Back to Journals » Infection and Drug Resistance » Volume 15
Plasmodium falciparum Malaria Susceptibility and Severity: Influence of MyD88-Adaptor-Like Gene (rs8177374) Polymorphism
Authors Ammar AI, El-Hefnawy SM, Shehab-Eldeen S ![]() , Essa A
, Essa A ![]() , ELnaidany SS, Mostafa RG, Alsalman MH, El-Refai SA
, ELnaidany SS, Mostafa RG, Alsalman MH, El-Refai SA
Received 24 August 2022
Accepted for publication 2 November 2022
Published 28 November 2022 Volume 2022:15 Pages 6815—6827
DOI https://doi.org/10.2147/IDR.S387463
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Suresh Antony
Amany I Ammar,1 Sally M El-Hefnawy,2 Somaia Shehab-Eldeen,3,4 Abdallah Essa,3,4 Sherin S ELnaidany,2 Rasha G Mostafa,5 Mortadah H Alsalman,4 Samar A El-Refai1
1Department of Medical Parasitology, Faculty of Medicine, Menoufia University, Shebin Elkom, Egypt; 2Department of Medical Biochemistry & Molecular Biology, Faculty of Medicine, Menoufia University, Shebin Elkom, Egypt; 3Tropical Medicine Department, Faculty of Medicine, Menoufia University, Shebin Elkom, Egypt; 4Internal Medicine Department, College of Medicine, King Faisal University, Al-Ahsa, Kingdom of Saudi Arabia; 5Department of Medical Microbiology and Immunology, Faculty of Medicine, Menoufia University, Shebin Elkom, Egypt
Correspondence: Somaia Shehab-Eldeen, Faculty of Medicine, Menoufia University, Yassen Abd Al Ghafar Street, Shebin Elkom, Menoufia, 32511, Egypt, Tel +20 1117251523, Email [email protected]
Background: MyD88-adapter-like (MAL), as an essential adapter protein for a variety of TLRs (Toll-like receptors), modulates the inflammatory response. Many infectious illnesses are influenced by single nucleotide polymorphisms (SNPs) that modify MAL function. We aimed to examine the influence of the MAL rs8177374 polymorphism on Plasmodium falciparum malaria susceptibility and severity.
Patients and Methods: Samples from 141 Plasmodium falciparum malaria patients and 147 healthy controls were used in the study. Patients were subdivided into mild and severe groups based on their clinical results, as defined by the World Health Organization (WHO). Genotypes for MAL rs8177374 were identified by allele-specific PCR technique, and TNF-alpha and IL-12 levels were measured using ELISA.
Results: The MAL rs8177374 (CT) genotype is associated with an increased risk of malaria (OR: 2.52; 95% CI: 1.44– 4.41). Furthermore, the CT and TT genotypes gave considerable protection against severe malaria (OR: 0.07; 95% CI: 0.03– 0.19 and OR: 0.03; 95% CI: 0.007– 0.1 respectively). And the T allele was linked to a higher risk of malaria (OR: 1.7; 95% CI: 1.18– 2.5), while protecting patients from severe malaria (OR: 0.135; 95% CI: 0.07– 0.3). Mutants (CT and TT) have greater TNF-alpha and IL-12 levels compared to wild-type (CC).
Conclusion: Malaria risk is linked to single nucleotide polymorphism in the MyD88-adaptor-like gene. People with the MAL rs8177374 mutant variant may be less likely to get severe malaria.
Keywords: MAL, malaria, P. falciparum, TNF-alpha, IL-12
Introduction
Malaria is a grave parasite disease spread by mosquitoes that is one of the most widespread and dangerous infectious diseases in the tropics and subtropics.1 Clinical symptoms of P. falciparum malaria differ from patient to patient, ranging from asymptomatic infections to life-threatening situations.2 Malaria episodes begin as a result of the interplay between malarial toxins released during red blood cell rupture and the innate immune system’s phagocytic cells.3 Toxins produced by malaria are referred to as pathogen-associated molecular patterns (PAMPs). They are identified primarily by toll-like receptors (TLRs).4
TLRs are a transmembrane protein family found in a variety of cells, including monocytes, macrophages, and dendritic cells. These receptors are critical components of the innate immune response to Plasmodium infection.5 They are classified as pattern recognition receptors tasked with the task of recognizing PAMPs.6 P. falciparum glycosylphosphatidylinositol (GPI) is recognized by TLR2 and TLR4.7 And once TLR4 detects the PAMPs of malaria, TLR4 promotes downstream signalling, which activates nuclear factor kappa-B (NF- kB). TNF-alpha, IL-1, IL-6, IL-8, and IL-12 are among the inflammatory cytokines produced as a result of this.8 The inflammatory cytokine response is critical for parasite control and clearance since it facilitates host immunity against the Plasmodium parasite. Excessive cytokine response, on the other hand, can have disastrous host effects, including tissue injury, which is frequently associated with severe complicated malaria.9 TLRs’ signalling, particularly TLR2 and TLR4, is mediated by a protein called MyD88 adaptor-like (MAL) encoded by the myeloid differentiation gene 88 (MyD88).10 When a TLR is activated, MyD88 facilitates the recruitment of various additional proteins, which leads to a downstream signaling cascade that results in the synthesis of pro-inflammatory proteins through MAPK and the NF-B pathway.11 MyD88 is thought to play a crucial role in the signalling cascades that are mediated by all TLRs, IL-1R, IL-18R, IFN, and IL-33.12–16
Single nucleotide polymorphisms (SNPs) in MAL-encoding genes impact their function, affecting the inflammatory process, cytokine generation, parasitaemia severity, clinical manifestations of the disease and Plasmodium clearance.17 The altered function of MAL has been linked to disease susceptibility or resistance, according to Belhaouane et al. These opposite effects demonstrate the paradoxical functions of MAL and its role in infectious diseases.18
The SNP (rs8177374) causes nucleotide substitution (C to T) at the position of 991bp, resulting in the production of leucine instead of serine in MAL protein. This is supposed to affect MAL function with reduced cytokine production that determines the severity of infection.19
Generally, genetic alterations and their effects remain controversial, with no conclusive evidence of their influence on disease outcome.5 This study aimed to explore the role of the rs8177374 polymorphism of the MyD88-adapter-like (MAL) gene in controlling the severity of P. falciparum infection. The link between the rs8177374 polymorphism in the MAL gene and blood levels of IL-12 and TNF-alpha was also examined.
Subjects and Methods
Study Design
The current case-control study was conducted between June 2017 and August 2021. Patients were recruited from Abbassia Fever Hospital, Cairo, Egypt.
Subjects included in this work were divided into two main groups:
Malaria Group
It included 141 Egyptian patients with P. falciparum infection. The diagnosis of P. falciparum malaria in this group was based on positive rapid diagnostic tests and positive blood films with more than 2 asexual stages of the parasite.
