Back to Journals » Journal of Inflammation Research » Volume 17
Revolutionizing Treatment Strategies for Autoimmune and Inflammatory Disorders: The Impact of Dipeptidyl-Peptidase 4 Inhibitors
Authors Rahim K, Shan M, Ul Haq I, Nawaz MN ![]() , Maryam S
, Maryam S ![]() , Alturki MS, Al Khzem AH
, Alturki MS, Al Khzem AH ![]() , Metwally K, Cavalu S, Alqifari SF
, Metwally K, Cavalu S, Alqifari SF ![]() , Yahya G
, Yahya G ![]()
Received 18 October 2023
Accepted for publication 9 March 2024
Published 23 March 2024 Volume 2024:17 Pages 1897—1917
DOI https://doi.org/10.2147/JIR.S442106
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Adam Bachstetter
Kashif Rahim,1 Muhammad Shan,2 Ihtisham Ul Haq,3– 5 Muhammad Naveed Nawaz,2 Sajida Maryam,3,4 Mansour S Alturki,6 Abdulaziz H Al Khzem,6 Kamel Metwally,7 Simona Cavalu,8 Saleh F Alqifari,9 Galal Yahya10
1School of Life Science and Technology, Beijing University of Chemical Technology, Beijing, 100029, People’s Republic of China; 2Department of Microbiology, Cholistan University of Veterinary and Animal Sciences, Bahawalpur, Punjab, Pakistan; 3Department of Physical Chemistry and Technology of Polymers, Silesian University of Technology, Gliwice, Poland; 4Joint Doctoral School, Silesian University of Technology, Gliwice, 44-100, Poland; 5Programa de Pós-graduação em Inovação Tecnológica, Universidade Federal de Minas Gerais, Belo Horizonte, MG, 31270-901, Brazil; 6Department of Pharmaceutical Chemistry, College of Clinical Pharmacy, Imam Abdulrahman Bin Faisal University, Dammam, 34212, Saudi Arabia; 7Department of Medicinal Chemistry, Faculty of Pharmacy, University of Tabuk, Tabuk, 71491, Saudi Arabia; 8Faculty of Medicine and Pharmacy, University of Oradea, Oradea, 410073, Romania; 9Department of Pharmacy Practice, Faculty of Pharmacy, University of Tabuk, Tabuk, 71491, Saudi Arabia; 10Department of Microbiology and Immunology, Faculty of Pharmacy, Zagazig University, Al Sharqia, 44519, Egypt
Correspondence: Kashif Rahim, School of Life Science and Technology, Beijing University of Chemical Technology, Beijing, 100029, People’s Republic of China, Email [email protected] Saleh F Alqifari, Department of Pharmacy Practice, Faculty of Pharmacy, University of Tabuk, Tabuk, 71491, Saudi Arabia, Email [email protected]
Abstract: DPP4 (Dipeptidyl-peptidase 4) a versatile protease, emerges as a prominent player in soluble and membrane-bound forms. Its heightened expression has been intimately linked to the initiation and severity of diverse autoimmune diseases, spanning rheumatoid arthritis, systemic lupus erythematosus, systemic sclerosis (SSc), inflammatory bowel disease, autoimmune diabetes, and even SARS-CoV-2 infection. Operating as a co-stimulator of T cell activity, DPP4 propels T cell proliferation by binding adenosine deaminase (ADA), thereby augmenting the breakdown of adenosine—an influential inhibitor of T cell proliferation. However, the discovery of a wide range of DPP4 inhibitors has shown promise in alleviating these diseases’ signs, symptoms, and severity. The available DPP4 inhibitors have demonstrated significant effectiveness in blocking DPP4 activity. Based on the characterization of their binding mechanisms, three distinct groups of DPP4 inhibitors have been identified: saxagliptin, alogliptin, and sitagliptin, each representing a different class. Elevated levels of angiotensin-converting enzyme 2 (ACE2) expression are associated with producing various coronavirus peptidases. With its anti-inflammatory properties, Sitagliptin may assist COVID-19 patients in preventing and managing cytokine storms. This comprehensive review delves into the burgeoning realm of DPP4 inhibitors as therapeutic interventions for diverse autoimmune diseases. With a discerning focus on their efficacy, the investigation sheds light on their remarkable capacity to alleviate the burdensome signs and symptoms intricately linked to these conditions.
Keywords: autoimmune diseases, DPP4 inhibitors, inflammatory disorders, MERS-CoV, diabetes mellitus, COVID-19
Introduction
Dipeptidyl-peptidase 4 (DPP4), also known as CD26, is a significant enzyme belonging to the SC serine peptidase clan, specifically within its subfamily 9b. The structure and function of DPP4 are characterized by a distinct enzymatic triad. This triad is composed of three amino acids: Serine (Ser), Aspartic acid (Asp), and Histidine (His). These are crucial for the enzyme’s activity and are situated within a specific structural motif known as the a/b-hydrolase loop.1 This combination of Ser, Asp, and His in DPP4’s active site is what sets it apart from other similar enzymes. For comparison, the catalytic triad in enzymes like chymotrypsin, another well-known serine protease, comprises Histidine (His), Aspartic acid (Asp), and Serine (Ser). Although the components are the same, the order and specific arrangement within the protein structure differ significantly, leading to differences in their enzymatic functions. DPP4 was first identified in 1966. Since its discovery, DPP4 has been extensively studied due to its role in various physiological processes and its implications in medical conditions like diabetes and cancer. As CD26, it is also known for its role as a cell surface protein involved in immune regulation and signal transduction.2
DPP4 predominantly exists as dimers on the cell surface and is widely distributed throughout the body, being present in epithelial cells, certain endothelial cells, leukocyte subsets, and fibroblasts.3,4 Human DPP4 comprises 766 amino acids, with a 738-amino-acid extracellular portion, a 6-amino-acid cytoplasmic tail, and a 22-amino-acid transmembrane hydrophobic segment.3,5,6 Cleavage of DPP4 from the cell membrane releases a soluble form of the enzyme, which maintains its catalytic activity as the active domain resides outside the cell.
Further studies revealed that DPP4 acts as a binding protein for adenosine deaminase (ADA) and serves as a T cell activation marker, transmitting co-stimulatory signals upon ADA stimulation.7 In 2013, DPP4 was identified as the entrance receptor for Middle East Respiratory Syndrome Coronavirus (MERS-CoV), facilitating viral entry into respiratory epithelial cells through binding to the spike protein on the virus’s surface.8–11 Additionally, DPP4 has been recognized as a co-receptor for human immunodeficiency virus (HIV) alongside CCR5, with the expression of DPP4+ T cell subsets possibly explaining the connection between HIV infection and DPP4.12–14 Recent studies have also unveiled new roles for DPP4 in wound healing and tissue scarring, with DPP4-expressing fibroblasts playing a significant role in connective tissue deposition in dermal scars.15
In acute viral infections such as SARS and MERS, individuals with severe illness exhibited higher viral loads and prolonged viral shedding in respiratory secretions compared to those with mild disease.16 This highlights the importance of the immunological response and viral efforts to evade it in determining disease outcomes, particularly in individuals with comorbidities. DPP4 inhibitors have shown significant effectiveness in blocking DPP4 activity, with clinical studies achieving over 80–90% inhibition.17 Moreover, by interacting with ligands such as ADA, DPP4 exerts noncatalytic functions, including co-stimulating T-cell activation and promoting lymphocyte proliferation, which can be beneficial in modulating immune responses.18
Considering the tissue distribution of DPP4, the expression levels of angiotensin-converting enzyme 2 (ACE2) become crucial, as they are associated with the production of coronavirus peptidases. Notably, a close relationship has been observed between the S1 protein of SARS-CoV-2 and DPP4, emphasizing the significance of DPP4’s tissue distribution in understanding viral interactions.19
DPP4 inhibitors have gained recognition as oral anti-diabetic medications for the treatment of type 2 diabetes. Sitagliptin was the pioneering agent, followed by linagliptin, vildagliptin, saxagliptin, alogliptin, anagliptin, gemigliptin, and teneligliptin.20 Three distinct groups of DPP4 inhibitors have been proposed based on the characterization of the binding mechanisms of the clinically most often used DPP4 inhibitors: Saxagliptin and vildagliptin, which belong to Class 1, engage with the S1- and S2 subsites of the active center and bind chemically to Ser630 of the DPP4 molecule. In addition to binding to S1 and S2, alogliptin and linagliptin also interact with S1 and S2’ and are members of class 2. Class 3 DPP4 inhibitors include sitagliptin, anagliptin, gemigliptin, and teneligliptin.21,22
Dipeptidyl peptidase-4 (DPP4), also known as CD26, plays a crucial role in both protective immunity and immune tolerance. In protective immunity, DPP4 is involved in T cell activation and proliferation, crucial for defending against pathogens, and influences cytokine production, affecting the immune response’s nature and strength. Additionally, it modifies chemokines, impacting immune cell migration and inflammation. Regarding immune tolerance, DPP4 regulates the intensity and duration of T cell responses to prevent excessive or inappropriate immune reactions, potentially aiding in the function of regulatory T cells (Tregs) which are pivotal in maintaining immune tolerance and preventing autoimmunity. This dual role in enhancing immune responses and promoting tolerance highlights DPP4’s significance in the immune system and its potential as a target for treating various immune-related conditions.
Dipeptidyl peptidase-4 (DPP4) plays a significant role in the development and exacerbation of autoimmune and autoinflammatory dermatoses, notably psoriasis and bullous pemphigoid. In psoriasis, DPP4 contributes to the inflammatory process by influencing the function and migration of key immune cells like T cells and dendritic cells, as well as modulating cytokines and chemokines, thereby exacerbating skin inflammation. In the case of bullous pemphigoid, an autoinflammatory blistering skin disease, DPP4’s role may involve impacting immune cell function and the formation of autoantibodies against skin components. These insights into DPP4’s involvement in skin-related immune responses underline its potential as a target for therapeutic interventions in these dermatological conditions.
DPP4 Inhibition
A well-established family of oral hypoglycemic medications for the treatment of T2DM is gliptins, a kind of DPP4 inhibitor (Figure 1). Gliptins are a diverse group of chemicals that interact differently with the active region of enzyme molecules.23 Gliptins are divided into three groups based on their binding mode.24 There are now subsets of DPP4 inhibitors licensed for T2DM, some of which are only available in Korea and Japan. Teneligliptin, for example, was introduced in Japan and had a unique shape with an anchor-lock domain and a J-shape that not only inhibits DPP4 but is also safe for individuals with end-stage renal illness.25
 |
Figure 1 This diagram explains how the use of DPP4 inhibitors. Excess glucagon can be avoided as an increased GLP-1 and 2 stimulates more insulin to reduce glucagon. Hence diabetes can be prevented.26 |
DPP4 inhibitors are classified according to chemical nature into two classes: the peptidomimetics and the non-peptidomimetics. Inhibitors of the peptidomimetic class such as saxagliptin and vildagliptin mimic the dipeptide terminal of DPP4 at the cleavage site of the incretin hormone substrates. By virtue of their peptide character, the peptidomimetic inhibitors have poor pharmacokinetics due to susceptibility to metabolic cleavage by peptidases. On the other hand, the non-peptidomimetic DPP4 inhibitors such as sitagliptin, linagliptin, and alogliptin were developed by high throughput screening and are not structural mimics of the dipeptide end of the enzyme. Non-peptidomimetic inhibitors are therefore generally more metabolically stable than their peptidomimetic counterparts.
DPP4 is a serine protease that cleaves a dipeptide from the N-terminal side of the incretin hormones adjacent to proline or alanine. Cleavage is accomplished by the nucleophilic amino acid ser630. The active site of the enzyme involves two binding pockets (S1 and S2). The S1 pocket harbors the catalytic triad namely, ser630, Asn710, and His740. The S2 pocket is larger and binds inhibitors and not substrates. The key polar interactions within the S2 pocket involve the amino acid residues Glu205, Glu206, and Arg125.23–25
Saxagliptin and vildagliptin are called covalent inhibitors because of their ability to bind covalently to the oxygen of ser630 through their electrophilic nitrile carbon. Such inhibitors bind reversibly but dissociate from the enzyme slowly thereby exhibiting extended duration despite their short t1/2 values. Other key interactions include a hydrogen bond between the carbonyl oxygen and Arg125 and a salt bridge between the amino group and residues Glu205 and Glu206 as illustrated in (Figure 2). Non-peptidomimetic inhibitors are structurally diverse but common key interactions with DPP4 can be observed. A primary or secondary amino group forms a salt bridge with Glu205 and Glu206. An aromatic moiety establishes hydrophobic interactions within the S1 pocket involving the amino acid residues of Tyr631, Val656, Trp659, Tyr662, Tyr666, and Val711. Other substituents engage in interactions with the amino acid residues of Phe357, Ser209, and Arg358 within the S2 pocket as shown in (Figure 2).23–25
 |
Figure 2 Depicts the possible role of DPP4 in COVID-19 infection and how the application of DPP4 inhibitors can minimize the disease severity. |
A Role of DPP4 in Autoimmune Disease
DPP4 in Rheumatoid Arthritis
Rheumatoid arthritis (RA) is an autoimmune disease characterized by joint stiffness, discomfort, loss of function, and swelling. While the exact cause and underlying mechanisms are not fully understood, T-cell activity is believed to play a significant role in the initiation and perpetuation of inflammation in RA.27 Various molecules involved in T cell activation, including CTLA4, CD40, CD28, IL-21, and ILRA/IL2, have been found to be elevated in RA.28 Among these molecules, DPP4 has gained considerable attention as a biomarker for T-cell stimulation in RA.
