development.
Results from CS indicated that both PRSBestFit and PRS10kBest were associated with ISQ sleep-onset latency at three (R2=0.022, β=0.081, p=0.007; and R2=0.021, β=−0.077, p=0.008, respectively) and 18 months (R2=0.020, β=0.087, p=0.009; and R2=0.018, β=−0.066, p=0.037, respectively), and with BISQ sleep-onset latency at 18 months (R2=0.028, β=0.096, p=0.005; and R2=0.025, β=−0.073, p=0.026, respectively). Further, PRSBestFit associated with BISQ sleep-onset latency at three months (R2=0.028, β=0.087, p=0.005), and PRS10kBest related to ISQ sleep-onset latency at 24 months (R2=0.025, β=−0.069, p=0.050). Interestingly, no significant associations with sleep-onset latency at eight months were observed. To further understand this lack of association, we conducted again this analysis in those children with actigraphy at eight months, and this new analysis confirmed this lack of association (see supplementary Table 3).
In FB, we found associations between PRS10kBest and BISQ sleep-onset latency at 12 months (R2=0.017, β=−0.061, p=0.035), while no associations at P<0.05 were found with PRSBestFit.
Genetic Risk for Diurnal Preference and Actigraphy-Based Sleep Measures in CS
There was significant association between PRSBestFit and the mean standard score for actigraphy-based sleep efficiency (R2=0.054, β=−0.150, p=0.008). In separate analysis of the time points, there were significant relations between PRSBestFit and sleep efficiency (R2=0.055, β=−0,154, p=0.008) and between PRSBestFit and sleep latency (R2=0.045, β=0.116, p=0.046) at eight months. Finally, no significant associations were found for PRS10kBest (see Table 3).
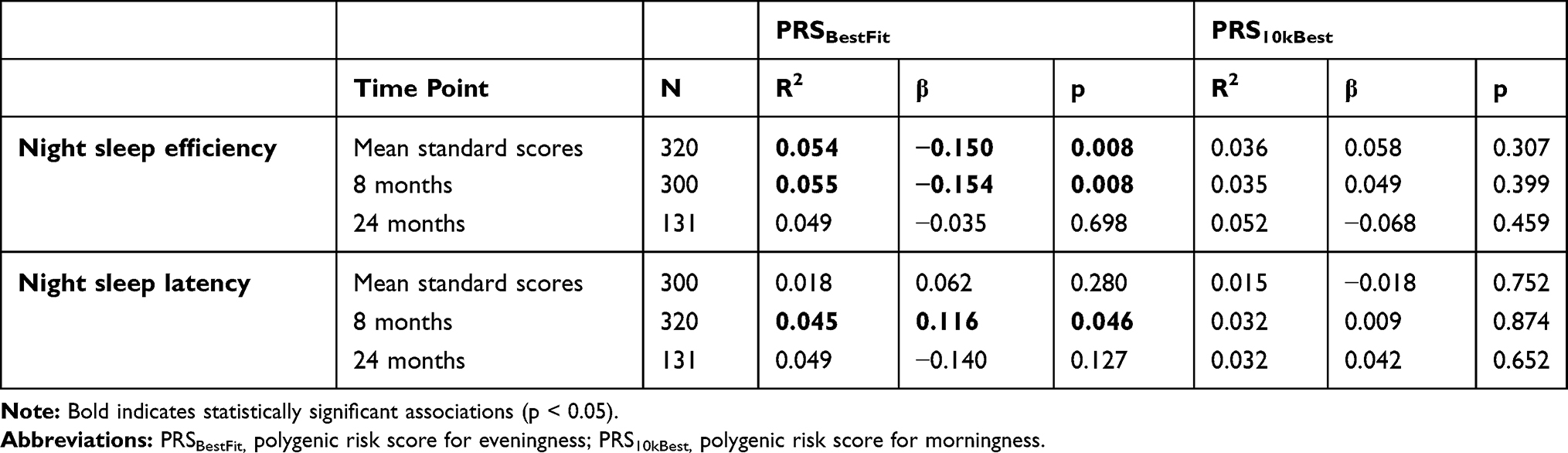 |
Table 3 Associations Between Genetic Risk for Diurnal Preference and Actigraphy-Based Sleep Variables, in CHILD-SLEEP |
Discussion
Our main findings indicate that genetic liability to eveningness associates with parent-reported longer sleep-onset latency and later bedtime in early childhood, while preference for morningness relates to shorter sleep-onset latency. This association between genetic risk for parent-reported diurnal preference and parent-reported sleep-onset latency was replicated in another birth cohort in Finland. Finally, we found that the polygenic background for eveningness related to longer actigraph-based sleep latency at eight months and lower sleep efficiency in early childhood.
