Back to Journals » Infection and Drug Resistance » Volume 17
Persistent Colonization of Ciprofloxacin-Resistant and Extended-Spectrum β-Lactamase (ESBL)-Producing Salmonella enterica Serovar Kentucky ST198 in a Patient with Inflammatory Bowel Disease
Authors Jiang Y, Yang H, Wang ZY, Lin DC, Jiao X ![]() , Hu Y, Wang J
, Hu Y, Wang J
Received 15 November 2023
Accepted for publication 1 April 2024
Published 12 April 2024 Volume 2024:17 Pages 1459—1466
DOI https://doi.org/10.2147/IDR.S447971
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Zhi Ruan
Yue Jiang,1,2,* Huilin Yang,3,* Zhen-Yu Wang,1,2 Da-Chuan Lin,4 Xinan Jiao,1,2 Yunlong Hu,3,4 Jing Wang1,2
1Jiangsu Key Laboratory of Zoonosis, Yangzhou University, Yangzhou, 225009, People’s Republic of China; 2Key Laboratory of Prevention and Control of Biological Hazard Factors (Animal Origin) for Agrifood Safety and Quality, Ministry of Agriculture of China, Yangzhou University, Yangzhou, 225009, People’s Republic of China; 3Department of Clinical Laboratory, Peking University Shenzhen Hospital, Shenzhen, 518036, People’s Republic of China; 4Guangdong Key Laboratory of Regional Immunity and Diseases, Shenzhen University School of Medicine, Shenzhen University, Shenzhen, 518060, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Yunlong Hu; Jing Wang, Email [email protected]; [email protected]
Objective: Salmonella enterica serovar Kentucky ST198 has emerged as a global threat to humans. In this study, we aimed to characterize the prolonged carriage of ciprofloxacin-resistant and extended-spectrum β-lactamase (ESBL)-producing S. Kentucky ST198 in a single patient with inflammatory bowel disease (IBD).
Methods: Three S. Kentucky strains were collected from a single patient with IBD on 11th January, 23rd January, and 8th February, 2022, respectively. Antimicrobial susceptibility testing, whole-genome sequencing, and phylogenetic analysis with 38 previously described Chinese S. Kentucky ST198 strains from patients and food were performed.
Results: All three S. Kentucky isolates belonged to ST198. They carried identical 16 resistance genes, such as blaCTX-M-55, tet(A), and qnrS1, and had identical mutations within gyrA (S83F and D87N) and parC (S80I). Therefore, they exhibited identical multidrug-resistant profiles, including the clinically important antibiotics cephalosporins (ceftazidime and cefepime), fluoroquinolones (ciprofloxacin and levofloxacin), and third-generation tetracycline (tigecycline). Our three S. Kentucky strains were classified into the subclade ST198.2– 2, and were genetically identical (2– 6 SNPs) to each other. They exhibited a close genetic similarity (15– 20 SNPs) to the isolate NT-h3189 from a patient and AH19MCS1 from chicken meat in China, indicating a possible epidemiological link between these S. Kentucky ST198 isolates from the patients and chicken meat.
Conclusion: Long-term colonization of ciprofloxacin-resistant and ESBL-producing S. Kentucky ST198 in a single patient is a matter of concern. Due to the potential transfer of S. Kentucky ST198 from food sources to humans, ongoing surveillance of this particular clone in animals, animal-derived food products, and humans should be strengthened.
Keywords: Salmonella, MDR, Kentucky, persistent colonization
Introduction
Salmonella is an important cause of food-borne disease in humans. In Europe, a total of 65,208 cases of salmonellosis in 2022, making it the second most common zoonotic pathogen in the European Union.1 Different types of animals, particularly poultry, pigs, and cattle, can serve as reservoirs for Salmonella.1 The consumption of contaminated food is a common route of infection for humans.1 Salmonellosis outbreaks linked to food have been previously reported in multiple countries with a variety of Salmonella serotypes. For example, multistate outbreak of Salmonella enterica serovar Thompson linked to the consumption of seafood was reported in the United States.2 Multiple countries have been affected by an outbreak of S. Typhimurium linked to chocolate products from Belgium in 2022.3 The rapid emergence and dissemination of antimicrobial resistance in Salmonella spp. from human cases pose a significant threat to clinical therapy. According to the World Health Organization (WHO), third-generation cephalosporin-resistant Enterobacteriaceae and fluoroquinolone-resistant Salmonella are classified as critical and high-priority pathogens, respectively. There is an urgent need for research and development of new antibiotics.4
Salmonella enterica serovar Kentucky is a common nontyphoid Salmonella serotype that could infect a variety of hosts, including humans.1 S. Kentucky has multiple sequence types (STs), such as ST198, ST314, and ST521.3 Among them, S. Kentucky ST198 is becoming a globally prevalent clone that caused human clinical infections and also frequently detected in various sources (eg, livestock, animal-derived food products, vegetable, the environment).5–10 Recently, there has been a growing prevalence of ciprofloxacin- and third-generation cephalosporin-resistant S. Kentucky ST198 strains worldwide.6,7,10–12
Inflammatory bowel diseases (IBDs) are chronic inflammatory diseases of the gastrointestinal tract characterized by a relapsing and remitting course, with Crohn’s disease and ulcerative colitis being the most common clinical manifestations.13,14 Although the etiology of IBD is not well defined, environmental, genetic, and dietary risk factors influence or contribute to the development and severity of this disease.15,16 Enteric infections with pathogens such as Salmonella or Campylobacter have been suggested to promote the onset of IBD.15,16
In this study, we characterized three ciprofloxacin-resistant and extended-spectrum β-lactamase (ESBL)-producing S. Kentucky ST198 strains from a single patient with IBD, to highlight the prolonged carriage of S. Kentucky ST198 in a single patient, and their potential epidemiological link to such organisms from chicken meat.
