Back to Journals » Drug Design, Development and Therapy » Volume 19
Preventive Effect of Oleanolic Acid on Ulcerative Colitis Caused by Dextran Sulfate Sodium via a PPARγ-Involved Mechanism in Mice
Authors Tian G, Dong F ![]() , Fu A
, Fu A ![]() , Lai S
, Lai S ![]() , Yu J
, Yu J ![]() , Ni J, Ding Q, Jin Y
, Ni J, Ding Q, Jin Y ![]()
Received 11 September 2025
Accepted for publication 13 December 2025
Published 25 December 2025 Volume 2025:19 Pages 11641—11657
DOI https://doi.org/10.2147/DDDT.S560422
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Professor Anastasios Lymperopoulos
Guojiang Tian,1,* Fan Dong,2,* Ai Fu,3,* Shanglei Lai,4 Jundi Yu,2 Jingjing Ni,5 Qinchao Ding,2 Yueping Jin5
1Department of Anus and Colorectal Surgery, Shaoxing People’s Hospital, Shaoxing, Zhejiang, People’s Republic of China; 2School of Public Health, Zhejiang Chinese Medical University, Hangzhou, Zhejiang, People’s Republic of China; 3Institute of Hepatology and Epidemiology, Affiliated Hangzhou Xixi Hospital, Zhejiang Chinese Medical University, Hangzhou, Zhejiang, People’s Republic of China; 4Department of Medical Research Center, Shaoxing People’s Hospital, Shaoxing, Zhejiang, People’s Republic of China; 5Department of Gastroenterology, Shaoxing People’s Hospital, Shaoxing, Zhejiang, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Qinchao Ding, School of Public Health, Zhejiang Chinese Medical University, No. 548, Binwen Road, Binjiang District, Hangzhou, Zhejiang, 310053, People’s Republic of China, Email [email protected] Yueping Jin, Department of Gastroenterology, Shaoxing People’s Hospital, No. 568, Zhongxing North Road, Yuecheng District, Shaoxing, Zhejiang, 312000, People’s Republic of China, Email [email protected]
Purpose: Oleanolic acid (OA), a naturally occurring pentacyclic triterpenoid, has been reported to possess anti-inflammatory and barrier-protective properties. However, its molecular mechanism in the context of ulcerative colitis (UC) remains unclear.
Methods: In this study, we evaluated the therapeutic potential of OA (25 and 50 mg/kg/day) in a dextran sulfate sodium (DSS)-induced acute colitis model in C57BL/6 mice.
Results: Oral administration of OA significantly alleviated clinical symptoms of colitis, including weight loss, colon shortening, increased disease activity index (DAI), and histopathological injury. Additionally, OA demonstrated potent anti-inflammatory effects by suppressing the production of pro-inflammatory cytokines and promoting anti-inflammatory cytokines. It also restored redox homeostasis by enhancing antioxidant defenses and reducing lipid peroxidation. OA also restored intestinal barrier integrity, as evidenced by increased tight junction protein expression and decreased intestinal permeability. Mechanistically, OA was found to inhibit the activation of the NF-κB signaling pathway, as evidenced by reduced phosphorylation of IKKα/β and p65, and decreased degradation of IκBα. A key finding was that OA restored the expression of PPARγ, a known negative regulator of NF-κB. OA promoted the accumulation of peroxisome proliferator-activated receptor gamma (PPARγ), a nuclear receptor essential for maintaining intestinal epithelial homeostasis and anti-inflammatory signaling. OA inhibited the ubiquitin-mediated proteasomal degradation of PPARγ, thereby stabilizing its protein levels in colonic tissue. Pharmacological inhibition of PPARγ with GW9662 abolished the protective effects of OA on intestinal inflammation and epithelial barrier function, confirming that PPARγ activation is required for OA-mediated protection.
Conclusion: These findings identify a previously unrecognized mechanism whereby OA protects against colitis through post-translational stabilization of PPARγ. Our study not only highlights the therapeutic value of OA as a potential intervention for UC but also provides mechanistic insight into the regulation of nuclear receptor homeostasis during intestinal inflammation.
Keywords: oleanolic acid, ulcerative colitis, inflammation, PPARγ, intestinal barrier function
Introduction
Ulcerative colitis (UC), a major subtype of inflammatory bowel disease (IBD), is a chronic relapsing inflammatory disorder characterized by continuous mucosal inflammation originating in the rectum and extending proximally through the colon.1 Patients commonly present with diarrhea, hematochezia, and, in severe cases, an elevated risk of colorectal cancer.2 Although the etiology of UC remains incompletely understood, accumulating evidence highlights the interplay among intestinal barrier disruption, dysregulated immune responses, oxidative stress, and genetic susceptibility.3,4
Accumulating evidence indicates that disruption of the intestinal epithelial barrier is a central event in UC pathogenesis. Impaired tight junction (TJ) integrity and mucus depletion lead to increased intestinal permeability, which facilitates luminal antigen infiltration and amplifies mucosal inflammation.5 Activated macrophages and neutrophils in UC lesions produce excessive reactive oxygen species (ROS) through respiratory burst, and mitochondrial dysfunction in epithelial cells further contributes to ROS overproduction.6,7 This oxidative stress induces lipid peroxidation, protein dysfunction, and epithelial injury, forming a vicious cycle that aggravates barrier damage.8–10 Concurrently, pro-inflammatory cytokines such as IL-1β, IL-6, and TNF-α activate the NF-κB signaling pathway, a key driver of immune–inflammatory responses in UC.11–13 NF-κB activation is consistently observed in UC patients and DSS-induced colitis models,14,15 whereas its upstream negative regulator PPARγ is reduced in colonic epithelium, contributing to sustained inflammation.16 Given the global rise of UC incidence and the limitations of current anti-inflammatory or immunosuppressive therapies—including long-term adverse effects17,18—there is an urgent need for safer and more effective therapeutic strategies.
Given the limitations and adverse effects of current UC therapies, there is growing interest in identifying natural compounds with improved safety profiles. Oleanolic acid (OA), a pentacyclic triterpenoid widely distributed in plants such as olives and apple peels, has been reported to exert anti-inflammatory, antioxidant, hepatoprotective, and metabolic regulatory effects.19–21 Increasing evidence suggests that OA can modulate systemic inflammation and oxidative stress, as demonstrated by its ability to reduce circulating IL-1β, IL-6, TNF-α, and MDA levels in metabolic disease models.20,22 Recent studies further highlight the potential relevance of OA in intestinal inflammation: OA activates the bile acid receptor TGR5 to inhibit NF-κB–mediated inflammatory signaling,23 and it may also rebalance mucosal immunity by suppressing Th17 differentiation while promoting Treg cell development.24 Although these findings suggest that OA is a promising candidate for IBD management, the precise mechanisms by which OA mitigates DSS-induced acute colitis—particularly in relation to oxidative stress, epithelial barrier protection, and inflammatory signaling—remain to be fully elucidated. Our study aims to clarify these mechanisms and provide new insights into the therapeutic potential of OA in UC.
In this study, we established a DSS-induced acute colitis model in C57BL/6 mice to evaluate the protective effects of OA and to elucidate its underlying mechanisms. We found that OA markedly alleviated DSS-induced disease severity, attenuated oxidative stress, improved epithelial barrier integrity, and reduced colonic inflammation. Mechanistic analyses further identified PPARγ as a key mediator contributing to the therapeutic actions of OA in UC. Together, these findings indicate that OA may serve as a promising natural compound for the treatment of ulcerative colitis.
Material and Methods
Animals and Experimental Design
The experimental subjects were 8-week-old male C57BL/6 mice, which were purchased from Shanghai SLAC Laboratory Animal Co. Ltd, Shanghai, China. The experiment was approved by the Animal Experimental Ethics Committee of Zhejiang Chinese Medical University (approval number IACUC-20250113-09) and following the ARRIVE guidelines. According to the Guide for the Care and Use of Laboratory Animals, all mice were housed in cages, with four mice per cage, in an environment maintained at 23 ± 2°C, 55 ± 5% relative humidity, and a 12-hour light-dark cycle. After a 7-day adaptation period, the mice were randomly divided into four groups: a) the control group; b) the DSS group; c) the DSS + OA25 (DSS + OA:25 mg/kg/day) group; d) the DSS + OA50 (DSS + OA:50 mg/kg/day) group. There were 8 mice in each group. A high-dose OA suspension (5 mg/mL) was prepared by sonicating OA in saline and combining it with 0.5% CMC-Na solution, and was then diluted with the 0.5%CMC-Na solution to yield a low-dose formulation (2.5 mg/mL). The experimental timeline of the UC model is shown in Figure 1A. Apart from the control group, the other three groups received 3% DSS diluted in drinking water for 7 consecutive days to induce colitis. OA was administered at doses of 25 mg/kg and 50 mg/kg once daily by gavage for 3 days before DSS treatment and continued at the same frequency throughout the DSS treatment period. Meanwhile, the control group and the DSS group were treated with physiological saline for 10 days by gavage before euthanasia. The body weights of mice in each group were recorded daily. All mice were sacrificed with pentobarbital solution (30 mg/kg body weight, ip) on day 10, and the colon tissue and plasma were collected from the mice for further analysis.
