Back to Journals » Journal of Pain Research » Volume 15
Proteomic and Phosphoproteomic Changes of MAPK-Related Inflammatory Response in an Animal Model of Neuropathic Pain by Differential Target Multiplexed SCS and Low-Rate SCS
Authors Cedeño DL, Tilley DM ![]() , Vetri F, Platt DC, Vallejo R
, Vetri F, Platt DC, Vallejo R
Received 10 November 2021
Accepted for publication 24 February 2022
Published 1 April 2022 Volume 2022:15 Pages 895—907
DOI https://doi.org/10.2147/JPR.S348738
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 4
Editor who approved publication: Professor E Alfonso Romero-Sandoval
David L Cedeño,1,2 Dana M Tilley,2 Francesco Vetri,3 David C Platt,1,2 Ricardo Vallejo1– 3
1Neuroscience, Illinois Wesleyan University, Bloomington, IL, USA; 2Research and Development, SGX Medical, Bloomington, IL, USA; 3Research Department, National Spine and Pain Centers, Bloomington, IL, USA
Correspondence: Ricardo Vallejo; David L Cedeño, Email [email protected]; [email protected]
Introduction: Neuropathic pain initiates an interplay of pathways, involving MAP kinases and NFκB-signaling, leading to expression of immune response factors and activation and inactivation of proteins via phosphorylation. Neuropathic pain models demonstrated that spinal cord stimulation (SCS) may provide analgesia by modulating gene and protein expression in neuroinflammatory processes. A differential target multiplexed programming (DTMP) approach was more effective than conventional SCS treatments at modulating these. This work investigated the effect of DTMP and low rate SCS (LR-SCS) on proteins associated with MAP kinases and NFκB-signaling relevant to neuroinflammation.
Methods: Animals subjected to the spared nerve injury model (SNI) of neuropathic pain were treated continuously (48h) with either DTMP or LR-SCS. No-SNI and No-SCS groups were included as controls. Proteomics and phosphoproteomics of stimulated spinal cord tissues were performed via liquid chromatography/tandem mass spectrometry. Proteins were identified from mass spectra using bioinformatics. Expression levels and fold changes (No-SCS/No-SNI and SCS/No-SCS) were obtained from spectral intensities.
Results: Analyses identified 7192 proteins, with 1451 and 705 significantly changed by DTMP and LR-SCS, respectively. Eighty-one proteins, including MAP kinases, facilitating NFκB-signaling as part of inflammatory processes were identified. The pain model significantly increased expression levels of complement pathway-related proteins (LBP, NRG1, APP, CFH, C3, C5), which were significantly reversed by DTMP. Expression levels of other complement pathway-related proteins (HMGB1, S100A8, S100A9, CRP, C4) were decreased by DTMP, although not significantly affected by SNI. Other proteins (ORM1, APOE, NG2, CNTF) involved in NFκB-signaling were increased by SNI and decreased by DTMP. Expression levels of phosphorylated protein kinases involved in NFκB-signaling (including MAP kinases, PKC, MARK1) were affected by the pain model and reverse modulated by DTMP. LR-SCS modulated inflammatory-related proteins although to a lesser extent than DTMP.
Conclusion: Proteomic analyses support the profound effect of the DTMP approach on neuroinflammation via MAP kinases and NFκB-mediated signaling to alleviate neuropathic pain.
Keywords: differential target multiplexed spinal cord stimulation, proteomics, neuropathic pain, neuroinflammation, mitogen activated protein kinase, nuclear factor-kappa B
Introduction
Pain is intended to protect species from harmful stimuli and to promote healing after injury. Processing in the spinal cord and dorsal root ganglia of acute pain resulting from tissue injury is accompanied by an inflammatory process concerted by immune and glial cells. In general, inflammation promotes healing and tissue regeneration, such that when healing occurs, both inflammation and pain go away. However, the immune response that led to local inflammation of the injured tissue is also signaled to the central nervous system (CNS) as a result of injury and may lead to chronic pain. Glial cells play a fundamental role in chronic neuropathic pain since they are actively involved in the regulation of immune and inflammatory responses. Once activated, microglia, the immune cells of the CNS, transition into activated states that alert (pro-inflammatory) and protect (anti-inflammatory) the CNS.1 The progression to chronic neuropathic pain involves the activation of microglia in response to aberrant neural activity in the periphery. Active microglia release signaling molecules, such as cytokines, chemokines and other moieties, such as glutamate, GABA, etc., that activate perisynaptic astrocytes, which in turn promote the disruption of fundamental biological processes which normally maintain neuron-glial interactions and are required for homeostatic balance of the CNS.2 Activated microglial cells and astrocytes play a pivotal role in neuroinflammation and chronic pain.3–5
Spinal cord stimulation (SCS) is an effective and safe treatment for intractable chronic neuropathic pain.6,7 Recent preclinical research based on high throughput transcriptomics and proteomics has revealed that SCS using a low-rate electrical signal, consisting of 50 Hz pulses, has an effect on immune and inflammatory processes.8–10 Further work implied that such a low rate SCS (LR-SCS) may be conducive to up regulate proinflammatory pathways via microglial activation, which may prevent optimal efficacy.11 In order to improve clinical outcomes, our group developed a SCS treatment based on a differential target multiplexed programming approach (DTMP) that uses multiple signals intended to modulate neurons and glial cells to balance their interactions toward a physiologic homeostatic state. DTMP provided significant relief of pain-like behavior in animals with the spared nerve injury (SNI) model of neuropathic pain while modulating gene and protein expression in biological processes such as immune system, synaptic transmission, and ion transport toward levels of naive animals.12,13 Furthermore, DTMP modulated cell-specific transcriptomes of both neurons and glial cells (microglia, astrocytes, and oligodendrocytes) more effectively than SCS programs which rely on a single signal at either low or high rate.14 Recent work on transcriptomes associated with active microglia states corroborated these findings.1
Besides transcriptomics, which evaluates changes in the expression levels of protein-coding mRNA, it is crucial to study the effects of SCS on proteins as well as their post-translational modifications, particularly those involved in signaling cascades. Phosphorylation and dephosphorylation of proteins at specific locations activate or deactivate them within regulatory pathways. At the intracellular molecular level, the activity of the NFκB cascade, known to promote inflammation and neuronal sensitization, is partially regulated by the MAP kinase superfamily.15 Relevant to the identification and quantitation of these proteins is the nature of the applied methodology. Proteomic-based screening provides information of thousands of proteins in a single experiment using liquid chromatography tandem mass spectrometry (LC/TMS). Various extraction and purification methods exist for LC/TMS proteomic analyses and have their own limitations depending on depth and breadth of desired observations to be made. The process we utilized may result in loss of non-anchored proteins and small peptides.16–18 Fortunately, multiple molecules involved in inflammatory signaling pathways, including MAP kinases, are membrane bound or intracellularly anchored to cytoskeletal proteins, and provide us with relevant insights into the overall effects of spinal cord stimulation (SCS) on those biological processes. This work offers a comparative look at the effect of conventional LR-SCS and DTMP on proteins involved in inflammatory pathways, particularly in relation to pro-inflammatory signaling via MAP kinases and indirectly the NFκB pathway in the early stages of an animal model of neuropathic pain.
