Back to Journals » Breast Cancer: Targets and Therapy » Volume 18
Salidroside-Based Nanomedicines for Triple-Negative Breast Cancer: From Molecular Mechanisms to Clinical Translation
Authors Ma X ![]() , Dang S, Wang H, Xia L, Ma P, Qi Y
, Dang S, Wang H, Xia L, Ma P, Qi Y
Received 26 February 2026
Accepted for publication 20 May 2026
Published 24 June 2026 Volume 2026:18 605051
DOI https://doi.org/10.2147/BCTT.S605051
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Professor Harikrishna Nakshatri
Xiaoying Ma,1– 3 Shengyan Dang,1,3,4 Hui Wang,3,4 Lei Xia,2 Ping Ma,2 Yujuan Qi1,3,4
1School of Medicine, Qinghai University, Xining, Qinghai, 810001, People’s Republic of China; 2Department of Breast and Thyroid, Qinghai Provincial People’s Hospital, Xining, Qinghai, 810001, People’s Republic of China; 3Qinghai Clinical Research Center for High Altitude Diseases, Xining, Qinghai, 810001, People’s Republic of China; 4Department of Medical Oncology, Qinghai Provincial People’s Hospital, Xining, Qinghai, 810001, People’s Republic of China
Correspondence: Yujuan Qi, Email [email protected]
Abstract: Triple-negative breast cancer (TNBC) presents a substantial therapeutic challenge due to its aggressive biological characteristics and the absence of actionable molecular targets. Salidroside, the principal bioactive compound of Rhodiola rosea, demonstrates significant anticancer potential; however, its clinical application is severely constrained by poor bioavailability (less than 12%) and a short half-life (2.3 hours). This review contributes to the field in three key ways. Firstly, it systematically elucidates the molecular mechanisms of salidroside in TNBC, emphasizing its distinctive dual role in redox homeostasis: inducing ferroptosis in cancer cells while safeguarding normal tissues. Secondly, it critically assesses advanced nanomedicine strategies such as PLGA-PEG nanoparticles, lipid-polymer hybrid nanoparticles, and biomimetic RBC membrane-camouflaged carriers-designed to address pharmacokinetic limitations. Thirdly, and most importantly, it proposes a biomarker-driven framework, centered on BRCA1 methylation and HIF-1α, along with a three-phase translational roadmap to advance salidroside into a precision nanomedicine for TNBC. The diagram illustrates the process of using salidroside for treating triple-negative breast cancer. Initially, salidroside targets the cancer, but faces clinical limitations such as low bioavailability, short half-life and no standardized formulation. A solution is proposed using salidroside nanoparticles to target tumors with precision. This involves a multi-molecular pathway with HIF-1α and macrophages to minimize toxicity while augmenting efficacy. The ultimate goal is reaching clinical application.A diagram illustrating the clinical limitations of free salidroside and the proposed nanoparticle-based strategy for addressing triple-negative breast cancer.
Keywords: salidroside, triple-negative breast cancer, nanomedicines, tumor immune microenvironment, ferroptosis
Introduction
Triple-negative breast cancer (TNBC) approximately accounts for 15–20% of all breast cancer cases and presents a formidable therapeutic challenge due to its aggressive clinical progression and lack of targetable estrogen receptor (ER), progesterone receptor (PR), human epidermal growth factor receptor 2 (HER2).1 Triple-negative breast cancer (TNBC) is not a singular entity; rather, it represents a highly heterogeneous group of diseases characterized by several distinct molecular subtypes, including basal-like 1 (BL1), basal-like 2 (BL2), immunomodulatory (IM), mesenchymal (M), mesenchymal stem-like (MSL), and luminal androgen receptor (LAR) subtypes. These subtypes demonstrate varying sensitivities to different therapeutic agents and possess unique immune microenvironment profiles. The standard therapeutic regimen for triple-negative breast cancer (TNBC) typically involves the administration of anthracyclines (eg., epirubicin), taxanes (eg., paclitaxel), and platinum-based agents, often followed by adjuvant capecitabine. Recent developments have highlighted the potential of antibody-drug conjugates, such as sacituzumab govitecan, especially when combined with immune checkpoint inhibitors like pembrolizumab, as promising strategies for the treatment of advanced TNBC. These findings are supported by data from the ASCENT-04 trials. Conventional endocrine and HER2-targeted therapies have proven ineffective in treating triple-negative breast cancer (TNBC). Furthermore, immune checkpoint inhibitors demonstrate limited efficacy, with clinical response rates below 20%.2,3 Within this evolving therapeutic landscape, salidroside emerges as a compound of interest due to its potential synergistic roles: it may mitigate epirubicin-induced cardiotoxicity, counteract paclitaxel resistance, and enhance the efficacy of anti-PD-1 immunotherapy. Consequently, salidroside is positioned as a potential adjunct to existing treatment regimens rather than as a replacement.
Salidroside (C14H20O7), the primary bioactive component of Rhodiola rosea L. (Figure 1), has emerged as a promising polypharmacological candidate. In addition to its extensively documented antioxidant and anti-inflammatory properties,4–12 it demonstrates multifaceted antitumor efficacy against breast cancer by inhibiting cellular proliferation, reprogramming metabolism pathways, and modulating the immune microenvironment. Elucidating how salidroside’s multi-targeted mechanisms-specifically its modulation of the Wnt/β-catenin and PI3K-AKT-mTOR pathways, as well as its induction of ferroptosis-may differentially impact specific TNBC subtypes is essential for the development of biomarker-driven therapeutic strategies. Nevertheless, similar to numerous natural compounds exhibiting pleiotropic anticancer properties, salidroside encounters a prevalent translational challenge: its high in vitro potency does not effectively translate into clinical efficacy, primarily due to suboptimal pharmacokinetic characteristics and the absence of standardized formulations. This gap presents an opportunity that nanomedicine is particularly well-suited to address.
 |
Figure 1 Chemical structure of salidroside (C14H20O7). The two-dimensional and three-dimensional chemical structures of salidroside are depicted, illustrating a benzene ring connected to a glucose moiety through a glycosidic bond. This configuration includes hydroxyl groups at critical positions, where the aromatic hydroxyl group is associated with redox regulation and antioxidant activity, while the glucosyl moiety hydroxyl groups contribute to water solubility and target binding affinity. Additionally, the O-glycosidic bond plays a role in the compound’s stability, metabolism, and bioavailability. This information has been adapted from PubChem (CID: 159278) with permission (https://pubchem.ncbi.nlm.nih.gov/). |
Despite strong preclinical evidence supporting its efficacy, the translational potential of salidroside is significantly constrained by its dependence on non-physiological experimental conditions, the use of supraphysiological concentrations in vitro, and limited clinical bioavailability, which is less than 12%.2 Nanomedicine presents a strategic approach to overcoming these limitations. The concept of “nanotheranostics” pertains to nanoscale platforms that amalgamate therapeutic and diagnostic (imaging) capabilities, facilitating the real-time monitoring of drug delivery and treatment efficacy. Advanced nanodelivery systems have the potential to enhance the pharmacokinetic properties of salidroside, facilitate targeted delivery, and enable real-time therapeutic monitoring, thereby establishing a foundation for precision therapy in TNBC.
To ensure a comprehensive synthesis, this review systematically incorporates 87 relevant studies. The foundational mechanistic literature encompasses the period from 2009 to 2023, while significant advancements in nanodelivery strategies and clinical translational frameworks from 2024 to 2025 have been included to maintain contemporary relevance. Relevant studies were identified through systematic searches of the PubMed, Web of Science, and Scopus databases (up to March 2025), employing combinations of keywords such as “salidroside,” “Rhodiola rosea,” “breast cancer,” “TNBC,” “nanomedicine,” “nanoparticle,” “ferroptosis,” and “tumor microenvironment.” The inclusion criteria were restricted to peer-reviewed, English-language original articles and reviews.
