Back to Journals » Journal of Inflammation Research » Volume 18
Single-Cell Transcriptomics Reveals CCL3+ Classical Monocyte Subset Linked to Autoimmune Pathogenesis
Authors Xu H, Yuan K, Chen G ![]() , Luo J
, Luo J ![]() , Yan A, Huang H, Yu X
, Yan A, Huang H, Yu X ![]() , Tao Q, Huang G, Xu A
, Tao Q, Huang G, Xu A
Received 2 July 2025
Accepted for publication 9 November 2025
Published 20 November 2025 Volume 2025:18 Pages 16273—16291
DOI https://doi.org/10.2147/JIR.S547283
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Dr Shouya Feng
Heng Xu,1,* Kai Yuan,1,* Guangyao Chen,2,* Jing Luo,2 Aimin Yan,1 Huaijuan Huang,1 Xinbo Yu,2 Qingwen Tao,2 Guangrui Huang,1 Anlong Xu1
1School of Life Sciences, Beijing University of Chinese Medicine, Beijing, People’s Republic of China; 2Traditional Chinese Medicine Department of Rheumatism, China-Japan Friendship Hospital, Beijing, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Anlong Xu, School of Life Sciences, Beijing University of Chinese Medicine, 11 Beisanhuandong Road, Chaoyang District, Beijing, People’s Republic of China, Email [email protected] Guangrui Huang, School of Life Sciences, Beijing University of Chinese Medicine, 11 Beisanhuandong Road, Chaoyang District, Beijing, People’s Republic of China, Email [email protected]
Objective: The diverse differentiation states of mononuclear macrophages are closely associated with the pathogenesis of autoimmune diseases. This study integrates single-cell RNA sequencing data from six autoimmune diseases to characterize shared and disease-specific alterations in mononuclear macrophages, with the aim of enhancing our understanding of the immune landscape in autoimmune diseases and refining clinical treatment strategies.
Methods: We collected single-cell RNA-sequencing data of autoimmune diseases including primary Sjogren’s syndrome (pSS), Behçet’s disease (BD), juvenile dermatomyositis (JDM), rheumatoid arthritis (RA), relapsing-remitting multiple sclerosis (RRMS), and systemic lupus erythematosus (SLE). We performed scRNA-seq analysis on 350,043 peripheral blood immune cells from autoimmune diseases patients and healthy controls, followed by validations with flow cytometry, immunohistochemical staining, and immunofluorescence.
Results: Fifteen mononuclear phagocyte subpopulations were clustered from peripheral blood mononuclear cells (PBMCs), we identified a new subpopulation named CCL3+ classical monocytes (cMo) that is co-amplified in multiple autoimmune diseases (BD, JDM, pSS, RRMS, SLE). The CCL3+ cMo cells are characterized by high M1-like score, exhibiting strong inflammatory characteristics and high chemotaxis toward other monocytes. In addition, CCL3+ cMo cells upregulated antigen presentation-related signaling pathways, and the cytotoxic CD8+ T or memory CD8+ T cells were strongly perturbed by their signaling crosstalk.
Conclusion: This study delineates a comprehensive landscape of mononuclear phagocyte heterogeneity in autoimmune diseases and reveals CCL3+ cMo as a commonly amplified immune subset associated with multiple autoimmune diseases. These findings highlight its potential role in disease mechanisms and nominate CCL3+ cMo as a candidate therapeutic target.
Keywords: autoimmune diseases, single-cell RNA sequencing, CCL3+ classical monocytes, mononuclear phagocytes, primary Sjogren’s syndrome
Introduction
Autoimmune diseases are caused by abnormal response of the immune system, which mistakenly attacks healthy, normally functioning parts of the body as if they were foreign organisms.1 The classic autoimmune diseases include systemic lupus erythematosus (SLE), rheumatoid arthritis (RA), primary Sjogren’s syndrome (pSS), Behcet’s disease (BD), etc. They are characterized by spontaneous overactivity of the immune system, resulting in the production of additional antibodies into the circulation.2–5 Persistent disease activity and comorbidities contribute to accrual mortality. However, the multiple initial symptoms of autoimmune disease are often ignored until these diseases cause organ complications, leading to irreversible organ damage. Therefore, finding specific targets for autoimmune diseases is of positive significance for the diagnosis and treatment of diseases.
Over the last decades, advances in the understanding of pathogenesis have led to improvements in the diagnosis and management of autoimmune diseases. Peripheral blood serves as the primary conduit for the immune system. Peripheral blood mononuclear cells (PBMCs) are responsible for initiating autoimmune pathogenesis against target organs, in which monocytes and the macrophages they differentiate into, can promote inflammatory responses.6 Monocytes function refined by their heterogeneity into classical, intermediate, and non-classical subsets.7 Their pro-inflammatory impact is critically mediated through dynamic interactions with other immune cells. Recruited monocytes differentiate into macrophages or dendritic cells (DCs), which present antigen and produce cytokines like IL-1β and TNF-α to activate resident DCs and T cells. Conversely, intermediate monocytes directly influence T helper cell polarization through antigen presentation and secretion of IL-12 and IL-23. In return, T-cell-derived signals such as IFN-γ amplify monocyte activation, establishing a feed-forward loop that sustains inflammation in autoimmune and metabolic diseases.8–10 Macrophages are a major source of pro-inflammatory cytokines and chemokines such as TNF and IL-1β, which activate a variety of immune and non-immune cells and drive chronic pro-inflammatory, tissue destruction, and pain responses.11,12 FABP4 is expressed in M1-polarized macrophages and promotes synovitis and angiogenesis, thereby exacerbating rheumatoid arthritis.13 Circulating monocytes exhibit NLRP3 inflammasome activation and demonstrate heightened responsiveness to NLRP3 stimuli, while macrophages infiltrating the salivary glands show signs of NLRP3 activation and pyroptosis. Systemic activation of the NLRP3 inflammasome is observed in patients with severe sjögren’s syndrome.14 Monocytes can infiltrate the ocular surface and differentiate into inflammation-associated M1 macrophages, which can be induced in mice through desiccating stress to trigger dry eye disease (DED). Local depletion of macrophages in the corneal stroma, limbal region, and lacrimal gland of DED mice significantly improves tear secretion.15 Beyond their detrimental pro-inflammatory effects, monocytes and macrophages also play immunoregulatory and tissue-repairing roles that can help prevent the progression of autoimmune diseases. In the synovial lining, CX3CR1+ tissue-resident macrophages (CX3CR1+ lining macrophages) form dynamic membrane-like structures through tight junctions, creating an immunological barrier that physically sequesters the joint and restricts inflammatory reactions.16 Thus, the commonalities and specificities of monocyte-macrophage functions across diverse autoimmune diseases merit further investigation. Single-cell RNA sequencing (scRNA-seq) enables specific analysis of cell populations and gene expression at the single-cell level.17–19 Therefore, scRNA-seq can provide insights into the composition of peripheral blood cells and find disease-specific cell subpopulations.
We collected and organized a large number of single-cell RNA sequencing data of peripheral blood immune cells isolated from patients with autoimmune diseases. This work provided a comprehensive transcriptome data map of normal and autoimmune disease peripheral blood mononuclear cell types. More importantly, we revealed alterations in gene expression at the level of cell type specificity and revealed the function of peripheral blood mononuclear cell subsets. In conclusion, we integrated multiple autoimmune disease peripheral blood mononuclear phagocyte gene profile characteristics, to identify potential biomarkers and relatively specific cell subpopulations. This study provided insights for improving the current diagnosis and therapeutic strategies of autoimmune diseases.