Methods
Animals, Surgical Manipulations, and SCS
A detailed description of the experimental design, including implementation of the SNI model and evaluation of the resultant hypersensitive behavior, is provided elsewhere.12 The proteomic analysis presented in this work has been conducted on samples from animals subjected to behavioral testing in reference 12. This study represents an extension of the analyses started with that study.
Briefly, the study was approved by the Institutional Animal Care and Use Committee at Illinois Wesleyan University according to the USDA Animal Welfare Act and the NIH Public Health Service Policy on the Humane Care and Use of Animals. Male Sprague-Dawley rats (Envigo RMS, Indianapolis, US) weighing in the 275–315 g range were housed individually in a temperature and humidity-controlled room with a 12-hour light/dark cycle. Food and water were supplied ad libitum. After acclimation to the environment, animals were randomly assigned to either No-SCS (untreated, n = 10), DTMP (n = 10), LR-SCS (n=10), or No-SNI (uninjured, n = 10). Animals in the No-SCS, DTMP, and LR-SCS groups were implanted with a miniaturized cylindrical quadrupolar SCS lead (0.62 mm diameter, 1 mm Pt/Ir electrodes, 1 mm spacing) at the L1-L2 level and subjected to the spared nerve injury (SNI) model of neuropathic pain as described previously.12 Lead position was chosen according to the anatomical dissections by Gelderd and Chopin19 showing that the sciatic nerve originates at L1-2 vertebral levels and exits the cord at L4-6. Five days after SNI surgery, animals in the DTMP and LR-SCS groups that successfully developed mechanical hypersensitivity (at least 30% reduction of paw withdrawal threshold, PWT) were stimulated continuously for 48 hours. No-SCS animals were connected to the SCS device but not stimulated for the same duration. No-SNI animals did not receive neither the SNI model nor any SCS. All animals were behaviorally assessed, as previously described,12 for mechanical and thermal hypersensitivity in parallel by a researcher blinded to the assignments before surgical intervention, as well as before starting and ending the stimulation period. DTMP uses multiplexed charge-balanced pulsed signals with components at 50 Hz (150 µs pulse width, PW) and 1200 Hz (50 µs PW) that are distributed over the contacts of the lead. LR-SCS uses single pulses at 50Hz (150 µs PW). Signal intensities corresponded to about 70% of the motor threshold and were in the 0.03–0.10 mA range. Signals were not duty cycled and initial intensities were unchanged throughout stimulation.
Protein Isolation and Quantification
After assessment and euthanization of animals following 48 hours of SCS, the ipsilateral dorsal quadrant of the spinal cord underneath the SCS lead was harvested, washed with cold saline, snap frozen and stored at −80 °C until analyses. Out of the available representative samples from responders to treatment (PWT increased to at least 30% of the pre-SNI measurement, 8 per SCS group), 4 were used in the previously reported transcriptomic analysis. Therefore, the 4 remaining samples per group were available for the proteomic analyses. Each dissected tissue was suspended in a buffer (9M urea) enriched with protease inhibitors and free of ionic detergents followed by sonication and centrifugation to separate the proteins out. After determination of protein concentration, proteins were digested with trypsin followed by alkylation of cysteine residues under appropriate buffering using standard methods.20 Tryptic peptides in a pool of biological specimens (n = 3–4) of each experimental group were isotopically labeled using a tandem mass tag (TMT) system, which allows for simultaneous identification, quantification, and comparison of a protein in various experimental groups.21 All peptides were combined and loaded onto a 50 cm x 100 µm PicoFrit capillary column packed with C18 reversed-phase resin and fractionated via reverse column liquid chromatography (LC) into 96 fractions. The column was developed with a 150-minute linear gradient of acetonitrile in 0.125% formic acid delivered at 280 nL/min. Fractions were combined non-sequentially to 12 fractions, and mass spectra obtained in a LC/TMS instrument, allowing the highest number of identifications possible and most accurate quantification via multi-notch MS3 methodology,20,22 with parameters optimized under protocols developed at Cell Signaling Technology (Danvers, MA). Mass spectra were evaluated using SEQUEST and the Core platform from Harvard University.23 The UniProt rat database (uniprot.org) was used to search for proteins. Search results were filtered with mass accuracy of ±5 ppm on precursor ions and further filtered for multiple comparisons to a 1% protein level false discovery rate (FDR). Fold changes were obtained from comparison of the normalized spectral intensities (log2 scale) of the tagged peptides uniquely assigned to each protein. Significance of fold changes was calculated using a two tailed t-test for each protein identified and quantified. Proteins significantly differentiated (p < 0.05) between No-SNI and No-SCS were isolated, while the effect of either DTMP or LR-SCS treatment relative to No-SCS was followed. Protein-protein interaction networks for significantly differentiated proteins were built using the StringDB v11.0 bioinformatics tool.24 Significantly enriched biological processes affected by the pain model and either DTMP or LR-SCS were obtained via gene ontology enrichment analyses (GOEA) within the online Panther database.25
For phosphoproteomics, proteins from the same 3–4 biological samples used in the whole proteomics were digested with trypsin and the peptides purified by reversed-phase solid-phase extraction, followed by phospho-enrichment using immobilized metal affinity chromatography (IMAC) with iron-based magnetic beads (PTMScan® Fe-IMAC, Cell Signaling Technology, Danvers MA).26 Unbound peptides were washed out, and immobilized phosphopeptides eluted with basic pH buffer. Reversed-phase purification was performed to purify peptides prior to LC/TMS analysis of pooled peptides from 3–4 biological samples, conducted as described above for whole proteomics following standard techniques developed at Cell Signaling Technology. Due to the limited amount of protein sample left for phosphoproteomics after the whole proteomic analysis, technical assays were run for each pool of biological samples in duplicate. Therefore, phosphoproteomic results based on spectral intensities of phosphoproteins filtered as described above, although reliable, could not be assessed for statistical significance. Instead, the coefficient of variation (CV) was used to measure the relative variability of fold changes from the variability of protein expression levels for individual groups. The CV, defined as the ratio of the standard deviation to the mean for spectral counts for a given experimental group, was used to obtain the coefficient of variation of the fold changes No-SCS/No-SNI and SCS/No-SCS. The CV of the ratio is given by the square root of the sum of the square of the individual CVs of the ratioed values.