Patients in this group were further divided into two subgroups:
Severe Malaria (SM): It includes 57 patients with severe or complicated P. falciparum malaria. Severe P. falciparum malaria patients were selected according to WHO criteria for severe malaria. They had at least one or more of the following: impaired consciousness (Glasgow score <11), generalized weakness (unable to sit, stand or walk without an aid), convulsions (> two episodes within 24 h), acidosis (plasma base excess < −3.3 mmol/L), hypoglycemia (blood glucose <2.2 mmol/L), severe malarial anaemia (haemoglobin concentration ≤7.0 g/dL in adults with parasite count >10,000/µL), renal impairment (serum creatinine >265 µmol/L), jaundice (serum bilirubin >50µmol/L), pulmonary oedema (confirmed by radiology or oxygen saturation <92% with a respiratory rate >30/min), significant bleeding (recurrent nose and gums bleeding, hematemesis or melena), shock (compensated without hypotension or decompensated with blood pressure <80 Hg) and hyperparasitemia (parasitemia >10% of RBCs or >250,000 parasites/µL in non-immune subjects).20
Mild Malaria (MM): It includes 84 patients with mild or uncomplicated P. falciparum malaria. They have symptoms of malaria with the absence of criteria for severe malaria.20
The Healthy Control Group (HC)
It included 147 blood donors. They have the following characteristics: Their birthplace was in the study area, they had no history of malaria infection, had no symptoms of the disease, and had negative results from thick blood film and rapid diagnostic test.
Patients with other Plasmodium species co-infection, patients with chronic diseases (liver cirrhosis, chronic renal failure, and tuberculosis), and those who have autoimmune diseases such as systemic lupus erythematosus or rheumatoid arthritis were excluded from the study.
Every patient in the study was assessed for the criteria of the severity of P. falciparum malaria through detailed history taking, full clinical examination, radiological and laboratory investigations.
Ethical Considerations
Written informed consent was taken from every subject who participated in this study. The study was approved by the local ethics committee of the Faculty of Medicine, Menoufia University. The personal information and human biological samples of study participants were de-identified and secured to protect their privacy, and the ethical guidelines set by the 1975 Helsinki Declaration were followed during this study.
Sampling
Venous blood samples were taken from each patient for rapid diagnostic tests and thin and thick blood films. The remaining blood samples were transferred to clean, sterile, dry tubes containing EDTA. Blood specimens were stored at −20°C for genotyping of the MAL gene.
Rapid Diagnostic Testing for Malaria
The RDT kit, SD BIOLINE Malaria Antigen P.F HRP2/PLDH (Cat no. 05FK9, Abbott Diagnostics, Korea Inc.) was used according to the manufacturer’s instructions. It is an immunochromatographic test that is used for the detection of histidine‐rich protein II (HRP‐II) and Plasmodium lactate dehydrogenase (pLDH) of P. falciparum in human whole blood.
Blood Films: Thin and Thick
Thin and thick blood films were prepared, stained with 10% Giemsa, and microscopically examined using a light microscope (100× oil immersion objective lens) to determine malaria species as shown in Figure 1. The examination was done by two independent examiners who were blinded to the results of each other.21 The negativity of the thick blood film was considered after examination of at least 100 fields, each containing around 20 WBCs.
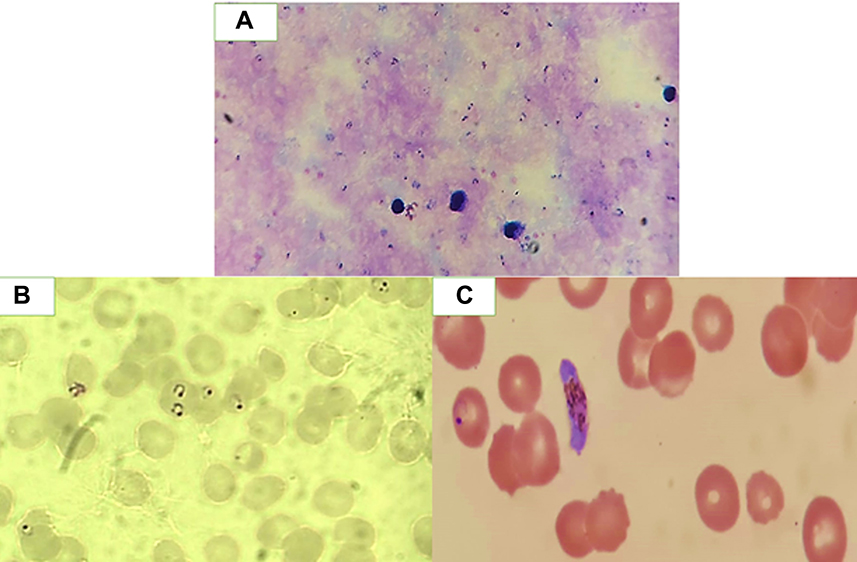 |
Figure 1 Photographs of P. falciparum (X 100) (A) Thick blood film showing ring stages of the parasite. (B) Thin blood film showing red blood cells containing ring stages. (C) Thin blood film showing gametocyte stage. |
Determination of Serum Levels of IL-12 and TNF-Alpha
Sandwich ELISA was performed to measure the serum levels of TNF-alpha and IL-12. The anti-cytokine antibodies were used by BD Pharmingen (Cat. No. 551220 and 551227, respectively, San Diego, USA). Recombinant cytokines were used as the standards. Briefly, ELISA Wells were coated with the capture antibodies (100μL). A blocking buffer to block the non-specific binding sites was used. Serum and standard samples were added. Then the biotinylated detection antibody was added. Then the conjugate (streptavidin-alkaline phosphatase) was added, followed by para-nitrophenyl phosphate (Sigma). Absorbance was then measured at 405 nm. Cytokine concentrations were calculated using standard curves (Microplate Manager Software, Bio-Rad).
Genotyping of MAL SNP (rs8177374)
First, DNA was extracted from whole blood by the GeneJET Whole Blood Genomic DNA Purification Mini Extraction Kit (Cat no. K0702, Thermo Fisher Scientific, USA) according to the manufacturer’s instructions. Then, Taqman® SNP Genotyping assays (Applied Biosystems by Life Technologies, Monza, Italy) were used to do SNP genotyping on genomic DNA. Genotyping of MAL gene rs8177374 variants was performed on genomic DNA by an allelic discrimination assay using readymade TaqMan probes, Applied Biosystems, USA.