Our findings are consistent with previous questionnaire-based studies, where toddlers exhibiting parent-reported morningness had earlier sleep onset times.18 We found that both PRS for eveningness and morningness were associated with longer and shorter parent-reported sleep-onset latency in early childhood, respectively; and PRS for eveningness was also linked to longer actigraphy-based sleep latency at eight months. These findings are also in line with our previous studies, where we reported self-reported maternal eveningness being related to longer sleep-onset latency at three, 18 and 24 months.20 Further, our current findings are also consistent with previous studies in adolescent twins showing that the influence of genetic factors was greatest for actigraphy-based sleep-onset latency.44
Secondly, we found associations between PRS for eveningness and later bedtime during early childhood. These findings support previous studies showing that self-reported maternal eveningness20 associated with later bedtimes in early childhood. Further, previous research has shown that parent-reported morningness in toddlers concurrently associated with earlier bedtimes.18 In our study, we did not observe significant associations between PRS for morningness and earlier bedtime. This might be explained by the fact that our sample was considerably younger than previous research using parent-reported chronotype in toddlers.
Thirdly, when we examined the associations between genetic risk for diurnal preference and parent-reported sleep-onset latency at different time points, we found that PRS for eveningness related to sleep-onset latency at three and 18 months, while no associations were found at eight months or 24 months. This suggests that the genetic contribution of chronotype may be masked by co-occurring environmental and developmental factors, challenging the parental evaluation of sleep parameters at these specific stages. This is supported in our finding that PRS for eveningness associated with sleep-onset latency in eight-month-old infants when measured with actigraphy. It could be that subjective evaluation of sleep onset latency is challenged in specific time periods like shown in findings of discrepancies45 and underreports46 of sleep onset times by parents of preschoolers compared to actigraphy measures.
Interestingly, we did not find association between genetic risk for diurnal preference and sleep duration in early childhood. These results contradict previous research using parent-reported chronotype.20,47 However, this lack of association is consistent with previous research conducted in young adult twins6 and school-aged children,48,49 as well as with a large study in adult population showing non-significant genetic association between sleep duration and chronotype.50 Furthermore, previous findings from 18 month-old twins support the idea that short sleep may be mainly explained by common shared environmental factors.51
This study has some limitations. First, the amount of bright light during the day could contribute to individual differences in sleep difficulties.52 However, we controlled for season of assessment, based on the changes in the amount of daily light. Second, there are some other confounding factors that might influence our results, such as environmental factors (ie daycare, especially at 18 and 24 months) or children´s developmental factors. Further, it is important to take into account the important role of environmental factors, in conjunction with genetic ones, when investigating the influences on sleep traits in early childhood. In fact, this is something that we have already reported in our previous research on the effects of parental chronotype20 and maternal risk factors53 in the development of sleep problems in early childhood. In this current study, we were interested in reporting the child´s genetic contribution (ie PRS for diurnal preference), but these findings should be interpreted with caution, as relevant parent-related environmental factors might also exert