Back to Journals » Pediatric Health, Medicine and Therapeutics » Volume 15
Oral Cavity Microbiome Impact on Respiratory Infections Among Children
Authors Crestez AM, Nechita A, Daineanu MP, Busila C, Tatu AL ![]() , Ionescu MA, Martinez JD, Debita M
, Ionescu MA, Martinez JD, Debita M ![]()
Received 6 June 2024
Accepted for publication 3 September 2024
Published 9 October 2024 Volume 2024:15 Pages 311—323
DOI https://doi.org/10.2147/PHMT.S471588
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Professor Laurens Holmes, Jr
Alexandra Mihaela Crestez,1,2 Aurel Nechita,1,2 Miruna Patricia Daineanu,1,2 Camelia Busila,1,2 Alin Laurentiu Tatu,2– 4,* Marius Anton Ionescu,5 Jose Dario Martinez,6,* Mihaela Debita7,8
1Pediatric Department, “Sf. Ioan” Emergency Clinical Pediatric Hospital, Galați, 800494, România; 2Clinical Medical Department, Faculty of Medicine and Pharmacy, “Dunărea de Jos” University, Galați, 800201, România; 3Dermatology Department, “Sfanta Cuvioasa Parascheva” Hospital of Infectious Diseases, Galați, 800179, România; 4Multidisciplinary Integrated Center of Dermatological Interface Research MIC-DIR “Dunărea de Jos” University, Galați, 800201, România; 5Department of Dermatology, University Hospital Saint Louis, Paris, 75475, France; 6Department of Internal Medicine, Faculty of Medicine, University Autonomous of Nuevo León, Monterrey, 66455, Mexico; 7Department of Infectious Disease, “Sfanta Cuvioasa Parascheva” Hospital of Infectious Diseases, Galați, 800179, România; 8Clinical Department, Faculty of Medicine and Pharmacy, “Dunărea de Jos” University, Galați, 800201, România
*These authors contributed equally to this work
Correspondence: Camelia Busila; Miruna Patricia Daineanu, Email [email protected]; [email protected]
Background: The respiratory system, traditionally considered antiseptic, harbors a diverse and dynamic bacterial microbiome. Recent advancements in microbiome research have revealed its significant influence on both innate and adaptive immunity, particularly in the context of respiratory infections in children. This article also provides an overview of the types of bacteria that commonly affect the respiratory system, including Streptococcus pneumoniae, Moraxella catarrhalis and Haemophilus influenzae. These bacteria are prevalent in pediatric populations and significantly contribute to the development and severity of respiratory tract infections (RTIs).
Purpose: This review aims to evaluate the impact of the oral cavity and upper respiratory microbiome on the susceptibility and severity of respiratory infections in pediatric populations. We specifically focus on how early colonization patterns of bacteria such as Moraxella and Streptococcus contribute to the development of respiratory tract infections in children from birth through adolescence.
Methods: A thorough literature review was performed, focusing on studies publishing between 2004 and 2023. The review included research exploring the role of the upper respiratory microbiome in pediatric populations, with a specific focus on children aged birth to 18 years. Emphasis was placed on microbial characterization, the modulation of immune responses in respiratory tract infections, and the potential therapeutic applications of microbiome-targeted interventions.
Results: The findings suggest that the composition and disruption of the upper respiratory microbiome significantly influence clinical outcomes in children with respiratory infections. Notably, dysbiosis in the microbiome has been linked to increased susceptibility to repeated infections, highlighting the importance of maintaining microbial balance for optimal respiratory health.
Conclusion: Understanding the impact of oral cavity and upper respiratory microbiome could lead to improved management and prevention strategies for respiratory infections in children. This review underscores the potential of microbiome modulation, including the use of probiotics as a therapeutic approach to enhance clinical outcomes in pediatric respiratory infections.
Keywords: microbiome, children, immunity, respiratory infections
Introduction
The collection of microorganisms found in a particular habitat, or “ecological niche”, is known as the microbiota; this term is similar to the earlier concept of bacterial flora. The entire ecological niche under consideration is represented by the microbiome in its broadest sense. This comprises both environment-influencing elements (cellular proteases, immune cells, etc). and inhabitant microorganisms (bacteria, viruses, parasites, and fungi). Traditional techniques for identifying microbiota relied heavily on time-consuming and insensitive cultivation methods. The last ten years have seen the amplification and sequencing of DNA encoding bacterial 16S RNA, which has enabled researchers to study the microbiota of skin bacteria, tracts, or organs in both quantitative and qualitative ways.1 This has also allowed researchers to define the microbiome because of the increased sensitivity of detection compared to culture methods.2
With the emergence of high-speed sequencing technologies throughput, or the so-called “new generation” sequencing (Next Generation Sequencing [NGS]), capable of producing a considerable amount of data per sample, an enormous number of studies from the Human Microbiome Project (HMP) have led to significant advancements in describing the different human microbiome. However, many questions about these microbiome still remain unanswered, especially with respect to their potential wealth.3
Some authors consider that there are up to 104 bacteria on the surface or in our body, ten times more than the number of our own cells. These data nevertheless remain controversial, going so far as to decrease by a factor of 10, according to the most recent studies.5
Like all the microbiota described in humans, the composition is better defined, and its physiological role. In particular, it is assumed that it participates in host’s defense.5 The microbiome of airways, whether upper or lower, could act as a true biological barrier, contributing to maintaining normal respiratory functions and protecting the individual from colonization or invasion by external pathogens.6
We will present and discuss our view on the respiratory microbiome in relation to viral respiratory infections in this paper. Because there is limited research on the respiratory mycobiome and virome (fungal and viral microbiome, respectively), we will concentrate on the data related to the bacterial microbiome.
Materials and Methods
We conducted a thorough literature review, focusing on studies published over the past 20 years, specifically from 2004 to 2023, without any language restrictions. The review targeted articles that examined the role of the oral cavity and upper respiratory microbiome in pediatric populations, particularly those investigating the impact of this microbiome on respiratory infections in children from birth through adolescence (ages 0–18 years). The research sought to identify studies involving children with recurrent respiratory infections, where the microbiome had been analyzed to explore the relationship between upper airway dysbiosis and secondary immune deficiencies, which may increase susceptibility to recurrent inflammatory processes. We utilized the PubMed database, using key terms such as “upper respiratory microbiome”, “oral cavity microbiome”, “pediatric respiratory infections”, “microbiome”, and “next-generation sequencing”.