Building on foundational evidence, this review initially elucidates the complex molecular mechanisms and subtype-specific pharmacological effects of salidroside in breast cancer. Subsequently, it identifies critical translational barriers and proposes the use of integrated nanomedicine approaches-merging advanced nanodelivery systems with real-time imaging-as a pivotal strategy to address pharmacokinetic limitations and enhance personalized therapy for triple-negative breast cancer (TNBC). Overall, our analysis provides a comprehensive roadmap for the development of salidroside as a precision oncology agent, offering novel insights to overcome existing therapeutic challenges in TNBC.
Molecular Mechanisms of Salidroside in Breast Cancer
Salidroside demonstrates a wide range of antitumor effects in breast cancer by influencing a complex network of molecular pathways that regulate uncontrolled proliferation, tumor metastasis, metabolic reprogramming, epigenetic dysregulation, immune evasion, and regulated cell death, including ferroptosis and apoptosis (Figure 2). These mechanisms display distinct specificity across different subtypes, offering a robust basis for pharmacological development. However, core translational limitations are primarily associated with non-physiological experimental conditions and insufficient disease models, which are central to the ongoing optimization of nanotheranostic approaches.
 |
Figure 2 Integrated molecular mechanisms of salidroside in breast cancer. Schematic diagram illustrating salidroside’s core molecular targets and pathways in breast cancer, including proliferation inhibition (PI3K-AKT-HIF-1α axis), ferroptosis induction (SCD1-NCOA4 pathway), angiogenesis suppression (HIF-1α/VEGF/STAT3 axis), EMT inhibition (Twist1/Zeb1-E-cadherin pathway), immune microenvironment remodeling (Treg/DC regulation), and epigenetic modulation (DNMT3B-BRCA1 axis). Note: Solid arrows are used to denote promotional regulatory effects, while T-shaped arrows signify inhibition. Blue dashed arrows illustrate indirect paracrine modulation of the tumor microenvironment, encompassing macrophages, dendritic cells, and regulatory T cells (Tregs). Orange dashed arrows indicate synergistic therapeutic effects. Notably, all solid arrows represent the direct regulatory impact of salidroside on intracellular signaling pathways within breast cancer cells. |
Regulation of Cell Proliferation and Apoptosis
Salidroside influences sustained proliferation and apoptosis resistance through subtype-specific mechanisms in breast cancer. In TNBC cell line MDA-MB-231, salidroside induced G1 phase cell cycle arrest by upregulating cyclin-dependent kinase inhibitors p21 and p27, while inhibiting the CDK4-cyclin D1 signaling pathway, resulted in 65% arrest in the G1 phase.13 Conversely, in luminal A breast cancer cell line MCF-7, it impeded proliferation by targeting the phosphatidylinositol 3-kinase (PI3K)-protein kinase B (AKT)-mammalian target of rapamycin (mTOR) signaling pathway, leading to a 58% reduction in malignant cell growth.14 The induction of apoptosis was consistently facilitated by caspase activation: salidroside activated caspase-3 in MCF-7 xenografts and caspase-9 in MDA-MB-231 cells by modulating Bcl-2 family proteins, thereby increased the apoptotic index by up to 3.1-fold.13,14
It is noteworthy that the effective in vitro concentrations (50–200 μM) exceeded clinically achievable plasma levels (1–5 μM) by a factor of 10–4010,16. Furthermore, the majority of studies are constrained by a limited selection of cell lines, such as MDA-MB-231, MCF-7, and 4T1, which do not adequately represent the heterogeneity of breast cancer.14,15 These limitations underscore the need for the development of nanodelivery systems to enhance in vivo bioavailability and targeted accumulation.
Inhibition of Epithelial-Mesenchymal Transition (EMT) and Metastasis
Salidroside interfered with tumor angiogenesis, adaptation to hypoxia, EMT, and metastatic spread in breast cancer. Under experimentally induced hypoxic conditions (1–2% O2), Rhodiola rosea extract reduced hypoxia-inducible factor 1α (HIF-1α)/HIF-2α expression by 51% in MCF-7 cells, which was associated with reduced cellular proliferation.16 In human umbilical vein endothelial cells (HUVECs), the extract inhibited the phosphorylation of signal transducer and activator of transcription 3 (STAT3) and subsequent matrix metalloproteinase 2 (MMP2) activity, leading to a 65% reduction in tube formation.17 Concurrent studies in MDA-MB-231 cells demonstrated suppression of the EGFR/Jak2/STAT3-MMP2 signaling pathway, thereby impaired migration, invasion, and angiogenic potential.17 Research conducted in a non-oncological setting, specifically utilizing a rat model of cerebral small vessel disease (CSVD), has demonstrated that the administration of salidroside (24 mg/kg, intraperitoneally) enhanced vascular integrity and function. This effect is associated with the upregulation of Notch1, Hes1/5, and ITGB1 signaling pathways.18 Though derived from non-breast cancer models, this finding offer mechanistic insights likely applicable to TNBC due to the conserved nature of these pathways. These findings collectively indicate that salidroside may have dual effects on tumor vasculature, potentially combining anti-angiogenic properties with vascular stabilization. Furthermore, Salidroside directly influences cellular plasticity and metastasis. In hypoxic MDA-MB-231 cells (1% O2), treatment with 50 µM salidroside results in a 62% reduction in Twist1 expression and a 58% reduction in Zeb1 expression, while simultaneously inducing a 2.1-fold increase in E-cadherin expression. This modulation leads to a 73% decrease in Transwell invasion.19,20 Beyond these cell-intrinsic effects, salidroside also disrupted the pre-metastatic niche by significantly reducing nicotine-induced N2 neutrophil infiltration by 81% and inhibiting STAT3-mediated LCN2 release by 75%, thereby decreased the lung metastatic burden by 69% in hormone receptor-negative breast cancer models.21
A significant translational challenge is that the experimental hypoxic conditions (1–2% O2) employed are more severe than those observed in clinical breast tumor hypoxia (5–10% O2).22 Furthermore, current metastasis models do not accurately replicate the organotropic metastasis of human triple-negative breast cancer (TNBC), particularly to bone and brain.23 The development of nanocarriers with tumor microenvironment-responsive targeting capabilities offers a potential solution by enabling the targeted delivery of salidroside to hypoxic tumor regions, thereby enhancing its anti-metastatic efficacy in clinically relevant settings.