some influence. Third, as is usual in birth cohort studies, we experienced loss of study cases across the time points. However, the attrition rate in our study was relatively low, which ensures the validity of the findings. Fourth, although we reported significant associations between genetic liability and subjective sleep pattern (ie parent-reported sleep-onset latency and bedtime), the effect size of these associations were relatively small; however, the effect size between genetic liability and objective sleep measures (ie actigraph-based sleep latency and sleep efficiency) was considerably bigger. Therefore, this suggests that when examining the associations between genetic liability and parent-reported sleep development in early childhood, several contributing variables, probably associated with parent-related factors, might also play an important role in these associations. However, the bigger effect size observed with the actigraphy indicates that these associations appear stronger when subjective factors are controlled. Fifth, some other sleep timing parameters of interest could have also been considered for this study, such as midpoint of sleep and/or getup time. However, getup time was not a sleep parameter included within the parent-reported questionnaire data, and thus we were not able to study this specific sleep timing parameter or to calculate the midpoint of sleep in the whole sample of this study. However, this is something to consider for future research on the topic. Finally, these results were solely conducted in Finnish population. Considering the unique genetic heritage in Finland,54 this study should be replicated in other geographical regions. It is, however, noteworthy that the PRSes used in this study were obtained from UK Biobank and 23andMe comprising mostly individuals with European descent. The polygenic risk score derived from UK Biobank (PRSBestFit) was based on information from SNPs of 301,143 individuals, which gave us a chance to test the best fit p-value threshold in an independent Finnish population sample. PRS10kBest was derived from a larger GWAS containing almost 700,000 individuals from the UK Biobank and 23andMe dataset but with information of only the 10,000 most significant variants. Thus, no population-specific threshold in the Finnish sample could be calculated.
Conclusion
PRS for eveningness and morningness are related to sleep-onset latency in early childhood. Furthermore, PRS for eveningness associates with later bedtime and to actigraphy-based lowered sleep efficiency. These findings add to the recent research on parental-reported chronotype and children´s sleep development, indicating that genetic background for diurnal preference contributes to sleep-wake development in early childhood. Further studies in later stages of childhood should be conducted, to understand whether this genetic influence mainly appears at early stages of life or whether it extends to later stages. Our study helps to clarify the physiological architecture of sleeping in early childhood and may eventually contribute to the development of personalized therapeutic approaches targeting the circadian system.
Acknowledgments
The authors would like to thank all the families that participated in the CHILD-SLEEP and FinnBrain birth cohorts. The authors are also grateful for the nurses at the maternity clinics who introduced the study to the families.
Funding
This work was supported by The Academy of Finland (#134880 and #253346 to TP; #308588 to EJP, #308589 to LK; # 134950 and #253270 to HK; #277557 to OSH; #315035 to IMM; and #326631 to AK), the Signe and Ane Gyllenberg foundation, the Yrjö Jahnsson Foundation, the Foundation for Pediatric Research, the Finnish Cultural Foundation, the Competitive Research Financing of the Expert Responsibility area of Tampere University Hospital, the Arvo ja Lea Ylppö Foundation, Finnish State Grants for Clinical Research (ERVA), and The Medical Society of Tampere.
Disclosure
The authors report no conflicts of interest in this work.
