Back to Journals » Neuropsychiatric Disease and Treatment » Volume 18
Orexin/Hypocretin System Dysfunction in ESSENCE (Early Symptomatic Syndromes Eliciting Neurodevelopmental Clinical Examinations)
Authors Knez R ![]() , Stevanovic D
, Stevanovic D ![]() , Fernell E
, Fernell E ![]() , Gillberg C
, Gillberg C ![]()
Received 14 January 2022
Accepted for publication 28 July 2022
Published 15 November 2022 Volume 2022:18 Pages 2683—2702
DOI https://doi.org/10.2147/NDT.S358373
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Dr Roger Pinder
Rajna Knez,1– 3 Dejan Stevanovic,1 Elisabeth Fernell,1 Christopher Gillberg1
1Gillberg Neuropsychiatry Centre, Institute of Neuroscience and Physiology, Sahlgrenska Academy, University of Gothenburg, Gothenburg, Sweden; 2Department of Pediatrics, Skaraborg Hospital, Skövde, Sweden; 3School of Health Sciences, University of Skövde, Skövde, Sweden
Correspondence: Rajna Knez, Gillberg Neuropsychiatry Centre, Institute of Neuroscience and Physiology, Sahlgrenska Academy, University of Gothenburg, Kungsgatan 12, vån 2, Göteborg, 41119, Sweden, Email [email protected]
Abstract: Early Symptomatic Syndromes Eliciting Neurodevelopmental Clinical Examinations (ESSENCE) is an umbrella term covering a wide range of neurodevelopmental difficulties and disorders. Thus, ESSENCE includes attention-deficit/hyperactivity disorder (ADHD), autism spectrum disorder (ASD), and other neurodevelopmental disorders (NDDs) and difficulties, with a variety of symptoms in cognitive, motor, sensory, social, arousal, regulatory, emotional, and behavioral developmental domains, frequently co-occurring and likely having partly common neurobiological substrates. The ESSENCE concept is a clinical paradigm that promotes organizing NDDs in everyday clinical practice according to their coexistence, symptom dimensions overlapping, and treatment possibilities. Despite increased knowledge regarding NDDs, the neurobiological mechanisms that underlie them and other ESSENCE-related problems, are not well understood. With its wide range of neural circuits and interactions with numerous neurotransmitters, the orexin/hypocretin system (Orx-S) is possibly associated with a variety of neurocognitive, psychobiological, neuroendocrine, and physiological functions and behaviors. Dysfunction of Orx-S has been implicated in various psychiatric and neurological disorders. This article provides an overview of Orx-S dysfunctions’ possible involvement in the development, presentation, and maintenance of ESSENCE. We provide a focused review of current research evidence linking orexin neuropeptides with specific clinical NDDs symptoms, mostly in ADHD and ASD, within the Research Domain Criteria (RDoC) framework. We propose that Orx-S dysfunction might have an important role in some of these neurodevelopmental symptom domains, such as arousal, wakefulness, sleep, motor and sensory processing, mood and emotional regulation, fear processing, reward, feeding, attention, executive functions, and sociability. Our perspective is presented from a clinical point of view. Further, more thorough systematic reviews are needed as well as planning of extensive new research into the Orx-S’s role in ESSENCE, especially considering RDoC elements.
Keywords: attention-deficit/hyperactivity disorder, autism spectrum disorder, developmental coordination disorder, neurodevelopmental disorders, research domain criteria, orexin, hypocretin
ESSENCE as a Clinical Paradigm
Early Symptomatic Syndromes Eliciting Neurodevelopmental Clinical Examinations (ESSENCE) is an umbrella term covering a wide range of neurodevelopmental difficulties and neurodevelopmental disorders (NDDs), such as attention-deficit/hyperactivity disorder (ADHD), developmental coordination disorder (DCD), and autism spectrum disorder (ASD), with a variety of symptoms in cognitive, motor, sensory, social, arousal, regulatory, emotional, and behavioral developmental domains, which frequently co-occur and likely have partly common neurobiological substrates.1 There is a variety of disorders, syndromes, and disabilities under the ESSENCE umbrella, as schematically presented in Figure 1. ESSENCE disorders are highly prevalent, together affecting about 10% of school-age children and more frequently males, and in many cases persist into adulthood.2
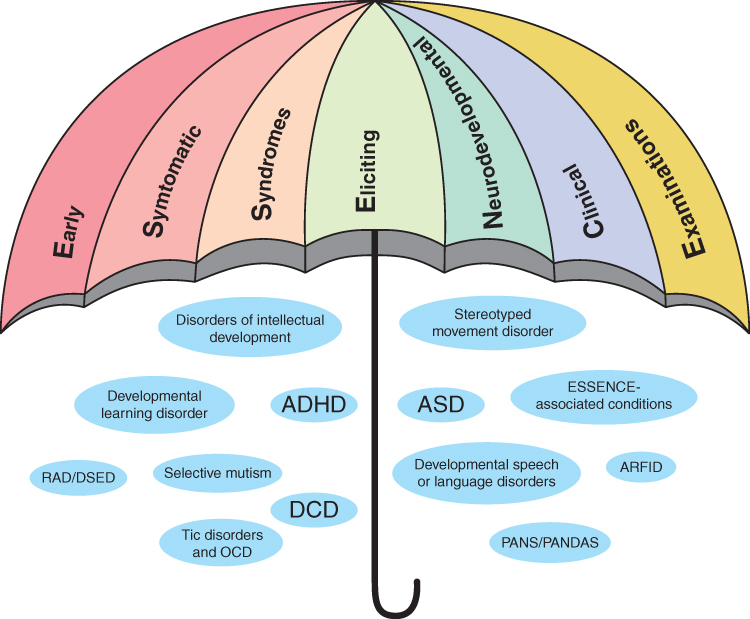 |
Figure 1 Early Symptomatic Syndromes Eliciting Neurodevelopmental Clinical Examinations (ESSENCE) diagnoses and conditions. Abbreviations: ADHD, attention-deficit/hyperactivity disorder; DCD, developmental (motor) coordination disorder; ASD, autism spectrum disorder; RAD, reactive attachment disorder; DSED, disinhibited social engagement disorder; ARFID, avoidant-restrictive food intake disorder; PANS, paediatric acute-onset neuropsychiatric syndrome; PANDAS, paediatric acute-onset neuropsychiatric disorder associated with streptococcal infection; OCD, obsessive-compulsive disorder. Notes: Developmental learning disorder, learning disorders with impairment in reading, written expression, or mathematics; ESSENCE-associated conditions, behavioural phenotype syndromes (eg Down’s syndrome, fragile X syndrome and premutation FraX, sex chromosome anomalies, tuberous sclerosis complex, 22q11 deletion syndrome, Prader-Willi and Angelman syndrome, neurofibromatosis, foetal alcohol spectrum disorder/syndrome, valproate acid syndrome), neurological conditions and disorders (eg epilepsy or seizures, cerebral palsy, disorders of muscles), severe early-onset mental disorders (eg early-onset bipolar disorder, schizophrenia, disruptive mood dysregulation disorder). |
On one hand, the concept behind ESSENCE could be viewed as a clinical paradigm as it promotes organizing diagnoses according to their coexistence seen in everyday clinical practice and their possible common underlying neurobiology. The use of categorical diagnoses and classifications, thus far, has failed NDDs.3 Instead, we should approach these disorders dimensionally. Applying a “spectral approach” to understand NDD symptoms, measure the impact that such symptoms have on an individual's functioning and quality of life, and suggest optimal intervention strategies requires that the paradigm used in clinical practice be both comprehensive and easy to understand. ESSENCE might also serve as a framework that helps researchers understand the need for a comprehensive approach to questions of possible common neurobiological substrates for the disorders under the ESSENCE umbrella. Rather than focusing on isolated or subspecialized parts that might provide only limited insight into research problems and partial answers, the ESSENCE conceptualization invites researchers to take a broader perspective.
On the other hand, the Research Domain Criteria (RDoC) represent a research paradigm with a possible clinical utility. The RDoC framework for psychopathology research relies on genetics and neuroscientific research tools to reveal biosignatures,4,5 representing an alternative to the more classical categorical approach to the conceptualization of psychopathologies. The dimensional psychopathology measures that the RDoC framework uses include “indicators of functional brain disruption”.6 The RDoC system is designed to develop a biologically oriented classification7 of mental and behavioral disorders, and it is organized in a matrix of six domains: cognitive systems, negative and positive valence systems, systems for social processes, arousal/regulatory systems, and sensorimotor systems, each with several constructs/subconstructs.8
In clinical practice, deconstructing NDDs down into their “target symptoms” based on the ESSENCE and RDoC paradigms combined, may guide practitioners in choosing optimal therapeutic strategies. Deconstructing a psychiatric disorder into its specific symptoms, which may involve unique neurobiological mechanisms, could lead to personalized psychopharmacological therapeutic strategies,9,10 which could be relevant for NDDs conditions, too.
Neurobiological Mechanisms That Underlie ESSENCE
There are several medical, neurological, and psychiatric disorders, conditions, and symptoms that coexist with ADHD and ASD, such as oppositional defiant disorder, conduct disorders, substance use disorder, anxiety, depression, DCD, tic disorders, obsessive-compulsive disorder (OCD), disorders of intellectual development and language, epilepsy, gastrointestinal problems, and sleep disorders.1,2,11 Finding a shared biological substrate of ESSENCE that might account for the overlap of symptoms could guide clinicians in choosing optimal treatment strategies.
Despite increased awareness of NDDs, the neurobiological mechanisms underlying NDDs are not well understood. Recently, orexin/hypocretin system (Orx-S) dysfunction has been implicated in a variety of psychiatric and neurological disorders,12 including ADHD and ASD.13–18 Kohyama was likely the first to connect the Orx-S with both ASD and ADHD, suggesting that Orx-S hyperactivity might play a role in insomnia in these two classical NDDs.17
The current paper aims at providing a new perspective and an overview of possible involvements of Orx-S dysfunction in the development, presentation, and maintenance of NDDs and some other ESSENCE conditions, focusing on specific neuropsychiatric symptoms—mostly in ADHD and ASD—in the context of the RDoC framework. Currently, there is no consensus regarding what kinds of symptom dimensions best capture ADHD and at which levels these dimensions should be assessed.19 Furthermore, ADHD is heterogenous in its presentation,20 which motivates the use of the RDoC approach when aiming to close the gap in our understanding of symptoms and causes.6 ADHD spans the entire RDoC matrix, and individuals with ADHD may differ in the extent to which their neurological processes are affected in the multiple brain systems involved.6,7 Regarding ASD’s phenotypic heterogeneity, the RDoC framework also offers an alternative way of characterizing ASD features, representing a continuum that extends into the “normative” span.4 Furthermore, Ibrahim and Sukhodolsky pointed out that “current research using the RDoC framework in ASD has suggested conceptualizing ASD symptoms along positive, negative, and cognitive dimensions, similar to a dimensional approach traditionally used for schizophrenia”.4
Autism is officially conceptualized as a spectrum disorder,21,22 whereas ADHD is still viewed categorically, although its symptomatology might also be better conceptualized as a spectrum than a discrete diagnostic entity.20,23 Finally, considering ADHD and ASD as related spectra may help identify their partly common neurobiological substrates, and Orx-S dysfunction might be one such common pathway.
