")
Back to Journals » Cancer Management and Research » Volume 12
Regulating DSC2 Expression Affects the Proliferation and Apoptosis of Prostate Cancer Cells
Received 22 July 2020
Accepted for publication 14 October 2020
Published 9 November 2020 Volume 2020:12 Pages 11453—11462
DOI https://doi.org/10.2147/CMAR.S273394
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Rudolph Navari
Feng Jiang,1 Pengfei Wu2
1Department of Urology, Shanghai Pudong New Area People’s Hospital, Shanghai 201200, People’s Republic of China; 2Department of Urology, Tenth People’s Hospital of Tongji University, Shanghai 200072, People’s Republic of China
Correspondence: Pengfei Wu
Department of Urology, Tenth People’s Hospital of Tongji University, 301 Yanchang Road, Jing’an District, Shanghai 200072, People’s Republic of China
Email [email protected]
Background: Prostate cancer threatens the life and health of men in China. Desmocollin-2 (DSC2) is a member of DSC family, abnormal expression of which can affect the invasion and metastasis of tumor cells. The aim of this study was to investigate the role of DSC2 in prostate cancer.
Materials and Methods: Regulating DSC2 expression in prostate cancer cells was conducted with transfection. The expression of DSC2, apoptosis-related proteins, cell cycle-related proteins and E-cadherin (E-cad)/β-catenin pathway was detected by Western blot analysis. The proliferation, clone formation ability, migration, invasion and apoptosis of transfected cells were in turn detected by cell counting kit-8 (CCK-8) assay, clone formation assay, wound healing assay, transwell assay and flow cytometry analysis.
Results: DSC2 expression was increased in prostate cancer cells compared with RWPE-1 cells. Inhibition of DSC2 promoted the proliferation, clone formation ability, migration and invasion while suppressed apoptosis of LNCaP cells and PC-3 cells. Inhibition of DSC2 affected the expression of apoptosis-related proteins and cell cycle-related proteins according to the changes of apoptosis and proliferation. Furthermore, inhibition of DSC2 up-regulated the expression of p-β-catenin and EGFR while down-regulated the expression of E-cad. DSC2 overexpression exerted the opposite effect of inhibition of DSC2 on LNCaP cells and PC-3 cells.
Conclusion: DSC2 expression was increased in prostate cancer cells. In addition, inhibition of DSC2 promoted the proliferation, clone formation ability, migration and invasion while suppressed apoptosis of LNCaP cells and PC-3 cells, which provided the fundamental basis for treatment of prostate cancer.
Keywords: DSC2, proliferation, apoptosis, prostate cancer cell
Introduction
Prostate cancer is the most common malignant tumor among men in western countries such as the United States and the United Kingdom, ranking second only to lung cancer in the global incidence.1 Although patients with early stage prostate cancer have a good prognosis, most of the patients are treated in the middle and late stage. The 5-year survival rate of patients with advanced stage prostate cancer is less than 30%, and about 90% of the patients die from tumor recurrence and metastasis.2 In the past 10 years, with the change of Chinese people’s western lifestyle, the incidence and death rate of male prostate cancer in China has been on a continuous and rapid rise.3 At present, prostate cancer has gradually become one of the common tumors affecting the life and health of men in China.
Desmosome is a unique intercellular adhesion structure of epithelial cells, which plays the role of intercellular connection. Desmosome is not only the main anchoring site of intermediate filaments on the cell surface, but also plays the role in conducting intra-cellular signaling regulating cell behaviors. Desmosome consists of two kinds of proteins, one is desmosome cadherin including desmoglein (DSG) and desmocollin (DSC) and the other is desmoplakin including desmoplakin and plakoglobin.4 DSC2, a member of DSC family, is expressed in a variety of normal human tissues, cancer tissues and cell lines.5 DSC2 has an interaction site of inter-cell adhesion, and structural integrity of tissues and normal adhesion between cells are damaged when the tumor occurs. It can be inferred that invasion and metastasis of tumor cells may be related to the abnormal expression of DSC2.6 DSC2 expression is abnormally down-regulated in pancreatic ductal adenocarcinoma,7 colorectal carcinoma,8 urothelial carcinoma9 and lung cancer.10
Through GEPIA (http://gepia.cancer-pku.cn/), DSC2 is abnormally expressed in many cancer tissues compared with adjacent tissues. Also, DSC2 expression is increased in tumor tissues compared with adjacent tissues in many other cancers. DSC2 expression was also increased in prostate cancer tissues and the lower the DSC2 expression is, the less conducive the disease-free survival (DFS) of patients is. Therefore, we aimed to investigate the role of DSC2 in prostate cancer and preliminarily explain the significance of its high expression in tumor tissues.
Materials and Methods
Cell Culture
RWPE-1, DU-145, VCaP, LNCaP and PC-3 cells were obtained from American Type Culture Collection (Rockville, MD, USA). RWPE-1 cells were cultured in F-12K medium (Gibco BRL Co. Ltd., USA) containing L-glutamine. DU-145, VCaP, LNCaP and PC-3 cells were cultured in RPMI-1640 medium (HyClone, Logan, UT, USA). All medium must be supplemented with 100 U/mL penicillin, 100 U/mL streptomycin and 10% fetal bovine serum (FBS). The cells were routinely cultured in a saturated humidity incubator at 37°C with 5% CO2 and had digestive transfer culture with 0.05% EDTA.