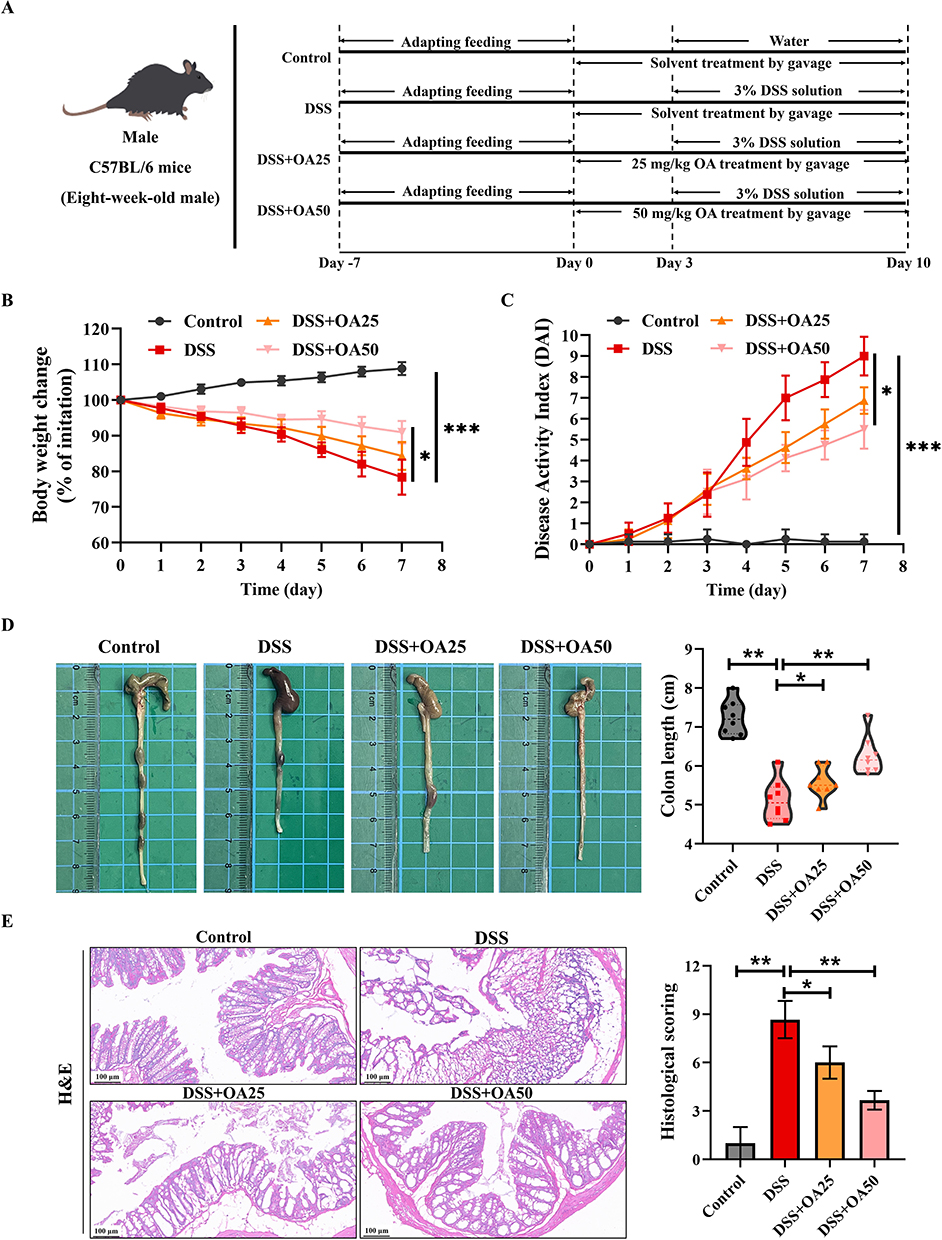 |
Figure 1 OA alleviates DSS-induced acute colitis in mice. (A) Schematic of the experimental design. Control, DSS, DSS+OA25 (25 mg/kg/day OA), and DSS+OA50 (50 mg/kg/day OA). (B) Daily changes in body weight during DSS treatment. (C) DAI scores recorded during the experiment. (D) Representative images and quantification of colon length in each group. (E) Representative H&E-stained sections of colon sections and corresponding histological scores. All data were presented as mean ± SD, n =8 per group. *P < 0.05, **P < 0.01, ***P < 0.001 vs corresponding control. |
To further explore the role of PPARγ in the amelioration of colitis with OA treatment, GW9662, an inhibitor of PPARγ, was used in the next experiment. The experiment was divided into the control group, the DSS group, DSS + OA50 group, DSS + OA50 + GW9662 group. OA was given at doses of 50 mg/kg for 10 days. GW9662 (1 mg/kg) was injected intraperitoneally every day for 7 days, starting on the same day as DSS administration. 0.1 mg/mL GW9662 working solution in 1% DMSO/saline was prepared by first dissolving the compound in DMSO (10 mg/mL) and then diluting it in physiological saline.
Oleanolic acid (OA, Cat. No: B20954; Purity ≥ 98%) was purchased from Shanghai Yuanye Biotechnology Co., Ltd (Shanghai, China). GW9662 (Cat. No: HY-16578) was purchased from MedChemExpress (State of New Jersey, United States). Dextran sulphate sodium (DSS, Cat. No: D491691-40k, MW40000) was purchased from Aladdin (Shanghai, China).
Disease Activity Index (DAI) Score
The DAI score is a composite index that combines the percentage of body weight loss, stool consistency, and fecal bleeding in mice and is used to assess the severity of colitis. Body weights and DAI scores of mice were recorded daily from the first day of DSS administration. The DAI scores were presented as a sum of the results for: body weight loss (0 for unchanged weight, 1 for 1–5%, 2 for 5–10%, 3 for 10–20%, and 4 for greater than 20%), stool consistency (0 for normal, 1 for feces adhering slightly, 2 for loose stools, 3 for slurry stools, and 4 for diarrhea), and stool bleeding (0 for normal, 1 for weak positive occult blood test, 2 for occult blood positivity, 3 for strong positive occult blood test, and 4 for overt bleeding).25
Histological Examination
Portions of the colonic tissues were fixed in paraformaldehyde and subsequently dehydrated and embedded in paraffin wax. The embedded tissues were cut into 5-µm thick sections and finally stained with hematoxylin and eosin (H&E). The images were observed under a light microscope at 200× magnification (Olympus, Japan). H&E staining kit was obtained from Beijing Solarbio Science Research Institute. The histopathological scores were calculated based on the extent of crypt injury, damage to epithelial cells, and inflammatory infiltration.26
Cell Culture
Caco-2 cells were purchased from the Procell Life Science Technology Co. Ltd (CL-0050) and cultured in Modified Eagle’s Medium (MEM) supplemented with 20% (v/v) fetal bovine serum (FBS), and 1% non-essential amino acids. The cells were maintained at 37°C in a humidified atmosphere containing 5% CO2.
Biochemical Analysis
Samples of colon tissue were homogenized using a homogenizer and then centrifuged at 3000 rpm for 15 min to collect the supernatants. Subsequently, the indicator of inflammation, myeloperoxidase (MPO) activity, was assayed using a commercial kit (Nanjing jiancheng, Nanjing, China) according to the manufacturer’s recommended protocol. Additionally, the status of oxidative stress in the colon, including superoxide dismutase (SOD), catalase (CAT), malondialdehyde (MDA), glutathione peroxidase (GSH-Px), glutathione (GSH), and GSH/oxidized glutathione (GSH/GSSG), was detected using commercial assay kits, which were obtained from Beyotime Biotechnology Research Institute (Nanjing, China).
Enzyme-Linked Immunosorbent Assay (ELISA)
The levels of cytokines, including IL-1β, IL-6, TNF-α, IL-4, and IL-10, were detected in both plasma and colon tissue utilizing ELISA kits (Abcam, Cambridge, UK), in accordance with the manufacturer’s instructions.
Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR)
Total RNA was extracted from colon tissues utilizing the TRIzol® reagent (Invitrogen, Carlsbad) according to standardized protocols. Subsequently, the extracted RNA was transcribed into cDNA following the rigorous guidelines provided by the reverse transcription kit (Promega, Madison, WI). Amplification of the cDNA was performed in a 384-well PCR plate utilizing the SYBR green/ROX qPCR premix (2X) within the Quantstudio™ 6 Pro real-time PCR system (Thermo Fisher Scientific), ensuring precision and reproducibility. The expression levels of the genes of interest were normalized against the level of 18s rRNA and quantitatively analyzed employing the 2−ΔΔCt method. All primers employed in this study were custom-synthesized by Sangon Biotechnology Co. Ltd., and their specific sequences are detailed in Supplementary Table 1, facilitating reproducibility and validation of the experimental results.
