Back to Journals » Infection and Drug Resistance » Volume 19
Serotype Distribution, Antibiotic Resistance Characteristics, and Virulence Factor Analysis of Group B Streptococcus Isolates in Guangzhou Based on Whole-Genome Sequencing
Authors Ling Y ![]() , Deng Y
, Deng Y ![]() , Zhang P, Yuan K
, Zhang P, Yuan K ![]() , Ye L
, Ye L ![]() , Zhang X, Chen X, Cui S, Liu J, Zhao Y
, Zhang X, Chen X, Cui S, Liu J, Zhao Y ![]()
Received 22 July 2025
Accepted for publication 17 December 2025
Published 15 January 2026 Volume 2026:19 549193
DOI https://doi.org/10.2147/IDR.S549193
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Sandip Patil
Yong Ling,1,* Yangxi Deng,1,* Peipei Zhang,1 Kaixuan Yuan,1 Long Ye,1 Xinqiang Zhang,1 Xiaoli Chen,1 Shanzhao Cui,1 Jie Liu,2 Yunhu Zhao1
1Department of Clinical Laboratory Medicine, Guangdong Provincial People’s Hospital (Guangdong Academy of Medical Sciences), Southern Medical University, Guangzhou, Guangdong, 510000, People’s Republic of China; 2Department of Reproductive Medicine, Guangdong Provincial People’s Hospital (Guangdong Academy of Medical Sciences), Southern Medical University, Guangzhou, Guangdong, 510000, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Jie Liu, Email [email protected] Yunhu Zhao, Email [email protected]
Introduction: Streptococcus agalactiae (Group B Streptococcus, GBS) is a Gram-positive opportunistic pathogen and a leading cause of invasive infections in neonates and pregnant women worldwide. Pathogenicity and transmission dynamics are shaped by capsular serotypes, clonal backgrounds, and virulence and resistance determinants, which vary over time and by region. Data from South China remain limited. This study characterized the serotype distribution, sequence types (STs), clonal complexes (CCs), antimicrobial resistance genes, and virulence factors of clinical GBS isolates from Guangzhou.
Methods: GBS-positive clinical isolates collected at Guangdong Provincial People’s Hospital (Guangzhou, China) from 2016 to 2022 were subjected to genomic epidemiology analyses. A total of 72 non-duplicate isolates were included. Multilocus sequence typing (MLST) and whole-genome sequencing (WGS) were used to determine capsular serotypes, sequence types (STs) and clonal complexes (CCs), and to identify antimicrobial resistance genes and virulence determinants, including pilus islands.
Results: Among 72 isolates, serotype III predominated (36.1%), followed by serotype V (25.0%). Isolates were assigned to 22 STs within 7 CCs, with ST19 being most common (15.3%). Resistance gene profiling showed tetM in 61.1% of isolates and ermB in 37.5%. Virulence factor analysis indicated universal presence of lmb, bca, and cpsJ; 97.2% carried the hemolysin gene hlyB. Pilus island genes PI-1 and PI-2a were detected in 73.6% of isolates.
Discussion: GBS circulating in Guangzhou shows a serotype distribution dominated by III and V and a diverse clonal structure with ST19 prominence. The high carriage of tetM and ermB suggests sustained selective pressure for tetracycline and macrolide resistance, while the near-ubiquitous virulence repertoire—including lmb, bca, cpsJ, and hlyB—and frequent PI-1/PI-2a may support colonization and invasiveness. These findings highlight the need for ongoing, region-specific genomic surveillance to track serotype/lineage shifts and resistance trends and to inform prophylaxis, empiric therapy, and future vaccine coverage in South China.
Keywords: Group B Streptococcus, serotype, multilocus sequence typing, resistance gene, virulence factor, whole-genome sequencing
Introduction
Streptococcus agalactiae (Group B Streptococcus, GBS) is a common Gram-positive bacterium that frequently colonizes the human reproductive and gastrointestinal tracts. The global colonization rate of GBS in pregnant women is approximately 18%, while in Chinese pregnant women, it is 10.5%.1,2 During pregnancy and childbirth, GBS can be transmitted vertically to neonates, leading to severe diseases such as sepsis, pneumonia, and meningitis. These infections are major contributors to the increasing morbidity and mortality rates among newborns worldwide.3 GBS colonization is typically transient and intermittent; however, under specific conditions, it can transform into pathogenic strains, significantly elevating the risk of infection.4 In the absence of effective preventive measures, the risk of neonatal GBS infection is markedly increased.5,6 Additionally, beyond neonates and pregnant women, elderly individuals and those with compromised immune systems are also susceptible to GBS infections.7 Recent studies have indicated that healthy adults may also experience GBS infections.8
Antibiotics are currently the primary means for preventing and treating GBS infections, particularly through intrapartum antimicrobial prophylaxis (IAP), which has been proven effective in reducing the incidence of neonatal sepsis, especially early-onset GBS infection (EOD) in GBS-colonized pregnant women.9 However, with the widespread use of antibiotics, the issue of antibiotic resistance in GBS has become increasingly prominent. Research has shown a decline in GBS sensitivity to certain antibiotics, especially macrolides represented by the ermB, ermTR, and mefA/E genes.10,11 A systematic review and meta-analysis of GBS infection rates and antibiotic resistance in Chinese infants revealed the highest resistance rates for tetracycline (median 98.0%, IQR 80.0–100.0%), followed by clindamycin (median 73.3%, IQR 62.6–78.7%), erythromycin (median 64.4%, IQR 56.6–75.0%), and ciprofloxacin (median 25.0%, IQR 9.1–35.2%).12 The increasing resistance poses a significant public health challenge, urgently necessitating the development of new prevention and treatment strategies to address resistant GBS strains.
The virulence factors of GBS are complex and diverse, among which capsular polysaccharides (CPS) are critical virulence determinants. Based on CPS variations, GBS isolates are classified into 10 serotypes: Ia, Ib, II-IX.13 This classification is of significant importance for the development of CPS-based vaccines. Currently, vaccines targeting different serotypes are under development, including monovalent vaccines, bivalent vaccines (serotypes II and III), trivalent vaccines (serotypes Ia, Ib, and III), and pentavalent vaccines (serotypes Ia, Ib, II, III, and V). With the inclusion of serotype IV, a hexavalent vaccine (I a, I b, II, III, IV, V) is also in the development stage.14,15 The hexavalent vaccine offers the broadest coverage; however, the presence of regionally rare serotypes may reduce vaccine efficacy, potentially leading to vaccine-driven serotype replacement. Therefore, continuous monitoring of serotype prevalence trends in different regions is crucial to promptly identify emerging serotypes and adjust vaccine strategies accordingly.16
In addition to CPS, GBS expresses various other virulence factors such as lmb, cpsM, and fbsA, which play important roles in bacterial adhesion, invasion, and immune evasion. Notably, pili (PI) are surface proteins involved in biofilm formation17 and facilitate bacterial invasion of host cells. These virulence factors contribute to the multifaceted roles GBS plays in invasive infections, making them potential targets for vaccine development.
Given the substantial heterogeneity of GBS serotypes and virulence factors, as well as their region- and time-dependent epidemiologic variation,1,18 systematic, region-specific data curation is warranted. Although global distributions of GBS serotypes have been reported,19,20 granular information for southern China—particularly clinical isolates from Guangdong Province—remains limited with respect to serotype composition, sequence types (STs), and antibiotic resistance spectra. In this study, we analyzed multiple GBS-positive clinical isolates collected at Guangdong Provincial People’s Hospital, describing serotype distributions, STs, resistance genes, and virulence-factor profiles, and documenting co-occurrence phenomena and potential distributional patterns across these features. These descriptive data provide a regional baseline for the genomic and resistance characteristics of GBS in Guangzhou, offer context for understanding local epidemiologic trends and clinical management, and generate hypotheses to inform subsequent mechanistic and association-focused investigations.
Materials and Methods
Source of Strains
This study collected 72 GBS-positive isolates obtained from the microbiology laboratory of Guangdong Provincial People’s Hospital between 2016 and 2022. Duplicate strains from the same patient were excluded.
Microbial Culture and Identification
Microbial culture and identification were conducted according to standard procedures in the hospital’s clinical laboratory. Specimens were inoculated onto blood agar plates and incubated at 35°C in a 5% CO2 environment for 24–48 hours. Selected colonies were purified by sub-culturing under the same conditions for an additional 24–48 hours. Isolates with β-hemolytic rings were identified using the VITEK MS mass spectrometer, and antimicrobial susceptibility testing was performed with the VITEK® 2 Compact system. Once identified as GBS, isolates were cultured on blood agar plates for subsequent whole-genome sequencing.