The allelic discrimination test uses two primer and probe pairs to identify variations of a single nucleic acid sequence in each reaction, allowing genotyping of the two variants at the SNP in a target template sequence using TaqMan Genotyping Master Mix (40×). Using the maxima probe qPCR Master Mix, the probe sequence (Thermo Fisher Scientific) was as follows:
(VIC/FAM)GAGGGCTGCACCATCCCCCTGCTGT[C/T]GGGCCTCAGCAGAGCTGCCTACCCA
The reaction was performed in a total volume of 20 μL by adding 10 μL of Master Mix, 1.25 μL of primer/probe mix, and 3.75 μL of DNAase-free water. For the negative control reaction, 5 μL of genomic DNA template and 5 μL of DNAase-free water were used for each sample.
The following were the cycling conditions: Denaturation was first performed at 95°C for 10 minutes, followed by 45 cycles at 92°C for 15s, primer annealing at 60°C for 1 minute, primer extension at 72°C for 2 minutes, and the last extension step at 72°C for 1 minute.
Fluorescence was detected. Analysis of the data was performed using the 7500 Real-Time PCR instrument, Applied Biosystems, version 2.0.1. Figures 2 and 3 show allelic discrimination and amplification plots.
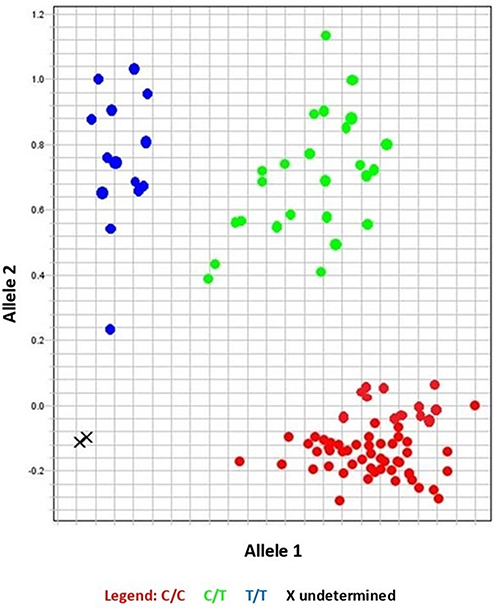 |
Figure 2 Allelic discrimination plot showing different sample genotypes. |
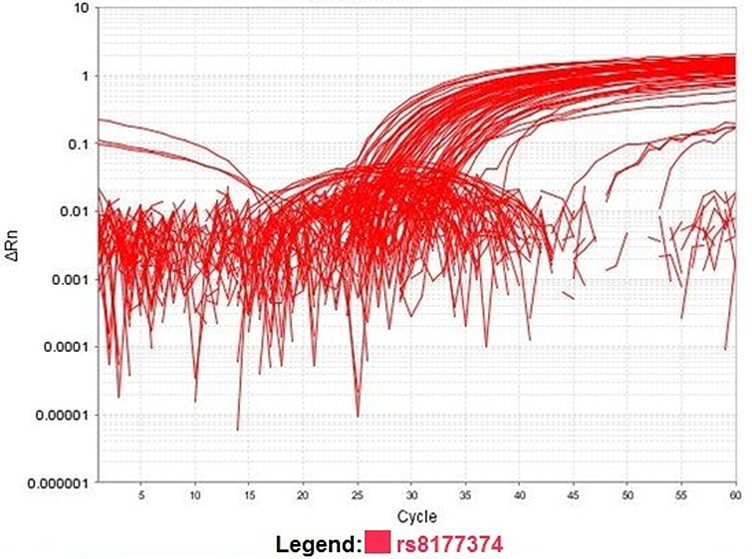 |
Figure 3 Amplification plot showing normalized reporter (Rn) dye fluorescence as a function of the cycle. |
Statistical Analysis
The t-test was used to compare quantitative parametric variables. Analysis of differences in quantitative and non-parametric variables (cytokine levels) between the clinical groups was conducted using the Mann–Whitney test, while the Chi-square test was used for categorical variables. To determine the Hardy-Weinberg equilibrium (HWE), the observed and expected genotype frequencies were compared, while to assess the association between various genotypes and risk, we calculated the odds ratio and 95% confidence interval using multiple logistic regression models. A P-value of less than 0.05 was considered significant.
Results
Study Population Characteristics
The characteristics of the studied groups and subgroups are shown in Table 1.
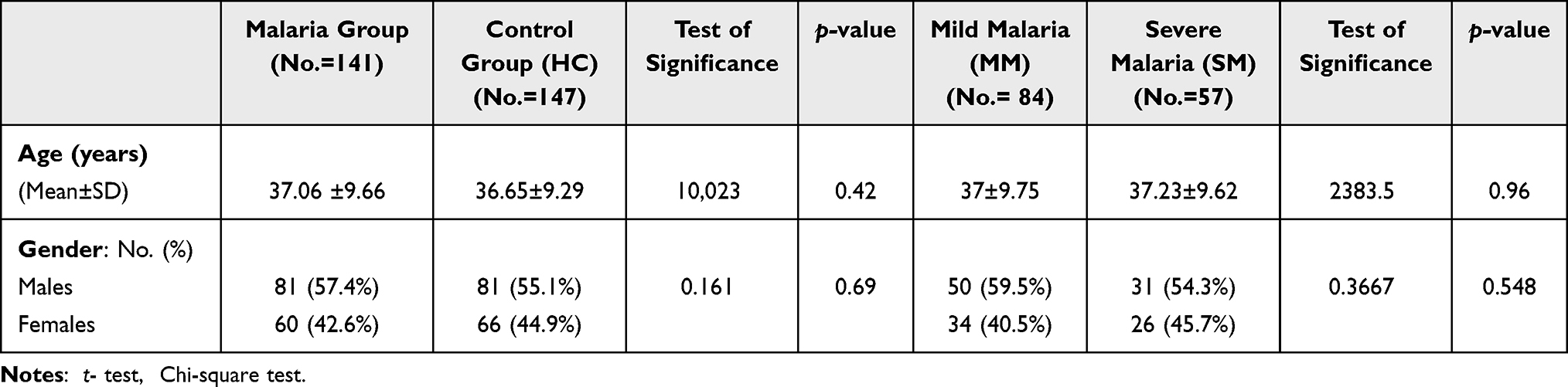 |
Table 1 Demographic Data of the Studied Groups and Subgroups |
Genotype and Allele Frequencies and HWE Estimation
There was a significant difference between patients with malaria and healthy controls regarding MAL genotype frequencies, where the CC genotype had a lower frequency in malaria patients compared to healthy controls. Also, there was a significant difference between both groups regarding allele frequencies, where the C allele had a significantly lower frequency in the malaria group compared to healthy controls (Table 2).
 |
Table 2 Genotype and Allele Frequencies of the MAL Gene (rs8177374) Polymorphism in the P. falciparum Group and Healthy Controls |
There was also a significant difference between mild and severe malaria patients regarding MAL genotypes where the CC genotype had a higher frequency in severe malaria compared to mild malaria patients. Similarly, there was a significant difference between both groups regarding allele frequencies. The C allele was more frequent in patients who had severe malaria than in those who had mild malaria (Table 3).
