Back to Journals » Neuropsychiatric Disease and Treatment » Volume 22
Parameter-Specific Effects of Low-Intensity Transcranial Focused Ultrasound Stimulation on Depression-Like Behaviors in a CUMS Mouse Model
Authors Zhang Y, Zhang Y ![]() , Zhang K, Wang L, Huang D, Zhen F, Wang R, An C
, Zhang K, Wang L, Huang D, Zhen F, Wang R, An C
Received 21 December 2025
Accepted for publication 27 February 2026
Published 12 March 2026 Volume 2026:22 586583
DOI https://doi.org/10.2147/NDT.S586583
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Roger Pinder
Yan Zhang,1– 3,* Yaxing Zhang,1– 3,* Kaiming Zhang,1– 3 Lan Wang,1– 4 Dongyang Huang,5– 7 Fengya Zhen,1– 3 Ran Wang,1– 3 Cuixia An1– 4
1Department of Psychiatry, The First Hospital of Hebei Medical University, Shijiazhuang, Hebei, 050031, People’s Republic of China; 2The Mental Health Center of Hebei Medical University, Shijiazhuang, Hebei, 050031, People’s Republic of China; 3The Mental Health Institute of Hebei Medical University, Shijiazhuang, Hebei, 050031, People’s Republic of China; 4Collaborative Innovation Center of Hebei Province for Mechanism, Diagnosis and Treatment of Neuropsychiatric Diseases, Shijiazhuang, Hebei, 050017, People’s Republic of China; 5The Department of Pharmacology, Hebei Medical University, Shijiazhuang, Hebei, 050017, People’s Republic of China; 6The Key Laboratory of Neural and Vascular Biology, Ministry of Education, Hebei Medical University, Shijiazhuang, Hebei, 050017, People’s Republic of China; 7The Key Laboratory of New Drug Pharmacology and Toxicology, Hebei Medical University, Shijiazhuang, Hebei, 050017, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Cuixia An, Email [email protected]
Purpose: Depression is a multifactorial disorder involving neurotransmitter dysregulation, gut microbiota imbalance, and metabolic disturbances. Low-intensity transcranial focused ultrasound stimulation (LIFUS) holds promise for treating depression. However, the effects of different LIFUS parameter settings on depression-like behaviors, and their potential associations with gut microbiota and fecal metabolite changes, remain largely unexplored. This study aims to investigate the parameter-specific effects of LIFUS on depression-like behaviors in a chronic unpredictable mild stress (CUMS) mouse model, and to explore potential associations with changes in gut microbiota and fecal metabolites.
Methods: To establish a depression model, C57BL/6 mice were subjected to CUMS, while a separate cohort was kept as a control (CON) group. The CUMS-exposed mice were then randomly divided into four groups: CUMSpo, LIFUS1, LIFUS2 and SHAM. Depressive-like behaviors were evaluated using the sucrose preference test (SPT) and forced swim test (FST). The levels of neurotransmitters and Fecal concentrations of metabolites were quantified using liquid chromatography-tandem mass spectrometry (LC-MS/MS). Gut microbiota composition was analyzed by metagenomic sequencing, and α-diversity was assessed using the ACE, Chao1, and Shannon indices. Histopathology was assessed via HE staining.
Results: LIFUS at 1.5 kHz PRF, but not 300 Hz, significantly attenuated CUMS-induced depressive-like behaviors, evidenced by increased sucrose preference and reduced immobility time, without affecting locomotor activity. This behavioral effect was accompanied by a significant increase in cortical glutamate. LIFUS2 protocol was associated with a significant increase in tryptamine, alongside a concurrent trend towards restoring the abundance of Clostridia and enhancing gut microbiota α-diversity. HE staining confirmed protocol safety.
Conclusion: The antidepressant-like effects of LIFUS appear to be associated with multi-systemic alterations, including changes in cortical glutamate, modulation of the gut microbiota, and specific changes in tryptophan metabolism.
Keywords: LIFUS, depression-like behavior, gut-brain axis, glutamate, neuromodulation
Introduction
Major Depressive Disorder (MDD) is a significant contributor to the global burden of disease, affecting approximately 300 million individuals worldwide.1 In China, the lifetime prevalence of depressive disorders among adults is reported to be 6.8%.2 The chronic unpredictable mild stress (CUMS) model represents one of the most rigorously validated animal paradigms for examining depression-associated behavioral and neurobiological modifications.3 This paradigm was originally developed by Willner et al in 1987,4 utilizing exposure to various unpredictable mild stressors over several weeks to induce core depressive-like behaviors in rodents. Importantly, CUMS animals exhibit neurobiological alterations that closely parallel those observed in patients with major depressive disorder.5 This model has been extensively utilized for evaluating antidepressant efficacy6 and has proven instrumental in advancing our understanding of the underlying neurobiological mechanisms of depression.
Accumulating evidence suggests that the gut-brain axis is a critical bidirectional communication system.7 Tryptophan metabolism represents a critical node in the gut-brain axis. Alterations in fecal tryptophan metabolites reflect intestinal metabolic status; upon entering the circulation, these microbial metabolites can cross the blood-brain barrier to directly influence neuronal function and are closely associated with the pathogenesis of depression.8,9 Gut microbiota regulates tryptophan-derived metabolites through multiple pathways, and microbial dysbiosis impairs this metabolic capacity.10 Metagenomic profiling can identify specific bacterial genes encoding enzymes involved in tryptophan metabolism, providing mechanistic insights into how microbiota composition influences central nervous system function.11
Traditional antidepressant treatments, such as medication and psychotherapy, although effective, often suffer from limited efficacy, significant side effects, and poor patient compliance.12,13 Consequently, neuromodulation presents a promising therapeutic alternative. However, transcranial direct current stimulation (tDCS) and repetitive transcranial magnetic stimulation (rTMS) have relatively low spatial resolution. Compared to LIFUS, tDCS14 and TMS15 have lower spatial resolution. For example, rTMS has limited magnetic field penetration, struggling to effectively reach deep brain areas.16 These limitations make it challenging to accurately modulate specific regions, particularly when the target area is small or adjacent to other critical functional zones.
Despite the availability of multiple therapeutic approaches, many patients experience inadequate symptom relief or treatment-related adverse effects, underscoring an urgent need for novel interventions. LIFUS offers substantial therapeutic potential owing to its superior spatial resolutionand flexible targeting capabilities.17 LIFUS enables frequency-dependent neuromodulation by varying PRF, thereby eliciting distinct neurobiological effects at the same stimulation site. The prefrontal cortex, serving as a central hub for emotional regulation, represents a classical therapeutic target for depressive disorders.18,19 However, the mechanisms underlying LIFUS-mediated antidepressant effects through prefrontal cortex targeting remain poorly understood, particularly regarding its potential impact on the gut-brain axis, microbiota composition, and tryptophan metabolism.
Previous studies have shown that LIFUS modulates synaptic plasticity and neurotransmitter levels, reducing depression-like behaviors in preclinical models.20–22 However, its effects appear highly sensitive to stimulation parameter settings, with considerable inconsistencies in effective parameters reported across existing literature, underscoring the need for systematic comparison of different parameter protocols.23–25 More critically, potential associations between LIFUS treatment and changes in gut microbiota composition and related metabolic pathways remain largely unexplored. Given the recognized involvement of gut microbiota26 and tryptophan metabolism27 in depression pathophysiology, investigating whether LIFUS-induced behavioral improvements are accompanied by changes in these may provide valuable mechanistic insight.
This study examines how LIFUS at varying parameters affects depressive-like behaviors in CUMS mice, alongside changes in cortical glutamatergic transmission, gut microbiota, and fecal tryptophan metabolism. We hypothesize that frequency-dependent LIFUS targeting the prelimbic cortex will reduce depressive-like behaviors, which may be associated with alterations in the brain-gut axis and metabolic pathways. This study will explore the relationship between neural, microbial, and behavioral changes induced by LIFUS.