Whole-Genome Sequencing
Genomic DNA was extracted using the MagPure Bacterial DNA Kit (D6361-02), and libraries were prepared using the Hieff NGS® MaxUp II DNA Library Prep Kit for Illumina® (YEASEN, 12200ES08). PCR products were purified with magnetic beads (YEASEN, 12601ES56) and checked for quality using 2% agarose gel electrophoresis. Quantification was performed with a Qubit fluorometer, and paired-end sequencing was conducted on the Novaseq 6000 platform (Illumina, San Diego, CA).
Molecular Subtype Analysis
Resistance genes and virulence factors were identified using the CARD and VFDB databases. Serotype determination was performed based on the GBS bioinformatics typing tool from the Centers for Disease Control and Prevention (CDC).
Multilocus Sequence Typing (MLST)
MLST was conducted by sequencing seven housekeeping genes (adhP, pheS, atr, glnA, sdhA, glcK, and tkt).21 Based on allele variations, GBS isolates were classified into different STs and further grouped into clonal complexes (CCs). All alleles and STs of the GBS isolates were compared against data in the S. agalactiae MLST database (http://pubmlst.org/sagalactiae, accessed October 9, 2022) to confirm the corresponding STs and CCs. A minimum spanning tree was constructed using Phyloviz software by integrating CC and serotype data22 and visually refined in Adobe Illustrator 2020.
Results
Source of Specimens
This study collected 72 GBS clinical isolates between 2016 and 2022. Specimen types included genital secretions, midstream clean-catch urine, placenta, conjunctival secretions, wound exudate, venous blood, rectal swabs, synovial fluid, and cerebrospinal fluid. Genital secretions were the most common sample type, accounting for 50% (36/72) of isolates, followed by placenta samples at 18.1% (13/72), and midstream clean-catch urine at 9.7% (7/72). Details of sample types and their distribution are presented in Table 1. These GBS clinical isolates were primarily collected from obstetrics and gynecology departments and outpatient clinics, accounting for 90.2% (65/72) of all samples. Additionally, three samples were obtained from the departments of orthopedics and trauma, accounting for 4.2% (3/72). Detailed departmental source distributions are provided in Table 2.
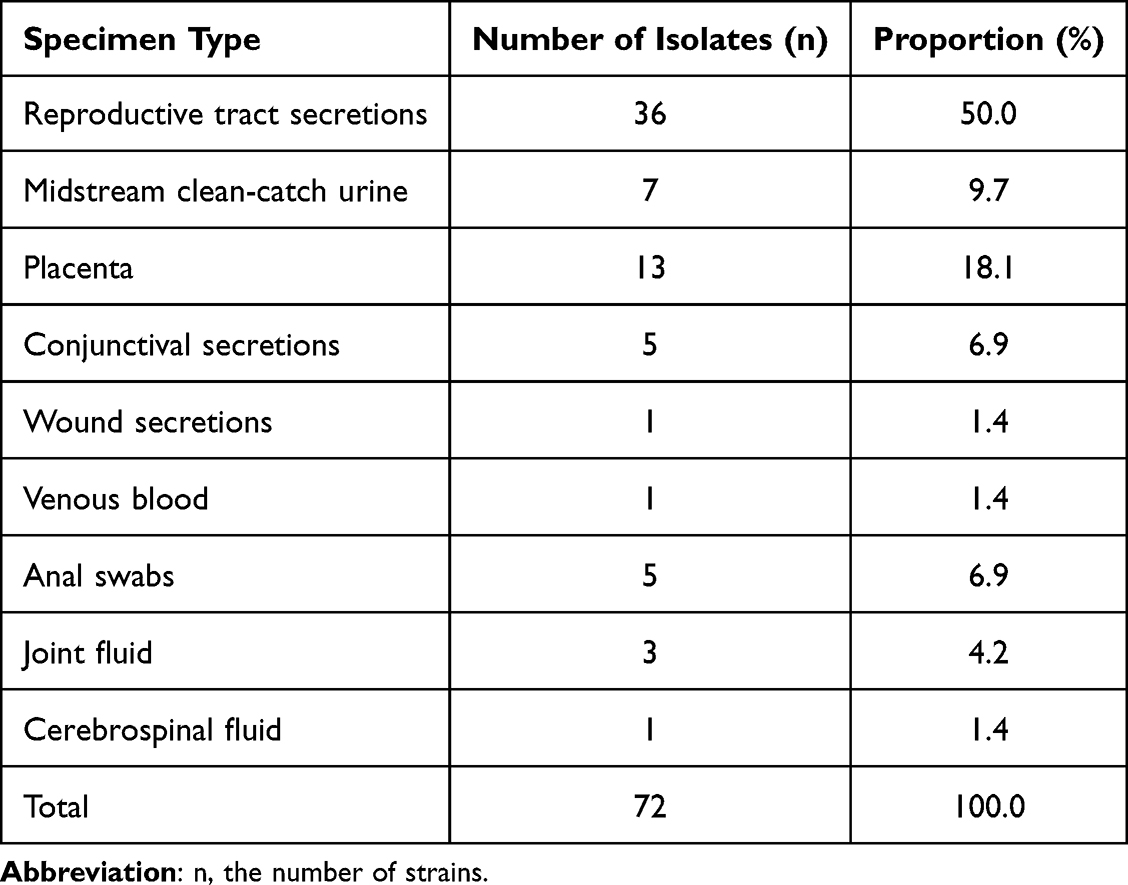 |
Table 1 Distribution of Specimen Types for 72 GBS Isolates |
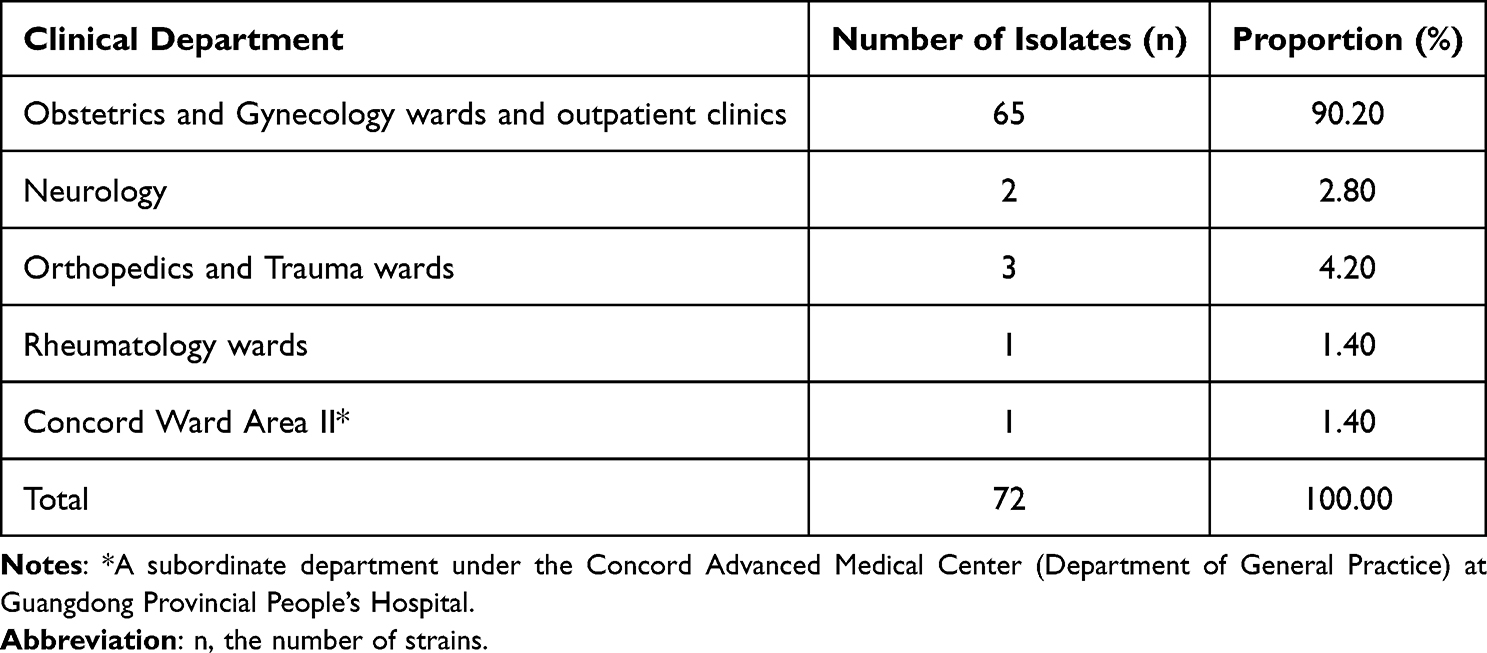 |
Table 2 Clinical Department Distribution of 72 GBS Isolates |
Serotype, Sequence Type, and Clonal Complex Analysis
Following an assessment of GBS isolates based on specimen origin, further analysis was conducted on serotype, ST, and clonal complex (CC) to comprehensively reveal their epidemiological patterns and genetic diversity. Results indicate six distinct serotypes among the isolates: Ia, Ib, II, III, V, and VI, with serotype III being the most prevalent (36.1%, 26/72), followed by V (25.0%, 18/72) and Ia (15.3%, 11/72). No isolates were identified for serotype IV. Detailed serotype distributions are shown in Table 3.