Methods
Data Collection and Data Processing
To evaluate the amplified cell subsets and genetic programs shared by multiple autoimmune diseases, we collected single-cell RNA-sequencing data of peripheral blood from publicly available data. A total of 51 samples were enrolled in the analysis, after screening the clinical characteristics of the patients. RNA sequencing data of patients with SLE (n = 3) were obtained from the GSE142016 dataset of the GEO database.20 RNA sequencing data of patients with pSS (n = 5) were obtained from the GSE157278 dataset.21 RNA sequencing data of patients with juvenile dermatomyositis (JDM, n = 4) were obtained from the GSE190684 dataset.22 RNA sequencing data of RA (n = 19) and healthy control (HC, n = 4) were obtained from Genome Sequence Archive in BIG Data Center, code HRA000155.23 RNA sequencing data of patients with relapsing-remitting multiple sclerosis (RRMS, n = 12) were obtained from the GSE133028 dataset.24 RNA sequencing data of BD (n = 4) was obtained from the GSE198616 dataset.25
Quality Control and Batch Integration of Single-Cell Sequencing Data
The “Seurat” R package (v4.3.1) was mainly used for quality control procedures and downstream bioinformatics analysis.26 The Seurat R software package was used to input and process the original gene expression matrix. We first filtered out low-quality cells that met any of the following criteria. For quality control of single-cell RNA sequencing data, genes expressed in at least three cells were retained. Given that mitochondrial content was below 5% in most cells, a more stringent mitochondrial threshold of 7.5% was applied to ensure consistency across datasets, and cells exceeding this threshold were excluded. Cells were retained only if they exhibited fewer than 6000 unique genes and UMI (Unique molecular identifier) counts below 20,000. To eliminate red blood cell contamination, the erythrocyte-specific gene set (HBA1, HBA2, HBB, HBD, HBE1, HBG1, HBG2, HBM, HBQ1, and HBZ) was used. The PercentageFeatureSet function in Seurat calculated the proportion of erythrocyte genes per cell, and cells with a red blood cell score below 3% were retained for downstream analysis. DoubletFinder (v2.0) package was then installed to remove potential double cells.27 To mitigate batch effects across distinct datasets and samples, we employed the Harmony algorithm (v1.2.0) to perform batch correction on dataset and sample ID. Subsequent dimensionality reduction clustering and UMAP visualization were conducted based on the corrected PCA embeddings. The integration of cells across different datasets and samples was evaluated using the Local Inverse Simpson’s Index (LISI) algorithm.
FindClusters Performs Graph-Based Clustering on the Neighbor Graph
Principal component analysis (PCA) was carried out by applying the functions “FindNeighbors” and “FindClusters” with a resolution of 0.9. This was followed by t-distribution stochastic neighbor embedding (t-SNE) and uniform manifold approximation and projection (UMAP) to identify the top 20 principal components (PC). The cells were clustered at a resolution of dims = 30, and the clustering results were shown in the form of a tSNE / UMAP scatter plot. Then, according to the typical marker genes of cells, different clusters are annotated to cell types. A total of 29 clusters were identified and were divided into 11 types of cell populations. Last, the ggplot2 software package was used to calculate and display the proportion of different cell types.
Differential Expression and Functional Enrichment Analysis
The Seurat function “FindMarkers” used the likelihood ratio test (“test.use: bimod”) to detect differentially expressed genes in scRNA-seq data between two groups. The significant threshold was set to log2-fold change ≥ 0.25, adjusted P value ≤ 0.05, min.pct parameter was set to 0.1. Finally, clusterProfiler was used to analyze the enrichment of functional GO and KEGG terms in the gene list, and the adjusted P value was less than 0.05.
Clinical Sample Collection and Processing
Clinical patients with autoimmune diseases were recruited at the China–Japan Friendship Hospital. Peripheral blood samples were collected from a total number of patients with pSS patients. The collection of tissues from pSS patients has been approved by the Clinical Research Ethics Committee of the China–Japan Friendship Hospital (No.: 2022-KY-74-1).
10 mL of blood was collected from donors in BD Vacutainer EDTA tubes (BD Biosciences). PBMCs were isolated from people (with and without autoimmune diseases) whole blood by Ficoll-Paque (GE Healthcare, Illinois, USA) gradient centrifugation. Cells were cryopreserved in fetal bovine serum (FBS) (Sigma-Aldrich) containing 10% dimethyl sulfoxide and stored below −135°C until tested. Salivary gland (SG) biopsies were collected from the people (with and without pSS) and were fixed with 4% paraformaldehyde and embedded with petroleum wax histological evaluation.
Flow Cytometry of PBMCs
PBMCs were obtained from peripheral blood, belonging to patients with autoimmune diseases and normal people. After the isolation of PBMCs, cells were cultured with complete medium alone. To evaluate CCL3, CCL4, and CXCL8 production from T cells, B cells, and monocytes, cells were stained with PE/Cyanine7 anti-human CD16 (Cat: 302016) and FITC anti-human CD14 (Cat: 325608), conjugated with the appropriate fluorochrome. After centrifugation, cells were incubated with CCL4 (MIP-1 beta) Monoclonal Antibody (FL34Z3L) and APC anti-human IL-8. As described above, the BD CantoII flow cytometry was used for data detection, and Diva analysis software was used to analyze the data.
Immunohistochemistry (IHC)
3-µm-thick paraffin-embedded sections from the patients with pSS and normal people were used to detect the expression of CCL3, CCL4, CXCL8, and DAPI. Control staining, without primary antibodies, was used as negative control. Afterward, the evaluation of CCL3/CCL4/CXCL8-expressing cells in SG tissue was assessed by two independent investigators on photomicrographs obtained from a high-resolution laser scanning confocal microscope (Abberior Instruments).
Tyramide Signal Amplification (TSA)-Based Multicolor Immunofluorescence
Multicolor immunofluorescence analyses were performed using 3-μm-thick sections of formalin-fixed paraffin-embedded (SG) tissues. The slides were deparaffinized in xylene and hydrated in a series of decreasing graded ethanol series. After heat-induced antigen retrieval in buffer (CCL3 and CXCL8, PH = 9, CD14, PH = 6), samples were permeabilized with 0.5% Triton X-100, blocked with 5% goat serum-phosphate-buffered saline (PBS), and sequentially co-stained with antibodies recognizing CCL3 (Abcam, ab259372), CXCL8 (proteintect, 27095–1), CD14 (Abcam, ab183322), and DNA. A TSA indirect kit (PerkinElmer) was used according to the manufacturer’s instructions. A TSA indirect kit from PerkinElmer was used according to the manufacturer’s instructions. Images were collected using the Vectra Polaris system from Akoya Biosciences, and HALO Image Analysis Software was used for analysis. A high-resolution laser scanning confocal microscope (Abberior Instruments) was used to collect images and analyze data.
Results
Single-Cell Transcriptome Profiles of PBMC from Multiple Autoimmune Diseases versus Normal Control Patients
To compare the difference in cell composition and gene regulation pattern activation in peripheral blood of patients with various autoimmune diseases and normal people (Figure 1A), we integrated and analyzed scRNA-seq of autoimmune diseases and normal people (Table S1). After strict quality control and filtration, the harmony led to a good integration (Figure S1A and B), We employed the LISI to evaluate the extent of cell mixing across different samples following data integration with Harmony. The results demonstrated effective batch effect removal across different datasets and samples (Figure S1C and D). A total of 350,043 PBMC cells were collected and multiple cell types (Figure 1B) were identified. According to the classical cell markers in PBMC (Figure S1E), T cell subsets (CD3D, CD8A, CD4) were identified. According to the expression of CD4 and CD8A, CD4 T cells, CD8 T cells, and DN T cells were identified (neither CD4 nor CD8A Express). NK cells (GNLY, NKG7), B cells (MS4A1), and plasma B cells (CD38, MZB1) were all labeled. In myeloid cells, monocyte (Mon) (CD14, CD16, LYZ) and macrophage (Mø) subpopulations (CD68, CD163, LYZ) were identified. Dendritic cell subsets (DCs) (HLA-DQA1, LYZ), specifically included plasmacytoid dendritic cells (pDC) (LILRA4, IRF7 marker) and myeloid dendritic cell subsets (mDCs) (CD1C, LYZ). Others also contain proliferting cells cells. Detailed cell markers are shown in Figure 1C. Different cell types were further clustered based on the expression of the top 1000 genes with the largest standard deviation differences among cell subpopulations. T cells and NK cells clustered together. Monocytes, macrophages, and dendritic cells (mDC, pDC) clustered together (Figure 1D). Of note, the proportion of cell subpopulations in different diseases showed that monocytes and macrophages accounted for the largest proportion of myeloid cells in each autoimmune disease. (Figure 1E). The heterogeneity of monocytes and macrophages may be an important factor in various autoimmune diseases. To evaluate the cellular inflammatory functions that were commonly and specifically upregulated in autoimmune diseases, based on the GSEA algorithm, the upregulation of pathways was calculated in DC cells and mononuclear phagocytes in all autoimmune diseases. In DC cells, multiple humoral immune responses and eosinophil migration-related pathways were significantly upregulated in pSS, RRMS, RA, BD, and JDM (Figure 1F). In mononuclear phagocytes, immune response and inflammatory response pathways were upregulated. Granulocyte and eosinophil migration pathways significantly up-regulated in pSS, RRMS, RA, BD, and JDM (Figure 1G).