Materials and Methods
Patient and Bacterial Isolates
A 28-year-old female patient was diagnosed with IBD in February 2019 at a tertiary hospital in Shenzhen, China, and experienced symptoms including diarrhea and rectal bleeding. The symptoms were alleviated by the administration of prednisone and mesalazine. In July 2021, the patient was hospitalized because of diarrhea, rectal bleeding, vomiting, and low-grade fever. Salmonella infection was detected and the symptoms improved following treatment with antibiotics and prednisone. The patient was discharged in September 2021 and prescribed a regimen of mesalazine (3 g/d) and azathioprine (50 mg/d) for ongoing treatment. Subsequent follow-up visits were conducted in October and November 2021, as well as in January and February 2022.
Three S. Kentucky strains (designated as SZ22HS1 to SZ22HS3) were isolated from three fecal samples collected from this patient on 11th January, 23rd January, and 8th February, 2022, respectively. Stool samples were cultured on Salmonella Shigella agar and then identified using matrix-assisted laser desorption/ionization time-of-flight mass spectrometry. The serotype was determined through the conventional slide agglutination assay with the somatic (O) and flagellar (H) antigens according to the White-Kauffmann scheme.17
Antimicrobial Susceptibility Testing
Three Salmonella isolates were tested for their susceptibility to 14 antimicrobial agents by the broth microdilution method using BD PhoenixTM susceptibility panels, including ceftazidime, cefepime, aztreonam, ticarcillin/clavulanic acid, piperacillin/tazobactam, imipenem, meropenem, ciprofloxacin, levofloxacin, doxycycline, minocycline, tigecycline, colistin, and sulfamethoxazole/trimethoprim. The results were interpreted according to the standards provided by the Clinical and Laboratory Standards Institute (CLSI) M100, 32nd edition.18 The interpretation of tigecycline (>0.5 mg/L) was based on the clinical breakpoints for Enterobacterales established by the European Committee on Antimicrobial Susceptibility Testing (EUCAST) (https://www.eucast.org/).
Whole-Genome Sequencing and Analysis
The genomic DNA of three S. Kentucky isolates was extracted using the TIANamp Bacterial DNA kit (Tiangen Biotech, Beijing, China), following the manufacturer’s instructions. Sequence libraries were prepared using the NEB Next® UltraTM DNA Library Prep Kit for Illumina (NEB, USA), and subsequently subjected to genome sequencing on the Illumina HiSeq 2500 platform. The resulting raw reads with less than 90% Q30 bases were trimmed and filtered using the NGSQC Toolkit (v2.3.3), and de novo genome assembly of 150-bp paired-end reads was performed using SPAdes v.3.8.2. The draft genomes have been deposited in GenBank under accession numbers JAVCBR000000000, JAVCBS000000000, and JAVCBT000000000. The genomic sequences were analyzed using MLST 2.0 (https://cge.food.dtu.dk/services/MLST/) to identify the sequence types. Additionally, the identification of resistance genes and chromosomal point mutations was performed using ResFinder and PointFinder, respectively (http://genepi.food.dtu.dk/resfinder). Plasmid replicons with >95% sequence identity and >60% coverage were identified using PlasmidFinder (https://cge.food.dtu.dk/services/PlasmidFinder/).
Phylogenetic Analysis
The genome sequences of 38 S. Kentucky ST198 strains from China were downloaded from the NCBI database (Table S1). This dataset consists of 27 strains collected from patients and food products in China from 2013 to 2017,7 one isolate from a patient in 2022,19 and our previously described ten strains from chicken meat in 2019.11 To analyze their genetic relatedness, a phylogenetic tree was constructed using Parsnp (https://harvest.readthedocs.io/en/latest/content/parsnp.html), based on core genome single-nucleotide polymorphisms (cgSNPs). The genome of S. Kentucky strain AH19MCS1 (accession number CP102719.1) was used as a reference for comparison. The bacterial source tracking analysis of S. Kentucky strains in this study was performed using BacWGSTdb webserver.20 The resulting tree and heatmap of the SNP count matrix were visualized and edited using Chiplot (https://www.chiplot.online/).