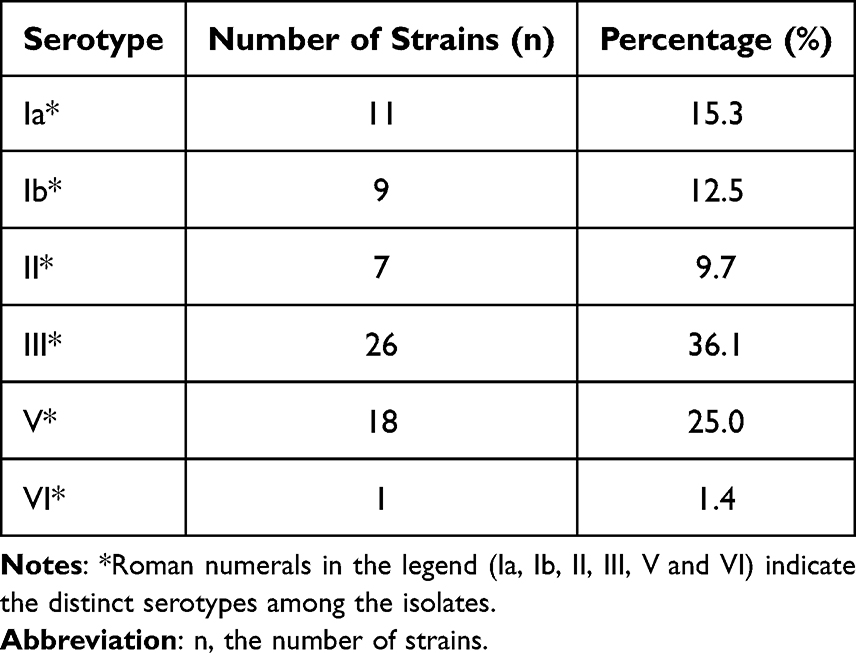 |
Table 3 Serotype Distribution of 72 GBS Isolates |
In the MLST typing analysis, 72 Group GBS isolates were assigned to 22 distinct sequence types (STs) and further grouped into seven clonal complexes (CCs); unknown clonal complexes are denoted as “Unk.” in subsequent figures. Five isolates could not be assigned to any known ST and were thus designated as new STs (NT). The most common STs were ST19 (15.3%, 11 isolates) and ST862 (9.2%, 7 isolates), with ST19 predominantly appearing in III and V serotypes and ST862 restricted to III serotypes. Additionally, ST17 and ST23 had six isolates each, playing significant roles in the detected STs, with ST17 and ST23 appearing in III and Ia serotypes, respectively. The distribution is detailed in Figures 1 and 2, and a minimum spanning tree is presented in Figure 3.
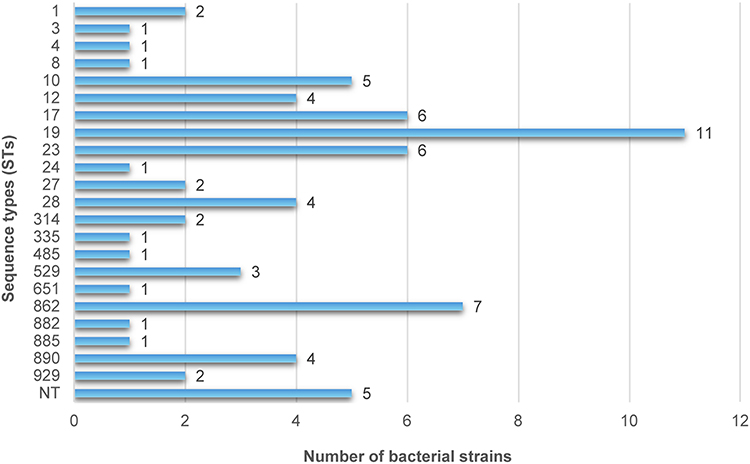 |
Figure 1 Distribution of GBS strains across sequence types (STs). The horizontal bar chart displays the number of strains on the X-axis and the corresponding STs on the Y-axis. ST19 (n = 11) and ST862 (n = 7) are the most prevalent sequence types, while five strains were non-typeable (NT). |
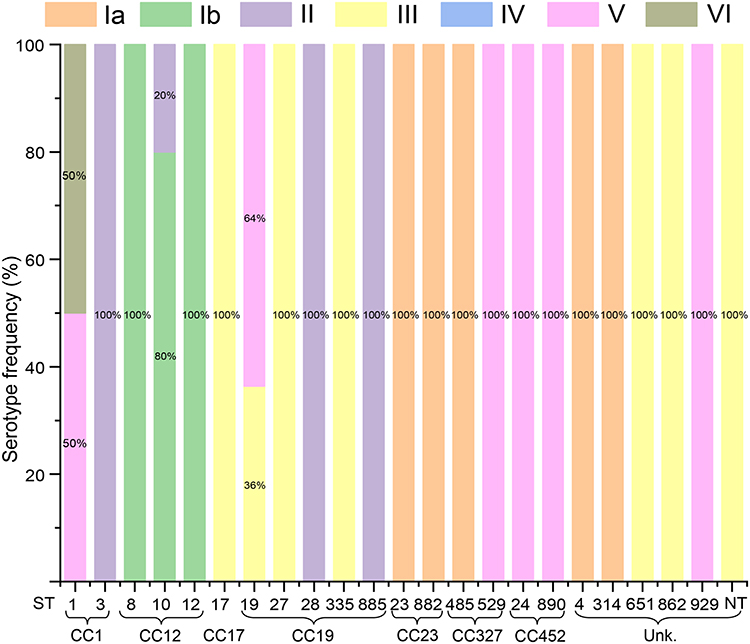 |
Figure 2 Distribution of serotypes among GBS sequence types (STs), multilocus sequence typing (MLST), and clonal complexes (CCs). The y-axis represents the serotype frequency (%), while the x-axis shows various STs and their associated CCs. Colored bars indicate different serotypes, including Ia, Ib, II, III, IV, V, and VI, with non-typeable (NT) strains also included. The percentage of each serotype within a given ST is labeled on the bars, illustrating the relationship between genetic profiles and serotype distributions. |
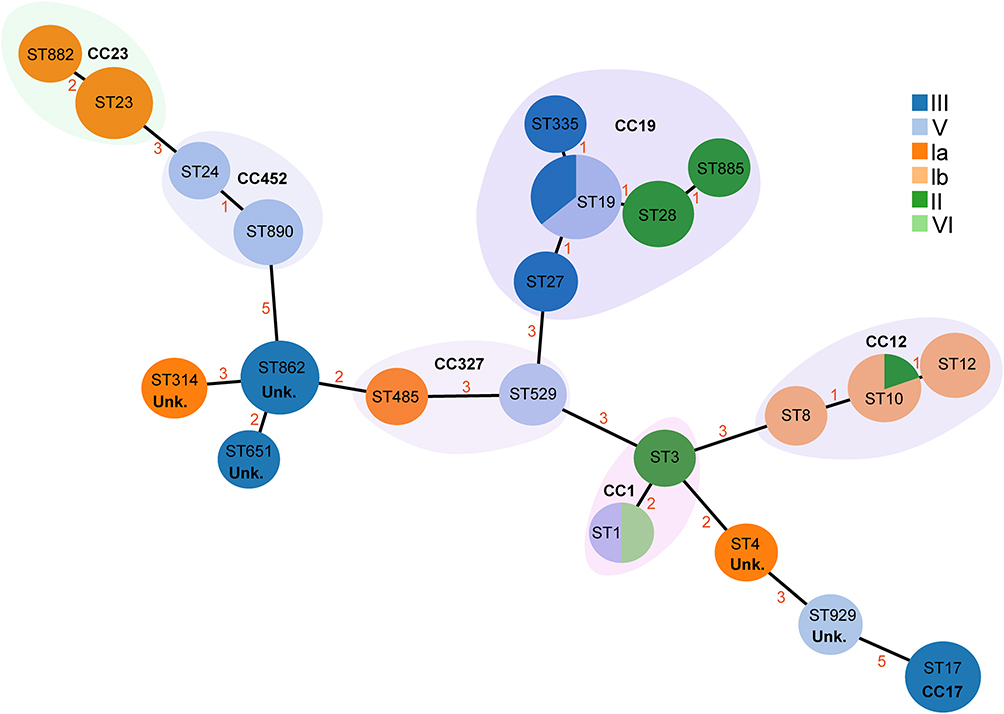 |
Figure 3 The minimum spanning tree of 67 GBS isolates illustrates the relationships between STs and CCs (classified by serotype). Circles represent ST types, with their sizes proportional to the number of isolates within each type. The ST with the most connections is considered the founder ST. Different colors indicate the serotypes of the GBS isolates, while shaded areas group STs belonging to the same CC. Roman numerals in the legend (Ia, Ib, II, III, V and VI) indicate the distinct serotypes among the isolates. Numbers (1–5) represent goeBURST distance scores, where higher scores indicate greater genetic distances between ST types. |
Antibiotic Susceptibility Results and Resistance Genes
The D-test identified four phenotypes, with the cMLSB phenotype being the most common (41 isolates), followed by iMLSB (10 isolates). M and L phenotypes were observed in 4 and 3 isolates, respectively. These findings suggest that cMLSB-type resistance is prevalent in GBS isolates, indicating an essential role for the macrolide-lincosamide-streptogramin B (MLSB) resistance mechanism. Detailed data are provided in Figure 4.