Inclusion and Exclusion Criteria
Studies were included based on the following criteria: population- focus on pediatric populations (children aged 0–18 years); microbiome data- detailed microbiome data, particularly regarding microbial diversity, taxonomic composition (eg, genus and species levels), and functional gene profiling; clinical focus- examination of the relationship between the oral cavity and upper respiratory microbiome and respiratory tract infections (RTIs), including both bacterial and viral infections; methodological approach- utilization of advanced microbiome analysis techniques, such as next-generation sequencing (NGS), with robust data presentation and control for confounding factors.
Exclusion criteria included: studies outside the age range of 0–18 years, research focusing on microbiome of other body sites unrelated to the upper respiratory tract, studies lacking sufficient microbiome characterization or not utilizing NGS methodologies, case studies and opinion pieces.
Data Extraction and Synthesis
Data from selected studies were extracted using a standardized extraction form. This included information on study characteristics (author, publication year, study design, sample size, and demographic details), microbiome characteristics (types of sequencing platforms, bioinformatics tools, and methods used for microbiome characterization), and clinical data (types of respiratory infections, clinical outcomes, and reported associations between microbiome composition and disease severity). Discrepancies in data extraction were resolved through consensus.
The synthesis focused on identifying common trends and key findings across studies regarding the composition of the oral cavity and upper respiratory microbiome and its association with respiratory infections in children. The review incorporated both quantitative synthesis (where applicable) and qualitative narrative synthesis to describe patterns, correlations, and potential mechanisms.
In selecting key species reported in the articles, we considered the frequency and consistency of these species’ occurrence across the various studies reviewed. Although complex statistical methods, such as correlation coefficients or network analyses, were not applied, we based the selection of key species on a consensus among the included studies, emphasizing those species repeatedly mentioned as significantly impacting respiratory health in children. This approach was supplemented by a qualitative analysis to assess the relevance of these species concerning our study’s objectives, ensuring they are representative of the pediatric population analyzed and the respiratory pathologies discussed.
Limitations
We acknowledged the limitations of the review process, including potential biases in study selection, variations in study designs, and differences in microbial analysis techniques.
Characteristics of the Respiratory Microbiome in Healthy People
The analysis of a large-scale studies has provided important information about the structure and composition of the microbiome in healthy subjects across different ecological niches (Figure 1).7
These studies, which also focus on the respiratory microbiome, have made it possible to distinguish two types of microbiome: “central” and “satellite”. The central respiratory microbiome (known as “core” microbiome) is composed of bacterial species that are consistently present in more than 95% of the sampled sites and are shared among healthy individuals.8
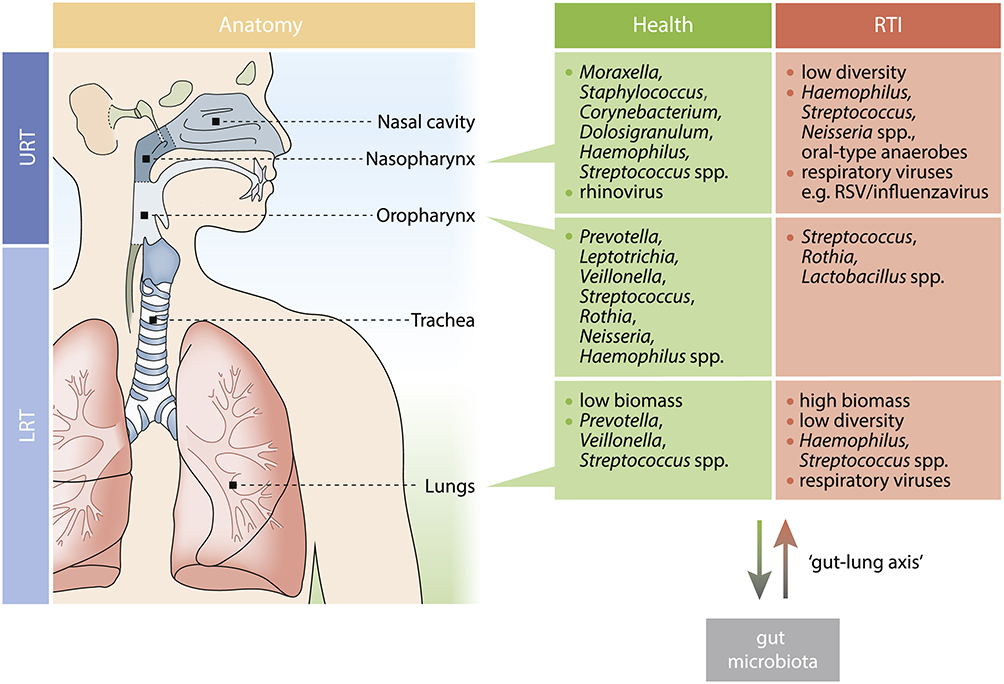 |
Figure 1 Key Microbial Community Members in Health and during RTI for Each Respiratory Niche. Notes: Reproduced from de Steenhuijsen Piters WAA, Binkowska J, Bogaert D. Early Life Microbiota and Respiratory Tract Infections. Cell Host Microbe. 2020;28(2):223–232, with permission from Elsevier.9. |
This microbiome comprises microbial genes and metabolic pathways that enable stable physiology. It should be noted that the central microbiome differs constitutively according to factors such as age, sex, social and/or ethnic origin of the individuals, but it remains influenced by environmental and/or extrinsic factors such as long-treatment courses or diet.10 The mode of childbirth is a fundamental factor influencing the variation (both quantitative and qualitative) of this respiratory microbiome at birth [8]. Infants born via caesarean section have respiratory microbiome profiles dominated by Staphylococcus aureus for a longer time, unlike children born vaginally, whose central respiratory microbiome evolves more quickly towards a more complex profile, including genera such as Moraxella, Corynebacterium or Dolosigranulum. This mature profile offers better protection against respiratory infections. It is also reinforced by breastfeeding. Additionally, the nasopharyngeal microbiome of preterm infants appears to be more heterogeneous than that of full-term infants.11
Although the composition of the microbiome varies considerably during the first year of life, the profiles of central bacteria gradually converge, very early, towards a mature profile by the age of two months, and remain stable thereafter. In contrast to the central microbiome, the peripheral microbiome, called “satellite” microbiome, corresponds to the remaining 5% and is subject to rapid and reversible variations during various physio-pathological alterations.12
Early Respiratory Microbiota Composition, Growth and RTIs in Children
