Back to Journals » Journal of Inflammation Research » Volume 16
Non-Coding RNAs Regulate Spinal Cord Injury-Related Neuropathic Pain via Neuroinflammation
Authors Zhu J, Huang F, Hu Y ![]() , Qiao W
, Qiao W ![]() , Guan Y, Zhang ZJ
, Guan Y, Zhang ZJ ![]() , Liu S, Liu Y
, Liu S, Liu Y
Received 20 March 2023
Accepted for publication 2 June 2023
Published 13 June 2023 Volume 2023:16 Pages 2477—2489
DOI https://doi.org/10.2147/JIR.S413264
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Adam Bachstetter
Jing Zhu,1,* Fei Huang,1,2,* Yonglin Hu,1,3,* Wei Qiao,1 Yingchao Guan,1 Zhi-Jun Zhang,1,* Su Liu,1 Ying Liu4
1Department of Rehabilitation Medicine, Affiliated Hospital of Nantong University, Medical School of Nantong University, Nantong, JiangSu Province, 226001, People’s Republic of China; 2Department of Rehabilitation Medicine, Nantong Health College of Jiangsu Province, Nantong, JiangSu Province, 226010, People’s Republic of China; 3Affiliated Nantong Rehabilitation Hospital of Nantong University, Nantong, JiangSu Province, 226001, People’s Republic of China; 4Department of Pathology, Affiliated Hospital of Nantong University, Nantong, JiangSu Province, 226001, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Ying Liu, Department of Pathology, Affiliated Hospital of Nantong University, Nantong, 226001, JiangSu Province, People’s Republic of China, Email [email protected] Su Liu, Department of Rehabilitation Medicine, Affiliated Hospital of Nantong University, Nantong, JiangSu Province, 226001, People’s Republic of China, Email [email protected]
Abstract: Secondary chronic neuropathic pain (NP) in addition to sensory, motor, or autonomic dysfunction can significantly reduce quality of life after spinal cord injury (SCI). The mechanisms of SCI-related NP have been studied in clinical trials and with the use of experimental models. However, in developing new treatment strategies for SCI patients, NP poses new challenges. The inflammatory response following SCI promotes the development of NP. Previous studies suggest that reducing neuroinflammation following SCI can improve NP-related behaviors. Intensive studies of the roles of non-coding RNAs in SCI have discovered that ncRNAs bind target mRNA, act between activated glia, neuronal cells, or other immunocytes, regulate gene expression, inhibit inflammation, and influence the prognosis of NP.
Keywords: non‐coding RNAs, neuroinflammation, neuropathic pain, spinal cord injury
Introduction
Spinal cord injury (SCI) is often accompanied by major impairments in motor, sensory, and autonomic functions.1 The molecular and biochemical pathways associated with the pathophysiology of SCI include intra-tissue hypoxia, oxidative stress, inflammation, apoptosis, and so on. SCI is classified as primary or secondary. Common symptoms of SCI include neuropathic pain (NP), spasticity, abnormal autonomic reflexes, and impairment of social, recreational, and occupational activities.2 Primary SCI can trigger secondary SCI, resulting in further chemical and mechanical damage to tissues. A common secondary complication of SCI is NP, which patients usually experience spontaneously, abnormally, or with hypersensitivity to pain.3 Severe SCI-related NP is associated with decreased quality of life. Conventional treatments have difficulty resolving severe NP, and many clinical needs remain unmet. Despite the many therapeutic strategies proposed and various breakthroughs achieved, cure remains elusive, probably due to the complex healing and protective mechanisms involved.4–6
Neuroinflammation (NI) is a coordinated response of the immune system against infection following SCI.7 In the past decade or so, NI following SCI was linked to intractable NP. Anatomical alterations associated with SCI include disruption of the blood-spinal cord barrier.8,9 During blood-spinal cord barrier disruption, immune cells will enter the spinal cord parenchyma and cause direct inflammation. Neurons, neutrophils, microglia, macrophages, astrocytes, and B and T lymphocytes, as well as cytokines contribute to this mechanistic process. SCI-induced NI produces NP and promotes NP-related behaviors.10–14 Disruption of the blood-spinal cord barrier and subsequent infiltration of leukocytes induces central NI, as well as activation, migration, and proliferation of microglia and astrocytes, which can promote the production and release of inflammatory cytokines, including tumor necrosis factor (TNF)-α, interleukin (IL)-1β, and IL-6, which have been associated with behavioral indicators of SCI-related NP.8,15–19 A growing body of evidence suggests that NI following SCI can prolong NP.7
Non-coding RNAs (ncRNAs), which include microRNAs (miRNAs), long non-coding RNAs (lncRNAs), and circular RNAs (circRNAs), play important roles in regulation of gene expression in central nervous system. In this review, we discuss how ncRNAs expression changes are associated with SCI-related NP. Except for miRNAs, IncRNAs and circRNAs, NP is rarely associated with other types of ncRNAs, and these are not discussed here. Notably, ncRNAs do not code for proteins, but rather regulate the expression of genes and proteins involved in inflammatory and immune responses following SCI through a variety of mechanisms. For example, miRNAs regulate the proliferation of astrocytes to mediate NI, while lncRNAs and circRNAs act as sponges of RNAs or miRNAs during the progression of NP, regulate expression epigenetically of NP-related molecules or modulate procession of miRNAs.20–24
The aim of this review article is to facilitate understanding of SCI-related NP by highlighting the etiological impact of ncRNAs and associations with cells and molecules involved in the pathophysiology of SCI as potential prognostic targets for SCI-related NP.
Epidemiology
Statistics show that the prevalence of NP ranges from 50.7% to 81% in patients with SCI.25–34 The International Association of the Study of Pain classifies SCI-related pain as nociceptive pain or NP.26,35 Among SCI patients, the incidence of nociceptive pain ranges from 40.6% to 59% and NP from 30.2% to 53%.5,25–27,32,36 Due to differences in the definition of pain, diagnostic tools, and study types, the prevalence of SCI-related NP widely varies.37 Nociceptive pain presents as musculoskeletal pain, which is associated with pathological changes to the bones, joints, and/or muscles, resulting in pain of the arm or shoulder, or visceral pain of the abdomen involving renal calculus, bowel, and sphincter dysfunction.35,38,39 NP is classified as at-level or below-level. At-level NP is referred to as the dermatome, which at the level of neuropathic damage and/or within three dermatomes below the level of neurological damage due to disease or injury to the nerve root or spinal cord, whereas below-level NP refers to pain beyond three dermatomes below the level of neuropathic damage due to direct injury to the spinal cord.40,41 SCI-related NP could be spontaneous or stimulus-evoked, continuous or intermittent. SCI patients describe NP as burning, sharp, cramping, cold, squeezing, stinging, or electric shock-like.2,42,43 SCI-related NP is severe in hyperacute time and it mainly occurs in the at-level. Compared to the chronic phase, acute NP is reported as intense electric and cold stimulation that is more prevalent after 6 months. After 1 year, below-level NP starts to increase.5,44 The severity of NP can gradually increase and cause progressive damage to the spinal cord over time.45,46 SCI-related NP negatively influences quality of life via disruption of daily activities, sleep disorders/insomnia, and anxiety/depression.29,33 In addition, SCI-related NP may cause more disabling than physical damage and bring about a sense of isolation and despair.47 Demographic variables related to NP include female sex, advanced age, and financial status.30,36,48 An analysis of clinical data in SCI-NP patients (spinal cord injury associated with neuropathic pain) showed significantly high TNF-α/IL-6 expression and a stronger association of TNF-α with NP in SCI patients.49
Pathophysiological Roles of miRNAs, IncRNAs, and circRNAs in SCI-Related NP via Neuroinflammation
Inflammation‑Linked miRNAs in SCI-Related NP
The mechanisms underlying SCI-related NP involve complex interactions among neuronal cells, glial cells, and non-neuronal cells. Most studies of the roles of miRNAs in SCI-related NP have focused on neurons, astrocytes, microglia, and macrophages. However, a previous study of the interactions of miRNAs in oligodendrocytes found that transplantation of precursor oligodendrocytes attenuated mechanical hypersensitivity reactions.50 During SCI, miRNAs regulate the functions of cells involved in tissue repair and regeneration together with active participation in messaging to maintain neurological homeostasis. These comprehensive summaries of research findings are shown in Figure 1.
