Back to Journals » Drug Design, Development and Therapy » Volume 19
Targeting Cellular Senescence for Healthy Aging: Advances in Senolytics and Senomorphics
Authors Alum EU ![]() , Izah SC, Uti DE
, Izah SC, Uti DE ![]() , Ugwu OPC
, Ugwu OPC ![]() , Betiang PA, Basajja M, Ejemot-Nwadiaro RI
, Betiang PA, Basajja M, Ejemot-Nwadiaro RI ![]()
Received 27 May 2025
Accepted for publication 11 September 2025
Published 19 September 2025 Volume 2025:19 Pages 8489—8522
DOI https://doi.org/10.2147/DDDT.S543211
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Tuo Deng
Esther Ugo Alum,1 Sylvester Chibueze Izah,2 Daniel Ejim Uti,1 Okechukwu Paul-Chima Ugwu,1 Peter A Betiang,3 Mariam Basajja,4 Regina Idu Ejemot-Nwadiaro5
1Department of Research Publications, Kampala International University, Kampala, Uganda; 2Department of Community Medicine, Faculty of Clinical Sciences, Bayelsa Medical University, Yenagoa, Bayelsa, Nigeria; 3Department of Access Early Childhood and Special Needs Education, Kampala International University, Kampala, Uganda; 4Leiden Institute of Advanced Computer Science (LIACS), Leiden University, Leiden, Netherlands; 5Directorate of Research, Innovation, Consultancy and Extension (RICE), Kampala International University, Kampala, Uganda
Correspondence: Esther Ugo Alum, Email [email protected]; [email protected]
Background: Cellular senescence is a fundamental characteristic of aging, marked by permanent cell cycle cessation and the release of pro-inflammatory mediators. Although senescence plays advantageous roles in tissue regeneration and tumor suppression, its accumulation leads to aging-related illnesses and functional deterioration.
Objective: This review examines the processes of cellular senescence, its effects on aging and age-related disorders, and emerging therapeutic strategies to modulate senescence for promoting healthy aging.
Methods: A thorough literature review was performed using peer-reviewed studies on cellular senescence, its molecular pathways, and therapeutic interventions. Emphasis was placed on senolytics, senomorphics, and lifestyle interventions that modulate senescence-associated pathways. Studies published in Scopus, Web of Science and PubMed between 2014– 2025 were selected.
Results: Recent discoveries underscore the dual function of cellular senescence in aging and pathology. The senescence-associated secretory phenotype (SASP) fosters chronic inflammation and tissue dysfunction, connecting senescence to age-related diseases including cardiovascular conditions, dementia, and metabolic disorders. Therapeutic strategies, including senolytics (drugs that specifically eradicate senescent cells) and senomorphics (compounds that suppress SASP without killing cells), show promise in preclinical and clinical studies. Notably, dosing interals (intermittent vs continuous) influence both therapeutic efficacy and adverse events such as thrombocytopenia. Additionally, the state and limitations of clinical validation of aging biomarkers (eg, p16^INK4a, β-galactosidase) remain major hurdles for translation. Lifestyle interventions such as calorie restriction and exercise have also been identified as natural modulators of senescence pathways.
Conclusion: Targeting cellular senescence offers a promising avenue for promoting healthy aging and mitigating age-linked diseases. Continued research into senescence-modulating interventions may lead to novel therapeutics designed to prolong healthspan and lifespan.
Plain Language Summary: As we age, some of our cells stop dividing in a process called cellular senescence. These cells do not die, but instead release harmful substances that can cause inflammation and damage nearby healthy cells. While this process can be helpful early in life (like preventing cancer), too many of these cells in old age contribute to diseases such as heart problems, diabetes, and dementia.
This study reviews recent research into ways to remove or control these “senescent” cells to support healthier aging. Two promising strategies are:Senolytics: drugs that kill senescent cells.Senomorphics: drugs that make senescent cells less harmful without killing them.
Scientists are testing both types of drugs in animals and humans. Some natural compounds, like quercetin (found in apples and onions) and fisetin (found in strawberries), show potential benefits. Others like metformin and rapamycin, which are already used for diabetes or immune issues, might also help slow aging by targeting senescent cells.
The study also emphasizes that healthy habits, like exercise and calorie restriction, naturally reduce the harmful effects of senescence.
Despite promising results, challenges remain. We need more human studies to understand:Which treatments are safest and most effectiveHow to deliver them to the right parts of the bodyHow often they should be used
The authors believe that combining senolytics, senomorphics, and lifestyle changes could significantly improve our health as we age.
Keywords: cellular senescence, aging, senolytics, senomorphics, healthy aging, SASP, age-related diseases
Graphical Abstract:

Introduction
Cellular senescence is a critical biological process that contributes to aging and the onset of age-related diseases. Defined as an irreversible arrest in cell division in response to stress or damage, senescence serves as a protective mechanism to prevent the propagation of damaged cells. However, over time, the accumulation of senescent cells in tissues leads to a decline in cellular function and the development of a range of age-related conditions, including cardiovascular diseases, neurodegenerative disorders, and cancers.1,2 The presence of these cells is often accompanied by the senescence-associated secretory phenotype (SASP), a pro-inflammatory state that can exacerbate tissue dysfunction and promote chronic disease progression. As such, cellular senescence has become a focal point in aging research, offering new opportunities for therapeutic intervention.3
Recent advances in the field have introduced novel strategies aimed at targeting senescent cells to mitigate their harmful effects. Among these strategies, senolytics and senomorphics have emerged as two promising therapeutic approaches.4 Senolytics are compounds that selectively induce the death of senescent cells, thereby reducing their detrimental impact on tissues. In contrast, senomorphics work by modulating the SASP or altering the cellular environment to alleviate the negative effects of senescence without eliminating the senescent cells themselves.5 Both approaches offer a unique opportunity to delay the onset of age-related diseases and extend the healthspan, the period of life spent in good health. Despite promising preclinical findings, challenges such as specificity, safety, and clinical translation remain unresolved.6
This review aims to explore recent advancements in senolytics and senomorphics, their therapeutic potential, and the challenges in translating these strategies into clinical applications for enhancing longevity and healthspan. This study is justified as it provides a comprehensive review of recent advancements in senescence-targeting therapies, highlighting their potential, limitations, and future directions for promoting healthy aging. Unlike previous reviews that often focus on either class of therapeutics, this study integrates recent advancements, compares their mechanisms, and discusses innovative combination strategies. Additionally, it explores emerging challenges in clinical translation, including precision targeting, safety concerns, and personalized interventions, offering new perspectives on optimizing these therapies for longevity and healthspan extension. We will discuss the underlying mechanisms of cellular senescence, the progress in developing senolytics and senomorphics, and the preclinical and clinical data supporting their potential as therapeutic agents. Additionally, we will examine the challenges faced in the translation of these therapies from the laboratory to the clinic, as well as the future prospects for their application in aging-related interventions. By exploring these therapeutic avenues, this review seeks to highlight the potential of targeting cellular senescence as a strategy to promote healthy aging and reduce the burden of age-associated diseases.
This review makes a distinct contribution to the field by integrating mechanistic insights into cellular senescence with a dual therapeutic focus on both senolytics and senomorphics. While previous reviews have often addressed these approaches separately, our synthesis brings together the most recent preclinical and clinical evidence, elucidates how oxidative stress is mechanistically linked to SASP activation, and critically evaluates pharmacokinetic and bioavailability challenges that influence therapeutic translation. By consolidating evidence from 2014–2025 into comprehensive comparative tables and highlighting combination therapy strategies, we offer a novel framework for understanding how targeted senescence modulation can be optimized for healthy aging. Furthermore, the review identifies underexplored research gaps such as tissue-specific senescence heterogeneity, biomarker-driven patient selection, and advanced delivery systems, that, if addressed, could substantially advance both scientific knowledge and clinical application in geroscience.
Methodology
This narrative review was conducted through a comprehensive literature search using databases such as PubMed, Scopus, and Web of Science. Peer-reviewed articles, review papers, and clinical studies published between 2014–2025 were prioritized. Keywords including cellular senescence, aging, senolytics, senomorphics, SASP, and age-related diseases were used to identify relevant studies. Articles were selected based on their relevance to the mechanisms of senescence, its role in aging and disease, and emerging therapeutic strategies. The findings of the study were presented and discussed concurrently. Thus, this review presents current knowledge, gaps in research, and discusses potential future directions for targeting cellular senescence in healthy aging. In total, our search initially retrieved 412 records. After removal of duplicates and screening titles and abstracts for relevance, 358 articles were selected for full-text review. Following eligibility assessment based on predefined inclusion criteria, that is, relevance to cellular senescence mechanisms, therapeutic targeting (senolytics or senomorphics), and availability of clear mechanistic or clinical outcome data, 261 peer-reviewed articles were finally included in this review. Only studies published in English between 2014–2025 were considered. Inclusion criteria prioritized mechanistic studies, preclinical and clinical trials, and high-quality reviews with explicit senescence-related endpoints, while those excluded were conference abstracts, editorials, and studies lacking primary data on senescence modulation.
Cellular Senescence: Mechanisms and Implications
Cellular senescence is a complex and highly regulated process that acts as a response to various cellular stresses, including DNA damage, oxidative stress, telomere shortening, and oncogenic signaling.7 This process, which can be triggered by a variety of internal and external factors, leads to a permanent cell cycle arrest. While senescence functions as a defense mechanism against the propagation of damaged cells and thus prevent the development of cancer and other diseases, it also contributes to the aging process when senescent cells accumulate over time in tissues.8
Mechanisms of Cellular Senescence
Cellular senescence, a state of stable cell cycle arrest that prevents the proliferation of damaged or stressed cells plays a crucial role in aging, tumor suppression, and tissue remodeling. Senescence is triggered by various intrinsic and extrinsic factors, including DNA damage, telomere attrition, oncogene activation, and oxidative stress, as illustrated in Figure 1. Once senescence is established, the cell enters a state of permanent arrest and exhibits specific markers, such as β-galactosidase activity, elevated levels of p53 and p16INK4a, and changes in the chromatin landscape that reinforce its non-proliferative state.1 Understanding the molecular mechanisms underlying senescence is critical for developing interventions in aging-related diseases and cancer. The primary molecular pathways involved in senescence include:
- DNA Damage Response (DDR) and Senescence Induction: When DNA damage occurs, the DDR pathway is activated to repair the damage. If the damage is too severe to repair, the p53-p21 pathway is triggered, inducing cell cycle arrest. This mechanism prevents the replication of damaged DNA and serves as an essential barrier against cancer.9 Additionally, the p16INK4a-Rb pathway is activated in response to cellular stress, causing another form of cell cycle arrest, particularly in older cells.10
- Telomere Shortening: Telomeres are repetitive DNA sequences at the ends of chromosomes that protect them from damage and prevent the loss of genetic material during cell division.11 However, each cell division results in the shortening of telomeres. Once telomeres become critically short, the cell enters senescence as a protective measure to avoid chromosomal instability and potential malignant transformation.12 Telomeres shorten with each cell division due to the end-replication problem in DNA synthesis. Critically short telomeres activate the p53/p21 pathway, leading to an irreversible cell cycle arrest. The enzyme telomerase can counteract this process, but its activity is limited in most somatic cells.13
- Oncogene-Induced Senescence (OIS): Activation of certain oncogenes, such as RAS or MYC, can induce senescence in cells, thereby preventing the transformation of these cells into tumors. OIS acts as a safeguard against the development of cancer by halting the growth of potentially malignant cells.14 In other words, oncogenic activation (eg, RAS, RAF, and MYC) can induce senescence as a protective mechanism against uncontrolled proliferation. Hyperactivation of oncogenes leads to excessive replication stress and activation of the DDR pathway.15
- Oxidative Stress and Mitochondrial Dysfunction: Reactive oxygen species (ROS) generated from mitochondrial dysfunction or environmental stress contribute to cellular senescence.16 Oxidative stress leads to DNA damage, lipid peroxidation, and protein dysfunction.17 ROS can activate the p38 MAPK pathway, which enhances p53 and p16 signaling, reinforcing senescence.18 Importantly, oxidative stress is not an isolated mechanism but a central upstream driver of the SASP. Elevated ROS levels, whether from mitochondrial dysfunction, environmental insults, or chronic inflammation, activate redox-sensitive transcription factors such as NF-κB and p38 MAPK, which directly enhance SASP gene expression. This mechanistic link explains why oxidative stress can amplify SASP-mediated tissue damage, creating a self-reinforcing cycle of inflammation and cellular dysfunction.18 Senolytics reduce SASP not only by eliminating the senescent cells that produce high ROS but also by disrupting pro-survival signaling pathways that sustain SASP activity. For example, dasatinib and quercetin reduce SASP cytokines by promoting apoptosis of ROS-rich senescent cells, while fisetin decreases oxidative stress burden and downregulates NF-κB signaling, thereby dampening SASP output. Understanding this oxidative stress–SASP interplay highlights the need for therapeutic approaches that concurrently target mitochondrial health, redox balance, and SASP modulation.
- Senescence-Associated Secretory Phenotype (SASP): Senescent cells develop a distinct secretory profile, known as the SASP. SASP includes pro-inflammatory cytokines (IL-6, IL-8, TNF-α), growth factors, and matrix metalloproteinases (MMPs), influencing the tissue microenvironment.19 While SASP plays roles in wound healing and tumor suppression, chronic SASP contributes to inflammation, fibrosis, and aging-related diseases.20
- Epigenetic Regulation of Senescence: Chromatin remodeling through histone modifications and DNA methylation influences senescence. H3K9me3 and H3K27me3 histone marks promote heterochromatin formation, reinforcing senescence-associated gene silencing.21 Senescent cells also exhibit DNA methylation changes, altering gene expression patterns.22
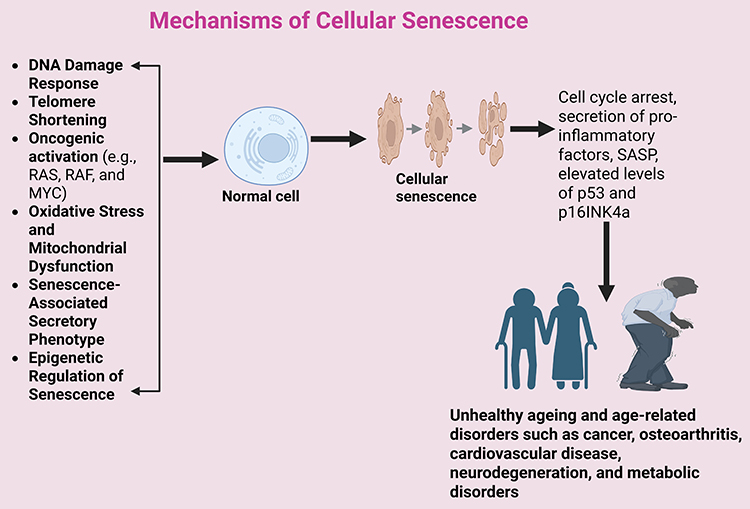 |
Figure 1 Mechanisms of Cellular Senescence (Created in BioRender. Basajja, M. (2025) https://BioRender.com/ q2r74mt). |
Senescence in Aging and Age-Related Diseases
The role of cellular senescence in aging is multifaceted. While senescence may initially serve as a protective mechanism, the long-term accumulation of senescent cells in tissues can contribute to various age-related pathological conditions. Over time, senescent cells secrete a variety of pro-inflammatory factors, collectively known as the senescence-associated secretory phenotype (SASP). The SASP includes cytokines, chemokines, growth factors, and extracellular matrix-degrading enzymes, which promote local inflammation and tissue degeneration.23
- Tissue Dysfunction: Senescent cells accumulate in various tissues as organisms age, including the skin, skeletal muscle, adipose tissue, liver, and vasculature. The presence of these cells in tissues disrupts the normal tissue architecture and impairs regenerative capacities, leading to functional decline.2 For example, in skeletal muscle, the accumulation of senescent cells has been linked to sarcopenia, the age-related loss of muscle mass and function.24
- Chronic Diseases: The inflammatory environment created by senescent cells contributes to the pathogenesis of several chronic diseases. In the cardiovascular system, senescent cells in the blood vessels promote arterial stiffness and endothelial dysfunction, increasing the risk of hypertension and atherosclerosis.25 In the brain, the buildup of senescent cells has been implicated in neurodegenerative diseases such as Alzheimer’s and Parkinson’s, where they exacerbate neuroinflammation and neuronal loss.26 Similarly, in adipose tissue, the accumulation of senescent cells contributes to insulin resistance and the development of metabolic disorders such as type 2 diabetes.27 Senescent chondrocytes promote cartilage degradation and joint inflammation. The SASP secreted by senescent chondrocytes increases matrix metalloproteinase (MMP) activity, leading to the breakdown of cartilage and the progression of osteoarthritis.28
- Cancer: While cellular senescence serves as a protective mechanism against tumorigenesis by halting the proliferation of damaged or oncogene-expressing cells, the chronic inflammatory environment created by senescent cells can promote cancer progression. The SASP can induce genomic instability in neighboring cells, creating a microenvironment conducive to tumor growth.29 Furthermore, the persistence of senescent cells in tissues can undermine the effectiveness of cancer therapies by altering immune surveillance and the tissue response to treatment.30
The Role of Senescence in Aging: A Double-Edged Sword
While senescence is essential for preventing the proliferation of damaged cells and the onset of cancer, its persistent presence in tissues throughout aging disrupts homeostasis. Initially protective, senescent cells transform into a detrimental factor that accelerates aging and facilitates the onset of specific chronic diseases.31 It functions as a double-edged sword, simultaneously inducing organismal deterioration and affecting health positively or negatively. Early life advantages from senescence partly by curtailing the proliferation of cancerous or damaged cells, promoting wound healing, and maintaining tissue homeostasis.32 The senescence-associated secretory phenotype (SASP) comprises bioactive substances generated by senescent cells that modify immune responses and tissue repair. Conversely, age-related senescence becomes detrimental when senescent cells accumulate due to diminished immune clearance. This results in chronic inflammation, tissue dysfunction, and age-associated diseases such as neurodegeneration, cardiovascular disorders, and cancer.3 The pro-inflammatory environment created by SASP exacerbates aging through systemic damage and diminished cellular function.33 The accumulation of senescent cells over time underscores the necessity of developing treatment strategies that selectively target these cells while preserving their beneficial roles in cancer suppression and tissue repair.34 The growing body of research on cellular senescence and its implications for aging has shifted the focus towards therapy techniques aimed at mitigating the adverse effects of senescent cells. Senolytic and senomorphic therapies have garnered significant interest as potential methods to promote healthy aging by either eliminating senescent cells or modifying their detrimental secretions.35 These medications represent a compelling domain in gerontology, aiming to restore tissue equilibrium, enhance organ functionality, and mitigate the impact of age-associated ailments.36 In greater detail, cellular senescence confers physiological benefits under specific conditions such as embryonic development, where transient senescence aids tissue patterning; wound healing, where short-term SASP signaling recruits immune cells for tissue repair; and fibrosis resolution, where senescent myofibroblasts limit excessive extracellular matrix deposition. It also serves as a robust tumor-suppressive mechanism by halting proliferation of potentially malignant cells. However, under pathological conditions such as chronic infections, metabolic dysregulation, or age-related immune decline, senescent cells persist beyond their beneficial window. This persistence is often due to reduced immune clearance efficiency, leading to prolonged SASP activity that fuels chronic inflammation, extracellular matrix degradation, and tissue dysfunction. Mechanistically, this shift from beneficial to detrimental involves sustained activation of NF-κB and p38 MAPK pathways, elevated ROS production from dysfunctional mitochondria, and unresolved DNA damage response signaling. The chronic inflammatory milieu created by these mechanisms accelerates degenerative changes in organs such as the brain, vasculature, joints, and adipose tissue. Understanding these situation-dependent roles is critical for designing interventions that selectively suppress detrimental aspects of senescence while preserving its protective functions.