 |
Figure 4 The D-test was performed to detect inducible resistance to clindamycin, showing the distribution of different phenotypes. The cMLSB phenotype (constitutive MLS_B) exhibits resistance to both clindamycin and erythromycin. The M phenotype is characterized by sensitivity to clindamycin but resistance to erythromycin. The L phenotype demonstrates resistance to clindamycin and sensitivity or intermediate resistance to erythromycin. The iMLSB phenotype (inducible MLS_B) is defined by resistance to erythromycin and inducible resistance to clindamycin (D-test positive). The donut chart illustrates the quantity and proportion of each phenotype. |
The antibiotic susceptibility testing showed that resistance rates varied across agents. Resistance rates for tetracycline and erythromycin were 80.6% and 72.2%, respectively, demonstrating high resistance levels. Clindamycin showed a resistance rate of 61.1% with an intermediate rate of 4.2%. All isolates remained 100% susceptible to penicillin, linezolid, vancomycin, ampicillin (Figure 5). These data indicate substantial resistance to tetracycline, erythromycin, and clindamycin, while β-lactams, linezolid, and vancomycin remain highly effective.
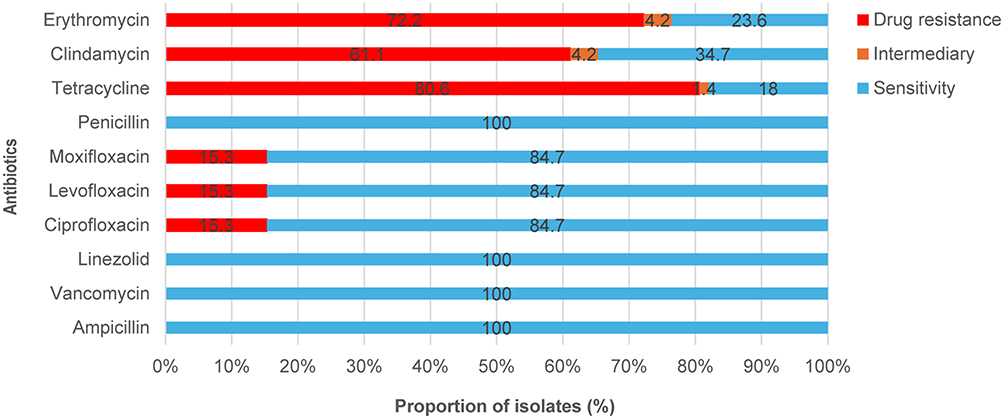 |
Figure 5 Antibiotic susceptibility results of the tested isolates. The horizontal bar chart shows, for each antimicrobial agent (Y-axis), the proportion of isolates that were resistant, intermediate, or susceptible (X-axis, %). Quinolone antibiotics include levofloxacin, moxifloxacin, and ciprofloxacin. The bar chart represents drug resistance rates (red), intermediary resistance rates (Orange), and susceptibility rates (blue). |
Antibiotic Susceptibility by Serotype
In the detection of macrolide resistance genes, results showed that 27 strains (37.5%) of GBS isolates carried the ermB gene, 9 strains (12.5%) carried the ermTR gene, and 15 strains (20.8%) carried the efflux determinant mefA/E gene. Additionally, 11 strains (15.3%) carried the clindamycin resistance gene lnuB. Among tetracycline resistance genes, 44 strains (61.1%) carried the tetM gene, while 18 strains (25.0%) carried the tetO gene. Across serotypes, these genes showed differing distributional patterns: ermB was more frequently observed in serotypes Ib and III, whereas tetM was more commonly noted in serotypes Ia and II. Cross-tabulations of resistance genes by serotype are provided in Table 4.
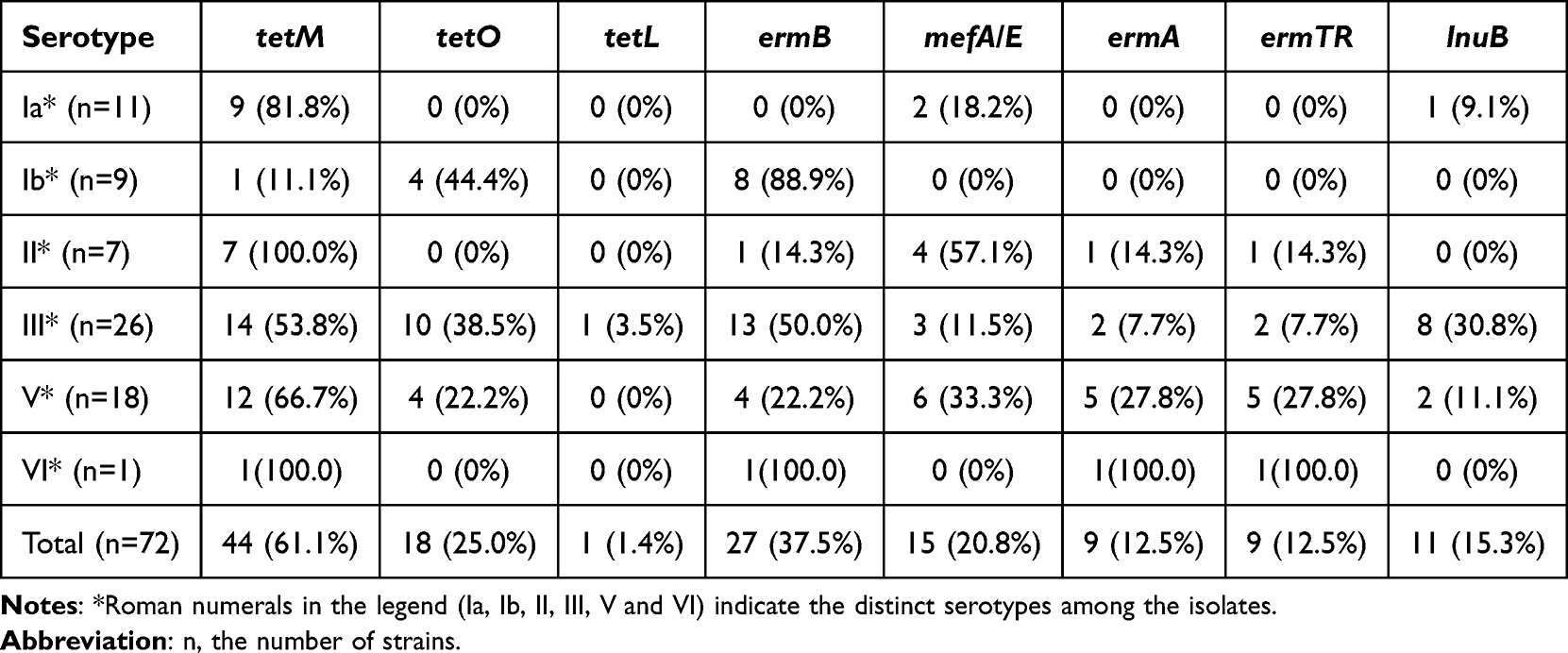 |
Table 4 Association of Antibiotic Resistance Genes with Serotypes |
Virulence Factors
Based on the cross-distribution of virulence factors by serotype (Figure 6 and Table 5), distinct serotype-specific virulence factor profiles were observed. For example, lmb, bca, and scpB were more commonly detected in serotype III, whereas cpsJ and cpsM were detected more frequently in serotypes Ia and III, respectively. Overall, serotype III exhibited a broader repertoire of virulence factors, encompassing nearly all factors assessed in this study, indicating that this serotype merits attention in GBS-related clinical contexts. In addition, specific virulence factors such as hylA and hylB showed differentiated distributions across serotypes.