Western-Blot Analysis
The appropriate quantities of colon tissues were excised and immersed in protein lysis buffer for homogenization. Following homogenization, the mixtures were centrifuged at 12000 rpm for a duration of 15 min at a temperature of 4°C, facilitating the segregation of insoluble components from the soluble fraction. Post-centrifugation, the supernatant, enriched with soluble proteins, was meticulously aspirated to avoid contamination with the sediment. Subsequently, the BCA assay was employed to quantitatively determine the protein concentration within the supernatant, enabling the precise preparation of loading samples commensurate with the measured concentration. Ultimately, Western-blot analysis was performed to assess the expression levels of diverse proteins within the samples. β-actin was used as the internal control. Bands were quantified using the ImageJ software. The antibodies are listed in Supplementary Tables 2 and 3.
Immunohistochemistry (IHC) Analysis
The expression of Zona Occludens 1 (ZO-1), Occludin, and Claudin-1 proteins in colon tissues were assessed using IHC staining. Tissue sections were deparaffinized in xylene and then hydrated in a graded alcohol series. After antigenic repair of the sections in sodium citrate buffer, endogenous peroxidase was inactivated by treatment with 3% H2O2 solution for 30 minutes. The sections were then blocked with goat serum for 30 min at room temperature. The sections were incubated with primary antibody overnight at 4°C and washed three times with phosphate-buffered saline (PBS). The sections are then incubated with biotin-labeled secondary antibody for 20 min. The sections were then stained with 3,3′-diaminobenzidine tetrahydrochloride (DAB) as a chromogen, counterstained with hematoxylin and dehydrated, sealed and visualized under a microscope (Olympus, Japan).
Molecular Docking
The mutual binding of PPARγ and oleanolic acid was explored by molecular docking. The 3D structure of PPARγ was retrieved from the AlphaFold Protein Structure Database (https://alphafold.ebi.ac.uk/).27,28 Hydrogen atoms were added to the protein structure utilizing Autodock Tools 1.5.7 software. The chemical structure of oleanolic acid was obtained from the PubChem database (https://pubchem.ncbi.nlm.nih.gov), subsequently imported into ChemBioDraw 3D to generate the 3D chemical structure and to calculate the minimum energy conformation, which was then saved for subsequent docking procedures. Thereafter, the PPARγ protein structure and the 3D chemical structure of oleanolic acid were imported into Autodock Tools 1.5.7 for docking analysis. The docking outcomes were subsequently analyzed and visualized using PyMOL.
Statistical Analysis
All data were analyzed using GraphPad Prism 8.0.1 software (GraphPad Software, CA) and were represented as mean ± standard deviation (SD). Two groups were compared by two-tailed Student t tests. One-way analysis of variance (ANOVA) followed by the Turkey multiple-comparison test was used to determine multiple groups. P < 0.05 was considered to represent statistical significance.
Results
OA Alleviates DSS-Induced Acute Colitis in Mice
A murine model of the UC was established by administering 3% DSS in drinking water for 7 consecutive days (Figure 1A). The results showed that mice in the DSS group exhibited significant weight loss and a progressive increase in DAI score compared to the control group (Figure 1B and C). Notably, the treatment with OA at a dosage of 50 mg/kg/day significantly alleviated weight loss, diarrhea, and rectal bleeding (Figure 1B and C). Additionally, DSS exposure also significantly reduced colon length relative to the control group. OA treatment markedly preserved colon length. (Figure 1D). These findings indicate that OA confers therapeutic benefit against DSS-induced acute colitis.
Representative H&E staining histological sections and histological scores are shown in Figure 1E. Colonic tissues from control mice exhibited normal architecture, including intact epithelial lining, organized mucosal structure, and absence of inflammatory cell infiltration. In contrast, DSS exposure led to hallmark pathological features, such as goblet cell depletion, epithelial erosion, crypt loss, submucosal edema, and extensive inflammatory cell infiltration. Meanwhile, the histological score of the DSS group was significantly higher than that of the control group (Figure 1E). OA treatment markedly alleviated these pathological alterations, as evidenced by reduced edema, improved epithelial integrity, diminished inflammatory cell infiltration, and lower histological scores.
OA Attenuates DSS-Induced Intestinal Inflammation and Cytokine Imbalance in Mice
To evaluate intestinal inflammation, MPO levels were measured in the colonic tissues of mice. The MPO levels were significantly elevated in the DSS group compared to controls, indicating increased neutrophil infiltration. OA treatment significantly suppressed this elevation (Figure 2A). Further analysis revealed that DSS markedly increased the colonic levels of pro-inflammatory cytokines IL-1β, IL-6, and TNF-α, while reducing anti-inflammatory cytokines IL-4 and IL-10 in colonic tissues compared to the control group (Figure 2B–F). Similar cytokine profiles were observed in plasma samples (Figure 2G–L), all of which were significantly reversed by OA administration. Consistent with these findings, qPCR analysis showed upregulated mRNA expression of Il-1β, Il-6, and Tnf-α, and downregulated mRNA expression of Il-4 and Il-10 in DSS-treated colons. OA supplementation restored these dysregulated inflammatory gene expressions (Supplementary Figure 1A). These results demonstrate that OA effectively mitigates DSS-induced intestinal inflammation, likely by rebalancing local and systemic pro- and anti-inflammatory cytokine responses.
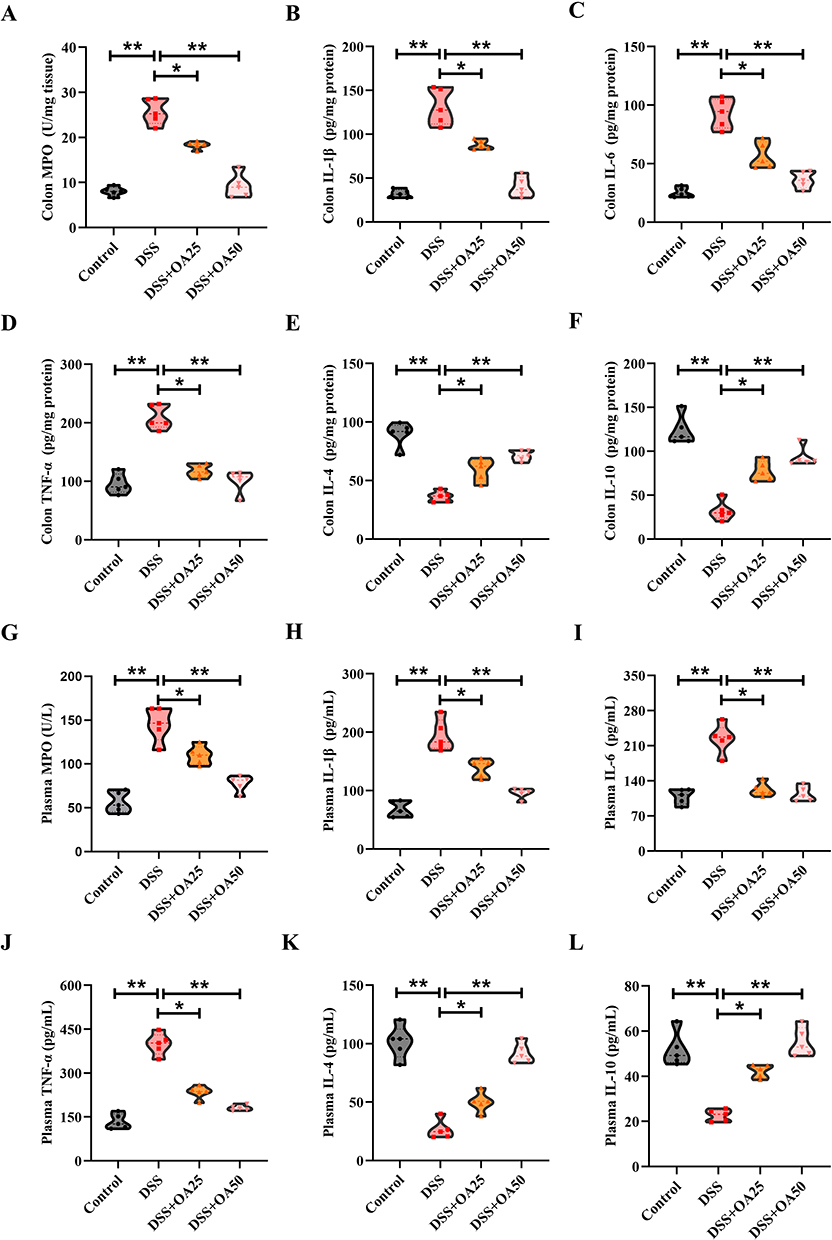 |
Figure 2 OA attenuates DSS-induced intestinal inflammation and cytokine imbalance in mice. (A) The activity of myeloperoxidase (MPO) in colonic tissues was assessed. The levels of (B)IL-1β, (C)IL-6, (D)TNF-α, (E)IL-4 and (F)IL-10 in colonic homogenates were quantitatively assessed using ELISA kits, following the manufacturer’s protocols. (G) The levels of MPO in plasma were assessed. The levels of (H) IL-1β, (I) IL-6, (J) TNF-α, (K) IL-4, and (L) IL-10 in plasma were quantitatively assessed using ELISA kits. All data were presented as mean ± SD, n =5 per group. *P < 0.05, **P < 0.01 vs corresponding control. |
OA Alleviates DSS-Induced Intestinal Oxidative Stress in Mice
Oxidative stress is a key pathological feature of DSS-induced acute colitis. To assess the effect of OA on redox homeostasis, we examined key oxidative stress markers and antioxidant defenses in colonic tissues. DSS exposure significantly impaired antioxidant capacity, as evidenced by reduced activities of SOD and CAT (Figure 3A and B) along with increased MDA levels, a marker of lipid peroxidation (Figure 3C). Moreover, DSS treatment markedly decreased GSH-Px activity, GSH level, and the GSH/GSSG ratio in the colon (Figure 3D–F), indicating compromised redox balance. Notably, OA supplementation significantly reversed these alterations, suggesting that OA protects against DSS-induced oxidative stress by restoring antioxidant defenses.