Senolytics: Targeting Senescent Cells
Senolytics represent a promising class of therapeutic agents that selectively induce the death of senescent cells, thereby reducing the harmful effects these cells have on surrounding tissues. Unlike traditional approaches aimed at mitigating the consequences of aging, senolytics directly target and eliminate senescent cells, potentially reversing the negative impact of their accumulation.37 This approach offers a novel means to delay the onset of age-related diseases and improve overall healthspan. A personalised strategy integrating senolytics with traditional therapies may provide optimal benefits for healthy aging and longevity while mitigating risks.38 Table 1 below compares senolytics and traditional methods targeting cellular senescence.
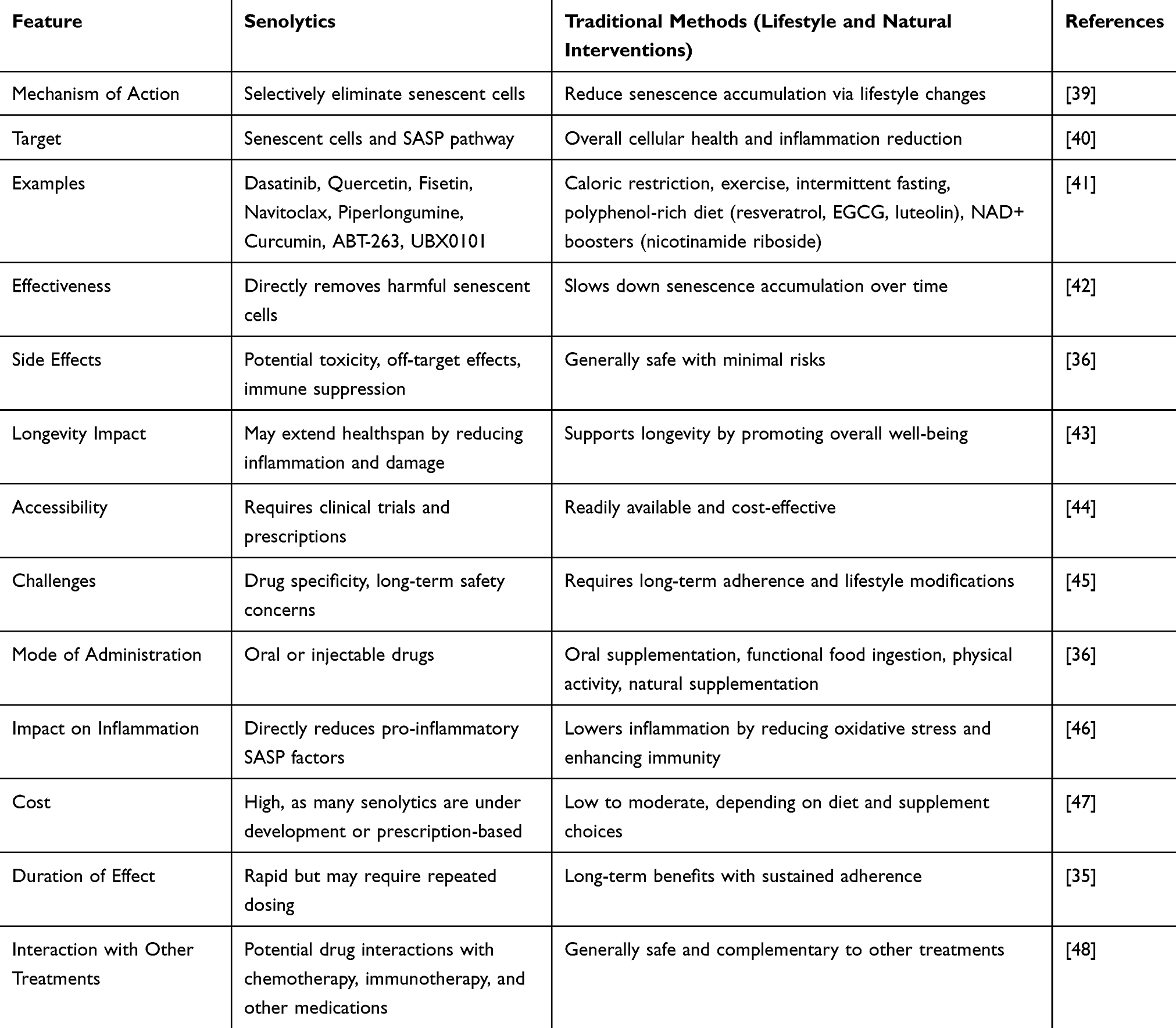 |
Table 1 Comparison Between Senolytics and Traditional Methods Targeting Cellular Senescence for Healthy Aging |
Overview of Senolytics
Senolytics are a class of therapeutic agents specifically designed to eliminate senescent cells, which are cells that have entered an irreversible condition of cell cycle arrest while remaining metabolically active.36 Numerous stressors, such as telomere attrition, oxidative stress, DNA damage, and oncogene activation, induce cellular senescence.1 Initially, senescence safeguards against cancer by inhibiting the proliferation of damaged cells; however, the continuous accumulation of senescent cells, through the secretion of pro-inflammatory molecules known as the senescence-associated secretory phenotype (SASP), ultimately leads to chronic inflammation, tissue dysfunction, and age-related diseases.34 Senolytic therapies are designed to exploit the vulnerabilities of senescent cells, particularly their altered molecular mechanisms that heighten their susceptibility to particular stressors.37 Distinctive features include modifications in the expression of pro-survival proteins and activation of specific survival pathways. Targeting these pathways enables senolytic therapies to specifically induce programmed cell death in senescent cells while sparing non-senescent, healthy cells, hence enhancing healthspan and mitigating age-related diseases.49 In contrast to conventional anti-aging therapies that just decelerate the aging process, senolytics actively eradicate dysfunctional cells, thereby rejuvenating tissues and improving physiological function.50
Mechanisms of Action of Senolytics
Senolytics exert their effects by targeting the key survival pathways that allow senescent cells to evade apoptosis. These pathways include anti-apoptotic proteins, metabolic regulators, and inflammatory mediators, as illustrated in Figure 2 below. Different senolytic agents act through distinct molecular mechanisms, including:
- Inhibition of Anti-Apoptotic Pathways: The BCL-2 family regulates apoptosis, and senescent cells often exhibit increased expression of anti-apoptotic (pro-survival proteins) proteins such as BCL-2, BCL-XL, and BCL-W. These proteins prevent the activation of caspases, which are essential for apoptosis. Senolytic compounds can act by inhibiting these proteins, promoting cell death in senescent cells.51,52 For instance, navitoclax (ABT-263), venetoclax, and ABT-737 are BCL-2 inhibitors that disrupt pro-survival signaling, leading to senescent cell death. These agents have shown promise in preclinical studies for reducing senescent cell burden in aging, fibrosis, and neurodegeneration.53,54
- Disruption of the FOXO4-p53 Interaction: While p53 is a tumor suppressor protein that plays a crucial role in cellular stress responses, its prolonged activation in senescent cells contributes to cellular dysfunction.55 Senolytics can modulate the p53 pathway to promote the selective elimination of senescent cells.56 Additionally, the FOXO4 transcription factor can interact with p53 in senescent cells. This interaction blocks p53-mediated apoptosis, allowing senescent cells to survive.57 FOXO4-DRI, a synthetic peptide, disrupts the FOXO4-p53 interaction, reactivating apoptosis and clearing senescent cells. This approach has demonstrated effectiveness in reversing age-related decline in animal models.58
- Suppression of Senescence-Associated Secretory Phenotype (SASP): Senescent cells secrete pro-inflammatory cytokines (IL-6, IL-8, TNF-α), growth factors, chemokines, and matrix metalloproteinases, which create a pro-inflammatory environment that accelerates aging and disease progression.59 For example, dasatinib and quercetin is a combination therapy that reduces SASP production and enhances the clearance of senescent cells.60 Similarly, fisetin and curcumin are flavonoids with senolytic properties that inhibits SASP secretion, reduce inflammation and improve tissue function.61,62
- Targeting Heat Shock Proteins (HSPs): Heat shock proteins (HSP90, HSP70) are molecular chaperones that stabilize key proteins involved in senescence.63 HSP90 inhibitors (eg, 17-AAG, Geldanamycin) cause degradation of senescence-associated proteins, leading to senescent cell death. This approach is being explored in fibrosis, neurodegenerative diseases, and cancer therapy.64,65
- Modulation of Metabolic Pathways: Senescent cells exhibit altered metabolism, particularly increased reliance on glycolysis and mitochondrial dysfunction. When senescent cells exhibit mitochondrial dysfunction, it leads to an altered cellular redox state. This provides an opportunity for senolytic agents to exploit the altered metabolism of these cells, inducing oxidative stress and cell death.66 For instance, metformin reduces mitochondrial oxidative stress and modulates AMPK signaling, helping prevent the accumulation of senescent cells.67,68 Resveratrol on the other hand activates SIRT1 and AMPK, which promote mitochondrial health and energy balance while NAD+ boosters (such as nicotinamide riboside, nicotinamide mononucleotide) improve mitochondrial function, counteract cellular senescence, and extend lifespan in animal models.69
- Enhancement of Autophagy and Cellular Clearance: Autophagy is a process that removes damaged organelles and proteins, but senescent cells often exhibit impaired autophagy, leading to metabolic dysfunction. This can serve as a therapeutic target against aging and age-related disorders.70 For example, rapamycin, an mTOR inhibitor restores autophagy and delays senescence-related diseases.71,72 Similarly, spermidine induces autophagy and improves cellular renewal, thus promoting longevity.73
- Regulation of Oxidative Stress and DNA Damage Response: Oxidative stress and DNA damage are major drivers of senescence. Some senolytics mitigate these effects by enhancing antioxidant defenses. A common example is sulforaphane, found in cruciferous vegetables, which activates Nrf2, a master regulator of antioxidant pathways.74 Similarly, coenzyme Q10 supports mitochondrial health and reduces oxidative stress-related senescence.75
 |
Figure 2 Mechanisms of Action of Senolytics (Created in BioRender. Basajja, M. (2025) https://BioRender.com/ td8igwu). |
Key Senolytic Compounds and Their Mechanisms of Action
A variety of compounds have been identified as potential senolytics, each with distinct mechanisms of action. These compounds have shown promise in preclinical models and are currently undergoing evaluation in clinical trials to assess safety, effectiveness, and long-term benefits (Table 2).
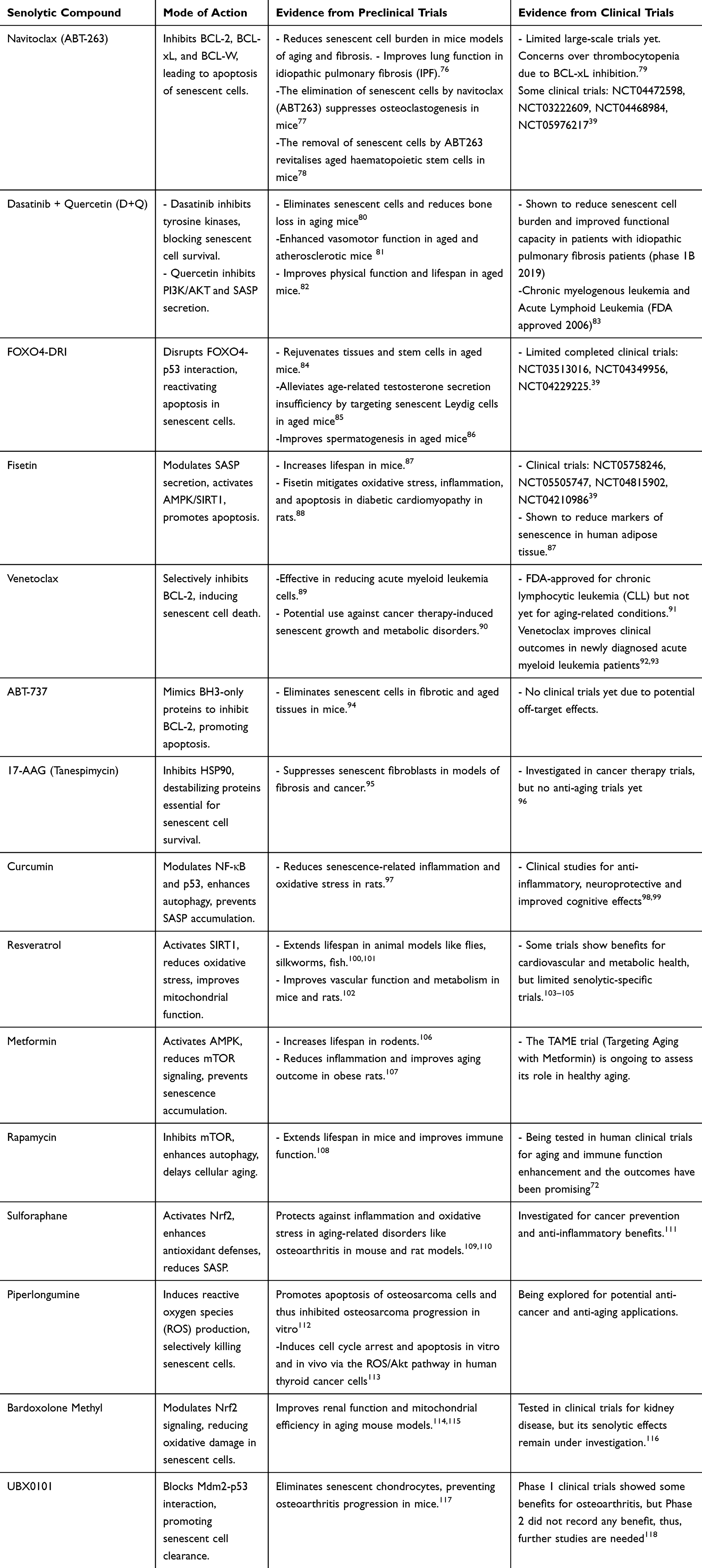 |
Table 2 Key Senolytic Compounds, Their Mode of Action, and Evidence from Preclinical and Clinical Trials |
Challenges in Senolytic Therapy
Despite the promising potential of senolytic therapy in combating aging and age-related diseases, several challenges must be addressed before widespread clinical application. These challenges include safety concerns, drug specificity, delivery methods, off-target effects, long-term consequences, and regulatory hurdles. Addressing these issues through better drug design, personalized treatment approaches, and rigorous clinical research will be crucial for making senolytics a viable therapeutic option in the future.
- Safety and Toxicity Concerns: Many senolytic drugs, such as navitoclax (ABT-263), target BCL-2 family proteins, which are also essential for the survival of non-senescent cells, including platelets and immune cells. This can lead to severe side effects, including thrombocytopenia (low platelet count) and immune suppression, increasing the risk of infections.36 Long-term use of senolytics could potentially cause tissue damage, impaired wound healing, or unintended apoptosis in healthy cells.119
- Lack of Senescent Cell-Specific Targets: One of the major challenges in senolytic therapy is ensuring the selective elimination of senescent cells without affecting healthy cells. While certain senolytic compounds have shown specificity for senescent cells in preclinical models, achieving this selectivity in humans remains a significant hurdle.120 For example, some senescence-associated proteins (eg, BCL-2, p53, SASP factors) are also expressed in normal, proliferating cells, leading to potential off-target effects.31 Therefore, optimizing senolytic agents to minimize side effects while maintaining efficacy is crucial.
- Drug Delivery Challenges: Senolytic drugs need to penetrate different tissues efficiently, but bioavailability and tissue specificity remain major hurdles. Some senolytics, such as FOXO4-DRI, require peptide-based delivery, which is complex and may limit clinical application.58 Blood-brain barrier penetration is another challenge, making it difficult for senolytics to target neurodegenerative diseases effectively.121
- Long-Term Consequences and Need for Repeated Treatment: The long-term effects of senolytic therapy remain unclear, as eliminating senescent cells could disrupt tissue homeostasis. Senescent cells accumulate over time, meaning that senolytic treatment may need to be repeated periodically, raising concerns about chronic drug administration and potential cumulative side effects.122
- Variability in Response and Individual Differences: Senescence is influenced by genetic factors, lifestyle, and environmental exposures, meaning that not all individuals may respond equally to senolytic therapy.123 Age-related changes in metabolism and drug clearance could also impact the effectiveness and safety of senolytics in older adults.124
- Dosing and Treatment Scheduling: Determining the ideal dose and treatment schedule for senolytic therapies is complex. Since the burden of senescent cells increases with age, repeated or long-term treatment may be necessary to maintain therapeutic benefits. However, frequent dosing of senolytics may increase the risk of adverse effects, making it essential to find the right balance between efficacy and safety.36 Emerging clinical and preclinical evidence suggests that intermittent senolytic dosing may reduce the risk of cumulative toxicity, including thrombocytopenia observed with agents such as navitoclax, while preserving efficacy.36 Continuous dosing, while potentially providing sustained clearance, increases exposure-related risks. Thus, optimizing interval schedules, potentially through biomarker-guided timing remains a critical translational challenge.
- Ethical and Regulatory Challenges: Regulatory approval for senolytic drugs is challenging because aging is not officially classified as a disease by the Food and Drug Agency (FDA) or European Medicine Agency EMA, making it harder to obtain approval for anti-aging treatments.125 Ethical concerns regarding the use of senolytics for life extension and enhancement rather than disease treatment could create societal and policy debates. High costs of development and intellectual property issues may limit access to senolytic therapies for wider populations.126
- Need for More Clinical Trials: While preclinical data are promising, large-scale, long-term clinical trials are needed to evaluate efficacy, safety, and optimal dosing of senolytics in humans. Ongoing trials (eg, TAME trial for Metformin, D+Q trials for aging-related diseases) will provide critical insights, but more diverse human studies are required.127
- Bioavailability Challenge: In terms of translational application, bioavailability remains a significant challenge for many senolytic compounds, particularly plant-derived polyphenols such as fisetin and quercetin, which exhibit poor oral absorption, rapid metabolism, and low systemic retention. Preclinical studies often administer fisetin at doses ranging from 20–100 mg/kg orally (PO) in mice, while dasatinib is typically dosed at 5–10 mg/kg PO. In early human trials, the Dasatinib plus Quercetin (D+Q) regimen has used 100 mg/day dasatinib combined with 1000 mg/day quercetin for 3 consecutive days per month, showing acceptable tolerability but variable pharmacokinetic profiles. These limitations underscore the need for delivery innovations such as nanoparticle encapsulation, liposomal formulations, and prodrug designs to enhance bioavailability, protect compounds from first-pass metabolism, and improve tissue-specific targeting. Moreover, pharmacokinetic studies should characterize parameters such as maximum plasma concentration (Cmax), half-life (t½), and area under the curve (AUC) to guide optimal dosing regimens. Integrating such data into clinical translation will facilitate the rational design of dosing schedules that balance efficacy with safety. Nanotechnology and advanced drug delivery systems can enhance the bioavailability, stability, and targeted delivery of senomorphic compounds by encapsulating them in nanoparticles or liposomal carriers.