Numerous longitudinal (birth) studies have shown a correlation between early respiratory microbiota and susceptibility to respiratory tract infections (RTIs), as well as later respiratory health.7,12,13 Alternatively, other events that coincide with the timing of microbiota colonization events may be partially responsible for the increased vulnerability to RTIs. One such event is the increasing loss of passive immunity caused by declining IgG levels.14 It is important to remember that these are not exclusive occurrences, and it is possible for declining maternal immunity to combine with infant-specific immunological and microbiota-mediated effects. The timing of colonization episodes appears to be crucial for the link between RTI susceptibility and microbiota development. The early presence of Moraxella spp. has been shown to trigger an inflammatory immune response of the respiratory mucosa and secondary wheezing. Accordingly, a greater chance of early lower respiratory tract infections (LRTIs) was linked to early Streptococcus enrichment (~9 weeks).13 It should be highlighted that, in addition to the date of colonization, these associations are made at the species or strain level, as evidenced by the variations in respiratory tract infection risk for those colonized in early childhood by Moraxella lincolnii or catarrhalis.7 Studies have shown that carrying certain infections, including S. pneumoniae, is a risk factor for RTIs, even beyond the role of commensals that may have immunomodulatory effects.15 In the past, H. influenzae and S. pneumoniae were closely linked to serious diseases such as bacterial pneumonia. Though these pathogens have a well-established and crucial function during overt bacterial RTIs, new research indicates that they may also be responsible for a broader variety of illness phenotypes, including intercurrent wheezing and bronchiolitis, which are typically thought to have a viral origin.4
Figure 1 illustrates how the anatomical niche affects the bacterial community’s makeup in the respiratory system. While a number of recent studies have demonstrated that a fetus is exposed to its mother’s microbiome throughout pregnancy, there is a chance that viable Micrococcus spp. is only sparsely dispersed in the fetal gut. Most of the newborn’s microbiome is acquired at birth; these bacteria are mostly derived from the mother. Crucially, throughout the perinatal and neonatal stages, host and environmental variables have a major influence on these microbial maturation patterns. The upper respiratory tract (URT) microbiota in children varies greatly depending on the delivery method; in infants delivered vaginally, Corynebacterium/Dolosigranulum spp. are typically abundant early and for a prolonged period of time, whereas in infants delivered by cesarean section, S. aureus persists, and anaerobes such as Prevotella, Veillonella, and Porphyromonas spp. emerge. Feeding technique also influences the microbiome, showing similarities between vaginal birth and communities supported by breastfeeding. There can be similarities between the makeup of the microbiota after vaginal delivery and nursing.16,17
Being exposed to congested situations, like attending daycare and having younger siblings, is another important factor that influences the microbiota’s composition and growth in the URT.18
These components, which are also established risk factors for the development of RTIs, might be explained by the related microbiological variables. Some observations have been made, including an early enrichment of Moraxella and an increase in the number of Haemophilus species.12 Isolating many variables that affect the microbiota’s composition is difficult because of their intimate relationships. For example, women who have had C-sections are more likely to experience early cessation of nursing.17 Furthermore, the composition of the bacteria in breast milk appears to be strongly impacted by the manner of birth, making it difficult to emphasise the research on their separate effects.19
The components of the lower respiratory tract (LRT) microbiota have not received much attention in research, which is very difficult considering the low bacterial density and restricted accessibility of this niche, particularly in healthy newborns. However, one study that examined the tracheal aspirations of premature newborn and full-term newborn revealed that composition of the microbiome varied most depending on the gestational age, with term children having adult-like anaerobic profile that included Fusobacterium, Prevotella, Veillonella, and preterm newborn having early Staphylococcus or Ureaplasma.20
Like other habitats, birth mode (only in preterm children) and postnatal age (only in term newborns) were variables influencing this early LRT microbiota.21
At several points in the development of RTI, the significance of respiratory microbiome composition has been proposed (Figure 2).7
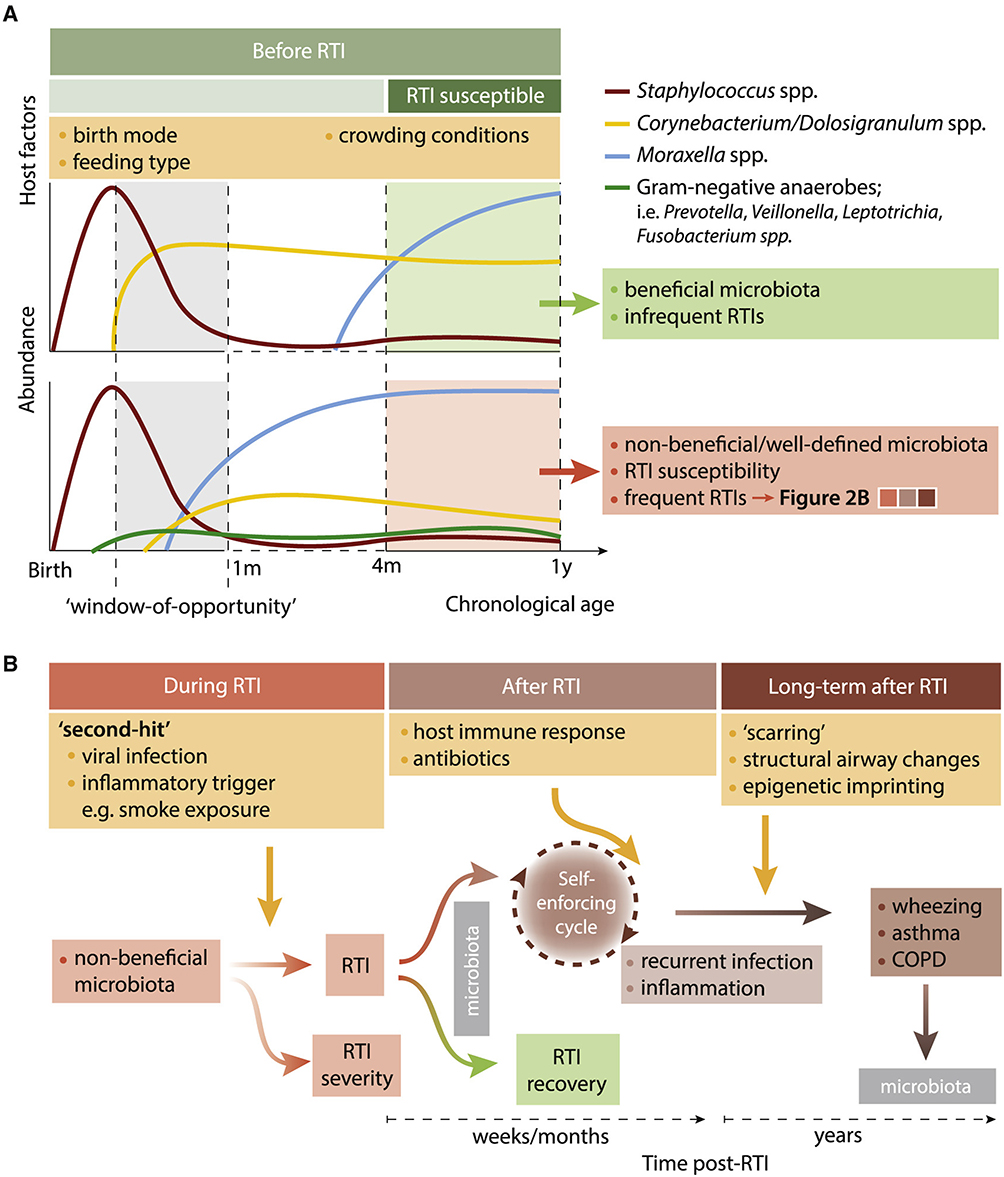 |
Figure 2 Impact of the Respiratory Microbiota before, during, and after RTIs (A). Prior to the first RTI episodes at the age of 4 months, the respiratory microbiota already shows distinct microbial maturation patterns related to consecutive RTI susceptibility (B). Notes: Reproduced from de Steenhuijsen Piters WAA, Binkowska J, Bogaert D. Early Life Microbiota and Respiratory Tract Infections. Cell Host Microbe. 2020;28(2):223–232, with permission from Elsevier.9. |
Impact of the Respiratory Microbiota on the Pathogenesis of Respiratory Viral Diseases
The respiratory system is always in communication with the outside world.22 Previous investigations on the microorganisms found in respiratory samples examined for pathogen research. High-throughput sequencing technologies have progressively made it possible to characterize and describe the commensal resident bacterial flora and examine how it affects innate or acquired immune responses throughout the respiratory tree.23 Only a tiny percentage of the over 7000 publications that returned results when the term “microbiome” was used in 2016 specifically discussed the pulmonary microbiome or the evolution of the microbiome in general during viral infections (Figure 3).
