Back to Journals » Journal of Inflammation Research » Volume 16
Resveratrol Ameliorates Imiquimod-Induced Psoriasis-Like Mouse Model via Reducing Macrophage Infiltration and Inhibiting Glycolysis
Authors Wang Y, Qi C, Feng F, Hu X, Zhao N, Zhao J ![]() , Di T, Meng Y, Yang D, Zhu H
, Di T, Meng Y, Yang D, Zhu H ![]() , Zhang X, Li P, Wang Y
, Zhang X, Li P, Wang Y
Received 1 May 2023
Accepted for publication 19 August 2023
Published 30 August 2023 Volume 2023:16 Pages 3823—3836
DOI https://doi.org/10.2147/JIR.S416417
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Adam Bachstetter
Yazhuo Wang,1,2 Cong Qi,1,2 Fang Feng,3 Xueqing Hu,3 Ning Zhao,1,2 Jingxia Zhao,1 Tingting Di,1 Yujiao Meng,1 Danyang Yang,1,2 Haoyue Zhu,1,2 Xiawei Zhang,1,2 Ping Li,1 Yan Wang1
1Beijing Hospital of Traditional Chinese Medicine, Capital Medical University, Beijing Institute of Traditional Chinese Medicine, Beijing Key Laboratory of Clinic and Basic Research with Traditional Chinese Medicine on Psoriasis, Beijing, People’s Republic of China; 2Capital Medical University, Beijing, 100069, People’s Republic of China; 3Beijing University of Chinese Medicine, Beijing, 100029, People’s Republic of China
Correspondence: Ping Li; Yan Wang, Beijing Hospital of Traditional Chinese Medicine, Capital Medical University, Beijing Institute of Traditional Chinese Medicine, Beijing Key Laboratory of Clinic and Basic Research with Traditional Chinese Medicine on Psoriasis, Beijing, 100010, People’s Republic of China, Email [email protected]; [email protected]
Purpose: Resveratrol (Res) is a natural polyphenol with anti-inflammatory and immunomodulatory effects. Alterations in metabolic pathways have been studied in psoriasis. This study is aimed to further explore the potential molecular mechanism of psoriasis improvement by Res.
Patients and Methods: Imiquimod (IMQ)-induced psoriasis-like mouse model was established to observe the effects of Res. NanoString nCounter Metabolic Pathways Panel was used to analyze the changed mRNA and qRT-PCR was used for validation. Flow cytometry was used to analyze immune cell subsets in skin lesions. In vitro, we observed the effects of Res on R848-stimulated macrophages glycolysis and inflammation.
Results: Res reduced the proliferation of keratinocytes and the secretion of inflammatory cytokines in IMQ-induced psoriasis-like mouse model. Psoriasis model skin lesions were in a state of hypoxia, with upregulated glycolysis and downregulated AMPK activity. Res inhibited the levels of hypoxia-related genes (hif1α, hif3α) and glycolysis-related genes (hk1, ldha), meanwhile increased the levels of AMPK genes (prkaa1, prkaa2). Flow cytometry analysis revealed that Res decreased the infiltration of macrophages in psoriasis-like lesions. In addition, Res decreased the secretion of macrophage-associated pro-inflammatory cytokines (IL-23, TNF-α, IL-1β). In vitro, Res diminished the secretion of IL-23, TNF-α, IL-1β, and lactate by R848-stimulated macrophages and activated AMPK.
Conclusion: This study suggested that Res diminished psoriasis symptoms by inhibiting macrophages infiltration and inhibiting glycolysis, which providing novel insights into the underlying mechanisms of therapeutic action of Res in the treatment of psoriasis.
Keywords: resveratrol, psoriasis, macrophage, AMPK, glycolysis
Introduction
Psoriasis is a common, chronic inflammatory skin disease that affects over 60 million adults and children worldwide. The most common type is psoriasis vulgaris, characterized by relapsing cutaneous erythema and scales.1 Although the pathogenesis of psoriasis remains unclear, it is generally assumed that psoriasis lesions result from hyperproliferation and disturbed differentiation of epidermal keratinocytes provoked by the IL-23/IL-17 pathway.2,3 Considerable immune cell infiltration has been observed, especially in macrophages, dendritic cells, and T cells, which are abundant in the dermis.1 T cells, which produce IL-17, IL-22, and TNF-a, are recognized as major pro-inflammatory factors in psoriasis. Metabolic pathways in these immune cells, also called “immunometabolism”, mainly consist of glycolysis, the tricarboxylic acid (TCA) cycle, the pentose phosphate pathway, fatty acid oxidation (FAO), fatty acid synthesis, and amino acid metabolism, which have been shown to shape the response of immune cells.4 In addition to T cells, accumulating recent evidence has highlighted the role of macrophages in psoriasis pathogenesis.5,6 Macrophages are a significant source of TNF-a and IL-23. Reportedly, macrophages account for 61.29% of infiltrating inflammatory cells in psoriasis lesions, and thus macrophages depletion could improve PASI scores.7 The extensive infiltration of macrophages in psoriasis lesions deserves further study to elucidate the relationship between macrophages and the pathogenesis of psoriasis.
Resveratrol (Res) (Figure 1) is a natural polyphenolic compound that exists in many plants belonging to the genera Vitis L., Veratrum L., Arachis, Polygonum, etc.8 It affects energy metabolism and mitochondrial function and improves glucose tolerance and insulin sensitivity in obese mice and humans. Moreover, it may be associated with the activation of AMPK (AMP-activated protein kinase) and the increase of PGC-1a (peroxisome proliferator-activated receptor-gamma coactivator-1 alpha) protein level.9–11 Recently, several studies showed that both oral and topical gel administration of Res improved the inflammatory response in psoriatic mice, possibly by inhibiting IL-17 secretion and keratinocyte proliferation.12–15 However, the target cells and the molecular mechanism underlying Res alleviation of psoriasis have not been fully elucidated.
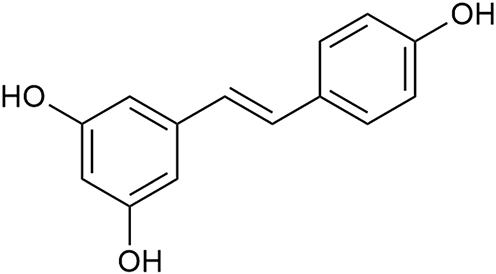 |
Figure 1 Chemical structure of resveratrol. |
In the present study, we evaluated the effects of Res on IMQ-induced psoriasis-like skin inflammation in mice. In addition to the basic disease-related cytokines, we focused on the changed metabolic pathways of lesions. Res reduced the proliferation of keratinocytes and attenuated psoriasis-related inflammatory responses. We found Res remarkable regulate glycolysis and AMPK, and inhibited the infiltration of macrophages in skin lesions. In vitro, Res diminished the levels of pro-inflammation cytokines secreted by macrophages, such as TNF-α, IL-23, and IL-1β, and inhibited macrophage glycolysis levels by AMPK activation. Our study provides novel insights into the mechanisms of therapeutic action of Res in the treatment of psoriasis.