Results
Behavioral analysis of the experimental groups was previously reported.12 Briefly, mean paw withdrawal thresholds (± standard error) normalized to pre-SNI baseline were 24.6 ± 2.6% for No-SCS, 62.9 ± 8.9% for DTMP, and 37.6 ± 6.0% for LR-SCS. Relief of mechanical hypersensitivity with DTMP and LR-SCS was significantly better than with No-SCS. DTMP also significantly reduced sensitivity to hot (44.9 ± 1.9 °C vs 39.4 ± 0.4 °C for pre-SCS) and cold (6.7 ± 1.4 °C vs 11.0 ± 0.4 °C for pre-SCS). In contrast, LR-SCS did not provide significant relief of hot (40.7 ± 2.2 °C) and cold (11.1 ± 1.6 °C) hypersensitivity relative to pre-SCS temperature thresholds.
The proteomic analysis uniquely identified 7192 proteins from the spinal cord tissue. Of these, 647 were significantly affected by the SNI pain model (as reflected by the No-SCS group) relative to uninjured animals (No-SNI), while 1451 and 705 proteins were found to be significantly affected by DTMP and LR-SCS respectively relative to No-SCS. The total breakdown depicting significant fold change in protein expression affected by SNI, DTMP, or LR-SCS is shown in Figure 1A. Figure 1B shows a heat map of the fold changes for the 1451 proteins significantly regulated by DTMP relative to No-SCS (DTMP/No-SCS) in comparison to the effect of LR-SCS (ie, LR-SCS/No-SCS) and the effect of the pain model (ie, No-SCS/No-SNI). Of the 1451 proteins regulated by DTMP, 472 (32.5%) were significantly changed by the pain model, while 521 (35.9%) were significantly regulated by LR-SCS.
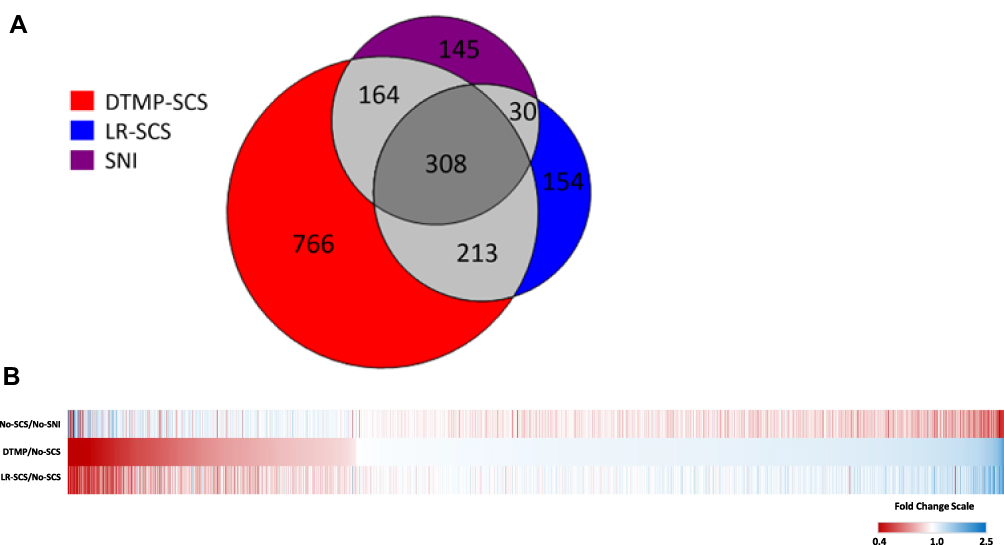 |
Figure 1 (A) Venn diagram accounting for proteins significantly affected by the pain model and both SCS treatment groups. The pain model significantly affected 647 proteins (purple circle), DTMP affected 1451 proteins (red circle), and LR-SCS affected 705 proteins (blue circle). Regions of overlap indicate the number of shared significantly affected proteins between the groups. (B) Heat map of proteins significantly regulated by DTMP relative to No-SCS (DTMP/No-SCS) and in comparison, to the effect of LR-SCS (LR-SCS/No-SCS) and the effect of the pain model relative to uninjured animals (No-SCS/No-SNI). Red color represents decreased expression, blue depicts increased fold changes, and white stands for no change. |
Proteomic Analysis
The GOEA revealed 85 proteins significantly regulated by either DTMP or LR-SCS which are categorized as part of the inflammatory response. Of these proteins, DTMP and LR-SCS significantly changed expression levels of 81 and 56, respectively. Figure 2A illustrates a heatmap of the 81 proteins significantly regulated by DTMP, showing expression level fold changes of these proteins for both SCS treatments relative to No-SCS, and for No-SCS relative to No-SNI (see Supplementary Table 1). These proteins were evaluated in terms of their role in inflammation and pain related pathways, with emphasis on MAP kinase and their related proteins that are involved in the NFκB cascade. The GOEA also identified 81 proteins as regulators of the MAP kinase cascade which are shown in the heatmap in Figure 2B (see Supplementary Table 2). Interestingly, only 18 proteins overlapped between inflammatory and MAP kinase pathways (Figure 2C).
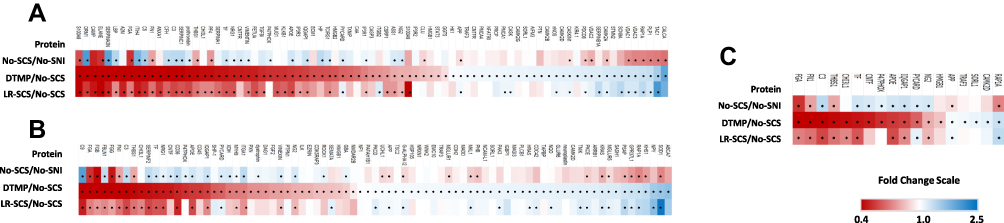 |
Figure 2 (A) Heat map of fold changes in expression levels of proteins enriched in regulation of inflammatory response significantly changed by DTMP. See Supplementary Tables 1 and 2 for corresponding data. (B) Heat map of proteins enriched in regulation of MAP kinase cascade significantly changed by DTMP. See supplementary files for corresponding table. (C) Heat map of proteins that overlap between the aforementioned heat maps. * denotes p < 0.05 for each fold change comparison. Red color represents decreased expression, blue depicts increased fold changes, and white stands for no change. |
Of the 7 MAP kinase upstream regulators, also known to play a role in the activation of the NFκB signaling pathway (CHI3L1, CNTF, HMGB1, NG2, APOE, SORL1, and C3), 5 were the signaling proteins: CHI3L1, CNTF, HMGB1, NG2, and APOE. Among those, CNTF, NG2, and APOE were upregulated by the pain model. DTMP reversed expression levels of all three, whereas LR-SCS reversed expression of only NG2 and APOE. SORL1, an APOE receptor, was only upregulated by DTMP. Multiple proteins involved in processes linked to the NFκB pathway, such as the complement system and the MAPK cascade, were affected. The pain model significantly changed expression levels of proteins upstream of the complement pathway such as LBP, NRG1, APP, CFH, C3, and C5, which were significantly reversed by both DTMP and LR-SCS. C3, a key component of the complement system, was upregulated by the pain model and downregulated by both DTMP and LR-SCS, although the effect was noticeably more pronounced for DTMP (Figure 2). Furthermore, DTMP significantly decreased expression levels of other complement pathway-related proteins, whose expression had not been affected by the pain model, such as HMGB1, S100A8, S100A9, CSRP1, and C4A. Of these, LR-SCS also changed expression levels of S100A8, S100A9, and CSRP1. Other proteins involved in the NFκB signaling pathway outside of the complement cascade that were increased by the pain model and decreased by DTMP were ORM1 and CNTF. Of these, CNTF was not affected by LR-SCS. Additional proteins that facilitate NFκB activation leading to induced expression identified as part of inflammation are depicted in Figure 2B and Figure 3. Figure 3A illustrates proteins significantly regulated by DTMP whereas Figure 3B illustrates the lesser effect of LR-SCS on the same proteins.