Interestingly, individuals with RA exhibit significantly lower serum levels of DPP4 compared to the control group,29 and this decrease is inversely correlated with the severity of the disease.30 Notably, there were no significant differences in DPP4 expression on the membrane of circulating monocytes and lymphocytes between healthy controls and patients with pre-clinical RA.30,31 However, DPP4 expression on CD4+ T cells in the peripheral blood was higher in active RA compared to inactive RA and controls. Interestingly, DPP4 expression was found to be higher in synovial fluid T cells in osteoarthritis patients than in RA patients.32 Moreover, DPP4 activity was found to be reduced in fluid mononuclear cells, synovial membrane, and synovial fluid in RA compared to osteoarthritis.33–35 Autoimmunity against DPP4 has also been observed in RA patients, suggesting a potential role for DPP4 in the autoimmune response associated with the disease.36
SDF-1, a substrate of DPP4, is believed to play a crucial role in the recruitment of inflammatory cells by interacting with its receptor, CXCR4. Depletion of DPP4 in mouse models of antigen-induced arthritis led to increased infiltration of CXCR4-positive inflammatory mediators into arthritic joints.37 Blood levels of DPP4 were lower in RA patients compared to osteoarthritis patients and showed an inverse correlation with the inflammatory marker C-reactive protein.37 The number of DPP4+ T lymphocytes in synovial fluid from RA patients was inversely associated with SDF-1 levels.33 The decrease in synovial DPP4 levels in RA may contribute to synovial inflammation through the SDF-1/CXCR4 axis, as the level of synovial SDF-1 was significantly correlated with inflammatory markers (serum C-reactive protein and IL-6) and disease activity score.37,38 These findings highlight the importance of the CXCR4/DPP4/SDF1 axis in synovial inflammation in RA.
In a recent study, exogenous DPP4 or upregulation of DPP4 in synovial fibroblasts was found to reduce the release of proinflammatory cytokines such as IL-6, IL-3, and IL-1β.12 Additionally, DPP4 was found to be significantly expressed in osteoclasts, and its activity was inhibited by the anti-resorptive drug denosumab, suggesting that DPP4 produced by osteoclasts could play a crucial role in bone remodeling and energy metabolism.12 In a mouse model of RA, inhibiting DPP4 was found to enhance the entry of synovial fibroblasts into cartilage cells.39 Moreover, DPP4 inhibitors have been shown to reduce bone loss and improve mechanical bone strength in a rat model of streptozotocin-induced diabetes, possibly by reducing CTX-I-dependent bone resorption.40
Clinical surveillance of patients taking DPP4 inhibitors could provide important insight into the impact of DPP4 proteolytic activity in RA. Several studies have linked DPP4 antagonists to recently diagnosed RA patients.41–43 However, compared to other antihyperglycemic medications, larger-scale population studies12,44–46 could not find a link between RA and DPP4i (Dipeptidyl peptidase-4 inhibitors) use. The specific effects on the development of RA by DPP4 enzymatic inhibition will need to be studied in future randomized controlled studies. A summary of DPP4 function and expression in different immune cells is illustrated in (Table 1).
 |
Table 1 DPP4 is Found in a Variety of Immune Cells. |
DPP4 and SLE (Systemic Lupus Erythematosus)
Many individuals diagnosed with systemic lupus erythematosus (SLE), a chronic autoimmune disease affecting multiple organ systems, develop a characteristic butterfly rash. Initially considered an adaptive immune system disorder, SLE is characterized by high disease activity and the presence of autoreactive antibodies.59
In lupus erythematosus-like syndrome mice, it was observed that serum DPP4 activity was significantly lower compared to the positive control group, suggesting a potential pathogenic role of DPP4 in SLE.5 Clinical studies have demonstrated that patients with Systemic Lupus Erythematosus (SLE), an autoimmune condition, exhibit lower levels of Dipeptidyl peptidase-4 (DPP4) in their serum and peripheral blood mononuclear cells compared to healthy individuals. This finding suggests that decreased DPP4 levels may be associated with the altered immune regulation in SLE, potentially contributing to its pathogenesis. The reduction in DPP4 could serve as a biomarker for disease activity, offering insights into the immune dysfunctions characteristic of SLE. This discovery is significant not only for understanding the disease’s mechanisms but also for exploring new therapeutic approaches that target DPP4 activity to restore immune balance and improve treatment outcomes for SLE patients.5,60,61 The study by Wong et al highlights significant immunological alterations in Systemic Lupus Erythematosus (SLE) patients, characterized by reduced surface expression of Dipeptidyl peptidase-4 (DPP4) on invariant natural killer T (iNKT) cells and CD4+ T cells. This reduction in DPP4 expression may contribute to the dysfunctional immune regulation observed in SLE. Additionally, the study notes an increased activation of T helper 1 (Th1) cells, which are associated with pro-inflammatory responses, and a decrease in the number of circulating iNKT cells, which are crucial for immune regulation and maintaining tolerance. These changes in the immune cell dynamics and function offer insights into the pathophysiology of SLE, suggesting a complex interplay between different immune cells and signaling pathways in the disease’s progression and may inform future therapeutic strategies.62 Additionally, qPCR analysis revealed higher DPP4 mRNA levels in the SLE group compared to healthy control mice, although no significant association with disease activity was found.61
Recent demographic studies conducted in Taiwan and Korea have suggested a potential link between DPP4 inhibitors and a reduced risk of developing SLE.63,64 However, limited therapeutic findings are currently available regarding the specific role of DPP4 suppression in treating SLE.
Systemic Sclerosis and DPP4
Scleroderma, also known as systemic sclerosis (SSc), is a severe autoimmune disease characterized by significant fibrosis affecting the skin and internal organs.65 Early investigations revealed that SSc patients exhibited higher levels of CD4+ DPP4+ T cells in their peripheral blood, and DPP4 activity in T cells was associated with disease severity.66 In comparison to healthy individuals, circulating soluble DPP4 levels were found to be significantly reduced in SSc patients.29 Moreover, in cases of widespread cutaneous SSc, soluble DPP4 levels were lower than in localized cutaneous SSc, suggesting a correlation between DPP4 activity and fibrosis progression in SSc.
Myofibroblasts are the primary producers of collagen in tissue fibrosis67,68 and play a role in tissue stiffness and the development of skin and joint contractures in SSc patients.69,70 As mentioned earlier, DPP4 is implicated in fibroblast activation. DPP4-positive fibroblasts exhibit elevated expression of collagen-1, collagen-3, and fibronectin.71 Additionally, stimulation with recombinant human DPP4 enhances fibronectin synthesis in lung fibroblasts, indicating the involvement of DPP4 in fibroblast activation and tissue remodeling.72 Recent research has shown that DPP4-expressing fibroblasts contribute to collagen accumulation in cutaneous scars, and inhibiting DPP4 reduces scar formation in a rat model of wound healing.15 The SFRP2/DPP4 fibroblast subpopulation serves as the progenitor for fibrogenic fibroblasts in SSc skin, and DPP4-induced NF-kB and SMAD signaling via PAR2 activation leads to dermal fibroblast activation.73,74
The anti-fibrotic effects of DPP4 inhibitors such as vildagliptin and sitagliptin have been demonstrated in a mouse model of bleomycin-induced skin fibrosis, characterized by improved dermal texture, reduced hydroxyproline content, and decreased myofibroblast deposition through the inhibition of TGF-β-induced ERK signaling.74 Vildagliptin has also shown efficacy in reducing fibrosis and inflammation in a mouse model of bleomycin-induced lung fibrosis.75 Although animal studies suggest the potential use of DPP4 inhibitors for SSc treatment, further clinical trials are needed to explore their effects on SSc patients.
DPP4 in Inflammatory Bowel Disease
Inflammatory bowel disease (IBD), including Crohn’s disease (CD) and ulcerative colitis (UC), is a common autoimmune-mediated digestive tract disease. Both CD and UC are characterized by chronic relapsing progressive conditions with the involvement of multiple organs. In patients with IBD, blood levels and enzymatic activity of DPP4, a key enzyme, were found to be significantly lower compared to healthy individuals or those with inactive diseases, and the degree of reduction correlated with disease activity.76–79 Additionally, in a mouse model of colitis, mice with elevated DPP4 levels exhibited higher levels of vasoactive intestinal peptide (VIP), IL-6, and neuropeptide Y (NPY), all of which are substrates of DPP4. During severe inflammatory phases of colitis, these mice showed increased levels of IL-6 and VIP in the brain and colon regions. Interestingly, wild-type mice had lower levels of IL-10 in the brain, whereas DPP4 mice had higher levels, suggesting a potential role of DPP4 in modulating neuroimmune responses during colitis onset.80 In another colitis model, DPP4 mice exhibited increased CD8+ T cells and NKT cells in the spleen, enhanced macrophage infiltration, and greater expression of the NF-kB p65 subunit in the colon mucosa.81
Furthermore, DPP4 cleavage hinders the beneficial effects of GLP-1 and GLP-2, which support T cell growth and behavior and aid in the healing of damaged intestinal epithelia.82,83 Animal studies have demonstrated the protective effects of DPP4 inhibitors in IBD. For instance, the DPP4 inhibitor ER319711 alleviated DSS-induced colitis in mice, as evidenced by reduced weight loss.82,83 Anagliptin therapy facilitated the healing of intestinal mucosal injury in a mouse model of DSS-induced colitis.84 However, clinical evidence regarding the impact of DPP4 inhibitors on IBD is limited and inconsistent. A cross-sectional study revealed a potential association between DPP4 inhibitor use and an elevated risk of Hashimoto’s thyroiditis, psoriasis, and Crohn’s disease.85 Similarly, Abrahami et al found a link between DPP4 inhibitor use and an increased risk of IBD.76 Nonetheless, a real-world investigation involving 895,747 individuals using DPP4 inhibitors or other anti-diabetic drugs did not find any association between DPP4 inhibitor use and a higher prevalence of IBD.86 A meta-analysis of 16 separate studies, employing a random-effects model, reported a non-significant increase in the prevalence of IBD following DPP4 inhibitor exposure. However, further research is required to investigate this potential impact, as the inclusion of a significant study influenced the overall result.87
DPP4 in Autoimmune Diabetes
Type 1 diabetes mellitus (T1DM), known as autoimmune diabetes, primarily affects children and young adults and is characterized by the immune-mediated destruction of pancreatic cells and insulin secretion.88 Serum DPP4 activity has been found to be elevated in individuals with T1DM, and this increase is associated with the duration of diabetes.89–91 While DPP4 inhibitors have been extensively used in the treatment of type 2 diabetes mellitus (T2DM) for over a decade, their efficacy in T1DM remains uncertain. Clinical trials investigating the impact of DPP4 inhibition on reducing HbA1c levels in T1DM patients have yielded inconsistent results.92–94
Despite the variability in clinical studies, preclinical research has focused on understanding the role of DPP4 in inflammation and the development of T1DM. T1DM is characterized by imbalances in Th17/Treg, Th1, and Th2 immune responses. Inhibition of DPP4 has been shown to increase Th2 cytokines, restore the balance between Tregs and Th17 cells, and reduce Th1 response.95 Additionally, in the pancreas and adipose tissue, GLP-1, a substrate of DPP4, has demonstrated anti-inflammatory properties by reducing the production of inflammatory cytokines and immune cell infiltration.96 Therefore, DPP4 inhibition may contribute to the anti-inflammatory processes in the pancreas. In NOD mice, treatment with the DPP4 inhibitor MK0431 prolonged the survival of islet grafts and reduced CD4+ T cell migration into the pancreas through the cAMP/PKA/Rac1 activation pathway.97 Another T1DM mouse model induced by low-dose streptozotocin showed decreased pancreatic inflammation with sitagliptin treatment, accompanied by an increase in CD4+ CD25hiFoxp3+ T cells and a reduction in inflammatory cells.98 Hence, preclinical research suggests that DPP4 inhibition may ameliorate T1DM through pathways involving both incretin-based mechanisms and anti-inflammatory effects.