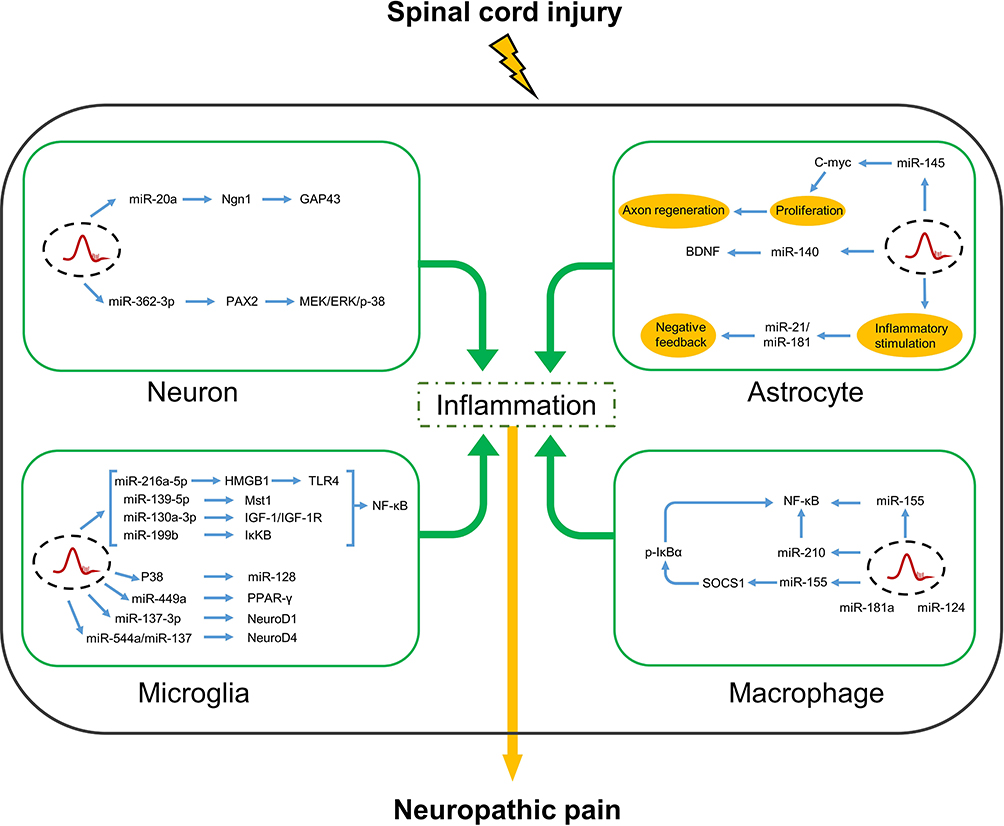 |
Figure 1 In neurons, microglia, astrocytes and macrophages, miRNAs regulate inflammatory factors by acting on various target genes after spinal cord injury, thereby relieving or worsening neuropathic pain. Abbreviations: PAX2, pair box gene2; MEK, mitogen-activated protein kinase kinase; ERK, extracellular signal regulated kinases; NF-κB, nuclear factor kappa-B; p-38, p38 MAPK; MAPK, mitogen-activated protein kinases; GAP43, growth-associated protein 43; Ngn1, neurogenin-1; C-myc, V-Myc avian myelocytomatosis viral oncogene homolog; BDNF, brain-derived neurotrophic factor; PPAR-γ, peroxisome proliferator-activated receptor γ; IκKβ, inhibitor of kappa B kinase β; NeuroD1, neuronal differentiation 1; NeuroD4, neuronal differentiation 4; Mst1, macrophage stimulating 1; HMGB1, high mobility group protein 1; TLR4, toll-like receptor 4; p-IκBα, phosphorylated NF-kappa-B inhibitor alpha; SOCS1, suppressor of cytokine signaling1. |
Roles of miRNAs in Neuron-Mediated Inflammation
NP occurs after nerve injury and the noxious reaction of neurons may persist long after tissue repair, suggesting a possible association with altered neuronal function.51 Neuronal responses to SCI are mediated by miR-20a via multiple pathways. Injection of miR20a into the spinal cord caused abnormal expression of miR20a and induced secondary injury, targeting Ngn-1 together with downregulated regeneration-related functional gene GAP43, upregulating IL-6, TNF-a, COX-2, with invasive inflammation of the spinal cord after 2 days and influenced the activation of endogenous neural cell regeneration, whereas the miR-20a-PDZ-Rhogef/RhoA/GAP43 axis promoted recovery of superior sensory function and inhibited the formation of lesions of the dorsal spinal cord via miR-20a modulation in spinal cord dorsal column injury.52,53 Subsequent studies revealed the involvement of miR-362-3p in the regulation of brain-derived neurotrophic factors by targeting PAX2, where overexpression of miR-362-3p downregulated expression of IL-1β, TNF-α, COX-2, and IL-6, which suppressed neuronal inflammation, thereby treating SCI-related NP.54 However, further studies are needed to explore the functional mechanisms of these various molecules as potential therapeutic targets for SCI-related inflammation and NP.
Roles of miRNAs in Microglia Mediated Inflammation
Microglia and astrocytes of the central nervous system strongly enhance neuronal excitability in response to injury.55 Microglia are quickly activated in response to harmful stimuli and directly express specific neurotransmitter receptors associated with NP and subsequently activate intracellular signaling pathways that promote the release of pro-inflammatory factors, such as TNF-α, IL-1β/6, and nitric oxide, which play key roles in the induction of NP.56–58 Therefore, it is important to clarify the molecular mechanisms underlying microglia-induced inflammation for treatment of SCI-related NP.
Many miRNAs that influence microglia-mediated inflammation after SCI participate in the regulation of downstream target genes. A previous study reported a correlation between TUSC7 and miR-449a expression in a rat model of SCI, where overexpression of TUSC7 inhibited miR-449a, while upregulation of PPAR-γ inhibited microglia activation and decreased expression of the pro-inflammatory factors TNF-a and IL-1β in the regulation of NP.59 Moreover, upregulation of miR-137-3p directly reduced expression of TNF-a and IL-1β via negative regulation of NeuroD1 in lipopolysaccharide (LPS)-treated BV2 cells (Gao et al, 2019).60 Both miR-137 and miR-544a can also target NeuroD4 and down-regulate pro-inflammatory cytokines to reduce inflammation caused by spinal cord injury. Activation of the P38 signaling pathway, following the downregulation of miRNA-128 promotes the transformation of microglia from M1 to M2 which is involved in NP and improves spinal cord injury.61 Collectively, these findings suggest that miRNAs can significantly ameliorate SCI-related inflammation and NP.62,63
It is well known that NF-κB is closely related to immunity as a transcription factor, and its mediated signaling pathway is considered to be the regulator of cell homeostasis. The signaling pathway is divided into three parts: (1) ligand and receptor binding; (2) receptor-related signaling; (3) Transcriptional activation and biological function of downstream genes. Three categories of protein families in the NF-κB pathway are the NF-κB transcription factor family (mainly divided into p50 (NF-κB1), p52 (NF-κB2), RelA (p65), c-Rel, and RelB), IκB family – an inhibitor of NF-κB transcription activity, and IκKB family – an inhibitor of IκB protein activity eliminate the inhibition of NF-κB. Under normal conditions, the inhibitory protein IκB forms a complex with NF-κB that inactivates NF-κB.64
Interestingly, NF-κB-dependent inflammation is reported to cause neuropathic nociceptive hyperalgesia in a mouse model of peripheral nerve injury, in response to SCI, inflammatory cytokines, and other antigens bind to receptors, and the NF-κB pathway is activated and involved in the immunomodulatory response caused by IκB inhibited by IκKB kinase.65 However, after the administration of IκKB inhibitors, the NF-κB pathway was inhibited, which negatively secrete pro-inflammatory factors IL-16, CCL2, TNF-α, MCP-1, and narrow NI to help prevent the development of chronic nerve pain.66 However, miRNAs can also modulate the inflammatory response by inhibiting activation of the NF-κB signaling pathway. MiR-199b is reported to mediate activation of the IκKβ-NF-κB signaling pathway in microglia, which attenuated inflammation in response to acute SCI, while overexpression of miR-199b reversed the upregulation of IκKβ resulting in the activity of p-p65 was inhibited and affected the microglia biological functions. MiR-216a-5p is reported to alleviate inflammation-related behaviors by inhibition of microglia-mediated inflammation via the HMGB1-TLR4-NF-κB pathway. In addition, the miRNAs mediated NF-κB signaling pathway plays a role in NP after spinal cord injury by regulating inflammatory responses. Upregulation of miR-216a-5p expression inhibits the activity of the p-65 transcription factor by targeting HMGB1 to bind to TLR4, thereby blocking the signaling pathway to reduce the inflammatory response and improve inflammatory pain. Thus, miR-216a-5p was proposed as a potential therapeutic target for SCI-related NP.67,68 A previous study demonstrated that miR-139-5P reduced expression levels of TNF-α and IL-1β by targeting Mst1, which inhibited NF-κB-related inflammatory responses and reduced nociceptive hypersensitivity, while downregulation of miR-130a-3p inhibited IGF-1 expression and reduced microglia activation by blocking NF-κB phosphorylation, leading to decreased expression of IL-1β, IL-6, and TNF-α, which significantly improved NP.69,70 These results confirm the important roles of miRNAs in NF-κB-mediated inflammation in response to SCI-NP.