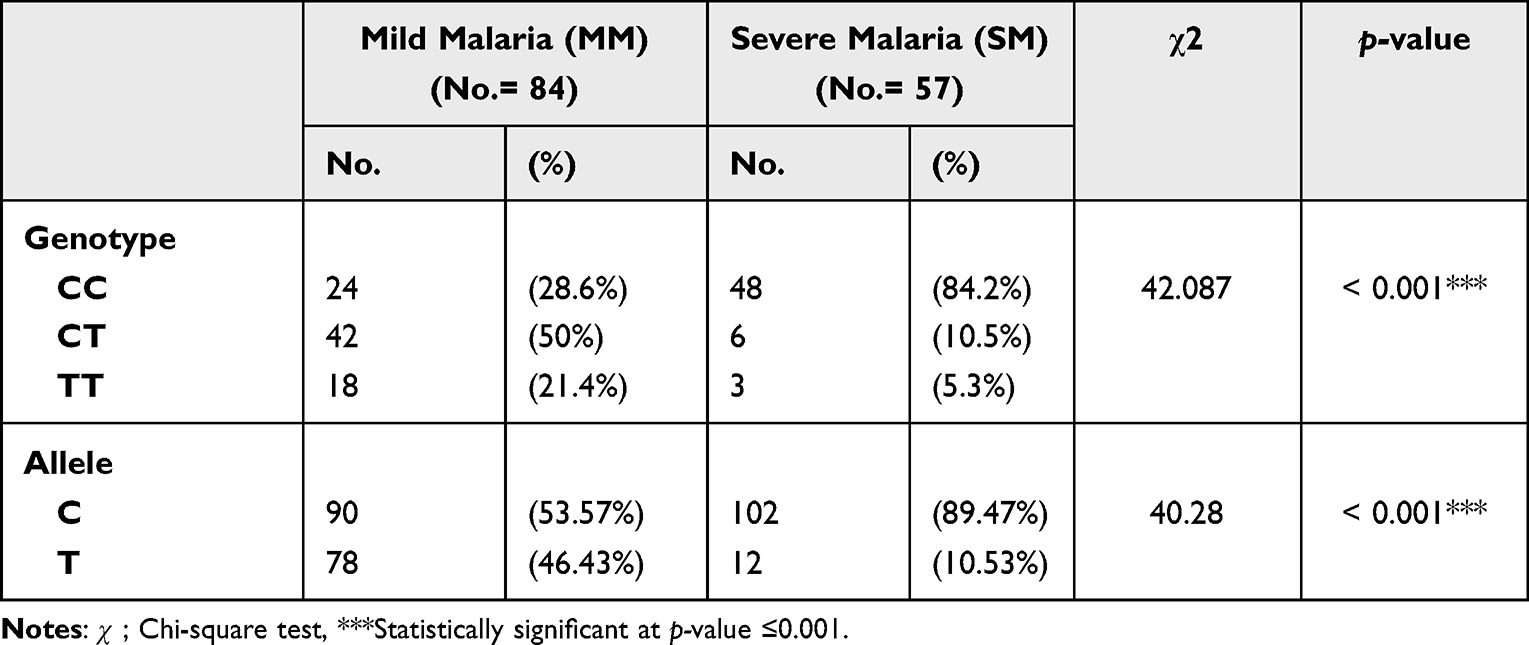 |
Table 3 Genotype and Allele Frequencies of the MAL Gene (rs8177374) Polymorphism in Malaria Subgroups |
HWE estimation showed that allele frequencies in the malaria group, as well as in patients with severe malaria, were deviant from HWE (Supplementary Table 1).
Association of MAL Gene (rs8177374) Polymorphism with Risk of Malaria
The CT genotype increases the risk of developing malaria. While the CT and TT genotypes protect patients from developing severe malaria.
In addition, the T allele was linked to a higher risk of malaria whereas it protects patients from severe malaria as shown in Table 4
 |
Table 4 The Association of the Mal rs8177374 Polymorphism with Malaria Based on Symptoms |
The Association of TNF-Alpha and IL-12 Levels with the MAL Gene (rs8177374) Polymorphism in Malaria Patients
Serum TNF-alpha levels were significantly higher in patients with malaria infection (538.15±284.37) than in healthy controls (40.9±14.9) (p < 0.001), and their levels were significantly higher in severe malaria (600.73±378.33) than in mild malaria (495.68±282.17) (p=0.032). The CC genotype was associated with significantly lower levels of TNF-alpha (414.79±221.37) when compared to the CT genotype (592.54± 258.95) and TT genotype (836.75±281.06) (p < 0.001 and <0.001 respectively) in patients with malaria infection as shown in Figure 4.
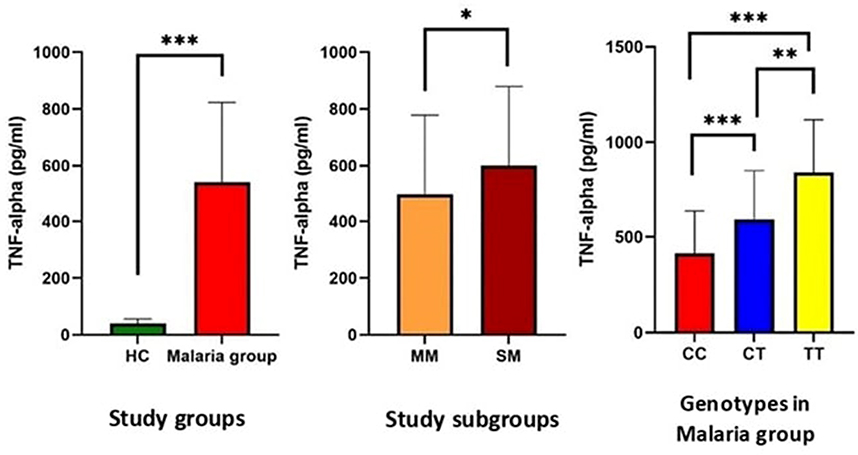 |
Figure 4 Serum levels of TNF-alpha in the studied groups and subgroups and in relation to Mal rs8177374 polymorphism in patients with malaria. Notes: *Statistically significant at p-value ≤ 0.05; **statistically significant at p-value ≤ 0.01; ***statistically significant at p-value ≤ 0.001.Abbreviations: HC; healthy controls, MM; mild malaria, SM; severe malaria. |
As regards IL-12 levels, there was a significant elevation in the malaria group (208.91±116.08) compared to healthy controls (42.8±12.1) (p < 0.001) and its levels were significantly lower in severe malaria (159.05±99.02) than in mild malaria (242.76±115.09) (p < 0.001). Moreover, patients with the CC genotype have significantly lower levels of IL-12 (165.67±93.8) compared to patients with the CT genotype (215.01±97.02) and the TT genotype (343.26 ± 123.25) (p = 0.003 and p < 0.001 respectively) as shown in Figure 5.