Materials and Methods
Reagents
Imiquimod cream was obtained from Sichuan Mingxin Pharmaceutical Co., Ltd. (Sichuan, China). R848 was bought from STEMCELL Technologies Inc. (Vancouver, BC, Canada). Resveratrol [>98% high-performance liquid chromatography (HPLC) purity] was purchased from Shanghai Yuanye Bio-Technology Co., Ltd (Shanghai, China). HyClone Dulbecco’s Modified Eagle Medium (high glucose, with L-glutamine, and sodium pyruvate) was purchased from Cytiva (Shanghai, China). Fetal bovine serum (FBS), L-glutamine, and penicillin-streptomycin were purchased from Gibco Life Technologies (Grand Island, NY, USA). Hanks’ Balanced Salt Solution 10x (no calcium, no magnesium, no phenol red), HEPES solution (1M, pH 7.0–7.6, sterile-filtered), and Calcium chloride were obtained from Sigma-Aldrich (UK). DNase I and Collagenase A from Clostridium histolyticum were bought from Roche (Mannheim, Germany). DDT was purchased from Beijing Solarbio Science&Technology Co., Ltd. (Beijing, China). CCK-8 kit and Lactate Assay Kit were obtained from Dojindo Laboratories (Japan). Ki67, CD11b, F4/80, and AMPKα1 antibodies were purchased from Abcam (Cambridge, UK). Fluorescent mounting medium with DAPI, and normal goat serum for blocking were obtained from Beijing Zhongshan Golden Bridge Biotechnology Co., Ltd. (Beijing, China). Bicinchoninic acid (BCA) protein assay kit was bought from Pierce (Rockford, IL, USA). P-AMPKα (Thr172) and HK II were purchased from Cell Signaling Technology (Danvers, MA, USA). β-tubulin was obtained from Beijing GuanXingYu Sci&Tech Co., Ltd. (Beijing, China). Anti-mouse CD45 APC/CY7, CD3 FITC, CD11b V500, CD11b BV605, CD11C PE/CY7, and F4/80 BV421 were obtained from BioLegend (San Diego, CA, USA). Ultra-pure total RNA extraction Kit and HiFi-MMLV cDNA first strand synthesis kit was bought from CoWin Biosciences (Jiangsu, China).
Animal Model and Groups
Male C57BL/6J mice, aged 6–8 weeks, were supplied by Beijing Huafukang Biological Technology Co., Ltd. (Beijing, China) (certification no. SCXK Jing 2019–0008) and kept in a specific pathogen-free environment. All experiments were conducted following the principles of Beijing Ethical Examination Guidelines for Experimental Animal Welfare approved by the Beijing Institute of Traditional Chinese Medicine (approval no.2020080203).
The mice were randomly divided into four groups of six mice each: Control, Model, Res-L (20 mg/kg), and Res-H (40 mg/kg). The animals were treated topically with 62.5 mg of imiquimod cream (5%) on the shaved back for six consecutive days to establish a psoriasiform lesion model, whereas the control group was treated with vaseline. Res was dissolved in 0.5% sodium carboxymethyl cellulose to achieve different concentrations for oral administration. The control and model groups received 0.5% sodium carboxymethyl cellulose. We observed and photographed the skin lesions of the mice every day. Six days later, the mice were sacrificed by cervical dislocation under sodium pentobarbital anesthesia, and the skin lesions were harvested.
Cell Culture
Raw264.7 cells were supplied by the National Laboratory Cell Resource Sharing Service Platform (Beijing, China) and cultured in DMEM medium supplemented with 10% FBS at 37°C in a humidified incubator with 5% CO2.
Histology, Immunohistochemistry, and Immunofluorescence Staining
Paraffin-embedded skin samples were cut into 5-μm tissue sections and subjected to hematoxylin and eosin (HE) staining. The histopathological changes were scanned using the Aperio CS2 Leica scanner (Leica, Germany). Epidermal thickness was measured using Image ScopeTM (Aperio Technologies, Vista, CA, USA). For immunofluorescence staining, the sections were stained with anti-Ki670, and DAPI was applied for counterstaining. Slides were observed under a Zeiss Axio Imager (Germany) and laser scanning confocal microscope (Zeiss LSM710, Germany). Under a 200x microscope, five fields were randomly selected for each slide and photographed.
Skin Cell Preparation
Hanks’ Balanced Salt Solution 10x diluted with ddH2O was mixed into 1x suspensions buffer, containing HEPES solution, DNase I, collagenase A, DDT, calcium chloride, L-glutamine, penicillin-streptomycin. The solution was store at 4°C before the experiment. Mice skin lesions were collected and transferred to wash off the remaining IMQ cream. We used scissors to cut the skin into crumbs to ensure adequate digestion. Then, we transferred the crumbs into a 50-mL conical centrifuge tube with 20 mL of digestive buffer, followed by incubation in a gas bath thermostatic oscillator for 1 h at 37°C and 220 rpm. Next, the suspension was filtered through a 70-μm cell strainer into a 50-mL tube. Then, the samples were centrifuged for 5 min at 1500 rpm and 4°C. Finally, the pellet was resuspended with 1 mL of cold staining buffer and kept on ice until staining.
Flow Cytometry
The cell densities of the prepared skin cells were adjusted to be within 106 cells per 100 μL after cell counting. Then, 1 μL Fc blocking buffer was added, followed by incubation for 5 min in the dark. Further, antibodies were added following the manual instructions, and incubated the samples for 15 min in the dark. Next, samples were washed twice with 2 mL of PBS and centrifugated at 1500 rpm for 10 min. CountBright beads quantified absolute cell count. Data were collected using a BD LSR II and analyzed using FACSDiVa 8.0 software.
Western Blot
The amounts of the protein were measured using the bicinchoninic acid assay kit. Samples were run on a sodium dodecyl sulfate-polyacrylamide gel and transferred onto polyvinylidene difluoride membranes. The membranes were then blocked using 5% skimmed milk at room temperature for 1 h and incubated with the primary antibodies against AMPKα1, P-AMPKα (Thr172), HK II, and β-tubulin at 4°C overnight. DyLight 800-Goat Anti-Mouse IgG (H+L), HSA (1:8000), DyLight 680-Goat Anti-Rabbit IgG (H+L), HSA (1:8000), for 1 h in the dark. The immunofluorescence intensities of the membranes were measured using the Odyssey® CLx Infrared Imaging System (LI-COR Biosciences, Lincoln, NE, USA).
RNA Isolation and Gene Expression Analysis
RNA was extracted from frozen mouse skin samples using the TransZol Up Plus RNA Kit (TransGen Biotech, BJS, CHN). RNA concentration was assessed with Nanodrop (Thermo Scientific, CA, USA). RNA fragmentation and quality were determined with a 2100 Bioanalyzer (Agilent, CA, USA). Systematic mRNA expression was measured on a nCounter FLEX Analysis System (NanoString, Seattle, WA, USA) using the nCounter Metabolic Pathways Panel (NanoString Technologies Inc.), covering 768 Mouse mRNAs associated with metabolic processes and immunometabolism. A minimum input of 100 ng of total RNA was utilized for each sample. Fluorescently color-coded reporter probes and biotin-labeled capture probes were hybridized to the mRNA on a thermal cycler overnight and automatically processed and loaded to the NanoString provided sample cartridge in the nCounter Prep Station following the manufacturer’s protocol. The row counts resulting from the analysis were normalized against reference genes (genes selected to have a minor variance with the geNorm algorithm). The normalized data were then analyzed using NanoString’s nSolver version 4.0 software with the Advanced Analysis application tool.
Quantitative RT-PCR Analysis
Total RNA was extracted from the skin lesions using an Ultra-pure total RNA extraction Kit. Reverse transcription was carried out with HiFi-MMLV cDNA first strand synthesis kit. Quantitative RT-PCR amplification was analyzed by ABI 7500 Real-time PCR systems (Applied Biosystems, USA). The RNA expression level was determined by 2−ΔΔCt. The actin was used as a reference to normalize the data. Beijing Invitrogen Company synthesized primers (Table 1).