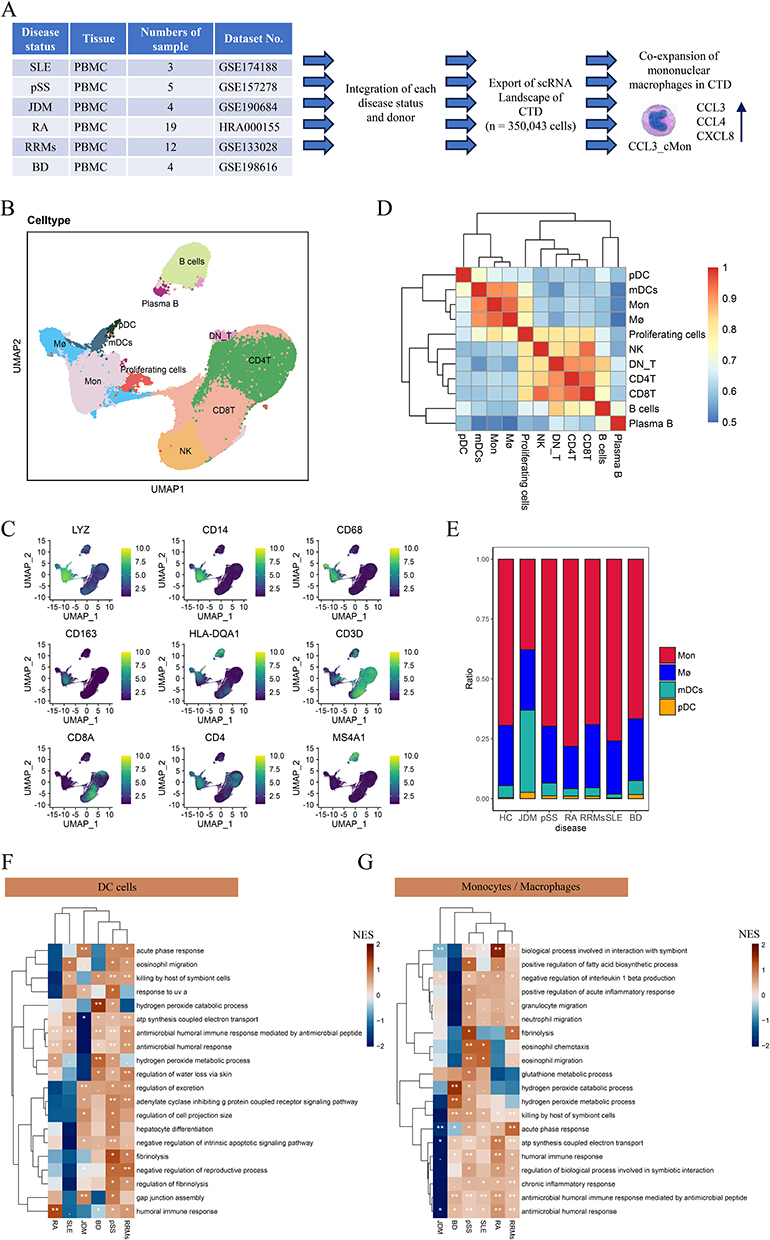 |
Figure 1 The cellular landscape of multiple autoimmune diseases. (A) Schematic diagram of the research process. (B) UMAP plot showing cell clustering in the peripheral blood of healthy people and autoimmune diseases. (C) Clustering diagram showing different cell subpopulations based on the expression of 1000 hypervariable genes. (D) UMAP plots showing the expression of the main markers of each cell type. (E) Histogram showing the proportion of peripheral blood myeloid cells in healthy people and patients with autoimmune diseases. (F) The pathways upregulated by DC cells in different autoimmune diseases calculated based on GSEA. (G) The pathways upregulated by mononuclear phagocytes in different autoimmune diseases calculated based on GSEA. Abbreviation: Mø, macrophages. |
Extensive Infiltration of CCL3+ Classical Monocytes in a Variety of Autoimmune Diseases
Mononuclear phagocytes played a key role in the pathogenesis of autoimmune diseases, and they influenced the systemic inflammatory status in patients with autoimmune diseases by releasing inflammatory factors, chemokines, and metabolites.28 To investigate the dynamic evolution of mononuclear phagocyte subpopulations, we extracted mononuclear phagocyte subsets and divided them into 15 cell subpopulations, including CCL3+ classical monocytes (CCL3+ cMo), CD16+ macrophages (CD16+ Mø), PPBP+ cMo, and other cell subpopulations (Figure 2A). CCL3+ cMo subsets highly express CCL3, CXCL8, CCL4, NFKB1A, IL1B, and other genes. The function involves positive regulation of interleukin 8 (CXCL8) production, positive regulation of wound healing, blood coagulation intrinsic pathway, and acute inflammatory response (Figures 2B and S2B).
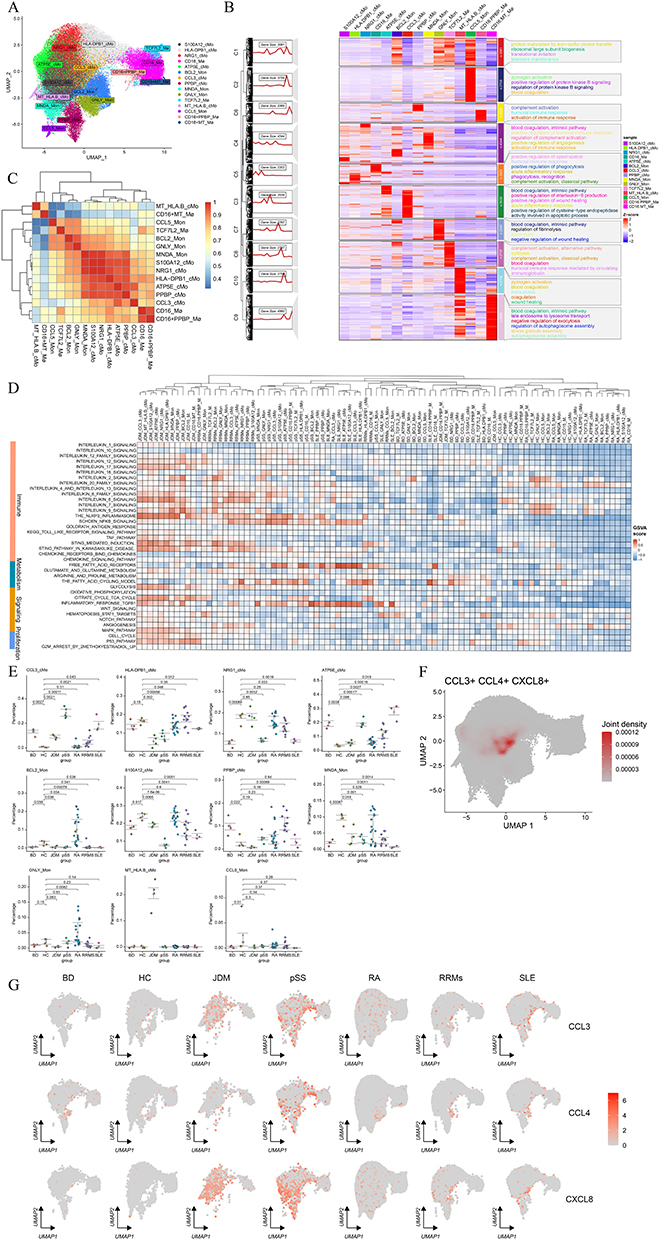 |
Figure 2 Differences and commonalities of mononuclear phagocytes in multiple autoimmune diseases. (A) UMAP plot showing the clustering of mononuclear phagocytes. (B) Heat map of specific up-regulated gene expression and subpopulation-specific functional enrichment of different mononuclear phagocyte subpopulations. (C) Cluster diagram showing mononuclear phagocytes calculated by the first 1000 genes based on standard deviation sorting. (D) Differences in inflammation, metabolism, proliferation, and functional program activation of common signaling pathways in mononuclear phagocytes under different disease states. (E) Differential analysis showing the proportion of identified mononuclear phagocyte subsets in the peripheral blood of healthy people and patients with connective tissue disease. (F) The expression of CCL3, CCL4, and CXCL8 in mononuclear phagocytes. (G) UMAP plots showing the expression of CCL3, CCL4, and CXCL8 genes in the mononuclear macrophage subpopulation under different disease states. Abbreviation: Mø, macrophages. |
We performed a cluster analysis of gene expression between mononuclear phagocyte types to find the subsets with similar characteristics. The results showed that the gene expression characteristics of CD16+ PPBP+ macrophages were similar to those of CD16+ macrophages. The gene expression profiles of MNDA+ Mon, S100A12+ cMo, NRG1+ cMo, HLA-DPB1+ cMo, ATP5E+ cMo, and PPBP+ cMo were also similar (Figure 2C). In addition, we performed a cluster analysis of the characteristics of functional activation of various mononuclear phagocytes under different disease states. The GSVA algorithm was used to estimate the score of each cell type pathway in different diseases. We found that the activation of related pathways in mononuclear phagocytes was disease-specific, but the same activation patterns of inflammatory and metabolic pathways were also shared between different diseases. For example, the NFKB signaling and fatty acid metabolism pathways were highly activated in mononuclear phagocyte subsets in SLE and pSS patients. The activation of interleukin-related signaling pathways (interleukin 6 signaling pathway, interleukin 7 signaling pathway, interleukin 8 signaling pathway, interleukin 9 signaling pathway, etc.) and glycolysis pathway was high in JDM patients, RRMs patients, and pSS patients. Monocyte phagocytes of pSS, JDM, and RRMs were highly activated in the inflammasome NLRP3 pathway (Figure 2D). We further examined the expression patterns of genes involved in inflammatory immune responses, metabolism, phagocytosis, angiogenesis, etc., in two distinct subsets of monocyte-macrophage populations. It was observed that CCL3+ cMo upregulated genes associated with the NF-κB pathway (such as NFKBIA, IL1A, and CXCL8) as well as numerous chemokines (including CXCL22, CXCL8, and CCL3). Meanwhile, ATP5E+ cMo upregulated genes in the interleukin-6-related pathway (such as IL6R, JAK1, and STAT3). Additionally, BCL2+ Mon cells exhibited robust glycolysis and fatty acid metabolism (Figure S2C). As indicated above, the secretion of chemokines by the CCL3+ cMo subset is significantly associated with the pronounced activation of its NF-κB pathway.