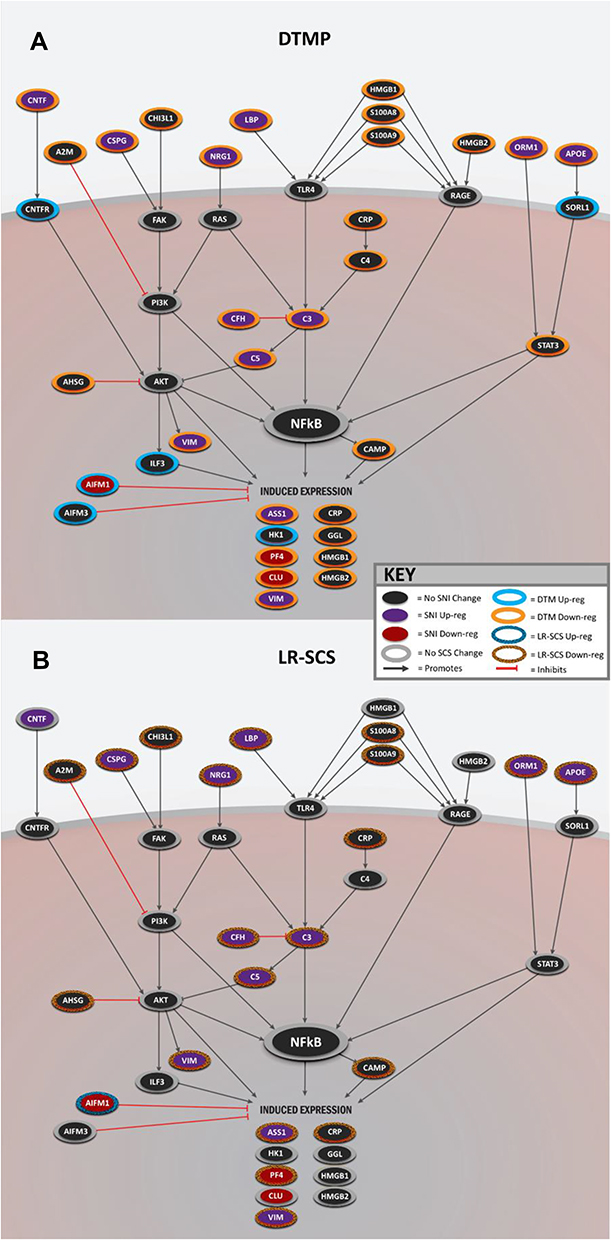 |
Figure 3 Illustrations summarizing changes in expression of proteins involved in the NFκB signaling pathway and neuroinflammation by (A) DTMP or (B) LR-SCS. Up-Reg: up regulation. Down-Reg: down regulation. |
Phosphoproteomic Analysis
Given the importance of signaling cascades and the need to respond rapidly to the environment, changes are reflected in the products of post-translational modifications, such as phosphorylation, rather than the translated expression levels of these proteins. Of the proteins identified to be involved in inflammatory processes through the NFκB signaling pathway, there were 90 unique phosphoproteins distributed in 254 different phosphorylated isoforms. Figure 4 shows the phosphorylated isoform with the greatest fold change between No-SNI, DTMP, and LR-SCS relative to No-SCS for each of the 90 phosphoproteins (see Supplementary Table 3). Figure 5 shows a breakdown of phosphoproteins relevant to the inflammatory process, in which MAP kinases play a fundamental role. The figure also indicates the number of phosphoprotein isoforms affected by the pain model (expression level changed by at least 10%) and the number of these in which SCS changes expression level in the direction of expression levels found in animals that were not subjected to the pain model. It is evident that DTMP has a larger effect on phosphorylated MAP kinases than LR-SCS.