Results
Analysis of Antimicrobial Susceptibility and Resistance Determinants
The results of antimicrobial susceptibility testing showed that three S. Kentucky isolates were resistant to ceftazidime, cefepime, aztreonam, ciprofloxacin, levofloxacin, doxycycline, minocycline, tigecycline, and sulfamethoxazole/trimethoprim, and intermediate to ticarcillin/clavulanic acid, but susceptible to piperacillin/tazobactam, imipenem, meropenem, and colistin (Tables 1 and S2). In accordance with their multidrug-resistant (MDR) profiles, three S. Kentucky strains possessed 16 resistance genes. These genes confer resistance to various antibiotics, including β-lactams (blaCTX-M-55 and blaTEM-1b), aminoglycosides [aadA7, aadA17, aph(3’)-Ia, aac(3)-Id, aac(3)-IId, and aac(6’)-Iaa], tetracyclines [tet(A)], phenicols (floR), quinolones (qnrS1), sulfonamide/trimethoprim (sul1 and dfrA14), lincosamide [lnu(F)], macrolides [mph(A)], and rifampicin (arr-2) (Table 1). The presence of the PMQR gene qnrS1 and mutations within gyrA (S83F and D87N) and parC (S80I) could explain their resistance to fluoroquinolone (ciprofloxacin and levofloxacin) (Table 1).
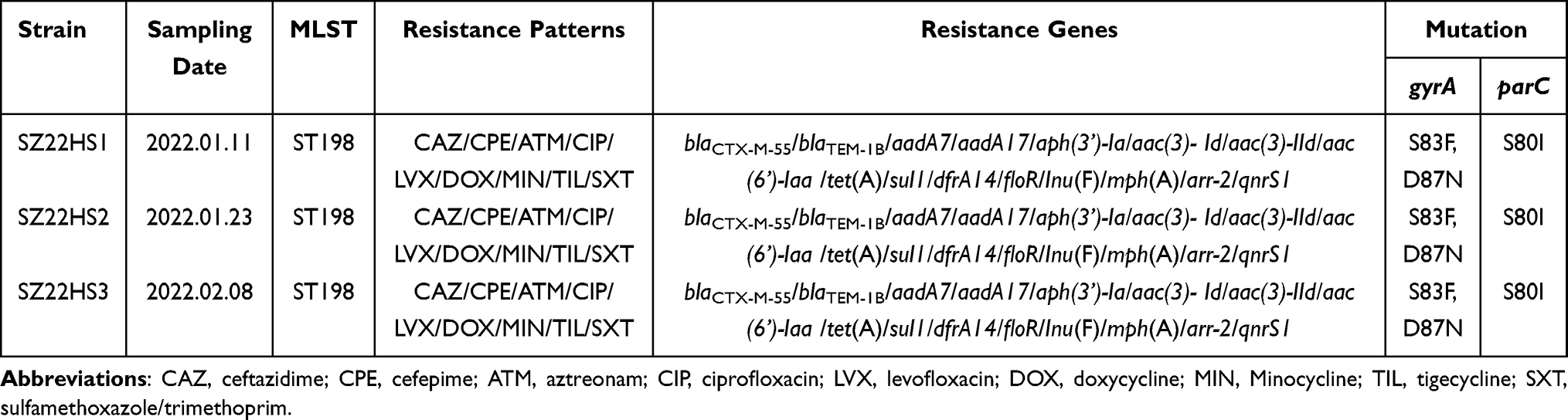 |
Table 1 Characterizations of Salmonella Kentucky Isolates from a Patient in This Study |
Intriguingly, all three S. Kentucky strains in this study exhibited resistance to tigecycline, with a minimum inhibitory concentration (MIC) of 8 mg/L. However, no tigecycline resistance genes [tet(X) and tmexCD-toprJ] or any alterations in the amino acid sequences of AcrAB-TolC, RamA, RamR, MarA, and MarR were identified in these strains. Nevertheless, these S. Kentucky isolates harbored the tetracycline resistance gene tet(A) variant, which was possibly linked to their tigecycline resistance.