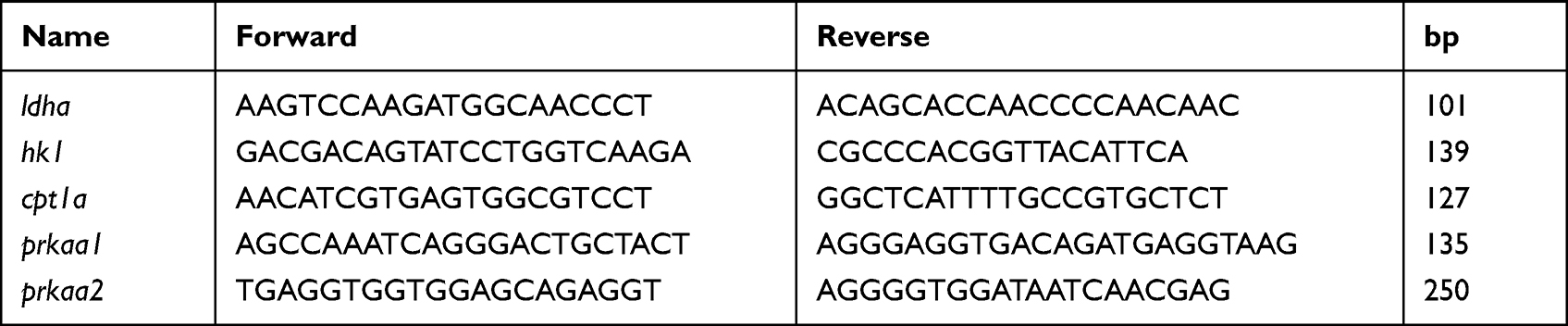 |
Table 1 The Primers Information of Genes |
Cell Viability Assay
The effects of Res on cell viability were assessed using the Cell Counting Kit-8 assay according to the directions provided by the manufacturer. Cells were seeded in 96-well plates and treated with different Res concentrations for 12 h. Next, 10 μL of CCK-8 reagent was added to each well and incubated at 37°C in 5% CO2 for 1 h. The mean optical density (OD) of the cells in each group was used to identify the non-toxic concentration of Res.
ProcartaPlex Multiplex Immunoassays
Skin tissue lysis mixtures and Raw264.7 cells supernatant were collected and assessed by 17-Plex Mouse ProcartaPlex™ Panel (EPX170-26087-901, Invitrogen, Carlsbad, CA, USA) according to the manufacturer’s instructions. The data were then read using a Luminex 200 analyzer. The data were fitted to the standard curve by the five-parameter nonlinear regression method, and the concentration value was calculated.
Statistical Analysis
The quantitative data are expressed as means ± SD. t-test was applied for comparison between the two groups. One-way analysis of variance (ANOVA) was employed for comparison between groups. Analysis for statistical significance and data display was conducted with GraphPad Prism 8.0. Correlation analysis was adopted using Pearson’s correlation. P < 0.05 was considered to indicate a statistically significant difference.
Results
Res Attenuated IMQ-Induced Psoriasis-Like Skin Inflammation
To explore the potential mechanism of Res in psoriasis treatment, we utilized an IMQ-induced psoriasis-like mouse model. Obvious scaling, erythema, and inflammatory infiltrate were observed in the IMQ-treated model mice. Notably, the mice treated with different concentrations of Res had lighter erythema, smoother skin, and fewer scales (Figure 2A). H&E staining of the skin lesions of the model mice revealed a thickened epidermal acanthocyte layer and downward digitation of the epidermis. In the Res groups, the pathological manifestations were significantly less pronounced, and the epidermis thickness was significantly lower than those in the model group (Figure 2B and C). To observe the Res-induced improvement in the pathological manifestations of psoriasis, we determined the expression of Ki67 in the mouse skin, a marker associated with cell proliferation, using immunofluorescence staining techniques. The levels of Ki67-positive cells were considerably higher in the skin epidermal basal layer of the model mice than in that of the control mice. Conversely, in the Res-treated mice, the expression of Ki67 was lower than that in the model mice, suggesting that Res reduces IMQ-induced proliferation and differentiation of keratinocytes (Figure 2D). As shown in Figure 2E–H, IL-23, IL-1β, IL-17A, and TNF-α secretion levels of model mice lesions were significantly upregulated compared with the control mice and Res inhibited these inflammatory factors secretion. These results suggest that Res attenuated IMQ-induced psoriasis-like skin inflammation by reducing the proliferation of keratinocytes and the secretion of inflammatory cytokines.
 |
Figure 2 Res improves IMQ-induced psoriasis-like skin inflammation. Notes: IMQ cream was applied on the back for six days in the model and Res groups, whereas the mice in the control group received Vaseline. (A) Photographs of IMQ-induced psoriasis-like skin on day 7. (B) Epidermal thickness evaluated by microscope. (C) H&E staining of the skin lesions. Scale bar =200μm. (D) Immunofluorescence staining for Ki67 (green) and DAPI (blue). Scale bar=50μm. (E–H) IL-23, IL-1β, IL-17A, TNF-α secretion levels of skin. Data are expressed as the mean ± SD (n=4–6). ##P<0.01, ###P<0.001 vs control group; **P<0.01, ***P<0.001 vs model group. |
Res Inhibited Glycolysis and Affected Multiple Biological Pathways
To extend and improve the existing understanding of the functional changes in Res-treated mice, we measured the mRNA using a nCounter FLEX Analysis System using the nCounter Metabolic Pathways Panel, covering 768 mouse mRNAs associated with metabolic processes and immunometabolism. Compared to the control group, 254 mRNAs in the model group were significantly upregulated (Figure 3A). In the Res group, 79 mRNAs were considerably more upregulated than in the model group (Figure 3B). Compared to the control group mice, metabolic pathways of psoriasis-like mice were significantly altered. The heatmap of pathway scores showed that the expression levels of glycolysis, arginine metabolism, and glutamine metabolism were higher. The expression levels of AMPK, mTOR, FAO, and amino acid transporters were lower (Figure 1A). In addition, the expression levels of PI3K, NF-κB, TLR signaling, reactive oxygen response, lysosomal degradation, and MAPK were up-regulated (Figure 3C). Res inhibited the levels of hypoxia-related genes (hif1α, hif3α) and glycolysis-related genes (hk1, hk3), and increased the levels of AMPK pathway-related genes (prkaa1, prkab2, pgk1) (Figure 3D).