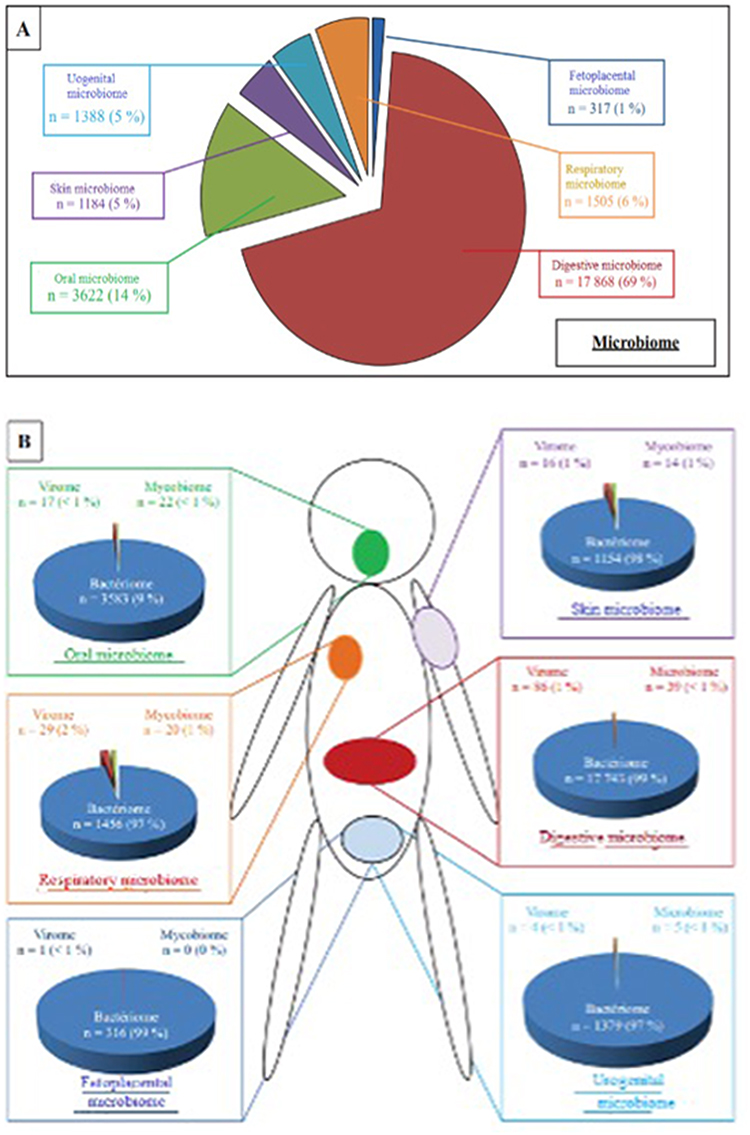 |
Figure 3 Distribution of scientific publications concerning microbiome by anatomical site. (A) Distribution of publications according to ecological niche. (B) Distribution of publications according to the different bacterial, viral and mycotic microbiome. |
With the exception of a few uncommon articles (one to two percent) that examined the fungal (mycobiome) and viral components (virome) microbiome, there is little data in this field of study (Figure 3B).
The most recent research has concentrated on the changes seen in so-called “pathological” scenarios, regardless of whether an infection is present or not, after an initial phase of characterizing the physiological fluctuations and composition of the “healthy” respiratory system microbiome (Figure 3A). Viral infections are the primary cause of acute respiratory infections. Less than thirty percent of the causes of low respiratory infections (bronchial or lung damage) are not viral.24 As a result, the scope of this review is restricted to descriptions of the changes that viral infections bring about in the respiratory system. According to this investigation, the lung microbiome of infected patients contain bacteria that express more genes, which enhances their motility (chemotaxis, flagella), as well as their ability to regulate transcription or metabolism, in comparison to bacteria found in the microbiome of “healthy” patients. The hypothesis that there is a process of metabolically based selection of bacteria that may adapt to a changing environment within an open microbiome is supported by these comparative metatranscriptomics data. These bacterial species would be able to travel into the lower airways and create immunomodulatory compounds that would restrict the host’s reaction to viral infection, especially by altering the mucociliary clearance. Therefore, these bacteria may have a significant effect on the therapeutic state, whether positive or negative, of hosts that are infected.25 A less explored role could be related to the existence of endosymbiotic bacteria at the level of the face or oral cavity, in connection with the presence of Demodex folliculorum and dental changes.26–29
A specific immunological tolerance fostered by a microbiome might prevent tissue damage from the heightened inflammatory response in the event of a recognised respiratory disease exacerbated by a viral infection. In a study, the authors were able to show the significance of a healthy respiratory microbiome on an organism’s ability to withstand the influenza virus using a mouse model. They achieved this by using mice that were known to be more sensitive to influenza infections than wild mice of the same genotype bred in a non-sterile natural setting (ie, greater weight loss, poorer survival rate, etc). These mice were called SPF (Specific Pathogen Free) mice. The colonization of commensal bacteria in the upper respiratory tract of the SPF mouse model allowed for a significant reduction in lung changes associated with infections.