Senomorphics: Modulating the Senescence-Associated Secretory Phenotype (SASP)
Unlike senolytic therapies that aim to directly eliminate senescent cells, senomorphics work by modulating the senescence-associated secretory phenotype (SASP), which consists of pro-inflammatory cytokines, chemokines, growth factors, and extracellular matrix-degrading enzymes secreted by senescent cells. By targeting the SASP, senomorphic therapies aim to alleviate the detrimental effects of senescence without necessarily removing the senescent cells.128 This approach provides a novel strategy to mitigate the harmful inflammation, tissue degradation, and chronic diseases associated with cellular senescence, while preserving the beneficial aspects of senescence, such as tumor suppression and tissue repair.129
Overview of Senomorphics
Senomorphic agents target the SASP or the molecular pathways that drive it. These compounds aim to reduce inflammation, restore tissue homeostasis, and promote tissue regeneration by modulating the environment surrounding senescent cells.130 Unlike senolytics, which focus on cell death, senomorphics maintain the senescent cell’s presence but modify its behavior, particularly the pro-inflammatory and tissue-damaging components of the SASP.129 By reducing the detrimental effects of senescent cells, such as the SASP, senomorphics hold promise for treating a range of diseases, including cancer, neurodegenerative disorders, cardiovascular diseases, and metabolic syndromes.26
At the molecular level, senomorphics exert their effects primarily by modulating the signaling pathways that govern SASP production without inducing apoptosis in senescent cells. This often involves inhibition of pro-inflammatory transcription factors such as NF-κB and C/EBPβ, suppression of p38 MAPK and mTOR signaling, and enhancement of autophagic flux to reduce the accumulation of damaged organelles and protein aggregates. For instance, rapamycin attenuates SASP by inhibiting mTORC1-dependent translation of IL-1α, a key upstream SASP regulator, while metformin activates AMPK to indirectly suppress NF-κB activity and lower pro-inflammatory cytokine secretion. Flavonoids like apigenin and luteolin downregulate MAPK and JAK/STAT pathways, leading to broad SASP suppression. By altering the secretory profile of senescent cells, senomorphics can mitigate tissue inflammation and promote a microenvironment more conducive to regeneration, while preserving the beneficial growth-arrest functions of senescence. This mechanistic distinction from senolytics is crucial for situations where complete removal of senescent cells may be undesirable, such as in wound healing or tissue repair.
Mechanisms of Actions of Senomorphics
Senomorphics target various molecular pathways involved in cellular senescence, effectively modulating the detrimental aspects of the senescent phenotype while preserving beneficial aspects. Simply put, senomorphics work by modulating key pathways that regulate the SASP. Some of the key mechanisms include the following and illustrated in Figure 3.
- NF-κB Pathway: The nuclear factor-kappa B (NF-κB) pathway plays a central role in regulating the expression of pro-inflammatory cytokines and other SASP factors. In senescent cells, NF-κB is often constitutively activated, driving chronic inflammation.131 Senomorphic agents can inhibit this pathway, reducing the secretion of inflammatory cytokines and alleviating the deleterious effects of chronic inflammation.132
- JAK/STAT Pathway: The Janus kinase (JAK) signal transducer and activator of transcription (STAT) pathway is involved in the inflammatory signaling of senescent cells. Inhibition of the JAK/STAT pathway can reduce the secretion of pro-inflammatory cytokines and mitigate the impact of senescence-induced inflammation.133
- mTOR Pathway: The mechanistic target of rapamycin (mTOR) pathway is implicated in aging and senescence. Activation of mTOR in senescent cells contributes to cellular dysfunction and the pro-inflammatory SASP.134 Inhibition of mTOR has been shown to reduce the expression of SASP factors and improve tissue function in animal models of aging.135
- Modulation of the SASP: Senescent cells secrete a variety of pro-inflammatory cytokines, chemokines, growth factors, and proteases collectively known as the SASP. This secretory profile contributes to chronic inflammation, which is a hallmark of aging and a driver of multiple diseases. Senomorphics reduce SASP activity, thereby lowering inflammation, preventing tissue damage, and mitigating age-related diseases.3
- Epigenetic and Transcriptomic Regulation: Cellular senescence is associated with widespread epigenetic changes, including alterations in DNA methylation, histone modifications, and chromatin remodeling. Senomorphics target these epigenetic regulators to reverse or slow down senescence.136 Some compounds reactivate silenced genes or suppress harmful gene expression, thus modifying the aging process at the molecular level.137
- Maintenance of Mitochondrial and Metabolic Homeostasis: Mitochondrial dysfunction is a major driver of cellular senescence, leading to increased reactive oxygen species (ROS) production, metabolic decline, and energy deficits.138 Senomorphics enhance mitochondrial function by stimulating mitophagy (removal of damaged mitochondria), improving oxidative phosphorylation, and reducing oxidative stress, thereby promoting healthier cellular metabolism.66
- Modulation of DNA Damage Response (DDR) and p53/p21 Pathways: Cellular senescence is often triggered by DNA damage, which activates the DNA damage response (DDR) and key regulatory pathways such as p53/p21 and p16INK4a/Rb.55 While these pathways prevent damaged cells from proliferating, they also promote inflammation and tissue dysfunction if senescent cells persist. Senomorphics modulate these pathways to restore cell cycle control and improve tissue regeneration while maintaining the tumor-suppressive benefits of senescence.139
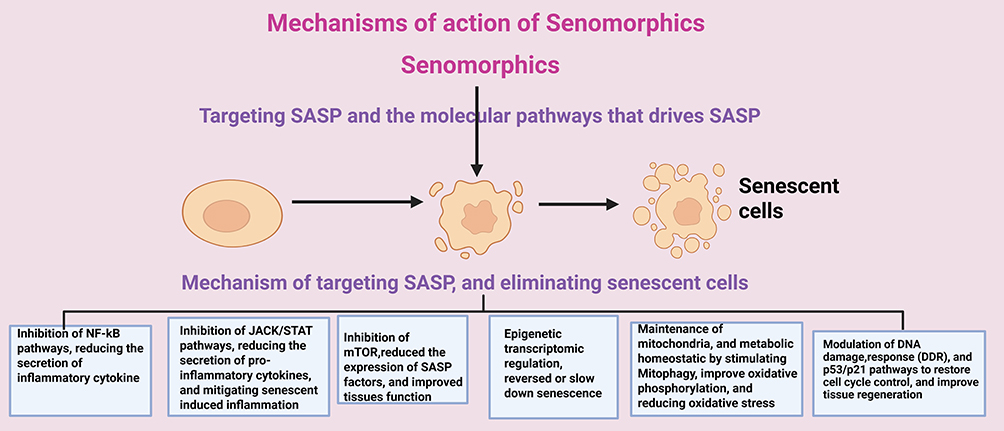 |
Figure 3 Mechanisms of Action of Senomorphics (Created in BioRender. Basajja, M. (2025) https://BioRender.com/ z3l9g0q). |
Differences Between Senolytics and Senomorphics
The major difference between senolytics and senomorphics lies in their mechanism of action on senescent cells. Senolytics selectively eliminate senescent cells by inducing apoptosis, thereby reducing their harmful effects on aging and disease. Senomorphics modulate the behavior of senescent cells, suppressing their harmful secretions (SASP) without killing them, aiming to restore function and reduce inflammation.36,37 In short, senolytics kill senescent cells, while senomorphics reprogram them to be less harmful. A combination of both strategies may provide a more balanced approach to mitigating aging-related pathologies while preserving essential cellular functions.140 Table 3 highlights some major differences between senolytics and senomorphics.
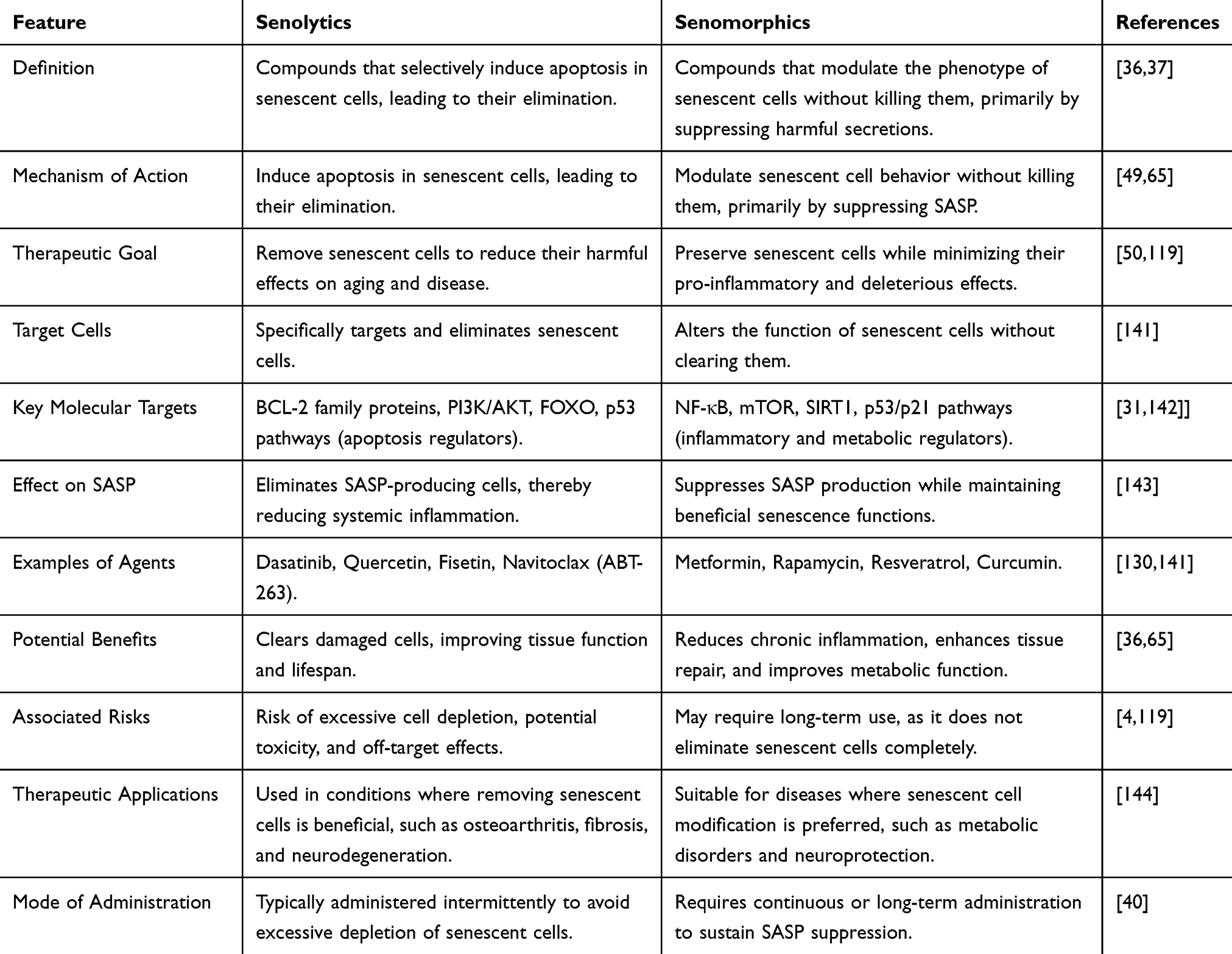 |
Table 3 Some Major Differences Between Senolytics and Senomorphics |
Key Senomorphic Compounds and Their Mechanisms of Action
Several compounds have been identified as potential senomorphics, each targeting different components of the SASP or senescence-associated signaling pathways. These agents aim to modify the senescent cell’s secretory phenotype, suppress inflammation, and promote tissue homeostasis. While animal models have shown encouraging results, the translation of these therapies to humans remains in early stages. Table 4 highlights some senomorphic compounds, their mode of action, and evidence from preclinical and clinical trials.
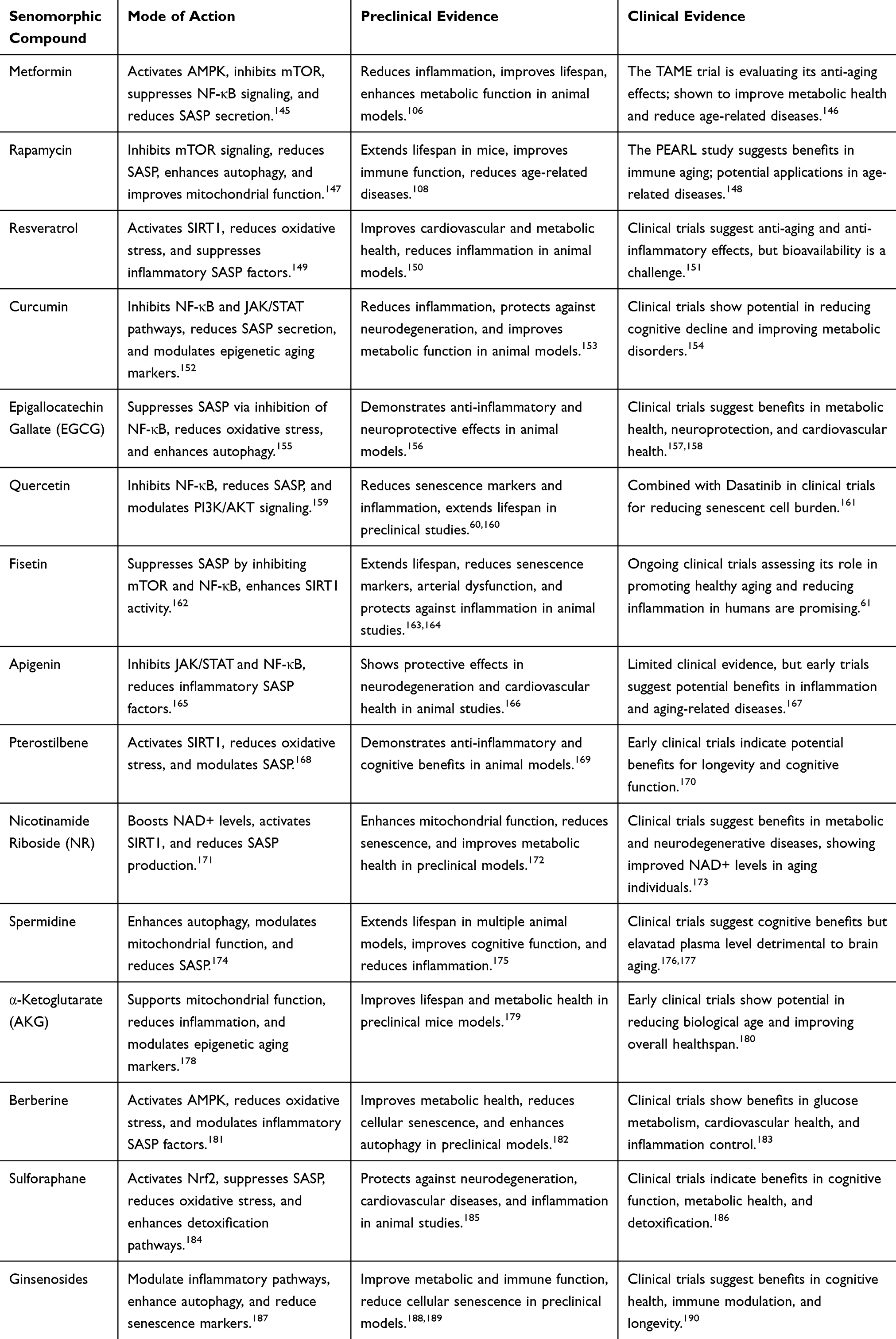 |
Table 4 Key Senomorphic Compounds, Their Mode of Action, and Evidence from Preclinical and Clinical Trials |
Challenges and Opportunities in Senomorphic Therapy
Senomorphic therapy offers a promising approach to mitigating the harmful effects of cellular senescence without eliminating the beneficial functions of senescent cells and thus holds great promise in combating age-related diseases. However, there are several challenges to overcome in its development and application. At the same time, there are significant opportunities that could accelerate progress and widen its therapeutic scope.
Challenges in Senomorphic Therapy
- Limited Clinical Evidence: While preclinical studies in animal models suggest that senomorphic compounds can effectively modulate the effects of senescence, clinical trials in humans are still relatively limited. Large-scale randomized controlled trials (RCTs) are needed to confirm the long-term safety and efficacy of these compounds, particularly in preventing or mitigating age-related diseases like Alzheimer’s disease, osteoarthritis, and cardiovascular disorders.123 Most current evidence comes from short-term studies or observational data, which makes it difficult to determine the real-world, long-term effects.191
- Translating Preclinical Findings to Humans: While animal models have shown encouraging results, the translation of these therapies to humans remains challenging.192 Species differences, variations in the aging process, and the complexity of human aging may complicate the successful application of senomorphic agents in clinical practice.193 Therefore, further clinical trials are needed to validate the efficacy and safety of these agents in human populations.
- Variability in Senescence Markers: Senescence is a complex biological process with different markers and hallmarks depending on tissue type, age, and the specific stressors that triggered senescence. Identifying consistent and reliable biomarkers to measure senescence in humans remains a significant hurdle. Moreover, SASP factors vary across tissues, making it difficult to design universal interventions. This variability complicates the development of a standardized approach for measuring therapeutic outcomes and assessing the effectiveness of senomorphic treatments.194
- Bioavailability and Pharmacokinetics: Many senomorphic compounds, such as curcumin and resveratrol, have poor bioavailability when administered orally, meaning they are not efficiently absorbed or utilized in the body.195 This is a critical challenge because without proper absorption, these compounds cannot exert their beneficial effects on senescence. Researchers are exploring various drug delivery strategies, such as liposomal formulations, nanoencapsulation, and prodrug strategies, to improve the bioavailability and targeted delivery of senomorphic compounds to specific tissues affected by senescence.196
- Potential Off-Target Effects: While senomorphic compounds are designed to modulate the senescence process without clearing senescent cells completely, there is a risk of unwanted off-target effects.197 Senescence plays a protective role in certain contexts, such as tumor suppression and wound healing. Chronic suppression of senescence or the SASP could potentially interfere with these beneficial processes. For instance, inhibiting SASP may reduce inflammation but could also impair the immune system’s ability to fight infections or prevent cancerous growths.194 Similarly, inhibition of mTOR can impair immune function, and restoring NAD+ levels could potentially affect metabolic processes.198 As such, careful monitoring of patients undergoing senomorphic therapy is necessary to prevent adverse effects.