Phylogenomic Analysis of S. Kentucky ST198 Strains
Genome sequencing analysis revealed that all three S. Kentucky isolates (SZ22HS1, SZ22HS2, and SZ22HS3) belonged to sequence type 198 (ST198). The genome sizes of SZ22HS1, SZ22HS2, and SZ22HS3 were 4,806,845 bp with a GC content of 52.21%, 4,804,955 bp with a GC content of 52.21%, and 4,841,148 bp with a GC content of 52.14%, respectively (Figure S1). Furthermore, no known plasmid replicons were identified in these three S. Kentucky strains. In order to assess the genetic variations among three S. Kentucky ST198 isolates in this study and additional 38 Chinese S. Kentucky ST198 strains, we constructed a phylogenetic tree using cgSNPs analysis (Figure 1). The findings demonstrated a relatively close genetic relationship (2–6 SNPs) among three S. Kentucky strains (SZ22HS1, SZ22HS2, and SZ22HS3) derived from a single patient in this study (Figure 2). Additionally, the bacterial source tracking analysis based on cgMLST showed that three S. Kentucky strains in this study were close to multiple S. Kentucky strains from humans, water, poultry tissue, and chicken carcass from the United Kingdom, USA, Nigeria, and Egypt.
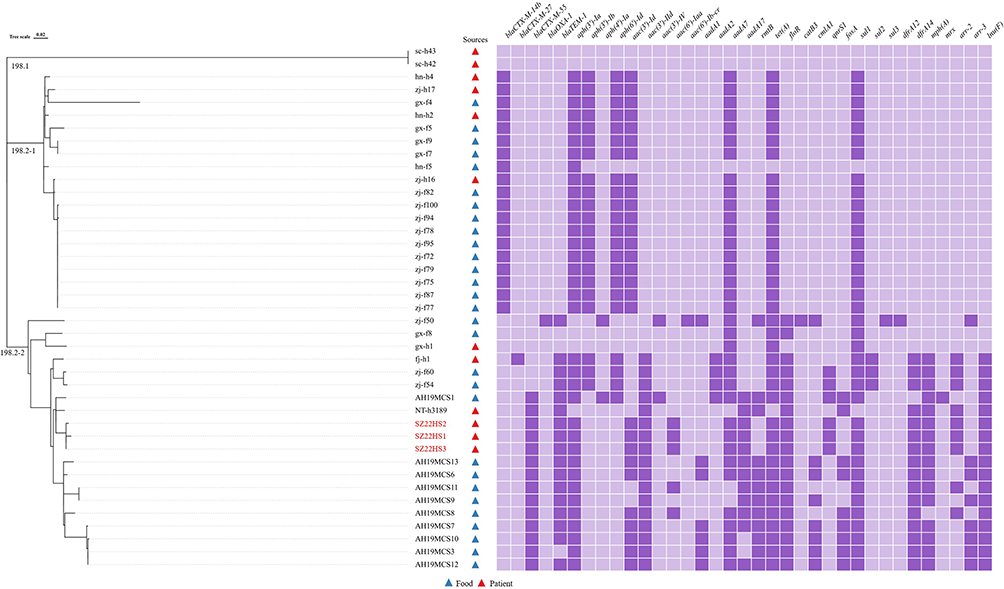 |
Figure 1 Maximum likelihood tree of S. Kentucky ST198 isolates in this study compared with 38 S. Kentucky ST198 isolates from China based on cgSNP analysis. The dark purple rectangle indicates the presence of this gene in the corresponding isolate, while the light purple rectangle represents its absence in the corresponding isolate. Antibiotic resistance genes with >95% sequence identity and >60% coverage are shown. The isolates obtained in this study were indicated in red. |
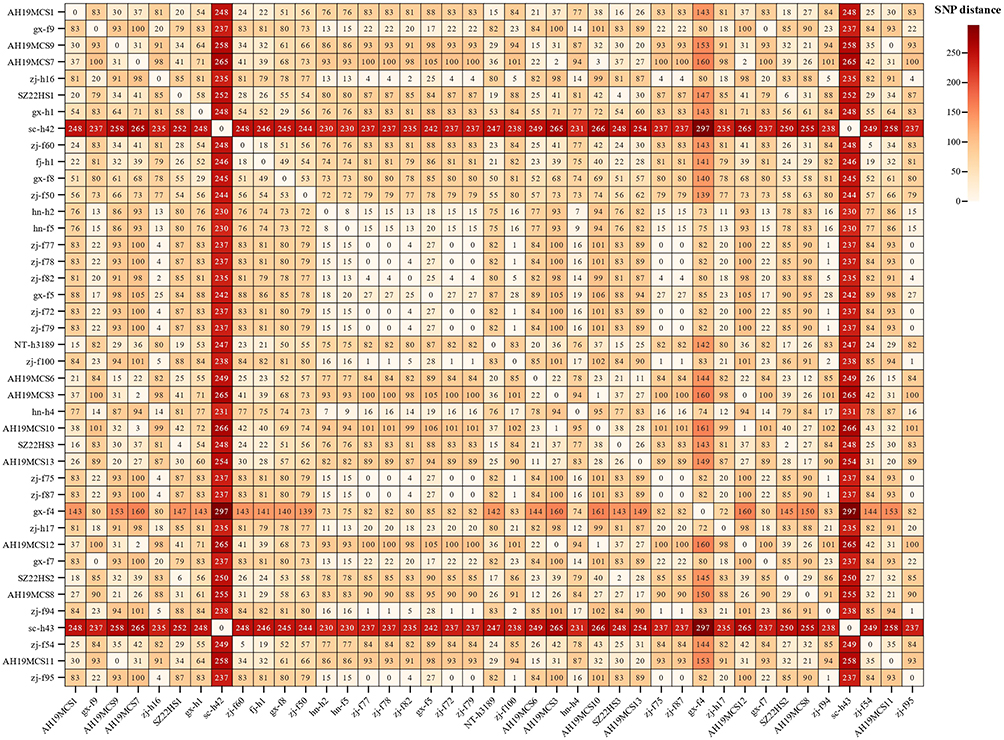 |
Figure 2 Pairwise SNP matrix of S. Kentucky ST198 isolates in this study compared with 38 S. Kentucky ST198 isolates from China. |
As shown in Figure 1, 41S. Kentucky ST198 strains were divided into two clades, namely ST198.1 and ST198.2. With the exception of two isolates (sc-h42 and sc-h43) collected from patients in Sichuan Province, China, in 2013, which lacked any resistance genes and were assigned to the clade ST198.1, the remaining 39 isolates were grouped into ST198.2. The clade ST198.2 was further divided into two subclades, namely ST198.2–1 and ST198.2–2. Within the subclade ST198.2–1, a total of 19 isolates were identified with 18 of them carrying the same resistance genes. The subclade 198.2–2 consisted of six isolates from humans and 14 isolates from animal-derived food products with close genetic relatedness (Figure 2). Our three S. Kentucky ST198 strains (SZ22HS1, SZ22HS2, and SZ22HS3) in the subclade ST198.2–2 exhibited a close genetic similarity to the isolates NT-h3189 from a patient in Nantong, Jiangsu Province, and AH19MCS1 from chicken meat in Anhui Province, differing by 15 to 20 SNPs (Figure 2). It indicates a possible epidemiological link between these S. Kentucky ST198 isolates from the patients and chicken meat.
Discussion
Many bacteria have the ability to establish long-term infections within their hosts despite antibiotic treatment, which can be attributed to several factors, including compromised immune system of the host, immune escape of pathogens, and/or inadequate eradication of pathogens by antibiotics.21 Although rare, previous studies have reported cases of long-term persistence of S. Kentucky in patients. Hawkey et al collected three S. Kentucky ST198 strains from a single patient who was infected in Egypt over a span of three consecutive years (2009–2011). These strains shared a most recent common ancestor that can be traced back to around 2005, indicating a prolonged colonization period of approximately six years.6 In a study conducted in Switzerland, it was observed that out of 156 patients infected