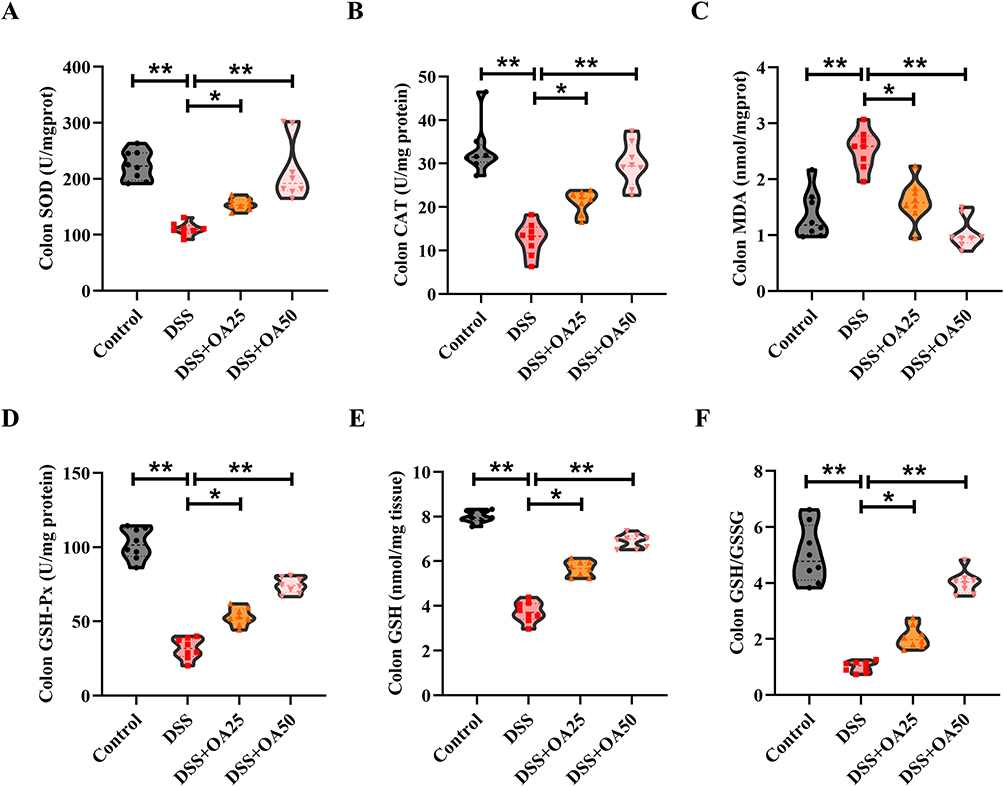 |
Figure 3 OA alleviates DSS-induced intestinal oxidative stress in mice. (A) The activities of SOD in colonic tissues. (B) The activities of CAT in colonic tissues. (C) The concentrations of MDA in colonic tissues. (D) The activities of GSH-Px in colonic tissues. (E and F) Colonic concentrations of GSH and the GSH/GSSG. All data were presented as mean ± SD, n =8 per group. *P < 0.05, **P < 0.01 vs corresponding control. |
OA Improves Intestinal Barrier Function in Mice with DSS-Induced Acute Colitis
Tight junction (TJ) proteins such as ZO-1, Occludin, and Claudin-1 are essential for maintaining intestinal epithelial barrier integrity. RT-qPCR analysis showed that DSS exposure significantly reduced the mRNA expression levels of ZO-1, Occludin, and Claudin-1 in colonic tissues, compared to the control group. Notably, OA treatment restored the expression of these TJ-related genes (Figure 4A–C). Consistently, immunohistochemical (IHC) analysis revealed a marked reduction in ZO-1, Occludin, and Claudin-1 protein expression in the DSS group, which was significantly reversed by OA administration (Figure 4D). These findings indicate that OA preserves intestinal barrier integrity during colitis, at least in part, by upregulating the expression of key TJ proteins.
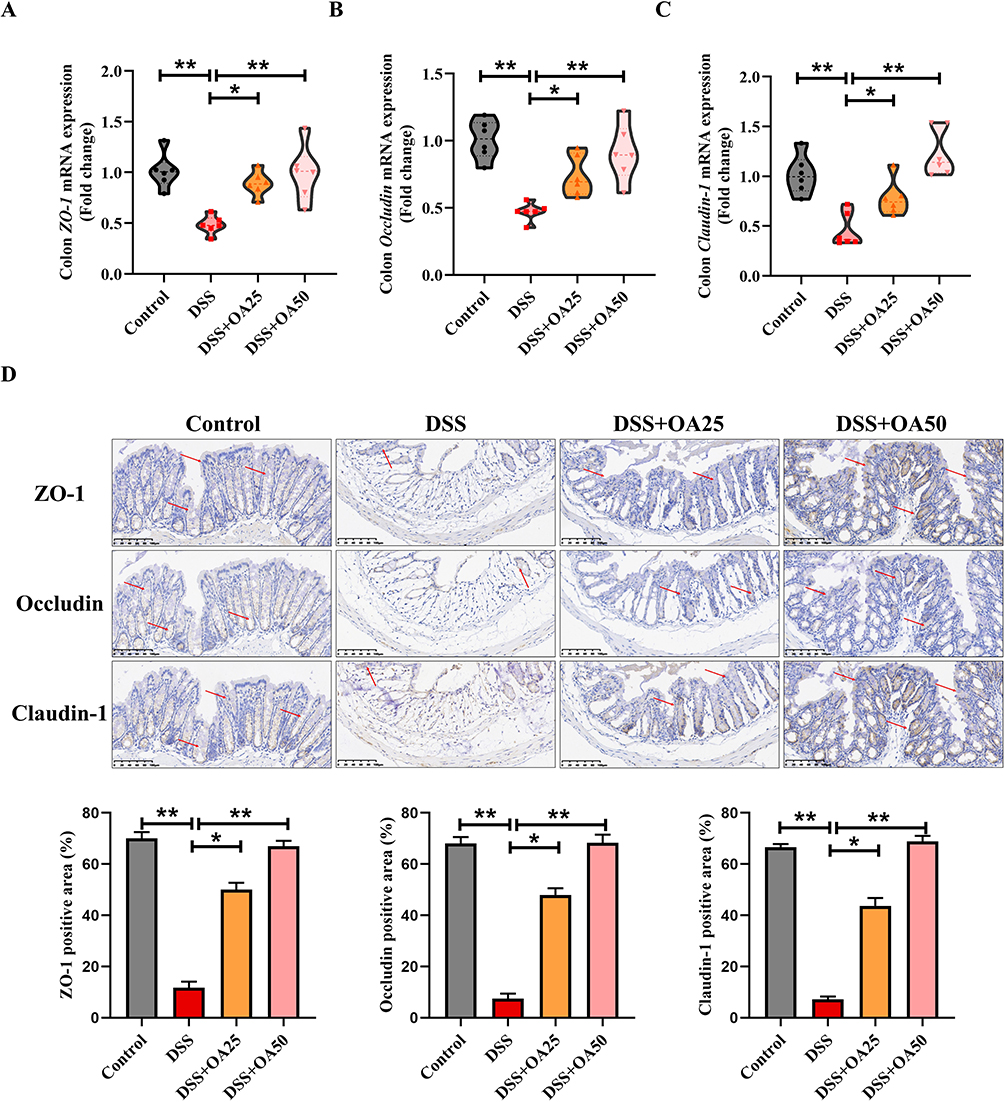 |
Figure 4 OA improves intestinal barrier function in mice with DSS-induced acute colitis. (A-C) The mRNA expression levels of ZO-1 (A), Occludin (B), and Claudin-1 (C) in colonic tissues were measured by qRT-PCR analysis. (D) Representative IHC images of ZO-1, Occludin, and Claudin-1 in colonic tissues. Semiquantitative analysis of ZO-1, Occludin, and Claudin-1 using ImageJ. All data were presented as mean ± SD, n =6 per group. *P < 0.05, **P < 0.01 vs corresponding control. |
OA Restores PPARγ Expression and Inhibits the Activation of the NF-κB Signaling Pathway
Activation of the NF-κB signaling cascade plays a central role in the pathogenesis of UC. To determine whether OA exerts anti-inflammatory effects via this pathway, we assessed the protein levels of phosphorylated p65 (p-p65), phosphorylated IKKα/β (p-IKKα/β), IKBα, and iNOS in the colonic tissues. Compared with the control group, DSS treatment significantly increased the expression of p-p65, p-IKK α/β, and iNOS, while markedly reducing IκBα levels. Notably, OA treatment reversed these alterations, indicating a suppressive effect on IKK/NF-κB activation (Figure 5A and B). Given that PPARγ is known to negatively regulate NF-κB signaling,29 we further investigated the effect of OA on PPARγ expression. Molecular docking was employed to elucidate the interaction between the protein and the drug. The analysis revealed that OA interacts with the catalytic domain of PPARγ via hydrogen bonding at LYS-184, with the binding energy of the docked PPARγ and OA complex being −8.3 kcal/mol (Figure 5C), suggesting potential ligand-receptor affinity. Consistent with this, DSS markedly downregulated PPARγ protein levels in colonic tissues, whereas OA treatment significantly restored its expression (Figure 5D). To explore the underlying mechanism of PPARγ downregulation, we examined the role of the ubiquitin-proteasome system (UPS), a known pathway for PPARγ degradation.30,31 Upon treatment with DSS, the expression of PPARγ was significantly downregulated in Caco-2 cells. However, this reduction was effectively abrogated by treatment with the proteasome inhibitor MG132, highlighting the involvement of the UPS in PPARγ degradation under inflammatory conditions (Figure 5E). Concurrently, we investigated the effects of OA on PPARγ expression in DSS-treated Caco-2 cells. Consistent with the results obtained from animal experiments, OA treatment significantly reversed the DSS-induced downregulation of PPARγ. Intriguingly, the presence of MG132 further potentiated this effect, suggesting that OA may modulate PPARγ stability through mechanisms that intersect with the UPS (Figure 5F). These findings suggest that OA not only upregulates PPARγ expression but may also stabilize it by attenuating UPS-mediated degradation, thereby contributing to its anti-inflammatory effects in DSS-induced colitis.