- Heterogeneity of Senescent Cells: Senescent cells are heterogeneous, meaning that not all senescent cells behave the same way in different tissues and organs. For example, senescence in muscle cells might cause dysfunction through a different mechanism compared to senescence in endothelial cells. This diversity complicates the development of broad-spectrum senomorphic therapies.31 Senomorphic compounds that work effectively in one tissue might not produce the same results in others, requiring tailored or tissue-specific therapies.199
- Regulatory Hurdles: Senomorphic therapies, particularly those derived from natural compounds (eg, resveratrol, curcumin), may face challenges in regulatory approval. These compounds are often classified as nutraceuticals or dietary supplements, which are not subject to the same rigorous testing and regulatory requirements as pharmaceutical drugs.200 However, for senomorphic therapies to be used as effective interventions, they will likely need to undergo extensive clinical testing and receive approval from health authorities like the FDA or EMA.126 Regulatory frameworks may need to be adapted to accommodate new therapeutic paradigms based on modulating senescence.
- Drug Synergy and Combinatorial Approaches: Combining senomorphic compounds with other therapeutic strategies, such as senolytics (which eliminate senescent cells), caloric restriction mimetics, or antioxidants, could potentially enhance therapeutic outcomes. However, the interactions between these treatments are not fully understood. There is a need for more systematic studies to determine how senomorphics and senolytics can be used in combination to optimize the treatment of aging and age-related diseases. Additionally, the long-term safety of combinatorial treatments remains unclear.36
Opportunities in Senomorphic Therapy
- Targeting Age-Related Diseases: Senomorphic therapy offers tremendous potential in treating a wide range of chronic age-related diseases. By modulating the harmful effects of senescence, these therapies could slow down or even reverse conditions such as neurodegenerative diseases (eg, Alzheimer’s, Parkinson’s), cardiovascular disease, arthritis, and metabolic disorders (eg, type 2 diabetes).193 Since many of these diseases are associated with the accumulation of senescent cells and their inflammatory secretions, senomorphic compounds have the potential to treat the root cause of these diseases, rather than just alleviating symptoms.201
- Personalized Medicine Approaches: As our understanding of individual aging profiles and senescence mechanisms deepens, there is an opportunity to design personalized treatments based on a person’s genetic makeup, lifestyle factors, and specific senescence-related conditions. Biomarker-based diagnostics will allow for more precise identification of individuals who would benefit most from senomorphic therapy.202 Personalized medicine has proven to help in disease diagnosis, minimize side effects and maximize the therapeutic approaches.203
- Combination with Senolytics: Senomorphic therapy could be used in combination with senolytic compounds (which selectively kill senescent cells). While senolytics eliminate the senescent cells that accumulate over time, senomorphics can help modulate the negative effects of the remaining senescent cells. The combination of these therapies could provide a synergistic effect, improving tissue function, reducing inflammation, and slowing down aging processes. This approach could enhance the overall effectiveness of treatments for age-related diseases.65
- Advancements in Drug Delivery Systems: Nanotechnology and advanced drug delivery systems offer the potential to improve the bioavailability, stability, and targeted delivery of senomorphic compounds. By encapsulating these compounds in nanoparticles or using liposomal carriers, researchers can enhance the efficiency of drug delivery to specific tissues or organs. These delivery technologies may allow for more precise treatment of senescence in key tissues (such as the brain, heart, or joints), thereby improving therapeutic outcomes.204
- Expansion into Preventive Medicine: Instead of focusing solely on treating diseases, senomorphic therapies could be used for preventive purposes. Early intervention with senomorphic compounds could delay the onset of age-related diseases by modulating senescence in at-risk individuals, potentially leading to an extension of healthspan (the period of life spent in good health).35,130 This preventive approach could be especially beneficial for populations with genetic predispositions to certain conditions or those already showing early signs of aging-related decline.
- Potential for Nutraceutical Development: Many senomorphic compounds are naturally occurring and are already consumed as part of a healthy diet (eg, resveratrol, curcumin, quercetin). These compounds could be developed as dietary supplements or functional foods that support healthy aging and reduce the risk of age-related diseases. Their natural origin may make them appealing to consumers seeking less invasive, more holistic health solutions.130,205 As research progresses, these compounds could be integrated into preventive health regimens.
- Integration with AI and Systems Biology: The application of artificial intelligence (AI) and systems biology approaches in drug discovery can help accelerate the identification of new senomorphic compounds. AI-driven analyses can uncover novel compounds and predict their senescence-modulating properties based on biological data.206 Moreover, systems biology approaches that analyze the complex networks of cellular aging processes could help uncover new targets for senomorphic therapies and optimize their clinical applications.207
Combination Therapies: Synergizing Senolytics and Senomorphics
While both senolytics and senomorphics offer promising approaches for targeting cellular senescence, recent research suggests that combining these two therapeutic strategies may enhance their individual benefits, leading to more robust outcomes in promoting healthy aging. By simultaneously eliminating senescent cells and modulating the harmful effects of their secretory phenotype, combination therapies hold the potential to address the multifaceted nature of aging and age-related diseases.39,119 This integrated approach offers a comprehensive strategy for improving healthspan, reducing chronic inflammation, and enhancing tissue regeneration.
Rationale for Combination Therapies
The rationale for combining senolytic and senomorphic therapies lies in the complementary mechanisms of action of these two approaches. Senolytic agents focus on clearing the burden of senescent cells, directly addressing the source of cellular dysfunction, while senomorphics target the pro-inflammatory and tissue-degrading effects of the SASP, which result from the senescence process. When used in combination, these therapies can not only reduce the number of senescent cells but also ameliorate the systemic inflammation and tissue damage that occurs as a result of senescence. This dual approach has the potential to enhance the overall therapeutic effect, improving the health of tissues, organs, and systems affected by aging.208,209 Furthermore, cellular senescence is a complex process that cannot be fully addressed by targeting a single aspect, such as cell clearance or SASP modulation. Senolytic therapies may eliminate senescent cells but may not fully resolve the long-term inflammatory consequences of their presence, while senomorphic agents may suppress the SASP but leave senescent cells intact, allowing them to persist and potentially cause harm.210 Combining both approaches addresses the complexity of senescence, offering a more holistic therapeutic strategy for aging-related conditions.45
Potential Applications of Combination Therapies
The combination of senolytic and senomorphic therapies has broad potential applications in age-related diseases and conditions. Some key areas where combination therapies may offer substantial benefits include:
- Cardiovascular Disease: Cellular senescence plays a key role in the development of cardiovascular diseases, particularly through the accumulation of senescent cells in blood vessels and the heart. By using senolytics to remove senescent endothelial and smooth muscle cells, combined with senomorphics like rapamycin to modulate the SASP, this dual approach could reduce arterial stiffness, improve vascular function, and lower the risk of heart attacks and stroke.211,212
- Neurodegenerative Diseases: Conditions like Alzheimer’s disease, Parkinson’s disease, and other forms of dementia are characterized by neuroinflammation and the accumulation of senescent cells in the brain.26 Combination therapies targeting both senescent cells and the neuroinflammatory SASP could slow disease progression, improve cognitive function, and promote neuronal repair.213 Senolytics like dasatinib and quercetin, combined with senomorphic agents such as NMN, could help rejuvenate neuronal tissue and prevent cognitive decline.214
- Osteoporosis and Sarcopenia: Aging is associated with a decline in bone density and muscle mass, partly driven by the accumulation of senescent cells in bone and muscle tissue. The combination of senolytics and senomorphics could rejuvenate both bone and muscle tissue by eliminating senescent cells and modulating the inflammatory environment that promotes bone resorption and muscle wasting.215 Such therapies could reduce the risk of fractures, improve muscle strength, and enhance mobility in the elderly.216
- Cancer: Cellular senescence is a double-edged sword in cancer. While senescence acts as a protective mechanism against tumorigenesis, the accumulation of senescent cells and their inflammatory SASP can promote tumor progression.53 Combination therapies that target senescent cells in the tumor microenvironment and modulate the SASP could offer new therapeutic options in cancer treatment, potentially inhibiting tumor growth while enhancing the efficacy of existing therapies like chemotherapy and immunotherapy.194,217
Preclinical Evidence Supporting Combination Therapies
In preclinical animal models, the combination of senolytic and senomorphic agents has shown promising results. For example, the combination of dasatinib and quercetin (senolytic agents) with fisetin (a senomorphic agent) has been tested in mice and humans and demonstrated enhanced effects compared to either treatment alone. In this combination, dasatinib and quercetin effectively reduced the number of senescent cells in adipose tissue and the vasculature, while fisetin modulated the inflammatory SASP, improving overall tissue function and reducing the risk of age-related diseases such as cardiovascular disease and osteoporosis.18 In another study, the combination of fisetin (a senolytic flavonoid) with sorafenib (a senomorphic agent) showed better synergistic effects in vitro and in vivo than either agent used alone against human cervical cancer.218 These findings highlight the potential of combination therapies to improve multiple aspects of aging simultaneously, rather than focusing on a single target. Table 5 highlights some preclinical animal models where the combination of senolytic and senomorphic agents have shown promising results.
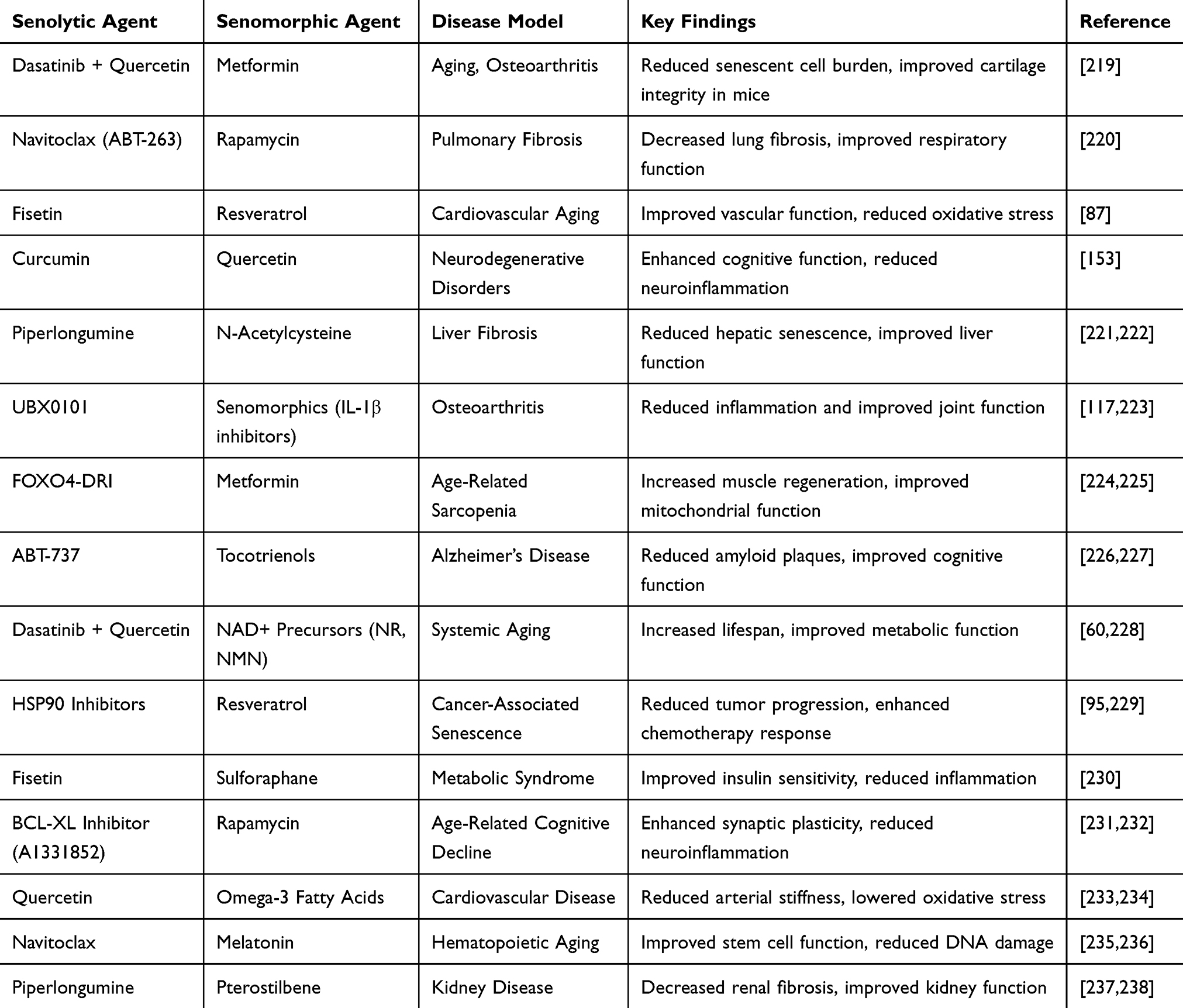 |
Table 5 Some Preclinical Evidence Supporting Combination Therapies of Senolytic and Senomorphic Agents |
Clinical Validation of Senescence Biomarkers
The translation of senescence-targeted therapies into clinical practice is hindered by the lack of fully validated, standardized biomarkers for senescence burden. Commonly used markers such as p16^INK4a, SA-β-galactosidase, and SASP cytokines (eg, IL-6, IL-8) show promise in experimental models but have variable sensitivity and specificity across tissues and disease contexts.1 Their expression can be transient, context-dependent, and influenced by non-senescent cellular states, complicating interpretation in human studies. Furthermore, the invasive nature of current tissue-based assays limits their clinical utility. Advancements in liquid biopsy technologies, molecular imaging, and multi-omics profiling may enable non-invasive, longitudinal tracking of senescence burden, improving patient selection and therapeutic monitoring. However, large-scale, prospective studies are urgently needed to establish reproducibility, predictive validity, and regulatory acceptance of these biomarkers.
Clinical Trials and Translational Research
Although preclinical data are promising, the translation of combination therapies into clinical practice is still in its early stages. A few early-phase clinical trials are exploring the use of senolytics and senomorphics in combination for various aging-related conditions. For instance, a clinical trial is currently investigating the effects of dasatinib and quercetin combined with rapamycin in patients with idiopathic pulmonary fibrosis (IPF), a condition marked by senescence-driven fibrosis and inflammation.239 Preliminary results suggest that this combination can reduce fibrosis and improve lung function, demonstrating the potential of this combined approach to target both cellular senescence and its deleterious effects in humans. Similarly, clinical trials have evaluated the effects of combining NMN with other senolytic compounds in elderly populations and proved improved muscle strength, metabolism, and overall health.240,241 These studies aimed to assess whether the synergistic effects of combining senolytics and senomorphics translate into meaningful clinical outcomes, such as improved mobility, cognitive function, and quality of life. Table 6 is a tabular representation of some clinical evidence supporting combination therapies of senolytic and senomorphic agents.
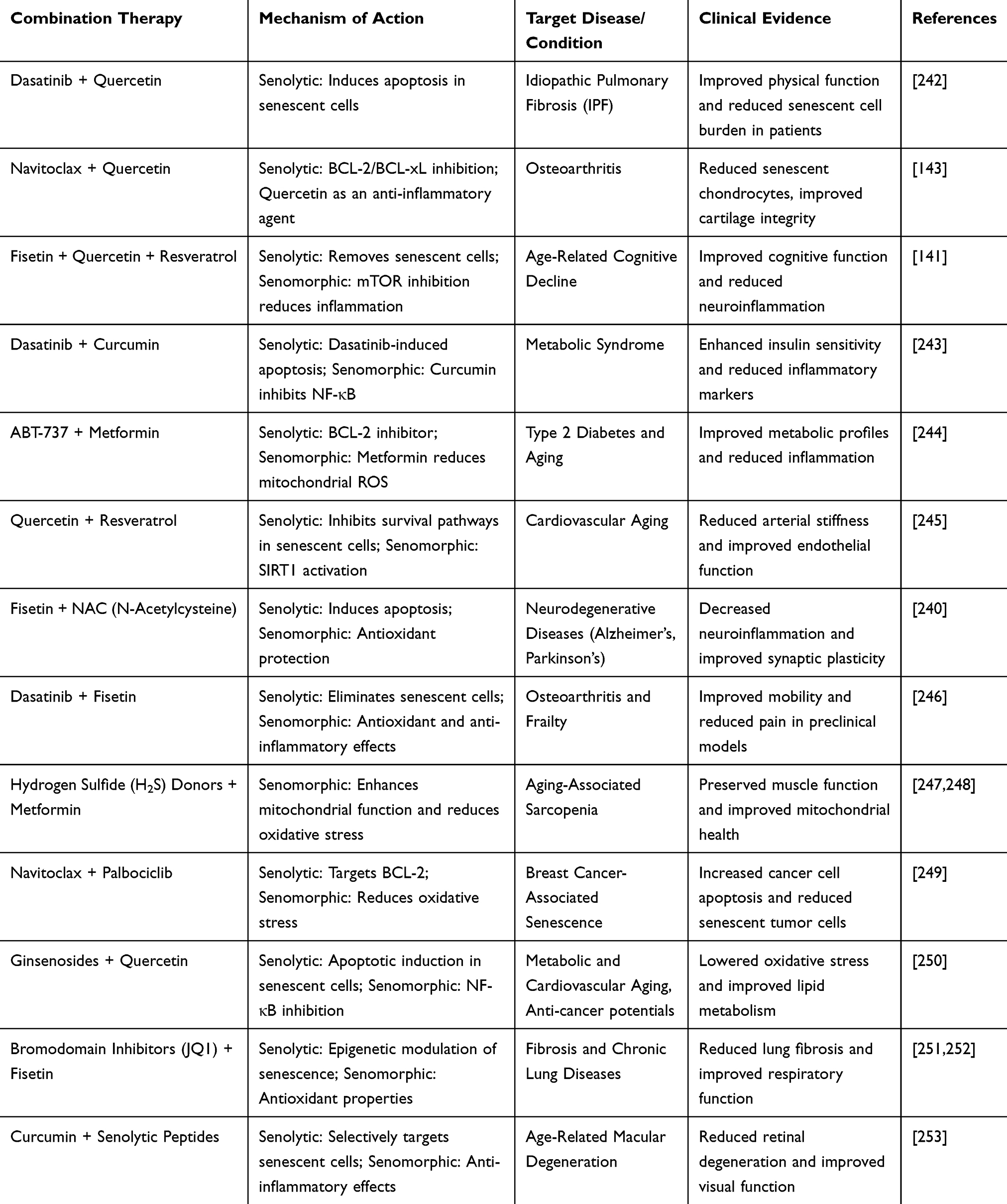 |
Table 6 Clinical Evidence Supporting Combination Therapies of Senolytic and Senomorphic Agents |
Clinical Translation and Challenges in the Development of Senolytic and Senomorphic Therapies
The clinical translation of senolytic and senomorphic therapies is a promising but challenging endeavor. Addressing safety concerns, developing reliable biomarkers, optimizing dosing regimens, and overcoming regulatory hurdles are key to the success of these therapies in clinical settings. As ongoing clinical trials continue to yield valuable insights, the path toward effective senescence-targeting interventions for healthy aging becomes clearer. The successful development of these therapies could revolutionize aging medicine, offering novel treatments for a range of age-related diseases and enhancing the quality of life for aging populations.