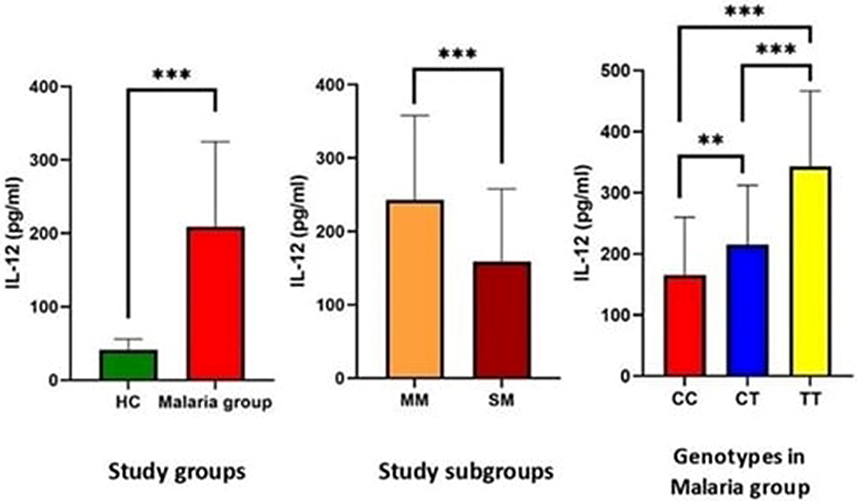 |
Figure 5 Serum levels of IL-12 in the studied groups and subgroups and in relation to Mal rs8177374 polymorphism in patients with malaria. Notes: **Statistically significant at p-value ≤ 0.01; ***statistically significant at p-value ≤ 0.001.Abbreviations: HC; healthy controls, MM; mild malaria, SM; severe malaria. |
While controlling for the malaria phenotype, we observed that the CC genotype was associated with significantly lower levels of TNF-alpha when compared to the CT and TT genotypes in both malaria subgroups. The TT genotype has the highest level of IL-12 in the severe malaria group, while the CT genotype has the highest level in the mild malaria group, as shown in Figure 6.
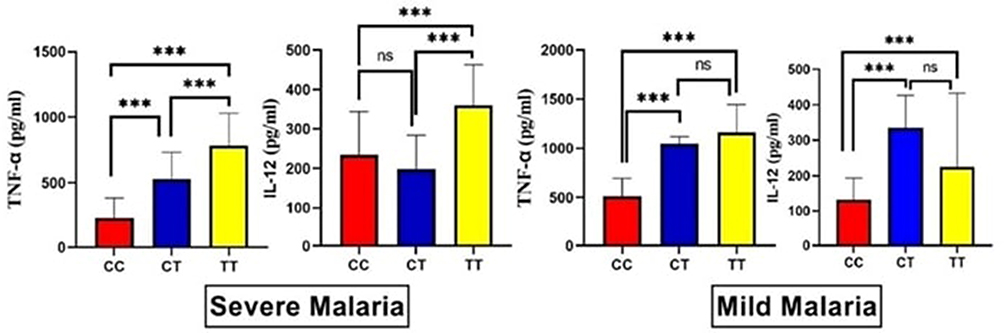 |
Figure 6 The levels of the circulating TNF-alpha and IL-12-malaria among both malaria subgroups in relation to Mal rs8177374 polymorphism. Notes: ns; non-significant; ***statistically significant at p-value ≤ 0.001. |
Discussion
Many countries still have a lot of malaria deaths, even though the disease is now under control. In 2020, there were an estimated 627,000 malaria deaths around the world, with more than 90% of these deaths occurring in sub-Saharan Africa.22 In Egypt, malaria is a well-known endemic infection that has been documented since ancient times.23 According to the WHO, Egypt is one of the countries that has successfully prevented malaria transmission and is currently in the stage of preventing the reappearance of the disease.22
Host genetic influences have been shown to explain approximately one-third of the diversity in the risk of severe or complicated malaria, and there are numerous genetic variations that have been linked to malaria protection.24 Mal (MyD88-adapter-like), alternatively referred to as TIRAP, has been identified as a key adaptor protein in the MYD88-dependent signalling pathway shared by TLR2 and TLR4.25 The genetic diversity in the TLRs signalling system has a significant effect on the disease susceptibility and prognosis.26 Polymorphisms in the MAL gene influence the function of this adaptor protein.1 This polymorphism has been linked to resistance to a variety of infectious illnesses, including tuberculosis (TB), bacterial septicaemia, invasive pneumococcal infection,27 and autoimmune disorders such as systemic lupus erythematosus (SLE) and Behcet’s disease.28
The main objective of this paper was to investigate the effect of the rs8177374 (C/T) polymorphism of the MyD88-adapter-like (MAL) gene on the severity of P. falciparum infection in Egyptian adults. Additionally, the effect of this polymorphism on serum TNF-alpha and IL-12 levels was examined.
The SNP rs8177374 encodes thymine (T) rather than cytosine (C). Inflammatory cytokines are overproduced in the presence of the CC genotype, while the TT genotype is associated with a weakened immunological response, and a balanced immunological response is produced by CT genotype carriers.26,29 Individuals with a balanced immune response are less likely to develop severe kinds of infectious diseases. The CT genotype has been shown to protect against a variety of illnesses, and this protective role of the heterozygous variant could be due to diminished TLR signalling or NF-kB activation.19,26,30 It has also been found that the Mal rs8177374 CT genotype is protective against chronic Chagas cardiomyopathy, as described by Ramasawmy et al.31
The results show that the frequency of Mal rs8177374 CC genotype was higher in patients with severe malaria compared to mild malaria, and the Mal rs8177374 CT genotype and T allele increase the risk of developing malaria. The Mal rs8177374 heterozygous (CT) mutation, homozygous (TT) mutation, and the T allele confer a protective effect against severe malaria. So, according to our findings, subjects with the CT and TT genotypes are unlikely to develop severe disease after being exposed to malaria, probably due to the balanced immunological response caused by this SNP.