By altering the monocyte response phenotype and generating an activation phenotype of less destructive macrophages, this colonization decreased the lethality of influenza infection.13 Additionally, mice treated with a combination of antibiotics, “naturally” denaturing their microbiome, exhibited greater morbidity when infected with the 2009 pandemic A (H1N1) virus, resulting in considerable oxygen desaturation and weight loss. Two characteristics were linked to the severity of these injuries: (1) a decrease in dendritic cell migratory capability, which is necessary for a successful immune response; and (2) a reduction in the number of tissue-specific T lymphocyte cells, which helps to sustain a high local viral load over time.30,31
Oral probiotic therapy dramatically improved the immune response of antibiotic-treated animals, enabling the improved activation of TLR-7 and NFB pathways.32 In this manner, viral clearance is aided by signaling, which is involved in the identification of single-stranded viral RNA. All of the information gathered from animal studies points to a potential connection between a weakened local immune response and the pathophysiology of influenza virus infection in a host with a modified microbiome. On the other hand, maintaining both a suitable response to the influenza virus and an efficient innate response is made feasible by the presence of an “integrous” microbiome.33
However, the viable microbes that comprise this “beneficial“ microbiome do not appear to be necessary. In fact, lipopolysaccharides (LPS) administered to a healthy host with an ”altered” microbiome could successfully reestablish an effective adaptive immune response in an experimental setting using a mouse model. According to the mechanistic concept, leukocytes are stimulated both locally and systemically by the microbiome, which also activates several molecular pattern recognition receptors produced by pathogens (Pathogen-Associated Molecular Patterns, or PAMPs).34,35
The impact of the microbiome in the pathogenesis of viral respiratory infections is therefore strongly suspected. By analogy with therapies of the gastrointestinal tract, it seems possible to modulate the respiratory microbiome. A mouse model was developed with direct administration of Escherichia coli into the airway tract. These inoculations showed a beneficial effect on local inflammation, which is strongly involved in the pathogenesis of respiratory infections, thus limiting clinical symptoms.36 Likewise, a clinical trial showed the positive influence of administering Lactobacillus rhamnosus (administered orally, far from the respiratory sphere) on the number and severity of cystic fibrosis exacerbations.37 Although requiring validation by cohort studies of sufficient size, the benefit of therapeutic modification of the respiratory microbiome in the context of chronic respiratory diseases seems strongly suggested by these studies. However, few studies are focused on the modification of the respiratory microbiome in the context of acute viral infections.
Some authors have nevertheless shown that the temporary relief of airway inflammation through the administration of bacterial antigens could be a treatment approach for influenza virus infections.34 In a murine model, it has also been shown that animals pretreated with aerosols of bacterial lysates containing LPS presented a faster and more efficient inflammatory response in obtaining viral clearance during a subsequent viral infection. The induced response by LPS reduced tissue damage linked to viral infection, improving the survival of infected animals.5 These data, however, remain contested in the specific context of influenza virus infection. In fact, it is admitted that the inflammation generated by the viral infection is responsible for the majority of the clinical manifestations observed; however, the use of immunomodulatory treatments has not been shown to be effective in clinical trials. The situation, therefore, seems more complex. As already studied for the management of severe sepsis, the control of the inflammatory response is only of interest in the early phase of the disease. A large part of the difficulty in controlling inflammation during influenza infection is thus linked to the fact that it varies over time, from early to late stages of the disease.38 Similarly, the activation of TLR-2, 6 and 9 by means of pretreatment with synthetic agonists made it possible to reduce the viral titer measured in infections by parainfluenza viruses in respiratory samples without helping to reduce symptoms of bronchial hyperreactivity.39 Finally, the administration of inactivated toxins of E. coli has demonstrated its effectiveness in providing protection against RSV infection and influenza viruses, as well as the administration of L. rhamnosus before contact with RSV.23,35 These lactobacilli would help protect against excess inflammation during the infection, modifying both the production of numerous pro-inflammatory mediators (interferon and interleukins) and activation of dendritic cells via the expression of CD103 and CD11b.40
Microbiota-Mediated Mechanism During RTIs
It was discovered that nicotine exposure in mice only affected the balance of the microbial population when the pathogen S. pneumoniae was present. In this case, the concurrent viral infection might also be considered the “second hit”. In line with this theory, kids with a Moraxella-enriched microbiome did not show any symptoms until a virus was found, which led to LRTI.13 Regardless of phenotype, including suspected (bacterial) pneumonia cases, the presence of a virus distinguished the healthy population from those with RTI in a mixed bacterial-viral diagnostic prototype of lower respiratory tract infections (LRTI).6 Viral infections may exacerbate the microbiome composition and affect disease severity and phenotype. Accordingly, hospitalization rates were higher, and genes linked to TLR signaling and neutrophil and monocyte recruitment were expressed more in infants infected with RSV who also had a Streptococcus/Haemophilus dominated microbiota profile.12 For kids whose microbiota was dominated by Staphylococcus, the opposite was true. A self-reinforcing loop of inflammation and microbial alterations may be initiated by subsequent pathogen-induced host inflammation, encouraging a more pathogenic expansion.20
Impact of Microbial Perturbation After RTIs
Research highlights changes in the makeup of the microbiota during RTIs and underscores the significance of early respiratory microbiome involvement in RTIs, as was previously mentioned. We hypothesize that this altered composition of the microbiota contributes to increased symptomatology during RTIs, decreased colonization resistance of the microbiome, symptom persistence, and increased risk of recurrences. Among other things, this is corroborated by recent research that revealed prolonged symptoms in the weeks following an infant’s RTI and the enrichment of Streptococcus three weeks later.15 Furthermore, antibiotic therapy, which is frequently used for resistant respiratory tract infections (RTIs), likely disrupts microbial communities that are already under stress, which might reduce their ability to fight off newly acquired pathogens after infection. Although this has not been investigated in a randomized controlled environment yet, it might help explain why acute otitis media recurrence was discovered more frequently following antibiotic therapy.21 Although the precise processes causing this phenomenon are still unknown, they most likely include a self-reinforcing loop in which particular microbial communities may sustain or foster an environment that is pro-inflammatory, thereby allowing pathogen breakouts. A vicious cycle is about to begin when these pathogens trigger an additional pro-inflammatory cascade. (Figure 2).