Materials and Methods
Experimental Paradigm
The experimental workflow is depicted in Figure 1. Male and female mice underwent a 7-day acclimation period before the experiments. All mice were subjected to an initial sucrose preference test (pre-SPT) to establish baseline anhedonia measurements. Based on the sucrose preference index (SPI) from pre-SPT, male and female mice were randomly assigned into two groups with comparable SPI distributions: CON group (n=20): 10 males and 10 females, no stress exposure; CUMS group (n=60): 30 males and 30 females, subjected to CUMS induction. Mice in the CON group were group-housed (5 per cage), while mice in the CUMS group were single-housed throughout the 4-week CUMS induction period. Following the 4-week CUMS protocol, behavioral tests (SPT, OFT, FST) were conducted to identify CUMS-vulnerable mice. A mouse was considered CUMS-vulnerable if it showed significant deficits in two or more tests compared to the CON group. Using this criterion, 17 males and 16 females from the CUMS group were identified as CUMS-vulnerable. Subsequently, these vulnerable mice were further randomized into four groups (n=8 per group, 4 males and 4 females per group): (1) CUMS group: received CUMS induction without stimulation; (2) LIFUS1 group: received CUMS induction with transcranial ultrasound stimulation at Protocol 1; (3) LIFUS2 group: received CUMS induction with transcranial ultrasound stimulation at Protocol 2; and (4) SHAM group: received CUMS induction with identical procedures as the LIFUS groups but with no ultrasound output. The CON group remained group-housed and was not stressed. In total, five experimental groups were established for the final phase of the study. LIFUS stimulation targeted the prelimbic cortex (PrL). After the stimulation period, all five groups underwent final behavioral assessments, including the SPT, OFT, and FST. Upon completion of behavioral testing, brain samples were collected for cortical neurotransmitter measurement and histopathological examination, while fecal samples were obtained for microbiota analysis and tryptophan metabolite detection.
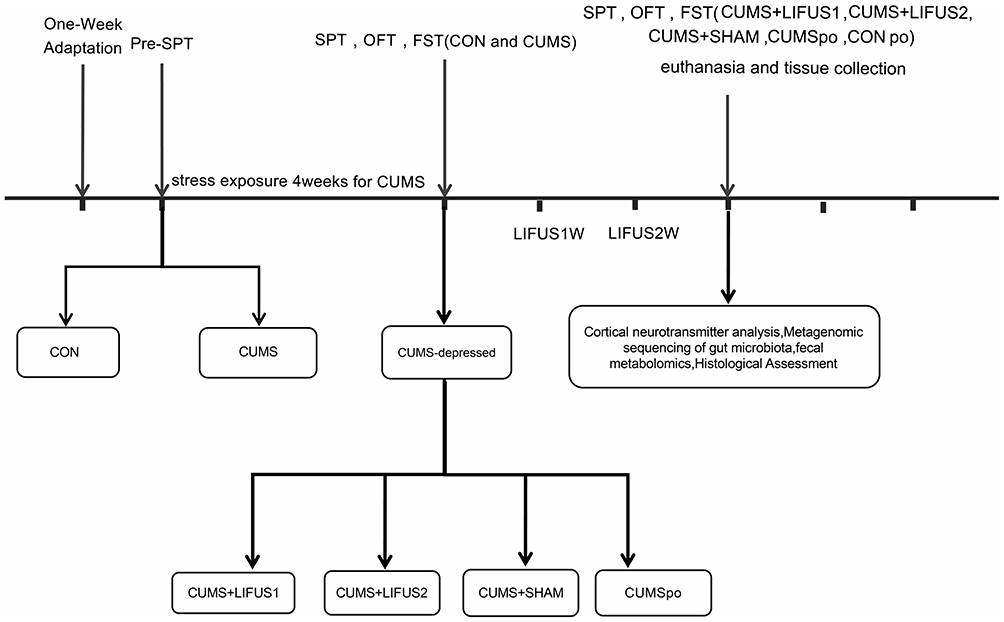 |
Figure 1 Schematic representation of the experimental procedure. Abbreviations: CUMS, Chronic Unpredictable Mild Stress; SPT, Sucrose Preference Test; OFT, Open Field Test; FST, Forced Swim Test; LIFUS1, Low-Intensity Transcranial Focused Ultrasound Stimulation with a PRF of 300 Hz; LIFUS2, Low-Intensity Transcranial Focused Ultrasound Stimulation with a PRF of 1.5 kHz. |
CUMS Model
The CUMS model was established using 80 adult C57BL/6 mice (40 males, 40 females; age: 6 weeks old; males: 18–20g, females:15–18g; Liaoning Changsheng Biotechnology Co., Ltd). Animals were housed in a controlled environment maintained at 24 ± 2°C, with a 12-hour light/dark cycle starting at 7:00 AM and provided with unrestricted access to food and water. The Laboratory Animal Ethical and Welfare Committee of Hebei Medical University granted approval for all animal procedures (Approval No. IACUC-Hebmu-2025031). The experiments were performed following the guidelines outlined in the National Institutes of Health Guide for the Care and Use of Laboratory Animals. After a one-week acclimatization period, mice were stratified by sex and randomly allocated to the control group (n=20) or CUMS group (n=60) in separate cages. Final intervention groups (n=8 per group) were determined based on previous studies20,21 showing adequate power to detect depression-related outcomes. The CUMS group was then subjected to a series of mild stressors, including: (1) 24 h water withholding, (2) 24 h food withholding, (3) cage tilt (45°,24 h), (4) soiled bedding exposure (24 h), (5) forced swimming in 40°C water (5 min), (6) forced swimming in 4°C water (5 min), (7) wet bedding exposure (24 h), (8) horizontal shaking (2 h), (9) physical restraint (2 h), and (10) tail clamping (5 min). Stressors were randomly scheduled each week to prevent repetition, and the CUMS protocol was maintained for 4 weeks. The control group was group-housed and not exposed to any stressors. Details are provided in Table 1.
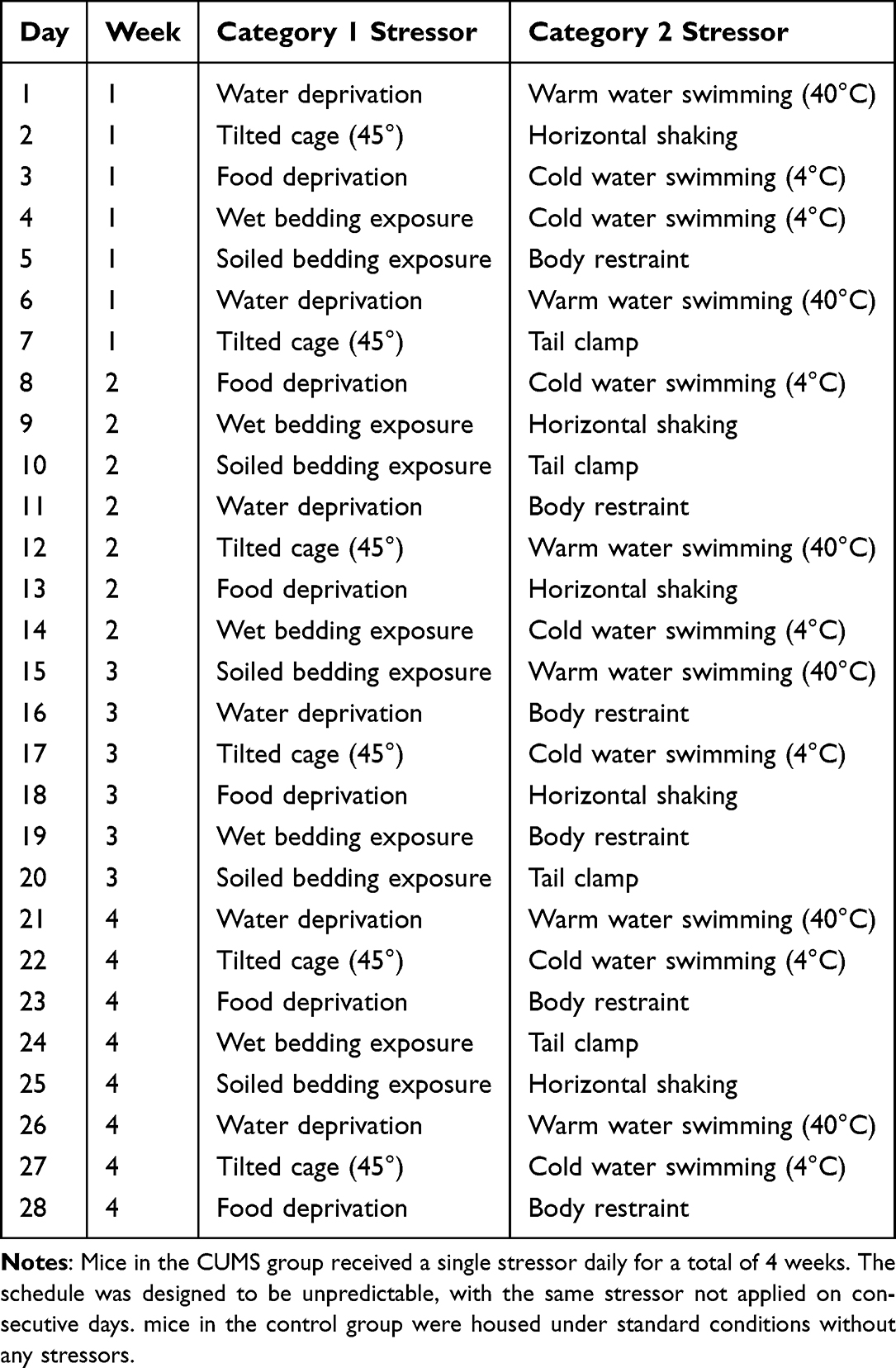 |
Table 1 Schedule of Chronic Unpredictable Mild Stress (CUMS) Procedures |
Behavioral Tests
A series of standardized behavioral tests, including the SPT, OFT, and FST, was conducted to evaluate behavioral phenotypes. All behavioral assessments were conducted under low-light, low-noise conditions.