Challenges in Clinical Translation
While preclinical studies have demonstrated promising results for both senolytic and senomorphic therapies, translating these findings into effective and safe clinical treatments presents several challenges. The path from bench to bedside is often complex and fraught with regulatory, safety, and efficacy concerns.
- Safety and Toxicity: The safety of senolytic and senomorphic therapies is one of the foremost concerns in clinical translation. Although many senolytic agents, such as dasatinib and quercetin, have shown efficacy in preclinical models, their long-term effects in humans are not yet fully understood. The risk of off-target effects, immune system disruption, or toxicity when these agents are used in combination or over extended periods is still a significant challenge.254 For example, the selective removal of senescent cells by senolytics may inadvertently affect healthy cells that share certain senescent-like characteristics or may lead to inflammatory responses in tissues as a result of the clearance process.194 Additionally, long-term inhibition of pathways such as mTOR, which is involved in regulating cell growth, immune function, and metabolism, could have unintended consequences that need to be carefully studied in clinical trials.255 Thus, a key focus for the clinical development of these therapies will be determining optimal dosing regimens and ensuring the safe use of these agents without disrupting vital physiological processes.
- Standardization of Senescence Biomarkers: A major challenge in both the development and application of senescence-targeting therapies is the identification of reliable biomarkers that can accurately assess the presence of senescent cells and their effects.256 Current biomarkers for senescence, such as p16^INK4a, β-galactosidase activity, and SASP factors, are useful in research settings, but their clinical applicability remains limited. There is a need for non-invasive, easily measurable biomarkers that can guide the use of senolytics and senomorphics, monitor treatment progress, and assess therapeutic efficacy. Furthermore, the heterogeneity of cellular senescence such as different tissues, types of senescent cells, and stages of senescence complicates the development of universal biomarkers. The presence of senescent cells in tissues may vary among individuals, with some aging populations showing greater senescence burden than others. Thus, tailored approaches for the identification and treatment of senescence in specific tissues will be necessary.257
- Dosing and Timing: Finding the right dosing schedule for senolytics and senomorphics is critical to their success in clinical practice. The balance between efficacy and safety requires careful modulation of dosage and treatment duration. Senolytic therapies, which target and eliminate senescent cells, may need to be administered intermittently, while senomorphics, which modulate the SASP, may be more suitable for continuous or long-term treatment.36 Additionally, in combination therapies, determining the timing and synergy of different agents is complex.258 The effects of senolytic and senomorphic agents may be enhanced or diminished depending on when and how they are administered.35 Therefore, clinical trials will need to explore various combinations and schedules to identify the most effective regimen for targeting senescence and promoting healthy aging without causing harm.
Strategies for Overcoming Developmental Challenges
Strategies for overcoming developmental challenges in order to bring these promising treatments into clinical practice include:
- Personalized Approaches: One way to address the challenges of clinical translation is through personalized medicine. Given the heterogeneity of aging and senescence across individuals, therapies may need to be tailored to specific patients based on biomarkers of senescence and their unique disease profile. Personalized approaches would allow for the more precise application of senolytic and senomorphic therapies, ensuring that individuals receive the right treatment at the right time and in the right dose.129 Advances in genomics, proteomics, and other omics technologies may facilitate the identification of biomarkers that predict senescence burden and SASP expression in different tissues. These biomarkers could guide treatment decisions, helping clinicians select the most appropriate therapies for patients based on their individual senescence profiles.259
- Developing Novel Delivery Systems: The efficacy of both senolytic and senomorphic therapies may be influenced by how these agents are delivered to target tissues. For example, nanoparticles, liposomes, or other drug delivery systems can be engineered to deliver these agents more specifically to tissues where senescence is a major contributor to dysfunction. This could improve the therapeutic outcomes by concentrating the treatment in the affected areas and minimizing systemic side effects.260 For instance, plant-derived nanocarriers have been shown to target specific organs or tissues in preclinical models of cancer, and similar strategies could be adapted to target senescent cells in tissues affected by aging.204 Developing such targeted delivery systems could enhance the effectiveness and reduce the side effects of senescence-targeting therapies.
- Collaboration Between Academia, Industry, and Regulatory Bodies: To accelerate the clinical translation of senolytic and senomorphic therapies, strong collaboration between academic researchers, pharmaceutical companies, and regulatory agencies is crucial. Academic research provides the foundational understanding of cellular senescence and aging, while industry partners have the resources and expertise to translate this knowledge into viable therapies.126 Regulatory bodies, such as the FDA, will play a critical role in establishing guidelines for the safe and effective use of these therapies in clinical practice. A collaborative effort can streamline the development process, reduce regulatory hurdles, and ensure that new therapies are both effective and safe.119
Future Directions in Senescence Research
As we move toward clinical application, targeting cellular senescence presents an exciting frontier in promoting healthy aging and mitigating the impact of aging-related diseases. The continuing development of senolytic and senomorphic therapies holds immense potential, but several areas require further research and refinement.
- Expanding the Senescence Landscape: Although much progress has been made in understanding the role of senescence in aging and disease, many unanswered questions remain. Future research should aim to further elucidate the various subtypes of cellular senescence, including differences between stress-induced senescence, oncogene-induced senescence, and developmental senescence, as each may present unique therapeutic opportunities. Investigating the molecular pathways that govern these senescent states could reveal new targets for senolytic and senomorphic therapies. Moreover, expanding our understanding of senescence in different tissue types, organs, and systems will be crucial. While much of the current research focuses on senescence in skin, muscle, and adipose tissue, the role of senescence in organs like the brain, liver, and heart remains underexplored. By addressing tissue-specific senescence and its relationship with local microenvironments, we can develop therapies that more effectively target aging in diverse tissues.
- Optimizing Combination Therapies: The synergistic use of senolytic and senomorphic agents could be an important avenue for improving therapeutic efficacy. As discussed earlier, senolytics focus on clearing senescent cells, while senomorphics aim to modify the SASP and other senescence-related pathways. Combining these therapies could enhance their individual effects, but careful optimization of dosing schedules, timing, and patient selection will be necessary. Emerging evidence suggests that a “cocktail” approach, that is, incorporating not only senolytics and senomorphics but also other therapeutic modalities, such as antioxidants, immune modulators, or metabolic enhancers may have a compounded effect in promoting healthy aging.261 Future clinical trials should explore these combination strategies, testing them in diverse age-related diseases to understand the potential for synergy and minimizing possible negative interactions between therapies.
- Personalized and Precision Medicine: Given the heterogeneity of aging, a one-size-fits-all approach to senescence-targeting therapies is unlikely to succeed. Future studies should focus on developing personalized medicine strategies, taking into account individual variations in genetics, epigenetics, microbiome composition, and lifestyle factors. Identifying biomarkers specific to the senescence burden in each patient could allow for the development of tailored treatment plans that maximize efficacy and minimize side effects. Furthermore, the use of “big data” approaches integrating genomic, proteomic, and phenotypic information could provide valuable insights into how senescence manifests across diverse populations. This information would be essential in designing personalized interventions and refining existing therapies to be more effective and individualized.
- Non-Invasive Monitoring Tools: The development of non-invasive technologies to monitor senescence burden and therapy efficacy will be pivotal in advancing clinical practice. Currently, many of the methods used to track cellular senescence are invasive, such as tissue biopsies or complex imaging techniques. Future research should focus on developing non-invasive biomarkers, such as blood tests or imaging technologies, that can assess senescence at the tissue level. This will enable clinicians to track treatment response in real time, adjust dosages accordingly, and determine the optimal time for intervention.
- Expanding the Scope of Senescence-Targeting Agents: While several promising senolytic and senomorphic agents have been identified, the search for new compounds should remain a priority. Natural products, particularly phytochemicals, remain an untapped resource for developing new therapeutic agents. The pharmacological properties of compounds derived from medicinal plants and other natural sources may offer unique mechanisms for modulating senescence. Exploring these avenues could lead to the discovery of novel, safer, and more effective senescence-targeting agents. In addition, understanding how existing pharmaceuticals, such as statins, metformin, and other widely used drugs, influence cellular senescence could open new therapeutic pathways. These drugs have already been shown to impact pathways related to aging and senescence, making them candidates for repurposing in the context of age-related disorders.
Conclusion
This review accentuates that targeting cellular senescence offers a transformative strategy for promoting healthy aging and mitigating age-related pathologies. By integrating recent advances in the understanding of senescence biology with detailed analyses of senolytic and senomorphic interventions, we highlight both the therapeutic promise and translational challenges in the field. Some of the challenges include issues related to safety, optimal dosing, patient stratification, and the development of reliable biomarkers. Oxidative stress and SASP are revealed as deeply interconnected processes, emphasizing the need for dual-targeting approaches that restore redox balance while modulating the senescent cell secretome. Furthermore, bioavailability and pharmacokinetic limitations remain key barriers, warranting innovation in formulation and delivery technologies.
Future success will depend not only on refining therapeutic targets but also on optimizing dosing strategies and establishing reliable, non-invasive biomarkers for senescence, enabling safe, effective, and personalized application in clinical aging medicine. Additionally, evaluating combination strategies that harness the synergistic potential of senolytics, senomorphics, and lifestyle interventions is paramount. As we continue to refine these approaches, it is likely that senolytic and senomorphic agents will become an integral part of clinical strategies aimed at enhancing the quality of life for aging populations and mitigating the impact of age-related disorders. Ultimately, the application of senescence-targeting therapies could revolutionize the way we approach aging and aging-related diseases, offering a pathway to healthier, more productive lives as we age. The primary aim of aging and gerontology research will continue to be not just to prolong lifespan but also to improve healthspan, hence fostering a more robust and fulfilling aging experience as advancements in this field progress.
Abbreviations
ATM, Ataxia Telangiectasia Mutated; CDK, Cyclin-Dependent Kinase; CGAS, Cyclic GMP-AMP Synthase; DDR, DNA Damage Response; DNA, Deoxyribonucleic Acid; IL-6, Interleukin-6; NF-κB, Nuclear Factor Kappa B; ROS, Reactive Oxygen Species; SASP, Senescence-Associated Secretory Phenotype; STING, Stimulator of Interferon Genes; TGF-β, Transforming Growth Factor Beta.
Data Sharing Statement
All used data is fully presented in the manuscript.
Acknowledgments
Authors are grateful to Kampala International University for its supports. The graphical abstract was Created in BioRender. Basajja, M. (2025) https://BioRender.com/ bigoce7.
Funding
No funding was received.
Disclosure
The authors declare no conflicts of interest in this work.
References
1. Kumari R, Jat P. Mechanisms of cellular senescence: cell cycle arrest and senescence associated secretory phenotype. Front Cell Dev Biol. 2021;9. doi:10.3389/fcell.2021.645593
2. Saito Y, Yamamoto S, Chikenji TS. Role of cellular senescence in inflammation and regeneration. Inflamm Regen. 2024;44(1):28. doi:10.1186/s41232-024-00342-5
3. Kaur J, Farr JN. Cellular senescence in age-related disorders. Transl Res. 2020;226:96–104. doi:10.1016/j.trsl.2020.06.007
4. von Kobbe C. Targeting senescent cells: approaches, opportunities, challenges. Aging. 2019;11(24):12844–12861. doi:10.18632/aging.102557
5. Sun Y, Li Q, Kirkland JL. Targeting senescent cells for a healthier longevity: the roadmap for an era of global aging. Life Med. 2022;1(2):103–119. doi:10.1093/lifemedi/lnac030
6. Wang C, Hao X, Zhang R. Targeting cellular senescence to combat cancer and aging. Mol Oncol. 2022;16(18):3319–3332. doi:10.1002/1878-0261.13266
7. Princilly J, Veerabhadrappa B, Rao NN, Dyavaiah M. Chapter one - cellular senescence in aging: molecular basis, implications and therapeutic interventions. In: Çakatay U, Atayik MC, editors. Advances in Protein Chemistry and Structural Biology. Academic Press; 2023:1–33.
8. Nadeem J, Sultana R, Parveen A, Kim SY. Recent advances in anti-aging therapeutic strategies targeting DNA damage response and senescence-associated secretory phenotype-linked signaling cascade. Cell Biochem Funct. 2025;43(3):e70046. doi:10.1002/cbf.70046
9. Steffens Reinhardt L, Groen K, newton C, Avery-Kiejda KA. The role of truncated p53 isoforms in the DNA damage response. Biochim Biophys Acta Rev Cancer. 2023;1878(3):188882. doi:10.1016/j.bbcan.2023.188882
10. Mas-Bargues C, Viña-Almunia J, Inglés M, et al. Role of p16INK4a and BMI-1 in oxidative stress-induced premature senescence in human dental pulp stem cells. Redox Biol. 2017;12:690–698. doi:10.1016/j.redox.2017.04.002
11. Shoeb M, Meier HCS, Antonini JM. Telomeres in toxicology: occupational health. Pharm Ther. 2021;220:107742. doi:10.1016/j.pharmthera.2020.107742
12. Shay JW. Role of telomeres and telomerase in aging and cancer. Cancer Discov. 2016;6(6):584–593. doi:10.1158/2159-8290.CD-16-0062
13. Victorelli S, Passos JF. Telomeres and cell senescence - size matters not. EBioMedicine. 2017;21:14–20. doi:10.1016/j.ebiom.2017.03.027
14. Afifi MM, Crncec A, Cornwell JA, et al. Irreversible cell cycle exit associated with senescence is mediated by constitutive MYC degradation. Cell Rep. 2023;42(9):113079. doi:10.1016/j.celrep.2023.113079
15. Sarni D, Kerem B. Oncogene-induced replication stress drives genome instability and tumorigenesis. Int J Mol Sci. 2017;18(7):1339. doi:10.3390/ijms18071339
16. Juan CA, de la Lastra JMP, Plou FJ, Pérez-Lebeña E. The Chemistry of Reactive Oxygen Species (ROS) revisited: outlining their role in biological macromolecules (DNA, Lipids and Proteins) and induced pathologies. Int J Mol Sci. 2021;22(9):4642. doi:10.3390/ijms22094642
17. Alum EU. Antioxidant effect of Buchholzia coriacea ethanol leaf-extract and fractions on freund’s adjuvant-induced arthritis in albino rats: a comparative study. Slov Vet Res. 2022;59(1). doi:10.26873/SVR-1150-2022
18. Prašnikar E, Borišek J, Perdih A. Senescent cells as promising targets to tackle age-related diseases. Aging Res Rev. 2021;66:101251. doi:10.1016/j.arr.2020.101251