Metabolic Reprogramming & Ferroptosis
Metabolic reprogramming is a critical adaptive mechanism in breast cancer, facilitating malignant proliferation and survival. Salidroside disrupted the metabolic reprogramming of breast cancer cells and induced ferroptosis, a form of iron-dependent regulated cell death intricately linked to metabolic processes. Preclinical studies have demonstrated that salidroside exerts dual modulation of energy metabolism and cell death pathways. At a concentration of 100 µM in vitro treatment, salidroside decreased the expression of HIF-1α by 62% and reduces the levels of glycolytic enzymes hexokinase 2 (HK2) and lactate dehydrogenase A (LDHA) by 45–58%, resulting in a 68% reduction in lactate production.24–27 Simultaneously, it enhanced the activity of mitochondrial complexes I and III, thereby promoting a metabolic shift towards oxidative phosphorylation and mitigating the Warburg effect.24–27 These metabolic changes were mechanistically associated with the suppression of the PI3K/AKT/mTOR signaling pathway, activation of AMPK, and engagement of the SIRT1-mediated PGC-1α/NRF-1/TFAM axis, which collectively promote mitochondrial biogenesis.28,29 This metabolic reprogramming induces ferroptosis in triple-negative breast cancer (TNBC) by inhibiting stearoyl-CoA desaturase-1 (SCD1) through salidroside, leading to a 73% reduction in oleic acid synthesis, and by activating nuclear receptor coactivator 4 (NCOA4)-mediated ferritinophagy, resulting in a 3.1-fold increase in lipid peroxidation.27 Notably, the induction of ferroptosis significantly enhances sensitivity to oxaliplatin by 30% through the inhibition of the SLC7A11/GPX4 pathway, demonstrating substantial chemosynergistic effects.27
In addition to its role in chemosensitization, the SLC7A11/GPX4 axis serves as a pivotal component of the ferroptotic defense network. SLC7A11, also referred to as xCT, acts as the light chain subunit of system Xc⁻, a cystine-glutamate antiporter responsible for importing cystine in exchange for intracellular glutamate. Once imported, cystine is reduced to cysteine, a rate-limiting precursor for glutathione (GSH) synthesis. GPX4 subsequently employs GSH as a cofactor to reduce phospholipid hydroperoxides, thereby averting lethal lipid peroxidation and ferroptosis. Salidroside disrupts this antioxidant defense by inhibiting SLC7A11/GPX4 signaling, resulting in the accumulation of lipid reactive oxygen species (ROS) and the induction of ferroptotic cell death. This mechanism is particularly pertinent in triple-negative breast cancer (TNBC), which frequently exhibits increased system Xc⁻ activity and GSH biosynthesis as an adaptive resistance strategy against oxidative stress. Recent studies have independently corroborated that pharmacological inhibition of SLC7A11 sensitizes TNBC cells to ferroptosis inducers, underscoring the therapeutic potential of targeting this axis.27,30
It is important to note that the quantitative data on SCD1 inhibition (73% reduction in oleic acid synthesis) and NCOA4-mediated ferritinophagy (3.1-fold increase in lipid peroxidation) are primarily derived from a single research group and require independent validation. The precise molecular interaction between salidroside and SLC7A11/GPX4, whether it is direct or indirect, has yet to be comprehensively elucidated. This represents a significant avenue for future mechanistic research.27,30
The principal obstacle in translation is that the effective in vitro concentration (≥100 µM) significantly surpasses the pharmacologically achievable levels in humans.30 This issue can be mitigated through the utilization of nanodelivery systems, which enhance intracellular drug accumulation within tumor cells.
Epigenetic Regulation and Immunomodulation of the Tumor Immune Microenvironment (TIME)
Salidroside modulates epigenetic dysregulation and remodels the immunosuppressive TIME to enhance the therapeutic efficacy against triple-negative breast cancer (TNBC). Specifically, in the context of epigenetic regulation, salidroside inhibits DNA methyltransferase 3B (DNMT3B) in MDA-MB-231 cells, leading to a reduction in BRCA1 promoter methylation. This results in a 5.1-fold increase in sensitivity to the poly (ADP-ribose) polymerase (PARP) inhibitor olaparib, representing a promising mechanism for overcoming resistance to PARP inhibitors in TNBC.31 Salidroside inhibits the histone acetyltransferase PCAF, reducing H3K9 acetylation.32 Though derived from non-breast cancer models, this finding offer mechanistic insights likely applicable to TNBC due to the conserved nature of the pathway. Recent studies have identified BPHL as a promoter of TNBC stemness through the resolution of R-loops.33 However, the potential influence of salidroside on this pathway has yet to be elucidated.
In the context of TIME remodeling, salidroside exerts its effects by inhibiting STAT5 phosphorylation in regulatory T cells (Tregs) and facilitating the maturation of dendritic cells (DCs) within humanized TNBC models. This process results in a 1.8-fold increase in CD80/CD86 expression and enhances the efficacy of programmed cell death protein 1 (PD-1) inhibitors by 37%.34 Additionally, salidroside influences macrophage polarization by decreasing NF-κB p65 expression, while liposomal salidroside activates the TLR4-NF-κB pathway, leading to an upregulation of co-stimulatory markers by 42–37% and a 1.78-fold improvement in antigen presentation.35 In preclinical models, salidroside promotes a 2.9/2.5-fold increase in CD8⁺/CD4⁺ T cell infiltration, reduces FOXP3⁺ Tregs by 43.5%, and achieves a tumor inhibition rate of 64.9% ± 5.1%.36 While this finding is derived from model not specific to breast cancer or triple-negative breast cancer (TNBC), they offer mechanistic insights that are likely applicable to TNBC, considering the conserved nature of these pathways. Moreover, clinical validation is sparse; a randomized trial indicated only modest peripheral immune alterations without direct evidence of enhanced intratumoral immunity in breast cancer patients.37,38 Salidroside enhances immune activity at concentrations up to 20 μM but may suppress T cell function at 50 μM or higher, with limited clinical evidence of its effects within tumors.34 Nanocarriers targeting tumors can sustain optimal salidroside levels for immune activation in the tumor microenvironment, minimizing systemic side effects.
Pharmacological Effects of Salidroside in Breast Cancer
Leveraging its molecular mechanisms, salidroside demonstrates three principal pharmacological effects in triple-negative breast cancer (TNBC) that are well-suited for integration with nanodelivery systems: enhancement of chemotherapy sensitivity and reduction of toxicity, radiosensitization, and synergy with immunotherapy and endocrine therapy. These effects are substantiated by preclinical evidence and initial findings from clinical randomized controlled trials (RCTs). The incorporation of nanodelivery systems further augments these effects by overcoming pharmacokinetic limitations (Figure 3 and Table 1).