In the Iranian Balochi population, the CT genotype has been linked to mild P. falciparum infection.6 Similarly, a study conducted in the Pakistani community indicated that the heterozygous genotype of Mal rs8177374 confers protection against severe malaria illness.1 Khor et al investigated the link between the Mal rs8177374 (S/L) gene polymorphism and pneumococcal illness, bacteraemia, malaria, and tuberculosis in the United Kingdom, Vietnam, and Africa. They demonstrated that Mal rs8177374 heterozygosity is protective against these infections.19 The association of MAL rs8177374 (S/L) heterozygosity with protection against severe malaria was later corroborated in the Indian population.32
In contrast, Esposito et al found no link between the rs8177374 (S/L) mutation in the MAL gene with malaria risk or severity in the Burundian population.33 And a study conducted by Nejentsev et al found that TIRAP (MAL) rs8177374 (S/L) heterozygosity did not have an effect on tuberculosis in the people of Ghana, Russia, and Indonesia.34 These contradictory findings could be explained by differences in ethnic populations with varying genetic backgrounds.
The link between the C allele and disease onset might be explained by enhanced MAL (TIRAP) signalling and activation of the NF-kB pathway.19 So, it could be concluded that the T allele is linked to mild malaria and protects against severe malaria because of a decreased activation of NF-kB. Ramasawmy et al also discovered a link between the C allele and chronic Chagas disease.31 It has been observed that the T allele protects against TB and SLE.30 On the contrary, Naderi et al showed that an increased risk of pulmonary TB is linked to the rs8177374 T allele.35
Several studies have shown that cytokines play a role in determining the severity and outcome of malaria.36–38 The optimal immune response to malaria infection is distinguished by early intensive proinflammatory cytokine-mediated mechanisms that clear the parasite-infected cells, which are then rapidly repressed by anti-inflammatory effectors after parasite replication is under control.39 Toll-like receptor (TLR) polymorphisms have been linked to altered cytokine responses in parasite infection, and a link between TLR9 SNPs and serum levels of the pro-inflammatory cytokine Interferon-gamma (IFN-γ) in children with cerebral malaria has been documented.40
We investigated the effect of MAL rs8177374 (C/T) gene polymorphism on TNF-alpha and IL-12 levels, and this is, to the best of our knowledge, the first study to look at cytokine levels in relation to MAL rs8177374 (C/T) gene variants. Our results revealed that there was a significant elevation of serum levels of TNF-alpha and IL-12 in the malaria group compared to healthy controls. TNF-alpha levels were significantly higher in severe malaria than in mild malaria. In the malaria group, patients with the CT and TT genotypes had significantly higher levels of TNF-alpha compared to patients with the CC genotype. IL-12 levels were significantly lower in severe malaria compared to mild malaria, and patients with the CT and TT genotypes had significantly higher levels of IL-12 compared to patients with the CC genotype.
Moreover, to attribute the causality of whether the MyD88 SNP is linked to the malaria phenotype or the cytokine phenotype, we looked at the association of the MyD88 SNP with the cytokine levels while controlling for the malaria phenotype. Interestingly, we found that the CC genotype was associated with significantly lower levels of TNF-alpha compared to the CT and TT genotypes in both severe and mild malaria subgroups. The TT genotype has the highest level of IL-12 in the severe malaria group, while the CT genotype has the highest level in the mild malaria group.
TNF-alpha was the first parasite-induced cytokine to be identified, and it was induced by the Plasmodium-infected erythrocytes, malarial pigment, and specific glycolipids such as the GPI moiety. TNF-alpha has a dual function in malaria pathogenesis: when present in sufficient quantities, it aids in parasite clearance, yet when present in excessive amounts, it may be detrimental to the outcome of the infection.39,41 Many studies have shown that high plasma TNF-alpha is linked to more severe malaria cases than less severe cases.42,43
Our results provide evidence that in MAL heterozygotes (C/T), the immunological response might be balanced, resulting in optimum TNF-alpha release that protects against severe illness. In agreement with our results, Ferwerda et al found that in vivo administering lipopolysaccharide of gram-negative bacteria in tiny quantities increased inflammatory molecule production in volunteers with Mal/TIRAP 180L (S/L) heterozygous compared to homozygous (S/S).44 Panda et al also found that plasma TNF-alpha levels were greater in malaria patients with heterozygous (S/L) mutants than in wild type (S/S) individuals.32
In the experimental research, IL-12 has been demonstrated to have a role in malaria protection via modulating IFN-γ, TNF-alpha, and nitric oxide responses.45 Additionally, it has been shown that in vivo administration of IL-12 protects monkeys against malaria.46 IL-12 plays a significant role in the adaptive immune response to malaria, increasing T helper 1 (Th1) cell formation, proliferation, and activity.47 The outcome of the infection, such as profound anaemia and other features of malarial pathophysiology, may be affected by host macrophages’ reaction to parasite products, which in turn may affect their ability to produce IL-12. Several studies48–50 found that low levels of IL-12 play a role in the worsening of anaemia and other clinical sequelae and are associated with severe disease, which is consistent with our findings.
In an in vivo study of IL-12-dependent liver damage caused by Plasmodium berghei (NK65) blood-stage, infected wild mice produced high levels of IL-12 and suffered significant liver damage, whereas MyD88-deficient animals produced much lower levels of IL-12 and were immune to liver damage.51 However, studies on IL-12 levels in relation to Mal gene polymorphism are still lacking.
There have been a large number of studies, including Genome-wide association studies (GWAS), searching for variants associated with decreased or increased incidence of malaria or severe malaria, and recent high-profile studies on variants in genes such as ATP2B4 and Piezo1.52–55 But none of these studies identify this or other variants in MyD88 associated with malaria. The likely explanation is that the SNP only has an effect in this specific population (Egyptians), which may have been omitted from previous global studies, or that the allele frequency of the MyD88 SNP may be different in the Egyptian population, which might explain such an observation.
The present study has some limitations like the relatively small sample size with one ethnicity (Egyptians), the G6PD status, blood types, and the levels of parasitaemia which are linked to the outcome in P. falciparum malaria, were not evaluated in the analysis, and finally, the focus of the present study was on MAL rs8177374 (C/T) polymorphism. Perhaps, polymorphisms in MAL upstream components may play a role in infection protection or susceptibility.
Conclusions
There is a link between the Mal rs8177374 (C/T) polymorphism and protection against severe P. falciparum malaria. Additionally, the Mal rs8177374 polymorphism is linked to increased TNF-alpha levels and IL-12 in the blood. Further research with a larger sample size of patients in malaria-endemic parts of the globe with different ethnicities might offer a clearer understanding of the involvement of Mal rs8177374 polymorphism in the outcome of Plasmodium falciparum infection.
Data Sharing Statement
The datasets used and/or analysed during the current study are available from the first author upon reasonable request.
Funding
This research did not receive any specific grant from funding agencies in the public, commercial, or not-for-profit sectors.
Disclosure
The authors declare that they have no conflicts of interest in relation to this work.