Cell Transfection
Cells from logarithmic growth stage were inoculated in a 24-well plate and cultured under conventional condition. Cell transfection was carried out when the confluence reached 60%-70%. Strictly following the construction of LipofectamineTM2000, cells were transfected with empty vector, ShRNA-DSC2-1, ShRNA-DSC2-2, OverExp-DSC2-1 and OverExp-DSC2-2. After transfection for 24 h, RT-qPCR analysis detected the DSC2 expression to determine the transfection effects. Cells in the control group received no treatment.
RT-qPCR Analysis
Total RNA of prostate cancer cells was extracted according to the instructions of TRIzol reagent kit (Invitrogen; Thermo Fisher Scientific, Inc.). After the purity and content of RNA were evaluated, cDNA was synthesized by reverse transcription. cDNA synthesis was performed according to the instructions of the reverse transcription kit (Takara Biotechnology Co., Ltd., Dalian, China). Using cDNA as template, the reaction system was conducted according to the RT-qPCR kit instructions for PCR reaction. PCR reaction procedure is as follows: 94°C for 30 s, 58°C for 30 s, 72°C for 30 s, a total of 40 cycles. Taking GAPDH as internal reference, relative RNA expression of DSC2 was calculated with 2−ΔΔCt method.
Cell Counting Kit-8 (CCK-8)
Transfected cells were collected at 24 h, 48 h and 72 h and respectively seeded into a 96-well plate with 5000 cells/well for 24 h culture. Ten microliter CCK-8 solution was added to each well and then cells were incubated at 37°C for another 4 h. The optical density (OD) value at 450 nm was determined by the Multiskan FC enzyme micro-plate reader.
Clone Formation Assay
Transfected cells were collected and seeded into a 6-well plate with 200 cells/well. After conventional culture for two weeks, the culture was terminated when visible clones appeared in the 6-well plate. After PBS washing for two times, the cells were fixed with 4% neutral methanol for 15 min, and then stained with Wright-Giemsa complex staining solution for 30 min. The cells were slowly washed by running water and then dried by natural air. The clones were observed and photographed by microscope.
Transwell Assay
Matrigel was smeared evenly at the bottom of upper chamber inside, which become a membrane overnight. The next day, transfected cells for 24 h were collected and re-suspended with serum-free medium to prepare single-cell suspension. Two hundred microliter single-cell suspension containing 2×105 cells was added to the upper chamber and 500 μL DMEM medium containing 10% FBS was added to the lower chamber. After continuous culture for 24 h, the upper chamber was taken out and a cotton swab gently wiped cells at the upper surface of the membrane. The remaining cells were fixed with 4% paraformaldehyde for 10 min and treated with crystal violet (10 g/L) staining for 30 min. After drying, the cells were observed and photographed under the microscope.
Wound Healing Assay
The cells were collected after transfection for 24 h and seeded in a 6-well plate for conventional culture. After cells were fused completely, 10 μL pipette tip was used to scratch in a straight line vertically in the bottom of wells. After PBS washing for three times, the cells were incubated for 24 h. The scratches at 0 h and 24 h were observed and photographed by a microscope.
Flow Cytometry Analysis
Transfected cells were collected, seeded into a 6-well plate and cultured for 24 h. After trypsin digestion, cells were washed with PBS for three times and re-suspended with 100 μL binding buffer. Each tube was added with 5 μL PI and 5 μL Annexin V-FITC to treat the cells on the ice for 15 min, followed by the addition of 400 μL binding buffer. Finally, the apoptotic cell rate was analyzed by flow cytometry within 1 h.
Western Blot Analysis
After transfected cells grew into 90% confluence, medium was removed and 200 μL RIPA lysis buffer was added to extract the protein on ice for 20 min. The mixture was transferred to a centrifuge tube which was centrifuged at 2000 r/min for 10 min to separate the supernatant. After quantitative analysis by BCA kit (Abcam, UK), 20 μg protein was uploaded, separated by 10% SDS-PAGE with electrophoresis. After protein separation by gel electrophoresis, protein was transferred to PVDF membrane. Five percent skim milk sealed the membrane at room temperature for 2 h. Next, PVDF membrane was incubated with primary antibodies including DSC2, Bax, B-cell lymphoma 2 (Bcl-2), Caspase-3 (Casp-3), c-Caspase-3 (c-Casp-3), Cyclin D1, Cyclin-dependent kinase 2 (CDK2), Cyclin B1, CDK1, β-catenin, p-β-catenin, Epidermal growth factor receptor (EGFR) and E-cadherin (E-cad) at 4°C overnight. The following day, after PVDF membrane was washed with PBS buffer for three times, PVDF membrane was incubated with horseradish peroxidase-linked IgG second antibody at 25°C for 2 h. After membrane exposed and developed, a quantitative gel imaging software Quantity One was applied to quantify each band in the electrophoresis map.
Statistical Analysis
Data were expressed as mean± standard deviation (SD). SPSS version 21.0 was used for statistical analysis and p<0.05 was considered statistically significant. The results were analyzed using the one-way ANOVA with Tukey’s test.
Results
DSC2 Protein Expression in Prostate Cancer Cells
DSC2 protein expression was increased in prostate cancer cells compared with RWPE-1 cells. The protein expression of DSC2 in LNCaP cells was the highest and in PC-3 cells was the lowest among prostate cancer cells. Therefore, LNCaP cells and PC-3 cells were chosen for the next study (Figure 1).