To identify disease-specific amplified subpopulations, the proportions of various types of peripheral blood mononuclear phagocytes were compared in different patients and healthy individuals. Some disease-specific subpopulations were identified, such as BCL2+ Mon and GNLY+ Mon subpopulations, which were independently amplified only in RA. MT+ HLA.B+ cMo cells were amplified in JDM disease. Importantly, we identified a subpopulation named CCL3+ cMo that is co-amplified in multiple autoimmune diseases (BD, JDM, pSS, RRMS, SLE) (Figures 2E and S2A). This subpopulation mainly expressed chemotaxis-related genes such as CCL3, CCL4, and CXCL8 in the UMAP map of mononuclear phagocytes (Figure 2F), may play an important chemotactic function for other cells. The expression of CCL3, CCL4, and CXCL8 was higher in Sjögren’s syndrome pSS and JDM than in other diseases (Figure 2G). CCL3+ cMo cells were significantly enriched in patients with autoimmune diseases, particularly pSS and JMD. Their gene expression signature, characterized by high chemokine production, suggests a potential role in recruiting immune cells and modulating inflammation, which may contribute to disease pathogenesis.
CCL3+ Classical Monocyte Subsets Have Both High Inflammatory Characteristics and Strong Chemokine Release Ability
To investigate the functional activation status of different mononuclear phagocyte subsets, we used Aucell to score the mononuclear phagocyte-related functions of different subsets. CCL3+ cMo, CD16+ PPBP+ Mø, and PPBP+ cMo subgroups had high activation of M1-type inflammatory features (Figure 3A). In most subpopulations, innate immune activation scores were high (Figure 3B). To characterize the chemotaxis of cells, we collected the genes of the chemokine family and used this chemokine signature to evaluate the chemotaxis of different cells. CCL3+ cMo and PPBP+ cMo exhibited higher chemotaxis scores and stronger chemotactic ability compared with other cells (Figure 3C). The antigen presentation ability was mainly activated in HLA-DPB1+ cMo, CD16+ PPBP+ Mø, and CCL3+ cMo subsets (Figure 3D). We further employed the FindMarkers function to analyze upregulated genes in the CCL3⁺ cMo subset. Genes with an average log2 fold change (avg_log2FC) > 0.25 were selected for Gene Ontology (GO) enrichment analysis. The results revealed that CCL3⁺ cMo primarily upregulated pathways involved in responses to lipopolysaccharide (LPS) and bacteria, leukocyte chemotaxis, and antigen presentation/activation. This indicates that CCL3⁺ cMo represent a highly inflammation-activated cellular subpopulation, characterized by their ability to chemoattract immune cells and activate T cells via antigen presentation (Figure 3E). Therefore, the CCL3+ cMo subsets have high inflammatory characteristics, strong chemokine release ability, and antigen presentation ability. As previous studies have shown, CCL3 promotes proinflammatory macrophage polarization through activation of the Ccl3-C-C motif chemokine receptor 5-p38/interferon regulatory factor 5 pathway.29 They were involved in the pathogenesis of RA by promoting the migration of inflammatory factors to synovial tissue, destruction of bone and joint, and angiogenesis.30
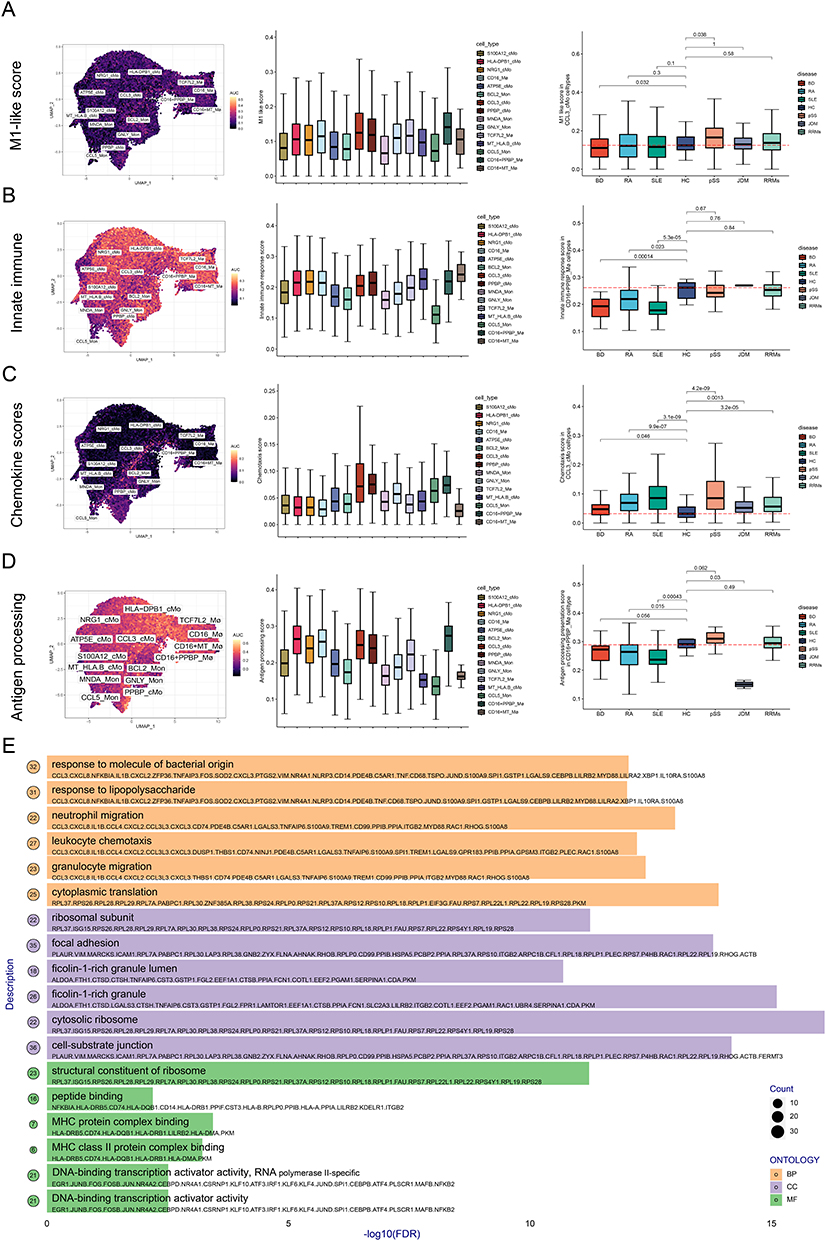 |
Figure 3 The Aucell algorithm scores the functional activation status of different mononuclear phagocyte subsets. (A) M1-like score of different cell subsets. (B) Innate immune scores of different cell subsets. (C) Migration ability scores of different cell subsets. (D) Antigen presentation ability scores of different cell subsets. (E) Gene Ontology (GO) enrichment analysis of upregulated genes (avg log2 fold change > 0.25) in the CCL3-positive classical monocyte (cMon) subset. Abbreviation: Mø, macrophages. |
The Communication Regulatory Network of CCL3+ Classical Monocytes in Autoimmune Diseases
To further investigate the major crosstalk of CCL3⁺ cMo cells with other cells, we observed communication networks between different subsets of monocyte phagocytes and T cells in diseased and healthy individuals. First, we compared the signal crosstalk network within the mononuclear phagocyte subpopulations. Compared with the normal group, CCL3+ cMo in pSS up-regulated HLA-DQB1-CD4, HLA-DQA1-CD4, MIF-(CD74-CD44), LGALS9-CD44, ANXA1-FPR1, CCL3-CCR1, CCL3L3-CCR1-related signal networks (Figure 4A).