References
1. Rani A, Nawaz SK, Irfan S, Arshad M, Bashir R, Shaheen N. Role of MyD88-adaptor-like gene polymorphism rs8177374 in modulation of malaria severity in the Pakistani population. Braz J Infect. 2017;21(4):418–423. doi:10.1016/j.bjid.2017.04.002
2. Arbour NC, Lorenz E, Schutte BC, et al. TLR4 mutations are associated with endotoxin hyporesponsiveness in humans. Nat Genet. 2000;25(2):187–191. doi:10.1038/76048
3. Oakley MS, Gerald N, McCutchan TF, Aravind L, Kumar S. Clinical and molecular aspects of malaria fever. Trends Parasitol. 2011;27(10):442–449. doi:10.1016/j.pt.2011.06.004
4. Gowda DC. TLR-mediated cell signaling by malaria GPIs. Trends Parasitol. 2007;23(12):596–604. doi:10.1016/j.pt.2007.09.003
5. De Mendonça VR, Goncalves MS, Barral-Netto M. The host genetic diversity in malaria infection. J Trop Med. 2012;2012:1–17. doi:10.1155/2012/940616
6. Zakeri S, Pirahmadi S, Mehrizi AA, Djadid ND. Genetic variation of TLR-4, TLR-9 and TIRAP genes in Iranian malaria patients. Malar J. 2011;10(1):1–7. doi:10.1186/1475-2875-10-77
7. Parroche P, Lauw FN, Goutagny N, et al. Malaria hemozoin is immunologically inert but radically enhances innate responses by presenting malaria DNA to Toll-like receptor 9. Proc Natl Acad Sci. 2007;104(6):1919–1924. doi:10.1073/pnas.0608745104
8. Akira S, Takeda K. Toll-like receptor signalling. Nat Rev Immunol. 2004;4(7):499–511. doi:10.1038/nri1391
9. Costa AG, Ramasawmy R, Ibiapina HNS, et al. Association of TLR variants with susceptibility to Plasmodium vivax malaria and parasitemia in the Amazon region of Brazil. PLoS One. 2017;12(8):e0183840. doi:10.1371/journal.pone.0183840
10. O’Neill LA, Bowie AG. The family of five: TIR-domain-containing adaptors in Toll-like receptor signalling. Nat Rev Immunol. 2007;7(5):353–364. doi:10.1038/nri2079
11. Kawai T, Akira S. Toll-like receptors and their crosstalk with other innate receptors in infection and immunity. Immunity. 2011;34(5):637–650. doi:10.1016/j.immuni.2011.05.006
12. Kakkar R, Lee RT. The IL-33/ST2 pathway: therapeutic target and novel biomarker. Nat Rev Drug Discov. 2008;7(10):827–840. doi:10.1038/nrd2660
13. Wesche H, Henzel WJ, Shillinglaw W, Li S, Cao Z. MyD88: an adapter that recruits IRAK to the IL-1 receptor complex. Immunity. 1997;7(6):837–847. doi:10.1016/S1074-7613(00)80402-1
14. Muzio M, Ni J, Feng P, Dixit VM. IRAK (Pelle) family member IRAK-2 and MyD88 as proximal mediators of IL-1 signaling. Science. 1997;278(5343):1612–1615. doi:10.1126/science.278.5343.1612
15. Burns K, Martinon F, Esslinger C, et al. MyD88, an adapter protein involved in interleukin-1 signaling. J Biol Chem. 1998;273(20):12203–12209. doi:10.1074/jbc.273.20.12203
16. Medzhitov R, Preston-Hurlburt P, Kopp E, et al. MyD88 is an adaptor protein in the hToll/IL-1 receptor family signaling pathways. Mol Cell. 1998;2(2):253–258. doi:10.1016/S1097-2765(00)80136-7
17. Guimarães da Costa A, Do Valle Antonelli LR, Augusto Carvalho Costa P, et al. The robust and modulated biomarker network elicited by the Plasmodium vivax infection is mainly mediated by the IL-6/IL-10 axis and is associated with the parasite load. J Immunol Res. 2014;2014. doi:10.1155/2014/318250
18. Belhaouane I, Hoffmann E, Chamaillard M, Brodin P, Machelart A. Paradoxical Roles of the MAL/Tirap Adaptor in Pathologies. Front Immunol. 2020;11:569127. doi:10.3389/fimmu.2020.569127
19. Khor CC, Chapman SJ, Vannberg FO, et al. A Mal functional variant is associated with protection against invasive pneumococcal disease bacteremia, malaria and tuberculosis. Nature Genetics. 2007;39(4):523–528.
20. World Health Organization. Guidelines for the Treatment of Malaria. World Health Organization; 2015.
21. Eticha T, Tamire T, Bati T. Performance evaluation of malaria Pf/Pv Combo test kit at highly malaria-endemic area, Southern Ethiopia: a cross-sectional study. J Trop Med. 2020;2020:1–7. doi:10.1155/2020/1807608