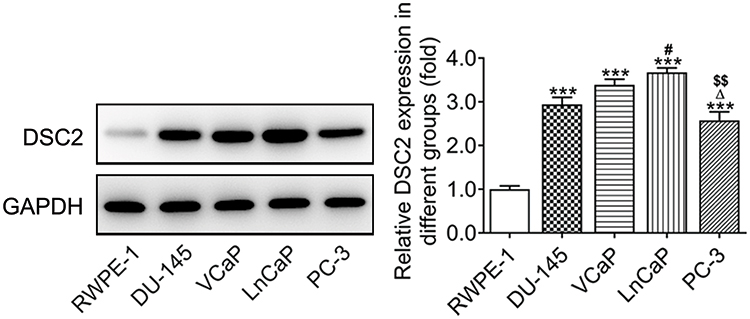 |
Figure 1 DSC2 protein expression in prostate cancer cells. DSC2 protein expression in RWPE-1 cells and prostate cancer cells was detected by Western blot analysis. ***P<0.001 vs. RWPE-1 group. #P<0.05 vs. DU-145 group. ∆P<0.05 vs. VCaP group. $$P<0.01 vs. LnCaP group. |
Plasmids of ShRNA-1 and OverExp-1 Were Chosen
After transfection, DSC2 expression was down-regulated in LNCaP cells (Figure 2A) and PC-3 cells (Figure 2B) transfected with ShRNA-DSC2-1/2 and up-regulated in LNCaP cells (Figure 2A) and PC-3 cells (Figure 2B) transfected with OverExp-DSC2-1/2. DSC2 expression in LNCaP cells and PC-3 cells transfected with ShRNA-DSC2-1 was lower than that transfected with ShRNA-DSC2-2, and DSC2 expression in LNCaP cells and PC-3 cells transfected with OverExp-DSC2-1 was higher than that transfected with OverExp-DSC2-2. Therefore, plasmids of ShRNA-1 and OverExp-1 were selected.
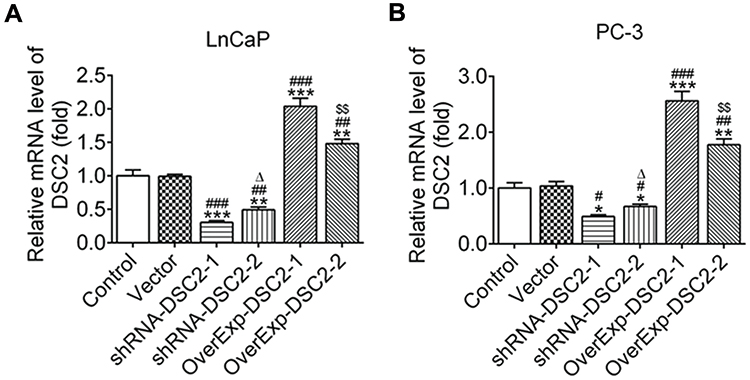 |
Figure 2 Plasmids of ShRNA-1 and OverExp-1 were chosen. (A) The mRNA expression of DSC2 in LNCaP cells was analyzed by RT-qPCR analysis. **P<0.01 and ***P<0.001 vs. Control group. ##P<0.01 and ###P<0.001 vs. Vector group. ∆P<0.05 vs. ShRNA-DSC2-1 group. $$P<0.01 vs. OverExp-DSC2-1 group. (B) The mRNA expression of DSC2 in PC-3 cells was analyzed by RT-qPCR analysis. *P<0.05, **P<0.01 and ***P<0.001 vs. Control group. #P<0.05, ##P<0.01 and ###P<0.001 vs. Vector group. ∆P<0.05 vs. ShRNA-DSC2-1 group. $$P<0.01 vs. OverExp-DSC2-1 group. |
DSC2 Expression Affects the Proliferation and Clone Formation Ability of LNCaP Cells and PC3 Cells
With the extension of transfection time from 24 h to 72 h, inhibition of DSC2 promoted the proliferation of LNCaP cells. DSC2 overexpression did not obviously affect the proliferation of LNCaP cells while proliferation of LNCaP cells showed a downward trend (Figure 3A). The proliferation of PC3 cells was also enhanced by inhibition of DSC2 while inhibited by DSC2 overexpression from 24 h to 72 h (Figure 3B). Inhibition of DSC2 increased the clone formation ability while DSC2 overexpression decreased the clone formation ability of LNCaP cells (Figure 3C and D) and PC3 cells (Figure 3E and F).