19. Fan H, Qiao Z, Li J, et al. Recent advances in senescence-associated secretory phenotype and osteoporosis. Heliyon. 2024;10(4):e25538. doi:10.1016/j.heliyon.2024.e25538
20. O’Reilly S, Markiewicz E, Idowu OC. Aging, senescence, and cutaneous wound healing—a complex relationship. Front Immunol. 2024;15:1429716. doi:10.3389/fimmu.2024.1429716
21. Becker JS, Nicetto D, Zaret KS. H3K9me3-dependent heterochromatin: barrier to cell fate changes. Trends Genet. 2016;32(1):29–41. doi:10.1016/j.tig.2015.11.001
22. Crouch J, Shvedova M, Thanapaul RJRS, Botchkarev V, Roh D. Epigenetic regulation of cellular senescence. Cells. 2022;11(4):672. doi:10.3390/cells11040672
23. Han Z, Wang K, Ding S, Zhang M. Cross-talk of inflammation and cellular senescence: a new insight into the occurrence and progression of osteoarthritis. Bone Res. 2024;12(1):1–17. doi:10.1038/s41413-024-00375-z
24. Larsson L, Degens H, Li M, et al. Sarcopenia: aging-related loss of muscle mass and function. Physiol Rev. 2019;99(1):427–511. doi:10.1152/physrev.00061.2017
25. Katsuumi G, Shimizu I, Yoshida Y, Minamino T. Vascular senescence in cardiovascular and metabolic diseases. Front Cardiovasc Med. 2018;5. doi:10.3389/fcvm.2018.00018
26. Wang Y, Kuca K, You L, et al. The role of cellular senescence in neurodegenerative diseases. Arch Toxicol. 2024;98(8):2393–2408. doi:10.1007/s00204-024-03768-5
27. Kruczkowska W, Gałęziewska J, Kciuk M, et al. Senescent adipocytes and type 2 diabetes – current knowledge and perspective concepts. Biomol Concepts. 2024;15(1). doi:10.1515/bmc-2022-0046
28. Coryell PR, Diekman BO, Loeser RF. Mechanisms and therapeutic implications of cellular senescence in osteoarthritis. Nat Rev Rheumatol. 2021;17(1):47–57. doi:10.1038/s41584-020-00533-7
29. Stoczynska-Fidelus E, Węgierska M, Kierasińska A, Ciunowicz D, Rieske P. Role of senescence in tumorigenesis and anticancer therapy. J Oncol. 2022;2022:5969536. doi:10.1155/2022/5969536
30. Wang L, Lankhorst L, Bernards R. Exploiting senescence for the treatment of cancer. Nat Rev Cancer. 2022;22(6):340–355. doi:10.1038/s41568-022-00450-9
31. Huang W, Hickson LJ, Eirin A, Kirkland JL, Lerman LO. Cellular senescence: the good, the bad and the unknown. Nat Rev Nephrol. 2022;18(10):611–627. doi:10.1038/s41581-022-00601-z
32. McHugh D, Gil J. Senescence and aging: causes, consequences, and therapeutic avenues. J Cell Biol. 2018;217(1):65–77. doi:10.1083/jcb.201708092
33. Li X, Li C, Zhang W, Wang Y, Qian P, Huang H. Inflammation and aging: signaling pathways and intervention therapies. Signal Transduct Target Ther. 2023;8(1):239. doi:10.1038/s41392-023-01502-8
34. Schmitt CA, Wang B, Demaria M. Senescence and cancer — role and therapeutic opportunities. Nat Rev Clin Oncol. 2022;19(10):619–636. doi:10.1038/s41571-022-00668-4
35. Imb M, Véghelyi Z, Maurer M, Kühnel H. Exploring senolytic and senomorphic properties of medicinal plants for anti-aging therapies. Int J Mol Sci. 2024;25(19):10419. doi:10.3390/ijms251910419
36. Rad AN, Grillari J. Current senolytics: mode of action, efficacy and limitations, and their future. Mech Ageing Dev. 2024;217:111888. doi:10.1016/j.mad.2023.111888
37. Robbins PD, Jurk D, Khosla S, et al. Senolytic Drugs: reducing senescent cell viability to extend health span. Annu Rev Pharmacol Toxicol. 2021;61(1):779–803. doi:10.1146/annurev-pharmtox-050120-105018
38. Song S, Tchkonia T, Jiang J, Kirkland JL, Sun Y. Targeting senescent cells for a healthier aging: challenges and opportunities. Adv Sci. 2020;7(23):2002611. doi:10.1002/advs.202002611
39. Lelarge V, Capelle R, Oger F, Mathieu T, Le Calvé B. Senolytics: from pharmacological inhibitors to immunotherapies, a promising future for patients. Treatment Npj Aging. 2024;10:1–8. doi:10.1038/s41514-024-00138-4
40. Chaib S, Tchkonia T, Kirkland JL. Cellular senescence and senolytics: the path to the clinic. Nat Med. 2022;28(8):1556–1568. doi:10.1038/s41591-022-01923-y
41. Kang C. Senolytics and senostatics: a two-pronged approach to target cellular senescence for delaying aging and age-related diseases. Mol Cells. 2019;42(12):821–827. doi:10.14348/molcells.2019.0298
42. Kirkland JL, Tchkonia T. Senolytic drugs: from discovery to translation. J Intern Med. 2020;288(5):518–536. doi:10.1111/joim.13141
43. Millar CL, Iloputaife I, Baldyga K, et al. A pilot study of senolytics to improve cognition and mobility in older adults at risk for Alzheimer’s disease. eBioMedicine. 2025;113:105612. doi:10.1016/j.ebiom.2025.105612
44. Blagosklonny MV. Anti-aging: senolytics or gerostatics (unconventional view). Oncotarget. 2021;12(18):1821–1835. doi:10.18632/oncotarget.28049
45. Davan-Wetton CSA, Pessolano E, Perretti M, Montero-Melendez T. Senescence under appraisal: hopes and challenges revisited. Cell Mol Life Sci. 2021;78(7):3333–3354. doi:10.1007/s00018-020-03746-x
46. Bogdanova DA, Kolosova ED, Pukhalskaia TV, Levchuk KA, Demidov ON, Belotserkovskaya EV. The differential effect of senolytics on SASP cytokine secretion and regulation of EMT by CAFs. Int J Mol Sci. 2024;25(7):4031. doi:10.3390/ijms25074031
47. Power H, Valtchev P, Dehghani F, Schindeler A. Strategies for senolytic drug discovery. Aging Cell. 2023;22(10):e13948. doi:10.1111/acel.13948
48. Short S, Fielder E, Miwa S, von Zglinicki T. Senolytics and senostatics as adjuvant tumour therapy. EBioMedicine. 2019;41:683–692. doi:10.1016/j.ebiom.2019.01.056
49. Calabrò A, Accardi G, Aiello A, Caruso C, Galimberti D, Candore G. Senotherapeutics to counteract senescent cells are prominent topics in the context of anti-aging strategies. Int J Mol Sci. 2024;25(3):1792. doi:10.3390/ijms25031792
50. Fu TE, Zhou Z. Senescent cells as a target for anti-aging interventions: from senolytics to immune therapies. J Transl Int Med. 2025;13(1):33–47. doi:10.1515/jtim-2025-0005
51. Vogler M, Braun Y, Smith VM, et al. The BCL2 family: from apoptosis mechanisms to new advances in targeted therapy. Sig Transduct Target Ther. 2025;10:1–31. doi:10.1038/s41392-025-02176-0
52. Qian S, Wei Z, Yang W, Huang J, Yang Y, Wang J. The role of BCL-2 family proteins in regulating apoptosis and cancer therapy. Front Oncol. 2022;12:985363. doi:10.3389/fonc.2022.985363
53. Jin P, Duan X, Li L, Zhou P, Zou C-G, Xie K. Cellular senescence in cancer: molecular mechanisms and therapeutic targets. MedComm. 2024;5(5):e542. doi:10.1002/mco2.542
54. Mohamad Anuar NN, Nor Hisam NS, Liew SL, Ugusman A. Clinical review: navitoclax as a pro-apoptotic and anti-fibrotic agent. Front Pharmacol. 2020;11:564108. doi:10.3389/fphar.2020.564108
55. Mijit M, Caracciolo V, Melillo A, Amicarelli F, Giordano A. Role of p53 in the regulation of cellular senescence. Biomolecules. 2020;10(3):420. doi:10.3390/biom10030420
56. Pawge G, Khatik GL. p53 regulated senescence mechanism and role of its modulators in age-related disorders. Biochem Pharmacol. 2021;190:114651. doi:10.1016/j.bcp.2021.114651
57. Mandal R, Kohoutova K, Petrvalska O, et al. FOXO4 interacts with p53 TAD and CRD and inhibits its binding to DNA. Protein Sci. 2022;31(5):e4287. doi:10.1002/pro.4287
58. Huang Y, He Y, Makarcyzk MJ, Lin H. Senolytic peptide FOXO4-DRI selectively removes senescent cells from in vitro expanded human chondrocytes. Front Bioeng Biotechnol. 2021;9. doi:10.3389/fbioe.2021.677576
59. Coperchini F, Greco A, Teliti M, et al. Inflamm-aging: how cytokines and nutrition shape the trajectory of aging. Cytokine Growth Factor Rev. 2025;82:31–42. doi:10.1016/j.cytogfr.2024.08.004
60. Islam MT, Tuday E, Allen S, et al. Senolytic drugs, dasatinib and quercetin, attenuate adipose tissue inflammation, and ameliorate metabolic function in old age. Aging Cell. 2023;22(2):e13767. doi:10.1111/acel.13767
61. Tavenier J, Nehlin JO, Houlind MB, et al. Fisetin as a senotherapeutic agent: evidence and perspectives for age-related diseases. Mech Aging Dev. 2024;222:111995. doi:10.1016/j.mad.2024.111995
62. Domaszewska-Szostek A, Puzianowska-Kuźnicka M, Kuryłowicz A. Flavonoids in skin senescence prevention and treatment. Int J Mol Sci. 2021;22(13):6814. doi:10.3390/ijms22136814
63. Hu C, Yang J, Qi Z, et al. Heat shock proteins: biological functions, pathological roles, and therapeutic opportunities. MedComm. 2022;3(3):e161. doi:10.1002/mco2.161
64. Dabravolski SA, Sukhorukov VN, Kalmykov VA, Orekhov NA, Grechko AV, Orekhov AN. Heat shock protein 90 as therapeutic target for CVDs and heart aging. Int J Mol Sci. 2022;23(2):649. doi:10.3390/ijms23020649
65. Zhang L, Pitcher LE, Prahalad V, Niedernhofer LJ, Robbins PD. Targeting cellular senescence with senotherapeutics: senolytics and senomorphics. FEBS J. 2023;290(5):1362–1383. doi:10.1111/febs.16350
66. Miwa S, Kashyap S, Chini E, von Zglinicki T. Mitochondrial dysfunction in cell senescence and aging. J Clin Invest. 2022;132(13):e158447. doi:10.1172/JCI158447
67. Cheng -F-F, Liu Y-L, Du J, Lin J-T. Metformin’s mechanisms in attenuating hallmarks of aging and age-related disease. Aging Dis. 2022;13(4):970–986. doi:10.14336/AD.2021.1213
68. Zhang T, Zhou L, Makarczyk MJ, Feng P, Zhang J. The anti-aging mechanism of metformin: from molecular insights to clinical applications. Molecules. 2025;30(4):816. doi:10.3390/molecules30040816
69. Morevati M, Fang EF, Mace ML, et al. Roles of NAD+ in acute and chronic kidney diseases. Int J Mol Sci. 2023;24(1):137. doi:10.3390/ijms24010137
70. Gómez-Virgilio L, Silva-Lucero M-C, Flores-Morelos D-S, et al. Autophagy: a key regulator of homeostasis and disease: an overview of molecular mechanisms and modulators. Cells. 2022;11(15):2262. doi:10.3390/cells11152262
71. Selvarani R, Mohammed S, Richardson A. Effect of rapamycin on aging and age-related diseases—past and future. GeroScience. 2020;43(3):1135–1158. doi:10.1007/s11357-020-00274-1
72. Lee DJW, Kuerec AH, Maier AB. Targeting aging with rapamycin and its derivatives in humans: a systematic review. Lancet Health Longevity. 2024;5(2):e152–e162. doi:10.1016/S2666-7568(23)00258-1
73. Hofer SJ, Daskalaki I, Bergmann M, et al. Spermidine is essential for fasting-mediated autophagy and longevity. Nat Cell Biol. 2024;26(9):1571–1584. doi:10.1038/s41556-024-01468-x
74. Alves I, Araújo EMQ, Dalgaard LT, Singh S, Børsheim E, Carvalho E. Protective effects of sulforaphane preventing inflammation and oxidative stress to enhance metabolic health: a narrative review. Nutrients. 2025;17(3):428. doi:10.3390/nu17030428
75. Aaseth J, Alexander J, Alehagen U. Coenzyme Q10 supplementation – in aging and disease. Mech Aging Dev. 2021;197:111521. doi:10.1016/j.mad.2021.111521
76. Pan J, Li D, Xu Y, et al. Inhibition of Bcl-2/xl with ABT-263 selectively kills senescent type II pneumocytes and reverses persistent pulmonary fibrosis induced by ionizing radiation in mice. Int J Radiat Oncol Biol Phys. 2017;99(2):353–361. doi:10.1016/j.ijrobp.2017.02.216
77. Su W, Hu Y, Fan X, Xie J. Clearance of senescent cells by navitoclax (ABT263) rejuvenates UHMWPE-induced osteolysis. Int Immunopharmacol. 2023;115:109694. doi:10.1016/j.intimp.2023.109694
78. Chang J, Wang Y, Shao L, et al. Clearance of senescent cells by ABT263 rejuvenates aged hematopoietic stem cells in mice. Nat Med. 2016;22(1):78–83. doi:10.1038/nm.4010
79. Zhu Y, Tchkonia T, Fuhrmann-Stroissnigg H, et al. Identification of a novel senolytic agent, navitoclax, targeting the Bcl-2 family of anti-apoptotic factors. Aging Cell. 2016;15(3):428–435. doi:10.1111/acel.12445
80. Farr JN, Xu M, Weivoda MM, et al. Targeting cellular senescence prevents age-related bone loss in mice. Nat Med. 2017;23(9):1072–1079. doi:10.1038/nm.4385
81. Roos CM, Zhang B, Palmer AK, et al. Chronic senolytic treatment alleviates established vasomotor dysfunction in aged or atherosclerotic mice. Aging Cell. 2016;15(5):973–977. doi:10.1111/acel.12458
82. Xu M, Pirtskhalava T, Farr JN, et al. Senolytics improve physical function and increase lifespan in old age. Nat Med. 2018;24(8):1246–1256. doi:10.1038/s41591-018-0092-9
83. Kellogg DL, Kellogg DL, Musi N, Nambiar AM. Cellular senescence in idiopathic pulmonary fibrosis. Curr Mol Bio Rep. 2021;7(3):31–40. doi:10.1007/s40610-021-00145-4
84. Baar MP, Brandt RMC, Putavet DA, et al. Targeted apoptosis of senescent cells restores tissue homeostasis in response to chemotoxicity and aging. Cell. 2017;169(1):132–147.e16. doi:10.1016/j.cell.2017.02.031
85. Zhang C, Xie Y, Chen H, et al. FOXO4-DRI alleviates age-related testosterone secretion insufficiency by targeting senescent leydig cells in aged mice. Aging. 2020;12(2):1272–1284. doi:10.18632/aging.102682
86. Li Y, Zhang C, Cheng H, et al. FOXO4-DRI improves spermatogenesis in aged mice through reducing senescence-associated secretory phenotype secretion from leydig cells. Exper Gerontol. 2024;195:112522. doi:10.1016/j.exger.2024.112522
87. Yousefzadeh MJ, Zhu Y, McGowan SJ, et al. Fisetin is a senotherapeutic that extends health and lifespan. eBioMedicine. 2018;36:18–28. doi:10.1016/j.ebiom.2018.09.015
88. Althunibat OY, Al Hroob AM, Abukhalil MH, Germoush MO, Bin-Jumah M, Mahmoud AM. Fisetin ameliorates oxidative stress, inflammation and apoptosis in diabetic cardiomyopathy. Life Sci. 2019;221:83–92. doi:10.1016/j.lfs.2019.02.017
89. Yuan F, Li D, Li G, Cheng C, Wei X. Synergistic efficacy of homoharringtonine and venetoclax on acute myeloid leukemia cells and the underlying mechanisms. Ann Transl Med. 2022;10(8):490. doi:10.21037/atm-22-1459
90. As Sobeai HM, Alohaydib M, Alhoshani AR, et al. Sorafenib, rapamycin, and venetoclax attenuate doxorubicin-induced senescence and promote apoptosis in HCT116 cells. Saudi Pharm J. 2022;30(1):91–101. doi:10.1016/j.jsps.2021.12.004
91. Molica S. Venetoclax: a real game changer in treatment of chronic lymphocytic leukemia. Int J Hematol Oncol. 2020;9(4):IJH31. doi:10.2217/ijh-2020-0010
92. Griffioen MS, De leeuw DC, Janssen JJWM, Smit L. Targeting acute myeloid leukemia with venetoclax; biomarkers for sensitivity and rationale for venetoclax-based combination therapies. Cancers. 2022;14(14):3456. doi:10.3390/cancers14143456
93. Bewersdorf JP, Giri S, Wang R, et al. Venetoclax as monotherapy and in combination with hypomethylating agents or low dose cytarabine in relapsed and treatment refractory acute myeloid leukemia: a systematic review and meta-analysis. Haematologica. 2020;105(11):2659–2663. doi:10.3324/haematol.2019.242826
94. Yosef R, Pilpel N, Tokarsky-Amiel R, et al. Directed elimination of senescent cells by inhibition of BCL-W and BCL-XL. Nat Commun. 2016;7(1):11190. doi:10.1038/ncomms11190
95. Fuhrmann-Stroissnigg H, Ling YY, Zhao J, et al. Identification of HSP90 inhibitors as a novel class of senolytics. Nat Commun. 2017;8(1):422. doi:10.1038/s41467-017-00314-z
96. Rastogi S, Joshi A, Sato N, et al. An update on the status of HSP90 inhibitors in cancer clinical trials. Cell Stress Chap. 2024;29(4):519–539. doi:10.1016/j.cstres.2024.05.005
97. Belviranlı M, Okudan N, Atalık KEN, Öz M. Curcumin improves spatial memory and decreases oxidative damage in aged female rats. Biogerontology. 2013;14(2):187–196. doi:10.1007/s10522-013-9422-y
98. Allison DJ, Josse AR, Gabriel DA, Klentrou P, Ditor DS. Targeting inflammation to influence cognitive function following spinal cord injury: a randomized clinical trial. Spinal Cord. 2017;55(1):26–32. doi:10.1038/sc.2016.96
99. Ghodsi H, Rahimi HR, Aghili SM, Saberi A, Shoeibi A. Evaluation of curcumin as add-on therapy in patients with Parkinson’s disease: a pilot randomized, triple-blind, placebo-controlled trial. Clin Neurol Neurosurg. 2022;218:107300. doi:10.1016/j.clineuro.2022.107300
100. Pallauf K, Rimbach G, Rupp PM, Chin D, Wolf IMA. Resveratrol and lifespan in model organisms. Curr Med Chem. 2016;23(41):4639–4680. doi:10.2174/0929867323666161024151233
101. Song J, Liu L, Hao K, et al. Resveratrol elongates the lifespan and improves antioxidant activity in the silkworm Bombyx mori. J Pharm Anal. 2021;11(3):374–382. doi:10.1016/j.jpha.2020.06.005
102. Dolinsky VW, Chakrabarti S, Pereira TJ, et al. Resveratrol prevents hypertension and cardiac hypertrophy in hypertensive rats and mice. Biochim Biophys Acta Mol Basis Dis. 2013;1832(10):1723–1733. doi:10.1016/j.bbadis.2013.05.018
103. Godos J, Romano GL, Gozzo L, et al. Resveratrol and vascular health: evidence from clinical studies and mechanisms of actions related to its metabolites produced by gut microbiota. Front Pharmacol. 2024;15. doi:10.3389/fphar.2024.1368949
104. Seyyedebrahimi S, Khodabandehloo H, Nasli Esfahani E, Meshkani R. The effects of resveratrol on markers of oxidative stress in patients with type 2 diabetes: a randomized, double-blind, placebo-controlled clinical trial. Acta Diabetol. 2018;55(4):341–353. doi:10.1007/s00592-017-1098-3
105. Khodabandehloo H, Seyyedebrahimi S, Esfahani EN, Razi F, Meshkani R. Resveratrol supplementation decreases blood glucose without changing the circulating CD14+CD16+ monocytes and inflammatory cytokines in patients with type 2 diabetes: a randomized, double-blind, placebo-controlled study. Nutr Res. 2018;54:40–51. doi:10.1016/j.nutres.2018.03.015