Roles of miRNAs in Astrocyte-Mediated Inflammation
Astrocytes adopt either a pro-inflammatory or anti-inflammatory phenotype.71 Under pathological conditions, activated astrocytes adopt an anti-inflammatory phenotype to protect the central nervous system against NI.72,73
For days to weeks after spinal cord injury (SCI), the proliferation and hypertrophy of astrocytes may be involved in the lesion site, inducing glial scar formation. In response to SCI, miR-146a expression is upregulated in spinal cord astrocytes. Although miR-146a was highly expressed in spinal cord astrocytes after spinal cord injury, it has anti-inflammatory properties and plays a part in inducing glial scar hyperplasia.74 An in vitro model of LPS-induced glial cell injury revealed that MiR-140 not only inhibited expression of BDNF but also downregulated expression of IL-6 and TGF-α by binding to the 3’-untranslated region of BDNF.75
In a mouse model of LPS-induced inflammation, downregulation of miR-181 enhanced astrocyte production of the pro-inflammatory cytokines TNF-α, IL-6, IL-1β, and IL-8, while overexpression of miR-181 significantly increased expression of the anti-inflammatory cytokine IL-10.76 An in vitro study proposed GFAP and c-Myc as potential targets of miR-145 in astrocytes and found that miR-145 expression was downregulated in astrocytes in response to LPS-induced inflammation. However, overexpression of astrocyte-specific miR-145 decreased the density, size, and number of protrusions of astrocytes, suggesting that the inflammatory signaling pathways activated by SCI may promote the proliferation of astrocytes via inhibition of miR-145 expression.77 In rat models of chronic compression injury and spinal nerve ligation, NP was associated with miR-21 and miR-21 expression was upregulated in neurons and astrocytes in the chronic phase of SCI, especially affecting astroglial cell proliferation to form scarring.78–80 The regulatory sequence of miR-21 contains a highly conserved 300-bp region consisting of two STAT3-binding sites.81 Ablation of STAT3 limited early intrapathological hypertrophy of astrocytes and increased inflammatory cell invasion in response to SCI.82 Nonetheless, further studies are warranted to confirm the link between miR-21 and SCI-related NP.83
Roles of miRNAs in Macrophage-Mediated Inflammation
Macrophages from the peripheral circulation and resident microglia of the central nervous system are among the major effector cells that participate in SCI-related inflammation.84 Macrophages play an important role in the early inflammatory response via production of various pro-inflammatory cytokines and chemokines, which activate specific signaling pathways that trigger the production of other cytokines to further recruit more macrophages, ultimately leading to NP and disease progression.85
In a rat model of SCI, miR-124 reduced inflammation of the CNS by inhibiting the recruitment of macrophages and monocytes.86 In macrophages, miR-181a can directly downregulate IL-1α levels to regulate the inflammatory response.87 Since miR-181 is an important anti-inflammatory factor, reduced expression of miR-181 could increase SCI-related inflammation.74 Moreover, as a negative feedback regulator of an inflammatory response, miR-210 targeting NF-κB1 reduces inflammatory levels by inhibiting IκKB kinase activity, which is manifested by decreased expression of pro-inflammatory cytokines, including IL-6 and TNF-α, in endotoxin-induced macrophages.88 AgomiR-210 is a chemically modified miRNA agonist that is reported to mediate inflammation in a rat model of SCI. Together, these findings suggest the importance of the regulatory role of miR-210 in SCI-related NI.89
Human miR-155 has been implicated in stem cell differentiation, immunity, inflammation, cancer, and other pathophysiological processes.90 The use of a chronic compression injury model of NP demonstrated that inhibition of miR-155 significantly reduced mechanical and thermal hypernociception and inhibited macrophage production of pro-inflammatory cytokines and NF-κB and p38 MAPK activity via SOCS1.91 Thus, miR-155 presents a potential therapeutic target for SCI-related NP, although further studies are needed to clarify the underlying mechanism.92
Inflammation‑Linked lncRNAs in SCI-Related NP
Recent studies have revealed that lncRNAs are involved in various physiological and pathological processes, including adipogenesis, bone development, tumor formation, diabetes, and neurological diseases.93–97 The biological functions of lncRNAs mainly include chromatin regulation, transcription, and translation. Although initially considered untranslatable, lncRNAs contain open reading frame sequences with translational functions.98 The development of microarray and sequencing technologies has allowed investigations of the roles of lncRNAs in SCI-related inflammation and NP.
A mouse model of SCI showed that lncRNA Neat1 was regulated by miR-124 and promoted neuronal differentiation and migration and inhibited apoptosis via the Wnt/β-catenin signaling pathway, thus contributing to recovery from SCI.99 Interestingly, Neat1 increased expression of the inflammatory factors IL-6 and CXCL10, enhanced inflammasome activation, and promoted cellular scorching in vivo.100,101 In addition, Neat1 is reported to directly target the miR-128-3p/AQP4 axis to modulate SCI-related inflammation and NP.102 Meanwhile, knockdown of NEAT1 inhibited expression of IL-6, IL-1β, TNF-α, and AQP4, whereas inhibition of miR-381 restored expression levels. In a rat model of SCI, lncRNA PVT1 was reported upregulated and acted as a competing endogenous RNA that suppressed expression of miR-186-5p, thereby increasing CXCL13/CXCR5 expression and NP.103 Surprisingly, the novel lncRNA PKIA-AS1 enhanced promoter activity of CDK6 by reducing DNMT1-catalyzed methylation.104 In addition, lncRNA PKIA-AS1 has been identified as a key regulator of nerve injury-induced NP.
Small nucleolar RNA host genes (SNHGs) are a recently discovered family of lncRNAs that are closely related to tumor formation. The lncRNAs SNHG6 and SNHG17 can promote the development and metastasis of colorectal cancer.105,106 SNHG family members also play important roles in SCI-related inflammation and NP. Inhibition of KLF4, which is upregulated in SCI, reduces expression of pro-inflammatory cytokines and inhibits activation of microglia.107,108 KLF4 was also shown to directly target lncRNA SNHG5, which promotes progression of SCI by increasing activities of astrocytes and microglia.109,110
In a rat model of SCI, upregulation of lncRNA SNHG1 influenced the degree of NP via regulation of CDK4.111 Meanwhile, lncRNA SNHG12 has been associated with SCI-related inflammation and NP. In a rat model of spare nerve injury, silencing of lncRNA SNHG12, which was upregulated in the dorsal root ganglia, attenuated NP and reduced expression of the inflammatory factors IL-1β, IL-6, and TNF-α via upregulation of miR-494-3p.112
Inflammation‑Linked circRNAs in SCI-Related NP
Among the known ncRNAs, circRNAs have been widely studied in various cancers, cardiovascular diseases, and diseases of the central nervous system, where these molecules mainly function as sponges for miRNAs and RNA-binding proteins, and can drive translation through internal ribosomal entry sites and N6-methyladenosine sites.113 However, further studies are needed to clarify the differential expression of circRNAs before and after SCI.
Numerous studies have shown that circRNAs play crucial roles in SCI-related NP. For instance, circPrkcsh is reportedly upregulated in astrocytes and microglia in response to SCI and acts as a sponge to competitively inhibit expression of miR-488.114,115 Knockdown of circPrkcsh can reduce CCL2 expression in astrocytes and regulate the MEKK1/JNK/p38 MAPK pathway via upregulation of miR-488 in microglia, thereby improving SCI-related inflammation. KLF4 was identified as a key regulator of SI-related NI. In a mouse model of traumatic SCI at T8–10, miR-135b-5p was significantly downregulated.116 KLF4 and circAbca1 have been identified as targets of miR-135b-5p and the circAbca1/miR-135b-5p/KLF4 axis was found to regulate progression of traumatic SCI. Both circ0001723 and circ003564 were closely associated with NLRP3 expression after SCI.117,118 Inhibition of the NLRP3 inflammasome is reported to reduce SCI-related NI and mitochondrial dysfunction, thereby limiting the extent of SCI.119 In a mouse model of SCI, circ-Usp10 was found to regulate activation of microglia via the miR-152-5p/CD84 axis.120 Similarly, exosome-mediated circZFHX3 enhanced microglia viability and subsequently exacerbated inflammation-induced secondary injury.121
The NF-κB signaling pathway has been implicated in the pathology of NI, neural regeneration, cellular scorching, and disruption of the blood-spinal cord barrier after SCI. In addition, circ0000962, circ014301, circHecw1, and circ-Ncam2 were shown to regulate SCI-related NI via the NF-κB pathway. Expression of circ0000962 is downregulated after SCI and overexpression of circ0000962 was shown to reduce expression of TNF-α, IL-1β, IL-6, and IL-18 via downregulation of miR-302b-3p and regulation by the PI3K/Akt/NF-κB signaling pathway.122 An in vitro model of SCI based on LPS-treated PC12 cells found that circHecw1 regulated expression of inflammatory factors via miR-3551-3p and NF-κB.123 Meanwhile, circRNA_014301 was found to regulate inflammation and apoptosis of PC-12 cells by upregulation of p-NF-κB/NF-κB, Bax, and cleaved caspase-3, and downregulation of Bcl-2.124 Silencing of circ-Ncam2 induced miR-544-3p expression, which inhibited activation of the TLR4/NF-κB pathway, reduced activation of LPS-treated microglia, and inhibited neuronal apoptosis, thereby promoting recovery from SCI.125
In a rat model of diabetes, circHIPK3 negatively regulated miR-124 expression and knockdown of circHIPK3 attenuated NI-related NP.126 On day 7 post SCI of rats, circRNA-2960 was significantly upregulated in tissues around the surgical site.127 Interestingly, circRNA-2960 was found to target and downregulate miR-124, which exacerbated the inflammatory response, similar to diabetes-induced NP, suggesting that the circRNA-2960/miR-124 complex is a potential target for treatment of SCI-related NP. A comprehensive summary of these findings is shown in Figure 2.