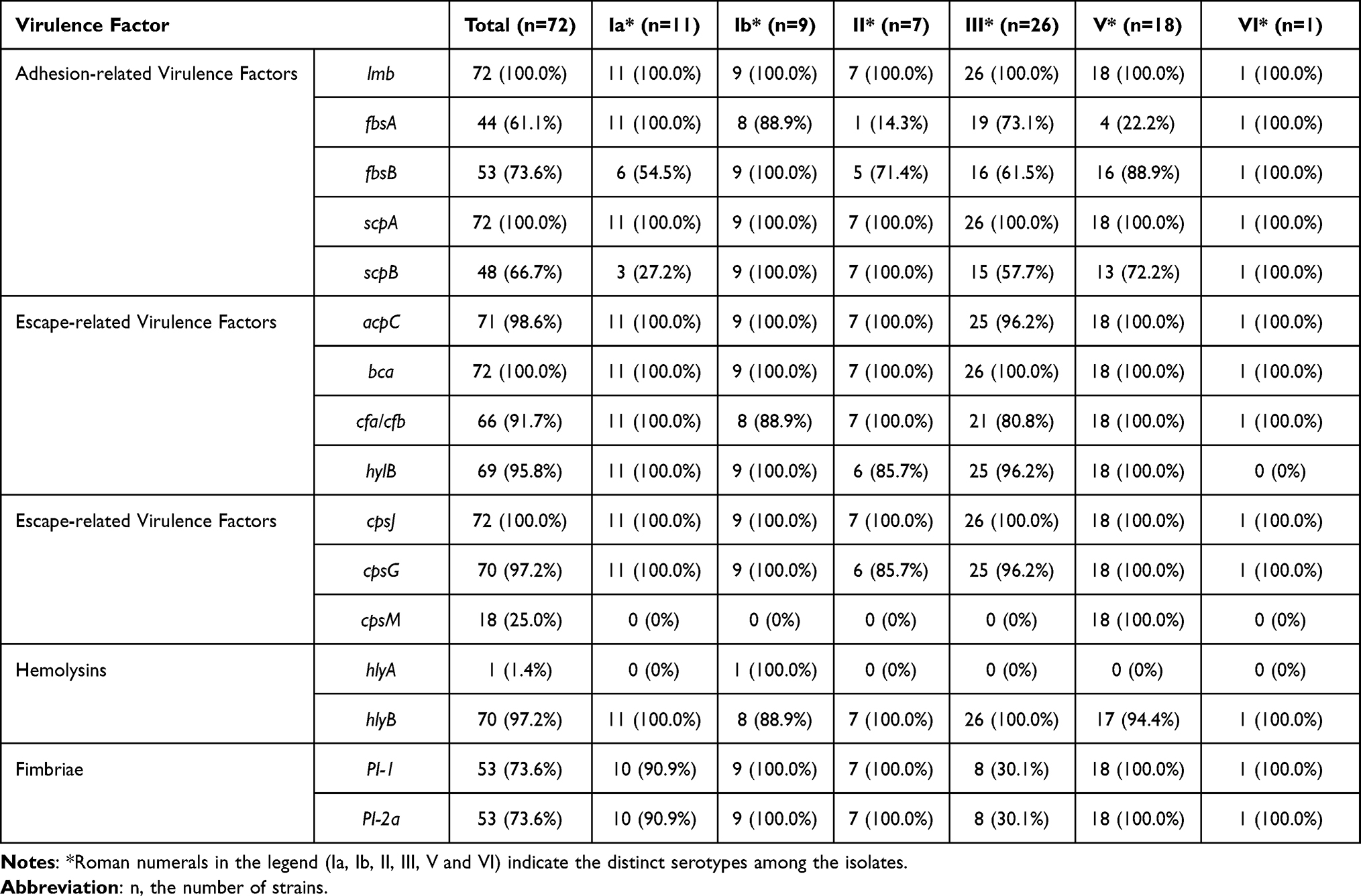 |
Table 5 Association of Virulence Factors with Serotypes |
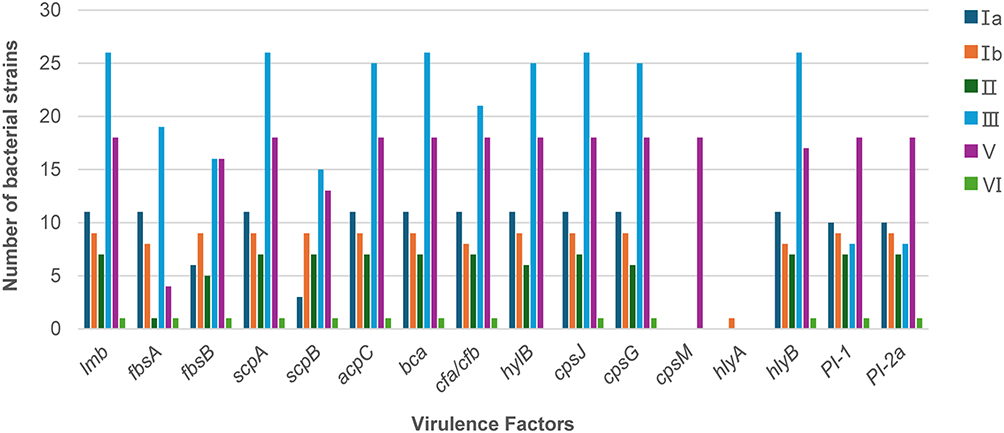 |
Figure 6 Association between virulence factors and serotypes in 72 GBS strains. The clustered bar chart displays the number of isolates (Y-axis) carrying individual virulence factors for each serotype (X-axis; Ia, Ib, II, III, V, and VI). Different bar colors represent different virulence factors, illustrating their distribution across serotypes. |
Discussion
Researching the virulence and antibiotic resistance of GBS provides insight into its pathogenicity and resistance mechanisms.23 A previous study reported a 9.61% GBS colonization rate among pregnant women in Guangzhou but did not include molecular characteristics such as serotypes, MLST typing, resistance genes, and virulence factors.24 In this report, we investigated the genetic characteristics of GBS-positive isolates collected over six years from a major tertiary hospital in Guangzhou. Using whole-genome sequencing, we analyzed the serotype distribution, sequence types, resistance genes, and virulence factors of GBS isolates. This study contributes to a better understanding of the pathogenicity and antibiotic resistance of GBS, aiding in clinical guidance for GBS infection treatment in the region.
The rich microbial flora in the vagina, with its self-purification capability, and bactericidal factors secreted by the endometrium work together to resist external bacterial infections. However, during pregnancy, women experience a unique immune modulation, entering an immunosuppressed state that protects the embryo by avoiding immune responses against it. This state reduces the mother’s immune response to external pathogens, making her more susceptible to bacterial infections.25,26 Accordingly, most of the 72 patient strains in this study were derived from pregnant women, with the main sample types being genital secretions and placenta. Both rectal swabs and conjunctival secretions originated from newborns, totaling 10 cases, accounting for 13.89% of all samples. Research indicates that GBS infections in neonates can lead to early-onset GBS disease.27 In immunocompromised infants, severe GBS infections can be life-threatening and may result in long-term sequelae such as intellectual disability, hydrocephalus, lung damage, and deafness even after recovery.28 This underscores the need for monitoring and treating GBS-positive pregnant women in Guangzhou to prevent vertical transmission and neonatal infections.