 |
Figure 4 CCL3 classical monocytes enhance antigen presentation to T cells and chemotaxis to other mononuclear phagocytes. (A) Communication signals within mononuclear phagocyte subsets up-regulated in pSS patients. (B) Heatmap showing up-regulated and down-regulated signaling pathways in pSS patients. (C) Violin plot showing gene expression of main signaling pathways in different mononuclear phagocytes and T cell subsets. (D) Differences in the information flow of cell communication signaling pathway between pSS patients and healthy people. (E) Difference of signal communication between pSS and healthy mononuclear phagocytes on T cells. Abbreviation: Mø, macrophages. |
In addition to internal crosstalk signaling, mononuclear macrophages are closely associated with the activation and proliferation of T cells.31 Furthermore, we observed the communication between mononuclear phagocyte subsets and T-cell subsets. The number of signals released by mononuclear phagocytes in pSS was generally significantly increased, especially in CD16+ PPBP+ Mø, GNLY+ Mon cells, and CCL3+ cMo cells (Figure 4B). CCL3+ cMo indeed up-regulated a series of chemokine signals such as CCL4, CCL3, CCL3L3, CXCL2, and CXCL8. Moreover, CCR1 was mainly expressed in mononuclear phagocyte subsets CCL3+ cMo, GNLY+ Mon, HLA-DPB1+ cMo, PPBP+ cMo, and S100A2+ cMo. It suggested that CCL3+ cMo has a chemotactic effect on other mononuclear macrophage subsets (Figure 4A and C).
We further analyzed the overall signaling network, patients with pSS up-regulated THBS, TNF, CXCL, MIF, MHC-I and other signaling pathways (Figure 4D). CCL3+ cMo in pSS patients have strong signal crosstalk with cytotoxic CD8+ T cells (MHC signaling pathway), indicating these cells have a strong ability of antigen presentation (Figure 4E). Furthermore, we compared the differences in communication signals emitted by CCL3 cMo across different autoimmune diseases. Based on the difference in signaling pathways in autoimmune diseases, the effect of CCL3 cMo on T cells differs in various diseases. CCL3+ cMo in peripheral blood of pSS, RRMS, and SLE patients significantly affect central memory CD8+ T cells and cytotoxic CD8+ T cell antigen-related functions (Figure S3B and C). CXCL chemotactic signaling is significantly upregulated in RRMS, SLE, BD, JDM, and pSS (Figure S4A and B). THBS, TNF, IL6, and MHC-I signaling are enhanced in the peripheral blood of RRMS patients compared to healthy individuals (Figure S4A and B). By contrast, the effect of CCL3+ cMo on T cells is not obvious in RA patients (Figure S3A). The whole-cell communication further showed that the intensity of signal crosstalk between mononuclear phagocytes and T cells in RA patients was lower than that in healthy people (Figure S4A and B). This further indicates that CCL3+ cMon orchestrate a more pronounced immune network in pSS, RRMS, and SLE, whereas their immunological contribution appears comparatively diminished in RA.
The Changes of Functional Pathways of CCL3 cMo Cells in Different Autoimmune Diseases
To clarify the changes in gene characteristics and functional activation of CCL3+ cMo cells in various autoimmune diseases, the up-regulated and down-regulated cell functions of CCL3+ cMo cells were tested in different diseased patients compared with normal people. According to these figures (Figures 5A and B, S5A and B), we found that CCL3+ cMo cells up-regulated the cytoplasmic translation pathway in multiple diseases (pSS, SLE, BD, and RRMS), indicating that CCL3+ cMo cells were in an active state of protein production. These cells also up-regulated antigen processing and presentation of peptide antigens in multiple disease states (pSS, SLE, and RRMs). CCL3+ cMo cells in pSS patients had the function of neutrophil chemotaxis and regulation of T cell activation, which was consistent with previous research.31 In addition, lipopolysaccharide response activation and ATP metabolism were also more vigorous, representing a state of high metabolism and immune-inflammatory stress (Figure 5A). SLE patients and pSS patients had the same CCL3+ cMo cell activation program, with up-regulated gene programs of ATP metabolism, leukocyte migration, and immune inflammation (Figure 5B). However, the up-regulated pathway in JMD (regulation of apoptotic signaling pathway, cellular response to chemical stress, and intrinsic apoptotic signaling pathway) and the up-regulated pathway in RA (negative regulation of transferase activity, response to gravity, and response to parathyroid hormone) were not associated with inflammatory activation (Figure S5C and D). In summary, CCL3+ cMo cells indeed played an important pro-inflammatory role in diseases such as pSS, SLE, BD, and RRMs. Conversely, the activation of the pro-inflammatory function of CCL3+ cMo cells in RA and JDM was weak.
 |
Figure 5 The internal differentiation trajectory and intercellular signal network of CCL3 cMo cells. (A) Radar plot showing the functional changes of CCL3 cMo cells in healthy people and pSS patients. (B) Radar plot showing the functional changes of CCL3 cMo cells in healthy people and SLE patients. (C) UMAP of various mononuclear phagocytes. (D) Intracellular evolution of mononuclear phagocytes based on velocity calculation. (E) The expression of CCL3, CCL4, and CXCL8 in the evolution of mononuclear phagocytes. (F) The signal network of CCL3 cMo cells releasing to other mononuclear phagocytes subsets under different autoimmune diseases. Abbreviation: Mø, macrophages. |
Analysis of the internal differentiation process suggested that there was an orderly differentiation process in classical monocytes. CCL3+ cMo cells were differentiated from HLA-DPB1+ cMo cells, but eventually differentiated into S100A12+ cMo cells, and may also transit to GNLY+ Mon and BCL2+ Mon cells. Therefore, we speculated that CCL3+ cMo cells are a transitional phase of classical monocyte differentiation (Figure 5C and D). As the classical monocyte transition stage, CCL3+ cMo cells showed high expression of CCL3, CCL4, and CXCL8 genes (Figure 5E). To further demonstrate the communication network of CCL3 cMo cells among all mononuclear phagocyte subsets, the communication signals of CCL3 cMo cells to other monocytes were analyzed under different disease states. In pSS, BD, JDM, and SLE, CCL3+ cMo cells and other monocytes had strong CCL3-CCR1, LGALS9-CD45/CD44, and ANXA1-FPR1 communication signals. Notably, compared with other diseases, only CCL3+ cMo cells in pSS patients can release CXCL8, CXCL2, and CXCL3, chemoattracting TCF7L2_Mø cells expressing CXCR2 (Figure 5F).
The CCL3+ Classical Monocytes Exhibit a Ubiquitous Activating Effect on T Cells Across Multiple Autoimmune Diseases
Based on the important link between macrophages and NK/T cells, NK and T cells were classified in detail to explain the cell communication relationship between CCL3+ cMo cells and different T cell subsets. First, the subsets of NKT cells were identified through marker genes, NK cells (NK_01, NK_02), CD8+ T cells (CD8T_GZMH, CD8T_ATXN1, CD8T_GZMK, CD8T_GZMB, CD8T_LEF1, CD8T_AOAH), CD4+ T cells (CD4T_CCR7, CD4_GPR183, CD4T_LEF1) and other T cells (AFF3_T and PPBP_T) were identified (Figures 6A and S7). CD4T_LEF1 and CD8T_LEF1 highly expressed LEF1 molecules, respectively, representing undifferentiated CD4+ T and CD8+ T initial cell states. The proportion of the former was relatively high in healthy people and RA patients, while the latter mainly existed in JDM patients. CD8T_GZMB cells accounted for a high proportion of JDM diseases and highly expressed GZMB, a protein that promotes cell disintegration and represents a state of cytotoxic CD8+ T cells. NK_01 cells accounted for a high proportion of pSS patients. Notably, healthy individuals had almost no PPBP+ T cells, while the patients in other autoimmune diseases showed amplified PPBP+ T cells and highly expressed PPBP chemokines (Figure 6B and C). Importantly, in patients with JDM, pSS, BD, and SLE, CCL3+ cMon primarily interacted with T cell subsets (CD4T_CCR7, CD4T_GPR183, CD8T_GZMH) via LGALS9-CD44/CD45 and MHC-I molecules (HLA-A/HLA-B/HLA-C) - CD8A/CD8B signaling pathways (Figure 6D). This also suggests that the interaction between CCL3+ cMo and T cells is primarily manifested in antigen presentation and the functional activation of T cells. Notably, this interaction is even more pronounced in diseases such as primary pSS, BD, and SLE.