SPT
Before the test, mice were acclimated for 24 hours with free access to two bottles, with one bottle providing 1% sucrose solution and the other providing tap water. Following the acclimation period, only food was deprived for 24 hours. Subsequently, the mice were given a free choice between the two bottles for 24 hours. Bottle positions were counterbalanced every 12 hours to eliminate position bias. Sucrose preference was calculated using the standardized formula: SP (%) = [sucrose consumption (g)/(sucrose consumption (g) + water consumption (g))]×100.
OFT
OFT was conducted on the day following the SPT. Each mouse was individually placed in the same corner of a white square arena (50 cm×50 cm×50 cm). After each trial, the testing area was thoroughly cleaned with 75% ethanol to eliminate any residual olfactory cues. The primary measures were total distance traveled (cm) to assess locomotor activity. Mice were allowed to freely explore the arena for 10 min, and their natural behaviors were monitored using video tracking software (Smart Maze, USA).
FST
In the FST, mice were placed in a transparent cylinder with a diameter of 15 cm filled with water maintained at 25–26°C for a 6-minute swimming session. Fresh water was replaced after each test. During the final 4 minutes of the 6-minute test, the duration of immobility was recorded. Tests were conducted in the following order, with a 24-hour interval between each: SPT, OFT, and FST.
Stereotactic Targeting and Animal Preparation
Anesthesia was induced in mice with 4% isoflurane and then maintained at 2.0%. The animals were then securely mounted onto a stereotaxic frame (RWD Life Science, China). After removing the hair from the scalp, a midline incision was made to reveal the underlying skull, and the bregma was identified as the reference point. According to The Mouse Brain in Stereotaxic Coordinates (second edition), the PrL was targeted using the following coordinates relative to bregma: anteroposterior (AP): +2.46 mm, mediolateral (ML):-0.25mm. As shown in Figure 2. Following a recovery period and a 3-day habituation protocol to the restraint apparatus, awake mice were gently secured in a custom-made holder for daily LIFUS stimulation.
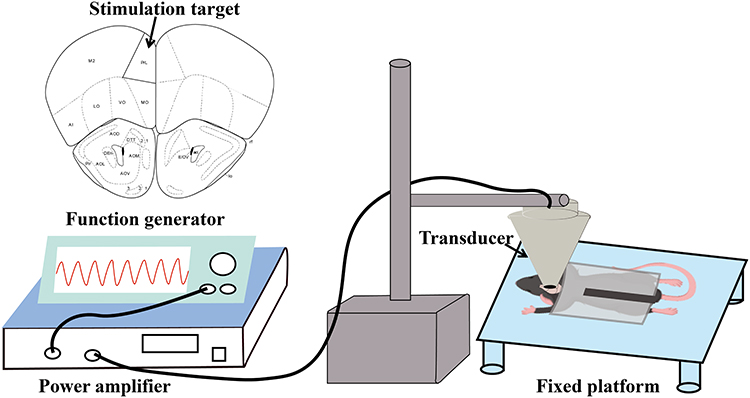 |
Figure 2 Schematic diagram of the experimental setup and principle. Abbreviation: PrL, prelimbic cortex. |
LIFUS System Setup
In the LIFUS system (Figure 2), a power amplifier (E&I240L, ENI Inc., USA) amplifies signals from function generators (DG1022U, RIGOL TECHNOLOGIES, INC., and AFG3022C, Tektronix, USA) and transmits them to an ultrasound transducer (V301-SU, Olympus, USA) to generate ultrasonic waves. A custom 3D-printed resin collimator, shaped like a cone and filled with coupling medium, was positioned to bridge the gap between the ultrasound transducer and the mouse skull, ensuring efficient acoustic energy delivery into the brain. These waves are delivered to the mouse’s PrL area via a collimator and a plastic tube filled with coupling gel. For the LIFUS group, the ultrasound parameters are as follows: Two LIFUS protocols were employed: Protocol 1 (Fundamental Frequency (FF) = 500 kHz, Pulse Repetition Frequency (PRF) = 300 Hz, Pulse Duration (PD) = 400 μs, Duty Cycle (DC) = 12%, ISPTA=0.26 W/cm2) and Protocol 2 (FF = 500 kHz, PRF = 1.5 kHz, PD = 400 μs, DC = 60%, ISPTA= 1.31 W/cm2). Both protocols used a sonication duration of 400 ms with 3-second inter-stimulus intervals. Each stimulation was conducted at the same time each day for a duration of 15 minutes. In the SHAM group, the power amplifier is turned off, with all other parameters remaining the same as in the LIFUS group. This measure was implemented to control for potential confounding factors, such as procedural noise and equipment sounds.
Anesthesia and Euthanasia
All procedures were performed under appropriate anesthesia to minimize animal pain and distress. Mice were anesthetized with isoflurane (RWD Life Science, Shenzhen, China) using an induction concentration of 4.0% in 100% oxygen, delivered via a precision vaporizer. Following induction, anesthesia was maintained at a concentration of 2.0% throughout the procedure. The depth of anesthesia was continuously monitored and confirmed by the lack of response to a toe pinch. Following the completion of the experimental procedures, euthanasia was performed by cervical dislocation while the animal remained under deep anesthesia, in strict accordance with the American Veterinary Medical Association (AVMA) Guidelines for the Euthanasia of Animals. Death was subsequently confirmed by the cessation of heartbeat and respiration.
Detection of Neurotransmitters
Neurotransmitter contents were detected based on the AB Sciex QTRAP 6500 LC-MS/MS platform.
Fecal Tissue
Fresh feces were collected in dry sterile 2 mL Eppendorf tubes and then transferred to a −80 °C refrigerator.
Metagenomic DNA Extraction and Library Preparation
Total microbial DNA was extracted using the Mag-Bind Soil DNA Kit (OMEGA, M5635-02). Metagenomic libraries were constructed following the Illumina TruSeq DNA Sample Preparation protocol. Briefly, DNA was fragmented using a Bioruptor, followed by end repair, A-tailing, and adapter ligation. Ligated products were purified with magnetic beads, and libraries were enriched by PCR with minimal cycles. Library quality was assessed using Qubit 4 and Agilent 2100 Bioanalyzer. Qualified libraries were normalized to 10 nM, pooled in equal volumes, and sequenced on the Illumina platform.