If the condition is not brought back to equilibrium, these alterations may persist and result in long-term health effects. Research on children who are prone to asthma, among other populations, indicates that the hypopharyngeal microbiota composition (Veillonella, Prevotella, and Gemella spp. at birth) is linked to a pro-inflammatory profile in the respiratory system and asthma diagnosis at six years old. This new study expands on earlier research that found a pro-inflammatory immunological reaction in the airways of asthma-prone newborns at one month of age, associated with M. catarrhalis and H. influenzae.20 Nevertheless, further research is required to determine the precise causal linkages behind these findings. Epigenetic imprinting is a conceivable mechanism by which microbial and immunological disturbances during early life might potentially modify physiological activities of the respiratory tract later. According to in vitro studies and molecular epidemiological research, RSV infection at an early age does cause histone and DNA methylation variations in a number of immune-altering genes connected to asthma. Additionally, as shown for gut commensals, beneficial microorganisms may modify the human epigenome by producing short-chain fatty acids (SCFAs). An in vitro investigation that showed that SCFAs generated from the nasal mucosa following exposure to C. accolens suppressed the development of S. pneumoniae indicated the importance of these SCFAs for the respiratory tract.21
There is a link between the presence of bacteria such as Haemophilus and Moraxella in the lower airways of children with bronchial asthma, where bacteria such as Bacteroidetes, Actinobacteria and Firmicutes should be predominant, and the development of bronchial hyperresponsiveness and bronchoconstriction. This process can be accentuated by respiratory allergens, air pollutants, antibiotic consumption and antacid medication. This creates dysbiosis in both the gut and the lung that bidirectionally affects the gut-lung axis, resulting in hypersensitivity and hyperresponsiveness to respiratory allergens and foods.41
Use in Biomarkers
It was quickly envisioned to use the microbiome as a risk marker for the occurrence of severe respiratory infection. Indeed, even before the concept of the microbiome was described, it had been observed that the detection and subsequent increase in the proportion of certain bacterial species (Pseudomonas aeruginosa, for example) in the respiratory tract signaled a negative turning point in the progression of the disease in patients with cystic fibrosis.42
Knowledge of an individual’s microbiome could make it possible to predict their level of risk concerning bacterial or viral respiratory attacks. Even though data is still limited, it has recently been described that nasal carriage of Pneumococcus was statistically positively correlated with Metapneumovirus seropositivity in children.43 Therefore, after verification in a cellular model, authors concluded the involvement of this bacterium in the susceptibility of an organism, in this case, a child, to infection by Metapneumovirus. In a published article, our team studied the microbiome of children with influenza infections of varying severity. We were able to highlight, in children infected with the influenza A or B virus, that several OTUs (S. aureus, Prevotella, Lachnospiracea incertae sedis, Streptobacillus, Porphyromonas, Granulicatella, Veillonella, Fusobacterium and Haemophilus spp). were present, from the start of the disease, in different proportions, and that this had consequences on the clinical outcome of the infected patient. With the exception of S. aureus, all these OTUs were found in more abundant quantities in patients progressing towards severe cases of influenza, from the initial sample.44 The detection of these OTUs together with that of the influenza virus could lead to offering appropriate support at the start of the infection. The initial orientation of patients, optimized due to the existence or absence of an identified risk of respiratory complications, would improve their management. A multicenter study, started in 2017, will make it possible to confirm the reality of this difference and especially the usefulness of such a biomarker for managing children with the flu. The microbiome could therefore serve, ultimately, as a prognostic marker by helping to take into account the early management of at-risk patients.
Although no causal relationship between the characteristics of the nasopharyngeal microbiome and the response to viral infections can be deduced from these studies, the data currently available nevertheless suggests that studying the microbiome could make it possible to obtain a predictive factor for the clinical consequences of this infection. The use of microbiome characteristics in the management of infected patients could therefore be considered. Conversely, a modification of the bacterial microbiome, through vaccination or other administration, could have consequences on infectious diseases, whether or not they are part of a strategy for vaccine prevention.7,23
Future Perspectives
The traditional “one pathogen-one disease” model has begun to give way to an ecosystem- wide explanation of RTI pathogenesis within the past few years. This hypothesis explains how immunity is modulated by microbe-host interactions and how resilience and susceptibility to RTIs are related. Current research has contributed to redefining a preventive, diagnostic, and treatment approaches in addition to deepening our understanding of the pathophysiology of RTIs.6 Additionally, real-time genetic analysis has been utilized to identify the microbiological cause of bacterial respiratory infection while also facilitating the identification of genes associated with bacterial resistance. This has allowed for the administration of customized antibiotics at the early stages of illness. The use of Neisseria lactamica to “replace” and stop the spread of the Meningococcal is one example of how this information may also result in innovative biotherapeutic techniques. While these initial instances are encouraging, more work has to be done to understand the basic mechanisms behind a stable balance between the host, microorganisms, and the external environment. We suggest using methodologies combining systems biology and model systems to achieve this.43
Even if it is crucial to identify correlations among the structure and the function of the microbiota and health outcomes at different phases of RTI pathogenesis, there are still a lot of unproven causal connections that should be taken into consideration. Some answers to such issues may be provided by using current developments in genetics and engineering. Microbiome interactions may be evaluated using gain or loss-of-function tests with genetically modified versions of model organisms in conjunction with next-generation sequencing methods.32 Moreover, sophisticated laboratory simulations, such as airway organoids, could be utilized to investigate microbe interactions in different environments. Gene-editing methods may be applied in vivo in animal models and organoid systems to modify the tissue host or the genome. Notwithstanding these developments, we think human challenge studies have potential since in vivo and in vitro models are by definition less complicated than human physiology and environmental variation.12
Unlike observational research, these investigations shed light on the dynamics of the microbiota surrounding pathogen acquisition and permit the modification of individual factors, including assessing vaccine interactions or co-colonizing bacteria or viruses.12 Similar to the field of vaccination, which has recently adopted a “systems science” approach, an analogous multi-omics approach may potentially be helpful in understanding the genesis of RTIs. Adding viruses, phages and fungi to the currently understudied non-bacterial microbial community will allow us to capture actual ecosystem-wide microbiota changes at even greater depth. This will enable us to investigate the interactions between the host, microorganisms, and environment in connection to the etiology and manifestations of respiratory disorders in greater detail.4
Conclusion
Recent studies have increasingly shown just how crucial the oral cavity and respiratory microbiome are in influencing the body’s response to respiratory viral infections. While much of the research so far has been done using in vitro models, chronic infection models, or specific populations like immunocompromised individuals and neonates, it’s becoming clear that the human microbiome significantly impacts both how these infections develop and how our bodies respond to them.
