Back to Journals » Journal of Asthma and Allergy » Volume 18
PP2A Promotes Epithelial-Mesenchymal Transition in Chronic Rhinosinusitis with Nasal Polyps via Wnt/β-Catenin Pathway: A Novel Insight
Authors Chen W, Zheng B, Shu L, Liu Y, Lu T, Shen Y ![]() , Yang Y
, Yang Y
Received 9 June 2025
Accepted for publication 22 October 2025
Published 4 November 2025 Volume 2025:18 Pages 1505—1522
DOI https://doi.org/10.2147/JAA.S542695
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Luis Garcia-Marcos
Wei Chen,* Bowen Zheng,* Longlan Shu, Yijun Liu, Tao Lu, Yang Shen, Yucheng Yang
Department of Otolaryngology, The First Affiliated Hospital of Chongqing Medical University, Chongqing, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Yucheng Yang, Department of Otolaryngology, The First Affiliated Hospital of Chongqing Medical University, Chongqing, People’s Republic of China, Email [email protected] Yang Shen, Department of Otolaryngology, The First Affiliated Hospital of Chongqing Medical University, Chongqing, People’s Republic of China, Email [email protected]
Purpose: Chronic rhinosinusitis with nasal polyps (CRSwNP) is a prevalent upper respiratory disease with eosinophilic infiltration. Epithelial-to-mesenchymal transition (EMT) has been considered an important pathological mechanism of CRSwNP. The Wnt signaling pathway is a well-established promoter of EMT, while Protein Phosphatase 2A (PP2A) exerts dual regulatory effects on Wnt pathway. However, the contribution of PP2A to CRSwNP has not been reported. This research aimed to explore the possible mechanisms by which PP2A modulates EMT in CRSwNP.
Patients and Methods: About 56 patients with CRSwNP and 20 control subjects were enrolled in this study. We collected nasal polyps (NPs) and inferior turbinate tissues; the expression of PP2A, EMT markers, and Wnt signaling-related mediators in tissues were analyzed by Western blotting, immunohistochemistry, and quantitative RT-PCR. Primary cells isolated from NPs were cultured with PP2A inhibitor LB-100. The rhWNT3A-induced EMT cell model using BEAS-2B cell line was established in vitro and treated with either LB-100 or the PP2A agonist DT-061. Murine NP model was developed in vivo and treated with LB-100. The expression changes of EMT markers and Wnt signaling mediators were detected using Western blotting, immunofluorescence staining, and hematoxylin-eosin staining to estimate the effect of PP2A in the pathogenesis of CRSwNP.
Results: Compared to controls, NPs exhibited increased PP2A expression and activity, activation of Wnt signaling, and evidence of EMT. These changes were more pronounced in eosinophilic NPs. PP2A inhibition alleviated, whereas PP2A activation promoted, EMT in epithelial cells by regulating Wnt pathway. PP2A inhibition could also suppress the formation of NPs and alleviate eosinophilic inflammation in the murine NP models.
Conclusion: PP2A promotes EMT through the activation of Wnt/β-catenin signaling pathway, leading to epithelial barrier dysfunction in NPs. PP2A exerts a positive regulatory effect on the modulation of Wnt signaling in CRSwNP. Targeting PP2A might represent a viable therapeutic option for treating CRSwNP.
Keywords: chronic rhinosinusitis, protein phosphatase 2A, epithelial-mesenchymal transition, nasal polyps, wnt
Graphical Abstract:

Introduction
Chronic rhinosinusitis with nasal polyps (CRSwNP) represents a common upper respiratory disease, characterized by persistent inflammation and nasal polyps (NPs) development,1 accounting for about 30% of chronic rhinosinusitis (CRS) patients.2 Common symptoms of CRSwNP include blocked nasal passages, facial pain, nasal discharge, and a diminished or lost sense of smell, leading to a reduced quality of life. Despite undergoing surgery and glucocorticoid therapy, a considerable number of patients experience poor therapeutic effects, due to the heterogeneity and indeterminacy of pathogenesis. Recent studies have demonstrated that dysfunctional epithelial barrier is linked with CRSwNP pathogenesis;3,4 the impairment of the nasal mucosal epithelial barrier leads to inflammatory cell infiltration and subsequent tissue remodeling.5 Epithelial-mesenchymal transition (EMT) is a well-documented process involved in tissue remodeling,6 where the normal epithelial adhesion junctions and polarities of epithelial cells are destroyed with gaining of mesenchymal phenotypes.7 The EMT process has been reported to play a crucial role in the development of CRS.8–10 Furthermore, EMT-induced epithelial dysfunction is believed to contribute to treatment resistance and recurrence in CRSwNP, while the exact mechanism that initiates EMT in CRSwNP is still unclear.
EMT could be regulated by several signaling pathways, one of which is the Wnt/β-catenin signaling pathway.11 Normal Wnt signaling is recognized as essential for embryonic development and maintenance of adult tissue,12 while abnormal Wnt signaling is associated with cancer development. A recent study has indicated that Wnt signaling is activated in CRSwNP,13 but the concrete regulation mechanism of this signaling pathway in NPs is still poorly understood.
Protein Phosphatase 2A (PP2A), a serine/threonine phosphoprotein phosphatase, is involved in modulating Wnt/β-catenin signaling pathway, exerting both stimulatory and inhibitory effects on this pathway.14 Early studies indicated that PP2A activity is linked to inflammation of the lower respiratory tract,15,16 but its involvement in upper respiratory tract inflammation is still unknown. So far, it remains unreported whether PP2A contributes to the pathogenesis of CRSwNP through Wnt pathway.
The study aimed to explore: (1) PP2A expression in normal sinonasal mucosa and its alteration in NPs; (2) whether the Wnt/β-catenin signaling pathway is involved in the development of CRSwNP through promoting EMT; (3) whether PP2A affects EMT process by regulating Wnt pathway and contributes to pathophysiology of CRSwNP; (4) whether there are differences in the expression of PP2A and Wnt signaling-associated mediators between different endotypes of CRSwNP. The experimental workflow is schematically illustrated in Figure 1.
 |
Figure 1 Schematic illustration of the research strategy to investigate the role of PP2A in promoting EMT in CRSwNP via regulating the Wnt/β-catenin pathway. The study was designed to validate the proposed mechanism across three levels. Part 1: The expression and activity levels of PP2A, key components of the Wnt/β-catenin pathway, and EMT markers were analyzed in nasal polyp tissues and inferior turbinate tissues. Part 2: The role of PP2A was investigated using primary nasal polyp epithelial cells and BEAS-2B cells through function experiments. Part 3: A murine model of eosinophilic nasal polyps was established and was intervened with a PP2A inhibitor. The effects of PP2A on EMT markers and eosinophil infiltration were evaluated. |
Materials and Methods
Study Subjects
This study included 20 control subjects, 20 eosinophilic CRSwNP (EosCRSwNP) patients, and 36 non-eosinophilic CRSwNP (non-EosCRSwNP) patients. The diagnosis of CRSwNP was made in accordance with EPOS 2020.1 Eosinophilic CRSwNP was determined on the basis of the histological quantification of eosinophils, with a count of 55 or more in a high-power field.17 The control subjects were diagnosed as having deviation of the nasal septum with hypertrophic inferior turbinate and without any sinonasal inflammatory diseases. Subjects with these conditions were excluded: (1) those under the age of 18; (2) those with a diagnosis of primary ciliary dyskinesia; (3) those who had been administered antibiotics, corticosteroids, or other immunomodulatory medications within 4 weeks preceding surgery. Informed consent was obtained from all the patients. During surgery, inferior turbinate tissues from control participants and nasal polyp tissues from CRSwNP patients were obtained. More information is provided in the supplementary material 1.