106. Martin-Montalvo A, Mercken EM, Mitchell SJ, et al. Metformin improves healthspan and lifespan in mice. Nat Commun. 2013;4(1):2192. doi:10.1038/ncomms3192
107. Yang L, Lu P, Qi X, et al. Metformin inhibits inflammatory response and endoplasmic reticulum stress to improve hypothalamic aging in obese mice. iScience. 2023;26(10):108082. doi:10.1016/j.isci.2023.108082
108. Baghdadi M, Nespital T, Monzó C, Deelen J, Grönke S, Partridge L. Intermittent rapamycin feeding recapitulates some effects of continuous treatment while maintaining lifespan extension. Mol Metab. 2024;81:101902. doi:10.1016/j.molmet.2024.101902
109. Chen M, Huang L, Lv Y, Li L, Dong Q. Sulforaphane protects against oxidative stress-induced apoptosis via activating SIRT1 in mouse osteoarthritis. Mol Med Rep. 2021;24(2):612. doi:10.3892/mmr.2021.12251
110. Ibrahim Fouad G. Sulforaphane, an Nrf-2 agonist, modulates oxidative stress and inflammation in a rat model of cuprizone-induced cardiotoxicity and hepatotoxicity. Cardiovasc Toxicol. 2023;23(1):46–60. doi:10.1007/s12012-022-09776-0
111. Qin Y, Zhang H, Liu Q, Jiang B, Chen J, Zhang T. Sulforaphane attenuates oxidative stress and inflammation induced by fine particulate matter in human bronchial epithelial cells. J Funct Foods. 2021;81:104460. doi:10.1016/j.jff.2021.104460
112. Zhou J, Huang Z, Ni X, Lv C. Piperlongumine induces apoptosis and G2/M phase arrest in human osteosarcoma cells by regulating ROS/PI3K/Akt pathway. Toxicol Vitro. 2020;65:104775. doi:10.1016/j.tiv.2020.104775
113. Kung F-P, Lim Y-P, Chao W-Y, et al. Piperlongumine, a potent anticancer phytotherapeutic, induces cell cycle arrest and apoptosis in vitro and in vivo through the ROS/Akt pathway in human thyroid cancer cells. Cancers. 2021;13(17):4266. doi:10.3390/cancers13174266
114. Nagasu H, Sogawa Y, Kidokoro K, et al. Bardoxolone methyl analog attenuates proteinuria-induced tubular damage by modulating mitochondrial function. FASEB J. 2019;33(11):12253–12263. doi:10.1096/fj.201900217R
115. Thompson LE, Tuey SM, Gonzalez PG, et al. Bardoxolone methyl improves survival and reduces clinical measures of kidney injury in tumor-bearing mice treated with cisplatin. AAPS Open. 2025;11(1):6. doi:10.1186/s41120-025-00107-5
116. Nangaku M, Kanda H, Takama H, Ichikawa T, Hase H, Akizawa T. Randomized clinical trial on the effect of bardoxolone methyl on GFR in Diabetic Kidney Disease Patients (TSUBAKI Study). Kidney Int Rep. 2020;5(6):879–890. doi:10.1016/j.ekir.2020.03.030
117. Jeon OH, Kim C, Laberge R-M, et al. Local clearance of senescent cells attenuates the development of post-traumatic osteoarthritis and creates a pro-regenerative environment. Nat Med. 2017;23(6):775–781. doi:10.1038/nm.4324
118. Lane N, Hsu B, Visich J, Xie B, Khan A, Dananberg J. A phase 2, randomized, double-blind, placebo-controlled study of senolytic molecule UBX0101 in the treatment of painful knee osteoarthritis. Osteoarthritis Cartilage. 2021;29:S52–S53. doi:10.1016/j.joca.2021.02.077
119. Raffaele M, Vinciguerra M. The costs and benefits of senotherapeutics for human health. Lancet Health Longevity. 2022;3(1):e67–e77. doi:10.1016/S2666-7568(21)00300-7
120. Vasilieva MI, Shatalova RO, Matveeva KS, et al. Senolytic vaccines from the central and peripheral tolerance perspective. Vaccines. 2024;12(12):1389. doi:10.3390/vaccines12121389
121. Yao M, Wei Z, Nielsen JS, et al. Senolytic therapy preserves blood-brain barrier integrity and promotes microglia homeostasis in a tauopathy model. Neurobiol Dis. 2024;202:106711. doi:10.1016/j.nbd.2024.106711
122. Khosla S. Senescent cells, senolytics and tissue repair: the devil may be in the dosing. Nat Aging. 2023;3(2):139–141. doi:10.1038/s43587-023-00365-6
123. Mansfield L, Ramponi V, Gupta K, et al. Emerging insights in senescence: pathways from preclinical models to therapeutic innovations. Npj Aging. 2024;10(1):1–33. doi:10.1038/s41514-024-00181-1
124. Lorenzo EC, Torrance BL, Haynes L. Impact of senolytic treatment on immunity, aging, and disease. Front Aging. 2023;4:1161799. doi:10.3389/fragi.2023.1161799
125. Rolland Y, Sierra F, Ferrucci L, et al. Challenges in developing Geroscience trials. Nat Commun. 2023;14(1):5038. doi:10.1038/s41467-023-39786-7
126. Saliev T, Singh PB. From bench to bedside: translating cellular rejuvenation therapies into clinical applications. Cells. 2024;13(24):2052. doi:10.3390/cells13242052
127. Zhang Z, Yang R, Zi Z, Liu B. A new clinical age of aging research. Trends Endocrinol Metab. 2024. doi:10.1016/j.tem.2024.08.004
128. Konstantinou E, Longange E, Kaya G. Mechanisms of senescence and anti-senescence strategies in the skin. Biology. 2024;13(9):647. doi:10.3390/biology13090647
129. Riessland M, Orr ME. Translating the biology of aging into new therapeutics for alzheimer’s disease: senolytics. J Prev Alzheimer Dis. 2023;10(4):633–646. doi:10.14283/jpad.2023.104
130. Costa CM, Pedrosa SS, Kirkland JL, Reis F, Madureira AR. The senotherapeutic potential of phytochemicals for age-related intestinal disease. Aging Res Rev. 2025;104:102619. doi:10.1016/j.arr.2024.102619
131. Liang Y, Liang N, Ma Y, Tang S, Ye S, Xiao F. Role of Clusterin/NF-κB in the secretion of senescence-associated secretory phenotype in Cr(VI)-induced premature senescent L-02 hepatocytes. Ecotoxicol Environ Saf. 2021;219:112343. doi:10.1016/j.ecoenv.2021.112343
132. Lim JS, Lee DY, Kim HS, et al. Identification of a novel senomorphic agent, avenanthramide C, via the suppression of the senescence-associated secretory phenotype. Mech Aging Dev. 2020;192:111355. doi:10.1016/j.mad.2020.111355
133. Sarapultsev A, Gusev E, Komelkova M, Utepova I, Luo S, Hu D. JAK-STAT signaling in inflammation and stress-related diseases: implications for therapeutic interventions. Mol Biomed. 2023;4(1):40. doi:10.1186/s43556-023-00151-1
134. Laberge R-M, Sun Y, Orjalo AV, et al. MTOR regulates the pro-tumorigenic senescence-associated secretory phenotype by promoting IL1A translation. Nat Cell Biol. 2015;17(8):1049–1061. doi:10.1038/ncb3195
135. Herranz N, Gallage S, Mellone M, et al. mTOR regulates MAPKAPK2 translation to control the senescence-associated secretory phenotype. Nat Cell Biol. 2015;17(9):1205–1217. doi:10.1038/ncb3225
136. Balamurli G, Liew AQX, Tee WW, Pervaiz S. Interplay between epigenetics, senescence and cellular redox metabolism in cancer and its therapeutic implications. Redox Biol. 2024;78:103441. doi:10.1016/j.redox.2024.103441
137. Pereira B, Correia FP, Alves IA, et al. Epigenetic reprogramming as a key to reverse aging and increase longevity. Aging Res Rev. 2024;95:102204. doi:10.1016/j.arr.2024.102204
138. Martini H, Passos JF. Cellular senescence: all roads lead to mitochondria. FEBS J. 2022;290(5):1186. doi:10.1111/febs.16361
139. Shreeya T, Ansari MS, Kumar P, et al. Senescence: a DNA damage response and its role in aging and neurodegenerative diseases. Front Aging. 2024;4:1292053. doi:10.3389/fragi.2023.1292053
140. Gasek NS, Kuchel GA, Kirkland JL, Xu M. Strategies for targeting senescent cells in human disease. Nat Aging. 2021;1(10):870–879. doi:10.1038/s43587-021-00121-8
141. Luís C, Maduro AT, Pereira P, Mendes JJ, Soares R, Ramalho R. Nutritional senolytics and senomorphics: implications to immune cells metabolism and aging – from theory to practice. Front Nutr. 2022;9. doi:10.3389/fnut.2022.958563
142. Yan J, Chen S, Yi Z, et al. The role of p21 in cellular senescence and aging-related diseases. Mol Cells. 2024;47(11):100113. doi:10.1016/j.mocell.2024.100113
143. Thompson EL, Pitcher LE, Niedernhofer LJ, Robbins PD. Targeting cellular senescence with senotherapeutics: development of new approaches for skin care. Plast Reconstr Surg. 2022;150:12S–19S. doi:10.1097/PRS.0000000000009668
144. Zheng L, He S, Wang H, Li J, Liu Y, Liu S. Targeting cellular senescence in aging and age-related diseases: challenges, considerations, and the emerging role of senolytic and senomorphic therapies. Aging Dis. 2024;15(6):2554–2594. doi:10.14336/AD.2024.0206
145. Abdelgawad IY, Agostinucci K, Sadaf B, Grant MKO, Zordoky BN. Metformin mitigates SASP secretion and LPS-triggered hyper-inflammation in doxorubicin-induced senescent endothelial cells. Front Aging. 2023;4:1170434. doi:10.3389/fragi.2023.1170434
146. Padki MM, Stambler I. Targeting Aging with Metformin (TAME). In: Gu D, Dupre ME, editors. Encyclopedia of Gerontology and Population Aging. Cham: Springer International Publishing; 2021:4908–4910.
147. Sheppard AJ, Delgado K, Barfield AM, et al. Rapamycin inhibits senescence and improves immunomodulatory function of mesenchymal stem cells through IL-8 and TGF-β signaling. Stem Cell Rev Rep. 2024;20(3):816–826. doi:10.1007/s12015-024-10682-x
148. Konopka AR, Lamming DW, Grasso BA. Blazing a trail for the clinical use of rapamycin as a geroprotecTOR. GeroScience. 2023;45(5):2769–2783. doi:10.1007/s11357-023-00935-x
149. Guan G, Chen Y, Dong Y. Unraveling the AMPK-SIRT1-FOXO pathway: the in-depth analysis and breakthrough prospects of oxidative stress-induced diseases. Antioxidants. 2025;14(1):70. doi:10.3390/antiox14010070
150. Bonnefont-Rousselot D. Resveratrol and cardiovascular diseases. Nutrients. 2016;8(5):250. doi:10.3390/nu8050250
151. Erdogan CS, Vang O. Challenges in analyzing the biological effects of resveratrol. Nutrients. 2016;8(6):353. doi:10.3390/nu8060353
152. Benameur T, Soleti R, Panaro MA, et al. Curcumin as prospective anti-aging natural compound: focus on brain. Molecules. 2021;26(16):4794. doi:10.3390/molecules26164794
153. Monroy A, Lithgow GJ, Alavez S. Curcumin and neurodegenerative diseases. Biofactors. 2013;39(1):122–132. doi:10.1002/biof.1063
154. Moldoveanu C-A, Tomoaia-Cotisel M, Sevastre-Berghian A, et al. A review on current aspects of curcumin-based effects in relation to neurodegenerative, neuroinflammatory and cerebrovascular diseases. Molecules. 2025;30(1):43. doi:10.3390/molecules30010043
155. Capasso L, De Masi L, Sirignano C, et al. Epigallocatechin Gallate (EGCG): pharmacological properties, biological activities and therapeutic potential. Molecules. 2025;30(3):654. doi:10.3390/molecules30030654
156. Wang J, Li P, Qin T, Sun D, Zhao X, Zhang B. Protective effect of epigallocatechin-3-gallate against neuroinflammation and anxiety-like behavior in a rat model of myocardial infarction. Brain Behav. 2020;10(6):e01633. doi:10.1002/brb3.1633
157. Zueva IV, Vasilieva EA, Gaynanova GA, et al. Can activation of acetylcholinesterase by β-amyloid peptide decrease the effectiveness of cholinesterase inhibitors? Int J Mol Sci. 2023;24(22):16395. doi:10.3390/ijms242216395
158. Meng J, Chen Y, Wang J, et al. EGCG protects vascular endothelial cells from oxidative stress-induced damage by targeting the autophagy-dependent PI3K-AKT-mTOR pathway. Ann Transl Med. 2020;8(5):200. doi:10.21037/atm.2020.01.92
159. Das D, Banerjee A, Mukherjee S, Maji BK. Quercetin inhibits NF-kB and JAK/STAT signaling via modulating TLR in thymocytes and splenocytes during MSG-induced immunotoxicity: an in vitro approach. Mol Biol Rep. 2024;51(1):277. doi:10.1007/s11033-024-09245-7
160. Ruggiero AD, Vemuri R, Blawas M, et al. Long-term dasatinib plus quercetin effects on aging outcomes and inflammation in nonhuman primates: implications for senolytic clinical trial design. GeroScience. 2023;45(5):2785–2803. doi:10.1007/s11357-023-00830-5
161. Hickson LJ, Langhi Prata LGP, Bobart SA, et al. Senolytics decrease senescent cells in humans: preliminary report from a clinical trial of dasatinib plus quercetin in individuals with diabetic kidney disease. EBioMedicine. 2019;47:446–456. doi:10.1016/j.ebiom.2019.08.069
162. Sun Y, Qin H, Zhang H, et al. Fisetin inhibits inflammation and induces autophagy by mediating PI3K/AKT/mTOR signaling in LPS-induced RAW264.7 cells. Food Nutr Res. 2021;65:
163. Mahoney SA, Venkatasubramanian R, Darrah MA, et al. Intermittent supplementation with fisetin improves arterial function in old mice by decreasing cellular senescence. Aging Cell. 2024;23(3):e14060. doi:10.1111/acel.14060
164. Kim JH, Kim M-Y, Kim J-H, Cho JY. Fisetin suppresses macrophage-mediated inflammatory responses by blockade of src and syk. Biomol Ther. 2015;23(5):414–420. doi:10.4062/biomolther.2015.036
165. Perrott KM, Wiley CD, Desprez P-Y, Campisi J. Apigenin suppresses the senescence-associated secretory phenotype and paracrine effects on breast cancer cells. GeroScience. 2017;39(2):161–173. doi:10.1007/s11357-017-9970-1
166. Yadav RK, Mehan S, Sahu R, et al. Protective effects of apigenin on methylmercury-induced behavioral/neurochemical abnormalities and neurotoxicity in rats. Hum Exp Toxicol. 2022;41:09603271221084276. doi:10.1177/09603271221084276
167. Jiang X, Huang H. The therapeutic potential of apigenin against atherosclerosis. Heliyon. 2025;11(1):e41272. doi:10.1016/j.heliyon.2024.e41272
168. Łanoszka K, Vlčková N. Natural sirtuin1 activators and atherosclerosis: an overview. Curr Atheroscler Rep. 2023;25(12):979–994. doi:10.1007/s11883-023-01165-4
169. Xu J, Liu J, Li Q, et al. Pterostilbene participates in TLR4- mediated inflammatory response and autophagy-dependent Aβ1–42 endocytosis in Alzheimer’s disease. Phytomedicine. 2023;119:155011. doi:10.1016/j.phymed.2023.155011
170. Dutta BJ, Rakshe PS, Maurya N, Chib S, Singh S. Unlocking the therapeutic potential of natural stilbene: exploring pterostilbene as a powerful ally against aging and cognitive decline. Aging Res Rev. 2023;92:102125. doi:10.1016/j.arr.2023.102125
171. Xu Y, Xiao W. NAD+: an old but promising therapeutic agent for skeletal muscle aging. Aging Res Rev. 2023;92:102106. doi:10.1016/j.arr.2023.102106
172. Zhou Q, Zhu L, Qiu W, et al. Nicotinamide riboside enhances mitochondrial proteostasis and adult neurogenesis through activation of mitochondrial unfolded protein response signaling in the brain of ALS SOD1 G93A mice. Int J Biol Sci. 2020;16(2):284–297. doi:10.7150/ijbs.38487
173. Lapatto HAK, Kuusela M, Heikkinen A, et al. Nicotinamide riboside improves muscle mitochondrial biogenesis, satellite cell differentiation, and gut microbiota in a twin study. Sci Adv. 2023;9(2):eadd5163. doi:10.1126/sciadv.add5163
174. Szabo L, Lejri I, Grimm A, Eckert A. Spermidine enhances mitochondrial bioenergetics in young and aged human-induced pluripotent stem cell-derived neurons. Antioxidants. 2024;13(12):1482. doi:10.3390/antiox13121482
175. Eisenberg T, Abdellatif M, Schroeder S, et al. Cardioprotection and lifespan extension by the natural polyamine spermidine. Nat Med. 2016;22(12):1428–1438. doi:10.1038/nm.4222
176. Schwarz C, Benson GS, Horn N, et al. Effects of spermidine supplementation on cognition and biomarkers in older adults with subjective cognitive decline. JAMA Netw Open. 2022;5(5):e2213875. doi:10.1001/jamanetworkopen.2022.13875
177. Wortha SM, Frenzel S, Bahls M, et al. Association of spermidine plasma levels with brain aging in a population-based study. Alzheimer Dementia. 2023;19(5):1832–1840. doi:10.1002/alz.12815
178. Trushina E, Nguyen TKO, Trushin S. Modulation of mitochondrial function as a therapeutic strategy for neurodegenerative diseases. J Prev Alzheimer Disease. 2023;10(4):675–685. doi:10.14283/jpad.2023.108
179. Rhoads TW, Anderson RM. Alpha-ketoglutarate, the metabolite that regulates aging in mice. Cell Metab. 2020;32(3):323–325. doi:10.1016/j.cmet.2020.08.009
180. Gyanwali B, Lim ZX, Soh J, et al. Alpha-ketoglutarate dietary supplementation to improve health in humans. Trends Endocrinol Metab. 2022;33(2):136–146. doi:10.1016/j.tem.2021.11.003
181. Mushtaq Z, Imran M, Saeed F, et al. Berberine: a comprehensive approach to combat human maladies. Int J Food Prop. 2023;26(1):787–807. doi:10.1080/10942912.2023.2184300
182. Dang Y, An Y, He J, et al. Berberine ameliorates cellular senescence and extends the lifespan of mice via regulating p16 and cyclin protein expression. Aging Cell. 2020;19(1):e13060. doi:10.1111/acel.13060
183. Nazari A, Ghotbabadi ZR, Kazemi KS, et al. The effect of berberine supplementation on glycemic control and inflammatory biomarkers in metabolic disorders: an umbrella meta-analysis of randomized controlled trials. Clin Ther. 2024;46(2):e64–e72. doi:10.1016/j.clinthera.2023.10.019
184. Cascajosa-Lira A, Prieto AI, Pichardo S, Jos A, Cameán AM. Protective effects of sulforaphane against toxic substances and contaminants: a systematic review. Phytomedicine. 2024;130:155731. doi:10.1016/j.phymed.2024.155731
185. Canto A, Martínez-González J, Miranda M, Olivar T, Almansa I, Hernández-Rabaza V. Sulforaphane modulates the inflammation and delays neurodegeneration on a retinitis pigmentosa mice model. Front Pharmacol. 2022;13. doi:10.3389/fphar.2022.811257
186. Hei G, Smith RC, Li R, et al. Sulforaphane effects on cognition and symptoms in first and early episode schizophrenia: a randomized double-blind trial. Schizophr Bull Open. 2022;3(1):sgac024. doi:10.1093/schizbullopen/sgac024