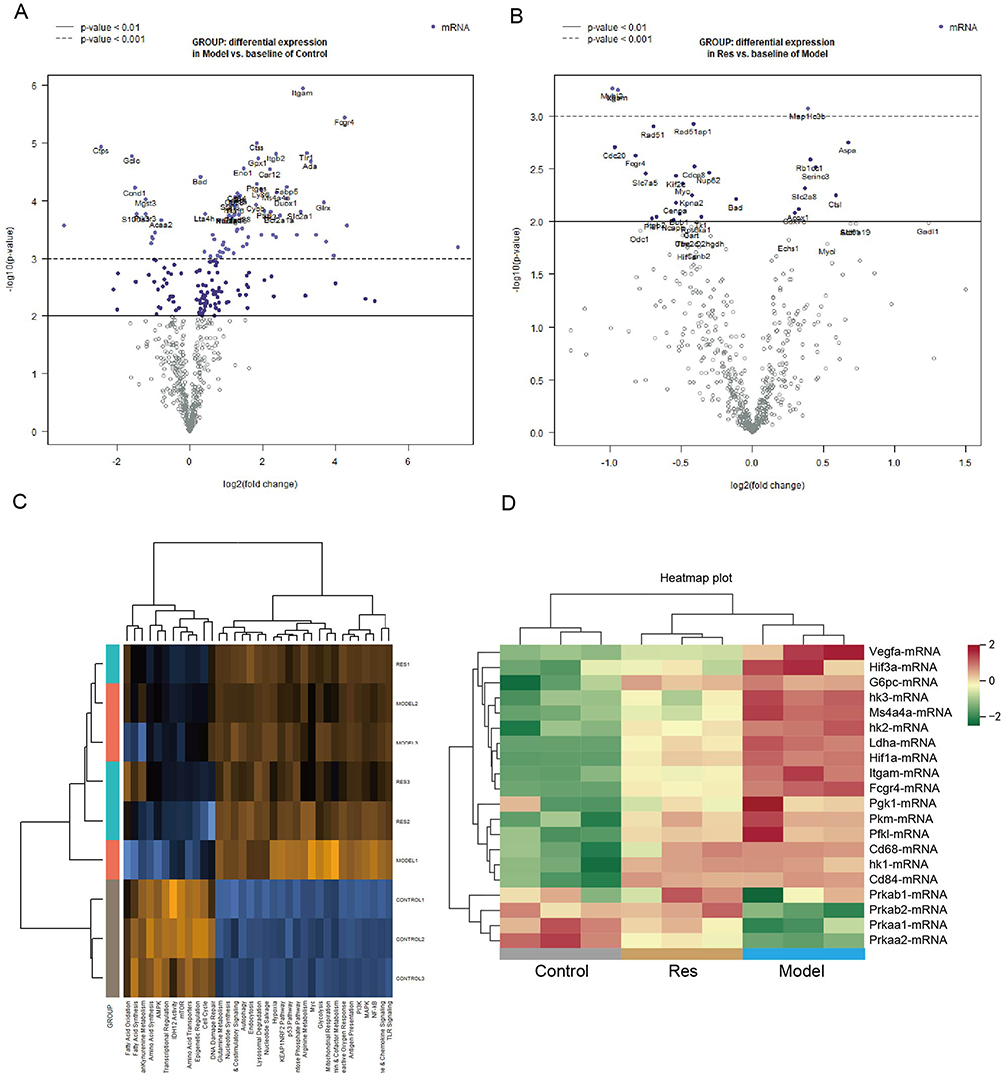 |
Figure 3 mRNA levels associated with metabolic processes and immunometabolism in mice of control, model, and Res groups. Notes: (A) Volcano plot displaying each gene’s -log10 (p-value) and log2 fold change with the selected covariate. The 40 most statistically significant genes are labeled in the plot. Control group vs model group. (B) Volcano plot displaying each gene’s -log10 (p-value) and log2 fold change with the selected covariate. The 40 most statistically significant genes are labeled in the plot. Res group vs model group. (C) Heatmap showing pathway score. Orange indicates high scores; blue indicates low scores. (D) The selected gene expression at log2 value among these three groups. Green represents gene down-regulation, and red represents gene up-regulation. Scores were displayed on the same scale via a Z-transformation. n = 3. |
Res Regulated the Expression of Metabolism-Related mRNA
To determine whether Res affects glycolysis, we measured the expression levels of mRNA in the skin by qRT-PCR. Consistent with previous mRNA sequencing results, the mRNA levels of glycolysis (ldha, hk1) were upregulated in the model mice, while Res down-regulated these mRNA (Figure 4A and B). The mRNA levels of FAO (cpt1a) and AMPK (prkaa1, prkaa2) were significantly decreased in the model mice, while Res increased the expression of these genes (Figure 4C–E). Spearman correlation analysis was performed to analyze the relationship between differential mRNA expression and the severity of psoriasis. The IL-17A had a positive association with ldha, and was negatively correlated with prkaa2 (Figure 4F and G). Therefore, these results underpin that glycolytic reprogramming has a crucial role in psoriasis. Res might regulate glycolytic reprogramming and activate AMPK to attenuate IMQ-induced psoriasis-like skin inflammation.
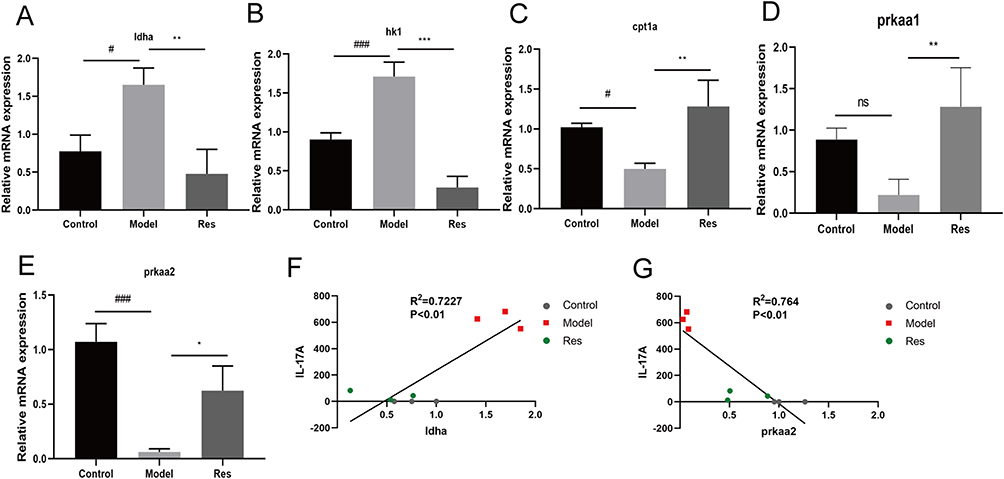 |
Figure 4 Res regulated the expression of mRNA. Notes: (A–E) Detection of ldha, hk1, cpt1a, prkaa1, and prkaa2 mRNA expression levels in skin lesions. (F and G) Spearman correlation analysis between IL-17A with ldha and prkaa2. Data are expressed as the mean ± SD. Ns, no significant, #P<0.05, ###P<0.001 vs control group. *P<0.05, **P<0.01, ***P<0.001 vs model group. |
Res Reduced the Infiltration of CD11b+cells, Especially Macrophages in the Lesion Skin
Interestingly, itgam, encoding CD11b, a differentiation marker for cells of the myeloid-monocytic lineage,16 was significantly regulated (Table 2 and Table 3), and Res inhibited the CD11b expression (Figure 5A). The obtained results suggested that the attenuation of inflammation may be associated with a decrease in the levels of infiltrating inflammatory cells expressing CD11b. We first analyzed CD45+CD11b+cells in mice skin lesions using flow cytometry. The percentage of CD11b+cells was significantly more upregulated in the model group than in the control group, whereas Res reduced the percentage of CD11b+cells in the mice skin as compared to the model group (Figure 5B).