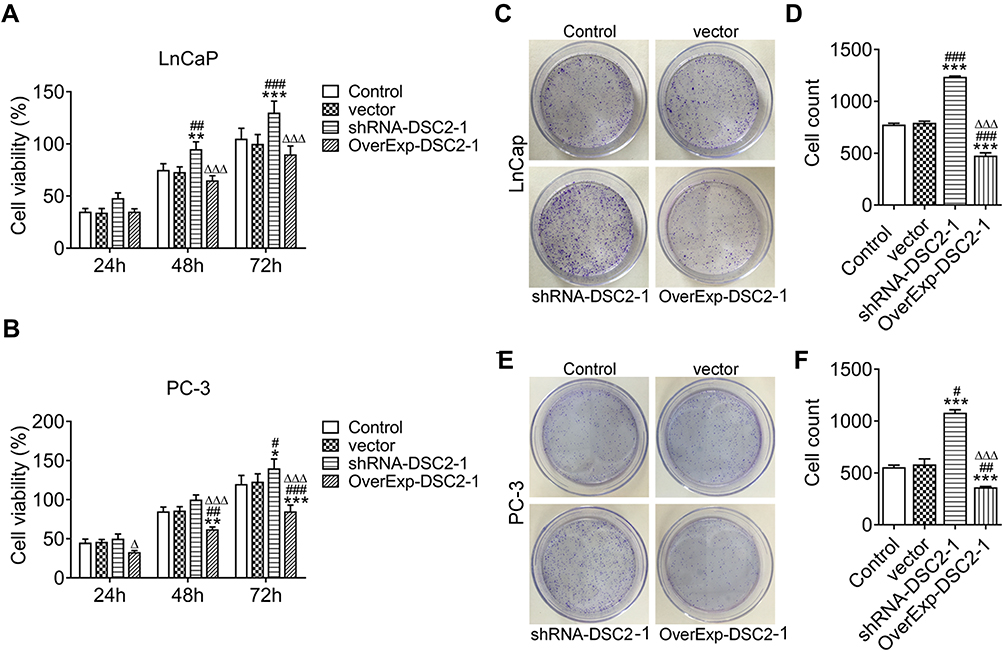 |
Figure 3 DSC2 expression affects the proliferation and clone formation ability of LNCaP cells and PC3 cells. (A) The proliferation of LNCaP cells after transfection was detected by CCK-8 assay. **P<0.01 and ***P<0.001 vs. Control group. ##P<0.01 and ###P<0.001 vs. Vector group. ∆∆∆P<0.001 vs. ShRNA-DSC2-1 group. (B) The proliferation of PC3 cells after transfection was detected by CCK-8 assay. *P<0.05, **P<0.01 and ***P<0.001 vs. Control group. ##P<0.01 and ###P<0.001 vs. Vector group. ∆P<0.05 and ∆∆∆P<0.001 vs. ShRNA-DSC2-1 group. (C/D) The clone formation ability of LNCaP cells after transfection was showed by clone formation assay. ***P<0.001 vs. Control group. ###P<0.001 vs. Vector group. ∆∆∆P<0.001 vs. ShRNA-DSC2-1 group. (E/F) The clone formation ability of PC3 cells after transfection was showed by clone formation assay. ***P<0.001 vs. Control group. #P<0.05 and ##P<0.01 vs. Vector group. ∆∆∆P<0.001 vs. ShRNA-DSC2-1 group. |
DSC2 Expression Affects the Invasion and Migration of LNCaP Cells and PC3 Cells
Inhibition of DSC2 promoted the invasion and migration of LNCaP cells and DSC2 overexpression suppressed the invasion and migration of LNCaP cells. Empty vector transfection exerted no obvious effect on the invasion and migration of LNCaP cells (Figure 4A). The invasion and migration of PC3 cells were enhanced in ShRNA-DSC2 group and inhibited in OverExp-DSC2 group while not obviously changed in vector group (Figure 4B).
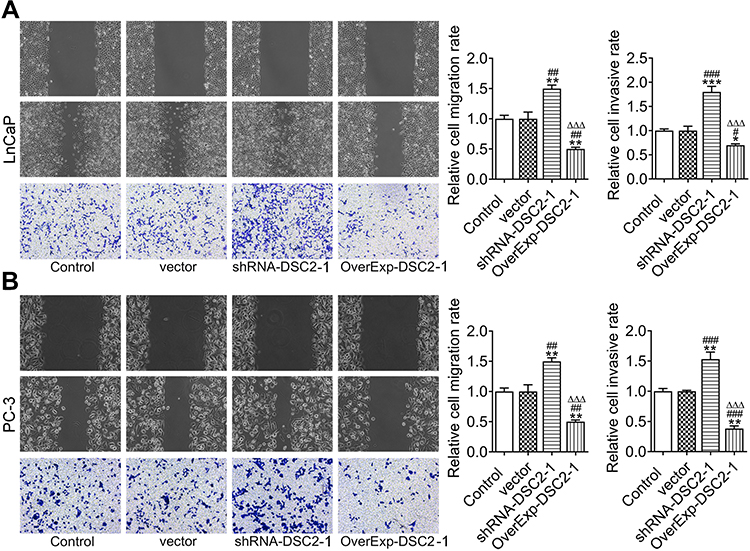 |
Figure 4 DSC2 expression affects the invasion and migration of LNCaP cells and PC3 cells. (A) The migration and invasion of LNCaP cells after transfection was respectively detected by wound healing assay and transwell assay. *P<0.05, **P<0.01 and ***P<0.001 vs. Control group. #P<0.05, ##P<0.01 and ###P<0.001 vs. Vector group. ∆∆∆P<0.001 vs. ShRNA-DSC2-1 group. (B) The migration and invasion of PC3 cells after transfection was respectively detected by wound healing assay and transwell assay. **P<0.01 vs. Control group. ##P<0.01 and ###P<0.001 vs. Vector group. ∆∆∆P<0.001 vs. ShRNA-DSC2-1 group. |
DSC2 Expression Affects the Apoptosis of LNCaP Cells and PC3 Cells
As shown in Figure 5A and B, inhibition of DSC2 suppressed the apoptosis while DSC2 overexpression stimulated the apoptosis of LNCaP cells. The result of Figure 5C and D indicated that apoptosis of PC3 cells was restrained by inhibition of DSC2 and accelerated by DSC2 overexpression.