References
1. Kalmbach DA, Schneider LD, Cheung J, et al. Genetic basis of chronotype in humans: insights from three landmark GWAS. Sleep. 2017;40:2. doi:10.1093/sleep/zsw048
2. Güldür TOH. Circadian rhythm in mammals: time to eat & time to sleep. Biol Rhythm Res. 2017;48(2):243–261. doi:10.1080/09291016.2016.1251968
3. Adan A, Archer SN, Hidalgo MP, Di Milia L, Natale V, Randler C. Circadian typology: a comprehensive review. Chronobiol Int. 2012;29(9):1153–1175. doi:10.3109/07420528.2012.719971
4. Rijo-Ferreira F, Takahashi JS. Genomics of circadian rhythms in health and disease. Genome Med. 2019;11(1):82. doi:10.1186/s13073-019-0704-0
5. Cox KH, Takahashi JS. Circadian clock genes and the transcriptional architecture of the clock mechanism. J Mol Endocrinol. 2019;63(4):R93–R102. doi:10.1530/JME-19-0153
6. Barclay NL, Eley TC, Buysse DJ, Rijsdijk FV, Gregory AM. Genetic and environmental influences on different components of the pittsburgh sleep quality index and their overlap. Sleep. 2010;33(5):659–668. doi:10.1093/sleep/33.5.659
7. Vink JM, Groot AS, Kerkhof GA, Boomsma DI. Genetic analysis of morningness and eveningness. Chronobiol Int. 2001;18(5):809–822. doi:10.1081/CBI-100107516
8. Koskenvuo M, Hublin C, Partinen M, Heikkila K, Kaprio J. Heritability of diurnal type: a nationwide study of 8753 adult twin pairs. J Sleep Res. 2007;16(2):156–162. doi:10.1111/j.1365-2869.2007.00580.x
9. Hu Y, Shmygelska A, Tran D, Eriksson N, Tung JY, Hinds DA. GWAS of 89,283 individuals identifies genetic variants associated with self-reporting of being a morning person. Nat Commun. 2016;7:10448. doi:10.1038/ncomms10448
10. Lane JM, Vlasac I, Anderson SG, et al. Genome-wide association analysis identifies novel loci for chronotype in 100,420 individuals from the UK Biobank. Nat Commun. 2016;7:10889. doi:10.1038/ncomms10889
11. Jones SE, Tyrrell J, Wood AR, et al. Genome-wide association analyses in 128,266 individuals identifies new morningness and sleep duration loci. PLoS Genet. 2016;12(8):e1006125. doi:10.1371/journal.pgen.1006125
12. Jones SE, Lane JM, Wood AR, et al. Genome-wide association analyses of chronotype in 697,828 individuals provides insights into circadian rhythms. Nat Commun. 2019;10(1):343. doi:10.1038/s41467-018-08259-7
13. Gelbmann G, Kuhn-Natriashvili S, Pazhedath TJ, Ardeljan M, Wober C, Wober-Bingol C. Morningness: protective factor for sleep-related and emotional problems in childhood and adolescence? Chronobiol Int. 2012;29(7):898–910. doi:10.3109/07420528.2012.686946
14. Roenneberg T, Allebrandt KV, Merrow M, Vetter C. Social jetlag and obesity. Curr Biol. 2012;22(10):939–943. doi:10.1016/j.cub.2012.03.038
15. Katzenberg D, Young T, Finn L, et al. A CLOCK polymorphism associated with human diurnal preference. Sleep. 1998;21(6):569–576. doi:10.1093/sleep/21.6.569
16. Lazar AS, Slak A, Lo JC-Y, et al. Sleep, diurnal preference, health, and psychological well-being: a prospective single-allelic-variation study. Chronobiol Int. 2012;29(2):131–146. doi:10.3109/07420528.2011.641193
17. Merikanto I, Paavonen EJ, Saarenpaa-Heikkila O, Paunio T, Partonen T. Eveningness associates with smoking and sleep problems among pregnant women. Chronobiol Int. 2017;34(5):650–658. doi:10.1080/07420528.2017.1293085
18. Simpkin CT, Jenni OG, Carskadon MA, et al. Chronotype is associated with the timing of the circadian clock and sleep in toddlers. J Sleep Res. 2014;23(4):397–405. doi:10.1111/jsr.12142
19. Zimmermann LK. The influence of chronotype in the daily lives of young children. Chronobiol Int. 2016;33(3):268–279. doi:10.3109/07420528.2016.1138120