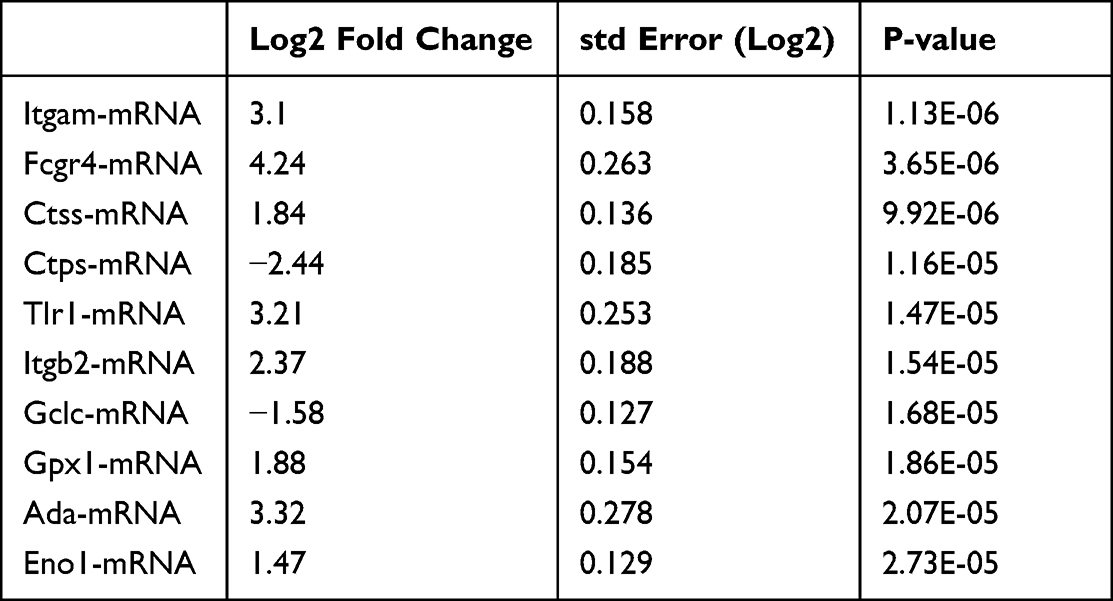 |
Table 2 Top 10 Most Statistically Significantly Differentially Expressed Genes Model vs Control |
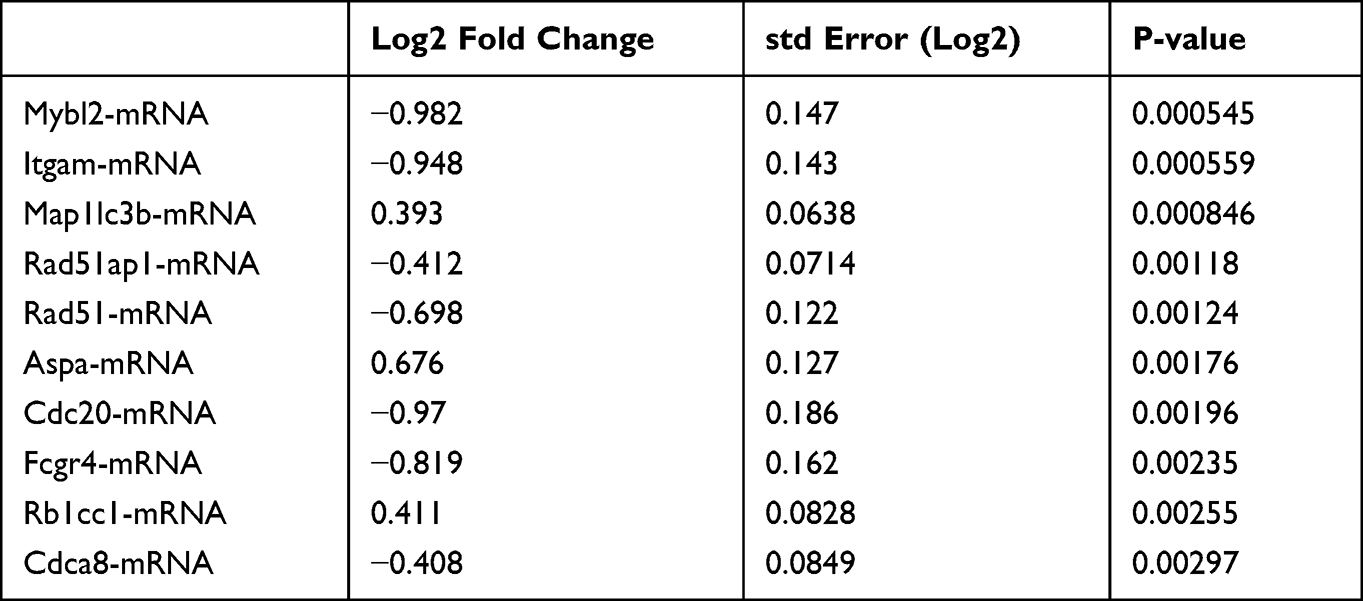 |
Table 3 Top 10 Most Statistically Significantly Differentially Expressed Genes Res vs Model |
 |
Figure 5 Res reduced the infiltration of macrophages. Notes: (A) The expression level of itgam mRNA and the representative Western blots of CD11b expression in the skin. (B) Percentage of CD45+CD11b+ cells of the skin in each group of mice. (C) Percentage of CD11b+F4/80+ macrophage of skin in each group of mice. (D) Percentage of dendritic cells of the skin in each group of mice. (E) Immunofluorescence staining of CD11b, F4/80, DAPI. Scale bar=100μm. (F) The expression level of ms4a4a mRNA in the skin. Data are expressed as mean ± SD (n = 3). Ns, no significant, ###P < 0.001 vs control group; *P < 0.05 and **P < 0.01 vs model group. |
Macrophages and dendritic cells play an essential role in the pathological process of psoriasis. Hence, CD11b+F4/80+ macrophages and F4/80−CD11c+dendritic cells were identified using flow cytometry. Compared with the control group, the percentage of macrophages in the model group was upregulated significantly, whereas Res inhibited the percentage of macrophages (Figure 5C). While the percentage of dendritic cells in the model group was also upregulated compared with the control group, Res had no obvious effect on the percentage of dendritic cells (Figure 5D). Immunofluorescence of CD11b and F4/80 in the skin lesions also indicated that Res reduced macrophages infiltration (Figure 5E). The plasma membrane tetraspan molecule ms4a4a, selectively expressed by macrophages,17 was upregulated in the model mice than control mice, and Res significantly inhibited its expression (Figure 5F). The aforementioned results suggested that the anti-psoriasis effect of Res might be related to reduced infiltration of macrophages in the lesion skin.
Res Inhibits Activation of Macrophages Stimulated by R848
Based on the flow cytometry experiment results, we assessed the effects of Res on macrophages in vitro. IMQ is an established TLR7/8 agonist used to induce a psoriasis-like skin inflammatory model. To maintain consistency between in vitro and in vivo studies, we chose R848, a TLR7/8 agonist, as a stimulant rather than routinely used LPS. First, to observe the toxicity of Res, Raw264.7 cells were treated with different concentrations of Res for 12 h, and their cell viability was measured using the CCK-8 assay. Res ranged from 3.125 to 200 μM, showing no effect on cell viability (Figure 6A). Therefore, 50, 100, and 200 μM were used in the following in vitro experiments. Increased secretion levels of pro-inflammatory cytokines were observed in R848-stimulated Raw264.7 cells. At 50, 100, and 200 μM, Res inhibited the secretion of IL-23, IL-6, and TNF-α to varying degrees (Figure 6B–D). In addition, Res inhibited the release of reactive oxygen species (ROS) in R848-induced Raw264.7 cells (Figure 6E).