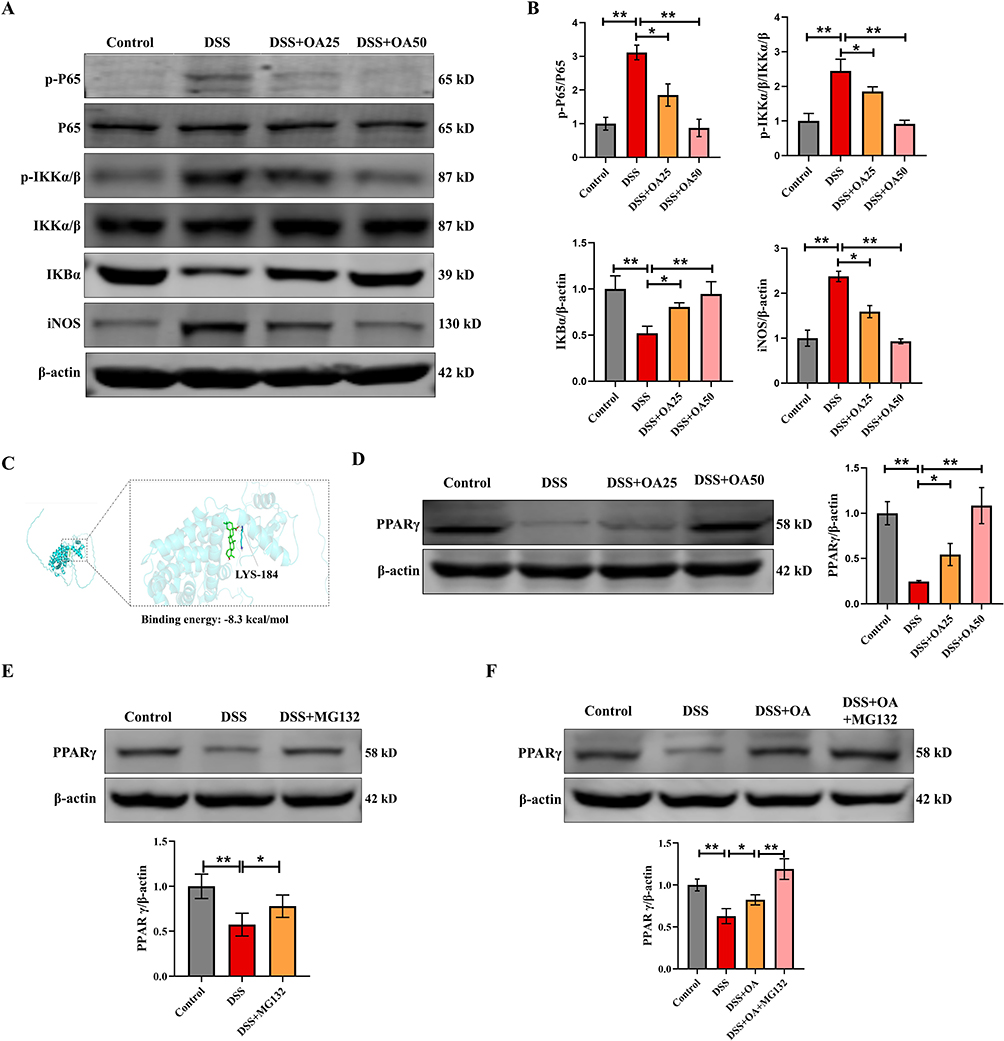 |
Figure 5 OA restores PPARγ expression and inhibits the activation of the NF-κB signaling pathway. (A) Western blot analysis of colonic NF-κB(P65), p-P65, p-IKKα/β, IKKα/β, IKBα, and iNOS protein expression levels. (B) P65, p-P65, p-IKKα/β, IKKα/β, IKBα, and iNOS were quantified using ImageJ software to provide a semiquantitative assessment of protein expression levels. (C) Molecular docking analysis illustrating the interaction of OA with the catalytic site of PPARγ. (D) The protein expression levels of colonic PPARγ were evaluated via Western blot. (E) Caco-2 cells were treated with DSS (50 mg/mL) for 6 h with or without a 1 h pretreatment with MG132 (10 μM). PPARγ protein expression levels were evaluated via Western blot. (F) Caco-2 cells were treated with DSS (50 mg/mL) for 6 h with or without a 1 h pretreatment with MG132 (10 μM) and OA (10 μM). PPARγ protein expression levels were evaluated via Western blot. β-actin was used as a loading control. All data were presented as mean ± SD, n =6 per group. *P < 0.05, **P < 0.01 vs corresponding control. |
GW9662 Eliminates the Inhibitory Effects of OA on DSS-Induced Ulcerative Colitis Symptoms in Mice
To further verify the involvement of PPARγ in the protective effects of OA against DSS-induced colitis, the specific PPARγ antagonist GW9662 was administered intraperitoneally (Figure 6A). Consistent with the results of previous experiments, DSS exposure led to significant body weight loss, colon shortening, and increased DAI scores compared to the control group. As expected, OA treatment (50 mg/kg/day) markedly alleviated these colitic symptoms, evidenced by improved weight maintenance, extended colon length, and reduced DAI scores (Figure 6B–D). However, co-administration of GW9662 significantly reversed these beneficial effects of OA. Specifically, mice in the GW9662 group showed markedly shortened colon length and increased DAI scores compared to those in the OA group (Figure 6B–D). Histopathological analysis further confirmed these findings. While OA administration notably improved DSS-induced mucosal damage, characterized by reduced epithelial disruption and inflammatory infiltration, the introduction of GW9662 abolished these improvements. Mice receiving GW9662 exhibited aggravated histological injury comparable to that observed in the DSS group (Figure 6E). Collectively, these results suggest that the therapeutic effects of OA against DSS-induced colitis are at least partially dependent on the activation of PPARγ, and that pharmacological inhibition of PPARγ negates the beneficial actions of OA in this context.
 |
Figure 6 GW9662 eliminates the inhibitory effects of OA on DSS-induced ulcerative colitis symptoms in mice. (A) Schematic illustration of the experimental design. Mice were divided into four groups: Control, DSS, DSS+OA50 (OA at 50 mg/kg/day), and DSS+OA50+GW9662 (OA at 50 mg/kg/day and intraperitoneal GW9662 at 1 mg/kg/day). (B) Quantitative measurements of colon length in mice at the end of the experiment. (C) Daily body weight changes throughout DSS treatment. (D) The DAI scores recorded during the experiment. (E) Representative H&E-stained sections of colonic tissues and histopathological score statistics of the colon in each group. All data were presented as mean ± SD, n =6 per group. *P < 0.05, **P < 0.01, ***P < 0.001 vs corresponding control. |
GW9662 Abolishes the Protective Effect of OA Against DSS-Induced Inflammation and Intestinal Barrier Damage in Mice
Consistent with previous findings, OA administration markedly suppressed the DSS-induced upregulation of p-p65, p-IKKα/β, and iNOS protein expression in colonic tissues, while restoring the decreased IκBα levels. Notably, co-treatment with GW9662 reversed these effects, leading to enhanced activation of the NF-κB signaling pathway (Figure 7A and B). Meanwhile, qPCR analysis revealed that OA supplementation significantly downregulated the mRNA expression of Il-1β, Il-6, and Tnf-α in the colon, while markedly upregulating the mRNA levels of Il-4 and Il-10. These results suggest that OA ameliorates DSS-induced colitis by modulating inflammatory responses. However, the beneficial anti-inflammatory effects of OA were abolished upon administration of GW9662 (Supplementary Figure 1B), supporting the notion that OA exerts anti-inflammatory effects through PPARγ activation. Moreover, immunohistochemical analysis revealed that OA treatment significantly elevated the expression of TJ proteins ZO-1, Occludin, and Claudin-1, indicating improved intestinal barrier integrity. However, the beneficial effects of OA on TJ protein expression were abolished by GW9662 administration, further confirming that PPARγ plays a crucial role in mediating the barrier-protective effects of OA in DSS-induced colitis (Figure 7C).