with S. Kentucky between 2010 and 2020, 16 patients (10.3%) exhibited persistent carriage of the bacteria for more than three weeks, and four of them were identified as chronic carriers, with the bacteria persisting for over 12 months.12 Three S. Kentucky strains collected from a single patient in this study share identical resistance genes and display a high degree of similarity with only minor cgSNP differences. It suggests that these strains are not only genetically identical but are likely to be duplicates. According to the available medical records, the patient experienced a Salmonella infection in July 2021. Unfortunately, due to the patient being transferred from a different medical facility, we did not obtain the Salmonella strain and are unsure of the specific serotype. At least, this particular clone successfully colonized in the patient’s intestinal tract for a duration exceeding four weeks.
These S. Kentucky strains detected in this patient contained numerous resistance genes and exhibited MDR phenotype, which may be one of the reasons for the failure of antimicrobial therapy. For instance, they carried resistance genes aadA7, sul1, tet(A), and blaTEM-1, which were known to be associated with Salmonella genomic island 1 (SGI-1) in S. Kentucky ST198 strains.10,11 Multiple resistance genes such as blaTEM-1b, aph(3’)-Ia, aadA17, aac(3)-IId, floR, qnrS1, dfrA14, lnu(F), mph(A), and arr-2 were previously detected within the multiresistance region on the chromosomes in S. Kentucky ST198 strains.10,11 Additionally, increasing dynamics were observed in several resistance genes including blaTEM-1b, aph(3’)-Ia, floR, and dfrA14 in Salmonella from China.22
Cephalosporins and fluoroquinolones are the first-line antimicrobials for the treatment of nontyphoidal Salmonella infections.23 However, our three S. Kentucky ST198 strains exhibited resistance to cephalosporins (ceftazidime and cefepime) and fluoroquinolones (ciprofloxacin and levofloxacin). This resistance significantly restricts the available treatment options, thereby impacting the effectiveness of treatment outcomes. In recent years, extended-spectrum β-lactamases (ESBLs)-producing S. Kentucky ST198 encoding by CTX-M-14b has been increasingly reported in Europe and China.7,9,10,24,25 Interestingly, the blaCTX-M-14b gene located within the chromosome was exclusively found in Chinese and European genomes within clade 198.2–1.25 However, the blaCTX-M-14b gene carried by an IncHI2 plasmid was previously described in S. Kentucky in Zimbabwe.26 Our three S. Kentucky strains contained the blaCTX-M-55 gene, which is frequently reported in both animals and humans in China27 and has been sporadically reported in subclade ST198.2–2 from China.10,11 Mutations in gyrA and parC genes and the presence of PMQR genes (eg, qnrS1) have been the primary causes of fluoroquinolone resistance in Salmonella, including S. Kentucky ST198 clone.7,10
Tigecycline is listed as a critically important antimicrobial for human medicine by the World Health Organization due to its effectiveness in treating MDR Gram-negative bacteria.23 Thus, resistance to tigecycline has become a global concern. Previous studies have identified the tet(A) variant as the reason for tigecycline resistance in Salmonella, Klebsiella pneumoniae, and Enterobacter hormaechei.24,28,29 A previous study found that most of the S. Kentucky ST198.2 isolates carried the tet(A) variant and exhibited resistance to tigecycline.10 Similarly, our three S. Kentucky ST198.2 isolates also displayed tigecycline resistance, potentially due to the presence of the tet(A) variant. Evidently, the tet(A) variant is prevalent among S. Kentucky ST198.2 strains and plays a significant role in the emergence and spread of tigecycline resistance in this particular subclade.