187. Che L, Zhu C, Huang L, et al. Ginsenoside Rg2 promotes the proliferation and stemness maintenance of porcine mesenchymal stem cells through autophagy induction. Foods. 2023;12(5):1075. doi:10.3390/foods12051075
188. Zhou B, Shi G, Xie D, et al. Ginsenoside Rb1 prevents age-related endothelial senescence by modulating SIRT1/caveolin-1/enos signaling pathway. Heliyon. 2024;10(3):e24586. doi:10.1016/j.heliyon.2024.e24586
189. Zong Y, Yu W, Hong H, et al. Ginsenoside Rg1 improves inflammation and autophagy of the pancreas and spleen in streptozotocin-induced type 1 diabetic mice. Int J Endocrinol. 2023;2023:3595992. doi:10.1155/2023/3595992
190. Bell L, Whyte A, Duysburgh C, et al. A randomized, placebo-controlled trial investigating the acute and chronic benefits of American Ginseng (Cereboost®) on mood and cognition in healthy young adults, including in vitro investigation of gut microbiota changes as a possible mechanism of action. Eur J Nutr. 2022;61(1):413–428. doi:10.1007/s00394-021-02654-5
191. Boccardi V, Orr ME, Polidori MC, Ruggiero C, Mecocci P. Focus on senescence: clinical significance and practical applications. J Int Med. 2024;295(5):599–619. doi:10.1111/joim.13775
192. Abrhaley A, Giday M, Hailu A. Challenges and opportunities of translating animal research into human trials in Ethiopia. BMC Med Res Methodol. 2024;24(1):211. doi:10.1186/s12874-024-02338-8
193. Guo J, Huang X, Dou L, et al. Aging and aging-related diseases: from molecular mechanisms to interventions and treatments. Sig Transd Target Ther. 2022;7:1–40. doi:10.1038/s41392-022-01251-0
194. Dong Z, Luo Y, Yuan Z, Tian Y, Jin T, Xu F. Cellular senescence and SASP in tumor progression and therapeutic opportunities. Mol Cancer. 2024;23(1):1–19. doi:10.1186/s12943-024-02096-7
195. Bertoncini-Silva C, Vlad A, Ricciarelli R, Giacomo Fassini P, Suen VMM, Zingg J-M. Enhancing the bioavailability and bioactivity of curcumin for disease prevention and treatment. Antioxidants. 2024;13(3):331. doi:10.3390/antiox13030331
196. Dejeu IL, Vicaș LG, Marian E, et al. Innovative approaches to enhancing the biomedical properties of liposomes. Pharmaceutics. 2024;16(12):1525. doi:10.3390/pharmaceutics16121525
197. Niklander SE, Lambert DW, Hunter KD. Senescent cells in cancer: wanted or unwanted citizens. Cells. 2021;10(12):3315. doi:10.3390/cells10123315
198. Fang J, Chen W, Hou P, et al. NAD+ metabolism-based immunoregulation and therapeutic potential. Cell Biosci. 2023;13(1):81. doi:10.1186/s13578-023-01031-5
199. Bousset L, Gil J. Targeting senescence as an anticancer therapy. Mol Oncol. 2022;16(21):3855–3880. doi:10.1002/1878-0261.13312
200. Posey KL. Curcumin and resveratrol: nutraceuticals with so much potential for pseudoachondroplasia and other ER-stress conditions. Biomolecules. 2024;14(2):154. doi:10.3390/biom14020154
201. Li Z, Zhang Z, Ren Y, et al. Aging and age‐related diseases: from mechanisms to therapeutic strategies. Biogerontology. 2021;22(2):165–187. doi:10.1007/s10522-021-09910-5
202. Zhou Y, Tao L, Qiu J, et al. Tumor biomarkers for diagnosis, prognosis and targeted therapy. Sig Transduct Target Ther. 2024;9:1–86. doi:10.1038/s41392-024-01823-2
203. Alum EU, Ugwu OP-C. Artificial intelligence in personalized medicine: transforming diagnosis and treatment. Discov Appl Sci. 2025;7(3):193. doi:10.1007/s42452-025-06625-x
204. Shi Y, Zhang Y, Zhang Y, et al. Advances in nanotherapy for targeting senescent cells. Int J Nanomed. 2024;19:8797–8813. doi:10.2147/IJN.S469110
205. Chen X, Walton K, Brodaty H, Chalton K. Polyphenols and diets as current and potential nutrition senotherapeutics in alzheimer’s disease: findings from clinical trials. J Alzheimer Disease. 2024;101(s1):S479–S501. doi:10.3233/JAD-231222
206. Hughes BK, Wallis R, Bishop CL. Yearning for machine learning: applications for the classification and characterisation of senescence. Cell Tissue Res. 2023;394(1):1–16. doi:10.1007/s00441-023-03768-4
207. Zhang L, Pitcher LE, Yousefzadeh MJ, Niedernhofer LJ, Robbins PD, Zhu Y. Cellular senescence: a key therapeutic target in aging and diseases. J Clin Invest. 2022;132(15):e158450. doi:10.1172/JCI158450
208. Liu H, Xu Q, Wufuer H, et al. Rutin is a potent senomorphic agent to target senescent cells and can improve chemotherapeutic efficacy. Aging Cell. 2024;23(1):e13921. doi:10.1111/acel.13921
209. Czajkowski K, Herbet M, Murias M, Piątkowska-Chmiel I. Senolytics: charting a new course or enhancing existing anti-tumor therapies? Cell Oncol. 2024. doi:10.1007/s13402-024-01018-5
210. Liao Z, Yeo HL, Wong SW, Zhao Y. Cellular senescence: mechanisms and therapeutic potential. Biomedicines. 2021;9(12):1769. doi:10.3390/biomedicines9121769
211. Han Y, Kim SY. Endothelial senescence in vascular diseases: current understanding and future opportunities in senotherapeutics. Exp Mol Med. 2023;55(1):1–12. doi:10.1038/s12276-022-00906-w
212. Abdelgawad IY, Agostinucci K, Zordoky BN. Cardiovascular ramifications of therapy-induced endothelial cell senescence in cancer survivors. Biochim Biophys Acta Mol Basis Dis. 2022;1868(4):166352. doi:10.1016/j.bbadis.2022.166352
213. Gaikwad S, Senapati S, Haque MA, Kayed R. Senescence, brain inflammation, and oligomeric tau drive cognitive decline in Alzheimer’s disease: evidence from clinical and preclinical studies. Alzheimers Dement. 2023;20(1):709–727. doi:10.1002/alz.13490
214. Krzystyniak A, Wesierska M, Petrazzo G, et al. Combination of dasatinib and quercetin improves cognitive abilities in aged male Wistar rats, alleviates inflammation and changes hippocampal synaptic plasticity and histone H3 methylation profile. Aging. 2022;14(2):572–595. doi:10.18632/aging.203835
215. Zhao S, Qiao Z, Pfeifer R, et al. Modulation of fracture healing by senescence-associated secretory phenotype (SASP): a narrative review of the current literature. Eur J Med Res. 2024;29(1):38. doi:10.1186/s40001-023-01604-7
216. Suda M, Paul KH, Tripathi U, Minamino T, Tchkonia T, Kirkland JL. Targeting cell senescence and senolytics: novel interventions for age-related endocrine dysfunction. Endocrine Rev. 2024;45(5):655–675. doi:10.1210/endrev/bnae010
217. Jiang B, Zhang W, Zhang X, Sun Y. Targeting senescent cells to reshape the tumor microenvironment and improve anticancer efficacy. Semin Cancer Biol. 2024;101:58–73. doi:10.1016/j.semcancer.2024.05.002
218. Lin M-T, Lin C-L, Lin T-Y, et al. Synergistic effect of fisetin combined with sorafenib in human cervical cancer HeLa cells through activation of death receptor-5 mediated caspase-8/caspase-3 and the mitochondria-dependent apoptotic pathway. Tumour Biol. 2016;37(5):6987–6996. doi:10.1007/s13277-015-4526-4
219. Zhu Y, Tchkonia T, Pirtskhalava T, et al. The Achilles’ heel of senescent cells: from transcriptome to senolytic drugs. Aging Cell. 2015;14(4):644–658. doi:10.1111/acel.12344
220. Hernandez-Gonzalez F, Faner R, Rojas M, Agustí A, Serrano M, Sellarés J. Cellular senescence in lung fibrosis. Int J Mol Sci. 2021;22(13):7012. doi:10.3390/ijms22137012
221. Yy C, Ji S, Ji H, Ds K. Co-administration of everolimus and N-acetylcysteine attenuates hepatic stellate cell activation and hepatic fibrosis. Abstract - Europe PMC.
222. Chilvery S, Bansod S, Saifi MA, Godugu C. Piperlongumine attenuates bile duct ligation-induced liver fibrosis in mice via inhibition of TGF-β1/Smad and EMT pathways. Int Immunopharmacol. 2020;88:106909. doi:10.1016/j.intimp.2020.106909
223. Faust HJ, Zhang H, Han J, et al. IL-17 and immunologically induced senescence regulate response to injury in osteoarthritis. J Clin Invest. 2020;130(10):5493–5507. doi:10.1172/JCI134091
224. Shang R, Miao J. Mechanisms and effects of metformin on skeletal muscle disorders. Front. Neurol. 2023;14. doi:10.3389/fneur.2023.1275266
225. Baar MP, Perdiguero E, Muñoz-Cánoves P, de Keizer PL. Musculoskeletal senescence: a moving target ready to be eliminated. Curr Opin Pharmacol. 2018;40:147–155. doi:10.1016/j.coph.2018.05.007
226. Ibrahim NF, Yanagisawa D, Durani LW, et al. Tocotrienol-rich fraction modulates amyloid pathology and improves cognitive function in AβPP/PS1 mice. J Alzheimers Dis. 2017;55(2):597–612. doi:10.3233/JAD-160685
227. Rachmian N, Medina S, Cherqui U, et al. TREM2-dependent senescent microglia conserved in aging and Alzheimer’s disease. 2023. Available from: https://www.biorxiv.org/content/10.1101/2023.03.20.533401v2.
228. Alegre GFS, Pastore GM, Ribeiro H. NAD+ Precursors Nicotinamide Mononucleotide (NMN) and Nicotinamide Riboside (NR): potential dietary contribution to health. Curr Nutr Rep. 2023;12(1):1–20. doi:10.1007/s13668-023-00475-y
229. Chen L, Yang S, Liao W, Xiong Y. Modification of antitumor immunity and tumor microenvironment by resveratrol in mouse renal tumor model. Cell Biochem Biophys. 2015;72(2):617–625. doi:10.1007/s12013-015-0513-z
230. Tian S, Wang Y, Li X, Liu J, Wang J, Lu Y. Sulforaphane regulates glucose and lipid metabolisms in obese mice by restraining JNK and activating insulin and FGF21 signal pathways. J Agric Food Chem. 2021;69(44):13066–13079. doi:10.1021/acs.jafc.1c04933
231. Wang L, Doherty GA, Judd AS, et al. Discovery of A-1331852, a first-in-class, potent, and orally-bioavailable BCL-XL inhibitor. ACS Med Chem Lett. 2020;11(10):1829–1836. doi:10.1021/acsmedchemlett.9b00568
232. Hou S-J, Zhang S-X, Li Y, Xu S-Y. Rapamycin responds to Alzheimer’s disease: a potential translational therapy. Clin Interv Aging. 2023;18:1629–1639. doi:10.2147/CIA.S429440
233. Mkhize SA, Nthlane RA, Xhakaza SP, et al. Decreased blood pressure with acute administration of quercetin in L-NAME-induced hypertensive rats. Basic Clin Pharmacol Toxicol. 2025;136(1):e14113. doi:10.1111/bcpt.14113
234. Yu F, Qi S, Ji Y, Wang X, Fang S, Cao R. Effects of omega-3 fatty acid on major cardiovascular outcomes: a systematic review and meta-analysis. Medicine. 2022;101(30):e29556. doi:10.1097/MD.0000000000029556
235. Ferreira SG, Peliciari-Garcia RA, Takahashi-Hyodo SA, et al. Effects of melatonin on DNA damage induced by cyclophosphamide in rats. Braz J Med Biol Res. 2013;46(3):278–286. doi:10.1590/1414-431X20122230
236. Greenberg EF, Voorbach MJ, Smith A, et al. Navitoclax safety, tolerability, and effect on biomarkers of senescence and neurodegeneration in aged nonhuman primates. Heliyon. 2024;10(16):e36483. doi:10.1016/j.heliyon.2024.e36483
237. Gu W, Yang L, Wang X, et al. Pterostilbene, a resveratrol derivative, improves ectopic lipid deposition in the kidneys of mice induced by a high-fat diet. Kidney Blood Press Res. 2022;47(8):514–522. doi:10.1159/000525385
238. Zhou L, Wu K, Gao Y, et al. Piperlonguminine attenuates renal fibrosis by inhibiting TRPC6. J Ethnopharmacol. 2023;313:116561. doi:10.1016/j.jep.2023.116561
239. Nambiar A, Kellogg D, Justice J, et al. Senolytics dasatinib and quercetin in idiopathic pulmonary fibrosis: results of a Phase I, single-blind, single-center, randomized, placebo-controlled pilot trial on feasibility and tolerability. EBioMedicine. 2023;90:104481. doi:10.1016/j.ebiom.2023.104481
240. Igarashi M, Nakagawa-Nagahama Y, Miura M, et al. Chronic nicotinamide mononucleotide supplementation elevates blood nicotinamide adenine dinucleotide levels and alters muscle function in healthy older men. NPJ Aging. 2022;8(1):5. doi:10.1038/s41514-022-00084-z
241. Song Q, Zhou X, Xu K, Liu S, Zhu X, Yang J. The safety and antiaging effects of nicotinamide mononucleotide in human clinical trials: an update. Adv Nutr. 2023;14(6):1416–1435. doi:10.1016/j.advnut.2023.08.008
242. Justice JN, Nambiar AM, Tchkonia T, et al. Senolytics in idiopathic pulmonary fibrosis: results from a first-in-human, open-label, pilot study. EBioMedicine. 2019;40:554–563. doi:10.1016/j.ebiom.2018.12.052
243. Pang P, Zhang S. Dasatinib enhances curcumin-induced cytotoxicity, apoptosis and protective autophagy in human schwannoma cells HEI-193: the role of Akt/mTOR/p70S6K signalling pathway. Acta Pharmaceutica. 2022;72(3):403–414. doi:10.2478/acph-2022-0025
244. Palmer AK, Xu M, Zhu Y, et al. Targeting senescent cells alleviates obesity‐induced metabolic dysfunction. Aging Cell. 2019;18(3):e12950. doi:10.1111/acel.12950
245. Rossman MJ, Kaplon RE, Hill SD, et al. Endothelial cell senescence with aging in healthy humans: prevention by habitual exercise and relation to vascular endothelial function. Am J Physiol Heart Circ Physiol. 2017;313(5):H890–H895. doi:10.1152/ajpheart.00416.2017
246. Colman R, Tchkonia T, Pirtskhalava T, et al. Effect of combined dasatinib and fisetin treatment on senescent cell clearance in monkeys. Innov Aging. 2020;4(Supplement_1):131–132. doi:10.1093/geroni/igaa057.432
247. Hu Y, Lu S, Xue C, et al. Exploring the protective effect of metformin against sarcopenia: insights from cohort studies and genetics. J Transl Med. 2025;23(1):356. doi:10.1186/s12967-025-06357-x
248. Guan R, Cai Z, Wang J, et al. Hydrogen sulfide attenuates mitochondrial dysfunction-induced cellular senescence and apoptosis in alveolar epithelial cells by upregulating sirtuin 1. Aging. 2019;11(24):11844–11864. doi:10.18632/aging.102454
249. Estepa-Fernández A, García-Fernández A, Lérida-Viso A, et al. Combination of palbociclib with navitoclax based-therapies enhances in vivo antitumoral activity in triple-negative breast cancer. Pharmacol Res. 2023;187:106628. doi:10.1016/j.phrs.2022.106628
250. Zhao L, Zhao H, Zhao Y, et al. Role of ginseng, quercetin, and tea in enhancing chemotherapeutic efficacy of colorectal cancer. Front Med. 2022;9:939424. doi:10.3389/fmed.2022.939424
251. Chen X, Yu T, Li S, Fang H. Inhibition of bromodomain regulates cellular senescence in pancreatic adenocarcinoma. Int J Clin Exp Pathol. 2024;17(10):360–370. doi:10.62347/BKNQ9812
252. Sun Y, Han J, Wang Z, Li X, Sun Y, Hu Z. Safety and efficacy of bromodomain and extra-terminal inhibitors for the treatment of hematological malignancies and solid tumors: a systematic study of clinical trials. Front. Pharmacol. 2021;11. doi:10.3389/fphar.2020.621093
253. Alsoudi AF, Wai KM, Koo E, Mruthyunjaya P, Rahimy E. Curcuma-based nutritional supplements and risk of age-related macular degeneration. JAMA Ophthalmol. 2024;142(12):1114–1121. doi:10.1001/jamaophthalmol.2024.4400
254. Lin A, Giuliano CJ, Palladino A, et al. Off-target toxicity is a common mechanism of action of cancer drugs undergoing clinical trials. Sci Transl Med. 2019;11(509):eaaw8412. doi:10.1126/scitranslmed.aaw8412
255. Zhang F, Cheng T, Zhang S-X. Mechanistic target of rapamycin (mTOR): a potential new therapeutic target for rheumatoid arthritis. Arthritis Research & Therapy. 2023;25(1):187. doi:10.1186/s13075-023-03181-w
256. Lucas V, Cavadas C, Aveleira CA. Cellular senescence: from mechanisms to current biomarkers and senotherapies. Pharmacol Rev. 2023;75(4):675–713. doi:10.1124/pharmrev.122.000622
257. Sanborn MA, Wang X, Gao S, Dai Y, Rehman J. Unveiling the cell-type-specific landscape of cellular senescence through single-cell transcriptomics using SenePy. Nat Commun. 2025;16(1):1884. doi:10.1038/s41467-025-57047-7
258. Plana D, Palmer AC, Sorger PK. Independent Drug Action in Combination Therapy: implications for Precision Oncology. Cancer Discov. 2022;12(3):606–624. doi:10.1158/2159-8290.CD-21-0212
259. Basisty N, Kale A, Patel S, Campisi J, Schilling B. The power of proteomics to monitor senescence-associated secretory phenotypes and beyond: toward clinical applications. Expert Rev Proteomics. 2020;17(4):297–308. doi:10.1080/14789450.2020.1766976
260. Elumalai K, Srinivasan S, Shanmugam A. Review of the efficacy of nanoparticle-based drug delivery systems for cancer treatment. Biomed Technol. 2024;5:109–122. doi:10.1016/j.bmt.2023.09.001
261. Nunkoo VS, Cristian A, Jurcau A, Diaconu RG, Jurcau MC. The quest for eternal youth: hallmarks of aging and rejuvenating therapeutic strategies. Biomedicines. 2024;12(11):2540. doi:10.3390/biomedicines12112540
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
How Does Chronobiology Contribute to the Development of Diseases in Later Life
Stowe TA, McClung CA
Clinical Interventions in Aging 2023, 18:655-666
Published Date: 20 April 2023
Xian Huang Fang Inhibits the PKCδ/MAPK Signaling Pathway to Attenuate Renal Tubular Epithelial Cell Senescence and Thereby Delay Renal Interstitial Fibrosis
Wang Y, Shao M, Li T, Du X, Chen K, Xu L
Drug Design, Development and Therapy 2025, 19:6991-7004
Published Date: 13 August 2025
Sex Differences in the Association Between Inflammatory Biomarkers (Hs-CRP, IL-6, IL-10, TNF-α) and Frailty in Hospitalized Adults Aged ≥65 in Taizhou, China: A Cross-Sectional Study
Yang H, Xiao J, Zhang W, Liu X, Mao H, Xu D
Clinical Interventions in Aging 2025, 20:2267-2281
Published Date: 1 December 2025
Immunosenescence in Idiopathic Pulmonary Fibrosis
Xiao S, Li Z, Zhang H, Cai W, Yu S, Song X
Journal of Inflammation Research 2026, 19:586167
Published Date: 5 June 2026