Our study complies with the Declaration of Helsinki. Approval for this study was granted by the ethics committee of The First Affiliated Hospital of Chongqing Medical University (2022-K301,17 June 2022). Due to constraints in sample availability, not all specimens were utilized in each experimental protocol. Table 1 displays the demographic information.
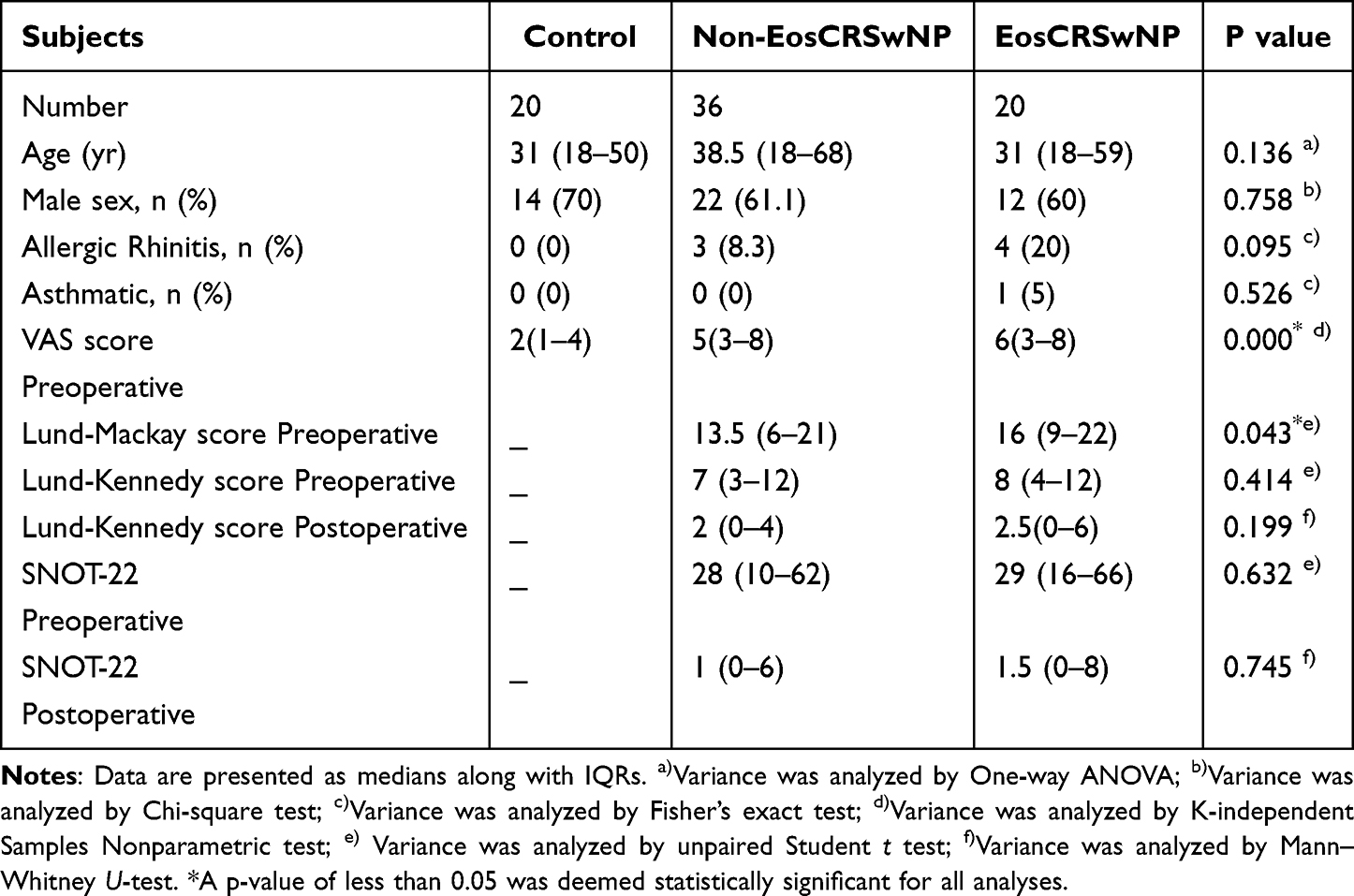 |
Table 1 Demographics of Subjects |
Quantitative Real-Time PCR
Total RNA was isolated using the Trizol reagent method, then cDNA was synthesized utilizing a reverse transcription kit (More information is provided in the supplementary method 1). The primer sequences were designed and synthesized by Sangon Biotech (Table 2). Relative expression levels of mRNA for Wnt3A, WIF1, CTNNB1, PPP2CA, and GAPDH were assessed using RT-qPCR assays. The 2−ΔΔCt method was used to calculate gene expression, which was then normalized to GAPDH.
 |
Table 2 Primer Sequences for PCR Amplification |
Western Blot Analysis
Specimens were lysed using RIPA buffer and subsequently quantified using BCA assay reagent. Following their separation using SDS-PAGE, the proteins were transferred onto PVDF membranes, then blocked with 6% skim milk for a duration of 2 hours. The membranes underwent incubation with primary antibodies specific to PP2A, phospho-PP2A alpha (Tyr307), E-Cadherin, Vimentin, GSK3β, phospho-GSK3 β(Ser9), Wnt3A, Cyclin D1, β-Catenin, phospho-β-Catenin (Ser33/Ser37), β-Catenin non-phospho(active)S45, and GAPDH overnight at 4°C. The next day, the membranes underwent an incubation step with an HRP-labeled secondary antibody, and then visualized by the ECL system. Finally, the images were analyzed by Image J software. More information is provided in the supplementary method 2.
Immunohistochemistry
Tissue specimens were fixed, then dehydrated, embedded in paraffin, and sectioned into 5 - μm - thick slices. Antigen was retrieved by citric acid buffer, and endogenous peroxidase activity was inhibited. Sections were permeabilized, subsequently blocked and incubated with the primary antibodies specific to PP2A, E- Cadherin, GSK3β, Wnt3A, β-Catenin, and Vimentin overnight at 4°C. The sections were subjected to three 5-minute washes with PBST, then incubated with secondary antibody, and treated with DAB on the following day. Ultimately, slides were counterstained with Hematoxylin, thoroughly rinsed under running tap water, and sealed for endoscopic examination. The average optical density (AOD) was used to measure the level of expression in IHC using Image J software, while the expressions of nuclear β - catenin and wnt3A were measured by cell counting in the epithelial layer by three blinded observers. More information is provided in the supplementary method 3.
Immunofluorescence Staining
Cell climbing films were first fixed, then permeabilized, thoroughly washed with PBS, subsequently blocked, incubated with primary antibodies, as outlined in the immunohistochemistry protocol. On the next day, cell climbing films were incubated with secondary antibodies, followed by DAPI counterstaining, then examined by fluorescence microscope, and the data were analyzed by Image J software, with the assessment based on mean fluorescence intensity. More information is provided in the supplementary method 4.
Primary Cell Culture
Nasal polyps obtained from patients diagnosed with CRSwNP were used for primary culture. Fresh tissue samples were washed in PBS, sectioned into smaller fragments, and digested with 0.1% collagenase type I and 0.25% trypsin. Monolayer primary nasal polyp epithelial cells were cultured in complete medium. After reaching about 80% confluence, primary cells were treated with 4 µM LB-100 (a small molecule PP2A inhibitor, SF1082-10Mm, Beyotime, China), or not as control for 48 h. Collected cells were used for Western blot analysis. More information is provided in the supplementary method 5.