Orexins/Hypocretins
The orexins, ie orexin-A/hypocretin-1 (Orx-A) and orexin-B/hypocretin-2 (Orx-B), are neuropeptides that originate in the lateral hypothalamus.24–27 Orexins act on orexin type 1 receptor (Orx1R) and orexin type 2 receptor (Orx2R), which are “widely distributed across the brain”, but their mRNA has also been determined in peripheral tissues.26–29 Outside of the central nervous system, the Orx-S could be linked to the peripheral nervous system and several peripheral tissue types (ie, the gastrointestinal tract, pituitary gland, pancreas, adrenal gland, adipose tissue, and male reproductive system).30–32 Orx1R shows a higher affinity for Orx-A than Orx-B, while Orx2R binds both orexins with similar affinity.24 The different distribution patterns of the two orexin receptor subtypes suggest different physiological roles33 and likely non-overlapping and/or opposite functions, such as anxiolytic versus anxiogenic responses.25,34
Orexin-producing cell bodies reside in the hypothalamus, but their projections are widely spread through the brain and spinal cord with dense projections throughout the hypothalamus and the basal forebrain, specifically in cholinergic areas.25,35 Orexins produced by cells in the hypothalamus are released in various brain areas.26 These areas include monoamine neurotransmitter centers in 1) the tuberomammillary nucleus (for histamine), 2) in the ventral tegmental area (for dopamine), 3) the locus coeruleus (for noradrenalin), 4) pedunculopontine tegmental and laterodorsal tegmental nuclei (for acetylcholine), and 5) the raphe nucleus (for serotonin).26,28,36,37 Orexin projections seem to also impart effects on 6) glutamate release from the thalamus and the prefrontal cortex and on 7) gamma-aminobutyric acid (GABA) release from the nucleus accumbens and the striatum.36 Orexin neurons receive numerous input signals linked to environmental, physiological, and emotional stimuli and from regions related to sleep–wake states, circadian phase, motivation, and visceral cues.24,38 Thus, the Orx-S may interact with the noradrenergic, dopaminergic, serotonergic, histaminergic, cholinergic, glutamatergic, and GABAergic systems.24–28,36–39 For example, manipulations of the Orx-S simultaneously modulate neurotransmitters associated with alertness and wakefulness (ie, dopamine, norepinephrine, acetylcholine, and histamine).27
GABA is the major inhibitory neurotransmitter in the brain.26,40 “Glutamate is the major excitatory neurotransmitter in the central nervous system”, sometimes considered the “master switch” due to its potential ability to excite almost all neurons in the brain.26 Interestingly, the effects of some neurotransmitters change during embryogenesis; for example GABA is excitatory in the embryo but inhibitory postnatal.41 The GABA shift seems to be deleyed or absent in some individuals with NDDs.40 In addition to GABA, glutamate seems important in ASD and ADHD, as evidence has implicated relative loss of GABA inhibitory action with “corresponding glutamate-mediated hyper-excitation in the development” of these NDDs.42 Orexin neurons may have capability of glutamate co-release and be “considered to have a glutamatergic phenotype”.43 However, a subpopulation of orexin neurons seems to have the ability to synthesize GABA.43 Thus, it is implied that the Orx-S may play a role in the complex interplay and balance of glutamatergic and GABAergic transmission.
Orexins' function in metabolic pathways is not yet fully understood,24 although it is hypothesized that the central Orx-S could modulate whole-brain activity.33 Orexin receptor signalling is more diversified than originaly considered.30,31 The Orx-S is associated with a variety of neurocognitive, psychobiological, neuroendocrine, physiological functions and behaviors. Suggested regulation/modulation roles of the Orx-S include arousal, vigilance states, sleep–wakefulness, somatic motor control, ventilation modulation, locomotion and spontaneous physical activity, stereotypical and obsessive-compulsive behaviors, enhanced hippocampal neurogenesis, various autonomic process, affect, mood and emotional regulation, fear and anxiety (anxious behaviors), motivation and reward (reward processing), reward-seeking/addictive behavior, stress processing (regulations of behavioral and neuroendocrine responses during stress) and stress resilience, appetite modulation (food intake) and feeding (feeding behaviors), gastrointestinal and reproductive functions, the micturition reflex, thermoregulation/thermogenesis, energy homeostasis and metabolism (metabolic regulation), cognitive function and cognitive flexibility, attention and decision making, memory, and (aversive) learning.12,24–28,31,33–36,38,39,44–53 Orx-S dysfunctions have been suggested to underlie various neurological and neuropsychiatric disorders, and psychopathological conditions.12,24,46 These include depressive and anxiety disorders, sleep and eating disorders, substance addictions, and post-traumatic stress disorder; whereas changes in orexin levels have been reported in depressive and bipolar disorders, narcolepsy, age-related cognitive decline, anorexia nervosa (AN), ADHD, schizophrenia, and neurodegenerative brain diseases, such as Parkinson’s disease, Alzheimer's disease (AD), Huntington's disease, amyotrophic lateral sclerosis, and multiple sclerosis.12,13,24,25,27,32,38,46,54–56 The Orx-S is also involved in metabolic syndrome, chronic inflammations, and cancers; moreover, orexin has a proposed role on “inverse comorbidity between cancer and neurodegenerative disorders”.32,56
The Orx-S is a potential therapeutic target for several disorders.27 Orexin receptor antagonists (ie, suvorexant and lemborexant) have been approved for the treatment of insomnia, while daridorexant is currently close to registration for the same indication.27 There are many registered clinical trials that explore suvorexant for the treatment of insomnia in different patient populations (ie, adolescents, those with fibromyalgia, and bipolar disorder).27 Of special interest is the suggestion that orexin receptor antagonists might be promising for treatment of insomnia in adolescents,57 and children with NDDs.58 In addition to insomnia, the potential and effectiveness of orexin-based therapeutics in the treatment of other conditions and disorders (ie major depressive disorders, anxiety disorders, panic disorder, post-traumatic stress disorder, eating disorder, addiction/substance-use disorder, ADHD, narcolepsy, some type of excessive daytime sleepiness, chronic fatigue syndrome, and cognitive impairments related to ageing, neurodegenerative disease and sleep deprivation) have also been implicated.25,27,34,57–61 Orexin antagonists may reduce motor impulsivity induced by psychostimulants and may diminish “sympathetic overactivity during withdrawl syndrome”.12,62
The Orexin System and ESSENCE
Cortese et al proposed that orexin neurons in the perifornical and dorsomedial hypothalamus may be hypoactive in ADHD, while those in the lateral hypothalamus may be overactivated.14 Recently, Baykal et al reported that children with ADHD had significantly lower Orx-A serum levels than neurotypical children.13 In addition, orexin levels were found significantly lowered in inattentive than in hyperactive and combined ADHD.
Kohyama presumed an increase in orexinergic system activity to be involved in insomnia symptoms in ADHD and in ASD.17 Messina et al, in a single-case, ASD study, showed that plasma Orx-A levels were higher than reference values,18 while Kobylinska et al found the distributions of plasma levels of Orx-A to be variable in children with ASD but similar to those of neurotypical children.16 The same study found that the lower the plasma levels of Orx-A, the greater the severity of ASD symptoms would be, and vice versa (moderate correlations). Thus, the role of orexin in ASD pathogenesis has also recently been implicated, although results are very preliminary.
Orx-S Dysfunction and Main Symptom Domain of ESSENCE Within the RDoC Framework
At present, direct connections between orexin and certain RDoC elements have been established for the negative valence system and positive valence system domains, and domain arousal/regulatory system.8 The Orx-S with its two peptides, two types of receptors, orexin neurons projections to many parts of the brain, and possible interactions with a range of neurotransmitters may affect many systems and behaviours in NDDs. Here, we focus on symptom domains that might be the most relevant for ADHD and ASD (Table 1). Some of these symptoms, such as fear, reward, and arousal already belong to RDoC domains connected with orexin.