A Role of DPP4 Inhibitors in Diabetic Kidney Disease
Cardiovascular Effects of DPP4 Inhibition
The CARMELINA trial, identified by its ClinicalTrials.gov Identifier NCT01897532, is currently underway with the enrollment of over 8000 patients using linagliptin. This clinical trial focuses on evaluating renal and cardiovascular endpoints in Type 2 Diabetes patients. The primary outcome being assessed is the time until the occurrence of the first cardiovascular mortality, non-fatal myocardial infarction, non-fatal stroke, or hospitalization for unstable angina pectoris. The development of renal failure is defined by the presence of end-stage renal disease, renal mortality, or a sustained decline in estimated glomerular filtration rate (eGFR) of 50% or more.
A recent randomized placebo-controlled trial published by Scirica et al, known as SAVOR-TIMI 53, found no significant difference in ischemic events but did observe higher hospitalizations for heart failure with saxagliptin.99 Despite preclinical studies and post hoc analyses of clinical trials suggesting a favorable cardiovascular risk profile for DPP4 inhibitors, this particular trial did not show such benefits. The study included 16,492 individuals with Type 2 diabetes who had either experienced a cardiovascular incident or were at risk for one, and they were followed for a median duration of 2.1 years.
Another study, the EXAMINE trial, investigated alogliptin medication in individuals with Type 2 diabetes who had experienced acute coronary syndrome and revealed no increase in major adverse cardiovascular events.100 This randomized placebo-controlled trial included 5380 patients and had a median follow-up of 18 months. The US Food and Drug Administration’s (FDA) independent advisory panel has recommended modifying the labeling of these two drugs to improve their cardiovascular safety profile.26
The TECOS trial, a major multicenter clinical study, compared saxagliptin to placebo in 14,671 individuals with Type 2 diabetes and established cardiovascular disease. Saxagliptin was administered in addition to the patients’ standard treatment.101 Additionally, the ongoing CAROLINA study (Cardiovascular Outcome Study of Linagliptin Against Glimepiride in Type 2 Diabetes Patients) is investigating the cardiovascular outcomes of linagliptin.101
It is essential to examine the cardiovascular profile of GLP-1 analogs as well since DPP4 inhibitors increase GLP-1 levels. A large retrospective study by Best et al compared patients treated with exenatide, a GLP-1 analog, to those treated with other glucose-lowering drugs, and found a 20% decrease in cardiovascular events.102 However, it should be noted that publications demonstrating the cardiovascular benefits of GLP-1 or its analogs have utilized supraphysiological dosages. DPP4 inhibitors, which work within the physiological range to enhance GLP-1 activity, may have additional pleiotropic effects beyond their GLP-1 activities (Figure 3). Examples of DPP4 inhibitors currently in clinical use include saxagliptin, vildagliptin, alogliptin, linagliptin, and sitagliptin. These inhibitors differ structurally in terms of pharmacokinetics, thus ongoing trials are crucial as no class effect can be assumed.
Lastly, an important ongoing clinical trial is examining hard renal endpoints. DPP4 inhibitors have shown potential as antifibrotic and anti-inflammatory agents in kidney disease. The impact of this class on the risk of cardiac failure is still under investigation.
 |
Figure 3 Explains the assumed structure of DPP4 according to different binding domains with ADA, SARS-CoV-2, and Gliptins (DPP4 inhibitor). |
DPP4 and the Kidney
The enzymatic activities of DPP4 in the kidney were investigated by analyzing renal tissue from DPP4 and DPP4/mice using liquid chromatography-mass spectrometry-based peptidomics. Over the past few decades, researchers have identified 10 peptides that are regulated by DPP4 in in vivo studies.26,103 Further research utilizing brush boundary membranes has shown that the aminopeptidase function of DPP4 is crucial for the generation of DPP4 precursors. This indicates that DPP4 plays a vital role in the extracellular protein metabolism of the kidney, particularly in the breakdown and hydrolysis of proline-containing peptides.104 Since DPP4 is located on the brush boundary (apical side) of renal tubular cells, inhibiting its activity is expected to affect the regulation and breakdown of peptides in the fluid, thereby influencing the morphology and activity of tubular cells in diabetes. Identifying the functionally important renal substrates of DPP4 is necessary to understand and predict the long-term effects of DPP4 inhibition on the kidney in patients with diabetes. However, the methods required to confirm specific receptors are limited, as antibodies need to be able to distinguish between cleaved and non-cleaved peptides, which differ by only two amino acids at the N-terminal end.
Transforming growth factor beta 1 (TGF1) plays a significant role in fibrosis associated with diabetic kidney disease. In human kidney proximal tubular cells,26 the DPP4 inhibitor linagliptin reduces the transcription and expression of phosphorylated smad2 and fibronectin by inhibiting the activation of TGF1 induced by high glucose. Linagliptin’s mechanism appears to involve preventing latent TGF1 from becoming activated, as it did not affect the total secreted TGF1 induced by high glucose. TGF1 is initially released in an inactive state and requires a complex interaction of soluble signaling molecules to activate and release it from the latency-related peptide (LAP).105 Once released from LAP, TGF1 can bind to its receptor and activate cell signaling through the Smad pathway. It has been observed that high glucose triggers an interaction between CIM6PR and DPP4, and linagliptin modulates this interaction in this context. The binding sites of CIM6PR become saturated when there is a significant amount of M6P, resulting in a reduced connection between CIM6PR and DPP4. This suggests that the interaction is mediated through an M6P residue on the DPP4 molecule. Notably, linagliptin’s ability to block this interaction in an in vitro system lacking GLP-1 suggests an M6P-mediated approach independent of GLP-1 (unpublished data). These findings contribute to the growing body of research indicating that DPP4 inhibition possesses antifibrotic properties beyond its proteolytic effects. It is important to consider the potential confounding effects of concurrent glucose reduction with DPP4 inhibitors when analyzing preclinical investigations. Two studies using vildagliptin and sitagliptin have examined the impact of DPP4 inhibition on the kidneys of hyperglycemic animal models, demonstrating renoprotective effects. Furthermore, diabetic rats treated with DPP4 inhibitors showed lower HbA1c levels compared to diabetic control animals.106,107 Hence, it is challenging to attribute the renal effects of DPP4 inhibitors solely to glucose reduction in in vivo trials.
In a model of Type 1 diabetic nephropathy, Kawasaki et al explored the antifibrotic properties of linagliptin and discovered that, after 4 weeks, it decreased diabetic kidney fibrosis, which was associated with the repair of microRNA29s and the suppression of EndMT.26 DPP4inh does not significantly lower blood sugar levels since there is no significant insulin response mediated by GLP-1, it can be evaluated independently of glycemic control in an insulin-deficient model of diabetes. As a result, the current findings reveal that linagliptin has renal benefits irrespective of glucose control.
DPP4 inhibitors’ anti-fibrotic activities have also been demonstrated in various renal fibrosis models, such as the unilateral ureteral obstruction (UUO) model.108 LC15-0444 (a DPP4inh) decreased inflammatory and fibrotic indicators like TGF1, toll-like receptor 4, HMGB1, NADPH oxidase 4, nuclear factor kappa B, and modified Smad2/3 in this TGF-driven model. However, there needed to be specific mechanistic data to interpret the reports in this investigation.
DPP4 in Diabetic Nephropathy
Research suggests that DPP4 may play a role in the receptor axis for advanced glycation end products (RAGE), which is crucial for the development of diabetic nephropathy. In animal models of Type 1 diabetic nephropathy, both DPP4 deficiency and linagliptin treatment have been shown to reduce kidney damage.109,110 The proposed mechanism involves a decrease in endothelial cell oxidative stress. Endothelial dysfunction is a characteristic feature of diabetic kidney disease. Studies have shown that DPP4 interacts with integrin 1 in endothelial cells. Shi et al demonstrated that the deletion of integrin 1 prevented the endothelial-to-mesenchymal transition associated with DPP4. Furthermore, DPP4 or integrin deficiency inhibited the TGF2-stimulated formation of heterodimers of TGF receptors. The interaction between DPP4 and integrin 1 also promoted the expression of vascular endothelial growth factor receptor (VEGF-R) 1 while suppressing VEGF-R2 levels.26 Linagliptin was found to reduce the direct interaction between integrin and DPP4 induced by TGF2. These findings suggest that the non-proteolytic interaction between DPP4 and integrin 1 is important in diabetic kidney fibrosis.
DPP4 Inhibitors and Mesenteric Arteries
Mechanistic studies have been conducted to investigate whether the effects of DPP4 inhibitors on glucose-induced endothelial impairment are associated or independent of GLP-1/GLP-1R. These studies found that DPP4 inhibitors, including linagliptin, have direct effects on mesenteric arteries regardless of their impact on glucose levels. The mechanism behind this effect has been investigated using direct reactive quenching and has been linked to the antioxidant activity of DPP4 inhibitors.26,111 Additionally, DPP4 inhibitors have been shown to possess anti-inflammatory properties. Alogliptin, for example, reduced macrophage infiltration in a non-diabetic animal model of glomerular damage, while PKF275-055 decreased nuclear factor kappa B activity and macrophage infiltration in a type 1 renal disease model.112,113
Although DPP4 inhibitors have similar abilities to improve glucose control, their pharmacokinetic and pharmacodynamic profiles differ significantly due to their binding to different sites on DPP4.114 While all DPP4 inhibitors can inhibit DPP4’s enzymatic function (cleaving), their capacity to alter DPP4’s co-stimulatory/receptor qualities varies.26,115 It is also important to consider the mechanism of clearance of DPP4 inhibitors.116 The strong expression of DPP4 on the proximal tubular brush border suggests that drugs primarily eliminated by the kidneys may have better access to tubular DPP4.