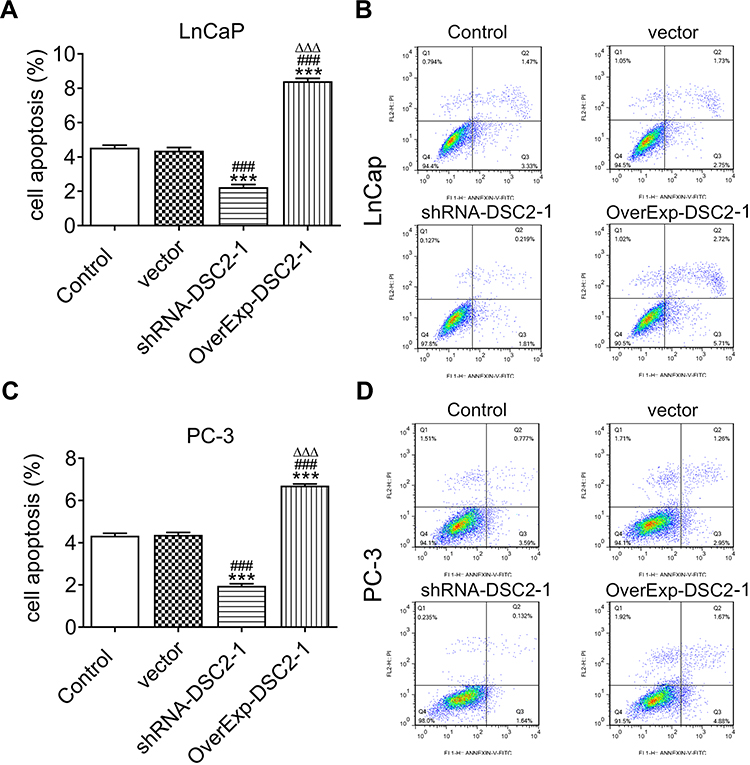 |
Figure 5 DSC2 expression affects the apoptosis of LNCaP cells and PC3 cells. (A) The apoptosis of LNCaP cells after transfection was analyzed by flow cytometry analysis. ***P<0.001 vs. Control group. ###P<0.001 vs. Vector group. ∆∆∆P<0.001 vs. ShRNA-DSC2-1 group. (B) Flow cytometry map of LNCaP cells after transfection. (C) The apoptosis of PC3 cells after transfection was analyzed by flow cytometry analysis. ***P<0.001 vs. Control group. ###P<0.001 vs. Vector group. ∆∆∆P<0.001 vs. ShRNA-DSC2-1 group. (D) Flow cytometry map of PC3 cells after transfection. |
DSC2 Expression Affects the Expression of Apoptosis-Related Proteins and Cell Cycle-Related Proteins in LNCaP Cells and PC3 Cells
Inhibition of DSC2 down-regulated the expression of Bax and c-Casp-3 and up-regulated the Bcl-2 expression while DSC2 overexpression up-regulated the expression of Bax and c-Casp-3 and down-regulated the Bcl-2 expression in LNCaP cells (Figure 6A) and PC3 cells (Figure 6B). Inhibition of DSC2 up-regulated the expression of Cyclin D1, CDK2, Cyclin B1 and CDK1 while DSC2 overexpression down-regulated the expression of Cyclin D1, CDK2, Cyclin B1 and CDK1 in LNCaP cells (Figure 6C) and PC3 cells (Figure 6D).