Targeted Quantitative Analysis of Tryptophan Pathway Metabolites
Metabolites were quantitatively analyzed using ultra-performance liquid chromatography-tandem mass spectrometry (UPLC-MS/MS). The detailed procedures for the quantitative analysis of tryptophan pathway metabolites are described in Supplementary Methods.
HE Staining
To evaluate the effects of LIFUS parameters on murine brain tissue, we initially performed HE staining. First, the brain tissue was harvested from anesthetized mice and fixed in 4% paraformaldehyde for 24 hours. The tissue was then embedded in paraffin, and 5μm-thick sections were cut. After deparaffinization and rehydration through a graded ethanol series, the sections were stained with hematoxylin for 15minutes, followed by three rinses in distilled water. Subsequently, the sections were differentiated in 1% hydrochloric alcohol for 10 seconds, rinsed three times in water, and then counterstained with 1% eosin for 5 minutes. Excess eosin was removed by washing with distilled water. Finally, the sections were dehydrated, cleared, and mounted. Stained sections were scanned and imaged using a digital slide scanner.
Statistical Analysis
Statistical analyses were conducted with GraphPad Prism 9 software. The normality of data distribution was assessed using the Kolmogorov–Smirnov test. The independent samples t-test was employed for two-group comparisons of normally distributed data. For comparisons involving more than two groups, a one-way analysis of variance (ANOVA) was applied, with Tukey’s post-hoc test used for subsequent pairwise analysis. For non-normally distributed data, the Mann–Whitney U-test was used. Although the study included both male and female mice, the data were pooled for analysis due to the limited sample size, which restricted the power to detect sex-specific differences. Statistical significance was defined as a P-value < 0.05 for all analyses.
Results
Behavioral Testing During CUMS Exposure
Before CUMS induction, baseline sucrose preference testing was conducted in all mice. The analysis showed no significant difference in SPI between the CON group (n=10) and CUMS group (n=30) in male mice (CON: 89.55±2.354%, CUMS: 89.34 ±1.423%, P = 0.860) (Figure 3A). Similarly, female mice showed comparable SPI values between CON (n=10) and CUMS groups (n=30) (CON: 84.4±2.519%, CUMS: 83.42±1.566%, P = 0.914) (Figure 3B). These findings demonstrate that hedonic responses were equivalent across groups before experimental manipulation. Following 4 weeks of CUMS exposure, pronounced depression-like behavioral alterations emerged in both sexes. Male mice subjected to CUMS exhibited a marked reduction in sucrose preference relative to controls (CON: 86.11±1.237%, CUMS: 79.13±2.019%, P = 0.011) (Figure 3C). Locomotor activity in the open field remained comparable between groups (CON: 1801±94.40cm, CUMS: 1802±72.80cm, P = 0.863) (Figure 3D). Stressed males displayed prolonged immobility in the forced swim test compared with controls (CON: 95.32±13.26 s, CUMS: 159.5±14.13 s, P = 0.006) (Figure 3E). Similarly, female mice treated with CUMS also exhibited significant depression-like behavioral changes (Figure 3F–H). Following the 4-week CUMS protocol, depression-like phenotypes were successfully induced in 17 male and 16 female mice, as evidenced by a significant reduction in sucrose preference and increased immobility in the forced swim test compared to controls.
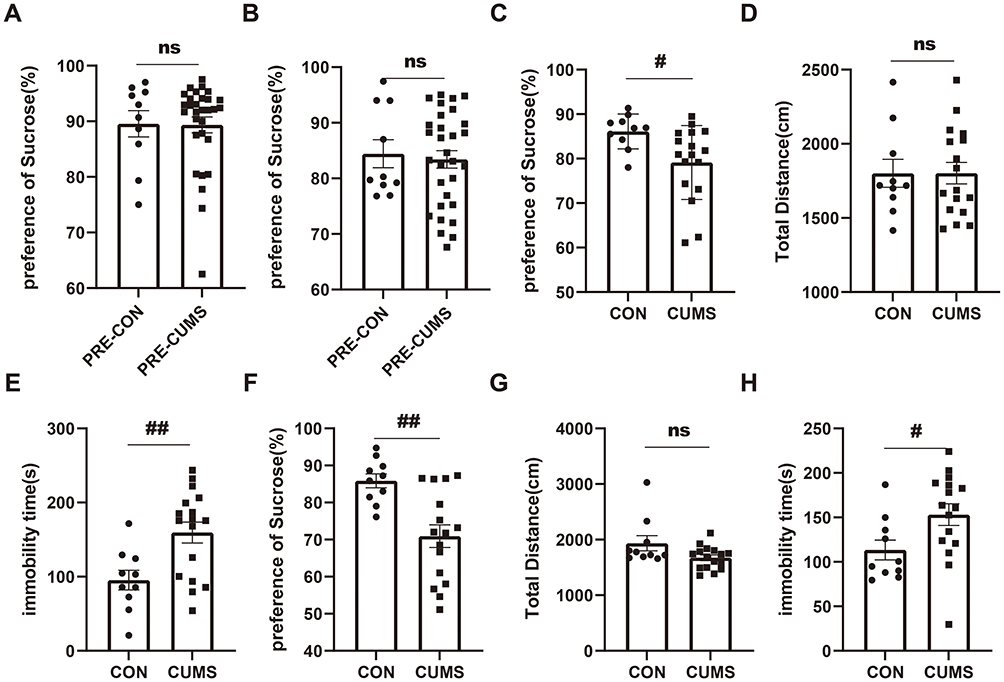 |
Figure 3 CUMS induces depression-like behaviors in mice. (A and B) Baseline sucrose preference in male (A) and female (B) mice before CUMS induction, showing no significant differences between groups. (C) Sucrose preference after 4 weeks of CUMS in male mice. (D) Total distance traveled in the OFT for male mice. (E) Immobility time in the FST for male mice. (F) Sucrose preference after 4 weeks of CUMS in female mice. (G) Total distance traveled in the OFT for female mice. (H) Immobility time in the FST for female mice. The initial group sizes were n=20 (male=10, female=10) for CON and n=60 (male=30, female=30) for CUMS. Following the 4-week CUMS protocol, mice were considered successfully modeled if they showed significant deficits in 2 or more tests compared with the CON group. Based on this criterion, 17 male and 16 female mice from the CUMS group were included in the final analysis. Data are presented as mean ± SEM. Abbreviations: CUMS, Chronic Unpredictable Mild Stress; ns, not significant. Notes: #P < 0.05, ##P < 0.01. |
Body Weight Dynamics During CUMS Exposure
Weight trajectories were monitored throughout the 4-week CUMS protocol using repeated measures ANOVA. In male mice, stress exposure significantly attenuated weight gain compared with controls (time × group interaction: F(5, 125) = 7.192 P<0.0001, Figure 4A). This divergence became apparent from week 2 onward, with statistically significant reductions observed in weeks 2, 3, and 5 (P < 0.001, Figure 4A). In female mice, CUMS-exposed animals showed significantly reduced body weight at weeks 2,4, and 5 relative to controls (P < 0.05, Figure 4B).