Role of DPP4 in COVID-19 Disease Severity
The clinical signs of COVID-19 are indicative of SARS-propensity Cov-2’s to infect several organs and their cells, which mimics the activation of viral fusion proteins.117 Although ACE2 is exhibited in enterocytes and pneumocytes of the alimentary canal,118 according to studies, the alveolar type 2 cells’ appearance is subdued when compared to other cells that express ACE2, indicating that SARS-CoV-2 utilizes co-receptors Various safe remedies are being considered to treat COVID-19 including phage-vaccine development, use of DPP4, application of herbs etc.119–121 Various safe remedies are being considered to treat COVID-19 including phage-vaccine development, use of DPP4, and application of Chloroquine phosphate, but not finding ample proof suggesting the effective consequences, Chloroquine use was stopped.122 ACE2 prevalence is prominent in the intestine and kidney and is connected to how many additional proteolytic enzymes coronaviruses employ as entry targets.123 DPP4 was discovered to be the first genotype to be grouped with ACE2, and because modeling reveals a close connection between DPP4 and SARS-CoV-2 S1,124 DPP4 region localization is essential. DPP4 is most highly expressed in the liver, salivary gland, kidney, and seminal fluid in human bodies (Figure 4). Enterocytes, particularly blood vessel capillaries, lung epithelium, immune cells, and intestinal enterocytes all have high DPP4 levels (activated myeloid cells, T, B, and natural killer cells).125
 |
Figure 4 Gliptins (DPP4 inhibitors) available in the drug market. |
In vitro, DPP4 activity is selectively produced on the surface of senescent, but non-regenerating, human diploid fibroblasts and is greater in older individuals than in younger individuals.126 DPP4 is largely present in alveolar type 2 cells in the epithelial tissue of the lower respiratory tract,126 but it is also found in alveolar macrophages in individuals with COPD and cystic fibrosis.127,128 Upregulation of DPP4 in type I pneumocytes may be a critical factor in MERS-CoV and SARS-CoV-2 infection severity (Figure 4).129
DPP4’s non-enzymatic activity as a signaling and coupling protein and its broad prevalence on myeloid cells, blood capillaries, myocardium, and imply a function in inflammation across a wide variety of animals, angiogenesis, and cardiovascular control.130,131
In patients with diabetes and obesity, DPP4 may also indirectly enhance COVID-19 development. It could explain why COVID-19 in these people has a more severe course, with higher vulnerability to cytokine storms and fulminant multi-organ failure. It has been demonstrated utilizing genetically engineered mouse models that a high-fat meal induces hyperglycemia and hyperinsulinemia, like human T2D. Still, mice missing the DPP4 gene were safeguarded against obesity and insulin resistance. Notably, compared to non-obese animals lacking human DPP4, diabetic mice challenged with MERS-CoV experienced a more severe course of the illness and a more extended period of recovery.132,133 The age of infected individuals is the next medically significant risk factor after T2D and overweight that is connected to a potential relationship between DPP4 and vulnerability to SARS-CoV-2 illness. Surprisingly, in vitro research found that growing diploid human fibroblasts did not express DPP4, whereas senescent human fibroblasts did.134 The DPP4 molecule was more active in older than younger people.135 The pulmonary epithelium of smokers and individuals with chronic obstructive pulmonary disease showed a considerable rise in DPP4 mRNA and protein expression, indicating that these people are more vulnerable to illness, particularly with MERS-CoV and probably also with SSARS-CoV-2 the second, though, has to be confirmed.136
DPP4 Inhibitors and COVID-19
Based on the factors mentioned, it has been proposed that DPP4 inhibitors could be beneficial for COVID-19 patients, potentially by preventing the entry of the virus.137,138 Structural models of DPP4 and its inhibitors, as well as the viral spike proteins of MERS-CoV and SARS-CoV-2, indicate that the binding sites for DPP4 inhibitors do not overlap with those for viral spike proteins.139,140 It is hypothesized that DPP4 inhibitors may modulate the endocytic cascade of SARS-CoV-2 by interacting with caveolin-1, a scaffold protein required for endosome formation.141 Recent evidence from a large-scale prospective cohort study involving 8.3 million individuals suggests that ACE inhibitors, which are often used in combination with DPP4 inhibitors, are safe and may even reduce the incidence of severe COVID-19 and its complications.142
However, it is crucial to assess the overall reliability and risk-benefit profile of using DPP4 inhibitors, particularly in the context of COVID-19 patients. There is a need to consider the possibility that DPP4 inhibitors may impact the immune response and potentially increase the risk of infections in general. The widespread distribution of DPP4 in various types of cells, especially immune cells, may contribute to this risk. The effects of DPP4 inhibitors on the immune system are not well understood.143 In studies involving healthy individuals, a 28-day treatment with sitagliptin did not affect lymphocyte and T cell subsets, plasma chemokine/cytokine concentrations, or cytokine release upon stimulation of peripheral blood mononuclear cells.144 A meta-analysis indicated that the use of DPP4 inhibitors was associated with a similar risk of infections, including respiratory diseases, compared to control groups.145–147 The anti-inflammatory properties of sitagliptin may help COVID-19 patients in avoiding and managing cytokine storms when administered alone or in combination with other medications such as dexamethasone, tocilizumab, or anti-IL-6 receptor antibodies.135,148 Clinical studies have shown that sitagliptin, when added to standard-of-care medication, was associated with significant clinical improvement and reduced mortality compared to standard-of-care alone.149
It is important to note that while there is evidence suggesting the potential benefits of using DPP4 inhibitors for COVID-19, further research and clinical trials are needed to establish their effectiveness and safety in this specific context. Medical decisions regarding the use of these medications should be made on an individual basis, considering the patient’s condition, medical history, and potential risks and benefits.
DPP4, Immunomodulation, and Infection
DPP4 interacts with ligands, notably ADA, to provide noncatalytic activities.150 DPP4 is a costimulator for T cell activation that binds to ADA and boosts T cell proliferation without ADA binding.150 Because adenosine is an effective inhibitor of T cell multiplication, increasing DPP4 activity can enhance T cell proliferation. DPP4 expression in dendritic/macrophage cell populations from visceral adipose tissue is higher in obese people, stimulating inflammatory response in obesity via dealing with ADA.54 As a result, elevated DPP4 in circumstances like obesity-induced MS (metabolic syndrome) leads to a failure to resolve inflammatory response and prolonged subclinical immunological stimulation. Some coronavirus diseases may find a “perfect habitat” amid this metabolic/immune imbalance. DPP4 consists mainly of three domains based on their binding sites (Figure 5).
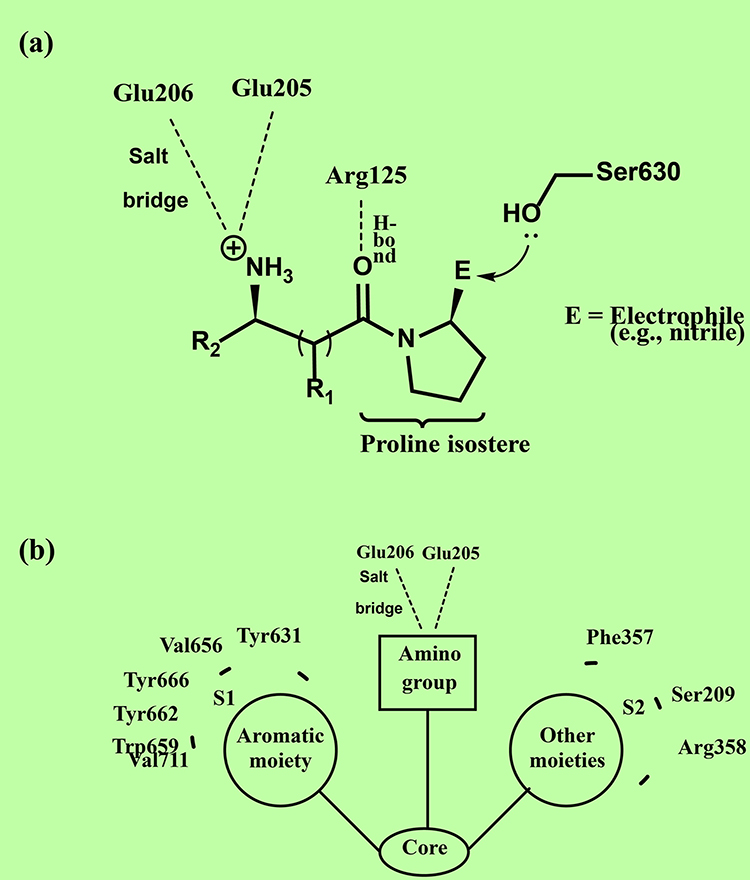 |
Figure 5 Chemistry of Gliptins. (a) Binding of covalent inhibitors to DPP4, (b) Common binding of non-peptidomimetic inhibitors to DPP4. |
Galidesivir (BCX4430), an adenosine nucleoside derivative with wide-ranging action towards RNA viruses, is now being tested in COVID-19 (ClinicalTrials.gov Identifier: NCT03891420) that was effective towards flaviviruses like Zika.151 It works by blocking viral RNA-dependent RNA polymerase, which is essential for viral replication. It’s unclear how such a treatment would react with adenosine’s immunostimulatory, primarily anti-inflammatory, effects in distinct immune cell types,152 or how elevated DPP4 activity and prolonged subclinical inflammation in obesity-induced MS would impact the therapeutic outcome.
Also, other pathogens have been linked to DPP4. DPP4 truncates the amino-terminal of the chemokine CXCL10 (also known as IP-10) in chronic hepatitis C virus (HCV) patients, resulting in an antagonist version of the chemokine, which can regulate immune responses through chemokine receptor antagonism.153 Truncated CXCL10 antagonism may also be a significant regulatory mechanism in malignancies154 and tuberculosis pathological locations.153
Covid-19 Comorbidities, Disease Manifestations, and Severity
The severity of COVID-19 is influenced by various factors, including comorbidities and age. In one study, diabetes mellitus (DM), hypertension, and cardio-cerebrovascular diseases were found to be 2–3 times higher in intensive care unit (ICU)/severe cases compared to non-ICU/severe cases.155 Individuals with diabetes and hyperglycemia had significantly higher levels of inflammatory mediators, such as IL-6, C-reactive protein, serum ferritin, and D-dimer, suggesting increased vulnerability to inflammation storms and higher mortality.156,157 Considering that approximately 425 million people worldwide have diabetes, diabetes therapy options are crucial.
Obesity also plays a significant role in COVID-19 severity, with severe obesity (BMI >35 kg/m2) and obesity (BMI >30 kg/m2) observed in a significant proportion of COVID-19 patients.158 Age, on the other hand, was not associated with diabetes, hypertension, or the need for invasive mechanical ventilation but was correlated with obesity (P<0.05) and male sex (P<0.05). There is a link between higher DPP4 activity and expression and metabolic syndrome (MS) and obesity.159–162 This suggests a direct metabolic and endocrine relationship to the viral pathogenesis, emphasizing the need for proper treatment in COVID-19 patients with these comorbidities.163
DPP4 is believed to be a co-receptor for the entrance of the SARS-CoV-2 virus and as DPP4 dysregulation is associated with disease severity in COVID-19, it is important to learn more about how DPP4 inhibition affects the course of the illness.164
Notwithstanding the fact that the DPP4 inhibitors do not attach to the SARS-anticipated CoV-2’s receptor binding site, studies on the DPP4 inhibitors over the last 20 years have given a depth of knowledge about the biology and chemistry of DPP4 and its binding relationships;24 this information could be used to build new anti-coronavirus medications. A medication may be developed to protect the human viral entrance protein(s) from the virus by binding to the human protein with an antibody or peptide. As a unique possible therapy for T2DM (Type-2 Diabetes Mellitus), a therapeutic vaccination targeting human DPP4 has already been produced.165 Pang et al selected three suitable DPP4 regions as vaccine candidate targets, conjugated these peptides to the adjuvant keyhole limpet hemocyanin, which presents a variety of T-cell epitopes to induce helper T-cell responses, and demonstrated its potency in enhancing the insulin-dependent phenotype in mouse models.
COVID-19 may benefit from modified DPP4 activity in comorbid diseases in ways that are unknown at the moment. Gliptins may affect the severity of COVID-19, although various DPP4 inhibitors may potentially have varied effects (s). A comparison of diabetes (and other) treatments for those with serious and non-serious diseases would be fascinating. In Japan, for example, most T2DM patients (1410/2666 patients) received a DPP4 inhibitor as the beginning treatment, a conclusion affected by adulthood.166
Multiple gliptins have been found in vitro models to have several actions that may be useful in COVID-19 disease. In an immediate model of an inflammatory process, saxagliptin and vildagliptin have anti-inflammatory efficacy similar to aspirin.167 Saxagliptin’s DPP4 inhibition can reduce the levels of CRP, tumor necrosis factor, IL-1, IL-18, and IL-6 in the blood and attenuate DM-induced activation of the NLRP3 inflammasome.168 Evidence of interplay between DPP4 and the renin-angiotensin system in the pathophysiology of cardiorenal syndromes suggests that gliptins are cardioprotective and may delay the onset of circulatory impairment in chronic kidney disease.169 The treatment of NAFLD, the hepatic symptom of the obesity-induced metabolic syndrome, may also involve gliptins.170,171
Gliptins have been demonstrated to have a broad and powerful anti-inflammatory therapy in humans; for example, sitagliptin therapy lowers plasma levels of IL-6 and C-reactive protein and decreases CD26/DPP4 mRNA expression in mononuclear cells by 16% after 2 hours after only one dosage (CRP).172 Mathematical simulations suggest that reducing DPP4 expression might decrease the propagation of the MERS-CoV infection.173 Hence, commercially available DPP4 inhibitors might be modified to treat COVID-19 patients, especially those with significant hyperglycemia and virally produced hyperinflammation symptoms, if DPP4 expression enhances SARS-CoV-2 infection,174,175 or cardio-renal issues.112
DPP4 inhibitors have been thoroughly reviewed by the FDA Adverse Event Reporting System (FAERS) to highlight the associated adverse effects. The ratio of DPP4 inhibitors’ adverse effects is common in the gastrointestinal tract and musculoskeletal system. These side effects are prevalent in non-insulin antidiabetics. The above study reported an understanding of the safety profiles and few side effects of DPP4 inhibitors however, it still recommended the further validation and interpretation of these findings.176 It is suggested that thorough studies on the side effects of DPP4 inhibitors need to be evaluated to provide a better understanding of DPP4 inhibitors’ mediated side effects.
Prospects of DPP4 Inhibitors in the Era of COVID-19 Therapeutics
In the context of COVID-19, it is crucial to conduct further research to fully understand the effectiveness, safety, and specific effects of DPP4 inhibitors. While there is evidence suggesting the potential benefits of DPP4 inhibitors in managing the disease, especially in individuals with comorbidities such as diabetes and obesity, comprehensive studies are needed to validate these findings.
Anti-inflammatory effects: Gliptins have demonstrated anti-inflammatory properties, which can be beneficial in managing the cytokine storms associated with severe COVID-19. By reducing the levels of inflammatory mediators, such as IL-6 and C-reactive protein, gliptins may help modulate the immune response and mitigate the excessive inflammation seen in severe cases.
Immunomodulatory effects: Gliptins have been found to affect immune cell function and cytokine production. These immunomodulatory effects could potentially help regulate the immune response to SARS-CoV-2 infection, preventing immune system dysregulation and excessive tissue damage.