 |
Figure 4 Heat map of the isoforms with the largest fold change for the 90 phosphoproteins involved in NFκB signaling. See Supplementary Table 3 for corresponding data. Highlighted names indicate those involved in the MAPK cascade pathway. |
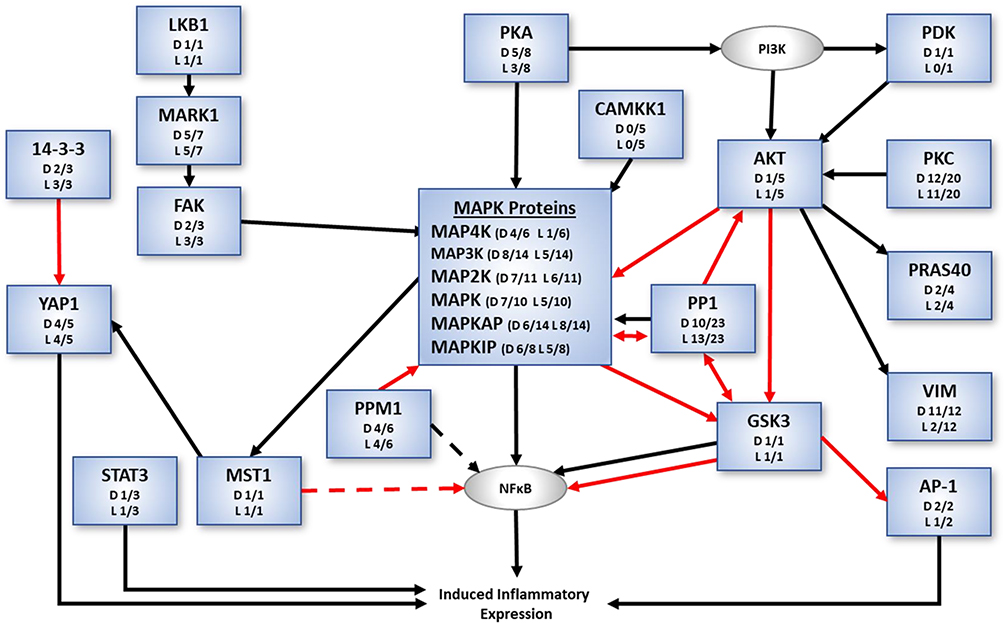 |
Figure 5 Phosphorylated protein classes (blue boxes) of the NFkB signaling cascade involved in inflammatory processes showing the general effect of the SCS in the direction of expression levels of animals without the pain model. Ratios shown for each phosphoprotein class indicate number of isoforms with back-regulation of expression levels due to DTMP (D) or LR-SCS (L) relative to the total number of isoforms in each class. Black arrows indicate activation whereas red arrows indicate inhibition. Protein classes in gray boxes were not differentially expressed. |
As MAP kinases are major regulatory components, their expression levels tend to be relatively stable Regulation of MAP kinases is mainly accomplished via post-translational modifications, such as phosphorylation, which will determine their levels of activity. Inflammatory pathways involving MAP kinases can be initiated by cytokines, chemokines, nerve growth factors, or other inflammatory stimuli. Relevant phosphoproteins of the MAPK cascade identified in the phosphorylation analysis is depicted in Figure 6 (see Supplementary Table 4). In total, 29 phosphorylated proteins of the MAPK cascade were identified. Of these, 21 are MAP kinases, 1 is a MAP kinase interacting protein, and 7 are MAP kinase activated proteins. From these 29 phosphoproteins, there were 63 different phosphorylated protein isoforms having changes in expression level in either the No-SNI control or treatments relative to the untreated animals (No-SCS). Setting a CV of 20% as the maximum threshold for reasonable variation of the calculated fold changes, the expression levels of 33 of these phosphorylated isoforms were affected by the pain model relative to No-SNI. Treatment with SCS changed expression levels towards expression levels found in the No-SNI group for many of these proteins.
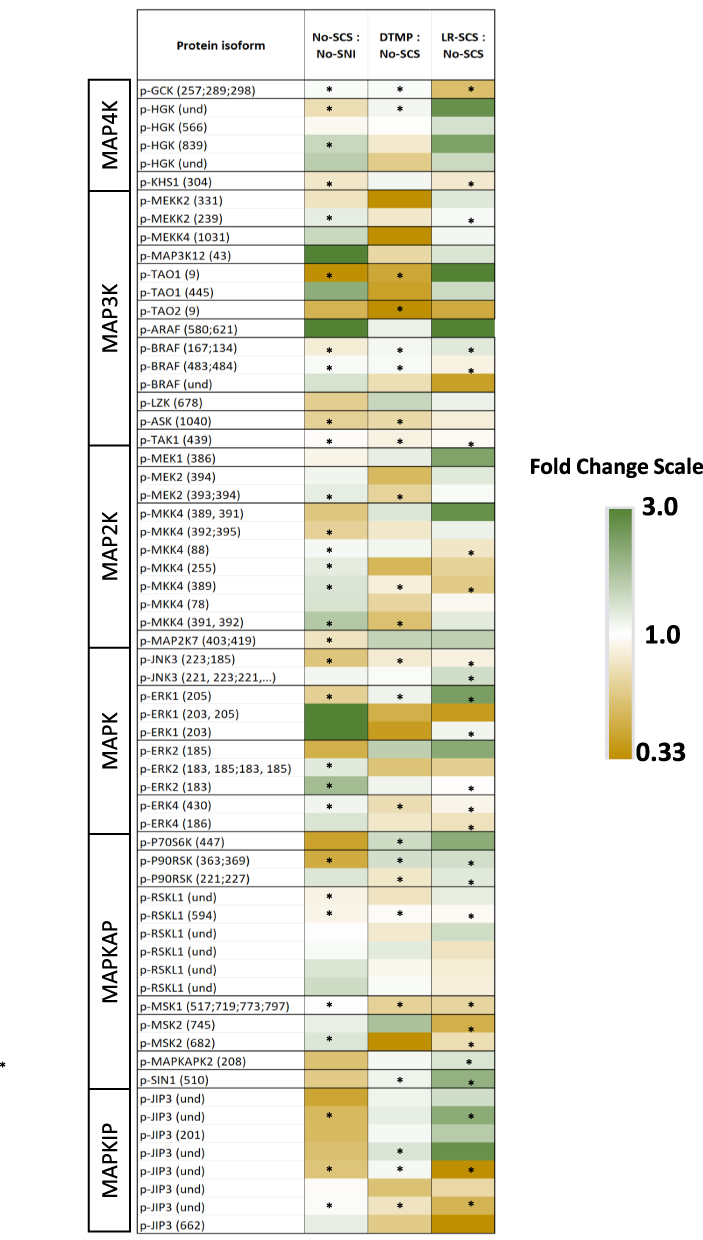 |
Figure 6 Heat map of fold changes of phosphoproteins associated with MAP kinase cascade. See Supplementary Table 4 for corresponding data. * denotes fold changes with ≤20% coefficient of variation. Content of parenthesis next to the protein isoform denotes the assignment of the phosphorylation site in the protein chain, und = undetermined. |
Of the MAP kinase kinase kinase kinases (MAP4Ks) 3 different phosphorylated proteins were identified (p-HGK, p-KHS1, and p-GCK) with 6 different isoforms (Figure 6). Three of these experienced more than 10% change in expression level due to the pain model. DTMP reversed expression levels of 2 of these (p-HGK and p-KHS1) while LR-SCS reversed none. Among the MAP kinase kinase kinases (MAP3Ks), there were 9 phosphoproteins found with 14 different isoforms. Five of them experienced more than 10% change in expression level due to the pain model, with p-BRAF being reversed by DTMP and LR-SCS. Of the MAP kinase kinases (MAP2Ks), 4 phosphoproteins with 11 phosphorylation states were found. Expression levels of 7 of these were affected by more than 10% by the pain model. DTMP reversed the effect in 3 of these (one p-MEK2, 2 p-MKK4), while LR-SCS reversed 2 p-MKK4. Four phosphorylated MAPKs (p-JNK3, p-ERK1, p-ERK2 and p-ERK4) with 10 phosphorylated isoforms were found, with expression levels in 5 of them changed by the pain model. DTMP and LR-SCS reversed expression levels of two of them (p-ERK1 and p-ERK4) and one of them (p-ERK1) respectively. Downstream of MAPKs are additional proteins and kinases, such as MAPK activated protein kinases (MAPKAPKs) and MAPK interacting proteins (MAPKIPs), which regulate diverse outcomes such as proliferation, apoptosis, sensitization, etc. Of the 7 MAPKAP phosphoproteins distributed in 14 isoforms, the pain model affected expression levels of 3 of them (p-p90RSK, p-MSK2, p-MAPKAPK2). DTMP reversed expression levels of 2 of these (p-p90RSK, p-MSK2), whereas LR-SCS reversed the 3 of them. There were 8 isoforms of one p-MAPKIP identified (p-JIP3), with 2 of these changed by the pain model. Expression levels of one of these was reversed by DTMP, while LR-SCS reversed levels of the other one.