Recombinant Human WNT3A and PP2A Inhibitor Treatment of Human Bronchial Epithelial Cell
Human bronchial epithelial cell line BEAS-2B (Pricella, China) was preserved at −80°C and recovered from the frozen state. After 2 generations, the third generation was seeded in a 10 cm culture dish and cultured in complete medium as previously described. After reaching about 80% confluence, cells were stimulated with 400 ng/mL of rhWNT3A (MedChemExpress, USA) to establish a Wnt-induced EMT cell group. Additionally, a combination treatment of rhWNT3A (400 ng/mL) and LB-100 (4 µM) was administered to create a PP2A inhibition group. The control group was cultured with complete medium only. After a 48-h incubation period, the harvested cells were used for Western blot analysis and immunofluorescence assays. More information is provided in the supplementary materials method 6.
Recombinant Human WNT3A and PP2A Agonist Treatment of Human Bronchial Epithelial Cell
The control group and Wnt-induced EMT cell group were established as previously described. Subsequently, BEAS-2B cells were treated with rhWNT3A (400 ng/mL) and PP2A agonist DT-061 (8 µM, Selleck, USA) for 48 h to create a group with PP2A stimulation, collected cells were used for Western blot analysis and immunofluorescence assays. More information is provided in the supplementary materials method 7.
Establishment of Murine NP Model and PP2A Inhibition Treatment
Fifteen female BALB/c mice (aged 5 weeks,16 to 20g) were maintained in a standard housing environment. The animal study was conducted following the standard protocol illustrated in Figure 2. All animal experiments received approval from the Institutional Animal Care and Use Committee of Chongqing Medical University (IACUC-CQMU-2023-0249) and adhered to the principles on ethical animal research outlined in the Basel Declaration and the ethical guidelines by the International Council for Laboratory Animal Science. Mice were randomly assigned to three groups (n = 5):18,19 the control group underwent PBS administration only; NP model group received treatments of ovalbumin (OVA) and staphylococcal enterotoxins B(SEB); the intervention group were also treated with LB-100 in addition to OVA and SEB. Histological examination of the nasal mucosal epithelium was conducted using Hematoxylin-Eosin staining. The presence of polyp-like lesions and eosinophilic infiltration in nasal mucosa was observed under light microscope by three blinded observers independently. Polyp-like lesions were defined as obvious nasal mucosal eminences with eosinophilic infiltration. The quantification of eosinophilic infiltration was determined by calculating the number of eosinophils per high power field (HPF). For each histological section, a total of 10 non-overlapping high power fields were examined. Subsequently, the mean of the eosinophil counts was computed to represent the degree of eosinophilic infiltration. The counts of polyp-like lesions and infiltrated eosinophils were recorded. Changes in the expressions of E-cadherin and Vimentin in the nasal mucosal epithelium of mice were assessed by Immunohistochemistry describe previously. More information is provided in the supplementary materials method 8.
 |
Figure 2 Protocol of the murine model of NPs in BALB/c mice with PP2A inhibitor intervention. (A) Control group: mice were injected intraperitoneally with 0.3 mL PBS on day 0 and 5, then 40μL PBS was instilled daily by nasal infusion from day 12 to day 19, next mice were received 40μL PBS 3 times a week by nasal infusion for 12 weeks. (B) NP model group: Mice were sensitized by intraperitoneal injection of OVA 25μg and Alum gel 2 mg on day 0 and 5, then 40μL of 3% VOA was instilled daily by nasal infusion from day 12 to day 19, next mice were received 40μL of 3% VOA 3 times a week by nasal infusion for 4 weeks, subsequently, mice were dripped nasally with 40μL of 3% OVA and 10 ng SEB for 8 weeks. (C) Intervention group: Same protocol as NP model group, with additional mice were injected intraperitoneally with LB-100 (1.5 mg/kg) twice a week during the final 8 week. |
Statistical Analysis
The normality of the data was evaluated by the Kolmogorov–Smirnov test and the Shapiro–Wilk test. For binary comparisons, unpaired Student t test was applied for testing if variables correspond to normally distribution; otherwise, the Mann–Whitney U-test was utilized. In the event of multi-group comparisons, one-way ANOVA was conducted, followed by the Tukey post hoc test for variables with normal distribution, while the Kruskal–Wallis test was used for variables that were not normally distributed. The data were illustrated using scatter dot plots and bar graphs, where the bars indicated the median values and the error bars represented the standard errors of the mean (SEMs). P <0.05 indicates statistical significance. The analyses were conducted using SPSS version 25.0 and GraphPad Prism software version 10.0.
Results
Increased Expression and Activity of PP2A in Nasal Polyps
The PP2A mRNA expression was increased in both EosCRSwNP and non-EosCRSwNP compared to control tissues. Moreover, a significantly higher mRNA expression of PP2A was observed in EosCRSwNP compared to non-EosCRSwNP (Figure 3A). The protein-level validation of these findings was performed using Western blotting. Additionally, the relative expression of Phospho-PP2A alpha (Tyr307) at the protein level was significantly decreased in NPs (Figure 3B), indicating that the activity of PP2A was also increased in nasal polyps compared with control.
 |
Figure 3 Increased expression and activity of PP2A in nasal polyp tissues. (A) the mRNA expression levels of PP2A in tissue samples from different study groups detected by quantitative RT-PCR. (B) the protein levels of PP2A and Phospho-PP2A alpha (Tyr307) in tissue samples from different study groups detected by Western blotting. (C), Immunohistochemical intensity of PP2A in the epithelium of different groups (x 400 magnification). For (A–C) data were analyzed by the one-way ANOVA with the Tukey post hoc test. *P <0.05; **P <0.01; and ***P < 0.001. |
Then we performed immunohistochemical staining of tissue sections to compare the levels of PP2A expression in NPs and inferior turbinate tissues. Result showed that the average optical density of PP2A in nasal polyps was significantly elevated compared to that in inferior turbinate tissues (Figure 3C), which is consistent with the result from Western blot analysis.
EMT Process and Wnt Signaling Are Activated in Nasal Polyps
The mRNA transcriptional level of Wnt3A was upregulated, and the Wnt inhibitor WIF1 was downregulated in NPs compared to control. Meanwhile, the mRNA expression level of β-catenin was upregulated only in eosinophilic NPs (Figure 4A).