Co-receptor inhibition: DPP4 has been identified as a co-receptor for SARS-CoV-2 entry into host cells. By inhibiting DPP4 activity, gliptins may interfere with the virus’s ability to infect cells, potentially reducing viral load and disease severity.
Combination therapies: Gliptins have been studied in combination with other COVID-19 therapeutics, such as dexamethasone and tocilizumab. Combining gliptins with these drugs may have synergistic effects in managing inflammation and improving clinical outcomes in severe cases.
Safety and tolerability: Gliptins are generally well-tolerated and have a favorable safety profile, as they have been extensively used for the treatment of diabetes. This existing safety data could facilitate their repurposing of COVID-19 treatment.
Personalized medicine: As gliptins act on the DPP4 pathway, which is involved in various physiological processes, including glucose metabolism and immune regulation, their use in COVID-19 could be tailored to individual patient profiles. Identifying specific patient subgroups that may benefit the most from gliptin therapy could enhance treatment efficacy.
Drug repurposing and new drug development: Gliptins are already approved for the treatment of diabetes, which facilitates their repurposing for COVID-19. Additionally, the knowledge gained from studying gliptins’ interactions with DPP4 could aid in the development of new drugs that target specific aspects of the SARS-CoV-2 infection process.
Additionally, it is important to consider the potential side effects of DPP4 inhibitors. FAERS has identified gastrointestinal and musculoskeletal system-related adverse effects as common with DPP4 inhibitors.176 To provide a better understanding of the safety profiles and potential side effects of these medications, thorough studies are required.
Furthermore, the interaction between DPP4 inhibitors and the SARS-CoV-2 virus needs to be explored in more detail. While gliptins do not bind to the receptor-binding site of the virus, their impact on DPP4 dysregulation and the disease severity of COVID-19 requires further investigation. The knowledge gained from studying DPP4 biology and chemistry over the years can potentially contribute to the development of new anti-coronavirus medications.
In summary, DPP4 inhibitors have shown promise in the management of COVID-19, particularly in individuals with specific comorbidities. However, comprehensive research is necessary to determine their effectiveness, safety, and potential side effects in the context of the disease. Understanding the interaction between DPP4 inhibitors and the SARS-CoV-2 virus can aid in the development of targeted therapies.
Conclusion
The impact of DPP4 expression in autoimmune diseases and COVID-19 infection is an area of ongoing research. While DPP4 is not a specific biomarker for any autoimmune disease, its widespread expression makes it less useful as a precise marker. The exact mechanisms by which DPP4 plays a role in disease onset and severity are still being investigated. However, there is evidence suggesting a connection between increased DPP4 concentration and disease severity. This observation has led to the exploration of DPP4 inhibitors as potential treatments for various diseases. DPP4 inhibitors are a diverse group of compounds that have shown promising results in reducing the severity of several autoimmune diseases. DPP4 itself consists of multiple domains that are involved in disease complications and provide binding sites for inhibitors. These inhibitors exert their effects through various mechanisms, including immunomodulatory and anti-inflammatory activities. Dipeptidyl-peptidase 4 (DPP4) inhibitors present a dual aspect in their impact on autoimmune and inflammatory diseases. While they have shown potential benefits by modulating the immune system, which can help in reducing inflammation and autoimmune reactions, there is also evidence that these drugs might induce or exacerbate such conditions. This paradoxical effect is due to the complex role of DPP4 in immune regulation its inhibition can both suppress overactive immune responses beneficial in autoimmune diseases, and potentially disrupt immune balance, leading to new or worsened autoimmune and inflammatory disorders. This highlights the need for personalized approaches and careful monitoring in the use of DPP4 inhibitors, particularly in individuals with predispositions to these conditions.
The specific functions of DPP4 inhibitors are still being elucidated, but they have shown encouraging results in several studies. Further research is needed to fully understand the role of DPP4 and its inhibitors in autoimmune diseases and COVID-19. Continued investigation into the mechanisms and potential therapeutic applications of DPP4 inhibitors may provide valuable insights for the management of these conditions.
Acknowledgments
The authors are grateful to their respective institutes.
Funding
The author Simona Cavalu acknowledges the support from University of Oradea, Romania, within the Grants Competition “Scientific Research of Excellence Related to Priority Areas with Capitalization through Technology Transfer: INO-TRANSFER-UO”, Project No. 324/2021.
Disclosure
The authors declare no conflicts of interest in this work.
References
1. Scanlan MJ, Raj B, Calvo B, et al. Molecular cloning of fibroblast activation protein alpha, a member of the serine protease family selectively expressed in stromal fibroblasts of epithelial cancers. Proc Natl Acad Sci. 1994;91:5657–5661.
2. Panevin TS, Zhelyabina OV, Eliseev MS, Shestakova MV. Urate-lowering effects of dipeptidyl peptidase-4 inhibitors. Diabetes mellitus. 2020;23:349–356. doi:10.14341/DM12412
3. De Meester I, Korom S, Van Damme J, Scharpé S. CD26, let it cut or cut it down. Immunol Today. 1999;20:367–375. doi:10.1016/S0167-5699(99)01486-3
4. Thul PJ, Åkesson L, Wiking M, et al. A subcellular map of the human proteome. Science. 2017;356:eaal3321. doi:10.1126/science.aal3321
5. Busso N, Wagtmann N, Herling C, et al. Circulating CD26 is negatively associated with inflammation in human and experimental arthritis. Am J Pathol. 2005;166:433–442. doi:10.1016/S0002-9440(10)62266-3
6. Klemann C, Wagner L, Stephan M, von Hörsten S. Cut to the chase: a review of CD26/dipeptidyl peptidase-4’s (DPP4) entanglement in the immune system. Clin Exp Immunol. 2016;185:1–21. doi:10.1111/cei.12781
7. Vanderheyden M, Bartunek J, Goethals M, et al. Dipeptidyl-peptidase IV and B-type natriuretic peptide. From bench to bedside. Clin Chem Lab Med. 2009;47:248–252. doi:10.1515/CCLM.2009.065
8. Raj VS, Mou H, Smits SL, et al. Dipeptidyl peptidase 4 is a functional receptor for the emerging human coronavirus-EMC. Nature. 2013;495:251–254. doi:10.1038/nature12005
9. Wang N, Shi X, Jiang L, et al. Structure of MERS-CoV spike receptor-binding domain complexed with human receptor DPP4. Cell Res. 2013;23:986–993. doi:10.1038/cr.2013.92
10. Lu G, Hu Y, Wang Q, et al. Molecular basis of binding between novel human coronavirus MERS-CoV and its receptor CD26. Nature. 2013;500:227–231. doi:10.1038/nature12328
11. Li K, Wohlford-Lenane C, Bartlett JA, McCray PB. Inter-individual variation in receptor expression influences MERS-CoV Infection and immune responses in airway epithelia. Front Public Health. 2021;2021:9.
12. Huang J, Liu X, Wei Y, et al. Emerging role of dipeptidyl peptidase-4 in autoimmune disease. Front Immunol. 2022;2022:13.
13. Cashin K, Gray LR, Jakobsen MR, Sterjovski J, Churchill MJ, Gorry PR. CoRSeq V3-C: a novel HIV-1 subtype C specific V3 sequence based coreceptor usage prediction algorithm. Retrovirology. 2013;10:1–10. doi:10.1186/1742-4690-10-24
14. Wu L, Paxton WA, Kassam N, et al. CCR5 levels and expression pattern correlate with infectability by macrophage-tropic HIV-1, in vitro. J Exp Med. 1997;185:1681–1692. doi:10.1084/jem.185.9.1681
15. Rinkevich Y, Walmsley GG, Hu MS, et al. Identification and isolation of a dermal lineage with intrinsic fibrogenic potential. Science. 2015;348:aaa2151. doi:10.1126/science.aaa2151
16. Oh M-D, Park WB, Choe PG, et al. Viral load kinetics of MERS coronavirus infection. N Engl J Med. 2016;375:1303–1305. doi:10.1056/NEJMc1511695
17. Abbott CA, McCaughan GW, Levy MT, Church WB, Gorrell MD. Binding to human dipeptidyl peptidase IV by adenosine deaminase and antibodies that inhibit ligand binding involves overlapping, discontinuous sites on a predicted β propeller domain. Eur J Biochem. 1999;266:798–810. doi:10.1046/j.1432-1327.1999.00902.x
18. Yu D, Slaitini L, Gysbers V, et al. Soluble CD26/dipeptidyl peptidase IV enhances human lymphocyte proliferation in vitro independent of dipeptidyl peptidase enzyme activity and adenosine deaminase binding. Scand J Immunol. 2011;73:102–111. doi:10.1111/j.1365-3083.2010.02488.x
19. Vankadari N, Wilce JA. Emerging COVID-19 coronavirus: glycan shield and structure prediction of spike glycoprotein and its interaction with human CD26. Emerging Microbes Infect. 2020;9:601–604. doi:10.1080/22221751.2020.1739565
20. Sesti G, Avogaro A, Belcastro S, et al. Ten years of experience with DPP-4 inhibitors for the treatment of type 2 diabetes mellitus. Acta Diabetologica. 2019;56:605–617. doi:10.1007/s00592-018-1271-3
21. Tomovic K, Lazarevic J, Kocic G, Deljanin‐Ilic M, Anderluh M, Smelcerovic A. Mechanisms and pathways of anti‐inflammatory activity of DPP‐4 inhibitors in cardiovascular and renal protection. Med Res Rev. 2019;39:404–422. doi:10.1002/med.21513
22. Lu Z, Doulabi BZ, Wuisman P, Bank R, Helder M. Differentiation of adipose stem cells by nucleus pulposus cells: configuration effect. Biochem Biophys Res Commun. 2007;359:991–996. doi:10.1016/j.bbrc.2007.06.002
23. Gallwitz B. Clinical use of DPP-4 inhibitors. Front Endocrinol. 2019;389. doi:10.3389/fendo.2019.00389
24. Kushwaha R, Haq W, Katti S. Sixteen-years of clinically relevant dipeptidyl peptidase-IV (DPP-IV) inhibitors for treatment of type-2 diabetes: a perspective. Curr Med Chem. 2014;21:4013–4045. doi:10.2174/0929867321666140915143309
25. Li X, Huang X, Bai C, et al. Efficacy and safety of teneligliptin in patients with type 2 diabetes mellitus: a systematic review and meta-analysis of randomized controlled trials. Front Pharmacol. 2018;9:449. doi:10.3389/fphar.2018.00449
26. Panchapakesan U, Pollock C. The role of dipeptidyl peptidase–4 inhibitors in diabetic kidney disease. Front Immunol. 2015;6:443. doi:10.3389/fimmu.2015.00443
27. Panayi G, Lanchbury J, Kingsley G. The importance of the T cell in initiating and maintaining the chronic synovitis of rheumatoid arthritis. Arthritis Rheum. 1992;35:729–735. doi:10.1002/art.1780350702
28. Novikov A, Aleksandrova Е, Lukina G. Serum cytokine profile in early and established rheumatoid arthritis. Alman Clin Med. 2019;47:393–399. doi:10.18786/2072-0505-2019-47-058