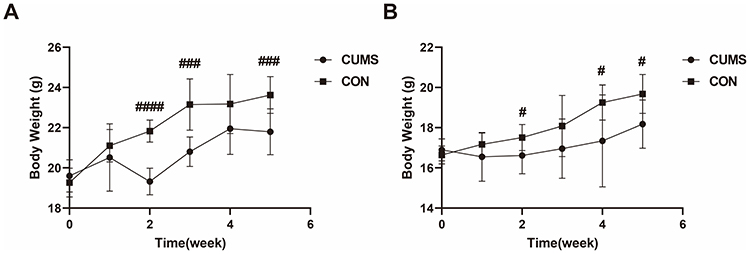 |
Figure 4 Body weight changes during CUMS exposure. Body weight of (A) male and (B) female mice measured weekly from baseline (Week 0) through 4 weeks of CUMS exposure. Male: CON (n=10), CUMS (n=17); Female: CON (n=10), CUMS (n=16). Data are presented as mean ± SD. Abbreviation: CUMS, Chronic Unpredictable Mild Stress. Notes: #P < 0.05, ###P < 0.001, ####P < 0.0001. |
Behavioral Improvement in Depressive-Related Behavior by LIFUS
To evaluate the antidepressant-like effects of ultrasound stimulation with different parameters, CUMS mice were allocated into a sham stimulation group (SHAM, n=8) and two active stimulation groups with distinct parameters (LIFUS1, n=8; LIFUS2, n=8), with each group comprising 4 male and 4 female mice, and compared against a control group (CON, n=8). One-way analysis of variance revealed that LIFUS2 treatment markedly ameliorated CUMS-induced anhedonia-like behavior, with the sucrose preference index showing a significant elevation relative to the SHAM group (LIFUS2: 90.96±1.113%, SHAM: 75.13±3.964%, Adjusted P value=0.033, Figure 5A). In the OFT, total locomotor distance showed no significant variation across five groups (Adjusted P value>0.999, Figure 5B), indicating that the antidepressant-like effects of ultrasound stimulation were not attributable to alterations in motor capacity. In FST, the LIFUS2 group demonstrated a significant reduction in immobility time compared to the SHAM group (Adjusted P value=0.036, Figure 5C), indicating an improvement in despair-like behavior. In contrast, the LIFUS1 group showed no significant difference from the Sham group (Adjusted P>0.999).
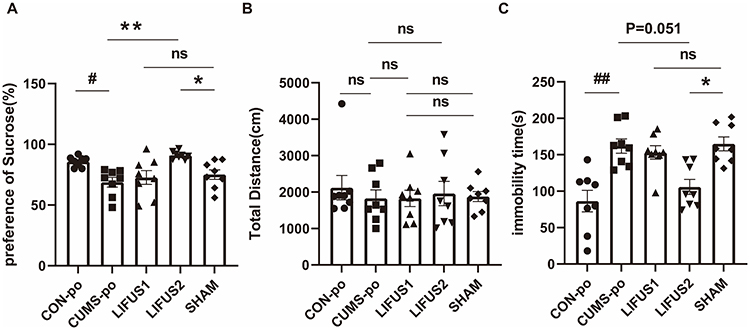 |
Figure 5 (A) SPT, (B) OFT, (C) FST in CON-po, CUMS-po, LIFUS1, LIFUS2, SHAM. LIFUS2 significantly improved SPT and reduced immobility time in FST compared to the SHAM group, while LIFUS1 showed no significant effects. Abbreviations: CUMSpo, Chronic Unpredictable Mild Stress; LIFUS1, Low-Intensity Transcranial Focused Ultrasound Stimulation with a PRF of 300 Hz; LIFUS2, Low-Intensity Transcranial Focused Ultrasound Stimulation with a PRF of 1.5 kHz; SPT, Sucrose Preference Test; OFT, Open Field Test; FST, Forced Swim Test; ns, no significant. Notes: #P < 0.05, ##P < 0.01, *P < 0.05, **P < 0. 01. |
Effect of LIFUS2 on Cortical Neurotransmitter Levels
We next assessed the levels of cortical neurotransmitters across the CON (n=6), CUMS (n=5), LIFUS2 (n=8), and SHAM (n=6) groups. As illustrated in Figure 6, the CUMS procedure resulted in a significant decrease in cortical glutamic acid levels compared with the CON group. Notably, LIFUS2 treatment effectively reversed the observed deficit. Furthermore, it elevated cortical glutamate content to a level that significantly surpassed that of the SHAM group.
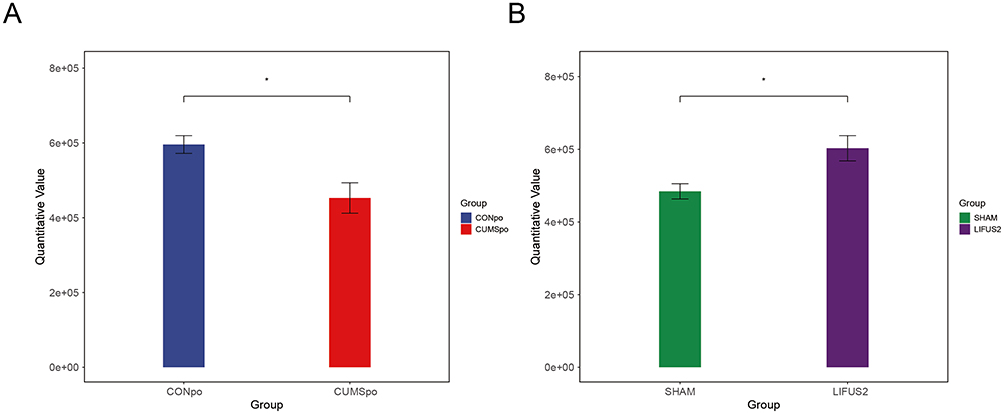 |
Figure 6 Effects of CUMS induction and LIFUS2 treatment on cortical neurotransmitter levels in mice. The bar chart illustrates the cortical glutamate concentrations across four groups: CONpo, CUMS, LIFUS2, and SHAM. (A) Results indicated that the CUMS group had significantly lower cortical glutamate levels compared to the Control group. (B) After LIFUS2 treatment, the cortical glutamate concentration was significantly higher than the SHAM group. Data are presented as mean ± SEM (ng/mg tissue). *P < 0.05 vs CON/SHAM group at the corresponding time point. Abbreviations: CUMSpo, Chronic Unpredictable Mild Stress; LIFUS2, Low-Intensity Transcranial Focused Ultrasound Stimulation with a PRF of 1.5 kHz. |
LIFUS Modulates α Diversity of the Gut Microbiota
A Kruskal–Wallis test confirmed a significant overall effect of treatment on gut microbiota alpha diversity (P < 0.05). The data exhibited a graded response consistent with our hypothesis: CUMS exposure was associated with the lowest alpha diversity, while LIFUS2 treatment was associated with intermediate, higher levels (Figure 7A). This CUMS-decreasing and LIFUS2-increasing trend was consistent across Chao1, pielou-e and Shannon indices, although post-hoc comparisons failed to achieve statistical significance (all P > 0.05). A similar non-significant trend was observed for the Clostridia class (Figure 7B and C). Beta diversity analysis, however, showed no significant alteration in the overall microbial community structure between the LIFUS2 and SHAM groups (Supplementary Table 1).