Discussion
In this work, we have used high-throughput techniques to assess proteomic changes and characterize the effect of SCS using DTMP or LR-SCS. We have utilized this approach for two main reasons: first, it was felt that it was important to move beyond transcriptomic analyses1,12,14 as proteins are the main effectors within tissues; second, instead of focusing on a very limited number of proteins determined a priori with conventional techniques, such as Western blot, we believe that a very complex phenomenon, such as neuropathic pain, requires a more comprehensive and powerful approach, as allowed by high-throughput proteomic analyses. Our previous transcriptomic analyses emphasized that the SNI pain model activated the immune system and inflammatory processes mediated by glial cells. It also showed that DTMP may alleviate neuropathic pain behavior by normalizing neuron-glial interactions in the spinal cord that have been disrupted by nerve injury.12 The effect of DTMP on the transcriptomics was found to be more profound that LR-SCS as DTMP significantly regulated more genes and biological processes associated with neuropathic pain toward the transcriptomic state of uninjured animals. The proteomic and phosphoproteomic analyses presented here corroborated the transcriptomics results. DTMP significantly regulated more proteins than LR-SCS, a similar occurrence in the transcriptomics. Similarly, many of the 81 proteins and genes, such as complement proteins (C3, C4A, C5, CFH), ITGB2, IP3R3, S100A8/9, and CNTF, found to be enriched in regulation by DTMP, are part of the inflammatory process associated with glia activation.
In this study, we also confirmed our previous observation that SCS regulated proteomic expression in an animal model of neuropathic pain. During the development and maintenance of chronic neuropathic pain, the activation of inflammatory cascades maintains hypersensitivity in the central nervous system. Here, we show that DTMP significantly affected expression levels of proteins involved in inflammatory states by reversing them toward those measured in uninjured animals, and by regulating other proteins that, although not significantly affected by the pain model, play an important role in inflammation regulation. Some healing-related or anti-inflammatory proteins may not be present in uninjured animals and may also remain unmodified by the SNI model. As such, the ability of DTMP to alter the expression of proteins unaffected by the pain model may indicate that the mechanism of action of DTMP occurs via activation of molecular pathways and not just inhibition of action potentials or neuronal signaling.
We directed our attention toward MAP kinases, which are a diverse group of intracellular proteins involved in a variety of pathways such as proliferation, differentiation, transformation, apoptosis, and inflammation. In particular, we studied MAP kinase proteins known to be involved in inflammation and, more specifically, modulation of NFκB pathway. Proteins identified and measured in this study are known to be involved in inflammation based on a GOEA (gene ontology enrichment analysis), that relies on curated information from published literature. Previous reports have emphasized the role of glial cells, particularly microglia and astrocytes, on inflammatory processes in chronic pain.27 For example, the activation of the complement system, specifically C3, is primarily observed in microglia cells within the CNS. Our results demonstrate that in the early stages of the SNI neuropathic pain model, DTMP down regulated key proteins of the complement system, such as C3, C4A and C5, which promote the production of pro-inflammatory agents as a response to nerve injury. These complement proteins primarily promote the NFκB pathway resulting in pro-inflammatory gene expression. CFH, which was increased by the SNI pain model and down regulated by DTMP, regulates C3 activity as does APP, which was decreased by the pain model and increased by DTMP. Additionally, C3 can be activated by TLR4 via cofactors such as LBP (increased by SNI and decreased by DTMP), HMGB1 (decreased by DTMP), and CAMP (decreased by DTMP), that were shown to be affected. Expression levels of CSRP1, a promoter of C4, were decreased by DTMP. Complement proteins, such as C3 and C4, activate the NFκB signaling pathway, which induces the production of proinflammatory agents including cytokines such as IL6, IL1β, TNFα, and the proteins CLU and CSRP1. The net effect of these changes could represent a decrease of the pro-inflammatory drive set in motion by the pain model.
Furthermore, activation of microglia and astrocytes is associated with calcium binding proteins S100A8 and S100A9, which can induce a pro-inflammatory response via membrane receptors such as TLR4 and RAGE that activate signaling cascades mediated by NFκB and AKT. Gliosis is also associated with neurotrophic factors such as CNTF, which is expressed by astrocytes. Expression levels of this protein were increased by the SNI model and decreased with DTMP. CNTF also activates the pro-inflammatory response by binding to the CNTF receptor, which then activates the AKT signaling pathway. This pathway plays a significant role in the generation and maintenance of chronic pain and has been shown to modulate activity of both MAPK as well as NFkB directly.28 As such, the inhibition of AKT by either reduced expression or phosphorylation is important in the therapeutic effect observed by SCS therapies. It is noteworthy that DTMP down regulates the expression of S100A8/9 as well as CNTF, thus potentially exerting a modulatory effect of the inflammatory response, likely through this AKT pathway.
Additionally, we have identified and quantified phosphorylated proteins, which are associated with the activation/deactivation of regulatory proteins related to inflammatory processes. Both DTMP and LR-SCS regulate expression levels of many of these phosphoproteins (see Figures 4 and 6) back toward expression levels found in uninjured animals. However, the effect of DTMP was more pronounced among the MAPK proteins. Out of the 63 isoforms identified, DTMP back-regulated 62% of them toward expression levels of uninjured animals, while LR-SCS back-regulated 48%. An interesting example is the activation of C3, which can lead to activation of ERK2 and ERK1 via phosphorylation, leading to increased transcription and cell survival.29 Phosphorylation of ERK2 can also promote cytoskeletal remodeling and cell migration.30 Regulation of phosphorylation via the MAPK cascade with DTMP may modulate the complement cascade and determine which pathway is activated.
Another remarkable difference in the effects of DTMP and LR-SCS was related to the expression levels of filament proteins like vimentin (VIM) and GFAP, which are markers of astrocyte activation and neuroinflammation in chronic pain.31 The pain model increased the expression levels of VIM and 12 p-VIMs, which were regulated back to levels found in uninjured animals by DTMP treatment in 11 of the p-VIMs. In contrast, LR-SCS only back-regulated levels of two of them. The pain model also increases expression levels of 14 p-GFAP isoforms. DTMP and LR-SCS back-regulated the expression levels of 7 and 6 of these isoforms, respectively.