29. Sinnathurai P, Lau W, Vieira de Ribeiro AJ, et al. Circulating fibroblast activation protein and dipeptidyl peptidase 4 in rheumatoid arthritis and systemic sclerosis. Internat J Rheum Dis. 2018;21:1915–1923. doi:10.1111/1756-185X.13031
30. Cordero OJ, Salgado FJ, Mera-Varela A, Nogueira M. Serum interleukin-12, interleukin-15, soluble CD26, and adenosine deaminase in patients with rheumatoid arthritis. Rheumatol Internat. 2001;21:69–74. doi:10.1007/s002960100134
31. Grujic M, Matic IZ, Crnogorac MD, et al. Activity and expression of dipeptidyl peptidase IV on peripheral blood mononuclear cells in patients with early steroid and disease modifying antirheumatic drugs naïve rheumatoid arthritis. Clin Chem Laborat Med. 2017;55:73–81. doi:10.1515/cclm-2015-1279
32. Muscat C, Bertotto A, Agea E, et al. Expression and functional role of 1F7 (CD26) antigen on peripheral blood and synovial fluid T cells in rheumatoid arthritis patients. Clin Exp Immunol. 1994;98:252–256. doi:10.1111/j.1365-2249.1994.tb06134.x
33. Sromova L, Mareckova H, Sedova L, Balaziova E, Sedo A. Dipeptidyl peptidase-IV in synovial fluid and in synovial fluid mononuclear cells of patients with rheumatoid arthritis. Clin Chim Acta. 2010;411:1046–1050. doi:10.1016/j.cca.2010.03.034
34. Buljevic S, Detel D, Pucar LB, et al. Levels of dipeptidyl peptidase IV/CD26 substrates neuropeptide Y and vasoactive intestinal peptide in rheumatoid arthritis patients. Rheumatol Internat. 2013;33:2867–2874. doi:10.1007/s00296-013-2823-z
35. Kamori M, Hagihara M, Nagatsu T, Iwata H, Miura T. Activities of dipeptidyl peptidase II, dipeptidyl peptidase IV, prolyl endopeptidase, and collagenase-like peptidase in synovial membrane from patients with rheumatoid arthritis and osteoarthritis. Biochem Med Metab Biol. 1991;45:154–160. doi:10.1016/0885-4505(91)90016-E
36. Cordero OJ, Varela-Calviño R, López-González T, et al. Anti-CD26 autoantibodies are involved in rheumatoid arthritis and show potential clinical interest. Clin. Biochem. 2017;50:903–910. doi:10.1016/j.clinbiochem.2017.06.001
37. Sedo A, Duke-Cohan JS, Balaziova E, Sedova LR. Dipeptidyl peptidase IV activity and/or structure homologs: contributing factors in the pathogenesis of rheumatoid arthritis? Arthritis Res Therapy. 2005;7:1–17. doi:10.1186/ar1852
38. Kanbe K, Chiba J, Inoue Y, Taguchi M, Yabuki A. SDF-1 and CXCR4 in synovium are associated with disease activity and bone and joint destruction in patients with rheumatoid arthritis treated with golimumab. Mod Rheumatol. 2016;26:46–50. doi:10.3109/14397595.2015.1054088
39. Sasaki T, Hiki Y, Nagumo S, et al. Acute onset of rheumatoid arthritis associated with administration of a dipeptidyl peptidase-4 (DPP-4) inhibitor to patients with diabetes mellitus. Diabetol Int. 2010;1:90–92. doi:10.1007/s13340-010-0010-y
40. Glorie L, Behets GJ, Baerts L, De Meester I, d’Haese PC, Verhulst A. DPP IV inhibitor treatment attenuates bone loss and improves mechanical bone strength in male diabetic rats. Am J Physiol Endocrinol Metab. 2014;307:E447–E455. doi:10.1152/ajpendo.00217.2014
41. Yokota K, Igaki N. Sitagliptin (DPP-4 inhibitor)-induced rheumatoid arthritis in type 2 diabetes mellitus: a case report. Internal Medicine. 2012;51:2041–2044. doi:10.2169/internalmedicine.51.7592
42. Padron S, Rogers E, Beckler MD, Kesselman M. DPP-4 inhibitor (sitagliptin)-induced seronegative rheumatoid arthritis. BMJ Case Reports CP. 2019;12:e228981. doi:10.1136/bcr-2018-228981
43. Mascolo A, Rafaniello C, Sportiello L, et al. Dipeptidyl peptidase (DPP)-4 inhibitor-induced arthritis/arthralgia: a review of clinical cases. Drug Safety. 2016;39:401–407. doi:10.1007/s40264-016-0399-8
44. Kathe N, Shah A, Said Q, Painter JT. DPP-4 inhibitor-induced rheumatoid arthritis among diabetics: a nested case–control study. Diabetes Therapy. 2018;9:141–151. doi:10.1007/s13300-017-0353-5
45. Douros A, Abrahami D, Yin H, et al. Use of dipeptidyl peptidase-4 inhibitors and new-onset rheumatoid arthritis in patients with type 2 diabetes. Epidemiology. 2018;29:904–912. doi:10.1097/EDE.0000000000000891
46. Charoenngam N, Rittiphairoj T, Ponvilawan B, Ungprasert P. Use of dipeptidyl peptidase-4 inhibitors is associated with a lower risk of rheumatoid arthritis in patients with type 2 diabetes mellitus: a systematic review and meta-analysis of cohort studies. Diabetes Metab Syndr. 2021;15:249–255. doi:10.1016/j.dsx.2020.12.042
47. Bengsch B, Seigel B, Flecken T, Wolanski J, Blum HE, Thimme R. Human Th17 cells express high levels of enzymatically active dipeptidylpeptidase IV (CD26). J Immunol. 2012;188:5438–5447. doi:10.4049/jimmunol.1103801
48. Yamada Y, Jang J-H, De Meester I, et al. CD26 costimulatory blockade improves lung allograft rejection and is associated with enhanced interleukin-10 expression. J Heart Lung Transplant. 2016;35:508–517. doi:10.1016/j.healun.2015.11.002
49. Willheim M, Ebner C, Kern W, et al. Cell surface characterization of T lymphocytes and allergen-specific T cell clones: correlation of CD26 expression with T H1 subsets. J Allergy Clin Immunol. 1997;100:348–355. doi:10.1016/S0091-6749(97)70248-3
50. Lun SW, Wong C, Ko FW, Hui DS, Lam CW. Increased expression of plasma and CD4+ T lymphocyte costimulatory molecule CD26 in adult patients with allergic asthma. J Clin Immunol. 2007;27:430–437. doi:10.1007/s10875-007-9093-z
51. Salgado FJ, Pérez‐Díaz A, Villanueva NM, Lamas O, Arias P, Nogueira M. CD26: a negative selection marker for human Treg cells. Cytometry Part A. 2012;81:843–855. doi:10.1002/cyto.a.22117
52. Shah Z, Kampfrath T, Deiuliis JA, et al. Long-term dipeptidyl-peptidase 4 inhibition reduces atherosclerosis and inflammation via effects on monocyte recruitment and chemotaxis. Circulation. 2011;124:2338–2349. doi:10.1161/CIRCULATIONAHA.111.041418
53. Gliddon DR, Howard CJ. CD26 is expressed on a restricted subpopulation of dendritic cells in vivo. Europ J Immunol. 2002;32:1472–1481. doi:10.1002/1521-4141(200205)32:5<1472::AID-IMMU1472>3.0.CO;2-Q
54. Zhong J, Rao X, Deiuliis J, et al. A potential role for dendritic cell/macrophage-expressing DPP4 in obesity-induced visceral inflammation. Diabetes. 2013;62:149–157. doi:10.2337/db12-0230
55. Bühling F, Kunz D, Reinhold D, et al. Expression and functional role of dipeptidyl peptidase IV (CD26) on human natural killer cells. Natural Immu. 1994;13:270–279.
56. Shingu K, Helfritz A, Zielinska-Skowronek M, et al. CD26 expression determines lung metastasis in mutant F344 rats: involvement of NK cell function and soluble CD26. Cancer Immunol Immunother. 2003;52:546–554. doi:10.1007/s00262-003-0392-9
57. Yan S, Marguet D, Dobers J, Reutter W, Fan H. Deficiency of CD26 results in a change of cytokine and immunoglobulin secretion after stimulation by pokeweed mitogen. Europ J Immunol. 2003;33:1519–1527. doi:10.1002/eji.200323469
58. Bühling F, Junker U, Reinhold D, Neubert K, Jäger L, Ansorge S. Functional role of CD26 on human B lymphocytes. Immunol Lett. 1995;45:47–51. doi:10.1016/0165-2478(94)00230-O
59. Adams DE, Shao W-H. Epigenetic alterations in immune cells of systemic Lupus erythematosus and therapeutic implications. Cells. 2022;11:506. doi:10.3390/cells11030506
60. Kobayashi H, Hosono O, Mimori T, et al. Reduction of serum soluble CD26/dipeptidyl peptidase IV enzyme activity and its correlation with disease activity in systemic lupus erythematosus. J Rheumatol. 2002;29:1858–1866.
61. Valizadeh M, Ahmadzadeh A, Behzadi M, Yeganeh F. CD26 mRNA expression in systemic lupus erythematosus. Rheumatol Res. 2018;3:77–82. doi:10.22631/rr.2018.69997.1045
62. Awadallah MG, Amer GA, Fawzy RM, El-Melouk MS, Egila SS. CD26 mRNA expression level in systemic lupus erythematosus patients attending Benha University Hospital. Egypt J MedMicrobiol. 2020;29(1):55–60. doi:10.21608/ejmm.2020.249857
63. Seong JM, Yee J, Gwak HS. Dipeptidyl peptidase‐4 inhibitors lower the risk of autoimmune disease in patients with type 2 diabetes mellitus: a nationwide population‐based cohort study. Br J Clin Pharmacol. 2019;85:1719–1727. doi:10.1111/bcp.13955
64. Chen Y-C, Chen T-H, Sun -C-C, et al. Dipeptidyl peptidase-4 inhibitors and the risks of autoimmune diseases in type 2 diabetes mellitus patients in Taiwan: a nationwide population-based cohort study. Acta Diabetologica. 2020;57:1181–1192. doi:10.1007/s00592-020-01533-5
65. Denton CP, Khanna D. Systemic sclerosis. Lancet. 2017;390:1685–1699. doi:10.1016/S0140-6736(17)30933-9
66. Fiocco U, Rosada M, Cozzi L, et al. Early phenotypic activation of circulating helper memory T cells in scleroderma: correlation with disease activity. Ann Rheumatic Dis. 1993;52:272–277. doi:10.1136/ard.52.4.272
67. Petroff A, Pena Diaz A, Armstrong JJ, Gonga-Cavé BC, Hutnik C, O’Gorman DB. Understanding inflammation-associated ophthalmic pathologies: a novel 3D co-culture model of monocyte-myofibroblast immunomodulation. Ocul Immunol Inflamm. 2021;1–12. doi:10.1080/09273948.2020.1862606
68. Bochaton-Piallat M-L, Gabbiani G, Hinz B. The myofibroblast in wound healing and fibrosis: answered and unanswered questions. F1000Research. 2016;2016:5.
69. Bosello S, De Santis M, Lama G, et al. B cell depletion in diffuse progressive systemic sclerosis: safety, skin score modification and IL-6 modulation in an up to thirty-six months follow-up open-label trial. Arthritis Res Therapy. 2010;12:1–10. doi:10.1186/ar2965
70. Ziemek J, Man A, Hinchcliff M, Varga J, Simms RW, Lafyatis R. The relationship between skin symptoms and the scleroderma modification of the health assessment questionnaire, the modified Rodnan skin score, and skin pathology in patients with systemic sclerosis. Rheumatology. 2016;55:911–917. doi:10.1093/rheumatology/kew003
71. Xin Y, Wang X, Zhu M, et al. Expansion of CD26 positive fibroblast population promotes keloid progression. Exp Cell Res. 2017;356:104–113. doi:10.1016/j.yexcr.2017.04.021
72. Shiobara T, Chibana K, Watanabe T, et al. Dipeptidyl peptidase-4 is highly expressed in bronchial epithelial cells of untreated asthma and it increases cell proliferation along with fibronectin production in airway constitutive cells. Respir Res. 2016;17:1–11. doi:10.1186/s12931-016-0342-7
73. Tabib T, Huang M, Morse N, et al. Myofibroblast transcriptome indicates SFRP2hi fibroblast progenitors in systemic sclerosis skin. Nat Commun. 2021;12:1–13. doi:10.1038/s41467-021-24607-6
74. Lee S-Y, Wu S-T, Liang Y-J, et al. Soluble dipeptidyl peptidase-4 induces fibroblast activation through proteinase-activated receptor-2. Front Pharmacol. 2020;1568. doi:10.3389/fphar.2019.01568
75. Zhang T, Tong X, Zhang S, et al. The roles of dipeptidyl peptidase 4 (DPP4) and DPP4 inhibitors in different lung diseases: new evidence. Front Pharmacol. 2021;3657.
76. Abrahami D, Douros A, Yin H, et al. Dipeptidyl peptidase-4 inhibitors and incidence of inflammatory bowel disease among patients with type 2 diabetes: population based cohort study. BMJ. 2018;2018:360.