 |
Figure 6 The signal communication network between CCL3 cMo cells and NK/T cells. (A) UMAP plot showing the subpopulations of NK and T cells. (B) The proportion of NK and T cell subsets in healthy people and different autoimmune diseases. (C) Differences in the proportions of different NK/T cell subsets among various disease groups. (D) Receptor-ligand signals from CCL3 cMo cells to different NK/T cells. |
CCL3+ Classical Monocytes Expand in the Peripheral Blood and Salivary Gland Tissues of Patients with pSS
The expression of CCL3, CCL4, and CXCL8 genes was significantly increased in peripheral blood mononuclear phagocytes from patients with pSS compared with healthy individuals (Figure 7A). Furthermore, these chemokines were broadly upregulated in the peripheral blood of patients with other autoimmune disorders (Figure S6A–E). We collected peripheral blood and salivary gland tissue samples from pSS patients exhibiting the highest CCL3 expression for experimental investigation, and examined the infiltration of CCL3+ classical monocytes in the salivary gland tissues of control group (The labial gland biopsy was performed due to xerostomia and xerophthalmia, but no pathological manifestations of pSS were observed) and patients with pSS. The immunohistochemistry images indicated the higher expression of CCL3, CCL4, and CXCL8 in gland tissues of pSS patients than in control group (Figure 7B). Tyramide signal amplification (TSA)-based multiple immunohistochemistry directly demonstrated that, compared with control group, the proportion of CCL3+ classical monocytes in the salivary glands of patients with pSS was significantly elevated (Figure 7C and D). To quantify the absolute proportion of CD14+ CCL3+ classical monocytes in the peripheral blood of patients with pSS, we employed two marker genes (CCL4 and CXCL8) to label this type of cells and conducted flow cytometry analysis. Flow cytometry analysis demonstrated a notable amplification of both CD14+ CCL4+ classical monocytes and CD14+ CXCL8+ classical monocytes within the PBMCs of patients with pSS, suggesting that the expansion of this subpopulation was most obvious in pSS diseases (Figure 7E–H). As indicated above, CCL3+ classical monocytes are strongly associated with the pathogenesis of pSS and are also a characteristic feature shared by various autoimmune diseases. Their central role makes them promising potential therapeutic targets for future clinical intervention.
 |
Figure 7 Salivary gland tissue of patients with pSS showed infiltration and amplification of CCL3+ CCL4+ CXCL8+ classical monocytes. (A) The different expression of CCL3, CCL4, and CXCL8 genes in peripheral blood mononuclear phagocytes of healthy people and patients with pSS. (B) Immunofluorescence staining of tissue sections showing CCL3, CCL4, and CXCL8 genes in salivary gland tissues of healthy people and patients with pSS. (C) The infiltration of CCL3+ CCL4+ CXCL8+ classical monocytes in the salivary gland tissue of healthy people. (D) The infiltration of CCL3+ CCL4+ CXCL8+ classical monocytes in the salivary gland tissue of patients with pSS. (E) Flow cytometric showing the proportion of CD14+ classical monocytes in peripheral blood of healthy individuals and pSS patients. (F) Patients with pSS significantly increased the proportion of CCL3+ classical monocytes of PBMC. (G) Flow cytometric showing the proportion of CXCL8+ classical monocytes in peripheral blood of healthy individuals and pSS patients. (H) Patients with pSS significantly increased the proportion of CXCL8+ classical monocytes of PBMC. **P < 0.01 compared with the control group; ***P < 0.001 compared with the control group; ****P < 0.0001 compared with the HC group. |
Discussion
The incidence of inflammation and autoimmune diseases has been gradually increasing. Currently, the treatment of autoimmune diseases calls for the identification of more novel therapeutic targets within immune cells.32–34 Monocyte-macrophages serve as the initiating factors in the pathogenesis of inflammation, and different subsets of monocyte-macrophages play a role in amplifying inflammatory responses in various inflammatory diseases.
Herein, we have obtained samples from patients with autoimmune diseases and normal people, and successfully mapped the first single-cell transcriptome atlas. This work provided high-quality data to reveal in-depth autoimmune disease-related alterations in gene expression in each cell type and subtype at the single-cell level. We identified a cell subset called CCL3+ cMo that may have high chemotaxis features and immune regulation ability. Monocytes can be used as a rapid inducer of inflammation,35 so we used a strategy to further resolve specific monocyte subsets and found that CCL3+ cMo cell subsets were significantly up-regulated in a variety of autoimmune diseases. CCL3 and its receptors constitute a key component of the immune system. Research indicates that CCL3 promotes the migration and infiltration of chemotactic immune cells into target organs, thereby contributing to local immune responses.36 Numerous investigations have documented elevated expression of CCL3 in various autoimmune diseases, where it influences both disease susceptibility and progression.37 Furthermore, evidence supports the involvement of CCL3 in the pathogenesis of multiple auto-inflammatory immune disorders.38 In addition, CCL3+ cMo cells highly expressed CCL4, CCL3, CCL3L3, CXCL2, and CXCL8. Our findings are consistent with evidence for expanded chemokines in peripheral blood from autoimmune diseases.39,40 Based on the GSEA algorithm, the upregulation of pathways were calculated in mononuclear phagocytes from all autoimmune diseases. We found that multiple immune response and inflammatory response pathways were up-regulated in mononuclear phagocytes. The granulocyte and eosinophil migration pathways were significantly up-regulated. The strong association between CCL3+ cMon cells and the upregulation of CCL3, CCL4, and CXCL8 suggests a potential role for this subset in the progression of autoimmune disease. This mechanism may contribute to the disease improvement of autoimmune diseases and needs to be explored in further studies.
Monocytes have strong migration, chemotaxis, and adhesion, and they can drive inflammation and fibrosis tissue remodeling by secreting pro-inflammatory and pro-fibrotic cytokines.41,42 In our study, CCL3+ classical monocytes (cMo) exhibit a marked transcriptional signature associated with M1-like genes, chemotaxis, and antigen presentation calculated by the Aucell algorithm. Cluster analysis also confirmed that mononuclear phagocytes had varying degrees of activation of inflammatory pathways and metabolic pathways. CCL3 mediated neutrophil migration in immune inflammation through the continuous release of TNF-α and LTB4), recruited CD14+ CD16+ circulating monocytes and worsened inflammation.42,43 Furthermore, CXCL8 regulated specific leukocyte infiltration in inflammation.44 Abnormal levels of CCL3 and CCL4 were confirmed to be characteristic of cytokine network disorders.28 This immune signature is evident in gene cluster analysis. Antigen presentation was mainly activated in CCL3+ cMo cells, and the chemotactic ability score showed that this subpopulation had high chemotactic ability. Furthermore, we found that CCR1 was mainly expressed in CCL3+ cMo, especially in pSS where signal crosstalk such as CCL3-CCR1 and CCL3L3-CCR1 was enhanced. The signal pathways that promoted disease progression were strengthened, such as the cytoplasmic translation pathway, antigen processing, and presentation of peptide antigen. These findings demonstrated that CCL3+ cMo cells had strong inflammatory characteristics and high chemotaxis to other monocytes.