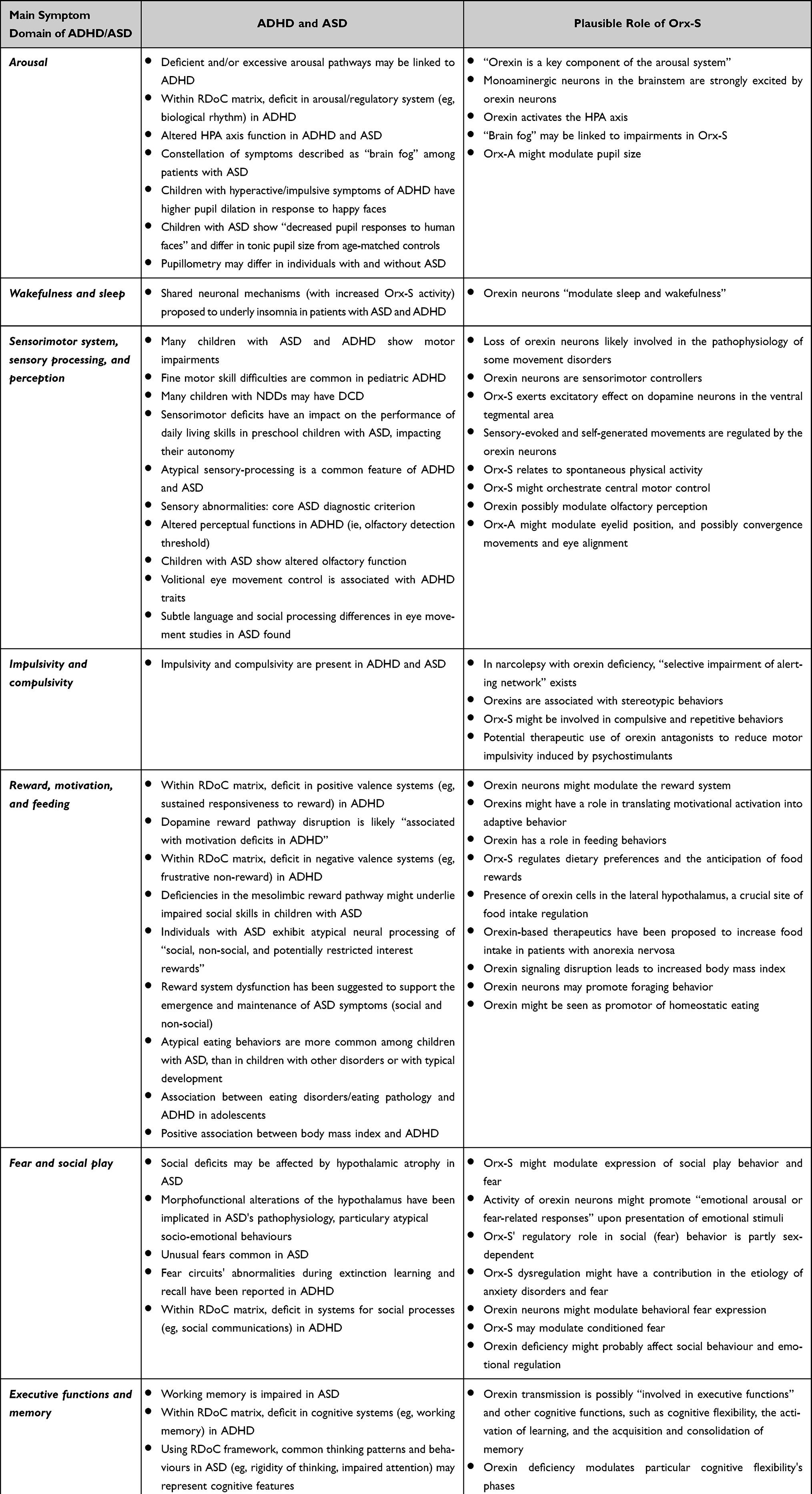 |
Table 1 ADHD and ASD Symptoms and Plausible Orexin/Hypocretin System (Orx-S) Involvement |
Arousal
Direct connection between orexin and the domain arousal/regulatory system (construct: arousal, and construct: sleep-wakefulness) is already established.8 “Orexin is a key component of the arousal system”,36 and Orx-S is a “key player in arousal stability”.27 Monoaminergic neurons in the brainstem are strongly excited by orexin neurons,69 and this might represent the main plausible explanation for orexin’s potential role in NDDs. On the other hand, the excitation of orexin neurons by external or internal stimuli leads to increased neurotransmission and activation of brain regions/circuits and peripheral responses, ie, including promotion of attention, cognition, and learning, as well as activation of the hypothalamus-pituitary-adrenal (HPA) axis.36
Deficient and/or excessive arousal pathways may be linked to ADHD,10 ie, “reduced HPA axis responsiveness to stress”.64 The intranasal administration of Orx-A may enhance arousal and, by consequence, alleviate cognitive impairments induced by hypoarousal (eg, attentional impairment due to sleep deprivation).25 Theoretically, based on estimated orexin levels in patients with ADHD and resultant impairments, orexin replacement (ie, agonists) or orexin antagonist therapy may be needed to target the dysfunctionality of the Orx-S. Baykal et al found that Orx-A levels were significantly lower in the inattentive ADHD subtype, compared to the hyperactive and combined subtypes.13 It seems reasonable to suggest that the choice of orexin-based therapeutic agents might be guided by the predominance of inattention or hyperactivity/impulsivity, but also by possible coexisting sleep problems, which might suggest whether or not imbalance in the arousal system is due to hypo- or hyperstimulation. Furthermore, pupil size reflects the arousal network's activity,106 and might be used as an indirect indicator of arousal level. Thus, pupillometry belongs to the diagnostic procedure in the RDoC domain arousal/regulatory system.8 Orx-A might modulate pupil size.71 This may be important in the context of NDDs as children with hyperactive/impulsive symptoms in ADHD have increased pupil dilation in response to happy faces,66 while children with ASD show “decreased pupil responses to human faces” and tonic pupil size differences compared to age-matched controls (ie, larger pupil size in ASD).67 A recently published meta-analysis revealed that measuring pupil size and reactivity differs in individuals with and without ASD.68
According to Taylor and Corbett, the HPA axis is slow to respond to physical or physiological manipulations in ASD.63 A key, common finding is hyper-responsiveness in otherwise benign social situations or in those involving unpleasant stimuli, and hypo-responsiveness to stressors involving social-evaluative threat.63 Overall cognitive dysfunctions may result from an increase in arousal (ie, overstimulation), but diminished arousal (ie, understimulation) may lead to inattentiveness and cognitive dysfunction.10,26,36 In summary, compounds that activate the Orx-S usually represent a treatment option for hypoarousal, while compounds that suppress this system are utilized in treatment of hypervigilance (ie, insomnia and addiction-related hyperarousal).28
The arousal spectrum of sleep and wakefulness26,36 could represent a spectrum of a possible dysfunctionality of the Orx-S,70 considering that both too much and too little stimulation may lead to different levels of arousal and, eventually, different levels of cognitive dysfunction. The constellation of various cognitive symptoms, including cognitive impairment, which can be described as “brain fog”,107 found also among patients with ASD,65 may be potentially linked to impairments in the Orx-S.70
Wakefulness and Sleep
In addition to two key neurotransmitters (ie, histamine from the tuberomammillary nucleus and GABA from the ventrolateral preoptic [VLPO] nucleus), the lateral hypothalamus' orexin-containing neurons and the suprachiasmatic nucleus' melatonin-sensitive neurons are also responsible for the regulation of the sleep/wake switch.26,35,36 Orexin neurons, as histamine neurons do, modulate weakfulness and sleep,35 and antihistamine agents block orexin’s excitatory effect.12 Benzodiazepines act by sensitizing GABA receptors,35 while orexin antagonists do not act directly on them.36 GABAB receptors on orexin-producing neurons are necessary to stabilize and consolidate sleep/wake states,108 and paradoxical reactions to benzodiazepines109 might reflect possible Orx-S abnormality in individuals with NDD. Suvorexant, used in treatment of insomnia, represents a novel mechanism of action, by “inhibiting the wakefulness-promoting orexin neurons”.35 The orexin receptor antagonists have more favourable adverse effects than conventional hypnotics,27 and may be treatment options if traditional hypnotics are contraindicated (ie, benzodiazepines).28
Kohyama proposed shared neuronal mechanisms underlying insomnia in ASD and ADHD, which possibly involve increased Orx-S activity, reduced serotonergic and melatonergic system activity, and rapid eye movement sleep reduction.17 Narcolepsy and neuropsychiatric disorders may share some clinical features, with an orexin deficiency proposed as a pathophysiological mechanism.48 Narcolepsy, one of hypersomnolence's central disorders, is classified into two types: narcolepsy type 1 (ie, NT1; narcolepsy with cataplexy [characterized by cataplexy and cerebrospinal fluid Orx-A deficiency], and narcolepsy type 2 (i.e, “narcolepsy without either cataplexy or orexin deficiency”).48,110 The profound loss of orexin neurons in NT1 may result from exposure to antigens and, consequently, autoimmune reactions that result in the selective loss or dysfunction of orexin neurons in the lateral hypothalamus of genetically predisposed individuals.26,36,50,110 NT1 may be considered as a hypothalamic disorder that affects “motor, psychiatric, emotional, cognitive, metabolic, and autonomic functions”; thus, it may not affect only sleep–wake regulation.110
Symptoms of ADHD frequently co-occur with narcolepsy, accounting for more than 30% of cases.111 On the other hand, children with narcolepsy experience treatment-resistent ADHD symptoms at high levels.112 Narcolepsy may be more common in ADHD than among the general population; however, it remains unclear what the proportion of undiagnosed narcolepsy is in patients with ADHD.113 Furthermore, ADHD and narcolepsy may be genetically related.114 Although based on a limited number of cases, a pathogenetic link between ASD and narcolepsy has been suggested.115 It has also been argued that social functioning is commonly impaired in children with NT1, especially in girls, and it has been suggested that orexin deficiency affects emotional regulation and social behavior, in addition to causing cataplexy and hypersomnolence in children with narcolepsy.103
Sensorimotor System, Sensory Processing, and Perception
The rapid loss of orexin neurons is likely “involved in the pathophysiology of the movement disorders”, frequently observed at disease onset in childhood NT1, while secondary dopaminergic abnormalities might also be involved.50 The orexin exerts an excitatory effect on dopamine neurons in the ventral tegmental area.81 According to Burdakov, sensory-evoked and self-generated movements are regulated by the orexin neurons.80 Even more, the Orx-S is related to spontaneous physical activity.24 Given that several motor deficits are associated with the loss of orexins and that Orx-S may orchestrate central motor control,51 motor phenomena might also be conceptualized as a spectrum, with the Orx-S playing an important role.70
Atypical sensory processing is a common feature of many neurodevelopmental disabilities, including ADHD and ASD.2,75 While sensory abnormalities have been included in the most recent edition of the Diagnostic and Statistical Manual of Mental Disorders in the core diagnostic criteria for ASD, this is not the case for ADHD.21,75 Nevertheless, the similarity in sensory symptoms imply that there may be a common neuronal mechanism of sensory dysfunction across developmental disabilities.75
Altered touch and pain processing have been tentatively confirmed in several psychiatric conditions, with pain processing in ASD and ADHD being a focus of research interest.116 While hypersensitivity to pain is reported in a single study on ADHD, the findings are inconsistent in several studies in ASD.116 Psychiatric conditions' underlying pathophysiology could be elucidated by a better understanding of pain processing in them, particularly due to the psychobiological overlap between these pathways.116 Pain regulation closely associates with the Orx-S.117,118 Manipulation of orexin neurons influences pain-related behaviors; therefore, activation of the Orx-S has been proposed as a novel approach to analgesic treatment.28 In addition, suvorexant has shown efficacy in improving sleep time and reducing next-day pain sensitivity, such as in patients with fibromyalgia and comorbid insomnia.119
All levels of rats' olfactory system include both orexins and orexin receptors, suggesting a possibility that these neuropeptides modulate olfactory perception.82 People with ADHD have been shown to have altered perceptual functions such as increased olfactory detection threshold.76 Relative to typically developing peers, children with ASD also demonstrate altered olfactory function.77
Orx-A might modulate eyelid position, and possibly convergence movement and eye alignment.71 Volitional eye movement control is associated with ADHD traits—more specifically, inattentive traits with premature anticipatory eye movements—and this phenotypic association appears attributable to shared genetic etiology.78 Several eye movement studies provide evidence of subtle processing differences in both language and social processing with potential to impact everyday communication in ASD.79 Attention levels during visual exploration might vary, and thus, “depending on intrinsic states”, eye movement may represent attentional load.120
Impulsivity and Compulsivity
Impulsivity and compulsivity are “forms of cognitive inflexibility”.26 Both impulsivity and compulsivity are symptoms of many NDDs, with an example of ADHD, ASD, Tourette’s syndrome/tic disorders, OCD, and stereotyped movement disorders to be possible categorized as impulsive–compulsive disorders, based on impulsivity and compulsivity endophenotypes present.26 Filardi et al showed more severe ADHD hyperactive–impulsive symptoms present in patients with NT1 versus in narcolepsy type 2 and healthy controls, indicating orexin's direct role in modulating impulsivity.83 Furthermore, stereotypical behaviors are also associated with orexins.12 The involvement of the Orx-S in compulsive and repetitive behavior is indicated by several preclinical findings,49,84 and study in rats has suggested the potential therapeutic use of orexin antagonists to reduce motor impulsivity induced by psychostimulants.62
Reward, Motivation, and Feeding
It has been indicated that orexin neurons modulate the reward system24,26 and proposed orexin’s role in translating motivational activation into adaptive behaviors.44 There is establish connection between orexin and RDoC positive valence system domains (construct: reward responsiveness; subconstruct: initial response to reward).8 In ADHD, a deficit in sustained responsiveness to reward (RDoC - positive valence system) and frustrative non-reward (RDoC – negative valence system) are present7 and dopamine reward pathway's disruption is thought to be in relation with motivation deficits.85 Regarding ASD, it has been shown that deficiencies in the mesolimbic reward pathway underlie impaired social skills in children with this disorder.86 Individuals with ASD also exibit atypical neural processing of “social, non-social, and potentially restricted interest rewards”.87 Moreover, reward system dysfunctions have been suggested to support the emergence as well as the maintenance of ASD symptoms, both social and non-social.88 The impaired motivational process, affecting both social and non-social rewards, has been shown to possibly characterize ASD.121
In the context of motivation and reward, it should be mentioned that orexin plays a role in feeding behaviors.27,36,92 Orexin neurons may promote “phenomena involved in successful foraging, including food-anticipatory locomotor behavior, olfactory sensitivity, visual attention, spatial memory, and mastication”.95 Orexin might stimulate food intake, motivation for food rewards and promote binge eating.27,28 Taking the orexin as promotor of homeostatic eating95 opens possibilities to consider the Orx-S involvement in the whole spectrum of eating disturbances observed in NDDs.