77. Pinto-Lopes P, Afonso J, Pinto-Lopes R, et al. Serum dipeptidyl peptidase 4: a predictor of disease activity and prognosis in inflammatory bowel disease. Inflammat Bow Dis. 2020;26(11):1707–1719. doi:10.1093/ibd/izz319
78. Magro DO, Kotze PG, Martinez CAR, et al. Changes in serum levels of lipopolysaccharides and CD26 in patients with Crohn’s disease. Intestinal Res. 2017;15:352. doi:10.5217/ir.2017.15.3.352
79. Moran G, O’Neill C, Padfield P, McLaughlin J. Dipeptidyl peptidase-4 expression is reduced in Crohn’s disease. Regul Pept. 2012;177:40–45. doi:10.1016/j.regpep.2012.04.006
80. Di Giovangiulio M, Verheijden S, Bosmans G, Stakenborg N, Boeckxstaens GE, Matteoli G. The neuromodulation of the intestinal immune system and its relevance in inflammatory bowel disease. Front Immunol. 2015;6:590. doi:10.3389/fimmu.2015.00590
81. Detel D, Buljevic S, Pucar LB, Kucic N, Pugel EP, Varljen J. Influence of CD26/dipeptidyl peptidase IV deficiency on immunophenotypic changes during colitis development and resolution. J Physiol Biochem. 2016;72:405–419. doi:10.1007/s13105-016-0491-7
82. Duan L, Rao X, Braunstein Z, Toomey AC, Zhong J. Role of incretin axis in inflammatory bowel disease. Front Immunol. 2017;8:1734. doi:10.3389/fimmu.2017.01734
83. Ning M-M, Yang W-J, Guan W-B, Gu Y-P, Feng Y, Leng Y. Dipeptidyl peptidase 4 inhibitor sitagliptin protected against dextran sulfate sodium-induced experimental colitis by potentiating the action of GLP-2. Acta Pharmacol Sin. 2020;41:1446–1456. doi:10.1038/s41401-020-0413-7
84. Mimura S, Ando T, Ishiguro K, et al. Dipeptidyl peptidase-4 inhibitor anagliptin facilitates restoration of dextran sulfate sodium-induced colitis. Scand J Gastroenterol. 2013;48:1152–1159. doi:10.3109/00365521.2013.832366
85. Kridin K, Amber K, Khamaisi M, Comaneshter D, Batat E, Cohen AD. Is there an association between dipeptidyl peptidase-4 inhibitors and autoimmune disease? A population-based study. Immunol Res. 2018;66:425–430. doi:10.1007/s12026-018-9005-8
86. Wang T, Yang JY, Buse JB, et al. Dipeptidyl peptidase 4 inhibitors and risk of inflammatory bowel disease: real-world evidence in US adults. Diabetes Care. 2019;42:2065–2074. doi:10.2337/dc19-0162
87. Radel JA, Pender DN, Shah SA. Dipeptidyl peptidase-4 inhibitors and inflammatory bowel disease risk: a meta-analysis. Ann. Pharmacother. 2019;53:697–704. doi:10.1177/1060028019827852
88. Malek Abrahimians E, Vander Elst L, Carlier VA, Saint-Remy J-M. Thioreductase-containing epitopes inhibit the development of type 1 diabetes in the NOD mouse model. Front Immunol. 2016;7:67. doi:10.3389/fimmu.2016.00067
89. Iwabuchi A, Kamoda T, Saito M, et al. Serum dipeptidyl peptidase 4 activity in children with type 1 diabetes mellitus. J Pediatr Endocrinol Metab. 2013;26:1093–1097. doi:10.1515/jpem-2013-0122
90. Varga T, Somogyi A, Barna G, et al. Higher serum DPP-4 enzyme activity and decreased lymphocyte CD26 expression in type 1 diabetes. Pathol Oncol Res. 2011;17:925–930. doi:10.1007/s12253-011-9404-9
91. Osawa S, Kawamori D, Katakami N, et al. Significant elevation of serum dipeptidyl peptidase-4 activity in young-adult type 1 diabetes. Diabetes Res Clin Pract. 2016;113:135–142. doi:10.1016/j.diabres.2015.12.022
92. Ellis S, Moser E, Snell‐Bergeon JK, Rodionova A, Hazenfield R, Garg S. Effect of sitagliptin on glucose control in adult patients with Type 1 diabetes: a pilot, double‐blind, randomized, crossover trial. Diabetic Med. 2011;28:1176–1181. doi:10.1111/j.1464-5491.2011.03331.x
93. Garg SK, Moser EG, Bode BW, et al. Effect of sitagliptin on post-prandial glucagon and GLP-1 levels in patients with type 1 diabetes: investigator-initiated, double-blind, randomized, placebo-controlled trial. Endocr Pract. 2013;19:19–28. doi:10.4158/EP12100.OR
94. Kumar KH, Shaikh A, Prusty P. Addition of exenatide or sitagliptin to insulin in new onset type 1 diabetes: a randomized, open label study. Diabetes Res Clin Pract. 2013;100:e55–e58. doi:10.1016/j.diabres.2013.01.020
95. Rahmani-Kukia N, Abbasi A. Physiological and immunological causes of the susceptibility of chronic inflammatory patients to COVID-19 infection: focus on diabetes. Front Endocrinol. 2021;12:84. doi:10.3389/fendo.2021.576412
96. Cechin S, Perez-Alvarez I, Fenjves E, et al. Anti-inflammatory properties of exenatide in human pancreatic islets. Cell Transplant. 2012;21:633–648. doi:10.3727/096368911X576027
97. Kim S-J, Nian C, Doudet DJ, McIntosh CH. Dipeptidyl peptidase IV inhibition with MK0431 improves islet graft survival in diabetic NOD mice partially via T-cell modulation. Diabetes. 2009;58:641–651. doi:10.2337/db08-1101
98. Davanso MR, Caliari-Oliveira C, Couri CEB, et al. DPP-4 inhibition leads to decreased pancreatic inflammatory profile and increased frequency of regulatory T cells in experimental type 1 diabetes. Inflammation. 2019;42:449–462. doi:10.1007/s10753-018-00954-3
99. Scirica BM, Bhatt DL, Braunwald E, et al. Saxagliptin and cardiovascular outcomes in patients with type 2 diabetes mellitus. N Engl J Med. 2013;369:1317–1326. doi:10.1056/NEJMoa1307684
100. Ferreira JP, Mehta C, Sharma A, Nissen SE, Rossignol P, Zannad F. Alogliptin after acute coronary syndrome in patients with type 2 diabetes: a renal function stratified analysis of the EXAMINE trial. BMC Med. 2020;18(1):1–10. doi:10.1186/s12916-020-01616-8
101. Green JB, Bethel MA, Armstrong PW, et al. Effect of sitagliptin on cardiovascular outcomes in type 2 diabetes. N Engl J Med. 2015;373:232–242. doi:10.1056/NEJMoa1501352
102. Best JH, Hoogwerf BJ, Herman WH, et al. Risk of cardiovascular disease events in patients with type 2 diabetes prescribed the glucagon-like peptide 1 (GLP-1) receptor agonist exenatide twice daily or other glucose-lowering therapies: a retrospective analysis of the LifeLink database. Diabetes Care. 2011;34:90–95. doi:10.2337/dc10-1393
103. Gu X, Al Dubayee M, Alshahrani A, et al. Distinctive metabolomics patterns associated with insulin resistance and type 2 diabetes mellitus. Front Mol Biosci. 2020;7:411. doi:10.3389/fmolb.2020.609806
104. Tiruppathi C, Miyamoto Y, Ganapathy V, Roesel RA, Whitford GM, Leibach FH. Hydrolysis and transport of proline-containing peptides in renal brush-border membrane vesicles from dipeptidyl peptidase IV-positive and dipeptidyl peptidase IV-negative rat strains. J Biol Chem. 1990;265:1476–1483. doi:10.1016/S0021-9258(19)40041-0
105. Miyazono K, Heldin C-H. Latent forms of TGF-: molecular structure and mechanisms of activation. Procee Clin Applic. 2008;2008:81–92.
106. Liu WJ, Xie SH, Liu YN, et al. Dipeptidyl peptidase IV inhibitor attenuates kidney injury in streptozotocin-induced diabetic rats. J Pharmacol Exp Ther. 2012;340:248–255. doi:10.1124/jpet.111.186866
107. Mega C, Teixeira de Lemos E, Vala H, et al. Diabetic nephropathy amelioration by a low-dose sitagliptin in an animal model of type 2 diabetes (Zucker diabetic fatty rat). Experim Diab Res. 2011;2011. doi:10.1155/2011/162092
108. Min HS, Kim JE, Lee MH, et al. Dipeptidyl peptidase IV inhibitor protects against renal interstitial fibrosis in a mouse model of ureteral obstruction. Lab Invest. 2014;94:598–607. doi:10.1038/labinvest.2014.50
109. Matsui T, Nakashima S, Nishino Y, et al. Dipeptidyl peptidase-4 deficiency protects against experimental diabetic nephropathy partly by blocking the advanced glycation end products-receptor axis. Lab Invest. 2015;95:525–533. doi:10.1038/labinvest.2015.35
110. Nakashima S, Matsui T, Takeuchi M, Yamagishi S-I. Linagliptin blocks renal damage in type 1 diabetic rats by suppressing advanced glycation end products-receptor axis. Hormone Metab Res. 2014;46:717–721. doi:10.1055/s-0034-1371892
111. Salheen S, Panchapakesan U, Pollock C, Woodman O. The DPP-4 inhibitor linagliptin and the GLP-1 receptor agonist exendin-4 improve endothelium-dependent relaxation of rat mesenteric arteries in the presence of high glucose. Pharmacol Res. 2015;94:26–33. doi:10.1016/j.phrs.2015.02.003
112. Nistala R, Savin V. Diabetes, hypertension, and chronic kidney disease progression: role of DPP4. Am J Physiol Renal Physiol. 2017;312:F661–F670. doi:10.1152/ajprenal.00316.2016
113. Kodera R, Shikata K, Takatsuka T, et al. Dipeptidyl peptidase-4 inhibitor ameliorates early renal injury through its anti-inflammatory action in a rat model of type 1 diabetes. Biochem Biophys Res Commun. 2014;443:828–833. doi:10.1016/j.bbrc.2013.12.049
114. Nabeno M, Akahoshi F, Kishida H, et al. A comparative study of the binding modes of recently launched dipeptidyl peptidase IV inhibitors in the active site. Biochem Biophys Res Commun. 2013;434:191–196. doi:10.1016/j.bbrc.2013.03.010
115. Makrantonaki E, Jiang D, Hossini AM, et al. Diabetes mellitus and the skin. Rev Endocr Metab Disord. 2016;17:269–282. doi:10.1007/s11154-016-9373-0
116. Röhrborn D, Wronkowitz N, Eckel J. DPP4 in diabetes. Front Immunol. 2015;6:386. doi:10.3389/fimmu.2015.00386
117. Maryam S, Ul Haq I, Yahya G, et al. COVID-19 surveillance in wastewater: an epidemiological tool for the monitoring of SARS-CoV-2. Front Cell Infect Microbiol. 2023;12:1743. doi:10.3389/fcimb.2022.978643
118. Elmorsy MA, El-Baz AM, Mohamed NH, Almeer R, Abdel-Daim MM, Yahya G. In silico screening of potent inhibitors against COVID-19 key targets from a library of FDA-approved drugs. Environ Sci Pollut Res Int. 2022;29(8):12336–12346. doi:10.1007/s11356-021-16427-4
119. Yang L, Tu L. Implications of gastrointestinal manifestations of COVID-19. Lancet Gastroent Hepat. 2020;5:629–630. doi:10.1016/S2468-1253(20)30132-1
120. Ul Haq I, Krukiewicz K, Yahya G, et al. The breadth of bacteriophages contributing to the development of the phage-based vaccines for COVID-19: an ideal platform to design the multiplex vaccine. Int J Mol Sci. 2023;24:1536. doi:10.3390/ijms24021536
121. Panther EJ, Lucke-Wold B. Subarachnoid hemorrhage: management considerations for COVID-19. Explorat Neuroprot Thera. 2022;2:65. doi:10.37349/ent.2022.00018
122. Egbuna C, Chandra S, Awuchi CG, et al. Myth surrounding the FDA disapproval of hydroxychloroquine sulfate and chloroquine phosphate as drugs for coronavirus disease 2019. In: Coronavirus Drug Discovery. Elsevier; 2022:153–168.
123. Haq IU, Krukiewicz K, Tayyab H, et al. Molecular Understanding of ACE-2 and HLA-conferred differential susceptibility to COVID-19: host-directed insights opening new windows in COVID-19 Therapeutics. J Clin Med. 2023;12:2645. doi:10.3390/jcm12072645
124. Levy E, Delvin E, Marcil V, Spahis S. Can phytotherapy with polyphenols serve as a powerful approach for the prevention and therapy tool of novel coronavirus disease 2019 (COVID-19)? Am J Physiol Endocrinol Metab. 2020;319:E689–E708. doi:10.1152/ajpendo.00298.2020
125. Sizemore G, Lucke-Wold B, Small C. Review of SARS-COV-2 systemic impact: building the case for sepsis via virus in the circulatory system. SM J Neurol Disord Strok. 2022;2022:6.