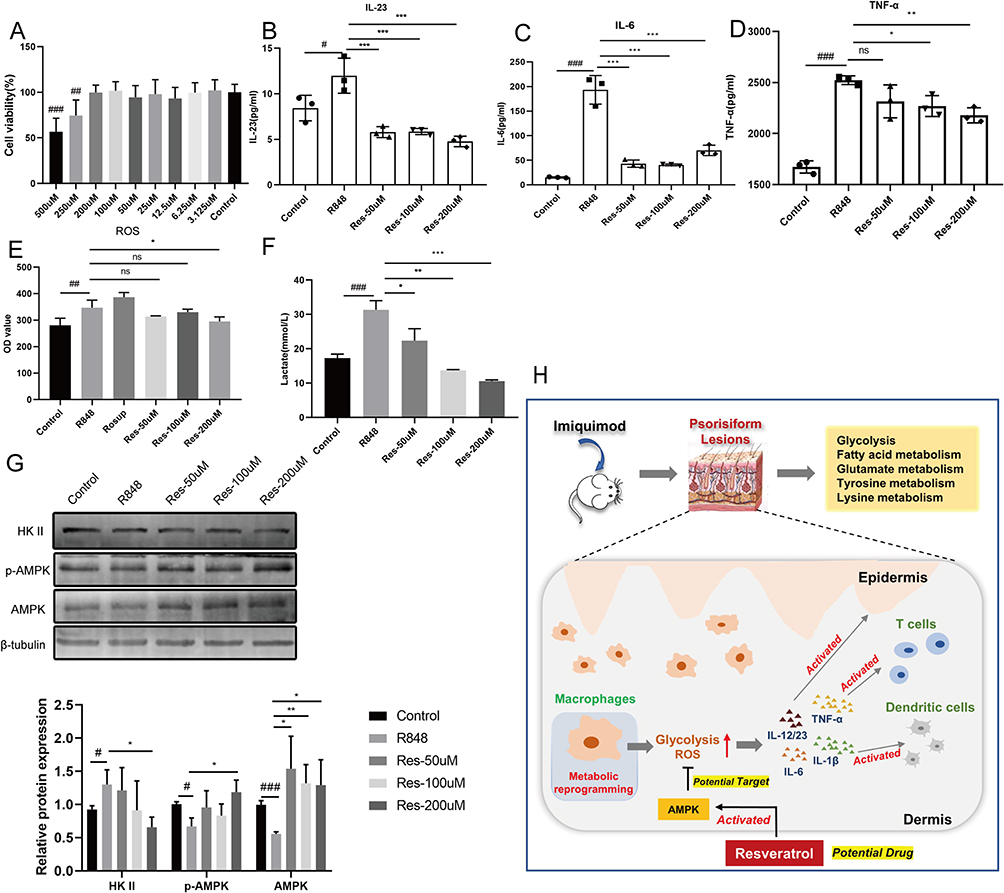 |
Figure 6 Res inhibits R848-stimulated activation of macrophages. Notes: (A) The effect of Res on the cell viability of Raw264.7 cells. (B–D) IL-23, IL-6, and TNF-α levels were shown in control, R848, and Res groups. (E) The levels of ROS. (F) The level of lactate production. (G) The expression of HK II, p-AMPK and AMPK. (H) Schematic view of glycolytic reprogramming of macrophages in IMQ-induced psoriasis-like mouse model and is a potential therapeutic target. Data were expressed as the mean ± SD. Ns, no significant, #P < 0.05, ##P < 0.01 and ###P < 0.001 vs control group; *P < 0.05, **P < 0.01 and ***P < 0.001 vs R848 group. |
Res Regulates Macrophages Glycolytic Reprogramming by Activating AMPK
Lactate is a terminal glycolysis product from pyruvate catalyzed by LDHA.18 The lactate production of R848-stimulated macrophages was higher than that in the control group, whereas Res attenuated lactate production (Figure 6F). The expression of HK II in the R848 group was increased, while Res-200 μM significantly inhibited the expression of HK II. AMPK is a negative regulator of aerobic glycolysis.19 In R848-stimulated macrophages, the expression levels of AMPK and p-AMPK were significantly inhibited compared to the control group. In contrast, Res increased AMPK and p-AMPK levels (Figure 6G). These results suggested that Res may regulate glycolysis reprogramming of macrophages by activating AMPK to reduce inflammation (Figure 6H).
Discussion
Psoriasis is a chronic, relapsing, inflammatory skin disease characterized by systemic immune dysregulation. Abundant macrophage infiltration has been observed in psoriasis lesions.20 As for the origin of macrophages, there is broad agreement that macrophages in health and disease originate from circulating monocytes.21 In mice, skin, lung, heart, and kidney tissue-resident macrophages (MTR) were found to originate from the yolk sac or fetal liver.22 In humans and mice, skin mononuclear phagocytes consist of Langerhans cells and dermal macrophages, a mix of cells derived from bone marrow monocytes and embryonic precursors.23,24 The primary function of macrophages is phagocytosis, which serves as the first defensive line of immunity. In addition, macrophages produce many inflammatory mediators that activate other immune cells to facilitate the induction of a robust immune response. In psoriasis, activated macrophages produce TNF-a, IL-23, and IL-1β to activate T cells, leading to abnormal proliferation in keratinocytes. Reportedly, macrophages are the most prominent population in normal and psoriatic skin and are the principal immune lymphocyte cells in hTNF-mediated psoriasis.7 Recently, single-cell RNA sequencing revealed that macrophages were the main group of immune cells in IMQ-induced psoriasis mice.25 Our study also found that the percentage of macrophages in skin lesions was prominently elevated, about 61.29%, which is similar to what was previously reported. The extensive infiltration of macrophages in psoriasis lesions deserves further investigation to elucidate the relationship between macrophage levels and activities and the pathogenesis of psoriasis.
Res has been shown to alleviate inflammation in skin lesions of psoriasis; however, its mechanism of action is still unclear. Studies have shown that Res decreased the expression of many M1 pro-inflammatory cytokines (TNF-α, IL-1β, and IL-6) but increased the regulatory T cells infiltration. In addition, Res elevated the level of p-AMPK.26 In LPS-induced murine acute lung injury, Res decreased CD45+CD206− subtype macrophage levels by the SOCS3 signaling pathway.27 Furthermore, the anti-inflammatory and antioxidant effects of Res have been widely explored and documented.26
Here, we found that Res alleviated the IMQ-induced psoriasis-like mouse model inflammation reaction. Based on customized NanoString panels analysis results, we found Itgam, which has been identified that associated with systemic lupus erythematosus.28,29 Itgam encodes CD11b (also known as integrin α M) which couples with CD18 (also known as integrin β2) to form macrophage antigen 1 (MAC1).30 According to literature reports, CD11b levels in patients with psoriasis were lower than in healthy controls.30 However, CD11b levels in patients with pustular psoriasis were also evidenced to be higher than in patients with psoriasis vulgaris.31 Hruska found that the MiR-21 binding site SNP within Itgam was associated with psoriasis susceptibility in women.32 Neutrophils in patients with acute psoriasis were associated with an increase in CD11b/CD18 expression,33 which may increase their migration capacity toward keratinocytes.34 We found that the Itgam/CD11b and ms4a4a in the IMQ-induced psoriasis-like mouse model were higher than in the control mice. Res significantly decreased the expression of CD11b and the percentage of CD11b+cells in the skin lesions. We further established that Res reduced the infiltration of CD11b+F4/80+macrophages in the skin lesions. Additionally, Res reduced the levels of the macrophage-related pro-inflammatory factors (IL-6, IL-1β, and TNF-α) secretion both in vivo and in vitro. These results suggest that Res alleviate psoriasis may by inhibiting macrophages infiltration and reducing the secretion of macrophage-associated cytokines. Further, we will consider incorporating depletion experiments to evaluate the impact of macrophages absence on the observed outcomes.