The Orx-S regulates both dietary preferences and the anticipation of food rewards.93 This is of particular interest, as more than 70% of children with ASD have atypical eating behaviors, with limited food preferences being the most common feature.89 In addition to ASD,89 few studies have found an association between eating disorders/pathology and ADHD.90,122,123 During childhood, individuals with ADHD could be at risk for disordered eating, while eating disorders - including AN, bulimia nervosa, and binge eating - may occur later on.122 Changes in dopamine receptors have been connected with reward system insufficiency.123 Reward deficiency syndrome might link genetic differences in the dopaminergic system to behavioral phenotypes and represent “a transdiagnostic feature between ADHD, eating disorders, obesity, and substance-abuse disorders”,122 all of which are associated with the Orx-S. Cortese et al have been suggested the Orx-S role in feeding behaviors in ADHD.14 Orexin-based therapeutics have been proposed to increase food intake in patients with AN.25 Neither AN nor bulimia nervosa are currently grouped within ESSENCE. On the other hand, avoidant-restrictive food intake disorder (ARFID), highly comorbid with ASD,124 is extremely common in ESSENCE,2 and ADHD might be associated with restrictive eating, too.125 Generally defined as restriction of food intake that leads to functional impairment,2 ARFID could be considered a disorder related to impairment of the Orx-S.
Some studies have shown increased rates of neurodevelopmental problems among children with obesity,126 and a higher prevalence of obesity in children and adolescents with ASD than in a neurotypical controls.127 In addition, 43% of the children who are overweight or obese normalized weight when receiving stimulant treatment for their ADHD.128 As orexin modulates obesity and reward,129 the findings of a recent study of a positive association between body mass index and ADHD,91 might be of interest. The authors proposed that genetic factors related to body mass index—particularly reward system-related ones—might be involved in this association. Conceptualizing eating pathologies as spectra and the role of the Orx-S in motivation and reward, Orx-S dysfunction could be considered a “common pathway” for associations of feeding/eating disturbances/disorders with NDDs.
As we have already mentioned, the role of orexin has been recognized within positive valence system.8 This is of special importance, as the complex neurobiology of anhedonia may involve dysfunction of the pathways regulating reward and motivation.26,130,131 In the context of anhedonic phenotype132 it might be of interest that the orexinergic innervation of the ventral palladium seems to be of importance in hedonic valence, during which processing, the orexin's signaling might be amplified by the insula.28 Reduced orexin-signaling might be implicated in anhedonia.27 Several psychopathologies (ie, ADHD and major depressive disorder) have been associated with low hedonic tone,131 with hedonic responsiveness being what depressive and ADHD's innatentive symptoms might have as a common endophenotype.133 Hedonic deficits are also reported in the context of a broader autism phenotype, as autistic traits were associated with social hedonic capacity being reduced in a general population.134 In addition, anhedonia is a component of depression and schizophrenia.135 Considering that disorders of intellectual development, ASD, and ADHD, as childhood NDDs, and schizophrenia and bipolar disorder, as adult psychiatric disorders, could be conceptualized as lying on a same neurodevelopmental continuum,3 a hedonic responsivity might be of interest for their etiology and differential diagnostic challenges. In summary, anhedonia or hedonic valence/tone and Orx-S function might represent an important research area, in this regard.
Fear and Social Play
Another RDoC domain, the negative valence system domain (construct: acute threat “fear”) has also been connected to orexin.8 In animal models, the Orx-S modulates the expression of social play behaviors and fear.45,100 In addition, orexin neurons fire maximaly during exploratory behavior.136 Moreover, the activity of orexin neurons might promote responses related to fear or emotional arousal upon presentation of emotional stimuli.24 Orx-S' regulatory role in social (fear) behavior is partly sex-dependent, with observed reduced sociability and decreased social novelty behavior in orexin-deficient female animal.45 However, the same study shows that “prolonged expression of social avoidance after social fear conditioning” could be attributed to orexin deficiency in both sexes.
Wolfe et al suggested that numerous neurophysiological functions and phenotypes connected to social deficits may be affected by hypothalamic atrophy and may explain symptomatologic commonalities in ASD and several other psychiatric diagnoses.96 The severe socioemotional dysfunctions following hypothalamic lesions, and morphofunctional alterations of the hypothalamus have been implicated in ASD's pathophysiology, particulary atypical socio-emotional behaviour.97
The Orx-S modulates fear (threat) learning by acting “upstream of the amygdala via the noradrenergic locus coeruleus”.60,101 Thus, Orx-S dysregulation might have a contribution to the etiology of anxiety disorders and fear.101 Orexin neurons in the hypothalamus, when activated by fearful stimuli, increase sympathetic nervous outflow,60 while at the same time, the lack of orexin might inhibit sympathetic activity.137 The Orx-S may modulate conditioned fear, and selective Orx1R antagonism might be a treatment option for disorders related to fear.102 Unusual fears have long been identified as common among patients with ASD.98 Recently, fear circuits' abnormalities during extinction learning and recall have been reported in medication-naïve, non-traumatized subjects with ADHD.99
Executive Functions and Memory
A role of orexin in cognitive processes and executive functions has been suggested.27,62 Its deficiency modulates particular cognitive flexibility's phases,105 as orexin plays an important part in activation of learning, and acquisition and consolidation of memory.12,94 Working memory and/or cognitive control—two RDoC subconstructs associated with cognitive system—are impaired in ADHD, ASD, mild intellectual disability, and borderline intellectual functioning.7,104,138,139 Several studies have shown an increased risk of developing neurodegenerative disease during later life for patients with a history of ADHD,140 including a large multi-generation cohort study in Sweden that associated ADHD with AD and any dementia across generations.141 However, well-designed prospective longitudinal studies are necessary to reassure the validity of estimated neurodegenerative risk for patients with ADHD.140 Future studies would preferably also investigate the possible role of an underlying Orx-S dysfunction.
The Orx-S Dysfunction in Some Conditions Within or Associated with ESSENCE
Other than ADHD and ASD, there are several other neurodevelopmental conditions under the ESSENCE umbrella—as well as ESSENCE associated conditions—in which Orx-S dysfunction may have a role (Figure 1; Table 2). This brings to mind the proposed diagnosis of deficit in attention, motor control, and perception that has been in clinical use in Scandinavia,142 although it is not recognized as an official diagnostic category. Nowadays, clinicians usually use DCD (or Developmental motor coordination disorder22) with associated ADHD, instead. However, DCD is still one of the most neglected group of problems under the ESSENCE umbrella as many clinicians fail to recognize it, and this has an impact on providing effective interventions.2 Other developmental abnormalities accompanying the sensorimotor disability of DCD143 may be a clinical manifestation of dysfunctionality in the Orx-S. In addition, deficits in attention, motor control, and perception may be co-occuring with a variety of problems and overlapping conditions, including depression and anxiety,142 diagnosis that already have been associated with Orx-S dysfuntion. Altogether, this underlies the need to focus more attention on DCD in clinical practice and in research.2 Evaluating the role of Orx-S in association across several symptom domains, viz, attention, motor control, and perception, might be of importance in understanding the basic neurobiology of DCD and associated ADHD.
 |
Table 2 Proposed Role of the Orx-S in Disorders and Conditions Associated with ESSENCE |
Plausible Explanation Models for Orx-S Dysfunction Pathways Within ESSENCE
Our understanding of Orx-S potential and implications in pathophysiology of ESSENCE is complicated by the several aspects. These include orexin receptor signaling's high level of diversity, the broad spread of orexin neurons projections within the brain, orexin’s ability to trigger responses in many tissues in addition to the central nervous system, its sexual dimorphisms, and the orexin physiology significantly differing among species.25,29–31,45,105,169,170 However, there are several hypothetical considerations for the Orx-S dysfunction pathways in ESSENCE, raised as research questions. In Table 3 are given a few areas that might be of interest, and were not mentioned previously in this paper.
 |
Table 3 Research Questions Regarding the Role of Orx-S Dysfunction in ESSENCE, that Require Further Studies |
Conclusion and Implications
We propose that Orx-S dysfunction might account for many symptoms in a variety of ESSENCE, especially in ADHD and ASD. As one of the key homeostatic neuropeptides linked to arousal, wakefulness, sleep, motor and sensory processing, mood and emotional regulation, fear processing, reward, feeding, attention, executive functions, and sociability – orexin might represent a missing puzzle-piece in the etiology of several disorders under the ESSENCE umbrella. In addition to the already proposed role of Orx-S dysfunction in ADHD and ASD, which we have attempted to highlight here, we would also like to stress the need for broader conceptualization of Orx-S dysfunction as a possible pathophysiological pathway in several other ESSENCE-associated conditions, potentially contributing to a new perspective in the whole field of neuropsychiatry/neurodevelopmental research.
More research is now needed to systematically review the available literature, and confirm the explanation model that our perspective suggests. Our paper has clear limitations, the main of which lies in the fact that this article is not a systematic study of all the available literature, and it could be affected by our own understanding of the topic and our subjectivity in selecting relevant evidence from published studies. Furthermore, our perspective has here been presented from a clinical point of view by clinicians; child and adolescent psychiatrists and child neurologists, in the future, this type of review should involve preclinicians and neuroscientists in the context of a broader neurotranslational research.
The main implications of our perspective paper are that there is (1) a need for a deeper, more systematically driven review that will include basic neurochemistry, neurobiology, and symptoms expressions; (2) a need for empirical, clinical studies testing proposed models, and (3) a need for testing available orexin agents in treatment studies of individuals affected by ESSENCE.
We hope that our perspective might serve as a glance through the window of opportunity to point researchers in directions that might provide new explanations and therapeutic possibilities to clinicians and, most importantly, generate new possibilities to help our patients obtain their maximal capacity and functionality and live a life of the best possible quality, which is essential in ESSENCE.