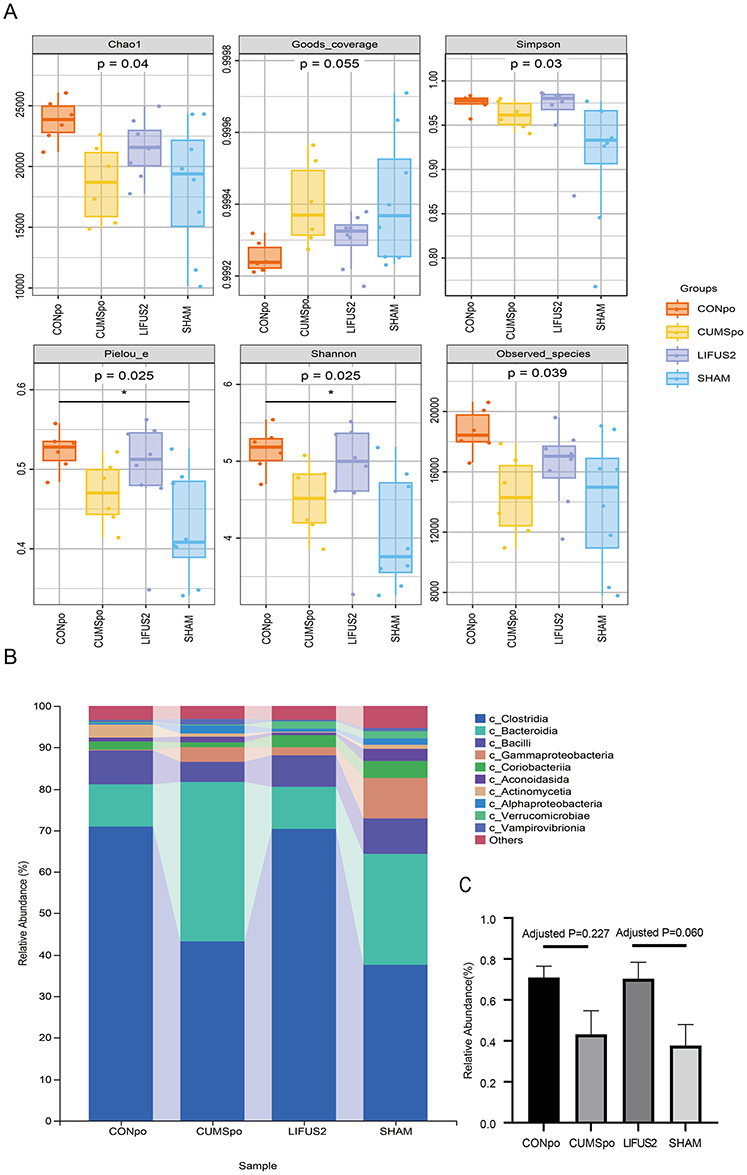 |
Figure 7 (A) Box-whisker plots showing alpha diversity indices. Chao1 and Observed_species reflect species richness; Shannon and Simpson indices reflect species diversity; Pielou_e represents species evenness; and Goods_coverage indicates sampling coverage. The box extends from the 25th to 75th percentiles (interquartile range, IQR), with a horizontal line at the median. Whiskers represent the most extreme values within 1.5×IQR. Individual points beyond the whiskers indicate outliers. CONpo (n=6), CUMSpo (n=6), LIFUS2 (n=8), and SHAM (n=8). (B) The x-axis shows the group names, while the y-axis shows the relative abundance of each taxon at a given taxonomic level. (C) Relative abundance of gut microbiota at the class level in different experimental groups. The bar chart shows the microbial composition at the class level in CONpo, CUMSpo, LIFUS2, and SHAM groups. Data are presented as mean ± SEM. Abbreviations: CONpo, Control; CUMSpo, Chronic Unpredictable Mild Stress; LIFUS2, Low-Intensity Transcranial Focused Ultrasound Stimulation with a PRF of 1.5 kHz. Notes: Kruskal–Wallis P-values are displayed below the diversity index labels, *P < 0.05. |
Targeted Analysis of Fecal Tryptophan Metabolites
To investigate alterations in the tryptophan metabolic pathway, we performed targeted metabolomic analysis on fecal samples. We identified tryptamine as one of the most significantly altered metabolites among the experimental groups (P < 0.05). Quantitative analysis (Figure 8) revealed that CUMS exposure significantly reduced fecal tryptamine levels compared to the control group (CON: 49.16 ± 27.310 ng/g, n = 6; CUMS: 15.80 ± 7.955 ng/g, n = 6, P < 0.05). Notably, this deficit was effectively reversed by LIFUS2 treatment. The tryptamine level in the LIFUS2 group (27.33 ± 8.02 ng/g, n = 5) was significantly higher than that in both the CUMS group (15.80 ± 7.955 ng/g, n = 6) and the SHAM group (13.84 ± 10.62 ng/g, n = 7, P < 0.05). These findings demonstrate that CUMS stress depletes fecal tryptamine and that LIFUS2 intervention can effectively restore its levels.
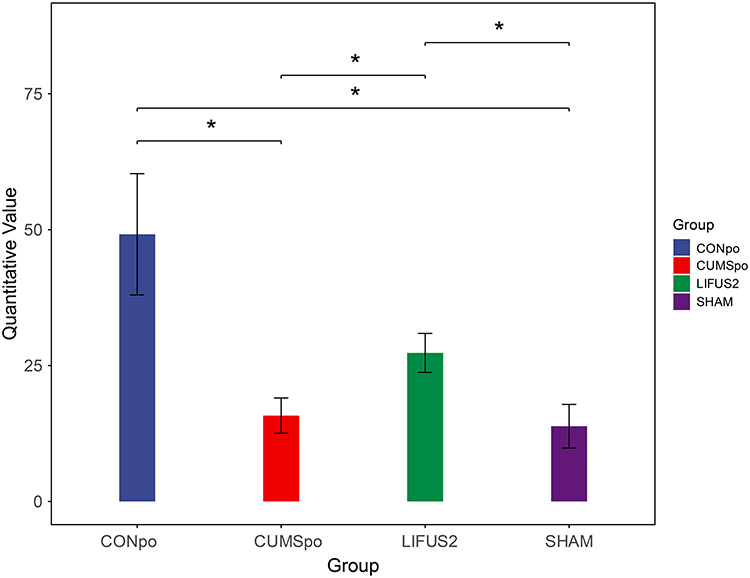 |
Figure 8 Distribution of fecal tryptamine, a differential tryptophan metabolite, in four experimental groups: CONpo (blue), CUMSpo (Orange), LIFUS2 (green), and SHAM (purple). Data are presented as Bar chart. Statistical significance was determined by one-way ANOVA followed by Tukey’s post hoc test. *P < 0.05. Abbreviations: CUMSpo, Chronic Unpredictable Mild Stress; LIFUS2, Low-Intensity Transcranial Focused Ultrasound Stimulation with a PRF of 1.5 kHz. |
Safety Assessment of LIFUS
To evaluate the safety of chronic LIFUS2 exposure, we performed H&E staining to screen for potential tissue damage or hemorrhage in the brain. After two weeks of stimulation, no signs of histopathological damage or hemorrhage were observed in the PrL of mice (Supplementary Figure 1). These results indicate that the LIFUS protocol used in this study is safe for chronic application to the PrL area.
Discussion
Glutamate, the primary excitatory neurotransmitter in the central nervous system, plays a critical role in brain function through its metabolic homeostasis.28 This neurometabolic feature has been observed in clinical patients,29 including those with first-episode MDD30,31 and chronic or relapsing-remitting MDD patients,32 more pronounced in the latter group. In this study, LIFUS2 treatment increases cortical glutamate levels and improves behavioral outcomes, as evidenced by SPT restoration and diminished despair in FST. Thus, glutamate elevation following LIFUS2 treatment may reflect recovery from pathological reduction toward the normal range.
The role of the brain-gut axis in depression is widely recognized.33–35 Previous studies show that CUMS-induced dysbiosis is accompanied by reduced α-diversity.36 In our study, although post-hoc pairwise comparisons did not reach statistical significance, the data present a consistent pattern. Across multiple alpha diversity metrics, CUMS exposure was associated with a reduction in diversity, while LIFUS2 treatment corresponded to a trend towards improvement. This directional consistency suggests that LIFUS2 may have the potential to counteract the dysbiotic effects of CUMS, although this interpretation remains limited by the current sample size. The consistency of this trend across different diversity indices and at the taxonomic level (for Clostridia) indicates that it may hold biological relevance, warranting further investigation with adequately powered cohorts. Our findings are partially supported by a previous study.37
Our observations showed that CUMS exposure was associated with a reduction in Clostridia abundance and significantly reduced fecal tryptamine, whereas LIFUS2 treatment appeared to counteract this trend and significantly elevate tryptamine concentrations. This parallel pattern is biologically plausible, as Clostridia are known tryptamine producers.38 We therefore postulate that the behavioral benefits of LIFUS2 may be related to the Clostridia abundance and subsequent modulation of tryptamine levels. However, our current data cannot establish a direct causal link. The Clostridia-tryptamine axis thus represents a promising therapeutic target for depressive-like behaviors that require future validation.
This suggests that LIFUS’s regulation of the gut-brain axis may involve mechanisms beyond tryptophan metabolism. Future studies should employ metagenomic and metabolomic approaches to systematically investigate LIFUS’s effects on microbial functional genes, other key metabolites (eg, bile acid metabolites). β-diversity remained relatively stable, suggesting LIFUS primarily affects microbial richness rather than community structure.
The most striking finding of this study is the parameter-dependent effect of LIFUS: LIFUS2 significantly improved depressive-like behaviors, accompanied by elevated cortical glutamate and brain-gut axis microecological alterations, whereas LIFUS1 showed no such effects. This contrast underscores the frequency specificity of LIFUS neuromodulation.