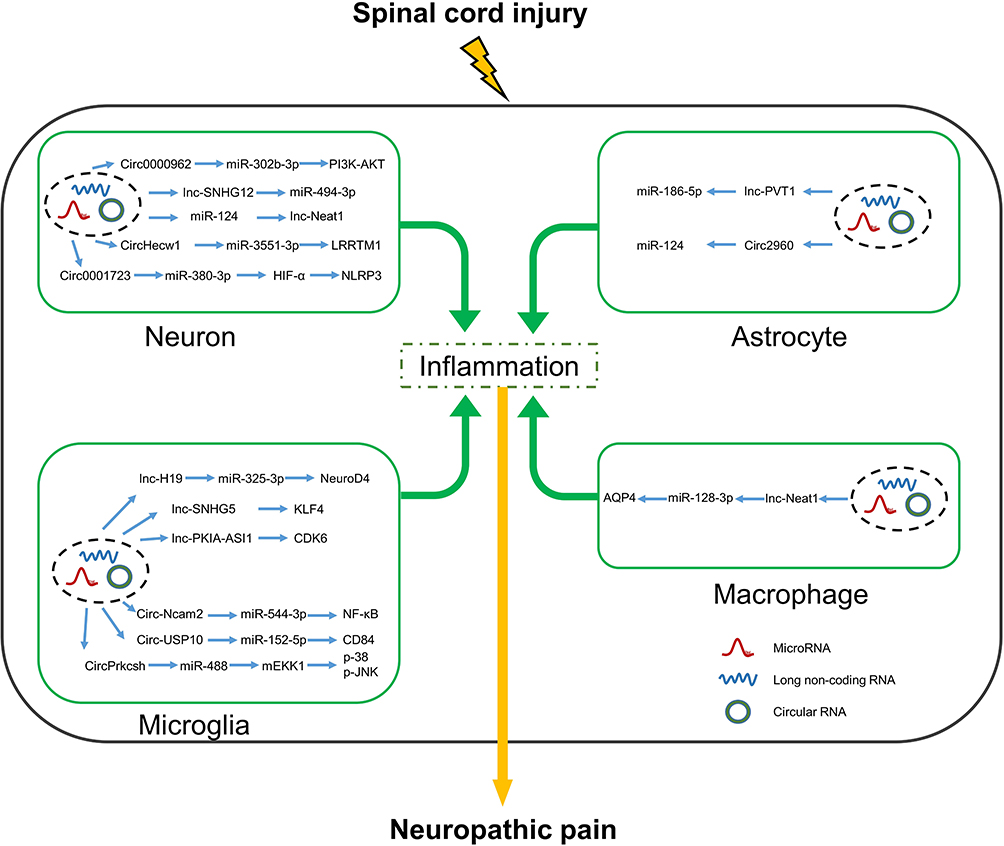 |
Figure 2 Interactions between lncRNAs/CircRNAs and miRNAs in neurons, microglia, astrocytes and macrophages alter the expression of inflammatory factors that affect neuropathic pain after spinal cord injury. Abbreviations: HIF-α, hypoxia-inducible factor-1α; PI3K, phosphoinositide 3-Kinase; AKT, protein kinase B; LRRTM1, leucine-rich repeat transmembrane neuronal protein 1; NLRP3, nod-like receptor thermal protein domain associated protein 3; CDK6, cyclin-dependent kinase 6; KLF4, kruppel-like factor 4; mEKK1, MAPK/ERK kinase kinase 1; p-38, p38 MAPK; MAPK, mitogen activated protein kinases; p-JNK, jun n terminal kinase phosphorylated; NeuroD4, neuronal differentiation 4; CD84 (SLAMF5), signaling lymphocytic Activation Molecule Family 5; AQP4, aquaporin-4. |
Although there are not many reports of circRNAs mediating SCI-related inflammation and NP via the NF-κB pathway, several studies have confirmed that the NF-κB pathway is closely associated with inflammation and NP following SCI. For example, combination therapy with minocycline and botulinum toxin can reduce SCI-induced NP and inflammation by activation of SIRT1 and inhibition of pAKT, P53, and p-NF-κB.128 SAFit2, an inhibitor of FKBP51, inhibits activation of the NF-κB pathway and reduces expression of inflammatory factors in the dorsal root ganglia and spinal cord to improve NP caused by nerve injury.129
Treatments
SCI-induced activation of the immune system is involved in tissue damage and repair, and activation of the inflammatory responses affects the prognosis of SCI-related NP. Recent studies have found that various miRNAs play essential roles in SCI-related inflammation and NP. Thus, the therapeutic roles of miRNAs have received much attention. Delivery of miRNA-124a via a chitosan multimeric system reduced microglia activation and TNF-α in vitro in a rat model of SCI, thereby providing a promising therapeutic approach for delivery of miRNAs.86
Several neuroprotective agents have been tested for treatment of SCI, including curcumin.60,130 A recent study of BV2 cells treated with curcumin found that LPS-induced inflammation was reduced via upregulation of miR-137-3p and downregulation of NeuroD1.60 In an in vivo study of SCI-related NP, miR-362-3p increased expression of TNF-α and IL-6 in neurons. Clinical data showed that SCI-NP is highly associated with TNF-α as a potential diagnostic biomarker in SCI patients. Recently, a TNF-α lentiviral shRNA vector relieved NP via TNF-α inhibition of downstream IL-6 expression, as a novel therapeutic option for NP.49,131
Perspectives and Conclusion
NP is a serious complication of SCI that negatively impacts quality of life and prognosis. Therefore, ncRNAs continue to attract attention for SCI-related NP. Our understanding of NP induced by spinal cord injury-mediated NI continues to advance. Researchers have tried a number of solutions in order to deliver ncRNAs into cells. Cui et al and Jee et al attempted to use intramyelin lentivirus injection in animal models, and Qi et al mostly used cell transfection to regulate the expression of ncRNAs in cell models.53,86,99 Louw et al found a cell selector-specific chitosan complex vector, miR-124 can be injected more stably in vivo and in vitro, while lncRNA and circRNA have not been found to have such more stable delivery system, and special materials such as nano-encapsulated materials can be considered as carriers.
Immune cells take on different roles during different phases of SCI, neurons respond to spinal cord injury, microglia are activated, and astrocyte proliferation promotes the repair of injured lesion sites but may also affect axon repair due to hyperplasia of glial scars, which is important to consider the duration of effect.80 Most commercially available reagents are now not cell type-specific, ensuring that targeted delivery of ncRNAs into cells and tissues to affect disease states without interfering with gene expression in other cells warrants consideration.
At present, gene therapy is gradually used in the clinical practice of various diseases. Recombinant adeno-associated virus (rAAV) is commonly used as a stable vector for gene delivery and is non-toxic and cannot be integrated into the human genome. Gene therapy would be a promising treatment approach, which can be used as gene vectors for ncRNAs to solve SCI-NP via NI, by silencing and increasing the regulation of pro-inflammatory and anti-inflammatory mediators. Further in-depth studies are warranted to explore the clinical application of ncRNAs for the treatment of SCI-related NP.
This review summarizes the mechanistic pathways mediated by non‐coding RNAs, NI, and NP, indicating that the regulation of NI by non-coding RNAs is a potential target and direction for the treatment of secondary neuropathic pain after spinal cord injury in the future.
Materials and Methods
We searched the online databases of NCBI Medical Database (PubMed), Web of Science, SpringerLink, Elsevier (ScienceDirect), OVID series of Medical Databases, and Open Access Library Databases for all valuable literature by manual search. To better control quality, articles published in all journals are subjected to a rigorous screening process. Keywords include non-coding RNAs, neuroinflammation, neuropathic pain, and spinal cord injury, and we searched highly relevant articles for a detailed research strategy. Conventional searches were supplemented by manual searches of all the relevant studies, review articles, and conference abstracts to avoid losing papers that might have been missed. We have no restrictions on the year and country of publication in our search.
Acknowledgments
This work was supported by the Health Commission of Jiangsu Province, China (M2022052), and the National Natural Science Foundation of China (No.31970938).
Disclosure
The authors declare that they have no conflicts of interest.