In the communication network between mononuclear phagocyte subsets and T cells in different diseases, the signal crosstalk increased significantly, except for rheumatoid arthritis. Notably, the communication network showed that CCL3+ cMo cells had strong signal interference on the antigen presentation-related pathways of Cytotoxic CD8+ T or memory CD8+ T cells. Memory CD8+ T cells promoted the delivery of chemokines and IFNγ, suggestive of relation to our findings.45 CCL3 cMo cells interacted with T cell subsets (CD4T CCR7, CD4T GPR183, CD8T GZMH) through LGALS9-CD44/CD45. These cells also interacted with CD8T GZMH and CD8T LEF1 through MHC class I receptors to promote the expansion and activation of T cells. CCL3 cMo cells highly expressed CCL3 and CCL4, which can directly recruit antigen-presenting cells to cause immune cell infiltration.46 CCL3+ cMo cells had a generally enhanced signal to PPBP T cells (a cross-cell subpopulation of myeloid cells and T cells with high inflammation scores). The expansion of PPBP T cells was observed in all autoimmune diseases, and PPBP chemokines were highly expressed. However, there were almost no PPBP T cells in healthy people. This finding suggested the correlation between CCL3+ cMo cells and inflammatory activation. We further observed the alterations of gene characteristics and functional activation of CCL3+ cMo cells in various diseases. It was found that the functional pathway changes of CCL3 cMo cells in different diseases were disease-specific. CCL3+ cMo cells up-regulated antigen presentation-related signaling pathways in multiple disease states (pSS/ SLE /RRMs). Only in JMD and RA, their pathway signals related to chemotaxis and antigen presentation were not significantly upregulated, and their proinflammatory functions were weak. A summary of our main findings was credible evidence that CCL3+ cMo cells promoted chemotaxis of T cells in multiple autoimmune diseases. CCL3+ classical monocytes exhibit distinct recruitment/chemotactic and antigen-presenting capabilities. They can chemoattract and prime T cells with antigen-specific responses, which may potentially contribute to the pathogenesis of autoimmune diseases.
Analysis of the internal differentiation process based on velocity calculations found that CCL3+ cMo was a transitional stage of classical monocyte differentiation. CCL3+ cMo differentiated from HLA-DPB1 cMo cells and eventually differentiated into S100A12 cMo cells. In different disease states (pSS, BD, JDM, and SLE), CCL3+ cMo cells and other monocytes had strong communication signals between CCL3-CCR1 receptors. In addition, there were communication signals of LGALS9-CD45/CD44 and ANXA1-FPR1. But only CCL3+ cMo cells in pSS patients released CXCL8, CXCL2, and CXCL3 and chemoattracted TCF7L2+ Mø cells expressing CXCR2. CXCR2 was the receptor for the CXC chemokine, and binding to it caused neutrophil influx and tissue damage. Therefore, testing to block the binding of chemokines (such as CXCL8) to receptors and treatments targeting CCL3+ cMo were of positive significance in experiments.
Especially in pSS, CCL3+ cMo cells increased, and their ability to promote chemotaxis and antigen presentation was particularly enhanced. The results of fluorescence staining of SG sections of patients with pSS showed that the tissues significantly expressed CCL3, CCL4, and CXCL8, consistent with our results from the signal network. Moreover, flow cytometry of peripheral monocytes showed that the expression of CCL4 and CXCL8 in pSS disease was higher than that of healthy people, and the level of pSS patients was the most significant, providing evidence for our findings. Due to the lack of specificity in chemokine receptors, a single receptor can bind multiple ligands and a single ligand may interact with multiple receptors. And different ligands can act as agonists or antagonists upon receptor binding. These challenges have hindered the development of effective compounds in human clinical trials. Our study may offer new perspectives for future drug development.
Limitations
While our study provides a comprehensive transcriptomic landscape of a shared monocyte subset across autoimmune diseases, we acknowledge several limitations. First, our analysis is restricted to peripheral blood samples, which may not fully reflect the functional and phenotypic states of monocytes and macrophages within affected tissues. Tissue-resident macrophages and infiltrating myeloid cells could exhibit context-dependent behaviors that contribute differently to disease pathogenesis. Second, our findings are primarily observational and based on transcriptomic data. While CCL3+ monocytes show strong association with disease states, their precise mechanistic role as pathogenic drivers requires validation through functional studies and protein-level assays. The disease-specific versus general relevance of this subset also warrants further investigation across distinct clinical contexts. Finally, while integration of public datasets enhances generalizability, inherent heterogeneity in sample processing and donor characteristics may introduce unresolved confounding factors. Future studies incorporating spatial transcriptomics, tissue-specific profiling, and mechanistic assays will be essential to extend these findings.
Conclusion
This study provided comprehensive single-cell transcriptome atlas for deciphering the gene expression landscapes of mononuclear phagocyte types in various autoimmune diseases. We identified CCL3+ classical monocytes as a shared transcriptional subset enriched in these diseases, supporting its potential role as a pan-disease inflammatory marker. Through cell–cell communication analysis, we revealed enhanced interactions between mononuclear phagocytes and T/NK cells, suggesting their involvement in autoimmune pathogenesis, providing new ideas for the pathogenesis and treatment of autoimmune diseases.
Ethics Approval and Consent to Participate
This study was conducted in accordance with the Ethical Review of Life Science and Medical Research Involving Human Subjects (China, February 18, 2023). The analysis of publicly available single-cell sequencing data falls under the exemption category, as it involves legally obtained, non-identifiable data without interference in public behavior. For the collection of labial gland pathology samples and peripheral blood, ethical approval was obtained from the Clinical Research Ethics Committee of the China–Japan Friendship Hospital (Approval No. 2022-KY-74-1) in compliance with the principles outlined in the Declaration of Helsinki. Written informed consent was obtained from all participants prior to sample collection.
Funding
This work was supported by Joint Funds of National Natural Science Foundation of China (Grant Number: U23A6012), National High Level Hospital Clinical Research Funding and Elite Medical Professionals Initiative of China–Japan Friendship Hospital (NO.ZRJY2025-QM05), National High Level Hospital Clinical Research Funding (2022-NHLHCRF-LX-02), Capital’s Funds for Health Improvement and Research (2024-1-4065), Elite Medical Professionals project of the China–Japan Friendship Hospital (ZRJY2021-QM14), and Rheumatology Branch of China Association of Chinese Medicine Youth Pei Ying Project (No.202327-007).
Disclosure
Heng Xu, Kai Yuan, and Guangyao Chen are co-first authors for this study. The authors report no conflicts of interest in this work.
References
1. Decker P, Moulinet T, Pontille F, et al. An updated review of anti-Ro52 (TRIM21) antibodies impact in connective tissue diseases clinical management. Autoimmunity Rev. 2022;21(3):103013. doi:10.1016/j.autrev.2021.103013
2. Fortuna G, Brennan MT. Systemic lupus erythematosus: epidemiology, pathophysiology, manifestations, and management. Dent Clin North Am. 2013;57(4):631–655. doi:10.1016/j.cden.2013.06.003
3. Lin Y-J, Anzaghe M, Schülke S. Update on the pathomechanism, diagnosis, and treatment options for rheumatoid arthritis. Cells. 2020;9(4):880.
4. Thorne I, Sutcliffe N. Sjögren’s syndrome. Br J Hosp Med. 2017;78(8):438–442. doi:10.12968/hmed.2017.78.8.438
5. Hatemi G, Seyahi E, Fresko I, Talarico R, Hamuryudan V. One year in review 2020: behçet’s syndrome. Clin Experiment Rheumatol. 2020;38(Suppl 127):5.