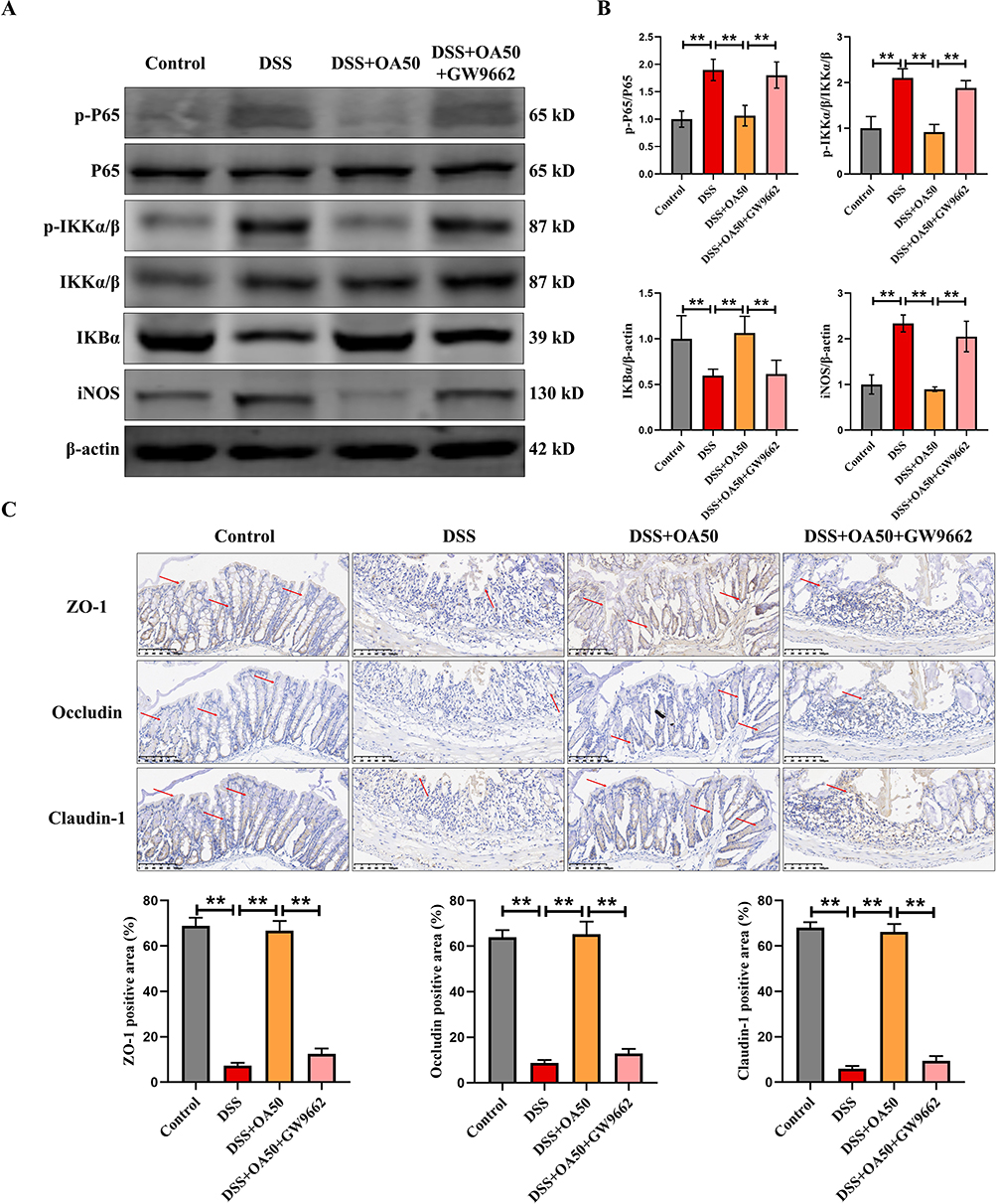 |
Figure 7 GW9662 abolishes the protective effect of OA against DSS-induced inflammation and intestinal barrier damage in mice. (A) The expression levels of colonic P65, p-P65, p-IKKα/β, IKKα/β, IKBα and iNOS were evaluated via Western-blot. (B) P65, p-P65, p-IKKα/β, IKKα/β, IKBα and iNOS were quantified using ImageJ software to provide a semiquantitative assessment of protein expression levels. (C) Representative IHC images of ZO-1, Occludin, and Claudin-1 in colonic tissues. Semiquantitative analysis of ZO-1, Occludin, and Claudin-1 using ImageJ. β-actin was used as a loading control. All data were presented as mean ± SD, n =6 per group. **P < 0.01 vs corresponding control. |
Discussion
In this study, OA administration (25 and 50 mg/kg/day) significantly alleviated DSS-induced acute UC in mice, as reflected by improvements in clinical parameters, reductions in DAI scores, and amelioration of histological damage. Mechanistically, OA protected against DSS-induced disruption of the intestinal barrier and aberrant immune responses, primarily through upregulation of PPARγ. These findings not only demonstrate the therapeutic potential of OA as a natural compound for UC management (Figure 8), but also provide novel insights into the molecular pathways through which OA exerts protective effects in colitis.
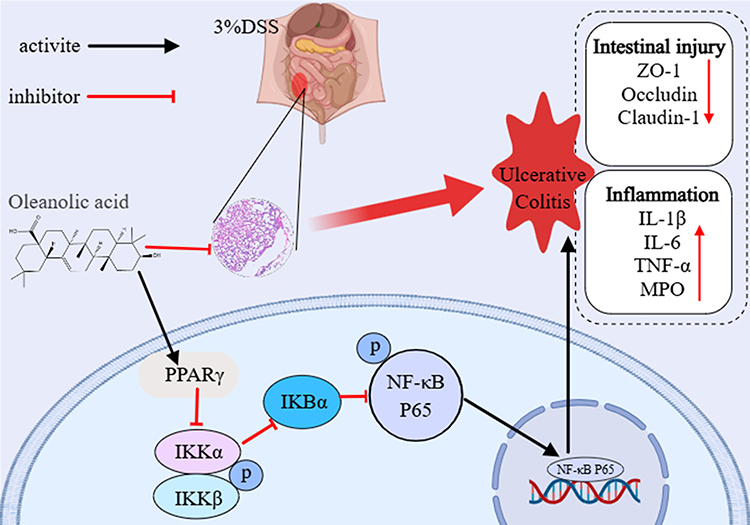 |
Figure 8 Schematic of proposed mechanism. |
Consistent with previous reports,32 DSS-treated mice exhibited classical colitis symptoms, including significant weight loss, increased DAI scores, shortened colon length, and pronounced histological damage. OA intervention, initiated 3 days prior to DSS exposure and continued for 10 days, significantly alleviated these phenotypes. At doses of 25 and 50 mg/kg/day,33 OA mitigated epithelial cell apoptosis, promoted mucosal regeneration, and restored the integrity of the intestinal barrier. DSS exposure compromised epithelial TJ integrity, downregulating ZO-1, Occludin, and Claudin-1. OA administration restored protein and mRNA expression of these TJ components, suggesting a robust protective effect on the epithelial barrier. These results underscore the critical role of epithelial integrity in UC pathogenesis and highlight OA’s ability to preserve barrier function under inflammatory stress, which may prevent further immune cell infiltration and tissue damage.
UC is characterized by dysregulated immune responses, with excessive pro-inflammatory cytokines (TNF-α, IL-1β, IL-6) promoting tissue injury, and reduced anti-inflammatory cytokines (IL-4, IL-10) failing to restrain inflammation.34–37 In our study, OA significantly suppressed neutrophil infiltration, as indicated by reduced MPO levels, and restored the balance of colonic and systemic cytokines. The mRNA expression of Tnf-α, Il-1β, and Il-6 was downregulated, whereas Il-4 and Il-10 were upregulated following OA treatment. These coordinated effects suggest that OA acts as an immunomodulatory agent, limiting excessive inflammatory cascades while promoting resolution of inflammation. Moreover, IL-1β-mediated ROS production contributes to smooth muscle dysfunction and diarrhea,38 implying that OA’s anti-inflammatory effects may indirectly preserve gut motility and function. Excessive inflammatory responses exacerbate oxidative stress, which is a critical driver of epithelial injury in UC.39,40 DSS exposure impaired antioxidant defenses, as evidenced by reduced SOD, CAT, and GSH-Px activities, decreased GSH content, and an imbalanced GSH/GSSG ratio, accompanied by elevated lipid peroxidation (MDA). OA supplementation effectively restored antioxidant enzyme activities and redox balance, highlighting its dual role in suppressing inflammation and counteracting oxidative damage. This interplay between anti-inflammatory and antioxidant effects suggests that OA can interrupt the self-amplifying cycle of inflammation–oxidative stress–barrier disruption in UC.