There is growing evidence that the microbiome could be useful not just as a diagnostic tool but also as a target for treatment in clinical practice. The respiratory microbiome plays a dual role: it not only affects the progression of diseases but also reflects their current state, making it an important area for developing new intervention strategies. With the continuous advancement of high-throughput sequencing technologies, we are likely to see more precise identification of biomarkers for disease severity, which could drive forward the development of personalized medicine.
Moreover, focusing on correcting imbalances within the satellite microbiome—variations of which can be influenced by environmental factors—could lead to better therapeutic and prognostic outcomes in the treatment of infectious diseases. This approach is already being explored in the context of inflammatory diseases. The microbiome, as a field of research, offers vast potential to deepen our understanding of the complex interactions between microbial communities, viruses, and fungi. Although our knowledge in these areas is still growing, they are likely to be central to significant scientific breakthroughs in the near future.
Abbreviations
CFU, Colony Forming Units; DNA, Deoxyribonucleic acid; HMP, Human Microbiome Project; LPS, Lipopolysaccharides; LRTI, Lower respiratory tract infections; NFB, Nuclear Factor B; NGS, Next Generation Sequencing; OTUs, Operational Taxonomic Units; PAMPS, Pathogen Associated Molecular Patterns; RSV, Respiratory Syncytial Virus; RTI, Respiratory Tract Infection; RTIs, Respiratory Tract Infections; RNA, Ribonucleic acid; SCTAs, Short Chain Fatty Acids; SPF, Specific Pathogen Free; TLR, Toll-Like Receptors; URT, Upper respiratory tract; LRT, Lower respiratory tract.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
The research was funded by “Dunarea de Jos” University, Galati, Romania.
Disclosure
The authors declare no conflict of interest.
References
1. Hofmann M, Schulz-Weidner N, Krämer N, Hain T. The bacterial oral microbiome in children with congenital heart disease: an extensive review. Pathogens. 2023;12:1269. doi:10.3390/pathogens12101269
2. Iwai S, Fei M, Huang D, et al. Oral and airway microbiota in HIV infected pneumonia patients. J Clin Microbiol. 2012;50:2995–3002. doi:10.1128/JCM.00278-12
3. Peterson J, Garges S, Giovanni M, et al.; The NIH HMP Working Group. The NIH human microbiome project. Genome Res. 19;2009:2317–2323. doi:10.1101/gr.096651.109
4. Teo SM, Mok D, Pham K, et al. The infant nasopharyngeal microbiome impacts severity of lower respiratory infection and risk of asthma development. Cell Host Microbe. 2015;17:704–715. doi:10.1016/j.chom.2015.03.008
5. Sender R, Fuchs S, Milo R. Revised Estimates for the number of human and bacteria cells in the body. PLOS Biol. 2016;14:e1002533. doi:10.1371/journal.pbio.1002533
6. Man WH, van Houten MA, Merelle ME, et al. Bacterial and viral respiratory tract microbiota and host characteristics in children with lower respiratory tract infections: a matched case-control study. Lancet Respir Med. 2019;7:417–426. doi:10.1016/S2213-2600(18)30449-1
7. Biesbroek G, Tsivtsivadze E, Sanders EAM, et al. Early respiratory microbiota composition determines bacterial succession patterns and respiratory health in children. Am J Respir Crit Care Med. 2014;190:1283–1292. doi:10.1164/rccm.201407-1240OC
8. Huse SM, Ye Y, Zhou Y, Fodor AA. A core human microbiome as viewed through 16S rRNA sequence clusters. PLoS One. 2012;7:e34242. doi:10.1371/journal.pone.0034242
9. de Steenhuijsen Piters WAA, Binkowska J, Bogaert D. Early life microbiota and respiratory tract infections. Cell Host Microbe. 2020;28(2):223–232. doi:10.1016/j.chom.2020.07.004
10. Erb-Downward JR, Thompson DL, Han MK, et al. Analysis of the lung microbiome in the “healthy” smoker and in COPD. PLoS One. 2011;6:e16384. doi:10.1371/journal.pone.0016384
11. Perez GF, Pérez-Losada M, Isaza N, et al. Nasopharyngeal microbiome in premature infants and stability during rhinovirus infection. J Investig Med. 2017;65:984–990. doi:10.1136/jim-2017-000414
12. Bosch AATM, de Steenhuijsen Piters WAA, van Houten MA, et al. Maturation of the infant respiratory microbiota, environmental drivers, and health consequences. a prospective cohort study. Am J Respir Crit Care Med. 2017;196:1582–1590. doi:10.1164/rccm.201703-0554OC
13. Wang J, Li F, Sun R, et al. Bacterial colonization dampens influenza mediated acute lung injury via induction of M2 alveolar macrophages. Nat Commun. 2013;2013:4.
14. Jennewein MF, Abu-Raya B, Jiang Y, Alter G, Marchant A. Transfer of maternal immunity and programming of the newborn immune system. Immunopathology. 2017;39:605–613. doi:10.1007/s00281-017-0653-x
15. Bogaert D, De Groot R, Hermans PWM. Streptococcus pneumoniae colonization: the key to pneumococcal disease. Lancet Infect Dis. 2004;144–154. doi:10.1016/S1473-3099(04)00938-7
16. Chu DM, Ma J, Prince AL, et al. Maturation of the infant microbiome community structure and function across multiple body sites and in relation to mode of delivery. Nat Med. 2017;23:314–326. doi:10.1038/nm.4272
17. Hobbs AJ, Mannion CA, McDonald SW, Brockway M, Rough SC. The impact of caesarean section on breastfeeding initiation, duration and difficulties in the first four months postpartum. BMC Pregnancy Childbirth. 2016;16:90. doi:10.1186/s12884-016-0876-1
18. Tatu AL, Nwabudike LC, Kubiak K, et al. Endosymbiosis and its significance in dermatology. J Eur Acad Dermatol Venereol. 2018;32(9). doi:10.1111/jdv.14921
19. Moosavi S, Sepehri S, Robertson B, et al. Composition and variation of the human milk microbiota are influenced by maternal and early-life factors. Cell Host Microbe. 2019;25:324–325. doi:10.1016/j.chom.2019.01.011
20. Folsgard NV, Schjoring S, Chawes BL, et al. Pathogenic bacteria colonizing the airways in asymptomatic neonates stimulates topical inflammatory mediator release. Am J Respir Crit Care Med. 2013;187:589–595. doi:10.1164/rccm.201207-1297OC
21. Pattaroni C, Watzenboeck ML, Schneidegger S, et al. Early life Formation of the microbial and immunological environment of the human airways. Cell Host Microbe. 2018;24:857–865. doi:10.1016/j.chom.2018.10.019
22. Lynch SV. Viruses and microbiome alterations. Ann Am Thorac Soc. 2014;11 Suppl 1:S57–60. doi:10.1513/AnnalsATS.201306-158MG
23. Williams AE, Edwards L, Humphreys IR, et al. Innate imprinting by the modified heat-labile toxin of Escherichia coli (LTK63) provides generic protection against lung infectious disease. J Immunol. 2004;173:7435–7443. doi:10.4049/jimmunol.173.12.7435
24. Jain P, Jain A, Prakash S, et al. Prevalence and genotypic characterization of human parvovirus B19 in children with hemato-oncological disorders in North India. J Med Virol. 2015;87:303–309. doi:10.1002/jmv.24028
25. Segal LN, Blaser MJ. A brave new world: the lung microbiota in an era of change. Ann Am Thorac Soc. 2014;11 Suppl 1:S21–7. doi:10.1513/AnnalsATS.201306-189MG
26. Tatu AL, Clatici VG, Nwabudike LC. Rosacea-like demodicosis (but not primary demodicosis) and papulopustular rosacea may be two phenotypes of the same disease - a microbioma, therapeutic and diagnostic tools perspective. J Eur Acad Dermatol Venereol. 2019;33(1). doi:10.1111/jdv.15166
27. Tatu AL, Ionescu MA, Cristea VC. Demodex folliculorum associated Bacillus pumilus in lesional areas in rosacea. Indian J Dermatol Venereol Leprol. 2017;83(5):610–611. doi:10.4103/ijdvl.IJDVL_921_16
28. Pricop R, Cristea VC, Gheorghe I, Tatu AL, Mihaescu G, Chifiriuc MC. Matrix-assisted laser desorption/ionization time-of-flight mass spectrometry (MALDI-TOF MS) reveals the anaerobic Slakia exigua as unique etiology of a dental abscess. Biointerf Res Appl Chem. 2017;7(2):1995–1997.