 |
Table 1 Core Molecular Mechanisms and Pharmacological Effects of Salidroside in Breast Cancer |
 |
Figure 3 Pharmacological effects and nanodelivery optimization of salidroside in TNBC. Therapeutic Effects: Chemosensitization utilizing epirubicin and paclitaxel; Radiosensitization via X-ray irradiation; Immuno/endocrine synergy achieved through anti-PD-1 and tamoxifen. Nanodelivery Advantages: Improved targeting and pharmacokinetics facilitated by PLGA-PEG and RBCm-ARISP. Evidence Level: Blue denotes clinical evidence from randomized controlled trials (RCTs), while Gray indicates preclinical evidence. Abbreviations: PLGA-PEG refers to poly(lactic-co-glycolic acid)-polyethylene glycol; RBCm-ARISP denotes red blood cell membrane-camouflaged salidroside/indocyanine green nanovesicles. |
Chemotherapy Sensitization and Toxicity Mitigation
Salidroside had been shown to counteract chemotherapy resistance in triple-negative breast cancer (TNBC) by downregulating drug efflux pumps and modulating pathways associated with resistance. Specifically, it decreased the half-maximal inhibitory concentration (IC50) of cisplatin from 18.6 μM to 11.2 μM by reducing HIF1α -mediated P-glycoprotein (P-gp) expression by 45%.47,48 Furthermore, it enhanced paclitaxel sensitivity by 78%, lowering the IC50 from 189 nM to 45 nM, through the suppression of the Wnt/β-catenin pathway and downregulation of ABCG2.39 While derived from non-breast cancer models, these findings offer mechanistic insights likely applicable to TNBC due to the conserved nature of these pathways. Salidroside also induced iron depletion via inhibition of the SLC7A11/GPX4 pathway, enhancing oxaliplatin sensitivity and reducing the IC50 by 30%.49 For anthracyclines, combining salidroside with epirubicin significantly enhances tumor cell apoptosis by 2.2-fold via caspase-3 activation.44
Concurrently, salidroside’s antioxidant and anti-inflammatory properties help mitigate chemotherapy-induced toxicities. In a cohort of 60 breast cancer patients, a daily dose of 600 mg salidroside in combination with epirubicin preserved left ventricular function (ΔSR +0.19) and maintained stable serum reactive oxygen species (ROS) levels.44 Additionally, in a group of 42 patients, Rhodiola rosea injection was found to reduce serum cardiac troponin I (cTnI) levels and improve left ventricular ejection fraction (LVEF), thereby alleviating anthracycline-induced cardiotoxicity.46 Salidroside also reduced the incidence of Grade III–IV myelosuppression by 57% and facilitates the recovery of white blood cell counts in patients undergoing chemotherapy.49 Additionally, an RCT (130 patients with various tumors) indicated Rhodiola rosea extract accelerated white blood cell recovery and reduced oral ulcer size and number.45
Radiosensitization and Immuno/Endocrine Therapy Synergy
Salidroside enhances the radiosensitivity of triple-negative breast cancer (TNBC) by modulating hypoxia and DNA repair pathways. At a concentration of 40 μM, salidroside decreases HIF-1α levels by 58%, elevates tumor partial oxygen pressure (pO2) by 2.3-fold, induces a 3.1-fold increase in reactive oxygen species (ROS) production, and inhibits nuclear factor erythroid 2-related factor 2 (Nrf2) nuclear translocation by 62%, resulting in a sensitizer enhancement ratio (SER) of 2.8.41 In the context of immunotherapy, salidroside synergized with anti-programmed cell death protein 1 (anti-PD-1) antibodies to transform “cold” TNBC tumors into “hot” tumors, as evidenced by a 2.9-fold increase in CD8⁺ T cell infiltration and a 43.5% reduction in regulatory T cells (Tregs), culminating in 64.9% tumor inhibition.36 For endocrine therapy, nanocarriers co-loaded with salidroside and tamoxifen overcame tamoxifen resistance by inhibiting the PI3K-AKT signaling pathway, resulting in a 2.8-fold increase in the area under the curve (AUC) and a 53% improvement in tumor inhibition compared to free tamoxifen.42
Nanodelivery Strategies for Salidroside in TNBC Therapy
Advanced nanodelivery systems constitute a fundamental strategy to address the challenges of salidroside’s limited bioavailability, short half-life, and inadequate targeting capability. In this study, we conduct a systematic evaluation of three types of salidroside-based nanoplatforms that exhibit significant translational potential for the treatment of triple-negative breast cancer (TNBC). Our analysis emphasizes their design principles, key performance metrics, and therapeutic efficacy, as detailed in Table 1.
Polymeric Nanoparticles: Poly(Lactic-Co-Glycolic Acid)-Polyethylene Glycol (PLGA-PEG)
PLGA-PEG polymeric nanoparticles represent the most advanced nanodelivery system for salidroside, utilizing FDA-approved materials that ensure excellent biocompatibility. Employing an optimized emulsion-solvent evaporation method, PLGA-PEG-PLGA (with a lactic acid to glycolic acid ratio of 3:1 and a molecular weight of 15 kDa) successfully co-encapsulates salidroside and tamoxifen. This results in nanoparticles with an average diameter of 275.3 ± 44.0 nm,42,50,51 encapsulation efficiencies of 32.63% for salidroside and 49.18% for tamoxifen, and a cumulative release of 82.3% over a 72-hour period.52 This delivery platform significantly extends the half-life of salidroside from 2.3 hours to 17.2 hours,10 enhances the area under the curve (AUC), and reduces the IC50 of tamoxifen by 1.7-fold through the enhanced permeability and retention (EPR) effect and targeted tumor accumulation.42 It demonstrates substantial efficacy in tamoxifen-resistant triple-negative breast cancer (TNBC) models, achieving a 53% greater inhibition of tumor volume compared to free tamoxifen. The system’s translational potential relies on the safety of FDA-approved materials like PLGA and PEG,53–55 but clinical translation is hindered by data gaps, such as the need for comprehensive in vivo safety assessments and manufacturing validation.
Lipid-Polymer Hybrid Nanoparticles (LPNPs)
LPNPs integrate the structural stability of polymeric cores with the biocompatibility of lipid shells, facilitating chemo-photothermal synergistic therapy for triple-negative breast cancer (TNBC). These nanoparticles are synthesized through the co-assembly of poly(lactic-co-glycolic acid)-polyethylene glycol/polypropylene glycol-polyethylene glycol (PLGA-PEG/PPG-PEG) with lecithin and cholesterol, resulting in a core-shell architecture capable of co-loading chemotherapeutic agents and photosensitizers. A notable example included cetuximab-targeted LPNPs, with an average size of 99.88 ± 2.51 nm, a polydispersity index (PDI) of 0.15 ± 0.03, and a zeta potential of −29.17 ± 3.09 mV, which encapsulated irinotecan (CPT-11) with an efficiency of 51.72% and indocyanine green (ICG) at 65.13%.56 These nanoparticles exhibited dual-responsive release triggered by pH and near-infrared (NIR) light; NIR irradiation significantly accelerates drug release, achieving 59.29% irinotecan release within 24 hours, and enhances synergistic chemo-photothermal effects.56 Moreover, surface modification using folate57 or cetuximab56 enhanced the specificity for cancer cells, especially in those overexpressing epidermal growth factor receptor (EGFR) antibodies or folate receptors, such as SW480 and MCF-7 cells. This modification increased specificity towards TNBC cells. Additionally, the enhanced permeability and retention (EPR) effect facilitated passive tumor targeting, thereby mitigating systemic toxicity.56
Biomimetic Nanocarriers: Red Blood Cell Membrane-Camouflaged Nanocarriers (RBCm-ARISP)
The RBCm-ARISP biomimetic nanocarriers proficiently incorporate mechanisms for immune evasion, active targeting of hypoxic environments, and controlled drug release. These features result in improved pharmacokinetics and notable anti-metastatic efficacy relative to traditional nanocarriers, thereby addressing the challenges associated with the limited in vivo circulation and insufficient accumulation of salidroside in hypoxic tumors. These nanocarriers encapsulate salidroside and indocyanine green (ICG) within poly(lactic-co-glycolic acid) (PLGA) cores, are camouflaged with rat red blood cell membranes achieving a coating efficiency of 92.3%, and are further modified with low-density lipoprotein receptor (LDLR) ligands to specifically target hypoxic triple-negative breast cancer (TNBC).43 The nanoparticles exhibited a mean size of 180 ± 15 nm, a zeta potential of −18.5 ± 3.2 mV, and demonstrated a 78% release of salidroside over a 72-hour period, with accelerated release at pH 6.5, simulating the tumor microenvironment.43 The RBC membrane coating significantly reduces macrophage uptake by 76% and extends the circulation half-life to 28.6 hours, compared to 4.2 hours for non-coated nanovesicles, thereby enhancing tumor targeting by a factor of 6.3. In 4T1 TNBC models, these nanocarriers reduce lung metastases by 78%, compared to 69% with free salidroside, demonstrating superior anti-metastatic efficacy. In addition to its therapeutic efficacy, the indocyanine green (ICG) component facilitates real-time near-infrared fluorescence imaging, which permits non-invasive monitoring of nanoparticle biodistribution and tumor accumulation. This theranostic capability establishes a feedback loop for optimizing dosing regimens and assessing treatment response, thereby offering a significant advantage over traditional nanotherapeutics.43
Despite demonstrating promising efficacy in preclinical studies, these nanoplatforms encounter several common challenges in translation to clinical application: (1) Scalable manufacturing: the intricate preparation processes, such as red blood cell membrane coating, result in batch inconsistencies and variable encapsulation efficiency.58–60 (2) Long-term safety: there is a lack of sufficient data regarding in vivo biodegradation, tissue accumulation, and immunogenicity.58 (3) Targeting specificity: the ligands currently in use, such as LDLR and folate, do not possess specificity for triple-negative breast cancer (TNBC), necessitating optimization with anti-EGFR/HER2 antibodies.56,59
Translational Challenges and Future Directions
Salidroside’s accumulation in Rhodiola rosea affects DNA methylation based on altitude (38.18% at 2594 meters vs. 25.00% at 763 meters).61 However, the link between altitude-dependent methylation in Rhodiola and salidroside accumulation is indirect and unproven in breast cancer models.