126. Qi F, Qian S, Zhang S, Zhang Z. Single cell RNA sequencing of 13 human tissues identify cell types and receptors of human coronaviruses. Biochem Biophys Res Commun. 2020;526:135–140. doi:10.1016/j.bbrc.2020.03.044
127. Meyerholz DK, Lambertz AM, McCray PB. Dipeptidyl peptidase 4 distribution in the human respiratory tract: implications for the Middle East respiratory syndrome. Am J Pathol. 2016;186:78–86. doi:10.1016/j.ajpath.2015.09.014
128. Seys LJ, Widagdo W, Verhamme FM, et al. DPP4, the Middle East respiratory syndrome coronavirus receptor, is upregulated in lungs of smokers and chronic obstructive pulmonary disease patients. Clinl Infect Dis. 2018;66:45–53. doi:10.1093/cid/cix741
129. Widagdo W, Sooksawasdi Na Ayudhya S, Hundie GB, Haagmans BL. Host determinants of MERS-CoV transmission and pathogenesis. Viruses. 2019;11:280. doi:10.3390/v11030280
130. Zhong J, Maiseyeu A, Davis SN, Rajagopalan S. DPP4 in cardiometabolic disease: recent insights from the laboratory and clinical trials of DPP4 inhibition. Circul Res. 2015;116:1491–1504. doi:10.1161/CIRCRESAHA.116.305665
131. Noels H, Bernhagen J. The CXCR4 ligand/receptor family and the DPP4 protease in high-risk cardiovascular patients. Front Immunol. 2016;7:58. doi:10.3389/fimmu.2016.00058
132. Conarello SL, Li Z, Ronan J, et al. Mice lacking dipeptidyl peptidase IV are protected against obesity and insulin resistance. Proc Natl Acad Sci. 2003;100:6825–6830. doi:10.1073/pnas.0631828100
133. Kulcsar K, Coleman C, Beck S, Frieman M. Comorbid diabetes results in immune dysregulation and enhanced disease severity following MERS-CoV infection. JCI Insight. 2019;4(20):e131774. doi:10.1172/jci.insight.131774
134. Kim KM, Noh JH, Bodogai M, et al. Identification of senescent cell surface targetable protein DPP4. Genes Dev. 2017;31:1529–1534. doi:10.1101/gad.302570.117
135. Atal S, Fatima Z. IL-6 inhibitors in the treatment of serious COVID-19: a promising therapy? Pharm Med. 2020;34:223–231. doi:10.1007/s40290-020-00342-z
136. Zea-Vera A, Cordova EG, Rodriguez S, et al. Parasite antigen in serum predicts the presence of viable brain parasites in patients with apparently calcified cysticercosis only. Clinl Infect Dis. 2013;57:e154–e159. doi:10.1093/cid/cit422
137. Krejner‐Bienias A, Grzela K, Grzela T. Do novel drugs for diabetes help in COVID‐19? Another brick in the wall? J Diab. 2020;12:703. doi:10.1111/1753-0407.13050
138. Inan M, Koyuncu A, Aydin C, Turan M, Gokgoz S, Sen M. Thyroid hormone supplementation in sepsis: an experimental study. Surgery Today. 2003;33:24–29. doi:10.1007/s005950300004
139. Arulmozhiraja S, Matsuo N, Ishitsubo E, Okazaki S, Shimano H, Tokiwa H. Comparative binding analysis of dipeptidyl peptidase IV (DPP-4) with antidiabetic drugs–an ab initio fragment molecular orbital study. PLoS One. 2016;11:e0166275. doi:10.1371/journal.pone.0166275
140. Wilce J, Wilce JA. Emerging WuHan (COVID-19) coronavirus: glycan shield and structure prediction of spike glycoprotein and its interaction with human CD26. Emerg Microbes Infect. 2020;9:601–604.
141. Bardaweel SK, Hajjo R, Sabbah A. Sitagliptin: a potential drug for the treatment of COVID-19? Acta Pharm. 2021;71:175–184. doi:10.2478/acph-2021-0013
142. Hippisley-Cox J, Young D, Coupland C, et al. Risk of severe COVID-1352 19 disease with ACE inhibitors and angiotensin receptor blockers: cohort study 1353 including 8.3 million people. Heart. 2020;106:1503–1511. doi:10.1136/heartjnl-2020-317393
143. van Riet E, Everts B, Retra K, et al. Combined TLR2 and TLR4 ligation in the context of bacterial or helminth extracts in human monocyte derived dendritic cells: molecular correlates for Th1/Th2 polarization. BMC Immunol. 2009;10:1–12. doi:10.1186/1471-2172-10-9
144. Price J, Linder G, Li W, et al. Effects of short-term sitagliptin treatment on immune parameters in healthy individuals, a randomized placebo-controlled study. Clin Exp Immunol. 2013;174:120–128. doi:10.1111/cei.12144
145. Yang W, Cai X, Han X, Ji L. DPP‐4 inhibitors and risk of infections: a meta‐analysis of randomized controlled trials. Diabetes/Metab Res Rev. 2016;32:391–404. doi:10.1002/dmrr.2723
146. Wvan der Zanden R, De Vries F, Lalmohamed A, et al. Use of dipeptidyl-peptidase-4 inhibitors and the risk of pneumonia: a population-based cohort study. PLoS One. 2015;10:e0139367. doi:10.1371/journal.pone.0139367
147. Gooßen K, Gräber S. Longer term safety of dipeptidyl peptidase‐4 inhibitors in patients with type 2 diabetes mellitus: systematic review and meta‐analysis. Diabetes Obesity Metab. 2012;14:1061–1072. doi:10.1111/j.1463-1326.2012.01610.x
148. Kupferschmidt K. A cheap steroid is the first drug shown to reduce death in COVID-19 patients. Science. 2020;2020:1.
149. Solerte SB, D’Addio F, Trevisan R, et al. Sitagliptin treatment at the time of hospitalization was associated with reduced mortality in patients with type 2 diabetes and COVID-19: a multicenter, case-control, retrospective, observational study. Diabetes Care. 2020;43:2999–3006. doi:10.2337/dc20-1521
150. Zhong J, Rao X, Rajagopalan S. An emerging role of dipeptidyl peptidase 4 (DPP4) beyond glucose control: potential implications in cardiovascular disease. Atherosclerosis. 2013;226:305–314. doi:10.1016/j.atherosclerosis.2012.09.012
151. Baz M, Boivin G. Antiviral agents in development for Zika virus infections. Pharmaceuticals. 2019;12:101. doi:10.3390/ph12030101
152. Ma J, Zhong M, Xiong Y, et al. Emerging roles of nucleotide metabolism in cancer development: progress and prospect. Aging. 2021;13:13349. doi:10.18632/aging.202962
153. Ragab D, Laird M, Duffy D, et al. CXCL10 antagonism and plasma sDPPIV correlate with increasing liver disease in chronic HCV genotype 4 infected patients. Cytokine. 2013;63:105–112. doi:10.1016/j.cyto.2013.04.016
154. Rainczuk A, Rao JR, Gathercole JL, et al. Evidence for the antagonistic form of CXC‐motif chemokine CXCL10 in serous epithelial ovarian tumours. Internat J Can. 2014;134:530–541. doi:10.1002/ijc.28393
155. Li B, Yang J, Zhao F, et al. Prevalence and impact of cardiovascular metabolic diseases on COVID-19 in China. Clin Res Cardiol. 2020;109:531–538. doi:10.1007/s00392-020-01626-9
156. Liu Z, Li J, Huang J, et al. Association between diabetes and COVID-19: a retrospective observational study with a large sample of 1880 cases in Leishenshan Hospital, Wuhan. Front Endocrinol. 2020;478. doi:10.3389/fendo.2020.00478
157. Wu Z, McGoogan JM. Asymptomatic and pre-symptomatic COVID-19 in China. Infect Diseases Poverty. 2020;9:1–2. doi:10.1186/s40249-020-00679-2
158. Mauvais-Jarvis F. Aging, male sex, obesity, and metabolic inflammation create the perfect storm for COVID-19. Diabetes. 2020;69:1857–1863. doi:10.2337/dbi19-0023
159. Ghorpade DS, Ozcan L, Zheng Z, et al. Hepatocyte-secreted DPP4 in obesity promotes adipose inflammation and insulin resistance. Nature. 2018;555:673–677. doi:10.1038/nature26138
160. Shekhar S, Copacino CE, Barrera FJ, Hall JE, Hannah-Shmouni F. Insights into the immunopathophysiology of severe COVID-19 in METABOLIC DISOrders. Anna Nat Acad Med Sci. 2020;56:112–115. doi:10.1055/s-0040-1713346
161. Kirino Y, Sei M, Kawazoe K, Minakuchi K, Sato Y. Plasma dipeptidyl peptidase 4 activity correlates with body mass index and the plasma adiponectin concentration in healthy young people. Endocr J. 2012;59:949–953. doi:10.1507/endocrj.EJ12-0158
162. Yang F, Zheng T, Gao Y, et al. Increased plasma DPP4 activity is an independent predictor of the onset of metabolic syndrome in Chinese over 4 years: result from the China national diabetes and metabolic disorders study. PLoS One. 2014;9:e92222. doi:10.1371/journal.pone.0092222
163. Cavalu S, Damian G, Dânşoreanu M. EPR study of non-covalent spin labeled serum albumin and hemoglobin. Biophys Chem. 2022;99(2):181–188. doi:10.1016/S0301-4622(02)00182-5
164. Metwally K, Pratsinis H, Kletsas D, et al. Novel quinazolinone-based 2,4-thiazolidinedione-3-acetic acid derivatives as potent aldose reductase inhibitors. Future Med Chem. 2017;9(18):2147–2166. doi:10.4155/fmc-2017-0149
165. Pang Z, Nakagami H, Osako MK, et al. Therapeutic vaccine against DPP4 improves glucose metabolism in mice. Proc Natl Acad Sci. 2014;111:E1256–E1263. doi:10.1073/pnas.1322009111
166. Fujihara K, Igarashi R, Matsunaga S, et al. Comparison of baseline characteristics and clinical course in Japanese patients with type 2 diabetes among whom different types of oral hypoglycemic agents were chosen by diabetes specialists as initial monotherapy (JDDM 42). Medicine. 2017;15:96. doi:10.1186/s12916-017-0859-8
167. Kagal UA, Angadi NB, Matule SM. Effect of dipeptidyl peptidase 4 inhibitors on acute and subacute models of inflammation in male Wistar rats: an experimental study. Int J App Basic Med Res. 2017;7:26. doi:10.4103/2229-516X.198516
168. Birnbaum Y, Bajaj M, Qian J, Ye Y. Dipeptidyl peptidase-4 inhibition by Saxagliptin prevents inflammation and renal injury by targeting the Nlrp3/ASC inflammasome. BMJ Open Diabetes Res Care. 2016;4:e000227. doi:10.1136/bmjdrc-2016-000227
169. Beraldo JI, Benetti A, Borges-Júnior FA, et al. Cardioprotection conferred by sitagliptin is associated with reduced cardiac angiotensin II/Angiotensin-(1-7) balance in experimental chronic kidney disease. Int J Mol Sci. 2019;20:1940. doi:10.3390/ijms20081940
170. Schuppan D, Gorrell MD, Klein T, Mark M, Afdhal NH. The challenge of developing novel pharmacological therapies for non‐alcoholic steatohepatitis. Liver Int. 2010;30:795–808. doi:10.1111/j.1478-3231.2010.02264.x
171. Tacelli M, Celsa C, Magro B, et al. Antidiabetic drugs in NAFLD: the accomplishment of two goals at once? Pharmaceuticals. 2018;11:121. doi:10.3390/ph11040121
172. Makdissi A, Ghanim H, Vora M, et al. Sitagliptin exerts an antinflammatory action. J Clin Endocrinol Metab. 2012;97:3333–3341. doi:10.1210/jc.2012-1544
173. Tang S, Ma W, Bai P. A novel dynamic model describing the spread of the MERS-CoV and the expression of dipeptidyl peptidase 4. Comput Math Methods Med. 2017;2017. doi:10.1155/2017/5285810
174. Mehta P, McAuley DF, Brown M, Sanchez E, Tattersall RS, Manson JJ. COVID-19: consider cytokine storm syndromes and immunosuppression. Lancet. 2020;395:1033–1034. doi:10.1016/S0140-6736(20)30628-0
175. Lagunas‐Rangel FA. Neutrophil‐to‐lymphocyte ratio and lymphocyte‐to‐C‐reactive protein ratio in patients with severe coronavirus disease 2019 (COVID‐19): a meta‐analysis. J med virol. 2020;92:1733–1734. doi:10.1002/jmv.25819
176. Sakaeda T, Tamon A, Kadoyama K, Okuno Y. Data mining of the public version of the FDA Adverse Event Reporting System. Int J Med Sci. 2013;10:796. doi:10.7150/ijms.6048
 © 2024 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2024 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.