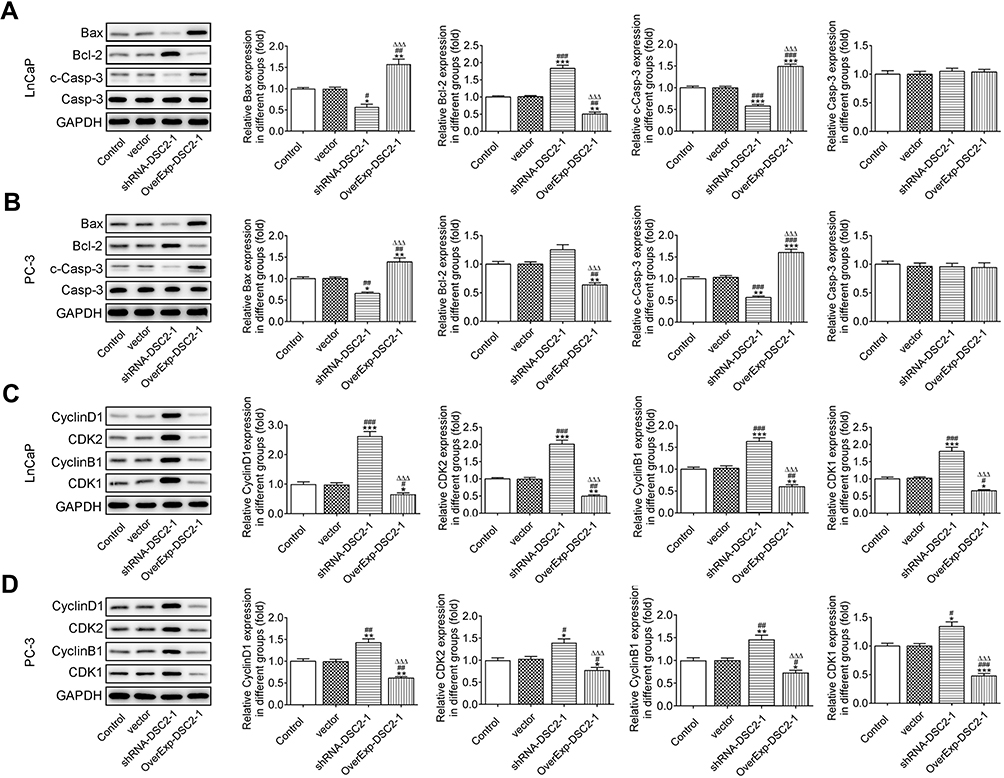 |
Figure 6 DSC2 expression affects the expression of apoptosis-related proteins and cell cycle-related proteins in LNCaP cells and PC3 cells. (A) The expression of Bax, Bcl-2, Casp-3 and c-Casp-3 in LNCaP cells after transfection was detected by Western blot analysis. *P<0.05, **P<0.01 and ***P<0.001 vs. Control group. ##P<0.01 and ###P<0.001 vs. Vector group. ∆∆∆P<0.001 vs. ShRNA-DSC2-1 group. (B) The expression of Bax, Bcl-2, Casp-3 and c-Casp-3 in PC3 cells after transfection was detected by Western blot analysis. *P<0.05, **P<0.01 and ***P<0.001 vs. Control group. ##P<0.01 and ###P<0.001 vs. Vector group. ∆∆∆P<0.001 vs. ShRNA-DSC2-1 group. (C) The expression of Cyclin D1, CDK2, Cyclin B1 and CDK1 in LNCaP cells after transfection was detected by Western blot analysis. *P<0.05, **P<0.01 and ***P<0.001 vs. Control group. #P<0.05, ##P<0.01 and ###P<0.001 vs. Vector group. ∆∆∆P<0.001 vs. ShRNA-DSC2-1 group. (D) The expression of Cyclin D1, CDK2, Cyclin B1 and CDK1 in PC3 cells after transfection was detected by Western blot analysis. *P<0.05, **P<0.01 and ***P<0.001 vs. Control group. #P<0.05, ##P<0.01 and ###P<0.001 vs. Vector group. ∆∆∆P<0.001 vs. ShRNA-DSC2-1 group. |
DSC2 Expression Affects the E-Cad/β-Catenin Signaling Pathway
The expression of p-β-catenin and EGFR was elevated and E-cad expression was reduced by the inhibition of DSC2 and DSC2 overexpression inhibited the expression of p-β-catenin and EGFR and promoted the E-cad expression in LNCaP cells (Figure 7A) and PC3 cells (Figure 7B).
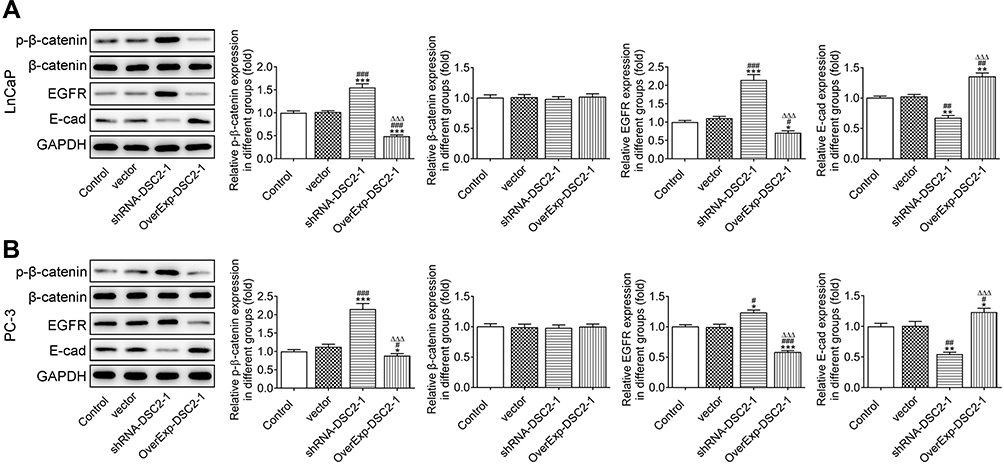 |
Figure 7 DSC2 expression affects the E-cad/β-catenin signaling pathway. (A) The expression of β-catenin, p-β-catenin, EGFR and E-cad in LNCaP cells after transfection was determined by Western blot analysis. *P<0.05, **P<0.01 and ***P<0.001 vs. Control group. #P<0.05, ##P<0.01 and ###P<0.001 vs. Vector group. ∆∆∆P<0.001 vs. ShRNA-DSC2-1 group. (B) The expression of β-catenin, p-β-catenin, EGFR and E-cad in PC3 cells after transfection was determined by Western blot analysis. *P<0.05, **P<0.01 and ***P<0.001 vs. Control group. #P<0.05, ##P<0.01 and ###P<0.001 vs. Vector group. ∆∆∆P<0.001 vs. ShRNA-DSC2-1 group. |
Discussion
Prostate cancer has now overtaken lung cancer as one of the most common tumors in men.11 In the United States, there were 164,690 new cases of prostate cancer and 29,430 deaths from prostate cancer in 2018.12 Due to the lack of effective diagnostic methods in the early stage of prostate cancer, many patients in China are diagnosed with prostate cancer at a later stage, thereby missing the chance of radical cure. To explore the molecular mechanism of the occurrence and development of prostate cancer, abnormalities of related genes will provide new ideas for the treatment of advanced prostate cancer.