20. Morales-Munoz I, Partonen T, Saarenpaa-Heikkila O, et al. The role of parental circadian preference in the onset of sleep difficulties in early childhood. Sleep Med. 2019;54:223–230. doi:10.1016/j.sleep.2018.10.039
21. Iglowstein I, Jenni OG, Molinari L, Largo RH. Sleep duration from infancy to adolescence: reference values and generational trends. Pediatrics. 2003;111(2):302–307. doi:10.1542/peds.111.2.302
22. Paavonen J, Saarenpää-Heikkilä O, Morales-Munoz I, et al. Normal sleep development in infants: findings from two large birth cohorts. Sleep Med. 2020;69:145–154. doi:10.1016/j.sleep.2020.01.009
23. Petit D, Touchette E, Tremblay RE, Boivin M, Montplaisir J. Dyssomnias and parasomnias in early childhood. Pediatrics. 2007;119(5):e1016–25. doi:10.1542/peds.2006-2132
24. Paavonen J, Saarenpaa-Heikkila O, Polkki P, Kylliainen A, Porkka-Heiskanen T, Paunio T. Maternal and paternal sleep during pregnancy in the Child-sleep birth cohort. Sleep Med. 2017;29:47–56. doi:10.1016/j.sleep.2016.09.011
25. Karlsson L, Tolvanen M, Scheinin NM, et al. Cohort profile: the finnbrain birth cohort study (FinnBrain). Int J Epidemiol. 2018;47(1):15–16j. doi:10.1093/ije/dyx173
26. Panagiotou OA, Willer CJ, Hirschhorn JN, Ioannidis JPA. The power of meta-analysis in genome-wide association studies. Annu Rev Genomics Hum Genet. 2013;14:441–465. doi:10.1146/annurev-genom-091212-153520
27. Chang CC, Chow CC, Tellier LC, Vattikuti S, Purcell SM, Lee JJ. Second-generation PLINK: rising to the challenge of larger and richer datasets. Gigascience. 2015;4:7. doi:10.1186/s13742-015-0047-8
28. Delaneau O, Zagury J-F, Marchini J. Improved whole-chromosome phasing for disease and population genetic studies. Nat Methods. 2013;10(1):5–6. doi:10.1038/nmeth.2307
29. Howie BN, Donnelly P, Marchini J. A flexible and accurate genotype imputation method for the next generation of genome-wide association studies. PLoS Genet. 2009;5(6):e1000529. doi:10.1371/journal.pgen.1000529
30. Loh P-R, Danecek P, Palamara PF, et al. Reference-based phasing using the haplotype reference consortium panel. Nat Genet. 2016;48(11):1443–1448. doi:10.1038/ng.3679
31. Euesden J, Lewis CM, O’Reilly PF. PRSice: polygenic Risk Score software. Bioinformatics. 2015;31(9):1466–1468. doi:10.1093/bioinformatics/btu848
32. Sadeh A. A brief screening questionnaire for infant sleep problems: validation and findings for an Internet sample. Pediatrics. 2004;113(6):e570–7. doi:10.1542/peds.113.6.e570
33. Morrell JM. The infant sleep questionnaire: a new tool to assess infant sleep problems for clinical and research purposes. Child Psychol Psychiatry Rev. 1999;4:20–26. doi:10.1017/S1360641798001816
34. Choi BCK, Pak AWP. A catalog of biases in questionnaires. Prev Chronic Dis. 2005;2(1):A13.
35. Romeo DM, Bruni O, Brogna C, et al. Application of the sleep disturbance scale for children (SDSC) in preschool age. Eur J Paediatr Neurol. 2013;17(4):374–382. doi:10.1016/j.ejpn.2012.12.009
36. Bruni O, Ottaviano S, Guidetti V, et al. The Sleep Disturbance Scale for Children (SDSC). Construction and validation of an instrument to evaluate sleep disturbances in childhood and adolescence. J Sleep Res. 1996;5(4):251–261. doi:10.1111/j.1365-2869.1996.00251.x
37. Tynjälä J, Kannas L, Levälahti E. Perceived sleep quality and its precursors in adolescents. J Sleep Res. 1997;6:189–198. doi:10.1046/j.1365-2869.1997.00048.x
38. Oakley NR. Validation with Polysomnography of the Sleepwatch Sleep/Wake Scoring Algorithm Used by the Actiwatch Activity Monitoring. Technical Report to Mini Mitter Co., Inc., System; 1997.