6. Wang Y, Xie X, Zhang C, et al. Rheumatoid arthritis, systemic lupus erythematosus and primary Sjögren’s syndrome shared megakaryocyte expansion in peripheral blood. Ann Rheumatic Dis. 2022;81(3):379–385. doi:10.1136/annrheumdis-2021-220066
7. Patel AA, Zhang Y, Fullerton JN, et al. The fate and lifespan of human monocyte subsets in steady state and systemic inflammation. J Exp Med. 2017;214(7):1913–1923. doi:10.1084/jem.20170355
8. Williams H, Mack C, Baraz R, et al. Monocyte differentiation and heterogeneity: inter-subset and interindividual differences. Int J Mol Sci. 2023;24(10):8757. doi:10.3390/ijms24108757
9. Vishnyakova P, Poltavets A, Karpulevich E, et al. The response of two polar monocyte subsets to inflammation. Biomed Pharmacother. 2021;139:111614. doi:10.1016/j.biopha.2021.111614
10. Lin L, Ren R, Xiong Q, Zheng C, Yang B, Wang H. Remodeling of T-cell mitochondrial metabolism to treat autoimmune diseases. Autoimmun Rev. 2024;23(6):103583. doi:10.1016/j.autrev.2024.103583
11. Siouti E, Andreakos E. The many facets of macrophages in rheumatoid arthritis. Biochem Pharmacol. 2019;165:152–169. doi:10.1016/j.bcp.2019.03.029
12. Ushio A, Arakaki R, Otsuka K, et al. CCL22-producing resident macrophages enhance T cell response in Sjögren’s syndrome. Front Immunol. 2018;9:2594. doi:10.3389/fimmu.2018.02594
13. Guo D, Lin C, Lu Y, et al. FABP4 secreted by M1-polarized macrophages promotes synovitis and angiogenesis to exacerbate rheumatoid arthritis. Bone Res. 2022;10(1):45. doi:10.1038/s41413-022-00211-2
14. Vakrakou AG, Boiu S, Ziakas PD, et al. Systemic activation of NLRP3 inflammasome in patients with severe primary Sjögren’s syndrome fueled by inflammagenic DNA accumulations. J Autoimmun. 2018;91:23–33. doi:10.1016/j.jaut.2018.02.010
15. Han Y, Zhang Y, Yuan K, Wu Y, Jin X, Huang X. Hyperosmolarity promotes macrophage pyroptosis by driving the glycolytic reprogramming of corneal epithelial cells in dry eye disease. Front Med. 2023;17(4):781–795. PMID: 37266854. doi:10.1007/s11684-023-0986-x
16. Culemann S, Grüneboom A, Nicolás-ávila JÁ, et al. Locally renewing resident synovial macrophages provide a protective barrier for the joint. Nature. 2019;572(7771):670–675. doi:10.1038/s41586-019-1471-1
17. Nehar-Belaid D, Hong S, Marches R, et al. Mapping systemic lupus erythematosus heterogeneity at the single-cell level. Nat Immunol. 2020;21(9):1094–1106. doi:10.1038/s41590-020-0743-0
18. Stubbington MJT, Rozenblatt-Rosen O, Regev A, et al. Single-cell transcriptomics to explore the immune system in health and disease. Science. 2017;358(6359):58–63. doi:10.1126/science.aan6828
19. Mulder K, Patel AA, Kong WT, et al. Cross-tissue single-cell landscape of human monocytes and macrophages in health and disease. Immunity. 2021;54(8):1883–1900.e5. doi:10.1016/j.immuni.2021.07.007
20. Perez RK, Gordon MG, Subramaniam M, et al. Single-cell RNA-seq reveals cell type-specific molecular and genetic associations to lupus. Science. 2022;376(6589):
21. Hong X, Meng S, Tang D, et al. Single-cell RNA sequencing reveals the expansion of cytotoxic CD4+ T lymphocytes and a landscape of immune cells in primary Sjögren’s syndrome. Front Immunol. 2020;11:594658. doi:10.3389/fimmu.2020.594658
22. Neely J, Hartoularos G, Bunis D, et al. Multi-modal single-cell sequencing identifies cellular immunophenotypes associated with Juvenile dermatomyositis disease activity. Front Immunol. 2022;13:902232. doi:10.3389/fimmu.2022.902232
23. Wu X, Liu Y, Jin S, et al. Single-cell sequencing of immune cells from anticitrullinated peptide antibody positive and negative rheumatoid arthritis. Nat Commun. 2021;12(1):4977. doi:10.1038/s41467-021-25246-7
24. Ramesh A, Schubert RD, Greenfield AL, et al. A pathogenic and clonally expanded B cell transcriptome in active multiple sclerosis. Proc Natl Acad Sci USA. 2020;117(37):22932–22943. doi:10.1073/pnas.2008523117
25. Zheng W, Wang X, Liu J, et al. Single-cell analyses highlight the proinflammatory contribution of C1q-high monocytes to Behçet’s disease. Proc Natl Acad Sci USA. 2022;119(26):e2204289119. doi:10.1073/pnas.2204289119
26. Stuart T, Butler A, Hoffman P, et al. Comprehensive integration of single-cell data. Cell. 2019;177(7):1888–1902.e21. doi:10.1016/j.cell.2019.05.031
27. McGinnis CS, Murrow LM, Gartner ZJ. DoubletFinder: doublet detection in single-cell RNA sequencing data using artificial nearest neighbors. Cell Systems. 2019;8(4):329–337.e4. doi:10.1016/j.cels.2019.03.003
28. Liu Y, Gong Y, Xu G. The role of mononuclear phagocyte system in IgA nephropathy: pathogenesis and prognosis. Front Immunol. 2023;14:1192941. doi:10.3389/fimmu.2023.1192941
29. Sheng D, Ma W, Zhang R, et al. Ccl3 enhances docetaxel chemosensitivity in breast cancer by triggering proinflammatory macrophage polarization. J Immunother Cancer. 2022;10(5):e003793. doi:10.1136/jitc-2021-003793
30. Yang Y-L, Li X-F, Song B, et al. The role of CCL3 in the pathogenesis of rheumatoid arthritis. Rheumatol Ther. 2023;10(4):793–808. doi:10.1007/s40744-023-00554-0
31. Ou Q, Power R, Griffin MD. Revisiting regulatory T cells as modulators of innate immune response and inflammatory diseases. Front Immunol. 2023;14:1287465. doi:10.3389/fimmu.2023.1287465
32. Chen G-Y, Liu X-Y, Yan X-E, et al. Total flavonoids of rhizoma drynariae treat osteoarthritis by inhibiting arachidonic acid metabolites through AMPK/NFκB pathway. J Inflamm Res. 2023;16:4123–4140. doi:10.2147/JIR.S418345
33. Zhao Y, Chen G-Y, Fang M. Research trends of rheumatoid arthritis and depression from 2019 to 2023: a bibliometric analysis. J Multidiscip Healthc. 2024;17:4465–4474. doi:10.2147/JMDH.S478748
34. Chen G-Y, Ji X-Y, Li Y, et al. Mechanisms of total glucosides of paeony in alleviating methotrexate-induced liver injury. Drug Des Devel Ther. 2025;19:3407–3423. doi:10.2147/DDDT.S521740
35. Lara S, Akula S, Fu Z, et al. The human monocyte-A circulating sensor of infection and a potent and rapid inducer of inflammation. Int J Mol Sci. 2022;23(7):3890. doi:10.3390/ijms23073890
36. Oliveira CR, Polonini H, Marcucci MC, Vieira RP. MiodesinTM positively modulates the immune response in endometrial and vaginal cells. Molecules. 2022;27(3):782. doi:10.3390/molecules27030782
37. M DLFL, Landskron G, Parada D, et al. The relationship between chemokines CCL2, CCL3, and CCL4 with the tumor microenvironment and tumor-associated macrophage markers in colorectal cancer. Tumour Biol. 2018;40(11):1010428318810059. doi:10.1177/1010428318810059
38. Yang YL, Li XF, Song B, et al. The role of CCL3 in the pathogenesis of rheumatoid arthritis. Rheumatol Ther. 2023;10(4):793–808.
39. Melamud MM, Ermakov EA, Boiko AS, et al. Multiplex analysis of serum cytokine profiles in systemic lupus erythematosus and multiple sclerosis. Int J Mol Sci. 2022;23(22):13829. doi:10.3390/ijms232213829
40. Chen X, Aqrawi LA, Utheim TP, et al. Elevated cytokine levels in tears and saliva of patients with primary Sjögren’s syndrome correlate with clinical ocular and oral manifestations. Sci Rep. 2019;9(1):7319. doi:10.1038/s41598-019-43714-5
41. Distler JHW, Györfi A-H, Ramanujam M, et al. Shared and distinct mechanisms of fibrosis. Nat Rev Rheumatol. 2019;15(12):705–730. doi:10.1038/s41584-019-0322-7
42. Ramos CDL, Canetti C, Souto JT, et al. MIP-1alpha[CCL3] acting on the CCR1 receptor mediates neutrophil migration in immune inflammation via sequential release of TNF-alpha and LTB4. J Leukoc Biol. 2005;78(1):167–177. doi:10.1189/jlb.0404237
43. Zhao X, Gu M, Xu X, et al. CCL3/CCR1 mediates CD14+CD16- circulating monocyte recruitment in knee osteoarthritis progression. Osteoarthritis Cartilage. 2020;28(5):613–625. doi:10.1016/j.joca.2020.01.009
44. Harada A, Mukaida N, Matsushima K. Interleukin 8 as a novel target for intervention therapy in acute inflammatory diseases. Mol Med Today. 1996;2(11):482–489.
45. Boutet M, Benet Z, Guillen E, et al. Memory CD8 + T cells mediate early pathogen-specific protection via localized delivery of chemokines and IFNγ to clusters of monocytes. Sci Adv. 2021;7(36):eabf9975. doi:10.1126/sciadv.abf9975
46. Allen F, Bobanga ID, Rauhe P, et al. CCL3 augments tumor rejection and enhances CD8 + T cell infiltration through NK and CD103 + dendritic cell recruitment via IFNγ. Oncoimmunology. 2018;7(3):e1393598. doi:10.1080/2162402X.2017.1393598
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Single-Cell RNA Sequencing Reveals Peripheral Immune Cell Senescence and Inflammatory Phenotypes in Patients with Premature Ovarian Failure
Liu J, Wang L, Zhong W, Cai J, Sun Y, Li S, Li J, Liu Y, Xiong F
Journal of Inflammation Research 2025, 18:2699-2715
Published Date: 25 February 2025