22. World Health Organization. World Malaria Report 2021. World Health Organization; 2021.
23. Nerlich AG, Schraut B, Dittrich S, Jelinek T, Zink AR. Plasmodium falciparum in ancient Egypt. Emerg Infect Dis. 2008;14(8):1317–1319. doi:10.3201/eid1408.080235
24. Kariuki SN, Williams TN. Human genetics and malaria resistance. Hum Genet. 2020;139(6–7):801–811. doi:10.1007/s00439-020-02142-6
25. Yamamoto M, Sato S, Hemmi H, et al. Essential role for TIRAP in activation of the signalling cascade shared by TLR2 and TLR4. Nature. 2002;420(6913):324–329. doi:10.1038/nature01182
26. Mansell A, Brint E, Gould JA, O’Neill LA, Hertzog PJ. Mal interacts with tumor necrosis factor receptor-associated factor (TRAF)-6 to mediate NF-κB activation by Toll-like receptor (TLR)-2 and TLR4. J Biol Chem. 2004;279(36):37227–37230. doi:10.1074/jbc.C400289200
27. Jenkins KA, Mansell A. TIR-containing adaptors in Toll-like receptor signalling. Cytokine. 2010;49(3):237–244. doi:10.1016/j.cyto.2009.01.009
28. Durrani O, Banahan K, Sheedy FJ, et al. TIRAP Ser180Leu polymorphism is associated with Behcet’s disease. Rheumatology. 2011;50(10):1760–1765. doi:10.1093/rheumatology/ker200
29. Miller LH, Baruch DI, Marsh K, Doumbo OK. The pathogenic basis of malaria. Nature. 2002;415(6872):673–679. doi:10.1038/415673a
30. Castiblanco J, Varela D-C, Castaño-Rodríguez N, Rojas-Villarraga A, Hincapié M-E, Anaya J-M. TIRAP (MAL) S180L polymorphism is a common protective factor against developing tuberculosis and systemic lupus erythematosus. Infect Genet Evol. 2008;8(5):541–544. doi:10.1016/j.meegid.2008.03.001
31. Ramasawmy R, Cunha‐Neto E, Fae KC, et al. Heterozygosity for the S180L variant of MAL/TIRAP, a gene expressing an adaptor protein in the Toll‐like receptor pathway, is associated with lower risk of developing chronic Chagas cardiomyopathy. J Infect Dis. 2009;199(12):1838–1845. doi:10.1086/599212
32. Panda AK, Das BK, Panda A, et al. Heterozygous mutants of TIRAP (S180L) polymorphism protect adult patients with Plasmodium falciparum infection against severe disease and mortality. Infect Genet Evol. 2016;43:146–150. doi:10.1016/j.meegid.2016.04.035
33. Esposito S, Molteni CG, Zampiero A, et al. Role of polymorphisms of toll-like receptor (TLR) 4, TLR9, toll-interleukin 1 receptor domain containing adaptor protein (TIRAP) and FCGR2A genes in malaria susceptibility and severity in Burundian children. Malar J. 2012;11(1):196. doi:10.1186/1475-2875-11-196
34. Nejentsev S, Thye T, Szeszko JS, et al. Analysis of association of the TIRAP (MAL) S180L variant and tuberculosis in three populations. Nat Genet. 2008;40(3):262–263. doi:10.1038/ng0308-261
35. Naderi M, Hashemi M, Pourmontaseri Z, Eskandari-Nasab E, Bahari G, Taheri M. TIRAP rs8177374 gene polymorphism increased the risk of pulmonary tuberculosis in Zahedan, southeast Iran. Asian Pac J Trop Med. 2014;7(6):451–455. doi:10.1016/S1995-7645(14)60073-0
36. Day N, Hien T, Schollaardt T, et al. The prognostic and pathophysiologic role of pro- and antiinflammatory cytokines in severe malaria. J Infect Dis. 1999;180(4):1288–1297. doi:10.1086/315016
37. Gogos CA, Drosou E, Bassaris HP, Skoutelis A. Pro- versus anti-inflammatory cytokine profile in patients with severe sepsis: a marker for prognosis and future therapeutic options. J Infect Dis. 2000;181(1):176–180. doi:10.1086/315214
38. Malaguarnera L, Pignatelli S, Musumeci M, Simporè J, Musumeci S. Plasma levels of interleukin-18 and interleukin-12 in Plasmodium falciparum malaria. Parasite Immunol. 2002;24(9–10):489–492. doi:10.1046/j.1365-3024.2002.00485.x
39. Artavanis-Tsakonas K, Tongren J, Riley E. The war between the malaria parasite and the immune system: immunity, immunoregulation and immunopathology. Clin Exp Immunol. 2003;133(2):145–152. doi:10.1046/j.1365-2249.2003.02174.x
40. Sam-Agudu NA, Greene JA, Opoka RO, et al. TLR9 polymorphisms are associated with altered IFN-gamma levels in children with cerebral malaria. Am J Trop Med Hyg. 2010;82(4):548–555. doi:10.4269/ajtmh.2010.09-0467
41. Boeuf PS, Loizon S, Awandare GA, et al. Insights into deregulated TNF and IL-10 production in malaria: implications for understanding severe malarial anaemia. Malar J. 2012;11(1):1–9. doi:10.1186/1475-2875-11-253
42. Kwiatkowski D, Hill AV, Sambou I, et al. TNF concentration in fatal cerebral non-fatal cerebral, and uncomplicated Plasmodium falciparum malaria. Lancet. 1990;336(8725):1201–1204.
43. Wellems TE, Hayton K, Fairhurst RM. The impact of malaria parasitism: from corpuscles to communities. J Clin Invest. 2009;119(9):2496–2505. doi:10.1172/JCI38307
44. Ferwerda B, Alonso S, Banahan K, et al. Functional and genetic evidence that the Mal/TIRAP allele variant 180L has been selected by providing protection against septic shock. Proc Natl Acad Sci. 2009;106(25):10272–10277. doi:10.1073/pnas.0811273106
45. Stevenson MM, Tam MF, Wolf SF, Sher A. IL-12-induced protection against blood-stage Plasmodium chabaudi AS requires IFN-gamma and TNF-alpha and occurs via a nitric oxide-dependent mechanism. J Immunol. 1995;155(5):2545–2556.
46. Hoffman SL, Crutcher JM, Puri SK, et al. Sterile protection of monkeys against malaria after administration of interleukin-12. Nat Med. 1997;3(1):80–83. doi:10.1038/nm0197-80
47. Trinchieri G. Interleukin-12 and its role in the generation of TH1 cells. Immunol Today. 1993;14(7):335–338. doi:10.1016/0167-5699(93)90230-I
48. Luty AJ, Perkins DJ, Lell B, et al. Low interleukin-12 activity in severe Plasmodium falciparum malaria. Infect Immun. 2000;68(7):3909–3915. doi:10.1128/IAI.68.7.3909-3915.2000
49. Perkins DJ, Weinberg JB, Kremsner PG. Reduced interleukin-12 and transforming growth factor—Β 1 in severe childhood malaria: relationship of cytokine balance with disease severity. J Infect Dis. 2000;182(3):988–992. doi:10.1086/315762
50. Malaguarnera L, Imbesi RM, Pignatelli S, Simpore J, Malaguarnera M, Musumeci S. Increased levels of interleukin-12 in Plasmodium falciparum malaria: correlation with the severity of disease. Parasite Immunol. 2002;24(7):387–389. doi:10.1046/j.1365-3024.2002.00478.x
51. Adachi K, Tsutsui H, Kashiwamura S, et al. Plasmodium berghei infection in mice induces liver injury by an IL-12- and toll-like receptor/myeloid differentiation factor 88-dependent mechanism. J Immunol. 2001;167(10):5928–5934.
52. Damena D, Denis A, Golassa L, Chimusa ER. Genome-wide association studies of severe P. falciparum malaria susceptibility: progress, pitfalls and prospects. BMC Med Genomics. 2019;12(1):120. doi:10.1186/s12920-019-0564-x
53. Ahouidi A, Ali M, Almagro-Garcia J, et al. An open dataset of Plasmodium falciparum genome variation in 7000 worldwide samples. Wellcome Open Res. 2021;6:42. doi:10.12688/wellcomeopenres.16168.2
54. Kwiatkowski DP. How malaria has affected the human genome and what human genetics can teach us about malaria. Am J Hum Genetics. 2005;77(2):171–192. doi:10.1086/432519
55. Nguetse CN, Purington N, Ebel ER, et al. A common polymorphism in the mechanosensitive ion channel PIEZO1 is associated with protection from severe malaria in humans. Proc Natl Acad Sci U S A. 2020;117(16):9074–9081. doi:10.1073/pnas.1919843117
 © 2022 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2022 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.