39. So K, Buckley P, Adamson TM, Horne RSC. Actigraphy correctly predicts sleep behavior in infants who are younger than six months, when compared with polysomnography. Pediatr Res. 2005;58(4):761–765. doi:10.1203/01.PDR.0000180568.97221.56
40. Meltzer LJ, Montgomery-Downs HE, Insana SP, Walsh CM. Use of actigraphy for assessment in pediatric sleep research. Sleep Med Rev. 2012;16(5):463–475. doi:10.1016/j.smrv.2011.10.002
41. Benjamini Y, Hochberg Y. Controlling the false discovery rate: a practical and powerful approach to multiple testing. J R Stat Soc Ser B. 1995;57(1):289–300.
42. R Core Team. R: A Language and Environment for Statistical Computing. Vienna: R Foundation for Statistical Computing; 2019.
43. Schwarzer G. meta: an R Package for Meta-Analysis. R News. 2007;7(3):40–45.
44. Sletten TL, Rajaratnam SMW, Wright MJ, et al. Genetic and environmental contributions to sleep-wake behavior in 12-year-old twins. Sleep. 2013;36(11):1715–1722. doi:10.5665/sleep.3136
45. Kushnir J, Sadeh A. Correspondence between reported and actigraphic sleep measures in preschool children: the role of a clinical context. J Clin Sleep Med JCSM off Publ Am Acad Sleep Med. 2013;9(11):1147–1151. doi:10.5664/jcsm.3154
46. Goodlin-Jones BL, Tang K, Liu J, Anders TF. Sleep patterns in preschool-age children with autism, developmental delay, and typical development. J Am Acad Child Adolesc Psychiatry. 2008;47(8):930–938. doi:10.1097/CHI.ObO13e3181799f7c
47. Jafar NK, Tham EKH, Eng DZH, et al. The association between chronotype and sleep problems in preschool children. Sleep Med. 2017;30:240–244. doi:10.1016/j.sleep.2016.11.015
48. Gedda L, Brenci G. Sleep and dream characteristics in twins. Acta Genet Med Gemellol. 1979;28(3):237–239. doi:10.1017/s0001566000009089
49. Gregory AM, Rijsdijk FV, Eley TC. A twin-study of sleep difficulties in school-aged children. Child Dev. 2006;77(6):1668–1679. doi:10.1111/j.1467-8624.2006.00966.x
50. Dashti HS, Jones SE, Wood AR, et al. Genome-wide association study identifies genetic loci for self-reported habitual sleep duration supported by accelerometer-derived estimates. Nat Commun. 2019;10(1):1100. doi:10.1038/s41467-019-08917-4
51. Brescianini S, Volzone A, Fagnani C, et al. Genetic and environmental factors shape infant sleep patterns: a study of 18-month-old twins. Pediatrics. 2011;127(5):e1296–302. doi:10.1542/peds.2010-0858
52. Swaminathan K, Klerman EB, Phillips AJK. Are individual differences in sleep and circadian timing amplified by use of artificial light sources? J Biol Rhythms. 2017;32(2):165–176. doi:10.1177/0748730417699310
53. Morales-Munoz I, Saarenpaa-Heikkila O, Kylliainen A, et al. The effects of maternal risk factors during pregnancy on the onset of sleep difficulties in infants at 3 months old. J Sleep Res. 2018;27(5):e12696. doi:10.1111/jsr.12696
54. Kääriäinen H, Muilu J, Perola M, Kristiansson K. Genetics in an isolated population like Finland: a different basis for genomic medicine? J Community Genet. 2017;8(4):319–326. doi:10.1007/s12687-017-0318-4
 © 2021 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2021 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.