Abbreviations
AD, Alzheimer's Disease; ADHD, Attention-Deficit/Hyperactivity Disorder; AN, Anorexia Nervosa; ARFID, Avoidant Restrictive Food Intake Disorder; ASD, Autism Spectrum Disorder; ASLO, Anti-Streptolysin O; BPS, Behavioral Phenotype Syndromes; CFS, Chronic Fatigue Syndrome; COVID-19, Coronavirus Disease 2019; DCD, Developmental Coordination Disorder; DSED, Disinhibited Social Engagement Disorder; ESSENCE, Early Symptomatic Syndromes Eliciting Neurodevelopmental Clinical Examinations; FAS, Fetal Alcohol Syndrome; FASD, Fetal Alcohol Spectrum Disorder; GABA, Gamma-Aminobutyric Acid; GAD65, Glutamic Acid Decarboxylase 65; hEDS, Hypermobile Ehlers-Danlos Syndrome; HLA, Human Leukocyte Antigen; HPA, Hypothalamus-Pituitary-Adrenal; HRV, Heart rate variability; HSD, Hypermobility Spectrum Disorder; LH, Lateral Hypothalamus; ME, Myalgic Encephalomyelitis; NDD, Neurodevelopmental Disorder; NT1, Narcolepsy Type 1; OCD, Obsessive-Compulsive Disorder; ODD, Oppositional-Defiant Disorder; Orx-A, Orexin-A/Hypocretin-1; Orx-B, Orexin-B/Hypocretin-2; Orx1R, Orexin/Hypocretin Type 1 Receptor; Orx2R, Orexin/Hypocretin Type 1 Receptor; Orx-S, Orexin/Hypocretin System; PANDAS, Paediatric Acute-Onset Neuropsychiatric Disorder Associated with Streptococcal Infection; PANS, Pediatric Acute-Onset Neuropsychiatric Syndrome; RAD, Reactive Attachment Disorder; RDoC, Research Domain Criteria; VLPO, Ventrolateral Preoptic.
Disclosure
The authors report no conflicts of interest in this work.
References
1. Gillberg C. The essence in child psychiatry: early symptomatic syndromes eliciting neurodevelopmental clinical examinations. Res Dev Disabil. 2010;31(6):1543–1551. doi:10.1016/j.ridd.2010.06.002
2. Gillberg C. The Essence of Autism and Other Neurodevelopmental Conditions. Rethinking Co-Morbidities, Assessment, and Intervention. Jessica Kingsley Publishers; 2021.
3. Morris-Rosendahl DJ, Crocq MA. Neurodevelopmental disorders-The history and future of a diagnostic concept. Dialogues Clin Neurosci. 2020;22:65–72.
4. Ibrahim K, Sukhodolsky D. Rdoc and autism. In: Encyclopedia of Autism Spectrum Disorders. New York: Springer; 2018.
5. Insel T, Cuthbert B, Garvey M, et al. Research domain criteria (rdoc): toward a new classification framework for research on mental disorders. Am J Psychiatry. 2010;167:748–751.
6. Brandeis D, Loo S, McLoughlin G, Heinrich H, Banaschewski T. Neurophysiology. In: Banaschewski T, Coghill D, Zuddas A, editors. Oxford Textbook of Attention Deficit Hyperactivity Disorder. Oxford University Press; 2018.
7. Rohde L, Kieling C, Salum G. Current diagnostic criteria dsm, icd, and future perspectives. In: Banaschewski T, Coghill D, Zuddas A, editors. Oxford Textbook of Attention Deficit Hyperactivity Disorder. Oxford University Press; 2018.
8. National Institute of Mental Health. Definitions of the RDoC Domains and Constructs. Available from: https://www.nimh.nih.gov/research/research-funded-by-nimh/rdoc. Accessed September 17, 2022.
9. Stahl SM. Deconstructing psychiatric disorders, part 2: an emerging, neurobiologically based therapeutic strategy for the modern psychopharmacologist. J Clin Psychiatry. 2003;64:1145–1146.
10. Stahl S, Mignon L. Stahl’s Illustrated Attention Deficit Hyperactivity Disorder. Cambridge University Press; 2009.
11. Gillberg C. ESSENCE samlar diagnoserna till en helhet [ESSENCE gathers the diagnoses into a whole]. Lakartidningen. 2014;111(39):1643–1646. Swedish.
12. Al-Kuraishy HM, Abdulhadi MH, Hussien NR, Al-Niemi MS, Rasheed HA, Al-Gareeb AI. Involvement of orexinergic system in psychiatric and neurodegenerative disorders: a scoping review. Brain Circ. 2020;6(2):70–80. doi:10.4103/bc.bc_42_19
13. Baykal S, Albayrak Y, Durankus F, et al. Decreased serum orexin a levels in drug-naive children with attention deficit and hyperactivity disorder. Neurol Sci. 2019;40(3):593–602. doi:10.1007/s10072-018-3692-8
14. Cortese S, Konofal E, Lecendreux M. Alertness and feeding behaviors in ADHD: does the hypocretin/orexin system play a role? Med Hypotheses. 2008;71(5):770–775. doi:10.1016/j.mehy.2008.06.017
15. Marotta R, Risoleo MC, Messina G, et al. The neurochemistry of autism. Brain Sci. 2020;10(3):163. doi:10.3390/brainsci10030163
16. Kobylinska L, Panaitescu AM, Gabreanu G, et al. Plasmatic levels of neuropeptides, including oxytocin, in children with autism spectrum disorder, correlate with the disorder severity. Acta Endocrinol. 2019;5:16–24.
17. Kohyama J. Possible neuronal mechanisms of sleep disturbances in patients with autism spectrum disorders and attention-deficit/hyperactivity disorder. Med Hypotheses. 2016;97:131–133.
18. Messina A, Monda V, Sessa F, et al. Sympathetic, metabolic adaptations, and oxidative stress in autism spectrum disorders: how far from physiology? Front Physiol. 2018;9:261.
19. Thapar A, Cooper M. Attention deficit hyperactivity disorder. Lancet. 2016;387:1240–1250.
20. Heidbreder R. ADHD symptomatology is best conceptualized as a spectrum: a dimensional versus unitary approach to diagnosis. Atten Defic Hyperact Disord. 2015;7:249–269.
21. American Psychiatric A. Diagnostic and Statistical Manual of Mental Disorders (Dsm-5®). Washington, United States: American Psychiatric Publishing; 2013.
22. World Health Organization. International Statistical Classification of Diseases and Related Health Problems.
23. Musser ED, Raiker JS. Attention-deficit/hyperactivity disorder: an integrated developmental psychopathology and research domain criteria (rdoc) approach. Compr Psychiatry. 2019;90:65–72.
24. Chieffi S, Carotenuto M, Monda V, et al. Orexin system: the key for a healthy life. Front Physiol. 2017;8:357.
25. Seigneur E, de Lecea L. Hypocretin (orexin) replacement therapies. Med Drug Discov. 2020;8:100070.
26. Stahl S. Stahl’s Essential Psychopharmacology: Neuroscientific Basis and Practical Applications.
27. Jacobson LH, Hoyer D, de Lecea L. Hypocretins (orexins): the ultimate translational neuropeptides. J Intern Med. 2022;291:533–556.
28. Li SB, de Lecea L. The hypocretin (orexin) system: from a neural circuitry perspective. Neuropharmacology. 2020;167:107993.
29. Jöhren O, Neidert SJ, Kummer M, Dendorfer A, Dominiak P. Prepro-orexin and orexin receptor mRNAs are differentially expressed in peripheral tissues of male and female rats. Endocrinology. 2001;142(8):3324–3431.
30. Kukkonen JP, Leonard CS. Orexin/hypocretin receptor signalling cascades. Br J Pharmacol. 2014;171:314–331.
31. Leonard CS, Kukkonen JP. Orexin/hypocretin receptor signalling: a functional perspective. Br J Pharmacol. 2014;171:294–313.
32. Alain C, Pascal N, Valérie G, Thierry V. Orexins/hypocretins and cancer: a neuropeptide as emerging target. Molecules. 2021;26:4849.
33. Zhang XY, Yu L, Zhuang QX, Zhu JN, Wang JJ. Central functions of the orexinergic system. Neurosci Bull. 2013;29:355–365.
34. Summers CH, Yaeger JDW, Staton CD, Arendt DH, Summers TR. Orexin/hypocretin receptor modulation of anxiolytic and antidepressive responses during social stress and decision-making: potential for therapy. Brain Res. 2020;1731:146085.
35. Higgins E, George M. The Neuroscience of Clinical Psychiatry: The Pathophysiology of Behavior and Mental Illness.
36. Stahl S, Morrissette D. Stahl’s Illustrated Sleep and Wake Disorders. Cambridge University Press; 2016.
37. Scammell TE, Winrow CJ. Orexin receptors: pharmacology and therapeutic opportunities. Annu Rev Pharmacol Toxicol. 2011;51:243–266.
38. Toor B, Ray LB, Pozzobon A, Fogel SM. Sleep, orexin and cognition. Front Neurol Neurosci. 2021;45:38–51.
39. Iranzo A. Sleep and neurological autoimmune diseases. Neuropsychopharmacology. 2020;45:129–140.
40. Peerboom C, Wierenga CJ. The postnatal gaba shift: a developmental perspective. Neurosci Biobehav Rev. 2021;124:179–192.
41. Herlenius E, Lagercrantz H. Neurotransmitters and neuromodulators during early human development. Early Hum Dev. 2001;65:21–37.
42. Purkayastha P, Malapati A, Yogeeswari P, Sriram D. A review on gaba/glutamate pathway for therapeutic intervention of asd and ADHD. Curr Med Chem. 2015;22(15):1850–1859.
43. Bonnavion P, Mickelsen LE, Fujita A, de Lecea L, Jackson AC. Hubs and spokes of the lateral hypothalamus: cell types, circuits and behaviour. J Physiol. 2016;594:6443–6462.
44. Mahler SV, Moorman DE, Smith RJ, James MH, Aston-Jones G. Motivational activation: a unifying hypothesis of orexin/hypocretin function. Nat Neurosci. 2014;17:1298–1303.
45. Faesel N, Kolodziejczyk MH, Koch M, Fendt M. Orexin deficiency affects sociability and the acquisition, expression, and extinction of conditioned social fear. Brain Res. 2021;1751:147199.
46. Chen Q, de Lecea L, Hu Z, Gao D. The hypocretin/orexin system: an increasingly important role in neuropsychiatry. Med Res Rev. 2015;35:152–197.
47. Mullington JM, Cunningham TJ, Haack M, Yang H. Causes and consequences of chronic sleep deficiency and the role of orexin. Front Neurol Neurosci. 2021;45:128–138.
48. BaHammam AS, Alnakshabandi K, Pandi-Perumal SR. Neuropsychiatric correlates of narcolepsy. Curr Psychiatry Rep. 2020;22:36.
49. Abounoori M, Maddah MM, Akbari E, Houshmand G, Ardeshiri MR. The effect of orexin receptor antagonism on quinpirole-induced compulsive-like checking behavior in rats. Neurotox Res. 2020;38:18–26.
50. Plazzi G, Pizza F, Palaia V, et al. Complex movement disorders at disease onset in childhood narcolepsy with cataplexy. Brain. 2011;134:3477–3489.
51. Hu B, Yang N, Qiao QC, Hu ZA, Zhang J. Roles of the orexin system in central motor control. Neurosci Biobehav Rev. 2015;49:43–54.
52. Kobayashi M, Nomura M, Fujihara H, et al. Involvement of orexin-A on micturition reflex in normal and cyclophosphamide-induced cystitis bladder in rat. Peptides. 2009;30:2348–2356.