Salidroside demonstrates significant anticancer potential in preclinical breast cancer models; however, the transition of salidroside from preclinical success to clinical use in TNBC faces three main obstacles: The absence of validated predictive biomarkers presents a significant hurdle. Although salidroside demonstrates subtype-specific effects, clinically actionable thresholds for potential biomarkers such as BRCA1 methylation, p-STAT3, and HIF-1α are yet to be established, and there is a lack of composite signatures for synergistic therapy.17,31,62 Machine learning frameworks have facilitated the discovery of biomarkers in breast cancer,63 and analogous methodologies could be employed to investigate salidroside-responsive subtypes of triple-negative breast cancer (TNBC). Additionally, the issue of non-standardized formulations is evident. The challenge of standardization is further complicated by the fact that the salidroside content in Rhodiola rosea is influenced by environmental factors, such as altitude. For instance, one study found a methylation level of 38.18% at an altitude of 2594 meters, compared to 25.00% at 763 meters.61 While this botanical variation does not directly apply to human breast cancer, it highlights the essential requirement for stringent quality control in the sourcing of raw materials, which is a fundamental prerequisite for any clinical formulation. Native salidroside is characterized by poor solubility (<1 mg/mL), a short half-life (2.3 hours in rats), and low bioavailability (<12%), which hinder the attainment of effective therapeutic concentrations.42,64 Current research often employs non-standardized Rhodiola extracts with varying salidroside content (5–30%), resulting in inconsistent experimental outcomes.45 Furthermore, there is insufficient clinical evidence to support its efficacy. Most available data are derived from simplistic cell lines and xenografts that do not adequately replicate the clinical heterogeneity of triple-negative breast cancer (TNBC). Preliminary clinical data suggest only a potential for toxicity mitigation, without robust evidence of direct antitumor efficacy.
Consequently, this study suggests a three-phase translational roadmap for salidroside-based nanomedicines (Figure 4), following a framework of formulation standardization → biomarker-driven trials → clinical therapy positioning.
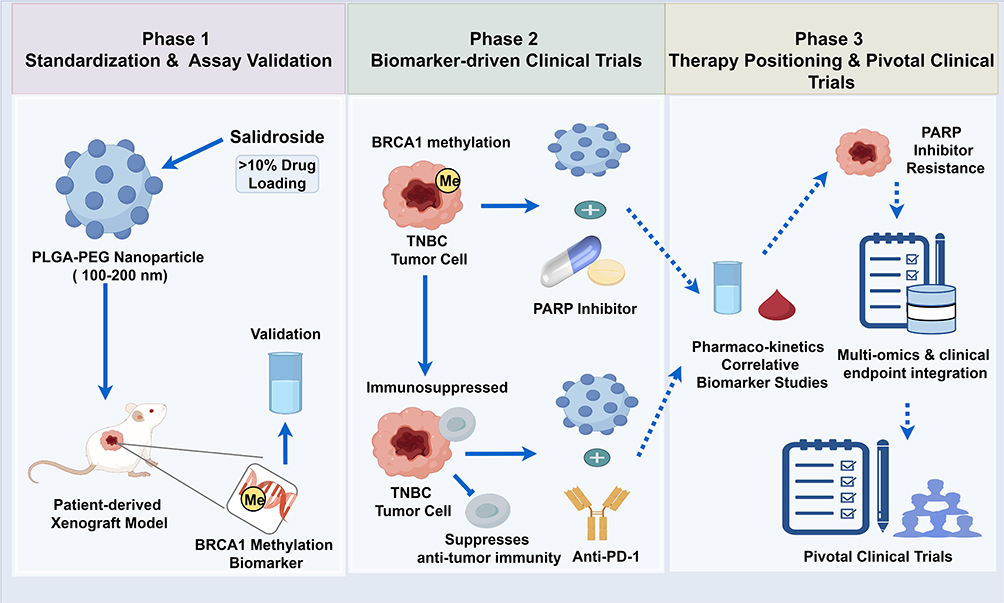 |
Figure 4 Three-phase translational roadmap for salidroside-based nanomedicines in TNBC. This schematic delineates a sequential clinical development strategy for salidroside-loaded nanomedicines in TNBC, with BRCA1 methylation serving as the central predictive biomarker. The yellow circle labeled “Me” represents the BRCA1 methylation biomarker, while the blue circles marked with a plus sign (+) signify combination treatment regimens, specifically nanoparticles in conjunction with PARP inhibitors or anti-PD-1 therapies. All arrows are standardized: solid arrows indicate direct, validated relationships; dashed arrows represent indirect or data-flow relationships; and T-shaped bars (—|) signify inhibition. Phase 1 (Preclinical Standardization and Validation): involves the formulation of PLGA-PEG nanoparticles with over 10% drug loading and a size range of 100–200 nm, validation using PDX models, and the identification of BRCA1 methylation as a biomarker. Phase 2 (Biomarker-Driven Adaptive Clinical Trials): focuses on BRCA1-driven adaptive trials that combine PARP inhibitors with anti-PD-1 therapy to address immunosuppressive TNBC. Phase 3 (Pivotal Trials and Clinical Positioning): encompasses pivotal trials that integrate pharmacokinetics/pharmacodynamics (PK/PD), correlative biomarkers, and multi-omics approaches to overcome resistance to PARP inhibitors. |
Three-Phase Translational Roadmap for Salidroside-Based Nanomedicines.
Phase 1 (Preclinical Standardization and Validation): This phase involves the development of standardized salidroside nanodelivery formulations, ensuring a drug loading efficiency greater than 10%.42,43 Concurrently, biomarker assays, specifically BRCA1 methylation and HIF-1α, would be validated using patient-derived xenograft (PDX) models that accurately represent the heterogeneity of TNBC.
Phase 2 (Biomarker-Driven Adaptive Clinical Trials): This phase could implement basket trials to assess the efficacy of salidroside nanomedicines in biomarker- defined subsets of TNBC, such as BRCA1-methylated TNBC resistant to PARP inhibitors31 and immunosuppressed TNBC with elevated p-STAT3 levels.3,65 The trials will incorporate pharmacokinetic monitoring and correlative biomarker studies.
Similarly, computational models that incorporate various cell death modalities have been developed for ovarian cancer.66 Adapting these models for TNBC could facilitate the rational design of combination treatment regimens.
Phase 3 (Pivotal Trials and Clinical Positioning): Based on data from early-phase studies, pivotal trials will be designed to establish the role of salidroside nanomedicines in overcoming resistance to PARP and PD-1 inhibitors in TNBC.67,68 This phase will also integrate multi-omics data to optimize personalized therapeutic strategies.
Conclusions
Salidroside emerges as a promising and versatile molecular regulator for triple-negative breast cancer (TNBC), demonstrating antitumor efficacy through its modulation of cellular proliferation, metabolic pathways, metastasis, epigenetic mechanisms, and the immune microenvironment. Importantly, its unique dual role in maintaining redox homeostasis-promoting ferroptosis in cancer cells while protecting normal tissues from oxidative damage-may reduce off-target toxicity. This potential advantage warrants further clinical investigation. However, the primary obstacle to its translational application lies in the misalignment between its polypharmacological actions and the prevailing single-target validation frameworks. This challenge is further compounded by suboptimal pharmacokinetic properties and the lack of standardized formulations.