References
1. Anjum A, Yazid MD, Fauzi Daud M, et al. Spinal cord injury: pathophysiology, multimolecular interactions, and underlying recovery mechanisms. Int J Mol Sci. 2020;21(20):7533. doi:10.3390/ijms21207533
2. Finnerup NB. Neuropathic pain and spasticity: intricate consequences of spinal cord injury. Spinal Cord. 2017;55(12):1046–1050. doi:10.1038/sc.2017.70
3. Jackson CM, Choi J, Lim M. Mechanisms of immunotherapy resistance: lessons from glioblastoma. Nat Immunol. 2019;20(9):1100–1109. doi:10.1038/s41590-019-0433-y
4. Burchiel KJ, Hsu FP. Pain and spasticity after spinal cord injury: mechanisms and treatment. Spine. 2001;26(24 Suppl):S146–60. doi:10.1097/00007632-200112151-00024
5. Burke D, Fullen BM, Stokes D, Lennon O. Neuropathic pain prevalence following spinal cord injury: a systematic review and meta-analysis. Eur J Pain. 2017;21(1):29–44. doi:10.1002/ejp.905
6. Widerstrom-Noga EG, Turk DC. Types and effectiveness of treatments used by people with chronic pain associated with spinal cord injuries: influence of pain and psychosocial characteristics. Spinal Cord. 2003;41(11):600–609. doi:10.1038/sj.sc.3101511
7. Walters ET. Neuroinflammatory contributions to pain after SCI: roles for central glial mechanisms and nociceptor-mediated host defense. Exp Neurol. 2014;258:48–61. doi:10.1016/j.expneurol.2014.02.001
8. Brooks TA, Hawkins BT, Huber JD, Egleton RD, Davis TP. Chronic inflammatory pain leads to increased blood-brain barrier permeability and tight junction protein alterations. Am J Physiol Heart Circ Physiol. 2005;289(2):H738–43. doi:10.1152/ajpheart.01288.2004
9. Beggs S, Liu XJ, Kwan C, Salter MW. Peripheral nerve injury and TRPV1-expressing primary afferent C-fibers cause opening of the blood-brain barrier. Mol Pain. 2010;6:74. doi:10.1186/1744-8069-6-74
10. Haroon F, Drogemuller K, Handel U, et al. Gp130-dependent astrocytic survival is critical for the control of autoimmune central nervous system inflammation. J Immunol. 2011;186(11):6521–6531. doi:10.4049/jimmunol.1001135
11. Yoshizaki A, Miyagaki T, DiLillo DJ, et al. Regulatory B cells control T-cell autoimmunity through IL-21-dependent cognate interactions. Nature. 2012;491(7423):264–268. doi:10.1038/nature11501
12. Francos-Quijorna I, Amo-Aparicio J, Martinez-Muriana A, Lopez-Vales R. IL-4 drives microglia and macrophages toward a phenotype conducive for tissue repair and functional recovery after spinal cord injury. Glia. 2016;64(12):2079–2092. doi:10.1002/glia.23041
13. Neirinckx V, Coste C, Franzen R, Gothot A, Rogister B, Wislet S. Neutrophil contribution to spinal cord injury and repair. J Neuroinflammation. 2014;11:150. doi:10.1186/s12974-014-0150-2
14. Tsuda M. Microglia in the spinal cord and neuropathic pain. J Diabetes Investig. 2016;7(1):17–26. doi:10.1111/jdi.12379
15. Detloff MR, Fisher LC, McGaughy V, Longbrake EE, Popovich PG, Basso DM. Remote activation of microglia and pro-inflammatory cytokines predict the onset and severity of below-level neuropathic pain after spinal cord injury in rats. Exp Neurol. 2008;212(2):337–347. doi:10.1016/j.expneurol.2008.04.009
16. Gordh T, Chu H, Sharma HS. Spinal nerve lesion alters blood-spinal cord barrier function and activates astrocytes in the rat. Pain. 2006;124(1–2):211–221. doi:10.1016/j.pain.2006.05.020
17. Joyce A, DeLeo RPY. The role of neuroinflammation and neuroimmune activation in persistent pain. PAIN. 2001;90(2001):16. doi:10.1016/s0304-3959(00)00490-5
18. Watkins LR, Milligan ED, Maier SF. Glial activation: a driving force for pathological pain. Trends Neurosci. 2001;24(8):450–455. doi:10.1016/s0166-2236(00)01854-3
19. Xanthos DN, Sandkuhler J. Neurogenic neuroinflammation: inflammatory CNS reactions in response to neuronal activity. Nat Rev Neurosci. 2014;15(1):43–53. doi:10.1038/nrn3617
20. Hong P, Jiang M, Li H. Functional requirement of dicer1 and miR-17-5p in reactive astrocyte proliferation after spinal cord injury in the mouse. Glia. 2014;62(12):2044–2060. doi:10.1002/glia.22725
21. Jiang M, Wang Y, Wang J, Feng S, Wang X. The etiological roles of miRNAs, lncRNAs, and circRNAs in neuropathic pain: a narrative review. J Clin Lab Anal. 2022;36(8):e24592. doi:10.1002/jcla.24592
22. Ma X, Wang X, Ma X, et al. An update on the roles of circular RNAs in spinal cord injury. Mol Neurobiol. 2022;59(4):2620–2628. doi:10.1007/s12035-021-02721-2
23. Wang F, Liu J, Wang X, et al. The Emerging Role of lncRNAs in Spinal Cord Injury. Biomed Res Int. 2019;2019:3467121. doi:10.1155/2019/3467121
24. Karthikeyan A, Patnala R, Jadhav SP, Eng-Ang L, Dheen ST. MicroRNAs Key players in microglia and astrocyte mediated inflammation in CNS pathologies. Curr Med Chem. 2016;23:3528–3546. doi:10.2174/0929867323666160814001040
25. Burke D, Lennon O, Fullen BM. Quality of life after spinal cord injury: the impact of pain. Eur J Pain. 2018;22(9):1662–1672. doi:10.1002/ejp.1248
26. Crul TC, Post MWM, Visser-Meily JMA, Stolwijk-Swuste JM. Prevalence and Determinants of Pain in Spinal Cord Injury During Initial Inpatient Rehabilitation: data From the Dutch Spinal Cord Injury Database. Arch Phys Med Rehabil. 2022. doi:10.1016/j.apmr.2022.07.005
27. Felix ER, Cardenas DD, Bryce TN, et al. Prevalence and Impact of Neuropathic and Nonneuropathic Pain in Chronic Spinal Cord Injury. Arch Phys Med Rehabil. 2022;103(4):729–737. doi:10.1016/j.apmr.2021.06.022
28. Khazaeipour Z, Ahmadipour E, Rahimi-Movaghar V, Ahmadipour F, Vaccaro AR, Babakhani B. Association of pain, social support and socioeconomic indicators in patients with spinal cord injury in Iran. Spinal Cord. 2017;55(2):180–186. doi:10.1038/sc.2016.160
29. Mann R, Schaefer C, Sadosky A, et al. Burden of spinal cord injury-related neuropathic pain in the United States: retrospective chart review and cross-sectional survey. Spinal Cord. 2013;51(7):564–570. doi:10.1038/sc.2013.34
30. Muller R, Brinkhof MW, Arnet U, et al. Prevalence and associated factors of pain in the Swiss spinal cord injury population. Spinal Cord. 2017;55(4):346–354. doi:10.1038/sc.2016.157
31. Ravenscroft A, Ahmed YS, Burnside IG. Chronic pain after SCI. A patient survey. Spinal Cord. 2000;38(10):611–614. doi:10.1038/sj.sc.3101073
32. Siddall PJ, McClelland JM, Rutkowski SB, Cousins MJ. A longitudinal study of the prevalence and characteristics of pain in the first 5 years following spinal cord injury. Pain. 2003;103(3):249–257. doi:10.1016/s0304-3959(02)00452-9
33. Widerstrom-Noga EG, Felipe-Cuervo E, Yezierski RP. Chronic pain after spinal injury: interference with sleep and daily activities. Arch Phys Med Rehabil. 2001;82(11):1571–1577. doi:10.1053/apmr.2001.26068
34. Wollaars MM, Post MW, van Asbeck FW, Brand N. Spinal cord injury pain: the influence of psychological factors and impact on quality of life. Clin J Pain. 2007;23(5):383–391. doi:10.1097/AJP.0b013e31804463e5
35. Siddall PJ. Management of neuropathic pain following spinal cord injury: now and in the future. Spinal Cord. 2009;47(5):352–359. doi:10.1038/sc.2008.136
36. Stampacchia G, Gerini A, Morganti R, et al. Pain characteristics in Italian people with spinal cord injury: a multicentre study. Spinal Cord. 2022;60(7):604–611. doi:10.1038/s41393-021-00656-y
37. van Gorp S, Kessels AG, Joosten EA, van Kleef M, Patijn J. Pain prevalence and its determinants after spinal cord injury: a systematic review. Eur J Pain. 2015;19(1):5–14. doi:10.1002/ejp.522
38. Cardenas DD, Felix ER. Pain after spinal cord injury: a review of classification, treatment approaches, and treatment assessment. PM R. 2009;1(12):1077–1090. doi:10.1016/j.pmrj.2009.07.002
39. Masri R, Keller A. Chronic pain following spinal cord injury. Adv Exp Med Biol. 2012;760:74–88. doi:10.1007/978-1-4614-4090-1_5
40. Chen J, Weidner N, Puttagunta R. The Impact of Activity-Based Interventions on Neuropathic Pain in Experimental Spinal Cord Injury. Cells. 2022;11(19). doi:10.3390/cells11193087
41. Finnerup NB, Norrbrink C, Trok K, et al. Phenotypes and predictors of pain following traumatic spinal cord injury: a prospective study. J Pain. 2014;15(1):40–48. doi:10.1016/j.jpain.2013.09.008
42. Attal N. Spinal cord injury pain. Rev Neurol (Paris). 2021;177(5):606–612. doi:10.1016/j.neurol.2020.07.003
43. Nakipoglu-Yüzer GF, Nermin Atçı N, Ozgirgin N. Neuropathic pain in spinal cord injury. Pain Physician. 2013;16:259–264.