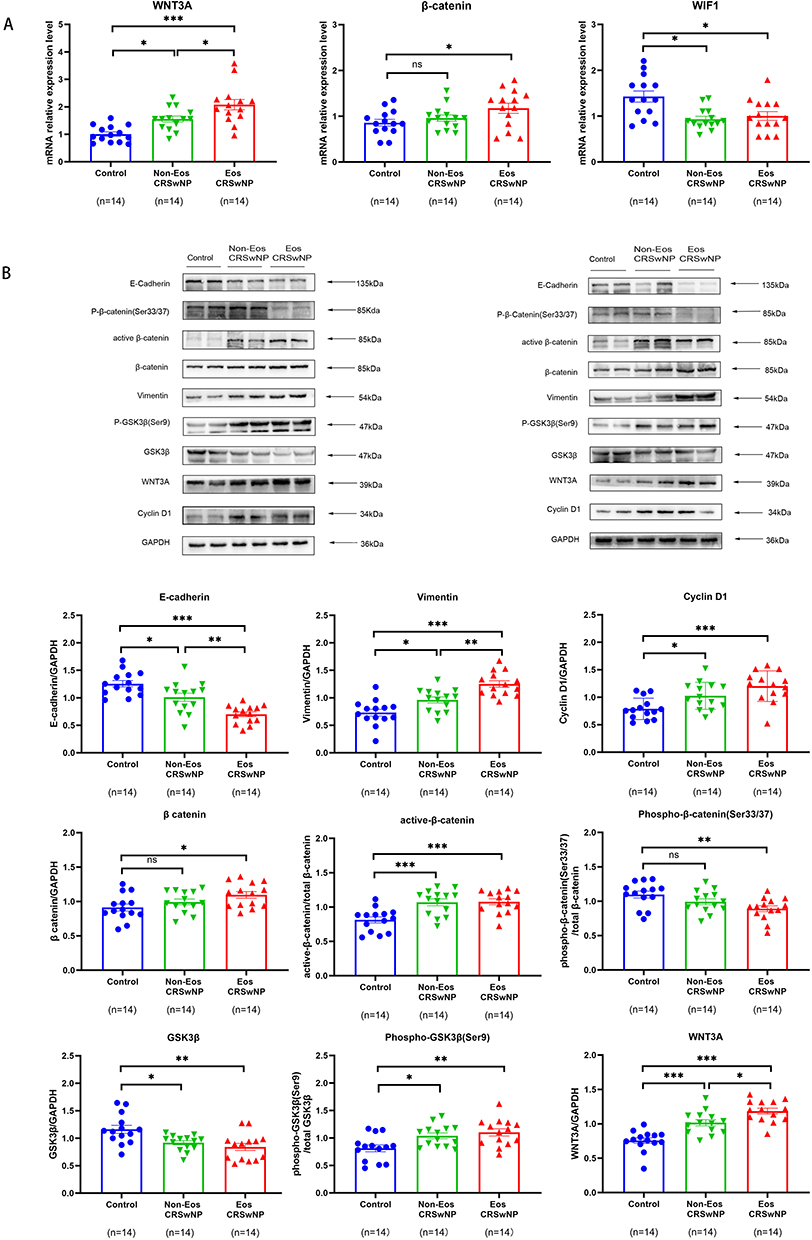 |
Figure 4 Wnt signaling and EMT are activated in nasal polyp tissues. (A) the mRNA expression levels of WNT3A, β-catenin and WIF1 in tissue samples from different study groups detected by quantitative RT-PCR. (B) the protein levels of E-cadherin, Vimentin, Cyclin D1, β-catenin, active β-catenin, Phospho-β-catenin (Ser33/37), GSK3β, Phospho- GSK3β (Ser9) and WNT3A in tissue samples from different study groups detected by Western blotting. For (A and B) Data were analyzed by the one-way ANOVA with the Tukey post hoc test. *P <0.05; **P <0.01; and ***P < 0.001. Abbreviation: ns, no significant difference. |
Western blot analysis was conducted to assess the expression of key regulator proteins of Wnt signaling pathway: Wnt3A, cyclin D1, β-catenin, and GSK3β. The outcomes indicated a reduction in the levels of GSK3β and phospho-β-catenin (Ser33/Ser37) expression, while the expression of Wnt3A, cyclin D1, phospho-GSK3β(Ser9), and active β-catenin was upregulated in nasal polyps in comparison to control. The level of total β-catenin expression was only increased in eosinophilic NPs (Figure 4B). The findings were additionally validated through conducting immunohistochemistry of nuclear β-catenin, WNT3A and GSK3β in tissue samples, and we found a notable elevation in the counts of nuclear β-catenin positive cells and WNT3A positive cells in the epithelium of nasal polyps (Figure 5E and F), while the average optical density of GSK3β in epithelium was lower in NPs compared with control (Figure 5D).
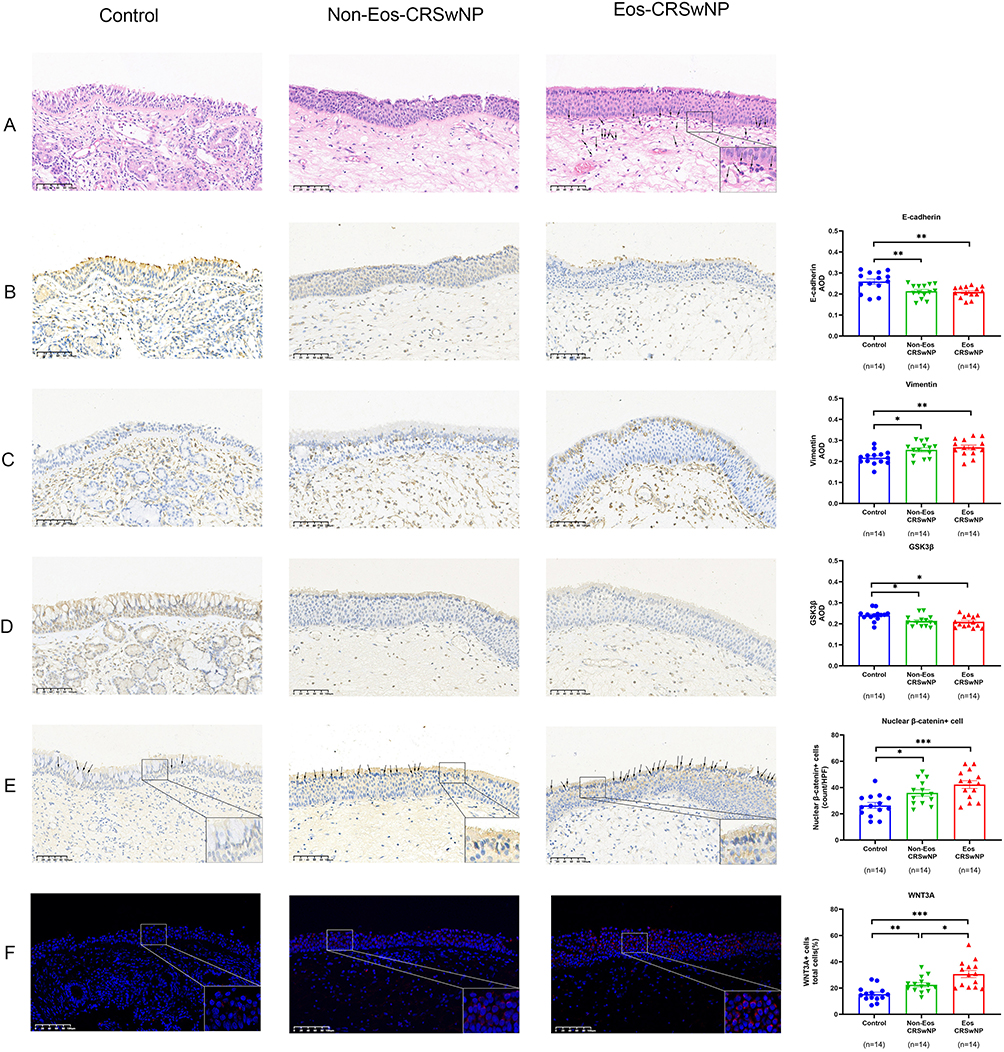 |
Figure 5 Wnt signaling and EMT are activated in nasal polyp tissues (x 200 magnification). (A) Eosinophilic infiltration in the different study groups by hematoxylin-eosin staining, →: eosinophils. (B–D), Immunohistochemical intensity of E-cadherin, Vimentin and GSK3β. (E) Nuclear β-catenin-positive cell numbers in the epithelium of different groups, →: nuclear β-catenin-positive cell. (F) Positive cell counts per total cells of WNT3A in the epithelium of different groups. Data were analyzed by the one-way ANOVA with the Tukey post hoc test. *P <0.05; **P <0.01; and ***P < 0.001. |
We also analyzed the expression levels of E-cadherin, a marker of epithelial tight junctions, as well as the mesenchymal marker Vimentin. We observed a significant increase in Vimentin expression and a decrease in E-cadherin expression in NPs compared to control by Western blot analysis (Figure 4B). Meanwhile, the same result was shown by immunohistochemical staining (Figure 5A–C).