53. Clark IA, Vissel B. Inflammation-sleep interface in brain disease: tnf, insulin, orexin. J Neuroinflammation. 2014;11:51.
54. Tsuchimine S, Hattori K, Ota M, et al. Reduced plasma orexin-A levels in patients with bipolar disorder. Neuropsychiatr Dis Treat. 2019;15:2221–2230.
55. Li H, Lu J, Li S, et al. Increased hypocretin (orexin) plasma level in depression, bipolar disorder patients. Front Psychiatry. 2021;12:676336.
56. Mogavero MP, Silvani A, DelRosso LM, Salemi M, Ferri R. Focus on the complex interconnection between cancer, narcolepsy and other neurodegenerative diseases: a possible case of orexin-dependent inverse comorbidity. In: Cancers; 2021:13.
57. Kawabe K, Horiuchi F, Ochi M, Nishimoto K, Ueno SI, Oka Y. Suvorexant for the treatment of insomnia in adolescents. J Child Adolesc Psychopharmacol. 2017;27:792–795.
58. Bruni O, Angriman M, Melegari MG, Ferri R. Pharmacotherapeutic management of sleep disorders in children with neurodevelopmental disorders. Expert Opin Pharmacother. 2019;20:2257–2271.
59. Shariq AS, Rosenblat JD, Alageel A, et al. Evaluating the role of orexins in the pathophysiology and treatment of depression: a comprehensive review. Prog Neuropsychopharmacol Biol Psychiatry. 2019;92:1–7.
60. Soya S, Takahashi TM, McHugh TJ, et al. Orexin modulates behavioral fear expression through the locus coeruleus. Nat Commun. 2017;8:1606.
61. Khairuddin S, Aquili L, Heng BC, Hoo TLC, Wong KH, Lim LW. Dysregulation of the orexinergic system: a potential neuropeptide target in depression. Neurosci Biobehav Rev. 2020;118:384–396.
62. Gentile TA, Simmons SJ, Watson MN, et al. Effects of suvorexant, a dual orexin/hypocretin receptor antagonist, on impulsive behavior associated with cocaine. Neuropsychopharmacology. 2018;43:1001–1009.
63. Taylor JL, Corbett BA. A review of rhythm and responsiveness of cortisol in individuals with autism spectrum disorders. Psychoneuroendocrinology. 2014;49:207–228.
64. Buske-Kirschbaum A, Trikojat K, Tesch F, et al. Altered hypothalamus-pituitary-adrenal axis function: a relevant factor in the comorbidity of atopic eczema and attention deficit/hyperactivity disorder? Psychoneuroendocrinology. 2019;105:178–186.
65. Theoharides TC, Stewart JM, Hatziagelaki E, Kolaitis G. Brain “fog”, inflammation and obesity: key aspects of neuropsychiatric disorders improved by luteolin. Front Neurosci. 2015;9:225.
66. Kleberg JL, Frick MA, Brocki KC. Increased pupil dilation to happy faces in children with hyperactive/impulsive symptoms of ADHD. Dev Psychopathol. 2021;33:767–777.
67. Anderson CJ, Colombo J. Larger tonic pupil size in young children with autism spectrum disorder. Dev Psychobiol. 2009;51:207–211.
68. de Vries L, Fouquaet I, Boets B, Naulaers G, Steyaert J. Autism spectrum disorder and pupillometry: a systematic review and meta-analysis. Neurosci Biobehav Rev. 2021;120:479–508.
69. Sakurai T, Saito YC, Yanagisawa M. Interaction between orexin neurons and monoaminergic systems. Front Neurol Neurosci. 2021;45:11–21.
70. Knez R. Commentary on the paper by Gagliano et al: artificial neural networks analysis of polysomnographic and clinical features in pediatric acute-onset neuropsychiatric syndrome (pans): from sleep alteration to “brain fog” [letter]. Nat Sci Sleep. 2021;13:1687–1688.
71. Schreyer S, Buttner-Ennever JA, Tang X, Mustari MJ, Horn AK. Orexin-A inputs onto visuomotor cell groups in the monkey brainstem. Neuroscience. 2009;164:629–640.
72. Albajara Saenz A, Villemonteix T, Van Schuerbeek P, et al. Motor abnormalities in attention-deficit/hyperactivity disorder and autism spectrum disorder are associated with regional grey matter volumes. Front Neurol. 2021;12:666980.
73. Mokobane M, Pillay BJ, Meyer A. Fine motor deficits and attention deficit hyperactivity disorder in primary school children. S Afr J Psychiatr. 2019;25:1232.
74. Jasmin E, Couture M, McKinley P, Reid G, Fombonne E, Gisel E. Sensori-motor and daily living skills of preschool children with autism spectrum disorders. J Autism Dev Disord. 2009;39:231–241.
75. Cardon GJ. Neural correlates of sensory abnormalities across developmental disabilities. Int Rev Res Dev Disabil. 2018;55:83–143.
76. Fuermaier ABM, Hupen P, De Vries SM, et al. Perception in attention deficit hyperactivity disorder. Atten Defic Hyperact Disord. 2018;10:21–47.
77. Sweigert JR, St John T, Begay KK, et al. Characterizing olfactory function in children with autism spectrum disorder and children with sensory processing dysfunction. Brain Sci. 2020;10:362.
78. Siqueiros Sanchez M, Falck-Ytter T, Kennedy DP, et al. Volitional eye movement control and ADHD traits: a twin study. J Child Psychol Psychiatry. 2020;61:1309–1316.
79. Howard PL, Zhang L, Benson V. What can eye movements tell us about subtle cognitive processing differences in autism? Vision. 2019;3:22.
80. Burdakov D. Subsecond ensemble dynamics of orexin neurons link sensation and action. Front Neurol Neurosci. 2021;45:52–60.
81. Muschamp JW, Dominguez JM, Sato SM, Shen RY, Hull EM. A role for hypocretin (orexin) in male sexual behavior. J Neurosci. 2007;27:2837–2845.
82. Caillol M, Aioun J, Baly C, Persuy MA, Salesse R. Localization of orexins and their receptors in the rat olfactory system: possible modulation of olfactory perception by a neuropeptide synthetized centrally or locally. Brain Res. 2003;960:48–61.
83. Filardi M, Pizza F, Tonetti L, Antelmi E, Natale V, Plazzi G. Attention impairments and ADHD symptoms in adult narcoleptic patients with and without hypocretin deficiency. PLoS One. 2017;12:e0182085.
84. Merlo Pich E, Melotto S. Orexin 1 receptor antagonists in compulsive behavior and anxiety: possible therapeutic use. Front Neurosci. 2014;8:26.
85. Volkow ND, Wang GJ, Newcorn JH, et al. Motivation deficit in ADHD is associated with dysfunction of the dopamine reward pathway. Mol Psychiatry. 2011;16:1147–1154.
86. Supekar K, Kochalka J, Schaer M, et al. Deficits in mesolimbic reward pathway underlie social interaction impairments in children with autism. Brain. 2018;141:2795–2805.
87. Clements CC, Zoltowski AR, Yankowitz LD, Yerys BE, Schultz RT, Herrington JD. Evaluation of the social motivation hypothesis of autism: a systematic review and meta-analysis. JAMA Psychiatry. 2018;75:797–808.
88. Kohls G, Antezana L, Mosner MG, Schultz RT, Yerys BE. Altered reward system reactivity for personalized circumscribed interests in autism. Mol Autism. 2018;9:9.
89. Mayes SD, Zickgraf H. Atypical eating behaviors in children and adolescents with autism, ADHD, other disorders, and typical development. Res Autism Spect Dis. 2019;64:76–83.
90. Curtin C, Pagoto S. The association between ADHD and eating disorders/pathology in adolescents: a systematic review. Open J Epidemiol. 2013;2013. doi:10.4236/ojepi.2013.34028
91. Martins-Silva T, Vaz JDS, Genro JP, et al. Obesity and ADHD: exploring the role of body composition, BMI polygenic risk score, and reward system genes. J Psychiatr Res. 2021;136:529–536.
92. Tyree SM, Borniger JC, de Lecea L. Hypocretin as a hub for arousal and motivation. Front Neurol. 2018;9:413.
93. Choi DL, Davis JF, Fitzgerald ME, Benoit SC. The role of orexin-A in food motivation, reward-based feeding behavior and food-induced neuronal activation in rats. Neuroscience. 2010;167:11–20.
94. Mavanji V, Butterick TA, Duffy CM, Nixon JP, Billington CJ, Kotz CM. Orexin/hypocretin treatment restores hippocampal-dependent memory in orexin-deficient mice. Neurobiol Learn Mem. 2017;146:21–30.
95. Barson JR. Orexin/hypocretin and dysregulated eating: promotion of foraging behavior. Brain Res. 2020;1731:145915.
96. Wolfe FH, Auzias G, Deruelle C, Chaminade T. Focal atrophy of the hypothalamus associated with third ventricle enlargement in autism spectrum disorder. Neuroreport. 2015;26:1017–1022.
97. Caria A, Ciringione L, Falco S. Morphofunctional alterations of the hypothalamus and social behavior in autism spectrum disorders. Brain Sci. 2020;10:435.
98. Mayes SD, Calhoun SL, Aggarwal R, et al. Unusual fears in children with autism. Res Autism Spect Dis. 2013;7:151–158.
99. Spencer AE, Marin MF, Milad MR, et al. Abnormal fear circuitry in attention deficit hyperactivity disorder: a controlled magnetic resonance imaging study. Psychiatry Res Neuroimaging. 2017;262:55–62.