44. Rosner J, Negraeff M, Belanger LM, et al. Characterization of Hyperacute Neuropathic Pain after Spinal Cord Injury: a Prospective Study. J Pain. 2022;23(1):89–97. doi:10.1016/j.jpain.2021.06.013
45. Fakhri S, Abbaszadeh F, Jorjani M. On the therapeutic targets and pharmacological treatments for pain relief following spinal cord injury: a mechanistic review. Biomed Pharmacother. 2021;139:111563. doi:10.1016/j.biopha.2021.111563
46. Viswanath O, Urits I, Burns J, et al. Central Neuropathic Mechanisms in Pain Signaling Pathways: current Evidence and Recommendations. Adv Ther. 2020;37(5):1946–1959. doi:10.1007/s12325-020-01334-w
47. Karran EL, Fryer CE, Middleton JW, Moseley GL. Pain and pain management experiences following spinal cord injury - a mixed methods study of Australian community-dwelling adults. Disabil Rehabil. 2022;1–14. doi:10.1080/09638288.2022.2034994
48. Norrbrink Budh C, Lund I, Ertzgaard P, et al. Pain in a Swedish spinal cord injury population. Clin Rehabil. 2003;17(6):685–690. doi:10.1191/0269215503cr664oa
49. Xu J, X E, Liu H, et al. Tumor necrosis factor-alpha is a potential diagnostic biomarker for chronic neuropathic pain after spinal cord injury. Neurosci Lett. 2015;595:30–34. doi:10.1016/j.neulet.2015.04.004
50. Yang J, Xiong LL, Wang YC, et al. Oligodendrocyte precursor cell transplantation promotes functional recovery following contusive spinal cord injury in rats and is associated with altered microRNA expression. Mol Med Rep. 2018;17(1):771–782. doi:10.3892/mmr.2017.7957
51. Hiraga SI, Itokazu T, Nishibe M, Yamashita T. Neuroplasticity related to chronic pain and its modulation by microglia. Inflamm Regen. 2022;42(1):15. doi:10.1186/s41232-022-00199-6
52. Wang T, Li B, Yuan X, et al. MiR-20a Plays a Key Regulatory Role in the Repair of Spinal Cord Dorsal Column Lesion via PDZ-RhoGEF/RhoA/GAP43 Axis in Rat. Cell Mol Neurobiol. 2019;39(1):87–98. doi:10.1007/s10571-018-0635-0
53. Jee MK, Jung JS, Im YB, Jung SJ, Kang SK. Silencing of miR20a is crucial for Ngn1-mediated neuroprotection in injured spinal cord. Hum Gene Ther. 2012;23(5):508–520. doi:10.1089/hum.2011.121
54. Hu Y, Liu Q, Zhang M, Yan Y, Yu H, Ge L. MicroRNA-362-3p attenuates motor deficit following spinal cord injury via targeting paired box gene 2. J Integr Neurosci. 2019;18(1):57–64. doi:10.31083/j.jin.2019.01.12
55. Grace PM, Hutchinson MR, Maier SF, Watkins LR. Pathological pain and the neuroimmune interface. Nat Rev Immunol. 2014;14(4):217–231. doi:10.1038/nri3621
56. Kang SY, Jung HW, Lee MY, Lee HW, Chae SW, Park YK. Effect of the semen extract of Cuscuta chinensis on inflammatory responses in LPS-stimulated BV-2 microglia. Chin J Nat Med. 2014;12(8):573–581. doi:10.1016/S1875-5364(14)60088-1
57. Zychowska M, Rojewska E, Makuch W, Przewlocka B, Mika J. The influence of microglia activation on the efficacy of amitriptyline, doxepin, milnacipran, venlafaxine and fluoxetine in a rat model of neuropathic pain. Eur J Pharmacol. 2015;749:115–123. doi:10.1016/j.ejphar.2014.11.022
58. Ji A, Xu J. Neuropathic Pain: biomolecular Intervention and Imaging via Targeting Microglia Activation. Biomolecules. 2021;11(9). doi:10.3390/biom11091343
59. Yu Y, Zhu M, Zhao Y, Xu M, Qiu M. Overexpression of TUSC7 inhibits the inflammation caused by microglia activation via regulating miR-449a/PPAR-gamma. Biochem Biophys Res Commun. 2018;503(2):1020–1026. doi:10.1016/j.bbrc.2018.06.111
60. Gao F, Lei J, Zhang Z, Yang Y, You H. Curcumin alleviates LPS-induced inflammation and oxidative stress in mouse microglial BV2 cells by targeting miR-137-3p/NeuroD1. RSC Adv. 2019;9(66):38397–38406. doi:10.1039/c9ra07266g
61. Yang Z, Xu J, Zhu R, Liu L. Down-Regulation of miRNA-128 Contributes to Neuropathic Pain Following Spinal Cord Injury via Activation of P38. Med Sci Monit. 2017;23:405–411. doi:10.12659/msm.898788
62. Yang L, Ge D, Chen X, Jiang C, Zheng S. miRNA-544a Regulates the Inflammation of Spinal Cord Injury by Inhibiting the Expression of NEUROD4. Cell Physiol Biochem. 2018;51(4):1921–1931. doi:10.1159/000495717
63. Dai J, Xu L-J, Han G-D, et al. MiR-137 attenuates spinal cord injury by modulating NEUROD4 through reducing inflammation and oxidative stress. Eur Rev Med Pharmacol Sci. 2018;22:1884–1890. doi:10.26355/eurrev_201804_14709
64. Yu H, Lin L, Zhang Z, Zhang H, Hu H. Targeting NF-kappaB pathway for the therapy of diseases: mechanism and clinical study. Signal Transduct Target Ther. 2020;5(1):209. doi:10.1038/s41392-020-00312-6
65. Grilli M, Memo M. Nuclear factor-kappaB/Rel proteins: a point of convergence of signalling pathways relevant in neuronal function and dysfunction. Biochem Pharmacol. 1999;57:1–7. doi:10.1016/s0006-2952(98)00214-7
66. Kanngiesser M, Haussler A, Myrczek T, et al. Inhibitor kappa B kinase beta dependent cytokine upregulation in nociceptive neurons contributes to nociceptive hypersensitivity after sciatic nerve injury. J Pain. 2012;13(5):485–497. doi:10.1016/j.jpain.2012.02.010
67. Zhou HJ, Wang LQ, Xu QS, et al. Downregulation of miR-199b promotes the acute spinal cord injury through IKKbeta-NF-kappaB signaling pathway activating microglial cells. Exp Cell Res. 2016;349(1):60–67. doi:10.1016/j.yexcr.2016.09.020
68. Zhenzhen Z, Fenghao L, Meina M, Rui L, Wenbo S, Qi W. Targeting HMGB1-TLR4 signaling by miR-216a-5p elevation alleviates the inflammatory behavioral hypersensitivity. Neurosci Lett. 2021;759:136043. doi:10.1016/j.neulet.2021.136043