This study offers novel experimental evidence for the multidimensional mechanisms underlying LIFUS. The observed alterations in cortical glutamate and the brain-gut axis suggest a new perspective wherein therapeutic effects from non-pharmacological neuromodulation are accompanied by shifts in cerebral metabolism and gut microecology. Notably, the efficacy of LIFUS may not be a simple local effect. This suggests that the efficacy of LIFUS may not be limited to local brain effects but may also involve systemic modulation that includes peripheral pathways.
From a clinical perspective, these findings hold promising translational potential. Unlike pharmacological treatments that often target single pathways, LIFUS offers a non-invasive alternative that shows promise in simultaneously modulating metabolic and microecological dysregulation associated with depression. This multimodal action suggests potential utility for treatment-resistant depression or patients intolerant to systemic drug side effects. Furthermore, given the potential of focused ultrasound to modulate the gut-brain axis, this approach suggests the possibility of developing personalized neuromodulation strategies based on a patient’s metabolic or microbial profile.
This study has several major limitations. First, the observational cross-sectional design with all measurements collected post-LIFUS treatment precludes establishing causality among the concurrent changes in cortical glutamate elevation, microbial diversity increase, and behavioral improvement. Second, findings from the mouse CUMS model39 may not directly translate to clinical depression, requiring validation in human patients. Physiological and anatomical differences between mice and humans, combined with the CUMS model’s inability to fully recapitulate human depression complexity, represent important limitations. While our data provides proof of preliminary evidence, clinical validation is necessary. Third, while LIFUS2 yielded significant behavioral effects and showed a trend toward gut microbiota changes, the precise neurobiological mechanisms underlying these effects are not yet fully understood and require further study. Future research needs to conduct long-term dynamic monitoring of all indicators while implementing interventions through microbiota transplantation and metabolite supplementation. On this basis, researchers need to systematically screen stimulation frequencies to identify the optimal therapeutic timing and determine the optimal parameters for stimulation intensity and duration. Subsequently, these optimized schemes must undergo rigorous validation, first in large animal models, then advancing to small-scale clinical trials, aimed at examining their safety and therapeutic efficacy, and exploring their synergistic effects with existing therapies. Fourth, this study was limited by a relatively small sample size, particularly for the microbiota and metabolomics analyses. While the observed trends in α-diversity and metabolite levels suggest potential biological effects, the limited statistical power may preclude the detection of subtle differences. Therefore, non-significant results in these exploratory outcomes should be interpreted with caution. Another limitation is that while the study included both sexes, the statistical analysis was performed on pooled data. Future studies with larger sample sizes are required to include sex as a biological variable in the statistical models to determine if there are sex-specific therapeutic effects. Finally, to comprehensively evaluate the systemic effects of LIFUS and lay the foundation for personalized treatment, the key lies in integrating advanced neuroimaging technologies.40
Conclusion
This study systematically evaluated the therapeutic effects of LIFUS on CUMS-induced depressive-like behaviors. We observed marked frequency dependence; LIFUS2 significantly ameliorated depressive-like behaviors, including anhedonia recovery and reduced despair, whereas LIFUS1 showed no effects. These findings suggest that the therapeutic benefits of LIFUS were accompanied by significant increases in cortical glutamate and the fecal metabolite tryptamine, as well as a trend towards greater gut microbiota α-diversity. This indicates that the therapeutic effects of LIFUS may involve multidimensional central and peripheral alterations, though the precise mechanistic links remain to be clarified. This study provides a theoretical basis for understanding the relationship between non-invasive neuromodulation and gut microbiota, warranting future long-term studies and clinical validation to further optimize LIFUS parameters and treatment protocols.
Data Sharing Statement
Data are available from the corresponding author upon reasonable request.
Ethical Statement
All animal procedures in this study were approved by the Laboratory Animal Ethical and Welfare Committee of Hebei Medical University (Approval No. IACUC-Hebmu-2025031) and were performed in strict accordance with relevant national and international guidelines for the care and use of laboratory animals. All efforts were made to minimize animal suffering.
Acknowledgments
We gratefully acknowledge Professor Yuan Yi’s research group for providing ultrasound therapy equipment and technical support. We gratefully acknowledge the proteomics experimental support from Shanghai Personal Biotechnology Co., Ltd, China.
Funding
This study is funded by the National Key Research and Development Program of China [No. 2023YFC2506201], the Government Funded Clinical Medicine Excellent Talents Training Project of Hebei Province [No. ZF2025052], the Beijing-Tianjin-Hebei Basic Research Cooperation Special Project [No. H2023206925], the Science and Technology Program of Hebei Province [No. SG2021189], the Medical Science Research Project of Hebei [No. 20240005] and the Natural Science Foundation of Hebei Province [No. H2023206491].
Disclosure
The authors declare that they have no conflicts of interest (financial or personal) that could have influenced the work described herein.
References
1. World Health Organization. Depression and Other Common Mental Disorders: Global Health Estimates. World Health Organization; 2017.
2. Lu J, Xu X, Huang Y, et al. Prevalence of depressive disorders and treatment in China: a cross-sectional epidemiological study. Lancet Psychiatry. 2021;8(11):981–15. doi:10.1016/S2215-0366(21)00251-0
3. Sharma S, Chawla S, Kumar P, Ahmad R, Verma PK. The chronic unpredictable mild stress (CUMS) Paradigm: bridging the gap in depression research from bench to bedside. Brain Res. 2024;1843:149123. doi:10.1016/j.brainres.2024.149123
4. Willner P, Towell A, Sampson D, Sophokleous S, Muscat R. Reduction of sucrose preference by chronic unpredictable mild stress, and its restoration by a tricyclic antidepressant. Psychopharmacology. 1987;93(3):358–364. doi:10.1007/BF00187257
5. Willner P. The chronic mild stress (CMS) model of depression: history, evaluation and usage. Neurobiol Stress. 2017;6:78–93. doi:10.1016/j.ynstr.2016.08.002
6. Ratajczak P, Martyński J, Zięba JK, et al. Comparative efficacy of animal depression models and antidepressant treatment: a systematic review and meta-analysis. Pharmaceutics. 2024;16(9):1144. doi:10.3390/pharmaceutics16091144
7. Loh JS, Mak WQ, Tan LKS, et al. Microbiota–gut–brain axis and its therapeutic applications in neurodegenerative diseases. Signal Transduct Target Ther. 2024;9(1):37. doi:10.1038/s41392-024-01743-1
8. Zhou M, Fan Y, Xu L, et al. Microbiome and tryptophan metabolomics analysis in adolescent depression: roles of the gut microbiota in the regulation of tryptophan-derived neurotransmitters and behaviors in human and mice. Microbiome. 2023;11(1):145. doi:10.1186/s40168-023-01589-9
9. Chen L-M, Bao C-H, Wu Y, et al. Tryptophan-kynurenine metabolism: a link between the gut and brain for depression in inflammatory bowel disease. J Neuroinflammation. 2021;18(1):135. doi:10.1186/s12974-021-02175-2