Rifampicin is an important first-line antibiotic for the treatment of tuberculosis.30 It also has clinical efficacy against Staphylococcus aureus, Streptococcus pneumoniae, Haemophilus influenzae, Neisseria, and some other bacteria.31 According to CLSI M100, 32th edition, Enterobacteriaceae are intrinsically resistant to rifampicin.18 However, Mumbo et al reported that rifampicin resistance was observed in 11.1% of Salmonella isolates, and clinical efficacy was also observed in Salmonella Typhi and E. coli.31,32 Furthermore, Salmonella was previously used as a model organism to investigate rifampicin resistance.33–35 The wild-type Salmonella enterica exhibited MIC against rifampicin was 12 mg/L, whereas its rifampicin-resistant mutants showed high rifampicin resistance with MICs of 1500 to >3000 mg/L.33 In this study, three S. Kentucky strains carried rifampicin resistance gene arr-2, which was previously found in 51.85% (56/108) of S. Kentucky ST198 isolates.10 Although the proportion of rifampicin resistance gene arr-3 has an increased trend over time in both human and non-human origins in 1713 Salmonella isolates from China between 2006 and 2019,22 it was only detected in one S. Kentucky ST198 strains.10
In recent years, S. Kentucky has become a common serotype causing human nontyphoidal salmonellosis.1 Significantly, MDR S. Kentucky ST198 strains have been increasingly reported in poultry, poultry products, and humans worldwide, including China.6–12,19,24,25 It has become a global epidemic clone, and poses a potential threat to both human and animal well-being. Furthermore, S. Kentucky ST198 has recently differentiated into two distinct clades, namely ST198.1 and ST198.2, with the latter being more prevalent on a global scale.10,25 Consistent with prior research,6,7,10,25 the ST198.2 clade analyzed in this study was further classified into two subclades ST198.2–1 and ST198.2–2. The close genetic relationship observed among isolates from patients and animal-derived food products within subclade ST198.2–2, particularly a strong correlation between our three patient-source isolates and an isolate originating from chicken meat, indicates the potential transfer of S. Kentucky ST198 from food sources to humans. Poultry and poultry productions are recognized to be the main reservoir of MDR S. Kentucky ST198 causing human infections.10 It should be a matter of concern in the monitoring and prevention of human salmonellosis in China.
However, our study has several limitations. As an important foodborne pathogen, humans generally become infected with Salmonella by eating poorly cooked, contaminated food.1 Our study provides a few information and insights into Salmonellosis surveillance since we only collected three S. Kentucky isolates from one patient with IBD within four weeks. Although poultry production was the main reservoir of ciprofloxacin- and tigecycline-resistant extended-spectrum β-lactamase-producing S. Kentucky ST198.2–2 causing human infections in China,10 and a possible epidemiological link between S. Kentucky strains from the patient and chicken meat was observed in the current study, the precise source of S. Kentucky strains colonized this patient is unclear. Broader and detailed studies involving more patients and strains are needed to surveillance the colonization of S. Kentucky ST198 in patients and to elucidate the transmission mechanism of this organism from animals to patients via the food chain. Additionally, it is crucial to perform long-term monitoring and evaluate the efficacy of current therapy for the eradication of this MDR S. Kentucky clone in this patient with IBD.