Inhibition of PP2A Inhibits EMT in Epithelial Cells of NPs via Regulating Wnt/β-Catenin Signaling Pathway
To determine whether PP2A affected EMT process in nasal polyps, we cultured primary epithelial cells isolated from NPs, and the primary cells were treated with LB-100. Then we analyzed variations in E-cadherin, Vimentin, β-catenin and active β-catenin expression by Western blot. The outcomes revealed an elevation in the expression level of E-cadherin, whereas the expression of β-catenin, active β-catenin, and Vimentin exhibited a decrease in primary cells treated with LB-100 compared to the control group (Figure 6). To ensure that the decreased active β-catenin was not triggered by the decreased total β-catenin expression, active β-catenin level was normalized to total β-catenin, we could confirm that PP2A inhibitor contributed to downregulation of active β-catenin, leading to inhibition of this pathway. Outcomes from the experiment indicate that inhibition of PP2A may inhibit EMT process in epithelial cells of nasal polyps by suppressing Wnt/β-catenin signaling pathway.
 |
Figure 6 Inhibition of PP2A inhibits EMT in epithelial cells of nasal polyps via regulating the Wnt/β-catenin signaling pathway. Primary epithelial cells of nasal polyps from CRSwNP patients were treated with LB-100, or not as control in three independent biological replicates (labeled as Rep 1, Rep 2, Rep 3). The protein levels of E-cadherin, Vimentin, β-catenin, active β-catenin, PP2A and Phospho-PP2A alpha (Tyr307) in cells treated with LB-100 and control as detected by Western blotting. Data were analyzed by the unpaired Student t test. *P <0.05; **P <0.01; and ***P < 0.001; ns: no significant difference. |
We further explored the molecular mechanism by LB-100 alleviating EMT in epithelial cells of NPs. We examined the activity and expression of PP2A. The findings indicated that there was no expression change of total PP2A, while there was significant increased expression of Phospho-PP2A (Tyr307) in epithelial cells after LB-100 treatment (Figure 6). Therefore, we conclude that LB-100 works through inhibiting activity but not decreasing the expression of PP2A.
Inhibition of PP2A Suppresses EMT Induced by rhWNT3A in Epithelial Cells
BEAS-2B cells underwent treatment with rhWNT3A as Wnt-induced EMT cell model group or only were cultured with complete medium as control. Then we conducted Western blot to analyze the changes in the expression levels of EMT markers. The findings indicated a notable reduction in the expression of E-cadherin, accompanied by an elevated level of vimentin expression in Wnt-induced EMT group (Figure 7A and B), as well as the morphological change of BEAS-2B cells, from a polygonal shape to a spindle - shaped morphology (Figure 7C).
 |
Figure 7 Inhibition of PP2A inhibits EMT and stimulation of PP2A promotes EMT induced by rhWNT3A in BEAS-2B cells. BEAS-2B cells were treated with rhWNT3A and LB-100 as PP2A inhibition group, stimulated with rhWNT3A as Wnt-induced EMT group, cultured with rhWNT3A and DT-061 as PP2A stimulation group, or only cultured with complete medium as control. (A) inhibition of PP2A inhibits EMT induced by rhWNT3A in BEAS-2B cells. The protein levels of E-cadherin, Vimentin, β-catenin, and active β-catenin in cells from different group as detected by Western blotting in three independent biological replicates (labeled as Rep 1, Rep 2, Rep 3). (B) stimulation of PP2A promotes EMT induced by rhWNT3A in BEAS-2B cells, the protein levels of E-cadherin, Vimentin, β-catenin, and active β-catenin in cells from different group as detected by Western blotting in three independent biological replicates (labeled as Rep 1, Rep 2, Rep 3). (C) contrast morphological images of BEAS-2B cells treated with WNT3A for 48 h and control (x 40 magnification). Data were analyzed by the one-way ANOVA with the Tukey post hoc test. *P <0.05; **P <0.01; ***P < 0.001; ns: no significant difference. |
At the same time, BEAS-2B cells were treated with a combination of rhWNT3A and LB-100 to establish the PP2A inhibition group. Next, we performed immunofluorescence and Western blot to analyze changes in the expression of EMT markers and essential regulatory proteins of Wnt pathway. The data obtained from Western blotting indicated a reduction in the levels of total β-catenin, activated β-catenin, and vimentin expression, while E-cadherin expression was increased in PP2A inhibition group compared to Wnt-induced EMT cell model group (Figure 7A). Consistently, immunofluorescence images showed similar changes of EMT markers (Figure 8). The results indicate that inhibition of PP2A can suppress EMT induced by Wnt signaling in airway epithelial cells.
 |
Figure 8 Inhibition of PP2A suppresses EMT and stimulation of PP2A promotes EMT induced by rhWNT3A in Human bronchial epithelial cells. (A) Immunofluorescence image of BEAS-2B cells stained for E-cadherin, fluorescence intensity of BEAS-2B cells in different group were compared. (B) Immunofluorescence image of BEAS-2B cells stained for Vimentin, fluorescence intensity of BEAS-2B cells in different group were compared. Scale bar:20 µm. Data were analyzed by the one-way ANOVA with the Tukey post hoc test. *P <0.05; **P <0.01; and ***P < 0.001, ns: no significant difference. |
Activation of PP2A Promotes EMT Induced by rhWNT3A in Epithelial Cells
We further validated the effect of PP2A in the function of the epithelial barrier by treating cells with PP2A agonists DT-061. We examined the changes in the expression levels of β-catenin and EMT markers after stimulation of PP2A in BEAS 2B cells. The findings from Western blotting showed that E-cadherin expression was reduced, while the expression of activated β-catenin and Vimentin were increased in the PP2A-stimulated group compared to the Wnt-induced EMT group (Figure 7B). The outcomes from immunofluorescence analysis demonstrated that the mean fluorescence intensity of E-cadherin expression was markedly decreased, whereas the mean fluorescence intensity of Vimentin was increased in epithelial cells after DT-061 treatment in comparison with Wnt-induced EMT group (Figure 8). The results show that activation of PP2A promotes EMT in airway epithelial cells.
PP2A Inhibitor Mitigate the Development of Nasal Polyps, EMT and Eosinophilic Inflammation in Murine NP Model
The murine nasal polyp model was developed through administration of SEB and OVA. LB-100 was administered to mice in the intervention group from day 47. The nasal mucosa of the NP model group showed significant polypoid lesions, increased mucosal thickening and folds, eosinophilic infiltration, as well as a disorganized arrangement of epithelial cilia compared with control group. In PP2A inhibitor treated group, although inflammatory changes were still present, the incidence of polypoid lesions and degree of eosinophilic infiltration were reduced relative to that observed in the NP model group (Figure 9A and B). We also analyzed the changes in expression levels of E-cadherin and Vimentin in the nasal mucosal epithelium of mice from different group by immunohistochemical staining. We observed a significant increase in Vimentin expression and a decrease in E-cadherin expression in NP model group compared to control. Meanwhile, E-cadherin expression was increased and Vimentin expression was decreased in PP2A inhibitor treated group compared to NP model group (Figure 9C). The results indicate that inhibition of PP2A may mitigate the development of nasal polyps, EMT and eosinophilic inflammation in murine NP model.