Activation of NF-κB is a central driver of inflammatory cytokine production and plays a critical role in the pathogenesis of UC.41,42 In colitis, NF-κB activation is primarily localized in macrophages and epithelial cells43,44 and can induce intestinal crypt apoptosis, leading to mucosal erosion.45 The p65 subunit of NF-κB is pivotal for the transcription of multiple inflammatory mediators,42,46 including iNOS, a hallmark of inflammation whose upregulation disrupts intestinal homeostasis and promotes dysbiosis.47,48 In our study, DSS exposure significantly increased p65 phosphorylation and iNOS protein expression in colonic tissues, whereas OA administration markedly attenuated these changes, indicating that OA mitigates colonic inflammation, at least in part, through suppression of NF-κB signaling.49,50 To further clarify the upstream mechanisms, we examined PPARγ, a nuclear receptor known to negatively regulate NF-κB and modulate inflammatory responses.51,52 Consistent with previous reports, DSS treatment significantly reduced PPARγ expression in the colon, whereas OA restored its protein levels. Notably, the use of GW9662, a PPARγ antagonist, largely abolished the protective effects of OA, as reflected by worsened clinical parameters, increased p65 phosphorylation and iNOS expression, and reduced TJ protein levels. These results suggest that OA exerts its anti-inflammatory and barrier-protective effects through PPARγ activation. Mechanistically, OA may alleviate intestinal inflammation via suppression of NF-κB signaling while maintaining mucosal integrity through upregulation of TJ proteins such as ZO-1, Occludin, and Claudin-1, thereby reinforcing barrier function and further limiting inflammatory responses.
Conclusion
In summary, our results demonstrated that OA significantly improved DSS-induced acute colitis by inhibiting inflammatory responses, alleviating intestinal oxidative stress, and ameliorating intestinal barrier function. Furthermore, PPARγ is a potential target for the improvement of UC in mice through OA intervention. This study indicates that OA may be a promising and natural strategy for prevention and improvement of UC.
Abbreviations
OA, Oleanolic acid; UC, ulcerative colitis; DSS, dextran sulfate sodium; DAI, disease activity index; PPARγ, peroxisome proliferator-activated receptor gamma; IBD, inflammatory bowel disease; NF-κB, Nuclear factor-kB; IL-1β, interleukin-1β; IL-6, interleukin-6; TNF-α, tumor necrosis factor-alpha; H&E, hematoxylin and eosin.
Acknowledgments
This work was supported by The Zhejiang Provincial Medical and Health Science and Technology Plan (2024KY1711), Shaoxing Bureau of Science and Technology (2023A14007, 2024A14025), China Postdoctoral Science Foundation funded project (2024M762958), research Project of Zhejiang Chinese Medical University (2024RCZXZK12), the medical and health research project of Zhejiang province (2025KY1135).
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Disclosure
The author(s) report no conflicts of interest in this work.
References
1. Ordas I, Eckmann L, Talamini M, Baumgart DC, Sandborn WJ. Ulcerative colitis. Lancet. 2012;380:1606–1619. doi:10.1016/S0140-6736(12)60150-0
2. Ungaro R, Mehandru S, Allen PB, Peyrin-Biroulet L, Colombel JF. Ulcerative colitis. Lancet. 2017;389:1756–1770. doi:10.1016/S0140-6736(16)32126-2
3. Shan Y, Lee M, Chang EB. The gut microbiome and inflammatory bowel diseases. Annu Rev Med. 2022;73:455–468. doi:10.1146/annurev-med-042320-021020
4. Wang Y, Zhang J, Zhang B, et al. Modified Gegen Qinlian decoction ameliorated ulcerative colitis by attenuating inflammation and oxidative stress and enhancing intestinal barrier function in vivo and in vitro. J Ethnopharmacol. 2023;313:116538. doi:10.1016/j.jep.2023.116538
5. Suzuki T. Regulation of the intestinal barrier by nutrients: the role of tight junctions. Anim Sci J. 2020;91:e13357. doi:10.1111/asj.13357
6. Wan Y, Yang L, Jiang S, Qian D, Duan J. Excessive apoptosis in ulcerative colitis: crosstalk between apoptosis, ROS, ER stress, and intestinal homeostasis. Inflamm Bowel Dis. 2022;28:639–648. doi:10.1093/ibd/izab277
7. Sahoo DK, Heilmann RM, Paital B, et al. Oxidative stress, hormones, and effects of natural antioxidants on intestinal inflammation in inflammatory bowel disease. Front Endocrinol. 2023;14:1217165.
8. Muro P, Zhang L, Li S, et al. The emerging role of oxidative stress in inflammatory bowel disease. Front Endocrinol. 2024;15:1390351. doi:10.3389/fendo.2024.1390351
9. Mo J, Ni J, Zhang M, et al. Mulberry anthocyanins ameliorate DSS-induced ulcerative colitis by improving intestinal barrier function and modulating gut microbiota. Antioxidants. 2022;11(9):1674. doi:10.3390/antiox11091674
10. Xu Y, Ou J, Zhang C, et al. Rapamycin promotes the intestinal barrier repair in ulcerative colitis via the mTOR/PBLD/AMOT signaling pathway. Biochim Biophys Acta Mol Basis Dis. 2024;1870:167287. doi:10.1016/j.bbadis.2024.167287
11. Papoutsopoulou S, Campbell BJ. Epigenetic modifications of the nuclear factor kappa B signalling pathway and its impact on inflammatory bowel disease. Curr Pharm Des. 2021;27:3702–3713. doi:10.2174/1381612827666210218141847
12. Grabinger T, Bode KJ, Demgenski J, et al. Inhibitor of apoptosis protein-1 regulates tumor necrosis factor-mediated destruction of intestinal epithelial cells. Gastroenterology. 2017;152:867–879. doi:10.1053/j.gastro.2016.11.019
13. Souza RF, Caetano MAF, Magalhaes HIR, Castelucci P. Study of tumor necrosis factor receptor in the inflammatory bowel disease. World J Gastroenterol. 2023;29:2733–2746. doi:10.3748/wjg.v29.i18.2733
14. Nguyen VQ, Eden K, Morrison HA, et al. Noncanonical NF-κB signaling upregulation in inflammatory bowel disease patients is associated with loss of response to anti-TNF agents. Front Pharmacol. 2021;12:655887. doi:10.3389/fphar.2021.655887
15. Lin Y, Wu Y, Su J, et al. Therapeutic role of d-pinitol on experimental colitis via activating Nrf2/ARE and PPAR-gamma/NF-κB signaling pathways. Food Funct. 2021;12:2554–2568. doi:10.1039/D0FO03139A
16. Venkataraman B, Almarzooqi S, Raj V, et al. alpha-bisabolol mitigates colon inflammation by stimulating Colon PPAR-gamma transcription factor: in vivo and in vitro study. PPAR Res. 2022;2022:5498115. doi:10.1155/2022/5498115
17. Peng Y, Yan Y, Wan P, et al. Gut microbiota modulation and anti-inflammatory properties of anthocyanins from the fruits of Lycium ruthenicum Murray in dextran sodium sulfate-induced colitis in mice. Free Radic Biol Med. 2019;136:96–108. doi:10.1016/j.freeradbiomed.2019.04.005
18. Mowat C, Cole A, Windsor A, et al. Gastroenterology, guidelines for the management of inflammatory bowel disease in adults. Gut. 2011;60:571–607. doi:10.1136/gut.2010.224154
19. Castellano JM, Ramos-Romero S, Perona JS. Oleanolic acid: extraction, characterization and biological activity. Nutrients. 2022;14:623. doi:10.3390/nu14030623
20. Gamede M, Mabuza L, Ngubane P, Khathi A. Plant-derived oleanolic acid ameliorates markers of subclinical inflammation and innate immunity activation in diet-induced pre-diabetic rats. Ther Adv Endocrinol Metab. 2020;11:2042018820935771. doi:10.1177/2042018820935771
21. Vrdoljak J, Kumric M, Vilovic M, et al. Effects of olive oil and its components on intestinal inflammation and inflammatory bowel disease. Nutrients. 2022;14:757. doi:10.3390/nu14040757
22. Iskender H, Dokumacioglu E, Kapakin KAT, et al. Effects of oleanolic acid on inflammation and metabolism in diabetic rats. Biotech Histochem. 2022;97:269–276. doi:10.1080/10520295.2021.1954691