Immune inflammation reactions are closely related to cellular metabolism. Activated immune cells undergo a metabolic switch to aerobic glycolysis, a phenomenon known as the Warburg effect.35 In psoriasis, cell proliferation and epidermal thickening increase oxygen consumption and supply. Psoriasis lesions are hypoxic, and thus hifs are strongly activated.36 Our experimental data also confirmed that the glycolysis levels and hypoxia were increased in psoriasis. Glycolysis is vital to Th17/Th1 cell differentiation and keratinocyte proliferation in psoriasis pathogenesis.37,38 Dimethyl fumarate, an immunomodulatory drug used to treat multiple sclerosis and psoriasis, downregulates aerobic glycolysis in activated myeloid and lymphoid cells.35 Thus, glycolysis is considered a potential therapeutic target in autoimmune diseases. AMPK, a central regulator of energy metabolism, is a drug target in different therapeutic areas, including metabolic diseases, cancer, pain, and chronic kidney disease.39 Metformin, aspirin, berberine, and Res activate AMPK, suggesting AMPK may mediate their effects, at least in part. AMPK is a negative regulator of aerobic glycolysis.40 AMPK activity was inhibited in both the psoriasis patients and the IMQ-induced psoriasis-like mouse model.41,42 It is noteworthy that impaired AMPK in macrophages has been shown to be associated with higher production of pro-inflammatory cytokines while salicylates ameliorate intestinal inflammation by activating macrophage AMPK.43 We also found that R848-stimulated macrophages had increased glycolytic activity and inhibited AMPK. However, Res could activate AMPK to regulate glycolysis to inhibit pro-inflammatory cytokine secretion.
This study has some limitations. First, the NanoString panel analysis was customized, including mRNAs associated with metabolic processes and immunometabolism rather than all mRNAs. Second, we detected only macrophages and dendritic cells in skin lesions but did not investigate neutrophils. In future studies, we will focus on the role of neutrophils in psoriasis and the related mechanisms of drug therapy. Furthermore, many metabolic pathways were changed in the psoriasis model and Res-treated mice, which we did not further investigate. Further exploration of these pathways may lead to the discovery of potential therapeutic targets.
Conclusion
Our study further elucidates the mechanism by which Res improves psoriasis, possibly by reducing macrophage infiltration and inhibiting glycolysis. Res showed promising prospects for further research and clinical application.
Abbreviations
AMPK, AMP-activated protein kinase; Arg1; Arginase-1; Cpt1a, carnitine palmitoyltransferase 1A; FAO, fatty acid oxidation; HK, hexokinase; IL17A, Interleukin-17A; IMQ, imiquimod; LDHA, lactate dehydrogenase A; OXPHOS, oxidative phosphorylation; PDEs, Phosphodiesterases; Res, resveratrol; TCA, tricarboxylic acid; TLR, Toll-like receptor; ROS, reactive oxygen species.
Acknowledgments
This work was supported by the National Natural Science Foundation of China (No. 82074180, 82074435) and supported by Beijing Key Laboratory of Clinic and Basic Research with Traditional Chinese Medicine on Psoriasis for technical assistance.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Disclosure
The authors report no conflicts of interest in this work.
References
1. Griffiths CEM, Armstrong AW, Gudjonsson JE, Barker J. Psoriasis. Lancet. 2021;397(10281):1301–1315. doi:10.1016/S0140-6736(20)32549-6
2. Nair RP, Duffin KC, Helms C, et al. Genome-wide scan reveals association of psoriasis with IL-23 and NF-kappaB pathways. Nat Genet. 2009;41(2):199–204. doi:10.1038/ng.311
3. Xing X, Liang Y, Sarkar MK, et al. IL-17 responses are the dominant inflammatory signal linking inverse, erythrodermic, and chronic plaque psoriasis. J Invest Dermatol. 2016;136(12):2498–2501. doi:10.1016/j.jid.2016.07.008
4. O’Neill LA, Kishton RJ, Rathmell J. A guide to immunometabolism for immunologists. Nat Rev Immunol. 2016;16(9):553–565. doi:10.1038/nri.2016.70
5. Stratis A, Pasparakis M, Rupec RA, et al. Pathogenic role for skin macrophages in a mouse model of keratinocyte-induced psoriasis-like skin inflammation. J Clin Invest. 2006;116(8):2094–2104. doi:10.1172/JCI27179
6. Wang H, Peters T, Kess D, et al. Activated macrophages are essential in a murine model for T cell-mediated chronic psoriasiform skin inflammation. J Clin Invest. 2006;116(8):2105–2114. doi:10.1172/JCI27180
7. Leite Dantas R, Masemann D, Schied T, et al. Macrophage-mediated psoriasis can be suppressed by regulatory T lymphocytes. J Pathol. 2016;240(3):366–377. doi:10.1002/path.4786
8. Ma Y, Li D, Liu W, et al. Resveratrol on the metabolic reprogramming in liver: implications for advanced atherosclerosis. Front Pharmacol. 2021;12:747625. doi:10.3389/fphar.2021.747625
9. Gong L, Guo S, Zou Z. Resveratrol ameliorates metabolic disorders and insulin resistance in high-fat diet-fed mice. Life Sci. 2020;242:117212. doi:10.1016/j.lfs.2019.117212
10. Tennen RI, Michishita-Kioi E, Chua KF. Finding a target for resveratrol. Cell. 2012;148(3):387–389. doi:10.1016/j.cell.2012.01.032
11. Timmers S, Konings E, Bilet L, et al. Calorie restriction-like effects of 30 days of resveratrol supplementation on energy metabolism and metabolic profile in obese humans. Cell Metab. 2011;14(5):612–622. doi:10.1016/j.cmet.2011.10.002
12. Cheng CY, Lin YK, Yang SC, et al. Percutaneous absorption of resveratrol and its oligomers to relieve psoriasiform lesions: in silico, in vitro and in vivo evaluations. Int J Pharm. 2020;585:119507. doi:10.1016/j.ijpharm.2020.119507
13. Khatoon K, Ali A, Ahmad FJ, et al. Novel nanoemulsion gel containing triple natural bio-actives combination of curcumin, thymoquinone, and resveratrol improves psoriasis therapy: in vitro and in vivo studies. Drug Deliv Transl Res. 2021;11(3):1245–1260. doi:10.1007/s13346-020-00852-y
14. Kjaer TN, Thorsen K, Jessen N, Stenderup K, Pedersen SB, Bobé P. Resveratrol ameliorates imiquimod-induced psoriasis-like skin inflammation in mice. PLoS One. 2015;10(5):e0126599. doi:10.1371/journal.pone.0126599
15. Oliveira ALB, Monteiro VVS, Navegantes-Lima KC, et al. Resveratrol role in autoimmune disease-a mini-review. Nutrients. 2017;9(12):1306. doi:10.3390/nu9121306
16. Abram CL, Lowell CA. The ins and outs of leukocyte integrin signaling. Annu Rev Immunol. 2009;27(1):339–362. doi:10.1146/annurev.immunol.021908.132554
17. Mattiola I, Tomay F, De Pizzol M, et al. The macrophage tetraspan MS4A4A enhances dectin-1-dependent NK cell-mediated resistance to metastasis. Nat Immunol. 2019;20(8):1012–1022. doi:10.1038/s41590-019-0417-y