Figure 9 Continued. Figure 9 PP2A inhibitor mitigate the development of nasal polyps, EMT and eosinophilic inflammation in murine NP model. (A) murine NP models were evaluated by H&E staining; dotted boxes represent polypoid lesions. The number of murine nasal polypoid lesion in each group were counted. (B) eosinophil infiltration in the submucosa in each group (x400 magnification). (C) Immunohistochemical intensity of E-cadherin and Vimentin in the nasal mucosal epithelium of mice from different groups (x 200 magnification). For A, data from NP model group and intervention group were analyzed by Mann–Whitney U-Test. For B, data were analyzed by the one-way ANOVA with the Tukey post hoc test. For C, data were analyzed by Brown-Forsythe test for expression of E-cadherin, data were analyzed by the one-way ANOVA with the Tukey post hoc test for expression of Vimentin, *P <0.05, **P <0.01; ***P < 0.001, ns: no significant difference.

Discussion
Previous studies have reported that Wnt/β-catenin signaling pathway is activated in lower airway diseases.20–24 The similar phenomena have been observed in NPs in recent studies.25 We assayed the expression levels of Wnt pathway modulators in tissues. Our experimental findings show a notable increased expression of active β-catenin in NPs. β-catenin serves a dual role in intracellular signal transduction and cell adhesion as a key mediator of Wnt/β-catenin signaling. On one hand, β-catenin constructs the cell membrane with E-cadherin, while on the other hand, it translocates into the nuclei where it associates with TCF/LEF under conditions of active Wnt signaling, resulting in the transcriptional activation of target genes.26 We also observed a downregulation of GSK3β expression and an upregulation of Phospho-GSK3β(Ser9) expression in NPs. GSK3β serves as another critical modulator of the Wnt pathway. In the absence of Wnt signaling, GSK3β associates with Axin, adenomatous polyposis coli (APC), CK1, and β-catenin to form a complex. GSK3β is activated and then phosphorylates β-catenin. This phosphorylation event leads to the ubiquitination of β-catenin, resulting in its degradation through the proteasomal pathway. When Wnt signaling is activated, the canonical Wnt-signaling pathway is initiated through the association of a Wnt ligand with Frizzled-related protein (FRP), leading to the activation of Dishevelled (Dsh) through phosphorylation, which then phosphorylates GSK3β.27 The inactivation of GSK3β leads to the dissociation of β-catenin from the degradation complex and its dephosphorylation, resulting in an aggregation of β-catenin within the cytoplasmic compartment. The outcomes of our experiments supported activation of Wnt/β-catenin pathway in CRSwNP. Aberrant Wnt signaling results in overactivation of Wnt pathway and promotes transcription of genes involved in EMT downstream of this pathway.
We also investigated the markers associated with EMT, and the findings indicated the presence of EMT process in CRSwNP. Previous research has substantiated the observation of a reduction in epithelial markers accompanied by an increase in mesenchymal markers in CRS.28–30 Differentiated epithelial cells lose their junctional attachments to one another during the EMT process, which causes epithelial barrier dysfunction. Epithelial barrier defects due to compromised cell-cell junctions have been suggested as significant contributors to the pathogenesis of NPs by allowing invasion of pathogens into the underlying mucosal tissues and subsequently elicit immune responses.31,32 As further proof of WNT pathway promoting EMT in CRSwNP, we cultured BEAS-2B cells with rhWNT3A, the experimental data showed that activation of Wnt pathway could induce EMT in epithelial cells. Jun-Sang Bae et al previously reported that rhWNT3A treatment induced EMT in human nasal epithelial cells(HNEpCs) and inhibition of Wnt signaling relieved EMT in murine NP models.33 Thus, it is reasonable to believe that abnormal Wnt signaling may exacerbate EMT of the nasal mucosa. Therefore, targeting key regulatory molecules of WNT pathway represents a promising therapeutic strategy for CRSwNP.
Although studies have shown that abnormal WNT signaling is associated with the pathogenesis of CRSwNP, the mechanisms by which the activated Wnt signaling leads to disruption of epithelial barrier integrity and promotion of inflammatory responses in NPs remain inadequately understood. Furthermore, it is unclear whether there is a difference in the regulatory mechanism of Wnt/β-catenin pathway between EosCRSwNP and non-EosCRSwNP.
PP2A is recognized as a modulator protein of Wnt pathway.34 Previous studies have focused on its role in Alzheimer’s disease35 and tumor immunity.36,37 It has been established that PP2A is integral to embryonic development and tumor suppression by modulating Wnt signaling.38,39 Nevertheless, the function of PP2A in CRSwNP remains unknown.
As far as we are aware, we first report the role of PP2A in the pathogenesis of nasal polyps, we have demonstrated increase of PP2A in NPs at both the transcriptional and expression levels, and the activity of PP2A was also found to be increased in NPs. Moreover, in vitro studies have provided evidence that, through the regulation of Wnt/β-catenin signaling, the inhibition of PP2A may suppress EMT, while stimulation of PP2A aggravates EMT in epithelial cells. Animal experiments reveal that inhibition of PP2A could reduce the number of polypoid lesions, mitigate EMT and alleviate eosinophilic inflammation of nasal mucosal epithelium in murine NP model. Our experiments show that PP2A may promote EMT through regulation of Wnt/β-catenin pathway and contribute to the formation of nasal polyps. PP2A is a heterotrimeric protein complex. Early studies have reported PP2A’s dual regulation of Wnt Signaling pathway. A study demonstrated that inhibition of PP2A through the administration of okadaic acid or silencing of the PP2Ac catalytic subunit led to elevated levels of phosphorylated-GSK3β,40 supporting a possible function of PP2A in the stimulation of GSK3β activity, which subsequently results in the phosphorylation and ultimately degradation of β-catenin. Another study provided evidence for the involvement of PP2A as a constituent of the β-catenin destruction complex,41 and indicated PP2A’s negative regulation of Wnt Signaling pathway. On the contrary, some studies suggested that PP2A exerts a positive regulatory effect on the Wnt signaling. Zhang et al showed that the regulatory subunit PR55α of PP2A could directly interact with β-catenin, dephosphorylate the main effector and increase its abundance.42 Strovel et al found that the overexpression of PP2Ac dephosphorylated scaffolding protein Axin,43 which serves as an inhibitory regulator of Wnt signaling by enhancing phosphorylation of β-catenin. Our study demonstrated that the inhibition of PP2A could reduce activated β-catenin expression levels, resulting in inhibition of Wnt signaling and alleviation of EMT in epithelial cells. Meanwhile, stimulation of PP2A increases expression of activated β-catenin, leading to activation of Wnt signaling and promoting EMT in epithelial cells. The results indicate that PP2A exerts positive regulation on Wnt signaling pathway in CRSwNP. We speculate that PP2A may regulate Wnt signaling in CRSwNP through dephosphorylating components of degradation complex and preventing phosphorylation of β-catenin, or directly interacting with and dephosphorylating β-catenin to activate Wnt signaling. Deeper research is needed to elucidate specific elements of Wnt pathway that are targeted by PP2A in CRSwNP.
We also compared the differences in expression of PP2A, Wnt signaling pathway modulators and EMT-related markers between two endotypes of CRSwNP. Compared to non-eosinophilic NPs, a notably enhanced expression of WNT3A and PP2A was observed, while E-cadherin expression was reduced in eosinophilic NPs. In animal experiments, we also observed a reduction in the number of eosinophilic infiltrations in PP2A inhibition group compared with that in NP model group. The results indicated that PP2A may play a more important role for promoting EMT in eosinophilic nasal polyps. Eosinophil infiltration serves as a manifestation of the type 2 inflammation characteristics in CRS. Eosinophilic inflammation has been recognized as a major pathological mechanism for poor efficacy of CRSwNP,44 PP2A may contribute to the eosinophilic inflammation in NPs, but more in-depth study is needed to explore mechanism by which PP2A contributes to eosinophilic inflammation in NPs.