Through GEPIA, we observed the expression of DSC1, DSC2, DSC3 and DSC4 in prostate cancer tissues and normal tissues and found that DSC2 expression between tumor tissues and normal tissues was significantly different, thereby choosing DSC2 to be investigated in this study. DSC2, a transmembrane protein, was connected with the other three kinds of DSC to ensure the integrity of myocardial structure and function, and also participates in cell signal transmission and apoptosis.13 Heuser et al14 knocked out DSC2 gene in zebrafish model, and observed arrhythmia, myocardial contractility damage and myocardial cell edema. The lipid droplet, vacuolar degeneration, mitochondrial swelling and cristae disappearance was showed in P19 cells with silencing of DSC2 and silencing of DSC2 also promoted the apoptosis of P19 cells.15 DSC2 expression was evaluated in 308 cases of esophageal squamous cell carcinoma and increased DSC2 expression was related to poor prognosis.16 From GEPIA database, DSC2 expression was increased in prostate cancer tissues compared with normal tissues. However, prostate cancer patients with relative low expression of DSC2 have poorer DFS. In this study, inhibition of DSC2 promoted the proliferation, clone formation ability, migration and invasion while suppressed the apoptosis of LNCaP cells and PC3 cells. Although DSC2 expression was demonstrated to be increased in prostate cancer cells which was consistent with the result from GEPIA database, inhibition of DSC2 promoted the proliferation, clone formation ability, migration and invasion while suppressed the apoptosis of prostate cancer cells. We suspect that the above phenomenon may be explained by the phenomenon that prostate cancer patients with relative low expression of DSC2 have poorer DFS.
It was found that DSC could regulate the β-catenin signaling pathway, change the proliferation and invasion of tumor cells, and promote the disease progression of malignant tumors. Kolegraff et al178 demonstrated that down-regulation of DSC2 activated the EGFR/Akt pathway, which caused the accumulation of phosphorylated β-catenin in the nucleus and phosphorylated β-catenin could activate the downstream target genes, resulting in the abnormal proliferation and invasion of tumor cells. Fang et al18 found that DSC2 interference reduced the DSC2/γ-catenin complex, which led to the increase of γ-catenin in cytoplasm. γ-catenin competed with β-catenin for E-catenin binding, which increased the amount of free β-catenin in the cell and nucleus, thus promoting the activation of the downstream target genes of β-catenin and the occurrence and development of malignant tumors. EGFR is associated with tumor cell proliferation, angiogenesis, tumor invasion, metastasis, and inhibition of apoptosis.19,20 Research indicated that EGFR overexpression was associated with the development of castration resistant prostate cancer (CRPC) in patients.21 WAP four-disulfide core domain 2 (WFDC2) inhibited the prostate cancer metastasis by suppressing EGFR signaling.22 Spautin-1 treated the prostate cancer by the inhibition of EGFR signaling.23 E-cad is a member of the cadherin family and widely exists in various epithelial cells.24 Decreased expression of E-cad would reduce the adhesion of cytoskeleton system, make tumor cells more easily dispersed, and accelerate their invasive growth to the surrounding tissue.25 The decreased E-cad promoted the prostate cancer chemoresistance via Notch signaling.26 In this study, inhibition of DSC2 up-regulated the expression of p-β-catenin and EGFR and down-regulated E-cad expression in LNCaP cells and PC3 cells, which was reversed by DSC2 overexpression.
In conclusion, DSC2 expression was increased in prostate cancer cells. Moreover, inhibition of DSC2 promoted the proliferation, clone formation ability, migration and invasion while suppressed the apoptosis of LNCaP cells and PC3 cells, which was reversed by DSC2 overexpression. GEPIA shows that patients with lower DSC2 expression have poor DFS. These data suggest that DSC2 may serve as a predictor for patient outcomes and provide the fundamental basis for treatment of prostate cancer.
Funding
There is no funding to report.
Disclosure
The authors declare they have no competing interests.
References
1. Siegel RL, Miller KD, Jemal A. Cancer statistics, 2020. CA Cancer J Clin. 2020;70(1):7–30. doi:10.3322/caac.21590
2. Patel S, Vanharanta S. Epigenetic determinants of metastasis. Mol Oncol. 2016;11(1):1–16.
3. Feng R-M, Zong Y-N, Cao S-M, Xu R-H. Current cancer situation in China: good or bad news from the 2018 global cancer statistics? Cancer Commun. 2019;39(1):22–34. doi:10.1186/s40880-019-0368-6
4. Chitaev NA, Troyanovsky SM. Direct Ca2+-dependent heterophilic interaction between desmosomal cadherins, desmoglein and desmocollin, contributes to cell-cell adhesion. J Cell Biol. 1997;138(1):193–201. doi:10.1083/jcb.138.1.193
5. Nuber UA, Schäfer S, Schmidt A, Koch P, Franke W. The widespread human desmocollin Dsc2 and tissue-specific patterns of synthesis of various desmocollin subtypes. Eur J Cell Biol. 1995;66(1):69–74.