100. Reppucci CJ, Gergely CK, Bredewold R, Veenema AH. Involvement of orexin/hypocretin in the expression of social play behaviour in juvenile rats. Int JPlay. 2020;9(1):108–127. doi:10.1080/21594937.2020.1720132
101. Sears RM, Fink AE, Wigestrand MB, Farb CR, de Lecea L, Ledoux JE. Orexin/hypocretin system modulates amygdala-dependent threat learning through the locus coeruleus. Proc Natl Acad Sci U S A. 2013;110(50):20260–20265. doi:10.1073/pnas.1320325110
102. Dustrude ET, Caliman IF, Bernabe CS, et al. Orexin depolarizes central amygdala neurons via orexin receptor 1, phospholipase c and sodium-calcium exchanger and modulates conditioned fear. Front Neurosci. 2018;12:934. doi:10.3389/fnins.2018.00934
103. Quaedackers L, van Gilst MM, van Mierlo P, et al. Impaired social functioning in children with narcolepsy. Sleep. 2019;42(2):zsy228. doi:10.1093/sleep/zsy228
104. Wang Y, Zhang Y-B, Liu -L-L, et al. A meta-analysis of working memory impairments in autism spectrum disorders. Neuropsychol Rev. 2017;27(1):46–61. doi:10.1007/s11065-016-9336-y
105. Durairaja A, Fendt M. Orexin deficiency modulates cognitive flexibility in a sex-dependent manner. Genes Brain Behav. 2021;20(3):e12707. doi:10.1111/gbb.12707
106. Wainstein G, Rojas-Libano D, Crossley NA, Carrasco X, Aboitiz F, Ossandon T. Pupil size tracks attentional performance in attention-deficit/hyperactivity disorder. Sci Rep. 2017;7(1):8228. doi:10.1038/s41598-017-08246-w
107. Deumer U-S, Varesi A, Floris V, et al. Myalgic encephalomyelitis/chronic fatigue syndrome (me/cfs): an overview. J Clin Med. 2021;10(20):4786. doi:10.3390/jcm10204786
108. Matsuki T, Nomiyama M, Takahira H, et al. Selective loss of GABA B receptors in orexin-producing neurons results in disrupted sleep/wakefulness architecture. Proc Natl Acad Sci U S A. 2009;106(11):4459–4464. doi:10.1073/pnas.0811126106
109. Mancuso CE, Tanzi MG, Gabay M. Paradoxical reactions to benzodiazepines: literature review and treatment options. Pharmacotherapy. 2004;24(9):1177–1185. doi:10.1592/phco.24.13.1177.38089
110. Bassetti CLA, Adamantidis A, Burdakov D, et al. Narcolepsy - clinical spectrum, aetiopathophysiology, diagnosis and treatment. Nat Rev Neurol. 2019;15(9):519–539. doi:10.1038/s41582-019-0226-9
111. Kim J, Lee G-H, Sung SM, Jung DS, Pak K. Prevalence of attention deficit hyperactivity disorder symptoms in narcolepsy: a systematic review. Sleep Med. 2020;65:84–88. doi:10.1016/j.sleep.2019.07.022
112. Lecendreux M, Lavault S, Lopez R, et al. Attention-deficit/hyperactivity disorder (ADHD) symptoms in pediatric narcolepsy: a cross-sectional study. Sleep. 2015;38(8):1285–1295. doi:10.5665/sleep.4910
113. Wilenius L, Partinen M. Attention-deficit/hyperactivity disorder patients may have undiagnosed narcolepsy. Cureus. 2020;12(6):e8436. doi:10.7759/cureus.8436
114. Takahashi N, Nishimura T, Harada T, et al. Polygenic risk score analysis revealed shared genetic background in attention deficit hyperactivity disorder and narcolepsy. Transl Psychiatry. 2020;10(1):284. doi:10.1038/s41398-020-00971-7
115. Posar A, Visconti P, Blunda V, Pizza F, Plazzi G. Autism spectrum disorder and narcolepsy: a possible connection that deserves to be investigated. Front Psychiatry. 2020;11:265. doi:10.3389/fpsyt.2020.00265
116. Vaughan S, Failla MD, Poole HM, et al. Pain processing in psychiatric conditions: a systematic review. Rev General Psychol. 2019;23(3):336–358. doi:10.1177/1089268019842771
117. Kang X, Tang H, Liu Y, Yuan Y, Wang M. Research progress on the mechanism of orexin in pain regulation in different brain regions. Open Life Sci. 2021;16(1):46–52. doi:10.1515/biol-2021-0001
118. Razavi BM, Hosseinzadeh H. A review of the role of orexin system in pain modulation. Biomed Pharmacother. 2017;90:187–193. doi:10.1016/j.biopha.2017.03.053
119. Roehrs T, Withrow D, Koshorek G, Verkler J, Bazan L, Roth T. Sleep and pain in humans with fibromyalgia and comorbid insomnia: double-blind, crossover study of suvorexant 20 mg versus placebo. J Clin Sleep Med. 2020;16(3):415–421. doi:10.5664/jcsm.8220
120. Montefusco-Siegmund R, Schwalm M, Rosales Jubal E, Devia C, Egana JI, Maldonado PE. Alpha EEG activity and pupil diameter coupling during inactive wakefulness in humans. eNeuro. 2022;9(2). doi:10.1523/ENEURO.0060-21.2022
121. Dichter GS, Rodriguez-Romaguera J. Anhedonia and Hyperhedonia in Autism and Related Neurodevelopmental Disorders. Berlin, Heidelberg: Springer Berlin Heidelberg; 2022:1–18.
122. Nazar BP, Bernardes C, Peachey G, Sergeant J, Mattos P, Treasure J. The risk of eating disorders comorbid with attention-deficit/hyperactivity disorder: a systematic review and meta-analysis. Int J Eat Disord. 2016;49(12):1045–1057. doi:10.1002/eat.22643
123. Svedlund NE, Norring C, Ginsberg Y, von Hausswolff-Juhlin Y. Behandling av ätstörningar vid samtidiga ADHD-symptom: Observerat samband kan ge nya möjligheter till medicinering, främst av Bulimia nervosa-men ännu finns kunskapsluckor [Treatment of eating disorders with concurrent ADHD symptoms: knowledge, knowledge gaps and clinical implications]. Lakartidningen. 2019;17:116. Swedish.
124. Koomar T, Thomas TR, Pottschmidt NR, Lutter M, Michaelson JJ. Estimating the prevalence and genetic risk mechanisms of arfid in a large autism cohort. Front Psychiatry. 2021;12:668297. doi:10.3389/fpsyt.2021.668297
125. Råstam M, Täljemark J, Tajnia A, et al. Eating problems and overlap with ADHD and autism spectrum disorders in a nationwide twin study of 9- and 12-year-old children. ScientificWorldJournal. 2013;2013:1–7. doi:10.1155/2013/315429
126. Hölcke M, Marcus C, Gillberg C, Fernell E. Paediatric obesity: a neurodevelopmental perspective. Acta Paediatrica. 2008;97(6):819–821. doi:10.1111/j.1651-2227.2008.00816.x
127. Sammels O, Karjalainen L, Dahlgren J, Wentz E. Autism spectrum disorder and obesity in children: a systematic review and meta-analysis. Obes Facts. 2022;15(3):305–320. doi:10.1159/000523943
128. Fast K, Bjork A, Strandberg M, Johannesson E, Wentz E, Dahlgren J. Half of the children with overweight or obesity and attention-deficit/hyperactivity disorder reach normal weight with stimulants. Acta Paediatr. 2021;110(10):2825–2832. doi:10.1111/apa.15881
129. Adeghate E, Lotfy M, D’Souza C, Alseiari S, Alsaadi A, Qahtan S. Hypocretin/orexin modulates body weight and the metabolism of glucose and insulin. Diabetes Metabol Res Rev. 2020;36(3):e3229. doi:10.1002/dmrr.3229
130. Sternat T, Fotinos K, Fine A, Epstein I, Katzman MA. Low hedonic tone and attention-deficit hyperactivity disorder: risk factors for treatment resistance in depressed adults. Neuropsychiatr Dis Treat. 2018;14:2379–2387. doi:10.2147/NDT.S170645
131. Sternat T, KAtzman M. Neurobiology of hedonic tone: the relationship between treatment-resistant depression, attention-deficit hyperactivity disorder, and substance abuse. Neuropsych Dis Treat. 2016;12:2149–2164. doi:10.2147/NDT.S111818
132. Pizzagalli DA, Jahn AL, O’Shea JP. Toward an objective characterization of an anhedonic phenotype: a signal-detection approach. Biol Psychiatry. 2005;57(4):319–327. doi:10.1016/j.biopsych.2004.11.026
133. Meinzer MC, Pettit J, Leventhal AM, Hill RM. Explaining the covariance between attention-deficit hyperactivity disorder symptoms and depressive symptoms: the role of hedonic responsivity. J Clin Psychol. 2012;68(10):1111–1121. doi:10.1002/jclp.21884
134. Novacek DM, Gooding DC, Pflum MJ. Hedonic capacity in the broader autism phenotype: should social anhedonia be considered a characteristic feature? Front Psychol. 2016;7:666. doi:10.3389/fpsyg.2016.00666
135. Billones RR, Kumar S, Saligan LN. Disentangling fatigue from anhedonia: a scoping review. Transl Psychiatry. 2020;10(1):273. doi:10.1038/s41398-020-00960-w
136. Mileykovskiy BY, Kiyashchenko LI, Siegel JM. Behavioral correlates of activity in identified hypocretin/orexin neurons. Neuron. 2005;46(5):787–798. doi:10.1016/j.neuron.2005.04.035
137. Fronczek R, Overeem S, Reijntjes R, Lammers GJ, van Dijk JG, Pijl H. Increased heart rate variability but normal resting metabolic rate in hypocretin/orexin-deficient human narcolepsy. J Clin Sleep Med. 2008;4(03):248–254. doi:10.5664/jcsm.27188
138. Schuchardt K, Gebhardt M, Mäehler C. Working memory functions in children with different degrees of intellectual disability. J Intellect Disabil Res. 2010;54(4):346–353. doi: 10.1111/j.1365-2788.2010.01265.x.