In this study, we have already identified the effect of Wnt/ β-catenin pathway and PP2A in CRSwNP. Nevertheless, several limitations should be acknowledged. First, although our in vivo data suggest that PP2A inhibition alleviates EMT and eosinophilic inflammation, the relatively small animal sample size limits the statistical power of these findings, necessitating future validation with larger cohorts. Second, whether the non-canonical Wnt pathway possesses similar regulatory mechanism remains unexplored. Third, while the BEAS-2B cell line is a well-established model for studying airway epithelial biology, future investigations employing HNEpCs are necessary to validate the role of the PP2A/Wnt/β-catenin signaling pathway in CRSwNP. Finally, the exact mechanism underlying the regulation of the Wnt/β-catenin pathway by PP2A remains to be elucidated. Our subsequent research will therefore focus on these aspects, especially on PP2A’s role in type 2 inflammation within eosinophilic polyps.
Conclusion
Our study has demonstrated overexpression of PP2A, as well as activation of Wnt/β-catenin signaling pathway in both endotypes of NPs, with a more pronounced effect observed in eosinophilic NPs. In vitro, Wnt can induce EMT in respiratory epithelial cells, while inhibition of PP2A could alleviate EMT and stimulation of PP2A could aggravate EMT in epithelial cells. We believe that PP2A promotes EMT in CRSwNP by exerting its positive influence on Wnt/β-catenin signaling pathway. Targeting PP2A might represent a viable therapeutic option for treating CRSwNP, while its therapeutic viability awaits a more comprehensive understanding of the mechanism and validation in larger-scale in vivo models.
Abbreviations
CRS, chronic rhinosinusitis; CRSwNP, Chronic rhinosinusitis with nasal polyps; PP2A, Protein Phosphatase 2A; EMT, epithelial-mesenchymal transition; NPs, nasal polyps; OVA, ovalbumin; SEB, staphylococcal enterotoxins B; APC, adenomatous polyposis coli; FRP, Frizzled-related protein; Dsh, Dishevelled; CK1: Casein Kinase1; EosCRSwNP, eosinophilic CRSwNP; non-EosCRSwNP, non-eosinophilic CRSwNP. HNEpCs: Human Nasal Epithelial Cells.
Acknowledgments
We would like to express our sincere gratitude to all participants who have contributed to the successful completion of this study.
Funding
This work was supported by Chongqing Health Commission and Science and Technology Bureau (grant number 2023GDRC003).
Disclosure
All authors declared no conflicts of interest in this work.
References
1. Fokkens WJ. EPOS2020: a major step forward. Rhinology J. 2020;58(Suppl 1):1–464. doi:10.4193/Rhin20.401
2. Kim DK, Eun KM, Kim MK, et al. Comparison between signature cytokines of nasal tissues in subtypes of chronic rhinosinusitis. Allergy Asthma Immunol Res. 2019;11(2):201–211. doi:10.4168/aair.2019.11.2.201
3. Bai J, Tan BK, Kato A. Endotypic heterogeneity and pathogenesis in chronic rhinosinusitis. Curr Opin Allergy Clin Immunol. 2024;24(1):1–8. doi:10.1097/aci.0000000000000954
4. Yan B, Lan F, Li J, et al. The mucosal concept in chronic rhinosinusitis: focus on the epithelial barrier. J Allergy Clin Immunol. 2024;153(5):1206–1214. doi:10.1016/j.jaci.2024.01.015
5. Schleimer RP. Immunopathogenesis of chronic rhinosinusitis and nasal polyposis. Annu Rev Pathol. 2017;12:331–357. doi:10.1146/annurev-pathol-052016-100401
6. Könnecke M, Burmeister M, Pries R, et al. Epithelial-mesenchymal transition in chronic rhinosinusitis: differences revealed between epithelial cells from nasal polyps and inferior turbinates. Arch Immunol Ther Exp. 2017;65(2):157–173. doi:10.1007/s00005-016-0409-7
7. Savagner P. The epithelial-mesenchymal transition (EMT) phenomenon. Ann Oncol. 2010;21(7):vii89–92. doi:10.1093/annonc/mdq292
8. He Y, Fu Y, Wu Y, et al. Pathogenesis and treatment of chronic rhinosinusitis from the perspective of sinonasal epithelial dysfunction. Front Med Lausanne. 2023;10:1139240. doi:10.3389/fmed.2023.1139240
9. Xia Y, Wang H, Yin J. The role of epithelial-mesenchymal transition in chronic rhinosinusitis. Int Arch Allergy Immunol. 2022;183(10):1029–1039. doi:10.1159/000524950
10. Yuan J, Wang M, Wang C, et al. Epithelial cell dysfunction in chronic rhinosinusitis: the epithelial-mesenchymal transition. Expert Rev Clin Immunol. 2023;19(8):959–968. doi:10.1080/1744666x.2023.2232113
11. Zhang J, Tian XJ, Xing J. Signal transduction pathways of EMT induced by TGF-β, SHH, and WNT and their crosstalks. J Clin Med. 2016;5(4). doi:10.3390/jcm5040041
12. Nusse R, Clevers H. Wnt/β-Catenin signaling, disease, and emerging therapeutic modalities. Cell. 2017;169(6):985–999. doi:10.1016/j.cell.2017.05.016
13. Böscke R, Vladar EK, Könnecke M, et al. Wnt signaling in chronic rhinosinusitis with nasal polyps. Am J Respir Cell Mol Biol. 2017;56(5):575–584. doi:10.1165/rcmb.2016-0024OC
14. Thompson J, Williams C. Protein phosphatase 2A in the regulation of wnt signaling, stem cells, and cancer. Genes. 2018;9(3). doi:10.3390/genes9030121
15. Rahman MM, Rumzhum NN, Hansbro PM, et al. Activating protein phosphatase 2A (PP2A) enhances tristetraprolin (TTP) anti-inflammatory function in A549 lung epithelial cells. Cell Signalling. 2016;28(4):325–334. doi:10.1016/j.cellsig.2016.01.009
16. Nair PM, Starkey MR, Haw TJ, et al. Targeting PP2A and proteasome activity ameliorates features of allergic airway disease in mice. Allergy. 2017;72(12):1891–1903. doi:10.1111/all.13212
17. McHugh T, Snidvongs K, Xie M, et al. High tissue eosinophilia as a marker to predict recurrence for eosinophilic chronic rhinosinusitis: a systematic review and meta‐analysis. Int Forum Allergy Rhinol. 2018;8(12):1421–1429. doi:10.1002/alr.22194
18. Shu L, Zheng B, Liu Y, et al. Piezo1 regulates TGF-β1 induced epithelial-mesenchymal transition in chronic rhinosinusitis with nasal polyps. Mol Immunol. 2024;175:63–73. doi:10.1016/j.molimm.2024.09.004
19. Wee JH, Ko Y-K, Khalmuratova R, et al. Effect of lipopolysaccharide and polyinosinic:polycytidylic acid in a murine model of nasal polyp. Sci Rep. 2021;11(1). doi:10.1038/s41598-020-80483-y
20. Conlon TM, John-Schuster G, Heide D, et al. Inhibition of LTβR signalling activates WNT-induced regeneration in lung. Nature. 2020;588(7836):151–156. doi:10.1038/s41586-020-2882-8