The activation of these signaling pathways, most likely in astrocytes,32,33 is manifested by the increase in the expression of cytoplasmic filament proteins such as VIM, which in turn may mediate further signaling via phosphorylated isoforms (p-VIMs). Studies have shown that VIM (increased by SNI and decreased by DTMP) can function as a receptor of cell adhesion proteins and can induce NFκB via CAMK2 activation.34 NFκB activation further promotes VIM expression in a feed-forward loop. When activated, kinases like AKT phosphorylates VIM which modulates cell motility and caspase proteolysis. For instance, the expression level of p-VIM (at S39) was upregulated by the pain model and further increased by DTMP and LR-SCS. Interestingly, levels of the other 11 p-VIMs were found to be elevated by the pain model, and were all reversed by DTMP, whereas only two were reversed by LR-SCS. This indicates that DTMP has an enhanced down-regulatory effect on expression level changes in p-VIMs caused by the SNI. This seems to imply a better effect of DTMP in astrocyte deactivation, given that VIM is a marker of astrocyte activation.35
The mTOR pathway also plays a regulatory role in NFκB pathway activation in neurons as well as glia. As such, regulation of mTOR pathway activation may lead to a reduction in inflammatory processes.36 The pain model affected nine phosphorylated mTOR-related proteins, with 8 of them back-regulated by DTMP and 7 of them by LR-SCS.
Activation of PKC via phosphorylation can increase calcium influx by phosphorylation of voltage gated calcium channels (VDCC).37 Also, PKC can reduce pre-synaptic inhibitory tone at the presynaptic terminal by phosphorylation of opioid receptors and inhibitory GABA receptors, leading to reduced presynaptic inhibition. Furthermore, chronic use of opioid analgesics increases activity and expression of PKC that correlates with a decrease in analgesia. The pain model affected 16 (4 p-PKCe, 3 p-PKCg, 3 p-PKCa, 2 p-PCKt, 1 p-PKCd, 1 p-PKCh, 1 p-PKCi, and 1 p-PKCz) out of 20 different phosphorylated PKC isoforms. DTMP and LR-SCS reversed expression levels of 12 and 11 of these, back to levels found in uninjured animals, respectively. This finding might relate to a possible mechanism for analgesia for electrical current applied via SCS.
Some classical inflammatory proteins (such as IL1b, TNFa, etc.) were not identified in the analysis. This is a limitation of the experimental design for protein quantification, which may miss transient proteins or small peptides. Large proteins or those embedded in a membrane or attached to the cytoskeleton were most likely to be identified and quantified.
Conclusions
Proteomic and phosphoproteomic changes observed indicate a net reduction in the expression of proteins involved in the proinflammatory cascade. All these changes, taken together, may have the net effect of decreasing the inflammatory phenotype triggered by the pain model as a result of SCS therapy. This was observed through opposing effects on MAP kinase and NFκB pathways between the injury model and DTMP and, to a lesser extent, LR-SCS therapy. This reduction of injury-evoked responses by DTMP, observed in both proteomic and phosphoproteomic analyses, although it does not constitute direct evidence, supports the potential role for electrical current to reverse-modulate biological processes triggered by pain.
Abbreviations
CNS, central nervous system; SCS, spinal cord stimulation; LR-SCS, low rate SCS; DTMP, differential target multiplexed programming; NFκB, nuclear factor kappa-light-chain-enhancer of activated B cells; MAP, mitogen-activated protein; SNI, spared nerve injury; GOEA, gene ontology enrichment analysis; LC/TMS, liquid chromatography/tandem mass spectrometry; mTOR, mammalian target of rapamycin.
Author Contributions
All authors made substantial contributions to conception and design, acquisition of data, or analysis and interpretation of data; took part in drafting the article or revising it critically for important intellectual content; agreed to submit to the current journal; gave final approval of the version to be published; and agree to be accountable for all aspects of the work.
Funding
This work was supported by SGX Medical.
Disclosure
David L Cedeño, Dana M Tilley, David C Platt, and Ricardo Vallejo are consultants of SGX Medical. DLC and RV are consultants and advisory board members of Medtronic and co-inventors of patents related to differential target multiplexed SCS. David L Cedeño reports personal fees from Medtronic, during the conduct of the study; and in addition, has a patent Method and Apparatus for Multimodal Electrical Modulation of Pain issued to Medtronic. Dana M Tilley reports personal fees from SGX Medical, during the conduct of the study. David C Platt reports personal fees from SGX Medical LLC, during the conduct of the study. Ricardo Vallejo reports grants from SGX Medical, during the conduct of the study; personal fees from Medtronic, outside the submitted work; and in addition, has a patent Multiplexing signals for SCS issued, no license. The authors report no other potential conflicts of interest for this work.
References
1. Smith WJ, Cedeño DL, Thomas SM, Kelley CA, Vetri F, Vallejo R. Modulation of microglial activation states by spinal cord stimulation in an animal model of neuropathic pain: comparing high rate, low rate, and differential target multiplexed programming. Mol Pain. 2021;17:1744806921999013. doi:10.1177/1744806921999013
2. Ji RR, Berta T, Nedergaard M. Glia and pain: is chronic pain a gliopathy? Pain. 2013;154:S10–28. doi:10.1016/j.pain.2013.06.022
3. Ellis A, Bennett DL. Neuroinflammation and the generation of neuropathic pain. Br J Anaesth. 2013;111(1):26–37. doi:10.1093/bja/aet128
4. Ji RR, Nackley A, Huh Y, Terrando N, Maixner W. Neuroinflammation and central sensitization in chronic and widespread pain. Anesthesiolog. 2018;129(2):343–366. doi:10.1097/ALN.0000000000002130
5. Sommer C, Leinders M, Uceyler N. Inflammation in the pathophysiology of neuropathic pain. Pain. 2018;159(3):595–602. doi:10.1097/j.pain.0000000000001122
6. Deer TR, Grider JS, Lamer TJ, et al. A systematic literature review of spine neurostimulation therapies for the treatment of pain. Pain Med. 2020;21(7):1421–1432. doi:10.1093/pm/pnz353
7. Vallejo R, Gupta A, Cedeno DL, et al. Clinical effectiveness and mechanism of action of spinal cord stimulation for treating chronic low back and lower extremity pain: a systematic review. Curr Pain Headache Rep. 2020;24(11):70. doi:10.1007/s11916-020-00907-2
8. Stephens KE, Chen Z, Sivanesan E, et al. RNA-seq of spinal cord from nerve-injured rats after spinal cord stimulation. Mol Pain. 2018;14:1744806918817429. doi:10.1177/1744806918817429
9. Tilley DM, Lietz CB, Cedeno DL, Kelley CA, Li L, Vallejo R. Proteomic modulation in the dorsal spinal cord following spinal cord stimulation therapy in an in vivo neuropathic pain model. Neuromodulation. 2021;24(1):22–32. doi:10.1111/ner.13103
10. Vallejo R, Tilley DM, Cedeño DL, Kelley CA, DeMaegd M, Benyamin R. Genomics of the effect of spinal cord stimulation on an animal model of neuropathic pain. Neuromodulation. 2016;19(6):576–586. doi:10.1111/ner.12465
11. Shu B, He SQ, Guan Y. Spinal cord stimulation enhances microglial activation in the spinal cord of nerve-injured rats. Neurosci Bull. 2020;36(12):1441–1453. doi:10.1007/s12264-020-00568-6
12. Vallejo R, Kelley CA, Gupta A, Smith WJ, Vallejo A, Cedeño DL. Modulation of neuroglial interactions using differential target multiplexed spinal cord stimulation in an animal model of neuropathic pain. Mol Pain. 2020;16:1744806920918057. doi:10.1177/1744806920918057
13. Tilley DM, Cedeño DL, Vetri F, Platt DC, Vallejo R. Differential target multiplexed spinal cord stimulation programming modulates proteins involved in ion regulation in an animal model of neuropathic pain. Mol Pain. 2022;18:17448069211060181. doi:10.1177/17448069211060181
14. Cedeño DL, Smith WJ, Kelley CA, Vallejo R. Spinal cord stimulation using differential target multiplexed programming modulates neural cell-specific transcriptomes in an animal model of neuropathic pain. Mol Pain. 2020;16:1744806920964360. doi:10.1177/1744806920964360
15. Saha RN, Jana M, Pahan K. MAPK p38 regulates transcriptional activity of NF-kappaB in primary human astrocytes via acetylation of p65. J Immunol. 2007;179(10):7101–7109. doi:10.4049/jimmunol.179.10.7101
16. Merrell K, Southwick K, Graves SW, Esplin MS, Lewis NE, Thulin CD. Analysis of low-abundance, low-molecular-weight serum proteins using mass spectrometry. J Biomol Tech. 2004;15(4):238–248.