69. Wang P, Zhang Y, Xia Y, et al. MicroRNA-139-5p Promotes Functional Recovery and Reduces Pain Hypersensitivity in Mice with Spinal Cord Injury by Targeting Mammalian Sterile 20-like Kinase 1. Neurochem Res. 2021;46(2):349–357. doi:10.1007/s11064-020-03170-4
70. Yao L, Guo Y, Wang L, et al. Knockdown of miR-130a-3p alleviates spinal cord injury induced neuropathic pain by activating IGF-1/IGF-1R pathway. J Neuroimmunol. 2021;351:577458. doi:10.1016/j.jneuroim.2020.577458
71. Chen M, Lai X, Wang X, et al. Long Non-coding RNAs and Circular RNAs: insights Into Microglia and Astrocyte Mediated Neurological Diseases. Front Mol Neurosci. 2021;14:745066. doi:10.3389/fnmol.2021.745066
72. Leng F, Edison P. Neuroinflammation and microglial activation in Alzheimer disease: where do we go from here? Nat Rev Neurol. 2021;17(3):157–172. doi:10.1038/s41582-020-00435-y
73. Ramirez AE, Gil-Jaramillo N, Tapias MA, et al. MicroRNA: a Linking between Astrocyte Dysfunction, Mild Cognitive Impairment, and Neurodegenerative Diseases. Life. 2022;12(9). doi:10.3390/life12091439
74. Martirosyan NL, Carotenuto A, Patel AA, et al. The Role of microRNA Markers in the Diagnosis, Treatment, and Outcome Prediction of Spinal Cord Injury. Front Surg. 2016;3:56. doi:10.3389/fsurg.2016.00056
75. Tu Z, Li Y, Dai Y, et al. MiR-140/BDNF axis regulates normal human astrocyte proliferation and LPS-induced IL-6 and TNF-alpha secretion. Biomed Pharmacother. 2017;91:899–905. doi:10.1016/j.biopha.2017.05.016
76. Hutchison ER, Kawamoto EM, Taub DD, et al. Evidence for miR-181 involvement in neuroinflammatory responses of astrocytes. Glia. 2013;61(7):1018–1028. doi:10.1002/glia.22483
77. Wang CY, Yang SH, Tzeng SF. MicroRNA-145 as one negative regulator of astrogliosis. Glia. 2015;63(2):194–205. doi:10.1002/glia.22743
78. Bhalala OG, Srikanth M, Kessler JA. The emerging roles of microRNAs in CNS injuries. Nat Rev Neurol. 2013;9(6):328–339. doi:10.1038/nrneurol.2013.67
79. Sakai A, Suzuki H. Nerve injury-induced upregulation of miR-21 in the primary sensory neurons contributes to neuropathic pain in rats. Biochem Biophys Res Commun. 2013;435(2):176–181. doi:10.1016/j.bbrc.2013.04.089
80. Bhalala OG, Pan L, Sahni V, et al. microRNA-21 regulates astrocytic response following spinal cord injury. J Neurosci. 2012;32(50):17935–17947. doi:10.1523/JNEUROSCI.3860-12.2012
81. Loffler D, Brocke-Heidrich K, Pfeifer G, et al. Interleukin-6 dependent survival of multiple myeloma cells involves the Stat3-mediated induction of microRNA-21 through a highly conserved enhancer. Blood. 2007;110(4):1330–1333. doi:10.1182/blood-2007-03-081133
82. Herrmann JE, Imura T, Song B, et al. STAT3 is a critical regulator of astrogliosis and scar formation after spinal cord injury. J Neurosci. 2008;28(28):7231–7243. doi:10.1523/JNEUROSCI.1709-08.2008
83. Sakai A, Suzuki H. Emerging roles of microRNAs in chronic pain. Neurochem Int. 2014;77:58–67. doi:10.1016/j.neuint.2014.05.010
84. David S, Kroner A. Repertoire of microglial and macrophage responses after spinal cord injury. Nat Rev Neurosci. 2011;12(7):388–399. doi:10.1038/nrn3053
85. Kuppa SS, Kim HK, Kang JY, Lee SC, Seon JK. Role of Mesenchymal Stem Cells and Their Paracrine Mediators in Macrophage Polarization: an Approach to Reduce Inflammation in Osteoarthritis. Int J Mol Sci. 2022;23(21). doi:10.3390/ijms232113016
86. Louw AM, Kolar MK, Novikova LN, et al. Chitosan polyplex mediated delivery of miRNA-124 reduces activation of microglial cells in vitro and in rat models of spinal cord injury. Nanomedicine. 2016;12(3):643–653. doi:10.1016/j.nano.2015.10.011
87. Xie W, Li M, Xu N, et al. MiR-181a regulates inflammation responses in monocytes and macrophages. PLoS One. 2013;8(3):e58639. doi:10.1371/journal.pone.0058639
88. Qi J, Qiao Y, Wang P, Li S, Zhao W, Gao C. microRNA-210 negatively regulates LPS-induced production of proinflammatory cytokines by targeting NF-kappaB1 in murine macrophages. FEBS Lett. 2012;586(8):1201–1207. doi:10.1016/j.febslet.2012.03.011
89. Cao Y, Wu TD, Wu H, et al. Synchrotron radiation micro-CT as a novel tool to evaluate the effect of agomir-210 in a rat spinal cord injury model. Brain Res. 2017;1655:55–65. doi:10.1016/j.brainres.2016.11.015
90. Zheng SR, Guo GL, Zhang W, et al. Clinical significance of miR-155 expression in breast cancer and effects of miR-155 ASO on cell viability and apoptosis. Oncol Rep. 2012;27(4):1149–1155. doi:10.3892/or.2012.1634
91. Gaudet AD, Mandrekar-Colucci S, Hall JC, et al. miR-155 Deletion in Mice Overcomes Neuron-Intrinsic and Neuron-Extrinsic Barriers to Spinal Cord Repair. J Neurosci. 2016;36(32):8516–8532. doi:10.1523/JNEUROSCI.0735-16.2016
92. Tan Y, Yang J, Xiang K, Tan Q, Guo Q. Suppression of microRNA-155 attenuates neuropathic pain by regulating SOCS1 signalling pathway. Neurochem Res. 2015;40(3):550–560. doi:10.1007/s11064-014-1500-2
93. Riva P, Ratti A, Venturin M. The Long Non-Coding RNAs in Neurodegenerative Diseases: novel Mechanisms of Pathogenesis. Curr Alzheimer Res. 2016;13(11):1219–1231. doi:10.2174/1567205013666160622112234
94. Feng SD, Yang JH, Yao CH, et al. Potential regulatory mechanisms of lncRNA in diabetes and its complications. Biochem Cell Biol. 2017;95(3):361–367. doi:10.1139/bcb-2016-0110
95. Tao T, Wu S, Sun Z, et al. The molecular mechanisms of LncRNA-correlated PKM2 in cancer metabolism. Biosci Rep. 2019;39(11):754.
96. Li D, Yang C, Yin C, et al. LncRNA, Important Player in Bone Development and Disease. Endocr Metab Immune Disord Drug Targets. 2020;20(1):50–66. doi:10.2174/1871530319666190904161707