10. Xie J, Wu W-T, Chen -J-J, et al. Tryptophan metabolism as bridge between gut microbiota and brain in chronic social defeat stress-induced depression mice. Front Cell Infect Microbiol. 2023;13:1121445. doi:10.3389/fcimb.2023.1121445
11. Lai W-T, Deng W-F, Xu S-X, et al. Shotgun metagenomics reveals both taxonomic and tryptophan pathway differences of gut microbiota in major depressive disorder patients. Psychol Med. 2021;51(1):90–101. doi:10.1017/S0033291719003027
12. Saelens J, Gramser A, Watzal V, Zarate CA, Lanzenberger R, Kraus C. Relative effectiveness of antidepressant treatments in treatment-resistant depression: a systematic review and network meta-analysis of randomized controlled trials. Neuropsychopharmacology. 2025;50(6):913–919. doi:10.1038/s41386-024-02044-5
13. Braund TA, Tillman G, Palmer DM, Gordon E, Rush AJ, Harris AW. Antidepressant side effects and their impact on treatment outcome in people with major depressive disorder: an iSPOT-D report. Transl Psychiatry. 2021;11(1):417. doi:10.1038/s41398-021-01533-1
14. Shi Y, Cai, G, Wu, W. A panoramic review of transcranial focused ultrasound neuromodulation: from basic research to clinical applications. J Neuroeng Rehabil. 2025;22(1):227. doi:10.1186/s12984-025-01753-2
15. Matt E, Radjenovic, S, Mitterwallner, M, Beisteiner, R. Current state of clinical ultrasound neuromodulation. Front Neurosci. 2024;18():1420255. doi:10.3389/fnins.2024.1420255
16. Khalifa A, Abrishami SM, Zaeimbashi M, et al. Magnetic temporal interference for noninvasive and focal brain stimulation. J Neural Eng. 2023;20(1):016002. doi:10.1088/1741-2552/acb015
17. Osada T, Konishi S. Noninvasive intervention by transcranial ultrasound stimulation: modulation of neural circuits and its clinical perspectives. Psych Clin Neurosci. 2024;78(5):273–281. doi:10.1111/pcn.13663
18. Marwood L, Wise T, Perkins AM, Cleare AJ. Meta-analyses of the neural mechanisms and predictors of response to psychotherapy in depression and anxiety. Neurosci Biobehav Rev. 2018;95:61–72. doi:10.1016/j.neubiorev.2018.09.022
19. Zhou X-T, Bao W-D, Liu D, Zhu L-Q. Targeting the neuronal activity of prefrontal cortex: new directions for the therapy of depression. Curr Neuropharmacol. 2020;18(4):332–346. doi:10.2174/1570159X17666191101124017
20. Wang F, Cai Q, Ju R, et al. Low-intensity focused ultrasound ameliorates depression-like behaviors associated with improving the synaptic plasticity in the vCA1-mPFC pathway. Cereb Cortex. 2023;33(12):8024–8034. doi:10.1093/cercor/bhad095
21. Zhang J, Zhou H, Yang J, et al. Low‐intensity pulsed ultrasound ameliorates depression‐like behaviors in a rat model of chronic unpredictable stress. CNS Neurosci Ther. 2021;27(2):233–243. doi:10.1111/cns.13463
22. Zhu Y, He J, Wu C, et al. Transcranial ultrasound stimulation relieves depression in mice with chronic restraint stress. J Neural Eng. 2023;20(3):036011. doi:10.1088/1741-2552/ac8bfd
23. Sherman J, Bortz E, Antonio ES, Tseng H-A, Raiff L, Han X. Ultrasound pulse repetition frequency preferentially activates different neuron populations independent of cell type. J Neural Eng. 2024;21(5):056008. doi:10.1088/1741-2552/ad731c
24. Manuel TJ, Kusunose J, Zhan X, et al. Ultrasound neuromodulation depends on pulse repetition frequency and can modulate inhibitory effects of TTX. Sci Rep. 2020;10(1):15347. doi:10.1038/s41598-020-72189-y
25. Yu K, Niu X, Krook-Magnuson E, He B. Intrinsic functional neuron-type selectivity of transcranial focused ultrasound neuromodulation. Nat Commun. 2021;12(1):2519. doi:10.1038/s41467-021-22743-7
26. Liu Y, Wang H, Gui S, et al. Proteomics analysis of the gut–brain axis in a gut microbiota-dysbiosis model of depression. Transl Psychiatry. 2021;11(1):568. doi:10.1038/s41398-021-01689-w
27. Gao K, Mu C-L, Farzi A, Zhu W-Y. Tryptophan metabolism: a link between the gut microbiota and brain. Adv Nutr. 2020;11(3):709–723. doi:10.1093/advances/nmz127
28. Sarawagi A, Soni ND, Patel AB. Glutamate and GABA homeostasis and neurometabolism in major depressive disorder. Front Psychiatry. 2021;12:637863. doi:10.3389/fpsyt.2021.637863
29. Hasler G, van der Veen JW, Tumonis T, Meyers N, Shen J, Drevets WC. Reduced prefrontal glutamate/glutamine and γ-aminobutyric acid levels in major depression determined using proton magnetic resonance spectroscopy. Arch Gen Psychiatry. 2007;64(2):193–200. doi:10.1001/archpsyc.64.2.193
30. Draganov M, Vives-Gilabert Y, de Diego-Adeliño J, Vicent-Gil M, Puigdemont D, Portella MJ. Glutamatergic and GABA-ergic abnormalities in First-episode depression. A 1-year follow-up 1H-MR spectroscopic study. J Affective Disorders. 2020;266:572–577. doi:10.1016/j.jad.2020.01.138
31. Shirayama Y, Takahashi M, Osone F, Hara A, Okubo T. Myo-inositol, glutamate, and glutamine in the prefrontal cortex, hippocampus, and amygdala in major depression. Biol Psychiatry. 2017;2(2):196–204. doi:10.1016/j.bpsc.2016.11.006
32. Portella MJ, de Diego-Adeliño J, Gómez-Ansón B, et al. Ventromedial prefrontal spectroscopic abnormalities over the course of depression: a comparison among first episode, remitted recurrent and chronic patients. J Psychiatr Res. 2011;45(4):427–434. doi:10.1016/j.jpsychires.2010.08.010
33. Morais LH, Schreiber IV HL, Mazmanian SK. The gut microbiota–brain axis in behaviour and brain disorders. Nat Rev Microbiol. 2021;19(4):241–255. doi:10.1038/s41579-020-00460-0
34. Valles-Colomer M, Falony G, Darzi Y, et al. The neuroactive potential of the human gut microbiota in quality of life and depression. Nat Microbiol. 2019;4:623–632. doi:10.1038/s41564-018-0337-x
35. Zhu Z, Cheng Y, Liu X, et al. The microbiota-gut-brain axis in depression: unraveling the relationships and therapeutic opportunities. Front Immunol. 2025;16:1644160. doi:10.3389/fimmu.2025.1644160
36. Gao L, Zhao X, Wu L, et al. Low-frequency transcranial magnetic stimulation ameliorates anhedonic behaviors and regulates the gut microbiome in mice exposed to chronic unpredictable mild stress. Alpha Psychiatr. 2024;25(4):493–501. doi:10.5152/alphapsychiatry.2024.241561
37. Liśkiewicz P, Pełka-Wysiecka J, Kaczmarczyk M, et al. Fecal microbiota analysis in patients going through a depressive episode during treatment in a psychiatric hospital setting. J Clin Med. 2019;8(2):164. doi:10.3390/jcm8020164
38. Williams BB, Van Benschoten AH, Cimermancic P, et al. Discovery and characterization of gut microbiota decarboxylases that can produce the neurotransmitter tryptamine. Cell Host Microbe. 2014;16(4):495–503. doi:10.1016/j.chom.2014.09.001
39. Willner P. Validity, reliability and utility of the chronic mild stress model of depression: a 10-year review and evaluation. Psychopharmacology. 1997;134(4):319–329. doi:10.1007/s002130050456
40. Pellow C, Pichardo S, Pike GB. A systematic review of preclinical and clinical transcranial ultrasound neuromodulation and opportunities for functional connectomics. Brain Stimul. 2024;17(4):734–751. doi:10.1016/j.brs.2024.06.005
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Early Clinical Experiences of Esketamine Nasal Spray in the UK in Adults with Treatment-Resistant Major Depressive Disorder: Advisory Panel Recommendations
Young AH, Abdelghani M, Juruena MF, Nikolova VL, Nilforooshan R
Neuropsychiatric Disease and Treatment 2023, 19:433-441
Published Date: 24 February 2023
A Systematic Review Assessing Patient-Related Predictors of Response to Transcranial Magnetic Stimulation in Major Depressive Disorder
Gonterman F
Neuropsychiatric Disease and Treatment 2023, 19:565-577
Published Date: 8 March 2023
Traditional Chinese Medicine Ameliorates Depression via the Gut-Brain Axis: A Review Focus on NLRP3/TLR4-Mediated Inflammatory Pathways and Gut Microbiota Modulation
Che L, Xie J, Xia C, Yu Q
Neuropsychiatric Disease and Treatment 2026, 22:579710
Published Date: 8 January 2026