17. Radulovic M, Godovac-Zimmermann J. Proteomic approaches to understanding the role of the cytoskeleton in host-defense mechanisms. Expert Rev Proteomics. 2011;8(1):117–126. doi:10.1586/epr.10.91
18. Tuli L, Ressom HW. LC-MS Based Detection of Differential Protein Expression. J Proteomics Bioinform. 2009;2:416–438. doi:10.4172/jpb.1000102
19. Gelderd JB, Chopin SF. The vertebral level of origin of spinal nerves in the rat. Anat Rec. 1977;188(1):45–47. doi:10.1002/ar.1091880106
20. Ting L, Rad R, Gygi SP, Haas W. MS3 eliminates ratio distortion in isobaric multiplexed quantitative proteomics. Nat Methods. 2011;8:937–940. doi:10.1038/nmeth.1714
21. Zhang L, Elias JE. Relative protein quantification using tandem mass tag mass spectrometry. Methods Mol Biol. 2017;1550:185–198.
22. McAlister GC, Nusinow DP, Jedrychowski MP, et al. MultiNotch MS3 enables accurate, sensitive, and multiplexed detection of differential expression across cancer cell line proteomes. Anal Chem. 2014;86(14):7150–7158. doi:10.1021/ac502040v
23. Eng JK, McCormack AL, Yates JR. An approach to correlate tandem mass spectral data of peptides with amino acid sequences in a protein database. J Am Soc Mass Spectrom. 1994;5(11):976–989. doi:10.1016/1044-0305(94)80016-2
24. Szklarczyk D, Gable AL, Lyon D, et al. STRING v11: protein-protein association networks with increased coverage, supporting functional discovery in genome-wide experimental datasets. Nucleic Acids Res. 2019;47(D1):D607–D613. doi:10.1093/nar/gky1131
25. Thomas PD, Campbell MJ, Kejariwal A, et al. PANTHER: a library of protein families and subfamilies indexed by function. Genome Res. 2003;13(9):2129–2141. doi:10.1101/gr.772403
26. Yue X, Schunter A, Hummon AB. Comparing multistep immobilized metal affinity chromatography and multistep TiO2 methods for phosphopeptide enrichment. Anal Chem. 2015;87(17):8837–8844. doi:10.1021/acs.analchem.5b01833
27. Vallejo R, Tilley DM, Vogel L, Benyamin R. The role of glia and the immune system in the development and maintenance of neuropathic pain. Pain Pract. 2010;10(3):167–184. doi:10.1111/j.1533-2500.2010.00367.x
28. Chen SP, Zhou YQ, Liu DQ, et al. PI3K/Akt pathway: a potential therapeutic target for chronic pain. Curr Pharm Des. 2017;23(12):1860–1868. doi:10.2174/1381612823666170210150147
29. Xu B, Yang L, Lye RJ, Hinton BT. p-MAPK1/3 and DUSP6 regulate epididymal cell proliferation and survival in a region-specific manner in mice. Biol Reprod. 2010;83(5):807–817. doi:10.1095/biolreprod.110.085613
30. Lavoie H, Gagnon J, Therrien M. ERK signaling: a master regulator of cell behaviour, life and fate. Nat Rev Mol Cell Biol. 2020;21(10):607–632. doi:10.1038/s41580-020-0255-7
31. Wilhelmsson U, Li L, Pekna M, et al. Absence of glial fibrillary acidic protein and vimentin prevents hypertrophy of astrocytic processes and improves post-traumatic regeneration. J Neurosci. 2004;24(21):5016–5021. doi:10.1523/JNEUROSCI.0820-04.2004
32. Cristovao JS, Gomes CM. S100 proteins in Alzheimer’s disease. Front Neurosci. 2019;13:463. doi:10.3389/fnins.2019.00463
33. Winter CG, Saotome Y, Levison SW, Hirsh D. A role for ciliary neurotrophic factor as an inducer of reactive gliosis, the glial response to central nervous system injury. Proc Natl Acad Sci U S A. 1995;92(13):5865–5869. doi:10.1073/pnas.92.13.5865
34. Huang SH, Chi F, Peng L, et al. Vimentin, a novel NF-κB regulator, is required for meningitic Escherichia coli k1-induced pathogen invasion and pmn transmigration across the blood-brain barrier. PLoS One. 2016;11(9):e0162641. doi:10.1371/journal.pone.0162641
35. Xue P, Chen L, Lu X, et al. Vimentin promotes astrocyte activation after chronic constriction injury. J Mol Neurosci. 2017;63(1):91–99. doi:10.1007/s12031-017-0961-6
36. Saxton RA, Sabatini DM. mTOR signaling in growth, metabolism, and disease. Cell. 2017;169(2):361–371. doi:10.1016/j.cell.2017.03.035
37. Velázquez KT, Mohammad H, Sweitzer SM. Protein kinase C in pain: involvement of multiple isoforms. Pharmacol Res. 2007;55(6):578–589. doi:10.1016/j.phrs.2007.04.006
 © 2022 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2022 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.