29. Abt. MC, Osborne LC, Monticelli LA, et al. Commensal bacteria calibrate the activation threshold of innate antiviral immunity. Immunity. 2012;37:158–170. doi:10.1016/j.immuni.2012.04.011
30. Leung R-K-K, Zhou J-W, Guan W, et al. Modulation of potential respiratory pathogens by pH1N1 viral infection. Raoult D, éditeur. Clin Microbiol Infect. 2013;19:930–935. doi:10.1111/1469-0691.12054
31. Ansari I, Raddatz G, Gutenkust J, et al. The microbiota programs DNAmetylation to control intestinal homeostasis and inflammation. Nat Microbiol. 2020;5:610–619. doi:10.1038/s41564-019-0659-3
32. Tuvim MJ, Evans SE, Clement CG, et al. Augmented lung inflammation protects against influenza A pneumonia. PLoS One. 2009;4:e4176. doi:10.1371/journal.pone.0004176
33. Ichinohe T, Pang IK, Kumamoto Y, et al. Microbiota regulates immune defense against respiratory tract influenza A virus infection. PNAS USA. 2011;108:5354–5359. doi:10.1073/pnas.1019378108
34. Norton EB, Clements JD, Voss TG, Cárdenas-Freytag L. Prophylactic administration of bacterially derived immunomodulators improves the outcome of influenza virus infection in a murine model. J Virol. 2010;84:2983–2995. doi:10.1128/JVI.01805-09
35. Nembrini C, Sichelstiel A, Kisielow J, et al. Bacterial-induced protection against allergic inflammation through a multicomponent immunoregulatory mechanism. Thorax. 2011;66:755–763. doi:10.1136/thx.2010.152512
36. Bruzzese E, Raia V, Spagnuolo MI, et al. Effect of Lactobacillus GG supplementation on pulmonary exacerbations in patients with cystic fibrosis: a pilot study. Clin Nutr. 2007;26:322–328. doi:10.1016/j.clnu.2007.01.004
37. Monneret G, Venet F. Sepsis-induced immune alterations monitoring by flow cytometry as a promising tool for individualized therapy. Cytometry B Clin Cytom. 2016;90:376–386. doi:10.1002/cyto.b.21270
38. Drake MG, Evans SE, Dickey BF, Fryer AD, Jacoby DB. Toll-like receptor-2/6 and Toll-like receptor-9 agonists suppress viral replication but not airway hyperreactivity in Guinea pigs. Am J Respir Cell Mol Biol. 2013;48:790–796. doi:10.1165/rcmb.2012-0498OC
39. Chiba E, Tomosada Y, Vizoso-Pinto MG, et al. Immunobiotic Lactobacillus rhamnosus improves resistance of infant mice against respiratory syncytial virus infection. Int Immunopharmacol. 2013;17:373–382. doi:10.1016/j.intimp.2013.06.024
40. Mayer-Hamblett N, Rosenfeld M, Gibson RL, et al. Pseudomonas aeruginosa in vitro phenotypes distinguish cystic fibrosis infection stages and outcomes. Am J Respir Crit Care Med. 2014;190:289–297. doi:10.1164/rccm.201404-0681OC
41. Hufnagl K, Pali-Schöll I, Roth-Walter F, Jensen-Jarolim E. Dysbiosis of the gut and lung microbiome has a role in asthma. Semin Immunopathol. 2020;42(1):75–93. PMID: 32072252; PMCID: PMC7066092. doi:10.1007/s00281-019-00775-y
42. Verkaik NJ, Nguyen DT, de Vogel CP, et al. Streptococcus pneumoniae exposure is associated with human metapneumovirus seroconversion and increased susceptibility to in vitro HMPV infection. Clin Microbiol Infect. 2011;17:1840–1844. doi:10.1111/j.1469-0691.2011.03480.x
43. Langevin S, Pichon M, Smith E, et al. Early nasopharyngeal microbial signature associated with severe influenza in children: a retrospective pilot study. J Gen Virol. 2017;98:2425–2437. doi:10.1099/jgv.0.000920
44. Tarabichi Y, Li K, Hu S, et al. The administration of intranasal live attenuated influenza vaccine induces changes in the nasal microbiota and nasal epithelium gene expression profiles. Microbiome. 2015;3:74. doi:10.1186/s40168-015-0133-2
 © 2024 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2024 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Five Years’ Experience with Respiratory Syncytial Virus Among Hospitalized Patients: A Retrospective Study from Jazan, Saudi Arabia
Dhayhi NS, Aqeel A, Ghazwani S, Gosadi IM, AlQassimi HM, Thubab A, Sairam IA, Hakami MA, Sawan FA, Asiry SM, Khalifa M, Daghreeri H, Badawy A, Ghawi AA, Arishi H, Almudeer A, Shrwani KJ, Alhazmi AH
Infection and Drug Resistance 2024, 17:5179-5187
Published Date: 23 November 2024