6. Chidgey M, Dawson C. Desmosomes: a role in cancer? Br J Cancer. 2007;96(12):1783–1787. doi:10.1038/sj.bjc.6603808
7. Hamidov Z, Altendorf-Hofmann A, Chen Y, Settmacher U, Petersen I, Knösel T. Reduced expression of desmocollin 2 is an independent prognostic biomarker for shorter patients survival in pancreatic ductal adenocarcinoma. J Clin Pathol. 2011;64(11):990–994. doi:10.1136/jclinpath-2011-200099
8. Knösel T, Chen Y, Hotovy S, Settmacher U, Altendorf-Hofmann A, Petersen I. Loss of desmocollin 1-3 and homeobox genes PITX1 and CDX2 are associated with tumor progression and survival in colorectal carcinoma. Int J Colorectal Dis. 2012;27(11):1391–1399. doi:10.1007/s00384-012-1460-4
9. Hayashi T, Sentani K, Oue N, et al. Desmocollin 2 is a new immunohistochemical marker indicative of squamous differentiation in urothelial carcinoma. Histopathology. 2011;59(4):710–721. doi:10.1111/j.1365-2559.2011.03988.x
10. Cui T, Chen Y, Yang L, et al. Diagnostic and prognostic impact of desmocollins in human lung cancer. J Clin Pathol. 2012;65(12):1100–1106. doi:10.1136/jclinpath-2011-200630
11. Mohler J, Armstrong A, Bahnson R. Prostate cancer, version 1.2016. J Natl Compr Canc Netw. 2016;14(1):19–30. doi:10.6004/jnccn.2016.0004
12. Siegel RL, Miller KD, Jemal A. Cancer statistics, 2018. CA Cancer J Clin. 2018;68(1):7–30.
13. Iyer VR, Chin AJ. Arrhythmogenic right ventricular cardiomyopathy/dysplasia (ARVC/D). Am J Med Genet C Semin Med Genet. 2013;163(3):185–197. doi:10.1002/ajmg.c.31368
14. Heuser A, Plovie ER, Ellinor PT, et al. Mutant desmocollin-2 causes arrhythmogenic right ventricular cardiomyopathy. Am J Hum Genet. 2006;79(6):1081–1088. doi:10.1086/509044
15. Li D, Yang B, Zhou X, et al. The effects of silencing desmocollin-2 gene on the cardiomyocyte-like cells differentiated from P19 cell lines. Chin J Cardiac Pacing Electrophysiol. 2014;28(5):427–431.
16. Fang W, Gu W, Li E, et al. Reduced membranous and ectopic cytoplasmic expression of DSC2 in esophageal squamous cell carcinoma: an independent prognostic factor. Hum Pathol. 2010;41(10):1456–1465. doi:10.1016/j.humpath.2010.04.003
17. Kolegraff K, Nava P, Helms MN, Parkos CA, Nusrat A. Loss of desmocollin-2 confers a tumorigenic phenotype to colonic epithelial cells through activation of Akt/β-catenin signaling. Molecular biology of the cell. 2011;22(8):1121–1134.
18. Fang WK, Liao LD, Li LY, et al. Down-regulated desmocollin-2 promotes cell aggressiveness through redistributing adherens junctions and activating beta-catenin signalling in oesophageal squamous cell carcinoma. J Pathol. 2013;231(2):257–270. doi:10.1002/path.4236
19. Lui VW, Grandis JR. EGFR-mediated cell cycle regulation. Anticancer Res. 2002;22(1a):1–11.
20. Liu W, Innocenti F, Chen P, Das S, Cook EH
21. Shah R, Ghosh D, Elder J. Epidermal growth factor receptor (ErbB1) expression in prostate cancer progression: correlation with androgen independence. Prostate. 2006;66(13):1437–1444. doi:10.1002/pros.20460
22. Xiong Y, Yuan L, Chen S, et al. WFDC2 suppresses prostate cancer metastasis by modulating EGFR signaling inactivation. Cell Death Dis. 2020;11(7):537. doi:10.1038/s41419-020-02752-y
23. Liao Y, Guo Z, Xia X, et al. Inhibition of EGFR signaling with Spautin-1 represents a novel therapeutics for prostate cancer. J Exp Clin Cancer Res. 2019;38(1):157. doi:10.1186/s13046-019-1165-4
24. Danyi Z, Tao W, Lei S, Kui J, Xiaoyan Y, Xian Z. The expression and regularive relationship of NDRG-1 and E-cadherin in hepatocellular carcinoma. J Mod Oncol. 2013;21(12):2691–2693.
25. Kokkinos MI, Murthi P, Wafai R, Thompson EW, Newgreen DF. Cadherins in the human placenta–epithelial-mesenchymal transition (EMT) and placental development. Placenta. 2010;31(9):747–755. doi:10.1016/j.placenta.2010.06.017
26. Wang W, Wang L, Mizokami A, et al. Down-regulation of E-cadherin enhances prostate cancer chemoresistance via notch signaling. Chin J Cancer. 2017;36(1):35. doi:10.1186/s40880-017-0203-x
 © 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.