GBS has various virulence factors, among which CPS is a key determinant in invasive infections with important epidemiological implications. Anti-CPS antibodies provide protective immunity and are crucial virulence factors for anti-phagocytosis.29 CPS is the main component of the GBS cell wall capsule, with serotypes Ia, Ib, II, and III as the primary strain types, of which serotype III can cause severe infections.30,31 This study identified six capsular serotypes, with the most prevalent being serotype III (36.1%, 26/72), followed by serotype V (18 strains, 25.0%) and serotype Ia (11 strains, 15.3%). Studies suggest that serotypes Ia, V, and III are the main serotypes causing invasive infections in neonates,27 while serotypes Ia and V are also prevalent in GBS infections among non-pregnant adults.32 The prevalent serotypes vary slightly by region; for instance, Suzhou has predominant serotypes III, V, and Ib,33 while Taiwan primarily reports serotypes VI and III.34 The six serotypes detected in this study are consistent with those primarily associated with invasive infections in neonates and non-pregnant adults, highlighting the need for close attention to infections caused by serotypes Ia, Ib, III, and V.
The primary GBS vaccines under development include polysaccharide vaccines covering multiple serotypes and polysaccharide-protein conjugate vaccines.35 The hexavalent conjugate vaccine, covering serotypes Ia, Ib, II, III, VI, and V, and recombinant protein vaccines using alpha-like protein fusion are already in Phase II clinical trials. Regardless of the vaccine type, they aim to provide broad coverage across various strains. This study identified six serotypes, with only one strain of serotype. The hexavalent conjugate vaccine aligns with the current GBS strain prevalence and can cover most pathogenic strains.
This study’s GBS isolates were found to contain 22 different ST types, with 5 strains unconfirmed for ST type; these 22 types formed 7 different CCs, with the most common being ST19/V and ST862/III. The distribution of serotypes within CCs also showed clinical diversity in GBS characteristics.36–38 Jiang Haoqin and others observed that serotype Ib mainly clustered in ST10 and ST12, while serotype III predominantly clustered in ST19 and ST17. In our study, serotype III was primarily distributed in ST862 and ST17, differing slightly from literature reports, which may be due to differences in GBS serotype prevalence and geographic location. CC17 is the primary clonal complex associated with neonatal GBS meningitis, consistent with reports,39 with six CC17 strains in our study, five of which were from the obstetrics ward. CC17 can acquire new mobile genes through recombination, replacing the original PI-1 locus, resulting in PI-1 deletion and carrying resistance genes like ermB and tetO against erythromycin and clindamycin. This gene acquisition leads to multidrug resistance in the bacteria it harbors.40 Additionally, ST23 is recognized as a zoonotic sequence type, initially identified in grey seals.41 In this study, six GBS isolates were classified as ST23, all of which belonged to serotype Ia. Among the ST23 strains, resistance rates to erythromycin, clindamycin, and tetracycline were 16.7% (1/6), 16.7% (1/6), and 33.3% (2/6), respectively, with erythromycin and clindamycin resistance phenotypes not entirely consistent with the presence of resistance genes. Therefore, investigating the ST types of animal-derived GBS strains in Guangzhou is warranted.
The antibiotic resistance of GBS presents challenges for therapeutic options. Penicillin and ampicillin are the primary drugs for preventing or treating GBS infections, and their susceptibility has remained consistently high;42 no strains in this study exhibited resistance to first-line treatments. However, resistance to erythromycin and clindamycin remains high.43 The antimicrobial susceptibility results show that some GBS strains exhibit varying levels of resistance to commonly used antibiotics, possibly due to the widespread presence of this bacterium in clinical environments and the inappropriate use of antibiotics. Additionally, resistance rates varied among isolates. In this cohort, tetracycline resistance was 80.6% (58/72), erythromycin 70.8% (51/72), and clindamycin 61.1% (44/72); all tested isolates were susceptible to penicillin, linezolid, vancomycin, and ampicillin (Figure 4). One study reported a slow decline in GBS resistance rates to erythromycin, clindamycin, and levofloxacin between 2018 and 2020, followed by a slight increase in 2021. Between 2017 and 2021, high resistance rates to tetracycline (75.1%) and erythromycin (65.4%) were observed. Resistance rates to clindamycin and levofloxacin varied significantly, with rates in southern China reported as 55.8% and 24.0%, respectively.44 Compared with previous reports, the resistance rates observed in this study were generally lower. Possible explanations include differences in sample composition, time frame, or regional antibiotic-use practices; confirmation will require larger, multi-center datasets. Multidrug resistance was also observed in this cohort: 7.0% (5/72) of isolates were resistant to tetracyclines, clindamycin, erythromycin, and quinolones simultaneously, and 50.0% (36/72) were resistant to erythromycin, clindamycin, and tetracyclines.
Macrolides and related drugs are recommended as alternative therapies for patients allergic to β-lactam antibiotics in GBS infections,45 but extensive use of erythromycin and other macrolides could increase resistance risks.46 Four GBS isolates in this study exhibited a positive D-test, indicating erythromycin resistance with inducible clindamycin resistance. In clinical management of GBS infections, comprehensive antimicrobial susceptibility testing is essential to avoid the misuse of antibiotics due to empirical treatment. Tetracycline, as a broad-spectrum antibiotic, is less commonly used in GBS infections due to its effects on pregnant women and fetuses, yet it retains high resistance rates. Additionally, serotypes Ib and II showed erythromycin and clindamycin resistance rates of up to 80%, while serotype V demonstrated a tetracycline resistance rate of 90%, suggesting differences in resistance mechanisms may differ across serotypes, leading to varying degrees of resistance to specific antibiotics.
Among resistance genes, the detection rate of the tetracycline-resistance determinant gene tetM was 61.1% (44/72), significantly higher than that of the macrolide resistance gene ermB (27/72, 37.5%). The resistance profile observed in this study is consistent with recent reports from various regions of China: macrolide and clindamycin resistance remain high, and tetracycline resistance is widespread.47 Antibiotic resistance appears to be associated with certain serotypes:48 strains carrying ermB were mostly of types Ib and III and commonly belonged to CC12/17/19; mefA/E was more frequently found in type II; and tetM was more often detected in types Ia and II, especially among CC19 strains. The clindamycin resistance gene lnuB was primarily found in type III strains. Previous research indicates that ermB and mefA/E genes are associated with macrolide resistance.49 Continuous monitoring of antimicrobial susceptibility patterns is needed to guide empirical antibiotic therapy.
Virulence genes such as lmb, scpA, bca, and cpsJ were found in all 72 strains, corresponding to virulence factors for adhesion, invasion, and immune evasion. Enzymes also represent critical virulence factors in GBS, as they degrade host tissue structures, compromising tissue integrity. The hyaluronidase encoded by hylB cleaves hyaluronic acid during infection, promoting GBS dissemination. Mutants lacking hylB induce more pro-inflammatory cytokines in vivo and in vitro, resulting in bacterial elimination by the immune system.50 The hylB detection rate reached 95.8% (69/72), suggesting it facilitates GBS spread. Pili (PI) are also related to pathogenicity, enhancing adhesion to host cells and strengthening GBS invasiveness. Pili play a crucial role in GBS pathogenicity, making them a potential target for GBS vaccine development. The two pilus loci, PI-1 and PI-2, are encoded in different genomic regions; PI-2 has two variants, PI-2a and PI-2b, with PI-2a most common in human GBS and PI-2b more frequent in animal isolates.51 Studies show that PI-1 and PI-2a are more prevalent in colonizing strains, while PI-2b is more likely found in invasive strains. This suggests that GBS strains carrying the PI-2b gene have stronger pathogenicity, making them more likely to cause invasive diseases such as septicemia, meningitis, and endocarditis.52 In contrast, GBS strains with PI-1 and PI-2a genes are relatively less pathogenic, and PI-2a and PI-2b do not coexist in the same strain, reducing the likelihood of strains that are both highly pathogenic and colonizing. In this study, 53 strains (73.6%) carried pili, all with PI-1 and PI-2a. Among the six serotypes, only 8 out of 26 strains (30.1%) of serotype III carried pili. As the most common serotype, the low frequency of pili in type III suggests that other virulence factors contribute to GBS colonization.
The hemolysin hly can damage host cell membranes, leading to cell death and eliciting host inflammatory responses, further aggravating tissue damage.53 Only one strain of hlyA was detected in type Ib; 70 strains (70/72, 97.2%) harbored hlyB. All strains demonstrated equal ability in hemolysin-mediated tissue damage, but the unique presence of hlyA merits further investigation. Other major virulence factors include cell surface proteins that mediate adhesion and invasion, such as fibrin binding surface protein (fbsA), C5a peptidase (scpA, scpB), which are closely related to GBS adhesion to host cells, epithelial invasion, promotion of platelet aggregation, and anti-phagocytic activities.