Integrated nanomedicine represents a promising strategy to current challenges, as advanced nanoplatforms such as PLGA-PEG, LPNPs, and RBCm-ARISP significantly enhance the bioavailability and targeted delivery of salidroside. These platforms also facilitate real-time in vivo monitoring of its mechanistic effects, including hypoxia modulation and immune remodeling, thereby validating its polypharmacological potential in clinically relevant settings. In future research, salidroside-based precision nanomedicines, guided by predictive biomarkers like BRCA1 methylation and HIF-1α, in conjunction with standardized formulations, could offer promising therapeutic strategies. These approaches merit further clinical investigation, particularly for biomarker-defined subsets of triple-negative breast cancer (TNBC) that demonstrate resistance to PARP and PD-1 inhibitors. Furthermore, the translational framework delineated in this review provides a generalizable model for the clinical advancement of other pleiotropic natural products, facilitating the integration of phytochemistry with systems-based precision oncology. This methodology has the potential to address unmet therapeutic needs in breast cancer treatment and to advance the development of natural product-based nanomedicine within the oncology domain.
Abbreviations
TNBC, triple-negative breast cancer; PI3K, phosphatidylinositol 3-kinase; AKT, protein kinase B; HIF-1α, hypoxia-inducible factor 1α; SCD1, stearoyl-CoA desaturase 1; NCOA4, nuclear receptor coactivator 4; VEGF, vascular endothelial growth factor; STAT3, signal transducer and activator of transcription 3; MMP2, matrix metalloproteinase 2; ECM, extracellular matrix; EMT, epithelial-mesenchymal transition; Treg, regulatory T cell; DC, dendritic cell; DNMT3B, DNA methyltransferase 3B; PDX, patient-derived xenograft; RCT, randomized controlled trial; PARP, poly(ADP-ribose) polymerase; RBCm, red blood cell membrane; ICG, indocyanine green; PD-1, programmed cell death protein 1; ROS, reactive oxygen species; Wnt/β-catenin, Wingless/Integrated-β-catenin; ABCG2, ATP-binding cassette subfamily G member 2; IC50, half-maximal inhibitory concentration; Nrf2, nuclear factor erythroid 2-related factor 2; SER, sensitizer enhancement ratio; PLGA, poly(lactic-co-glycolic acid); PEG, polyethylene glycol; AUC, area under the curve; WBC, white blood cell; cTnI, cardiac troponin I; LVEF, left ventricular ejection fraction; MTD, maximum tolerated dose; PLGA-PEG, poly(lactic-co-glycolic acid)-polyethylene glycol; RBCm-ARISP, red blood cell membrane-camouflaged salidroside/ICG co-loaded.
Data Sharing Statement
Data sharing is not applicable to this article as no datasets were generated or analyzed during the current study.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This research was supported by the Leading Talent Project of “Kunlun Talents High-end Innovation and Entrepreneurship Talents” Program in Qinghai Province (Qing Wei Jian Ban [2021] No. 104) and the Development of Outstanding Individuals initiative of “Kunlun Talents High-end Innovation and Entrepreneurship Talents” Program in Qinghai Province (Qing Talent Document [2025] No. 17).
Disclosure
The authors declare no competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
References
1. Kinowaki K, Takeiwa T, Ikeda K. et al. Positive immunoreactivities of TM9SF1 and EBAG9 are associated with Poor Prognosis in patients With triple-negative breast cancer. Clin Breast Cancer. 2026;26(2):259–267.e6. doi:10.1016/j.clbc.2025.09.007
2. Alonso-Ron C, Vethencourt A, González-Suárez E, et al. Triple-negative breast cancer systemic treatment: disruptive early-stage developments for overcoming stagnation in the advanced pipeline. Cancers. 2025;17(4):633. doi:10.3390/cancers17040633
3. Ciavattone NG, Bevoor A, Farfel A, et al. Inhibiting CXCR4 reduces immunosuppressive effects of myeloid cells in breast cancer immunotherapy. Sci Rep. 2025;15(1):5204. doi:10.1038/s41598-025-89882-5
4. Yu X, Wei P, Chen Z, et al. Comparative analysis of the organelle genomes of three Rhodiola species provide insights into their structural dynamics and sequence divergences. BMC Plant Biol. 2023;23(1):156. doi:10.1186/s12870-023-04159-1
5. Wang S, Feng Y, Zheng L, et al. Rosavin: research advances in extraction and synthesis, pharmacological activities and therapeutic effects on diseases of the characteristic active ingredients of Rhodiola rosea L. Molecules. 2023;28(21):7412. doi:10.3390/molecules28217412
6. Mattioli L, Funari C, Perfumi M. Effects of Rhodiola rosea L. extract on behavioural and physiological alterations induced by chronic mild stress in female rats. J Psychopharmacol. 2009;23(2):130–14. doi:10.1177/0269881108089872
7. Li C, Li Y, Huang X, et al. Traditional Tibetan medicine: therapeutic potential in lung diseases. Front Pharmacol. 2024;15:1365911. doi:10.3389/fphar.2024.1365911
8. Voronin MV, Abramova EV, Verbovaya ER, et al. Chaperone-dependent mechanisms as a pharmacological target for neuroprotection. Int J Mol Sci. 2023;24(1):823. doi:10.3390/ijms24010823
9. Wang J, Yang Z, Feng Z, et al. Synthesis of salidroside derivatives at C4 position of benzene ring and its effect on Hep3B cell viability. Chem Biol Drug Des. 2025;105(6):e70131. doi:10.1111/cbdd.70131
10. Liang K, Ma S, Luo K, et al. Salidroside: an overview of its promising potential and diverse applications. Pharmaceuticals. 2024;17(12):1703. doi:10.3390/ph17121703
11. Hong Y, Li Y, Liu X, et al. Salidroside inhibits the invasion and migration of colorectal cancer cells by regulating MMP-12 and WNT signaling pathway. Am J Cancer Res. 2025;15(3):929–945. doi:10.62347/KELA7583
12. Wojdasiewicz P, Brodacki S, Cieślicka E, et al. Salidroside: a promising agent in bone metabolism modulation. Nutrients. 2024;16(15):2387. doi:10.3390/nu16152387
13. Hu X, Zhang X, Qiu S, Yu D, Lin S. Salidroside induces cell-cycle arrest and apoptosis in human breast cancer cells. Biochem Biophys Res Commun. 2010;398(1):62–67. doi:10.1016/j.bbrc.2010.06.033
14. Sun AQ, Ju XL. Inhibitory effects of salidroside on MCF-7 breast cancer cells in vivo. J Int Med Res. 2020;48(11):300060520968353. doi:10.1177/0300060520968353
15. Stöger H, Wilders-Truschnig M, Samonigg H, et al. The presence of immunosuppressive ‘p15E-like’ factors in the serum and urine of patients suffering from malign and benign breast tumours. Clin Exp Immunol. 1993;93(3):437–441. doi:10.1111/j.1365-2249.1993.tb08197
16. Qi YJ, Cui S, Lu DX, et al. Effects of the aqueous extract of a Tibetan herb, Rhodiola algida var. tangutica on proliferation and HIF-1α, HIF-2α expression in MCF-7 cells under hypoxic condition in vitro. Cancer Cell Int. 2015;15:81. doi:10.1186/s12935-015-0225-x