21. Kaya-Yasar Y, Engin S, Barut EN, et al. The contribution of the WNT pathway to the therapeutic effects of montelukast in experimental murine airway inflammation induced by ovalbumin and lipopolysaccharide. Drug Dev Res. 2024;85(2):e22178. doi:10.1002/ddr.22178
22. Kwak HJ, Park DW, Seo JY, et al. The Wnt/β-catenin signaling pathway regulates the development of airway remodeling in patients with asthma. Exp Mol Med. 2015;47(12):e198. doi:10.1038/emm.2015.91
23. Yao L, Zhao H, Tang H, et al. Blockade of β-catenin signaling attenuates toluene diisocyanate-induced experimental asthma. Allergy. 2017;72(4):579–589. doi:10.1111/all.13045
24. Li P, Han X, Li J, et al. Aerobic exercise training engages the canonical wnt pathway to improve pulmonary function and inflammation in COPD. BMC Pulm Med. 2024;24(1):236. doi:10.1186/s12890-024-03048-z
25. Dobzanski A, Khalil SM, Lane AP. Nasal polyp fibroblasts modulate epithelial characteristics via wnt signaling. Int Forum Allergy Rhinol. 2018;8(12):1412–1420. doi:10.1002/alr.22199
26. Silva-García O, Valdez-Alarcón JJ, Baizabal-Aguirre VM. The Wnt/β-catenin signaling pathway controls the inflammatory response in infections caused by pathogenic bacteria. Mediators Inflamm. 2014;2014:310183. doi:10.1155/2014/310183
27. Clevers H, Nusse R. Wnt/β-catenin signaling and disease. Cell. 2012;149(6):1192–1205. doi:10.1016/j.cell.2012.05.012
28. Zhang N, Van Crombruggen K, Gevaert E, et al. Barrier function of the nasal mucosa in health and type-2 biased airway diseases. Allergy. 2016;71(3):295–307. doi:10.1111/all.12809
29. Jang YJ, Kim HG, Koo TW, et al. Localization of ZO-1 and E-cadherin in the nasal polyp epithelium. Eur Arch Otorhinolaryngol. 2002;259(9):465–469. doi:10.1007/s00405-002-0500-z
30. Rogers GA, Den Beste K, Parkos CA, et al. Epithelial tight junction alterations in nasal polyposis. Int Forum Allergy Rhinol. 2011;1(1):50–54. doi:10.1002/alr.20014
31. Min HJ, Choe JW, Kim KS, et al. High-mobility group box 1 protein induces epithelialmesenchymal transition in upper airway epithelial cells. Rhinology. 2020;58(5):495–505. doi:10.4193/Rhin18.281
32. Xiao C, Puddicombe SM, Field S, et al. Defective epithelial barrier function in asthma. J Allergy Clin Immunol. 2011;128(3):549–556.e541–512. doi:10.1016/j.jaci.2011.05.038
33. Bae J-S, Ryu G, Kim JH, et al. Effects of Wnt signaling on epithelial to mesenchymal transition in chronic rhinosinusitis with nasal polyp. Thorax. 2020;75(11):982–993. doi:10.1136/thoraxjnl-2019-213916
34. Ratcliffe MJ, Itoh K, Sokol SY. A positive role for the PP2A catalytic subunit in wnt signal transduction. J Biol Chem. 2000;275(46):35680–35683. doi:10.1074/jbc.C000639200
35. Arribas RL, Viejo L, Bravo I, et al. C-glycosides analogues of the okadaic acid central fragment exert neuroprotection via restoration of PP2A-phosphatase activity: a rational design of potential drugs for Alzheimer’s disease targeting tauopathies. Eur J Med Chem. 2023;251:115245. doi:10.1016/j.ejmech.2023.115245
36. Johnson H, Narayan S, Sharma AK. Altering phosphorylation in cancer through PP2A modifiers. Cancer Cell Int. 2024;24(1):11. doi:10.1186/s12935-023-03193-1
37. Cazzoli R, Romeo F, Pallavicini I, et al. Endogenous PP2A inhibitor CIP2A degradation by chaperone-mediated autophagy contributes to the antitumor effect of mitochondrial complex I inhibition. Cell Rep. 2023;42(6):112616. doi:10.1016/j.celrep.2023.112616
38. Park D-S, Yoon G-H, Lee H-S, et al. Capsaicin inhibits the Wnt/β-catenin signaling pathway by down-regulating PP2A. Biochem Biophys Res Commun. 2016;478(1):455–461. doi:10.1016/j.bbrc.2016.06.075
39. Zong Z, Dang Y, Zhang Y, et al. Promotion effect on liver tumor progression of microcystin-LR at environmentally relevant levels in female krasV12 transgenic zebrafish. Aquat Toxicol. 2022;252:106313. doi:10.1016/j.aquatox.2022.106313
40. Yokoyama N, Malbon CC. Phosphoprotein phosphatase-2A docks to dishevelled and counterregulates wnt3a/beta-catenin signaling. J Mol Signal. 2007;2:12. doi:10.1186/1750-2187-2-12
41. Li X, Yost HJ, Virshup DM, et al. Protein phosphatase 2A and its B56 regulatory subunit inhibit wnt signaling in xenopus. EMBO j. 2001;20(15):4122–4131. doi:10.1093/emboj/20.15.4122
42. Zhang W, Yang J, Liu Y, et al. PR55 alpha, a regulatory subunit of PP2A, specifically regulates PP2A-mediated beta-catenin dephosphorylation. J Biol Chem. 2009;284(34):22649–22656. doi:10.1074/jbc.M109.013698
43. Strovel ET, Wu D, Sussman DJ. Protein phosphatase 2Calpha dephosphorylates axin and activates LEF-1-dependent transcription. J Biol Chem. 2000;275(4):2399–2403. doi:10.1074/jbc.275.4.2399
44. Bachert C, Hicks A, Gane S, et al. The interleukin-4/interleukin-13 pathway in type 2 inflammation in chronic rhinosinusitis with nasal polyps. Front Immunol. 2024;15:1356298. doi:10.3389/fimmu.2024.1356298
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Improvement in Health-Related Quality of Life with Dupilumab in Patients with Moderate-to-Severe Asthma with Comorbid Chronic Rhinosinusitis with/without Nasal Polyps: An Analysis of the QUEST Study
Hopkins C, Buchheit KM, Heffler E, Cohen NA, Olze H, Khan AH, Msihid J, Siddiqui S, Nash S, Jacob-Nara JA, Rowe PJ, Deniz Y
Journal of Asthma and Allergy 2022, 15:767-773
Published Date: 7 June 2022
Profiling Disease and Economic Burden in CRSwNP Using Machine Learning
Bhattacharyya N, Silver J, Bogart M, Kponee-Shovein K, Cheng WY, Cheng M, Cheung HC, Duh MS, Hahn B
Journal of Asthma and Allergy 2022, 15:1401-1412
Published Date: 30 September 2022
Tezepelumab Efficacy in Patients with Severe, Uncontrolled Asthma with Comorbid Nasal Polyps in NAVIGATOR

Laidlaw TM, Menzies-Gow A, Caveney S, Han JK, Martin N, Israel E, Lee JK, Llanos JP, Martin N, Megally A, Parikh B, Vong S, Welte T, Corren J
Journal of Asthma and Allergy 2023, 16:915-932
Published Date: 4 September 2023
Remodeling of Paranasal Sinuses Mucosa Functions in Response to Biofilm-Induced Inflammation
Kaliniak S, Fiedoruk K, Spałek J, Piktel E, Durnaś B, Góźdź S, Bucki R, Okła S
Journal of Inflammation Research 2024, 17:1295-1323
Published Date: 26 February 2024
Minimal Clinically Important Difference of the Taiwan Smell Test in Patients with Bilateral Chronic Rhinosinusitis and Nasal Polyps Post-Sinus Surgery
Wu PW, Shen PH, Lee YS, Chang PH, Huang CC, Huang CC
Journal of Asthma and Allergy 2025, 18:1347-1353
Published Date: 1 October 2025