This study utilized whole-genome sequencing to evaluate the serotypes, sequence types, resistance genes, and virulence factors of GBS isolates, revealing the prevalent serotypes and sequence types of GBS strains in Guangzhou and analyzing the main resistance genes carried by each serotype. A limitation of this study is its single-center design and relatively small sample size; accordingly, the findings are presented descriptively and no inferential statistics were performed. Nevertheless, these data provide a regional baseline for GBS epidemiology and resistance characteristics and help to orient subsequent investigations. Subsequent studies will expand to establish a multi-center GBS sample repository across major hospitals in Guangzhou, and conduct transcriptomic or proteomic analyses to explore how gene expression changes influence GBS pathogenicity and resistance. In-depth research on GBS is anticipated to clarify why GBS exhibits strong colonization ability but relatively weak pathogenicity, potentially aiding in clinical decision-making for GBS infections in Guangzhou.
Ethical Statement
This study was approved by the Ethics Committee of Guangdong Provincial People’s Hospital (Approval No. KY2023-682-01) and was conducted in accordance with the principles outlined in the Declaration of Helsinki. Informed consent was obtained from all participants prior to sample collection. All data were de-identified after collection and analyzed anonymously to ensure the protection of participants’ privacy and data confidentiality.
Funding
This work is funded by the National Natural Science Foundation of China (82302571); Basic and Applied Basic Research Foundation of Guangdong (2024A1515011037); Medical Science and Technology Research Foundation of Guangdong (B2025152).
Disclosure
The authors declare that they have no conflicts of interest in this work.
References
1. Raabe VN, Shane AL. Group B Streptococcus (Streptococcus agalactiae). Microbiol Spectr. 2019;7(2):10–13. doi:10.1128/microbiolspec.GPP3-0007-2018
2. Furfaro LL, Chang BJ, Payne MS. Perinatal Streptococcus agalactiae epidemiology and surveillance targets. Clin Microbiol Rev. 2018;31(4):e00049–18. doi:10.1128/CMR.00049-18
3. de Goffau MC, Lager S, Sovio U, et al. Human placenta has no microbiome but can contain potential pathogens. Nature. 2019;572(7769):329–334. doi:10.1038/s41586-019-1451-5
4. Gendrin C, Vornhagen J, Ngo L, et al. Mast cell degranulation by a hemolytic lipid toxin decreases GBS colonization and infection. Sci Adv. 2015;1(6):e1400225. doi:10.1126/sciadv.1400225
5. Furuta A, Brokaw A, Manuel G, et al. Bacterial and host determinants of Group B Streptococcal infection of the neonate and infant. Front Microbiol. 2022;13:820365. doi:10.3389/fmicb.2022.820365
6. Nanduri SA, Petit S, Smelser C, et al. Epidemiology of invasive early-onset and late-onset Group B Streptococcal disease in the United States, 2006 to 2015: multistate laboratory and population-based surveillance. JAMA Pediatr. 2019;173(3):224–233. doi:10.1001/jamapediatrics.2018.4826
7. Edwards MS, Baker CJ. Group B streptococcal infections in elderly adults. Clin Infect Dis. 2005;41(6):839–847. doi:10.1086/432804
8. van Kassel MN, van Haeringen KJ, Brouwer MC, Bijlsma MW, van de Beek D. Community-acquired group B streptococcal meningitis in adults. J Infect. 2020;80(3):255–260. doi:10.1016/j.jinf.2019.12.002
9. Vornhagen J, Adams Waldorf KM, Rajagopal L. Perinatal Group B Streptococcal infections: virulence factors, immunity, and prevention strategies. Trends Microbiol. 2017;25(11):919–931. doi:10.1016/j.tim.2017.05.013
10. Hayes K, O’Halloran F, Cotter L. A review of antibiotic resistance in Group B Streptococcus: the story so far. Crit Rev Microbiol. 2020;46(3):253–269. doi:10.1080/1040841X.2020.1758626
11. Gizachew M, Tiruneh M, Moges F, Tessema B. Streptococcus agalactiae maternal colonization, antibiotic resistance and serotype profiles in Africa: a meta-analysis. Ann Clin Microbiol Antimicrob. 2019;18(1):14. doi:10.1186/s12941-019-0313-1
12. Ding Y, Wang Y, Hsia Y, Russell N, Heath PT. Systematic review and meta-analyses of incidence for Group B Streptococcus disease in infants and antimicrobial resistance, China. Emerg Infect Dis. 2020;26(11):2651–2659. doi:10.3201/eid2611.181414
13. Lo CW, Liu HC, Lee CC, et al. Serotype distribution and clinical correlation of Streptococcus agalactiae causing invasive disease in infants and children in Taiwan. J Microbiol Immunol Infect. 2019;52(4):578–584. doi:10.1016/j.jmii.2017.09.002
14. Pena JMS, Lannes-Costa PS, Nagao PE. Vaccines for Streptococcus agalactiae: current status and future perspectives. Front Immunol. 2024;15:1430901. doi:10.3389/fimmu.2024.1430901
15. Thorn N, Karampatsas K, Le Doare K, Heath PT. Progress towards a Group B streptococcal vaccine - where are we now? Vaccine. 2025;62:127575. doi:10.1016/j.vaccine.2025.127575
16. Madrid L, Seale AC, Kohli-Lynch M, et al. Infant Group B Streptococcal disease incidence and serotypes worldwide: systematic review and meta-analyses. Clin Infect Dis. 2017;65(suppl_2):S160–s72. doi:10.1093/cid/cix656
17. Konto-Ghiorghi Y, Mairey E, Mallet A, et al. Dual role for pilus in adherence to epithelial cells and biofilm formation in Streptococcus agalactiae. PLoS Pathog. 2009;5(5):e1000422. doi:10.1371/journal.ppat.1000422
18. Russell NJ, Seale AC, O’Driscoll M, et al. Maternal colonization with Group B Streptococcus and serotype distribution worldwide: systematic review and meta-analyses. Clin Infect Dis. 2017;65(suppl_2):S100–s11. doi:10.1093/cid/cix658
19. Hall J, Adams NH, Bartlett L, et al. Maternal disease with Group B Streptococcus and serotype distribution worldwide: systematic review and meta-analyses. Clin Infect Dis. 2017;65(suppl_2):S112–s24. doi:10.1093/cid/cix660
20. Ren X, Liu X, Che Y, et al. Serotype distribution, virulence, and antibiotic resistance genomic characterization of Group B Streptococcus - China, 1998–2024. China CDC Wkly. 2025;7(13):413–421. doi:10.46234/ccdcw2025.069
21. Jones N, Bohnsack JF, Takahashi S, et al. Multilocus sequence typing system for group B streptococcus. J Clin Microbiol. 2003;41(6):2530–2536. doi:10.1128/JCM.41.6.2530-2536.2003
22. Ali MM, Woldeamanuel Y, Asrat D, et al. Features of Streptococcus agalactiae strains recovered from pregnant women and newborns attending different hospitals in Ethiopia. BMC Infect Dis. 2020;20(1):848. doi:10.1186/s12879-020-05581-8
23. Alzayer M, Alkhulaifi MM, Alyami A, et al. Genomic insights into the diversity, virulence, and antimicrobial resistance of group B Streptococcus clinical isolates from Saudi Arabia. Front Cell Infect Microbiol. 2024;14:1377993. doi:10.3389/fcimb.2024.1377993
24. Liu Y, Liu W, Zhuang G, Liu W, Qiu C. Colonisation of Group B Streptococcus and its effects on pregnancy outcomes in pregnant women in Guangzhou, China: a retrospective study. BMJ Open. 2023;13(11):e078759. doi:10.1136/bmjopen-2023-078759
25. Lu J, Moore RE, Spicer SK, et al. Streptococcus agalactiae npx is required for survival in human placental macrophages and full virulence in a model of ascending vaginal infection during pregnancy. mBio. 2022;13(6):e0287022. doi:10.1128/mbio.02870-22
26. Yockey LJ, Iwasaki A. Interferons and proinflammatory cytokines in pregnancy and fetal development. Immunity. 2018;49(3):397–412. doi:10.1016/j.immuni.2018.07.017
27. Joubrel C, Tazi A, Six A, et al. Group B streptococcus neonatal invasive infections, France 2007–2012. Clin Microbiol Infect. 2015;21(10):910–916. doi:10.1016/j.cmi.2015.05.039
28. Verani JR, McGee L, Schrag SJ. Prevention of perinatal group B streptococcal disease--revised guidelines from CDC, 2010. MMWR Recomm Rep. 2010;59(Rr–10):1–36.