17. Kang DY, Sp N, Kim DH, et al. Salidroside inhibits migration, invasion and angiogenesis of MDA-MB231 TNBC cells by regulating EGFR/Jak2/STAT3 signaling via MMP2. Int J Oncol. 2018;53(2):877–885. doi:10.3892/ijo.2018.4430
18. Zhilan T, Zengyu Z, Pengpeng J, et al. Salidroside promotes pro-angiogenesis and repair of blood brain barrier via Notch/ITGB1 signal path in CSVD Model. J Adv Res. 2024;68:429–444. doi:10.1016/j.jare.2024.02.019
19. Perez-Oquendo M, Manshouri R, Tian Y, et al. ZEB1 is regulated by K811 acetylation to promote stability, NuRD complex interactions, EMT, and NSCLC metastasis. Mol Cancer Res. 2023;21(8):779–794. doi:10.1158/1541-7786.Mcr-22-0503
20. Chen J, Imanaka N, Chen J, et al. Hypoxia potentiates Notch signaling in breast cancer leading to decreased E-cadherin expression and increased cell migration and invasion. Br J Cancer. 2010;102(2):351–360. doi:10.1038/sj.bjc.6605486
21. Tyagi A, Sharma S, Wu K, et al. Nicotine promotes breast cancer metastasis by stimulating N2 neutrophils and generating pre-metastatic niche in lung. Nat Commun. 2021;12(1):474. doi:10.1038/s41467-020-20733-9
22. Majmundar AJ, Wong WJ, Simon MC. Hypoxia-inducible factors and the response to hypoxic stress. Mol Cell. 2010;40(2):294–309. doi:10.1016/j.molcel.2010.09.022
23. Bos PD, Zhang XHF, Nadal C, et al. Genes that mediate breast cancer metastasis to the brain. Nature. 2009;459(7249):1005–1009. doi:10.1038/nature08021
24. Hou Y, Zhang Y, Jiang S, et al. Salidroside intensifies mitochondrial function of CoCl2-damaged HT22 cells by stimulating PI3K-AKT-MAPK signaling pathway. Phytomedicine. 2022;109:154568. doi:10.1016/j.phymed.2022.154568
25. Fan F, Xu N, Sun Y, et al. Uncovering the metabolic mechanism of salidroside alleviating microglial hypoxia inflammation based on microfluidic chip-mass spectrometry. J Proteome Res. 2021;21(4):921–929. doi:10.1021/acs.jproteome.1c00647
26. Liao W, Liu J, Wang S, et al. Metabolic profiling reveals that salidroside antagonizes hypoxic injury via modulating energy and lipid metabolism in cardiomyocytes. Biomed Pharmacother. 2019;122:109700. doi:10.1016/j.biopha.2019.109700
27. Huang G, Cai Y, Ren M, et al. Salidroside sensitizes triple-negative breast cancer to ferroptosis by SCD1-mediated lipogenesis and NCOA4-mediated ferritinophagy. J Adv Res. 2025;74:589–607. doi:10.1016/j.jare.2024.09.027
28. Cui L, Ye C, Luo T, et al. Exploring the pharmacological action and molecular mechanism of salidroside in inhibiting MCF-7 cell proliferation and migration. J Vis Exp. 2023;196:e65634. doi:10.3791/65634
29. Iqbal MA, Chattopadhyay S, Siddiqui FA, et al. Silibinin induces metabolic crisis in triple-negative breast cancer cells by modulating EGFR-MYC-TXNIP axis: potential therapeutic implications. FEBS J. 2021;288(2):471–485. doi:10.1111/febs.15353
30. Zhang X, Zhang M, Zhang Z, et al. Salidroside induces mitochondrial dysfunction and ferroptosis to inhibit melanoma progression through reactive oxygen species production. Exp Cell Res. 2024;438(1):114034. doi:10.1016/j.yexcr.2024.114034
31. Lonning PE, Nikolaienko O, Pan K, et al. Constitutional BRCA1 methylation and risk of incident triple-negative breast cancer and high-grade serous ovarian cancer. JAMA Oncol. 2022;8(11):1579–1587. doi:10.1001/jamaoncol.2022.3846
32. Chen D, Lu D, Liu H, et al. Pharmacological blockade of PCAF ameliorates osteoarthritis development via dual inhibition of TNF-α-driven inflammation and ER stress. EBioMedicine. 2019;50:395–407. doi:10.1016/j.ebiom.2019.10.054
33. Yuan Q, Zhang H, Zhang N, et al. BPHL promotes TNBC stemness by resolving R-loops via POLR2A lactylation inhibition and BARD1-mediated ubiquitination. Cancer Lett. 2026;645:218388. doi:10.1016/j.canlet.2026.218388
34. Zhao X, Lu Y, Tao Y, et al. Salidroside liposome formulation enhances the activity of dendritic cells and immune responses. Int Immunopharmacol. 2013;17(4):1134–1140. doi:10.1016/j.intimp.2013.10.016
35. Wang X-L, Sun R-X, Li D-X, et al. Salidroside regulates mitochondrial homeostasis after polarization of RAW264.7 macrophages. J Cardiovasc Pharmacol. 2023;81(1):85–92. doi:10.1097/FJC.0000000000001362
36. Wen Z, Liu T, Zhang Y, et al. Salidroside regulates tumor microenvironment of non-small cell lung cancer via Hsp70/Stub1/Foxp3 pathway in Tregs. BMC Cancer. 2023;23(1):717. doi:10.1186/s12885-023-11036-5
37. Guan S, He J, Guo W, et al. Adjuvant effects of salidroside from Rhodiola rosea L. on the immune responses to ovalbumin in mice. Immunopharmacol Immunotoxicol. 2011;33(4):738–743. doi:10.3109/08923973.2011.567988
38. Lu L, Yuan J, Zhang S. Rejuvenating activity of salidroside (SDS): dietary intake of SDS enhances the immune response of aged rats. Age. 2012;35(3):637–646. doi:10.1007/s11357-012-9394-x
39. Pi GL, Cheng LH, Li BF, et al. Salidroside Enhances the Sensitivity of Lung Cancer Cells to Paclitaxel by Regulating the Wnt/β-catenin Signaling Pathway. Curr Med Sci. 2025;45(1):104–113. doi:10.1007/s11596-025-00005-7
40. Yu S, Liu W. Protective effects of Salidroside against ferroptosis through PPARG-dependent mechanism in diabetes-related cognitive impairment. Exp Neurol. 2025;390:115261. doi:10.1016/j.expneurol.2025.115261
41. Li Q, Chen Q, Xiao S, et al. A salidroside-based radiosensitizer regulates the Nrf2/ROS pathway for X-ray activated synergistic cancer precise therapy. Adv Mater. 2025;37(24):e2413226. doi:10.1002/adma.202413226
42. Yu X, Sun L, Tan L, et al. Preparation and Characterization of PLGA-PEG-PLGA Nanoparticles Containing Salidroside and Tamoxifen for Breast Cancer Therapy. AAPS Pharm Sci Tech. 2020;21(3):85. doi:10.1208/s12249-019-1523-8
43. Pan Y, He Y, Zhao X, et al. Engineered red blood cell membrane-coating salidroside/indocyanine green nanovesicles for high-efficiency hypoxic targeting phototherapy of triple-negative breast cancer. Adv Healthc Mater. 2022;11(17):e2200962. doi:10.1002/adhm.202200962
44. Zhang H, Shen W, Gao C, et al. Protective effects of salidroside on epirubicin-induced early left ventricular regional systolic dysfunction in patients with breast cancer. Drugs R D. 2012;12(2):101–106. doi:10.2165/11632530-000000000-00000
45. Loo WTY, Jin LJ, Chow LWC, et al. Rhodiola algida improves chemotherapy-induced oral mucositis in breast cancer patients. Expert Opin Investig Drugs. 2010;19(Suppl 1):S91–S100. doi:10.1517/13543781003727057
46. Shen WS, Gao CH, Zhang H. Effect of rhodiola on serum troponin 1, cardiac integral backscatter and left ventricle ejective fraction of patients who received epirubicin-contained chemotherapy [article in Chinese]. Zhongguo Zhong Xi Yi Jie He Za Zhi. 2010;30(12):1250–1252. PMID: 21302483.