18. Xu K, Yin N, Peng M, et al. Glycolytic ATP fuels phosphoinositide 3-kinase signaling to support effector T helper 17 cell responses. Immunity. 2021;54(5):976–987 e977. doi:10.1016/j.immuni.2021.04.008
19. Rutherford C, Speirs C, Williams JJ, et al. Phosphorylation of Janus kinase 1 (JAK1) by AMP-activated protein kinase (AMPK) links energy sensing to anti-inflammatory signaling. Sci Signal. 2016;9(453):ra109. doi:10.1126/scisignal.aaf8566
20. Nakamizo S, Dutertre CA, Khalilnezhad A, et al. Single-cell analysis of human skin identifies CD14+ type 3 dendritic cells co-producing IL1B and IL23A in psoriasis. J Exp Med. 2021;218(9). doi:10.1084/jem.20202345
21. van Furth R, Raeburn JA, van Zwet TL. Characteristics of human mononuclear phagocytes. Blood. 1979;54(2):485–500. doi:10.1182/blood.V54.2.485.485
22. Locati M, Curtale G, Mantovani A. Diversity, mechanisms, and significance of macrophage plasticity. Annu Rev Pathol. 2020;15:123–147. doi:10.1146/annurev-pathmechdis-012418-012718
23. Kolter J, Feuerstein R, Zeis P, et al. A subset of skin macrophages contributes to the surveillance and regeneration of local nerves. Immunity. 2019;50(6):1482–1497 e1487. doi:10.1016/j.immuni.2019.05.009
24. Nobs SP, Kopf M. Tissue-resident macrophages: guardians of organ homeostasis. Trends Immunol. 2021;42(6):495–507. doi:10.1016/j.it.2021.04.007
25. Jin S, Wang Y, Qie C, et al. Single-cell RNA sequencing reveals the immune cell profiling in IMQ induced psoriasis-like model. J Inflamm Res. 2022;15:5999–6012. doi:10.2147/JIR.S379349
26. Shabani M, Sadeghi A, Hosseini H, et al. Resveratrol alleviates obesity-induced skeletal muscle inflammation via decreasing M1 macrophage polarization and increasing the regulatory T cell population. Sci Rep. 2020;10(1):3791. doi:10.1038/s41598-020-60185-1
27. Hu L, Chen Z, Li L, Jiang Z, Zhu L. Resveratrol decreases CD45(+) CD206(-) subtype macrophages in LPS-induced murine acute lung injury by SOCS3 signalling pathway. J Cell Mol Med. 2019;23(12):8101–8113. doi:10.1111/jcmm.14680
28. Nath SK, Han S, Kim-Howard X, et al. A nonsynonymous functional variant in integrin-alpha(M) (encoded by ITGAM) is associated with systemic lupus erythematosus. Nat Genet. 2008;40(2):152–154. doi:10.1038/ng.71
29. Rhodes B, Furnrohr BG, Roberts AL, et al. The rs1143679 (R77H) lupus associated variant of ITGAM (CD11b) impairs complement receptor 3 mediated functions in human monocytes. Ann Rheum Dis. 2012;71(12):2028–2034. doi:10.1136/annrheumdis-2012-201390
30. Cao LY, Soler DC, Debanne SM, et al. Psoriasis and cardiovascular risk factors: increased serum myeloperoxidase and corresponding immunocellular overexpression by Cd11b(+) CD68(+) macrophages in skin lesions. Am J Transl Res. 2013;6(1):16–27.
31. Sjogren F, Ljunghusen O, Baas A, Coble BI, Stendahl O. Expression and function of beta 2 integrin CD11B/CD18 on leukocytes from patients with psoriasis. Acta Derm Venereol. 1999;79(2):105–110. doi:10.1080/000155599750011291
32. Hruska P, Kuruczova D, Vasku V, Bienertova-Vasku J, Garcia de Frutos P. MiR-21 binding site SNP within ITGAM associated with psoriasis susceptibility in women. PLoS One. 2019;14(6):e0218323. doi:10.1371/journal.pone.0218323
33. Kapuscinska R, Wysocka J, Zelazowska B, Niczyporuk W. [Evaluation of neutrophils adhesive molecules CD11b/CD18 in the course of photochemotherapy (PUVA) for psoriasis vulgaris]. Pol Merkur Lekarski. 2004;16(96):547–550. Polish.
34. Liu XT, Shi ZR, Lu SY, et al. Enhanced migratory ability of neutrophils toward epidermis contributes to the development of psoriasis via crosstalk with keratinocytes by releasing IL-17A. Front Immunol. 2022;13:817040. doi:10.3389/fimmu.2022.817040
35. Kornberg MD, Bhargava P, Kim PM, et al. Dimethyl fumarate targets GAPDH and aerobic glycolysis to modulate immunity. Science. 2018;360(6387):449–453. doi:10.1126/science.aan4665
36. Rosenberger C, Solovan C, Rosenberger AD, et al. Upregulation of hypoxia-inducible factors in normal and psoriatic skin. J Invest Dermatol. 2007;127(10):2445–2452. doi:10.1038/sj.jid.5700874
37. Lin Y, Xue K, Li Q, et al. Cyclin-dependent kinase 7 promotes Th17/Th1 cell differentiation in psoriasis by modulating glycolytic metabolism. J Invest Dermatol. 2021;141(11):2656–2667 e2611. doi:10.1016/j.jid.2021.04.018
38. Liu YZ, Xu MY, Dai XY, et al. Pyruvate kinase M2 mediates glycolysis contributes to psoriasis by promoting keratinocyte proliferation. Front Pharmacol. 2021;12:765790. doi:10.3389/fphar.2021.765790
39. Steinberg GR, Carling D. AMP-activated protein kinase: the current landscape for drug development. Nat Rev Drug Discov. 2019;18(7):527–551. doi:10.1038/s41573-019-0019-2
40. Faubert B, Boily G, Izreig S, et al. AMPK is a negative regulator of the Warburg effect and suppresses tumor growth in vivo. Cell Metab. 2013;17(1):113–124. doi:10.1016/j.cmet.2012.12.001
41. Garcin G, Guiraud I, Lacroix M, et al. AMPK/HuR-driven IL-20 post-transcriptional regulation in psoriatic skin. J Invest Dermatol. 2015;135(11):2732–2741. doi:10.1038/jid.2015.282
42. Yan K, Xu W, Huang Y, et al. Methotrexate restores the function of peripheral blood regulatory T cells in psoriasis vulgaris via the CD73/AMPK/mTOR pathway. Br J Dermatol. 2018;179(4):896–905. doi:10.1111/bjd.16560
43. Banskota S, Wang H, Kwon YH, et al. Salicylates ameliorate intestinal inflammation by activating macrophage AMPK. Inflamm Bowel Dis. 2021;27(6):914–926. doi:10.1093/ibd/izaa305
 © 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Study on the Key Genes and Molecular Mechanisms of IL-27 Promoting Keratinocytes Proliferation Based on Transcriptome Sequencing
Wu Z, Yang Q, Xu K, Wu J, Yang B
Clinical, Cosmetic and Investigational Dermatology 2023, 16:1457-1472
Published Date: 7 June 2023
Psoriasis and Leprosy: An Arcane Relationship
Ge G, Shang J, Gan T, Chen Z, Pan C, Mei Y, Long S, Wu A, Wang H
Journal of Inflammation Research 2023, 16:2521-2533
Published Date: 14 June 2023
Inhibition of Key Glycolytic Enzyme Hexokinase 2 Ameliorates Psoriasiform Inflammation in vitro and in vivo
Zhuang L, Ma W, Jiao J
Clinical, Cosmetic and Investigational Dermatology 2023, 16:3229-3239
Published Date: 9 November 2023
Layered Double Hydroxide Nanoparticles Loaded with Resveratrol Inhibit Glycolysis and Show Efficacy in the Treatment of Breast Cancer
Geng C, Yan L, Li Y, Li H, Ji Y, Xiao Y, Wang Z, Chen X, Chen C, Yang Q, Tang B, Wang W
International Journal of Nanomedicine 2025, 20:3423-3444
Published Date: 17 March 2025