97. Zhang P, Wu S, He Y, et al. LncRNA-Mediated Adipogenesis in Different Adipocytes. Int J Mol Sci. 2022;23(13):78.
98. Kong S, Tao M, Shen X, Ju S. Translatable circRNAs and lncRNAs: driving mechanisms and functions of their translation products. Cancer Lett. 2020;483:59–65. doi:10.1016/j.canlet.2020.04.006
99. Cui Y, Yin Y, Xiao Z, et al. LncRNA Neat1 mediates miR-124-induced activation of Wnt/beta-catenin signaling in spinal cord neural progenitor cells. Stem Cell Res Ther. 2019;10(1):400. doi:10.1186/s13287-019-1487-3
100. Zhang F, Wu L, Qian J, et al. Identification of the long noncoding RNA NEAT1 as a novel inflammatory regulator acting through MAPK pathway in human lupus. J Autoimmun. 2016;75:96–104. doi:10.1016/j.jaut.2016.07.012
101. Zhang P, Cao L, Zhou R, Yang X, Wu M. The lncRNA Neat1 promotes activation of inflammasomes in macrophages. Nat Commun. 2019;10(1):1495. doi:10.1038/s41467-019-09482-6
102. Xian S, Ding R, Li M, Chen F. LncRNA NEAT1/miR-128-3p/AQP4 axis regulating spinal cord injury-induced neuropathic pain progression. J Neuroimmunol. 2021;351:577457. doi:10.1016/j.jneuroim.2020.577457
103. Zhang P, Sun H, Ji Z. Downregulating lncRNA PVT1 Relieves Astrocyte Overactivation Induced Neuropathic Pain Through Targeting miR-186-5p/CXCL13/CXCR5 Axis. Neurochem Res. 2021;46(6):1457–1469. doi:10.1007/s11064-021-03287-0
104. Hu J-Z, Rong Z-J, Li M, et al. Silencing of lncRNA PKIA-AS1 Attenuates Spinal Nerve Ligation-Induced Neuropathic Pain Through Epigenetic Downregulation of CDK6 Expression. Front Cell Neurosci. 2019;13:50. doi:10.3389/fncel.2019.00050
105. Shao Q, Xu J, Deng R, et al. SNHG 6 promotes the progression of Colon and Rectal adenocarcinoma via miR-101-3p and Wnt/β-catenin signaling pathway. BMC Gastroenterol. 2019;19(1):163. doi:10.1186/s12876-019-1080-3
106. Bian Z, Zhou M, Cui K, et al. SNHG17 promotes colorectal tumorigenesis and metastasis via regulating Trim23-PES1 axis and miR-339-5p-FOSL2-SNHG17 positive feedback loop. J Exp Clin Cancer Res. 2021;40(1):360. doi:10.1186/s13046-021-02162-8
107. Liu J, Zhang C, Liu Z, Zhang J, Xiang Z, Sun T. Honokiol downregulates Kruppel-like factor 4 expression, attenuates inflammation, and reduces histopathology after spinal cord injury in rats. Spine. 2015;40(6):363–368. doi:10.1097/BRS.0000000000000758
108. Kaushik DK, Gupta M, Das S, Basu A. Krüppel-like factor 4, a novel transcription factor regulates microglial activation and subsequent neuroinflammation. J Neuroinflammation. 2010;7(1):68. doi:10.1186/1742-2094-7-68
109. Zhao L, Han T, Li Y, et al. The lncRNA SNHG5/miR-32 axis regulates gastric cancer cell proliferation and migration by targeting KLF4. FASEB J. 2017;31(3):893–903. doi:10.1096/fj.201600994R
110. Jiang ZS, Zhang JR. LncRNA SNHG5 enhances astrocytes and microglia viability via upregulating KLF4 in spinal cord injury. Int J Biol Macromol. 2018;120(Pt A):66–72. doi:10.1016/j.ijbiomac.2018.08.002
111. Zhang JY, Lv DB, Su YN, et al. LncRNA SNHG1 attenuates neuropathic pain following spinal cord injury by regulating CDK4 level. Eur Rev Med Pharmacol Sci. 2020;24(23):12034–12040. doi:10.26355/eurrev_202012_23992
112. Zhang J, Zhao H, Zhang A, et al. Identifying a novel KLF2/lncRNA SNHG12/miR-494-3p/RAD23B axis in Spare Nerve Injury-induced neuropathic pain. Cell Death Discov. 2022;8(1):272. doi:10.1038/s41420-022-01060-y
113. Qu X, Li Z, Chen J, Hou L. The emerging roles of circular RNAs in CNS injuries. J Neurosci Res. 2020;98(7):1485–1497. doi:10.1002/jnr.24591
114. Chen JN, Zhang YN, Tian LG, Zhang Y, Li XY, Ning B. Down-regulating Circular RNA Prkcsh suppresses the inflammatory response after spinal cord injury. Neural Regen Res. 2022;17(1):144–151. doi:10.4103/1673-5374.314114
115. Li X, Kang J, Lv H, et al. CircPrkcsh, a circular RNA, contributes to the polarization of microglia towards the M1 phenotype induced by spinal cord injury and acts via the JNK/p38 MAPK pathway. FASEB J. 2021;35(12):e22014. doi:10.1096/fj.202100993R
116. Wang WZ, Li J, Liu L, et al. Role of circular RNA expression in the pathological progression after spinal cord injury. Neural Regen Res. 2021;16(10):2048–2055. doi:10.4103/1673-5374.308100
117. Li X, Lou X, Xu S, Du J, Wu J. Hypoxia inducible factor-1 (HIF-1alpha) reduced inflammation in spinal cord injury via miR-380-3p/ NLRP3 by Circ 0001723. Biol Res. 2020;53(1):35. doi:10.1186/s40659-020-00302-6
118. Zhao Y, Chen Y, Wang Z, et al. Bone Marrow Mesenchymal Stem Cell Exosome Attenuates Inflammasome-Related Pyroptosis via Delivering circ_003564 to Improve the Recovery of Spinal Cord Injury. Mol Neurobiol. 2022;59(11):6771–6789. doi:10.1007/s12035-022-03006-y
119. Jiang W, Li M, He F, Zhou S, Zhu L. Targeting the NLRP3 inflammasome to attenuate spinal cord injury in mice. J Neuroinflammation. 2017;14(1):207. doi:10.1186/s12974-017-0980-9
120. Tong D, Zhao Y, Tang Y, Ma J, Wang Z, Li C. Circ-Usp10 promotes microglial activation and induces neuronal death by targeting miRNA-152-5p/CD84. Bioengineered. 2021;12(2):10812–10822. doi:10.1080/21655979.2021.2004362
121. Tian F, Yang J, Xia R. Exosomes Secreted from circZFHX3-modified Mesenchymal Stem Cells Repaired Spinal Cord Injury Through mir-16-5p/IGF-1 in Mice. Neurochem Res. 2022;47(7):2076–2089. doi:10.1007/s11064-022-03607-y
122. He R, Tang GL, Niu L, et al. Quietness Circ 0000962 promoted nerve cell inflammation through PIK3CA/Akt/NF-kappaB signaling by miR-302b-3p in spinal cord injury. Ann Palliat Med. 2020;9(2):190–198. doi:10.21037/apm.2020.02.13
123. Ban D, Xiang Z, Yu P, Liu Y. Circular RNA Hecw1 Regulates the Inflammatory Imbalance in Spinal Cord Injury via miR-3551-3p/LRRTM1 Axis. Appl Biochem Biotechnol. 2022;194(11):5151–5166. doi:10.1007/s12010-022-03999-1
124. Xie X, Xiao Y, Xu K. Mechanism underlying circularRNA_014301-mediated regulation of neuronal cell inflammation and apoptosis. Exp Ther Med. 2021;22(6):1432. doi:10.3892/etm.2021.10867
125. Guo K, Chang Y, Jin Y, Yuan H, Che P. circ-Ncam2 (mmu_circ_0006413) Participates in LPS-Induced Microglia Activation and Neuronal Apoptosis via the TLR4/NF-kappaB Pathway. J Mol Neurosci. 2022;72(8):1738–1748. doi:10.1007/s12031-022-02018-6
126. Wang L, Luo T, Bao Z, Li Y, Bu W. Intrathecal circHIPK3 shRNA alleviates neuropathic pain in diabetic rats. Biochem Biophys Res Commun. 2018;505(3):644–650. doi:10.1016/j.bbrc.2018.09.158
127. Chen J, Fu B, Bao J, Su R, Zhao H, Liu Z. Novel circular RNA 2960 contributes to secondary damage of spinal cord injury by sponging miRNA-124. J Comp Neurol. 2021;529(7):1456–1464. doi:10.1002/cne.25030
128. Yu Z, Liu J, Sun L, Wang Y, Meng H. Combination of Botulinum Toxin and minocycline Ameliorates Neuropathic Pain Through Antioxidant Stress and Anti-Inflammation via Promoting SIRT1 Pathway. Front Pharmacol. 2020;11:602417. doi:10.3389/fphar.2020.602417
129. Wedel S, Mathoor P, Rauh O, et al. SAFit2 reduces neuroinflammation and ameliorates nerve injury-induced neuropathic pain. J Neuroinflammation. 2022;19(1):254. doi:10.1186/s12974-022-02615-7
130. Sanivarapu R, Vallabhaneni V, Verma V. The potential of curcumin in treatment of spinal cord injury. Neurol Res Int. 2016;2016:9468193. doi:10.1155/2016/9468193
131. Ogawa N, Kawai H, Terashima T, et al. Gene therapy for neuropathic pain by silencing of TNF-alpha expression with lentiviral vectors targeting the dorsal root ganglion in mice. PLoS One. 2014;9(3):e92073. doi:10.1371/journal.pone.0092073
 © 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Complement Receptor 3 Pathway and NMDA Receptor 2B Subunit Involve Neuropathic Pain Associated with Spinal Cord Injury
Li Y, Fang SC, Zhou L, Mo XM, Guo HD, Deng YB, Yu HH, Gong WY
Journal of Pain Research 2022, 15:1813-1823
Published Date: 25 June 2022
Dexmedetomidine Alleviates Neuropathic Pain via the TRPC6-p38 MAPK Pathway in the Dorsal Root Ganglia of Rats
Xu S, Yi Y, Wang Y, Wang P, Zhao Y, Feng W
Journal of Pain Research 2022, 15:2437-2448
Published Date: 19 August 2022
Cerium Oxide Nanoparticles Alleviate Neuropathic Pain by Modulating Macrophage Polarization in a Rat SCI Model
Ban D, Yu H, Xiang Z, Li C, Yu P, Wang J, Liu Y
Journal of Pain Research 2022, 15:3369-3380
Published Date: 25 October 2022
A Silver Lining of Neuropathic Pain: Predicting Favorable Functional Outcome in Spinal Cord Injury
Xu ML, Wu XB, Liang Y, Li N, Hu X, Lin XD, Sun MQ, Dai CQ, Niu D, Zhang YR, Cao H, Zhao CG, Sun XL, Yuan H
Journal of Pain Research 2023, 16:2619-2632
Published Date: 27 July 2023
Mesenchymal Stem Cell Transplantation: Neuroprotection and Nerve Regeneration After Spinal Cord Injury
Chen SY, Yang RL, Wu XC, Zhao DZ, Fu SP, Lin FQ, Li LY, Yu LM, Zhang Q, Zhang T
Journal of Inflammation Research 2023, 16:4763-4776
Published Date: 20 October 2023