47. Qin Y, Liu HJ, Li M, et al. Salidroside improves the hypoxic tumor microenvironment and reverses the drug resistance of platinum drugs via HIF-1α signaling pathway. EBioMedicine. 2018;38:25–36. doi:10.1016/j.ebiom.2018.10.069
48. Qin Y, Liu HJ, Li M, et al. Corrigendum to “Salidroside improves the hypoxic tumor microenvironment and reverses the drug resistance of platinum drugs via HIF-1α signaling pathway. EBioMedicine. 2024;106:105237. doi:10.1016/j.ebiom.2024.105237
49. Liu S, Li Y, Li Z. Salidroside suppresses the activation of nasopharyngeal carcinoma cells via targeting miR-4262/GRP78 axis. Cell Cycle. 2022;21(7):720–729. doi:10.1080/15384101.2021.2019976
50. El-Hammadi MM, Arias JL. Recent advances in the surface functionalization of PLGA-based nanomedicines. Nanomaterials. 2022;12(3):354. doi:10.3390/nano12030354
51. Kesharwani P, Kumar V, Goh KW, et al. PEGylated PLGA nanoparticles: unlocking advanced strategies for cancer therapy. Mol Cancer. 2025;24(1):205. doi:10.1186/s12943-025-02410-x
52. Sanna V, Siddiqui IA, Sechi M, et al. Resveratrol-loaded nanoparticles based on poly(epsilon-caprolactone) and poly(D,L-lactic-co-glycolic acid)-poly(ethylene glycol) blend for prostate cancer treatment. Mol Pharm. 2013;10(10):3871–3881. doi:10.1021/mp400342f
53. Dashti M, Amir A, Yousefi M. Cost-optimized placenta-targeted nanoparticle for localized immune cloaking in recurrent pregnancy loss. Reprod Fertil. 2025;6(4):e250077. doi:10.1530/RAF-25-0077
54. Bhattacharya S, Shinde P, Page A, et al. Poly lactic co-glycolic acid d-α-tocopheryl polyethylene glycol 1000 succinate fabricated polyethylene glycol hybrid nanoparticles of imatinib mesylate for the treatment of glioblastoma multiforme. Curr Med Chem. 2025;32(37):8350–8370. doi:10.2174/0109298673323270241118103546
55. Lee CK, Atibalentja DF, Yao LE, et al. Anti-PD-L1 F(ab) Conjugated PEG-PLGA Nanoparticle Enhances Immune Checkpoint Therapy. Nanotheranostics. 2022;6(3):243–255. doi:10.7150/ntno.65544
56. Fang F, Chen YY, Zhang X-M, et al. EGFR-targeted and NIR-triggered lipid-polymer hybrid nanoparticles for chemo-photothermal colorectal tumor therapy. Int J Nanomed. 2024;19:9689–9705. doi:10.2147/IJN.S473473
57. Gu L, Shi T, Sun Y, et al. Folate-modified, indocyanine green-loaded lipid-polymer hybrid nanoparticles for targeted delivery of cisplatin. J Biomater Sci Polym Ed. 2017;28(7):690–702. doi:10.1080/09205063.2017.1296347
58. Alawi M, Hilles AR, Kumar M, et al. Lipid-polymer hybrid nanoparticles: a cutting-edge frontier in breast cancer treatment strategies. Nanomedicine. 2025;20(14):1775–1798. doi:10.1080/17435889.2025.2523730
59. Graván P, Marchal JA, Galisteo-González F. Improving tumor treatment: cell membrane-coated nanoparticles for targeted therapies. Mater Today Bio. 2025;32:101716. doi:10.1016/j.mtbio.2025.101716
60. Fang DL, Chen Y, Xu B, et al. Development of lipid-shell and polymer core nanoparticles with water-soluble salidroside for anti-cancer therapy. Int J Mol Sci. 2014;15(3):3373–3388. doi:10.3390/ijms15033373
61. Zhao W, Shi X, Li J, et al. Genetic, epigenetic, and HPLC fingerprint differentiation between natural and ex situ populations of Rhodiola sachalinensis from Changbai Mountain. China PLoS One. 2014;9(11):e112869. doi:10.1371/journal.pone.0112869
62. Zheng XD, Li HY, Gao SY, et al. High hypoxia inducible factor-1α expression is associated with reduced survival in patients with breast cancer: a meta-analysis. World J Clin Oncol. 2025;16(6):105691. doi:10.5306/wjco.v16.i6.105691
63. Yuan Q, Sha Y, Ye R, et al. Machine learning-based identification of kbhb-affected tumor cell subsets as prognostic and therapeutic targets in breast cancer. J Transl Med. 2025;24(1):71. doi:10.1186/s12967-025-07555-3
64. Zhou F, Huang W, Li M, et al. Bioaccessibility and absorption mechanism of phenylethanoid glycosides using simulated digestion/Caco-2 intestinal cell models. J Agric Food Chem. 2018;66(18):4630–4637. doi:10.1021/acs.jafc.8b01307
65. Panagi M, Voutouri C, Mpekris F, et al. TGF-β inhibition combined with cytotoxic nanomedicine normalizes triple negative breast cancer microenvironment towards anti-tumor immunity. Theranostics. 2020;10(4):1910–1922. doi:10.7150/thno.36936
66. Yu G, Yuan Q, Sun Z, et al. A machine learning-driven framework integrating cell death and senescence signatures for multi-target drug design and immunotherapy optimization in ovarian cancer. NPJ Precis Oncol. 2026. doi:10.1038/s41698-026-01448-4
67. Zhang X, Wang Y, G A, et al. Pan-cancer analysis of PARP1 alterations as biomarkers in the prediction of immunotherapeutic effects and the association of its expression levels and immunotherapy signatures. Front Immunol. 2021;12:721030. doi:10.3389/fimmu.2021.721030
68. Mehta AK, Cheney EM, Hartl CA, et al. Targeting immunosuppressive macrophages overcomes PARP inhibitor resistance in BRCA1-associated triple-negative breast cancer. Nat Cancer. 2021;2(1):66–82. doi:10.1038/s43018-020-00148-7
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Iron-Dependent Cell Death: Exploring Ferroptosis as a Unique Target in Triple-Negative Breast Cancer Management
Tan LK, Liu J, Ma CZ, Huang S, He FH, Long Y, Zheng ZS, Liang JL, Xu N, Wang G, Liu YF
Cancer Management and Research 2025, 17:625-637
Published Date: 19 March 2025
An Experimental Study into the Photodynamic/Sonodynamic Therapy of Triple-Negative Breast Cancer Mediated by the Novel Nano-Photonic Sensitizer NBDPBr Nanoparticles
Huang Q, Wang L, Zhang Y, Zhao L, Zhang T
International Journal of Nanomedicine 2026, 21:576927
Published Date: 28 March 2026
Diselenide-Bridged Iron-Porphyrin MOF for MRI-Guided Radiotherapy via Triple-Pathway Ferroptosis
Huang W, Zheng G, Mo B, Jing G, Gao M, Zhao J, Liu H, Luo S, Yu S
International Journal of Nanomedicine 2026, 21:610544
Published Date: 3 June 2026