In conclusion, the prolonged colonization of ciprofloxacin-resistant and ESBL-producing S. Kentucky ST198 in a single patient is of concern. It is recommended to conduct ongoing surveillance of this particular clone in humans, as well as poultry and poultry products in China. More importantly, exploring effective strategies such as phage therapy for bacterial decolonization in humans is urgently needed to prevent hosts from developing infections and reduce the spread of MDR organisms.36
Ethics Statement
This research was reviewed and approved by Yangzhou University. The patient was informed and consented to participate in this study, and the patient gave informed consent to publish the case details. This study complies with the Declaration of Helsinki.
Funding
This work was supported by the fifth phase of “333 project” scientific research project in Jiangsu Province (BRA2020002), the Priority Academic Program Development of Jiangsu Higher Education Institutions (PAPD), and the 111 Project (D18007).
Disclosure
The authors report no conflicts of interest in this work.
References
1. EFSA and ECDC (European Food Safety Authority and European Centre for Disease Prevention and Control). The European Union One Health 2022 Zoonoses Report. EFSA J. 2023;21(12):e8442. doi:10.2903/j.efsa.2023.8442
2. Shen AQ, Dalen A, Bankers L, et al. Multistate outbreak of Salmonella Thompson infections linked to seafood exposure - United States, 2021. MMWR Morb Mortal Wkly Rep. 2023;72(19):513–516. doi:10.15585/mmwr.mm7219a2
3. Samarasekera U. Salmonella Typhimurium outbreak linked to chocolate. Lancet Infect Dis. 2022;22(7):947. doi:10.1016/S1473-3099(22)00351-6
4. World Health Organization. Prioritization of Pathogens to Guide Discovery, Research and Development of New Antibiotics for Drug-Resistant Bacterial Infections, Including Tuberculosis. Geneva, Switzerland: World Health Organization; 2017.
5. Haley BJ, Kim SW, Haendiges J, et al. Salmonella enterica serovar Kentucky recovered from human clinical cases in Maryland, USA (2011-2015). Zoonoses Public Health. 2019;66(4):382–392. doi:10.1111/zph.12571
6. Hawkey J, Le Hello S, Doublet B, et al. Global phylogenomics of multidrug-resistant Salmonella enterica serotype Kentucky ST198. Microb Genom. 2019;5(7):e000269. doi:10.1099/mgen.0.000269
7. Chen H, Song J, Zeng X, et al. National prevalence of Salmonella enterica serotype Kentucky ST198 with high-level resistance to ciprofloxacin and extended-spectrum cephalosporins in China, 2013 to 2017. mSystems. 2021;6(1):e00935–20. doi:10.1128/mSystems.00935-20
8. Saraiva MMS, Benevides VP, da Silva NMV, et al. Genomic and evolutionary analysis of Salmonella enterica serovar Kentucky sequence type 198 isolated from livestock in East Africa. Front Cell Infect Microbiol. 2022;12:772829. doi:10.3389/fcimb.2022.772829
9. Coipan CE, Westrell T, van Hoek AHAM, et al. Genomic epidemiology of emerging ESBL-producing Salmonella Kentucky blaCTX-M-14b in Europe. Emerg Microbes Infect. 2020;9(1):2124–2135. doi:10.1080/22221751.2020.1821582
10. Wang Z, Jiang Y, Xu H, et al. Poultry production as the main reservoir of ciprofloxacin- and tigecycline-resistant extended-spectrum β-lactamase (ESBL)-producing Salmonella enterica serovar Kentucky ST198.2-2 causing human infections in China. Appl Environ Microbiol. 2023;89(9):e0094423. doi:10.1128/aem.00944-23
11. Jiang Y, Wang ZY, Li QC, et al. Characterization of extensively drug-resistant Salmonella enterica serovar Kentucky sequence type 198 isolates from chicken meat products in Xuancheng, China. Microbiol Spectr. 2023;11(2):e0321922. doi:10.1128/spectrum.03219-22
12. Biggel M, Horlbog J, Nüesch-Inderbinen M, et al. Epidemiological links and antimicrobial resistance of clinical Salmonella enterica ST198 isolates: a nationwide microbial population genomic study in Switzerland. Microb Genom. 2022;8(10):mgen000877. doi:10.1099/mgen.0.000877
13. Ungaro R, Mehandru S, Allen PB, et al. Ulcerative colitis. Lancet. 2017;389(10080):1756–1770. doi:10.1016/S0140-6736(16)32126-2
14. Torres J, Mehandru S, Colombel J-F, Peyrin-Biroulet L. Crohn′s disease. Lancet. 2017;389(10080):1741–1755. doi:10.1016/S0140-6736(16)31711-1
15. Jess T, Simonsen J, Nielsen NM, et al. Enteric Salmonella or Campylobacter infections and the risk of inflammatory bowel disease. Gut. 2011;60(3):318–324. doi:10.1136/gut.2010.223396
16. Schultz BM, Paduro CA, Salazar GA, et al. A potential role of Salmonella infection in the onset of inflammatory bowel diseases. Front Immunol. 2017;8:191. doi:10.3389/fimmu.2017.00191