139. Alloway TP. Working memory and executive function profiles of individuals with borderline intellectual functioning. J Intellect Disabil Res. 2010;54(5):448–456. doi:10.1111/j.1365-2788.2010.01281.x
140. Becker S, Sharma MJ, Callahan BL. ADHD and neurodegenerative disease risk: a critical examination of the evidence. Front Aging Neurosci. 2022;13:826213. doi:10.3389/fnagi.2021.826213
141. Zhang L, Du Rietz E, Kuja‐Halkola R, et al. Attention-deficit/hyperactivity disorder and Alzheimer’s disease and any dementia: a multi-generation cohort study in Sweden. Alzheimers Dement. 2022;18(6):1155–1163. doi:10.1002/alz.12462
142. Gillberg C. Deficits in attention, motor control, and perception: a brief review. Arch Dis Child. 2003;88(10):904–910. doi:10.1136/adc.88.10.904
143. Deng S, Li W-G, Ding J, et al. Understanding the mechanisms of cognitive impairments in developmental coordination disorder. Pediatr Res. 2014;75(1–2):210–216. doi:10.1038/pr.2013.192
144. Cikriklar HI, Kotan D, Yucel M, et al. The role of orexin-A levels in epileptic seizure. Neurosci Lett. 2020;734:135097. doi:10.1016/j.neulet.2020.135097
145. Ozsoy S, Erdem DS, Abdulrezzak U, Basturk M. Serum orexin a and oxytocin levels and their relationships with attachment and childhood trauma in patients with depression. Eur Neuropsychopharm. 2019;29:S374–S375. doi:10.1016/j.euroneuro.2018.11.573
146. Portelius E, Soininen H, Andreasson U, et al. Exploring Alzheimer molecular pathology in down’s syndrome cerebrospinal fluid. Neurodegener Dis. 2014;14(2):98–106. doi:10.1159/000358800
147. Zhang B, Guo D, Han L, Rensing N, Satoh A, Wong M. Hypothalamic orexin and mechanistic target of rapamycin activation mediate sleep dysfunction in a mouse model of tuberous sclerosis complex. Neurobiol Dis. 2020;134:104615. doi:10.1016/j.nbd.2019.104615
148. Cataldi M, Arnaldi D, Tucci V, et al. Sleep disorders in Prader-Willi syndrome, evidence from animal models and humans. Sleep Med Rev. 2021;57:101432. doi:10.1016/j.smrv.2021.101432
149. Olateju OI, Bhagwandin A, Ihunwo AO, Manger PR. Changes in the cholinergic, catecholaminergic, orexinergic and serotonergic structures forming part of the sleep systems of adult mice exposed to intrauterine alcohol. Front Neuroanat. 2017;11:110. doi:10.3389/fnana.2017.00110
150. Collier AD, Halkina V, Min SS, et al. Embryonic ethanol exposure affects the early development, migration, and location of hypocretin/orexin neurons in zebrafish. Alcohol Clin Exp Res. 2019;43(8):1702–1713. doi:10.1111/acer.14126
151. Stettner GM, Kubin L, Volgin DV. Antagonism of orexin 1 receptors eliminates motor hyperactivity and improves homing response acquisition in juvenile rats exposed to alcohol during early postnatal period. Behav Brain Res. 2011;221(1):324–328. doi:10.1016/j.bbr.2011.03.028
152. Gagliano A, Puligheddu M, Ronzano N, et al. Artificial neural networks analysis of polysomnographic and clinical features in pediatric acute-onset neuropsychiatric syndrome (pans): from sleep alteration to “brain fog”. Nat Sci Sleep. 2021;13:1209–1224. doi:10.2147/NSS.S300818
153. Wang JY, Han F, Dong SX, et al. Cerebrospinal fluid orexin a levels and autonomic function in Kleine-Levin syndrome. Sleep. 2016;39(4):855–860. doi:10.5665/sleep.5642
154. Moskowitz AK. ”Scared Stiff”: catatonia as an evolutionary-based fear response. Psychol Rev. 2004;111(4):984–1002. doi:10.1037/0033-295X.111.4.984
155. Soya S, Shoji H, Hasegawa E, et al. Orexin receptor-1 in the locus coeruleus plays an important role in cue-dependent fear memory consolidation. J Neurosci. 2013;33(36):14549–14557. doi:10.1523/JNEUROSCI.1130-13.2013
156. Hansen N, Bartels C, Teegen B, Wiltfang J, Malchow B. Catatonic schizophrenia associated with cerebrospinal gad65 autoantibodies: case report and literature review. Front Immunol. 2022;13:829058. doi:10.3389/fimmu.2022.829058
157. McKeon A, Robinson MT, McEvoy KM, et al. Stiff-man syndrome and variants: clinical course, treatments, and outcomes. Arch Neurol. 2012;69(2):230–238. doi:10.1001/archneurol.2011.991
158. Kosse C, Schone C, Bracey E, Burdakov D. Orexin-driven gad65 network of the lateral hypothalamus sets physical activity in mice. Proc Natl Acad Sci U S A. 2017;114(17):4525–4530. doi:10.1073/pnas.1619700114
159. Komaroff AL, Lipkin WI. Insights from myalgic encephalomyelitis/chronic fatigue syndrome may help unravel the pathogenesis of postacute covid-19 syndrome. Trends Mol Med. 2021;27(9):895–906. doi:10.1016/j.molmed.2021.06.002
160. Wang EY, Mao T, Klein J, et al. Diverse functional autoantibodies in patients with covid-19. Nature. 2021;595:283–288.
161. Bileviciute-Ljungar IA-O, Maroti D, Bejerot S. Patients with chronic fatigue syndrome do not score higher on the autism-spectrum quotient than healthy controls: comparison with autism spectrum disorder. Scand J Psychol. 2018;59(4):428–432.
162. Miyamae T, Chiba Y, Kato I, Tani Y, Yamanaka H. Neurodevelopmental disorders associated with juvenile fibromyalgia. Pediatr Int. 2018;60:1034–1035.
163. Okumura T, Nozu T, Ishioh M, Igarashi S, Kumei S, Ohhira M. Brain orexin improves intestinal barrier function via the vagal cholinergic pathway. Neurosci Lett. 2020;714:134592.
164. Mediavilla C. Bidirectional gut-brain communication: a role for orexin-A. Neurochem Int. 2020;141:104882.
165. Yilmaz E, Tamam L. Attention-deficit hyperactivity disorder and impulsivity in female patients with fibromyalgia. Neuropsychiatr Dis Treat. 2018;14:1883–1889.
166. Baeza-Velasco C, Grahame R, Bravo JF. A connective tissue disorder may underlie essence problems in childhood. Res Dev Disabil. 2017;60:232–242.
167. Yew KS, Kamps-Schmitt KA, Borge R. Hypermobile Ehlers-Danlos syndrome and hypermobility spectrum disorders. Am Fam Physician. 2021;103:481–492.
168. Kindgren E, Quinones Perez A, Knez R. Prevalence of ADHD and autism spectrum disorder in children with hypermobility spectrum disorders or hypermobile Ehlers-Danlos syndrome: a retrospective study. Neuropsychiatr Dis Treat. 2021;17:379–388.
169. Sagi D, de Lecea L, Appelbaum L. Heterogeneity of hypocretin/orexin neurons. Front Neurol Neurosci. 2021;45:61–74.
170. Kukkonen JP. Orexin/hypocretin signaling. Curr Top Behav Neurosci. 2017;33:17–50.
171. Dunn GA, Nigg JT, Sullivan EL. Neuroinflammation as a risk factor for attention deficit hyperactivity disorder. Pharmacol Biochem Behav. 2019;182:22–34.
172. Keil A, Daniels JL, Forssen U, et al. Parental autoimmune diseases associated with autism spectrum disorders in offspring. Epidemiology. 2010;21:805–808.
173. Nielsen TC, Nassar N, Shand AW, et al. Association of maternal autoimmune disease with attention-deficit/hyperactivity disorder in children. JAMA Pediatr. 2021;175:e205487.
174. McCaffrey TA, St Laurent G, Shtokalo D, et al. Biomarker discovery in attention deficit hyperactivity disorder: RNA sequencing of whole blood in discordant twin and case-controlled cohorts. BMC Med Genomics. 2020;13:160.
175. Nudel R, Benros ME, Krebs MD, et al. Immunity and mental illness: findings from a Danish population-based immunogenetic study of seven psychiatric and neurodevelopmental disorders. Eur J Hum Genet. 2019;27:1445–1455.
176. Tamouza R, Fernell E, Eriksson MA, et al. Hla polymorphism in regressive and non-regressive autism: a preliminary study. Autism Res. 2020;13:182–186.
177. Rout UK, Mungan NK, Dhossche DM. Presence of gad65 autoantibodies in the serum of children with autism or ADHD. Eur Child Adolesc Psychiatry. 2012;21:141–147.
178. Bellavance MA, Rivest S. The hpa - immune axis and the immunomodulatory actions of glucocorticoids in the brain. Front Immunol. 2014;5:136.
179. Sehgal A, Mignot E. Genetics of sleep and sleep disorders. Cell. 2011;146:194–207.
180. Ollila HM, Ravel JM, Han F, et al. Hla-dpb1 and hla class i confer risk of and protection from narcolepsy. Am J Hum Genet. 2015;96:136–146.
181. Checa-Ros A, Jerez-Calero A, Molina-Carballo A, Campoy C, Munoz-Hoyos A. Current evidence on the role of the gut microbiome in ADHD pathophysiology and therapeutic implications. Nutrients. 2021;13:249.
182. Xu M, Xu X, Li J, Li F. Association between gut microbiota and autism spectrum disorder: a systematic review and meta-analysis. Front Psychiatry. 2019;10:473.
183. Breugelmans T, De Winter B, Smet A. The microbiota-gut-brain axis in gastrointestinal inflammation and neurological comorbidities. Microb Health Dis. 2019;1:e201.
184. Gillberg C, Fernell E, Kocovska E, et al. The role of cholesterol metabolism and various steroid abnormalities in autism spectrum disorders: a hypothesis paper. Autism Res. 2017;10:1022–1044.
185. Gao Q, Kou T, Zhuang B, Ren Y, Dong X, Wang Q. The association between vitamin d deficiency and sleep disorders: a systematic review and meta-analysis. Nutrients. 2018;10:1395.
186. Zheng D, Zhao Y, Shen Y, Chang X, Ju S, Guo L. Orexin a-mediated stimulation of 3beta-hsd expression and testosterone production through MAPK signaling pathways in primary rat leydig cells. J Endocrinol Invest. 2014;37:285–292.
187. Rukmani MR, Seshadri SP, Thennarasu K, Raju TR, Sathyaprabha TN. Heart rate variability in children with attention-deficit/hyperactivity disorder: a pilot study. Ann Neurosci. 2016;23:81–88.
188. Griffiths KR, Quintana DS, Hermens DF, et al. Sustained attention and heart rate variability in children and adolescents with ADHD. Biol Psychol. 2017;124:11–20.
189. Sarovic D. A unifying theory for autism: the pathogenetic triad as a theoretical framework. Front Psychiatry. 2021;12:2081.
190. An CE, Nolty AAT, Amano SS, Rizzo AA, Buckwalter JG, Rensberger J. Heart rate variability as an index of resilience. Mil Med. 2020;185:363–369.
191. Carrive P. Orexin, stress and central cardiovascular control A Link with Hypertension?. Neurosci Biobehav Rev. 2017;74:376–392.
192. Dergacheva O, Wang X, Huang ZG, et al. Hypocretin-1 (orexin-A) facilitates inhibitory and diminishes excitatory synaptic pathways to cardiac vagal neurons in the nucleus ambiguus. J Pharmacol Exp Ther. 2005;314:1322–1327.
193. Li A, Nattie E. Orexin, cardio-respiratory function, and hypertension. Front Neurosci. 2014;8:22.
194. Kumar D, Dedic N, Flachskamm C, Deussing J, Kimura M. Cav1.2 calcium channel is involved in the circadian regulation of sleep. Sleep Med. 2013;14:e175.
195. Buddell T, Friedman V, Drozd CJ, Quinn CC. An autism-causing calcium channel variant functions with selective autophagy to alter axon targeting and behavior. PLoS Genet. 2019;15:e1008488.
 © 2022 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2022 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Detecting Time Concept Competence in Children with Autism Spectrum and Attention Disorders
Hus Y
Neuropsychiatric Disease and Treatment 2022, 18:2323-2348
Published Date: 14 October 2022
The ESSENCE-Questionnaire in Medical Records Screening for Neurodevelopmental Symptoms/Problems: Utility and Clinical Validity
Landgren V, Raanan Soltis Z, Svensson E, Theodosiou M, Landgren M, Knez R
Neuropsychiatric Disease and Treatment 2022, 18:2559-2574
Published Date: 3 November 2022
A Systematic Review of the Relationship Between Neurodiversity and Psychosexual Functioning in Individuals with Autism Spectrum Disorder (ASD) or Attention-Deficit/Hyperactivity Disorder (ADHD)
Young S, Cocallis K
Neuropsychiatric Disease and Treatment 2023, 19:1379-1395
Published Date: 2 June 2023
Vitamin Interventions in ASD and ADHD: Systematic Review and Meta-Analysis
Shen Y, Xie Y, Zheng Y, Zheng Y, Liu Y
Neuropsychiatric Disease and Treatment 2025, 21:1845-1855
Published Date: 30 August 2025
Maternal Diabetes and Attention Deficit/Hyperactivity Disorder Association: A Narrative Review on Emerging Evidence, Research Gaps and Future Directions
Zhao C, Liu Y
International Journal of General Medicine 2026, 19:602877
Published Date: 22 May 2026