29. Fabbrini M, Rigat F, Tuscano G, et al. Functional activity of maternal and cord antibodies elicited by an investigational group B Streptococcus trivalent glycoconjugate vaccine in pregnant women. J Infect. 2018;76(5):449–456. doi:10.1016/j.jinf.2018.01.006
30. Creti R, Imperi M, Berardi A, et al. Invasive Group B Streptococcal disease in neonates and infants, Italy, years 2015–2019. Microorganisms. 2021;9(12):2579. doi:10.3390/microorganisms9122579
31. Wu B, Su J, Li L, et al. Phenotypic and genetic differences among group B Streptococcus recovered from neonates and pregnant women in Shenzhen, China: 8-year study. BMC Microbiol. 2019;19(1):185. doi:10.1186/s12866-019-1551-2
32. Martins ER, Do Ó D N, Marques Costa AL, Melo-Cristino J, Ramirez M. Characteristics of Streptococcus agalactiae colonizing nonpregnant adults support the opportunistic nature of invasive infections. Microbiol Spectr. 2022;10(3):e0108222. doi:10.1128/spectrum.01082-22
33. Guo Y, Deng X, Liang Y, Zhang L, Zhao GP, Zhou Y. The draft genomes and investigation of serotype distribution, antimicrobial resistance of group B Streptococcus strains isolated from urine in Suzhou, China. Ann Clin Microbiol Antimicrob. 2018;17(1):28. doi:10.1186/s12941-018-0280-y
34. Tsai MH, Hsu JF, Lai MY, et al. Molecular characteristics and antimicrobial resistance of Group B Streptococcus strains causing invasive disease in neonates and adults. Front Microbiol. 2019;10:264. doi:10.3389/fmicb.2019.00264
35. Gent V, Lu YJ, Lukhele S, et al. Surface protein distribution in Group B Streptococcus isolates from South Africa and identifying vaccine targets through in silico analysis. Sci Rep. 2024;14(1):22665. doi:10.1038/s41598-024-73175-4
36. Bisharat N, Crook DW, Leigh J, et al. Hyperinvasive neonatal group B streptococcus has arisen from a bovine ancestor. J Clin Microbiol. 2004;42(5):2161–2167. doi:10.1128/JCM.42.5.2161-2167.2004
37. Manning SD, Springman AC, Lehotzky E, Lewis MA, Whittam TS, Davies HD. Multilocus sequence types associated with neonatal group B streptococcal sepsis and meningitis in Canada. J Clin Microbiol. 2009;47(4):1143–1148. doi:10.1128/JCM.01424-08
38. van Kassel MN, de Boer G, Teeri SAF, et al. Molecular epidemiology and mortality of group B streptococcal meningitis and infant sepsis in the Netherlands: a 30-year nationwide surveillance study. Lancet Microbe. 2021;2(1):e32–e40. doi:10.1016/S2666-5247(20)30192-0
39. Shabayek S, Spellerberg B. Group B Streptococcal colonization, molecular characteristics, and epidemiology. Front Microbiol. 2018;9:437. doi:10.3389/fmicb.2018.00437
40. Campisi E, Rosini R, Ji W, et al. Genomic analysis reveals multi-drug resistance clusters in Group B Streptococcus CC17 hypervirulent isolates causing neonatal invasive disease in Southern Mainland China. Front Microbiol. 2016;7:1265. doi:10.3389/fmicb.2016.01265
41. Delannoy CM, Crumlish M, Fontaine MC, et al. Human Streptococcus agalactiae strains in aquatic mammals and fish. BMC Microbiol. 2013;13:41. doi:10.1186/1471-2180-13-41
42. Tan J, Wang Y, Gong X, et al. Antibiotic resistance in neonates in China 2012–2019: a multicenter study. J Microbiol Immunol Infect. 2022;55(3):454–462. doi:10.1016/j.jmii.2021.05.004
43. Ma A, Thompson LA, Corsiatto T, Hurteau D, Tyrrell GJ. Epidemiological characterization of Group B Streptococcus Infections in Alberta, Canada: an update from 2014 to 2020. Microbiol Spectr. 2021;9(3):e0128321. doi:10.1128/Spectrum.01283-21
44. Wang J, Zhang Y, Lin M, et al. Maternal colonization with group B Streptococcus and antibiotic resistance in China: systematic review and meta-analyses. Ann Clin Microbiol Antimicrob. 2023;22(1):5. doi:10.1186/s12941-023-00553-7
45. Barros RR. Antimicrobial resistance among beta-hemolytic Streptococcus in Brazil: an overview. Antibiotics. 2021;10(8):973. doi:10.3390/antibiotics10080973
46. Dinos GP. The macrolide antibiotic renaissance. Br J Pharmacol. 2017;174(18):2967–2983. doi:10.1111/bph.13936
47. Mai W, Wang H, Meng Q, et al. Molecular epidemiology and antibiotic resistance of group B Streptococcus in pregnant women and neonates from Haikou, China: implications for vaccine development and antimicrobial stewardship. Front Cell Infect Microbiol. 2025;15:1655649. doi:10.3389/fcimb.2025.1655649
48. Lopes E, Fernandes T, Machado MP, et al. Increasing macrolide resistance among Streptococcus agalactiae causing invasive disease in non-pregnant adults was driven by a single capsular-transformed lineage, Portugal, 2009 to 2015. Euro Surveill. 2018;23(21):1700473. doi:10.2807/1560-7917.ES.2018.23.21.1700473
49. Gergova R, Boyanov V, Muhtarova A, Alexandrova A. A review of the impact of Streptococcal infections and antimicrobial resistance on human health. Antibiotics. 2024;13(4):360. doi:10.3390/antibiotics13040360
50. Coleman M, Armistead B, Orvis A, et al. Hyaluronidase impairs neutrophil function and promotes group B streptococcus invasion and preterm labor in nonhuman primates. mBio. 2021;12(1):e03115–20. doi:10.1128/mBio.03115-20
51. Springman AC, Lacher DW, Waymire EA, et al. Pilus distribution among lineages of group b streptococcus: an evolutionary and clinical perspective. BMC Microbiol. 2014;14:159. doi:10.1186/1471-2180-14-159
52. Schindler Y, Rahav G, Nissan I, et al. Group B streptococcus virulence factors associated with different clinical syndromes: asymptomatic carriage in pregnant women and early-onset disease in the newborn. Front Microbiol. 2023;14:1093288. doi:10.3389/fmicb.2023.1093288
53. Liu Y, Liu J. Group B Streptococcus: virulence factors and pathogenic mechanism. Microorganisms. 2022;10(12):2483. doi:10.3390/microorganisms10122483
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Late-Onset Sepsis in a Premature Infant Mediated by Breast Milk: Mother-to-Infant Transmission of Group B Streptococcus Detected by Whole-Genome Sequencing
Li A, Fang M, Hao D, Wu Q, Qian Y, Xu H, Zhu B
Infection and Drug Resistance 2022, 15:5345-5352
Published Date: 9 September 2022
Molecular and Clinical Characteristics of Carbapenem-Resistant Klebsiella pneumoniae Isolates at a Tertiary Hospital in Wuhan, China
Hu F, Lin ML, Mou JL, Feng JH, Huang K, Lao YJ, Cheng J, Lin J
Infection and Drug Resistance 2023, 16:65-76
Published Date: 5 January 2023
Characterization of Group B Streptococcus Recovered from Pregnant Women and Newborns Attending in a Hospital in Beijing, China
Li X, Gao W, Jia Z, Yao K, Yang J, Tong J, Wang Y
Infection and Drug Resistance 2023, 16:2549-2559
Published Date: 27 April 2023
Genomic Characterization of Antibiotic-Resistant Enterococcus Spp. Strains in Clinical Settings of Yichang, China
Yu H, Xiong P, Zhang H, Lu Z, Chen X, Yan R, Wang L, Yu L, Zhang Q, Zou L, Wang L
Infection and Drug Resistance 2026, 19:573082
Published Date: 14 April 2026
Molecular Epidemiology and Antimicrobial Resistance of Group B Streptococcus in Hainan, China: Genomic Insights from Perinatal and Adult Clinical Isolates
Zhao HM, Song XD, Li J, Miao BB, Zhang JY, Gong XY, He Y, Wang YJ, Fu YS, Wu H
Infection and Drug Resistance 2026, 19:617393
Published Date: 15 July 2026