23. Xue C, Jia H, Cao R, et al. Oleanolic acid improved intestinal immune function by activating and potentiating bile acids receptor signaling in E. coli-challenged piglets. J Anim Sci Biotechnol. 2024;15:79. doi:10.1186/s40104-024-01037-0
24. Kang GD, Lim S, Kim DH. Oleanolic acid ameliorates dextran sodium sulfate-induced colitis in mice by restoring the balance of Th17/Treg cells and inhibiting NF-κB signaling pathway. Int Immunopharmacol. 2015;29:393–400. doi:10.1016/j.intimp.2015.10.024
25. Yuan Z, Yang L, Zhang X, Ji P, Hua Y, Wei Y. Huang-lian-jie-du decoction ameliorates acute ulcerative colitis in mice via regulating NF-κB and Nrf2 signaling pathways and enhancing intestinal barrier function. Front Pharmacol. 2019;10:1354. doi:10.3389/fphar.2019.01354
26. Liu K, Yin Y, Shi C, et al. Asiaticoside ameliorates DSS-induced colitis in mice by inhibiting inflammatory response, protecting intestinal barrier and regulating intestinal microecology. Phytother Res. 2024;38:2023–2040. doi:10.1002/ptr.8129
27. Jumper J, Evans R, Pritzel A, et al. Highly accurate protein structure prediction with AlphaFold. Nature. 2021;596:583–589. doi:10.1038/s41586-021-03819-2
28. Varadi M, Anyango S, Deshpande M, et al. AlphaFold protein structure database: massively expanding the structural coverage of protein-sequence space with high-accuracy models. Nucleic Acids Res. 2022;50:D439–D444. doi:10.1093/nar/gkab1061
29. Kong Y, Gao Y, Lan D, et al. Trans-repression of NFkappaB pathway mediated by PPARgamma improves vascular endothelium insulin resistance. J Cell Mol Med. 2019;23:216–226. doi:10.1111/jcmm.13913
30. Li JJ, Wang R, Lama R, et al. Ubiquitin ligase NEDD4 regulates PPARgamma stability and adipocyte differentiation in 3T3-L1 cells. Sci Rep. 2016;6:38550. doi:10.1038/srep38550
31. Kim JH, Park KW, Lee EW, et al. Suppression of PPARgamma through MKRN1-mediated ubiquitination and degradation prevents adipocyte differentiation. Cell Death Differ. 2014;21:594–603. doi:10.1038/cdd.2013.181
32. Jin C, Liu J, Jin R, et al. Linarin ameliorates dextran sulfate sodium-induced colitis in C57BL/6J mice via the improvement of intestinal barrier, suppression of inflammatory responses and modulation of gut microbiota. Food Funct. 2022;13:10574–10586. doi:10.1039/D2FO02128E
33. Zhai Z, Liu Y, Niu K, et al. Oleanolic acid alleviate intestinal inflammation by inhibiting Takeda G-coupled protein receptor (TGR) 5 mediated cell apoptosis. Food Funct. 2024;15:1963–1976. doi:10.1039/D3FO04882A
34. Zhang Z, Cui Y, Liu S, et al. Short-term treatment with zingerone ameliorates dextran sulfate sodium-induced mouse experimental colitis. J Sci Food Agric. 2022;102:4873–4882. doi:10.1002/jsfa.11850
35. Scuruchi M, D’Ascola A, Avenoso A, Mandraffino G, Campo S, Campo GM. Endocan, a novel inflammatory marker, is upregulated in human chondrocytes stimulated with IL-1 beta. Mol Cell Biochem. 2021;476:1589–1597. doi:10.1007/s11010-020-04001-4
36. Ilchovska DD, Barrow DM. An Overview of the NF-kB mechanism of pathophysiology in rheumatoid arthritis, investigation of the NF-kB ligand RANKL and related nutritional interventions. Autoimmun Rev. 2021;20:102741. doi:10.1016/j.autrev.2020.102741
37. Wu P, Guo Y, Jia F, Wang X. The effects of armillarisin A on serum IL-1beta and IL-4 and in treating ulcerative colitis. Cell Biochem Biophys. 2015;72:103–106. doi:10.1007/s12013-014-0413-7
38. Cao W, Vrees MD, Potenti FM, Harnett KM, Fiocchi C, Pricolo VE. Interleukin 1beta-induced production of H2O2 contributes to reduced sigmoid colonic circular smooth muscle contractility in ulcerative colitis. J Pharmacol Exp Ther. 2004;311:60–70. doi:10.1124/jpet.104.068023
39. Wang R, Luo Y, Lu Y, et al. Maggot extracts alleviate inflammation and oxidative stress in acute experimental colitis via the activation of Nrf2. Oxid Med Cell Longev. 2019;2019:4703253. doi:10.1155/2019/4703253
40. Lv T, Shen L, Yang L, et al. Polydatin ameliorates dextran sulfate sodium-induced colitis by decreasing oxidative stress and apoptosis partially via Sonic hedgehog signaling pathway. Int Immunopharmacol. 2018;64:256–263. doi:10.1016/j.intimp.2018.09.009
41. Thompson WL, Van Eldik LJ. Inflammatory cytokines stimulate the chemokines CCL2/MCP-1 and CCL7/MCP-3 through NFkB and MAPK dependent pathways in rat astrocytes [corrected. Brain Res. 2009;1287:47–57. doi:10.1016/j.brainres.2009.06.081
42. Li Z, Zhang DK, Yi WQ, Ouyang Q, Chen YQ, Gan HT. NF-kappaB p65 antisense oligonucleotides may serve as a novel molecular approach for the treatment of patients with ulcerative colitis. Arch Med Res. 2008;39:729–734. doi:10.1016/j.arcmed.2008.08.001
43. Schreiber S, Nikolaus S, Hampe J. Activation of nuclear factor κB inflammatory bowel disease. Gut. 1998;42:477–484. doi:10.1136/gut.42.4.477
44. Rogler G, Brand K, Vogl D, et al. Nuclear factor κB is activated in macrophages and epithelial cells of inflamed intestinal mucosa. Gastroenterology. 1998;115:357–369. doi:10.1016/S0016-5085(98)70202-1
45. Wong J, Garcia-Carbonell R, Zelic M, et al. RIPK1 mediates TNF-induced intestinal crypt apoptosis during chronic NF-κB activation. Cell Mol Gastroenterol Hepatol. 2020;9:295–312. doi:10.1016/j.jcmgh.2019.10.002
46. Cui L, Feng L, Zhang ZH, Jia XB. The anti-inflammation effect of baicalin on experimental colitis through inhibiting TLR4/NF-κB pathway activation. Int Immunopharmacol. 2014;23:294–303. doi:10.1016/j.intimp.2014.09.005
47. El-Ashmawy NE, Khedr NF, El-Bahrawy HA, El-Adawy SA. Roflumilast, type 4 phosphodiesterase inhibitor, attenuates inflammation in rats with ulcerative colitis via down-regulation of iNOS and elevation of cAMP. Int Immunopharmacol. 2018;56:36–42. doi:10.1016/j.intimp.2018.01.004
48. Luo S, He J, Huang S, et al. Emodin targeting the colonic metabolism via PPARgamma alleviates UC by inhibiting facultative anaerobe. Phytomedicine. 2022;104:154106. doi:10.1016/j.phymed.2022.154106
49. Zhang H, Zhuo S, Song D, et al. Icariin inhibits intestinal inflammation of DSS-induced colitis mice through modulating intestinal flora abundance and modulating p-p65/p65 molecule. Turk J Gastroenterol. 2021;32:382–392. doi:10.5152/tjg.2021.20282
50. Schreiber O, Petersson J, Walden T, et al. iNOS-dependent increase in colonic mucus thickness in DSS-colitic rats. PLoS One. 2013;8:e71843. doi:10.1371/journal.pone.0071843
51. Dubuquoy L, Dharancy S, Nutten S, Pettersson S, Auwerx J, Desreumaux P. Role of peroxisome proliferator-activated receptor gamma and retinoid X receptor heterodimer in hepatogastroenterological diseases. Lancet. 2002;360:1410–1418. doi:10.1016/S0140-6736(02)11395-X
52. Han Y, Liu J, Ahn S, et al. Diallyl Biphenyl-Type Neolignans Have a Pharmacophore of PPARalpha/gamma Dual Modulators. Biomol Ther. 2020;28:397–404. doi:10.4062/biomolther.2019.180
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Quzhou Aurantii Fructus Flavonoids Ameliorate Inflammatory Responses, Intestinal Barrier Dysfunction in DSS-Induced Colitis by Modulating PI3K/AKT Signaling Pathway and Gut Microbiome

Wang H, Huang W, Pan X, Tian M, Chen J, Liu X, Li Q, Qi J, Ye Y, Gao L
Journal of Inflammation Research 2025, 18:1855-1874
Published Date: 6 February 2025
NLRP3 Mediates NOXs-Induced Iron Overload and Inflammation but Not Oxidative Damage in Colons of DSS-Treated Mice
Yu L, Wang M, Zhou Y, Qi J, Zheng Q, Song Z
Journal of Inflammation Research 2025, 18:4695-4708
Published Date: 4 April 2025
Elevated Serum Podoplanin Levels Correlate with Disease Severity and Inflammation in Ulcerative Colitis: A Cross-Sectional Study
Huang X, Tang C
Journal of Inflammation Research 2025, 18:12989-13000
Published Date: 18 September 2025