17. Grimont PAD, Weill FX. Antigenic Formulae of the Salmonella Serovars.
18. CLSI. Performance Standards for Antimicrobial Susceptibility Testing, CLSI Supplement M100.
19. Xu H, Cao X, Zhang W, et al. Co-existence of two ciprofloxacin-resistant Salmonella enterica serovar Kentucky strains in the urine of a bladder cancer patient. J Glob Antimicrob Resist. 2022;30:354–356. doi:10.1016/j.jgar.2022.07.011
20. Feng Y, Zou S, Chen H, et al. BacWGSTdb 2.0: A One-Stop Repository for Bacterial Whole-Genome Sequence Typing and Source Tracking. Nucleic Acids Res;2021;49(D1):D644–D650. doi:10.1093/nar/gkaa821
21. Fisher RA, Gollan B, Helaine S. Persistent bacterial infections and persister cells. Nat Rev Microbiol. 2017;15(8):453–464. doi:10.1038/nrmicro.2017.42
22. Wang Y, Liu Y, Lyu N, et al. The temporal dynamics of antimicrobial-resistant Salmonella enterica and predominant serovars in China. Natl Sci Rev. 2022;10(3):nwac269. doi:10.1093/nsr/nwac269
23. World Health Organization. Critically Important Antimicrobials for Human Medicine, 6th Revision. Geneva, Switzerland: World Health Organization; 2019.
24. Wang J, Jiang Y, Ji RY, et al. Colistin- and tigecycline-resistant CTX-M-14-producing Salmonella enterica serovar Kentucky ST198 from retail chicken meat, China. Int J Antimicrob Agents. 2022;59(2):106504. doi:10.1016/j.ijantimicag.2021.106504
25. She Y, Jiang Y, Luo M, et al. Emergence of chromosomally located blaCTX-M-14b and qnrS1 in Salmonella enterica serotype Kentucky ST198 in China. Int J Antimicrob Agents. 2023;62(3):106896. doi:10.1016/j.ijantimicag.2023.106896
26. Mashe T, Thilliez G, Chaibva BV, et al. Highly drug resistant clone of Salmonella Kentucky ST198 in clinical infections and poultry in Zimbabwe. Npj Antimicrobials and Resistance. 2023;1(1):6. doi:10.1038/s44259-023-00003-6
27. Bevan ER, Jones AM, Hawkey PM. Global epidemiology of CTX-M β-lactamases: temporal and geographical shifts in genotype. J Antimicrob Chemother. 2017;72(8):2145–2155. doi:10.1093/jac/dkx146
28. Yao H, Qin S, Chen S, Shen J, Du XD. Emergence of carbapenem-resistant hypervirulent Klebsiella pneumoniae. Lancet Infect Dis. 2018;18(1):25. doi:10.1016/S1473-3099(17)30628-X
29. Yu R, Chen Z, Li D, Schwarz S, Wang X, Du XD. Studies on the transmission of a tigecycline resistance-mediating tet(A) gene variant from Enterobacter hormaechei via a two-step recombination process. Microbiol Spectr. 2022;10(3):e0049622. doi:10.1128/spectrum.00496-22
30. Conde MB, Lapa E, Silva JR. New regimens for reducing the duration of the treatment of drug-susceptible pulmonary tuberculosis. Drug Dev Res. 2011;72(6):501–508. doi:10.1002/ddr.20456
31. Vesely JJ, Pien FD, Pien BC. Rifampin, a useful drug for nonmycobacterial infections. Pharmacotherapy. 1998;18(2):345–357. doi:10.1002/j.1875-9114.1998.tb03861.x
32. Mumbo MT, Nyaboga EN, Kinyua JK, et al. Antimicrobial resistance profiles of Salmonella spp. and Escherichia coli isolated from fresh Nile tilapia (Oreochromis niloticus) fish marketed for human consumption. BMC Microbiol. 2023;23(1):306. doi:10.1186/s12866-023-03049-8
33. Brandis G, Wrande M, Liljas L, et al. Fitness-compensatory mutations in rifampicin-resistant RNA polymerase. Mol Microbiol. 2012;85(1):142–151. doi:10.1111/j.1365-2958.2012.08099.x
34. Brandis G, Hughes D. Mechanisms of fitness cost reduction for rifampicin-resistant strains with deletion or duplication mutations in rpoB. Sci Rep. 2018;8(1):17488. doi:10.1038/s41598-018-36005-y
35. Sánchez-Romero MA, Casadesús J. Contribution of phenotypic heterogeneity to adaptive antibiotic resistance. Proc Natl Acad Sci U S A. 2014;111(1):355–360. doi:10.1073/pnas.1316084111
36. Fang Q, Yin X, He Y, et al. Safety and efficacy of phage application in bacterial decolonisation: a systematic review. Lancet Microbe. 2024. doi:10.1016/S2666-5247(24)00002-8
 © 2024 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2024 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.