")
Back to Journals » Drug Design, Development and Therapy » Volume 17
New Target for Minoxidil in the Treatment of Androgenetic Alopecia
Authors Shen Y, Zhu Y, Zhang L, Sun J , Xie B , Zhang H , Song X
Received 20 July 2023
Accepted for publication 19 August 2023
Published 24 August 2023 Volume 2023:17 Pages 2537—2547
DOI https://doi.org/10.2147/DDDT.S427612
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Prof. Dr. Tin Wui Wong
Yuqing Shen,1 Yuqi Zhu,1 Li Zhang,1 Jiayi Sun,1 Bo Xie,2 Hongyan Zhang,2 Xiuzu Song2
1Department of Dermatology, Hangzhou Third People’s Hospital, Zhejiang Chinese Medical University, Hangzhou, People’s Republic of China; 2Department of Dermatology, Hangzhou Third People’s Hospital, Affiliated Hangzhou Dermatology Hospital of Zhejiang University School of Medicine, Hangzhou, People’s Republic of China
Correspondence: Xiuzu Song, Hangzhou Third People’s Hospital, Hangzhou, People’s Republic of China, Email [email protected]
Objective: To investigate the mechanism of minoxidil in treating androgenetic alopecia (AGA).
Methods: The mechanism of action of minoxidil on AGA was first systematically investigated from the viewpoint of network pharmacology, including minoxidil-AGA target prediction, protein-protein interaction (PPI) network analysis, molecular docking and enrichment analysis of targets related to minoxidil and AGA, and dermal papilla cell assays to confirm the viability of prediction.
Results: The combined analysis revealed that minoxidil treatment of AGA not only acts on androgenic receptors (AR) but also on 2 new targets, steroid 17-alpha-hydroxylase/17,20 lyase (CYP17A1) and aromatase (CYP19A1). The biological processes linked to these targets were concentrated on several pathways, including enzymes and hormones. Further experiments have revealed that minoxidil suppresses the expression of AR and CYP17A1, boosts the activity of CYP19A1, decreases the formation and binding of dihydrotestosterone, and enhances the production of estradiol. Through these changes, minoxidil acts as a treatment for AGA.
Conclusion: Minoxidil may act by altering hormonal and enzymatic pathways. Our study finds two new targets (CYP17A1, CYP19A1) of minoxidil and demonstrates that minoxidil inhibits AR. These targets may provide new ideas for drug research.
Keywords: minoxidil, androgenetic alopecia, targets
Background
Androgenetic alopecia (AGA) is the most frequent kind of clinical alopecia, accounting for 95% of all hair loss, and is characterized by symmetrical progressive hair thinning.1,2 Minoxidil was approved for the topical treatment of AGA in 1988.3 Many possibilities have been proposed, including activation of the Wnt/β-catenin signaling pathway in dermal papilla cell (DPC),4 promotion of follicular epithelial cell proliferation and differentiation, prolongation of the follicular anagen phase, and facilitation of the conversion of vellus hair to terminal hair. Upregulation of vascular endothelial growth factor (VEGF) mRNA and protein expression in DPC stimulates angiogenesis and increases local blood supply.5 Opening potassium channels allows more potassium ions into the cell and prevents calcium ions from entering, lowering the inhibitory impact of the epidermal growth factor on hair development.6 Minoxidil significantly reduces the expression of the IL-1 gene in human immortalized keratinocytes, probably via slowing the inflammatory process of AGA.7 In addition to the above effects, it has also been suggested that minoxidil effects are also related to sulfotransferase activity, and that minoxidil is only a prodrug that needs to be metabolised by follicular sulfotransferase to minoxidil sulfate (the active form) in order to be effective.8 Although many possibilities have been suggested, the therapeutic mechanism of minoxidil in AGA is uncertain.
Network pharmacology is a novel field based on building multi-layered networks of disease phenotypes, genes, and drugs to predict drug targets with a holistic view, allowing the discovery of additional mechanisms behind the disease.9,10 It connects artificial intelligence with medicine and the analysis of massive biomedical data, allowing the transformation of data completion into knowledge. Our curiosity about the causes of hair loss and the rejuvenating benefits of minoxidil on hair growth has been awakened. In this study, network pharmacology—which includes drug target prediction, network analysis, and experimental validation—was used to study the effects of minoxidil on AGA. In-depth studies on the mechanism of action of minoxidil may identify new therapeutic targets and provide new ideas for the development of new drugs.
Methods
Possible Target Proteins for Minoxidil
We wanted to obtain more potential target proteins for minoxidil and searched different databases (SwissTargetPrediction, DrugBank, and PharmMapper); we limited the species to “Homo sapiens”. Cytoscape software visualizes the relationship between minoxidil and target proteins, with nodes representing minoxidil and target proteins and edges representing their interaction.11
Collection of Related Genes Associated with AGA
A search of the Genecards and OMIM databases using the keywords “androgenetic alopecia” and “Homo sapiens” yielded a dataset containing 414 genes associated with androgenetic alopecia disorders.12
Screening of Crucial Target Proteins and GO and KEGG Analysis
The Minoxidil-AGA common target was imported into Cytoscape 3.9.0 software, and the “Network Analyzer” feature was used to examine the network topology of PPI data and build protein-protein interaction networks. The Cytoscape 3.9.0 plug-ins to NCA and cytohubba were used to do network topology analysis by “degree”, “betweenness”, and “closeness” of critical targets. “Nearness” for screening essential targets.13,14 For the intersection targets, Gene Ontology (GO) functional annotation and Kyoto Encyclopedia Of Genes and Genomes (KEGG) pathway analysis were done using the DAVID 6.8 database (https://david.Ncifcrf.gov/) enrichment analysis.
Molecular Docking
The screened core targets’ 3D structures were retrieved from the RCSB-PDB database (http://www.rcsb.org/), the proteins were dehydrated and hydrogenated using AutoDock 4.2 software, and molecular docking was performed using ChemBioDraw 3D software and Molegro Virtual Docker. ChemBioDraw 3D software displayed the 3D chemical structures and minimizes their energy, and Molegro Virtual Docker predicts the docking partners by comparing the predicted conformation with the observed crystal structure. The docking mode with the lowest binding energy to minoxidil was selected for each protein separately.15
Experimental Validation
Cell Culture and Drug Treatment
Human epidermal dermal papilla cell (CTCC-106-HUM) were purchased from MeisenCTCC (Zhejiang Meisen Cell Technology Co., Ltd). They were grown in Dulbecco’s modified Eagle’s medium (DMEM), 10% FBS (Corning), with 1% antibiotics (100 U/mL penicillin and 100 μg/mL streptomycin). The cell lines were cultured in 6-well plates and the medium was changed to DMEM with various additives except serum when 80% confluent cells were present. The cell incubation solution containing only 10 nM testosterone (MCE, HY-B1269) was used as the control group, and the group in which cells were incubated with 100 nM minoxidil (MCE, HY-B0112) in addition to 10 nM testosterone was called the minoxidil group. Cells were treated for 48 hours according to the above grouping. The doses of testosterone and minoxidil were chosen based on the cell culture methods used in previous studies and the results of the CCK-8 cell activity analysis that we performed (Supplementary Material 1).16–18
RT-PCR Analysis
DPC was inoculated in 6-well plates at 1×105 cells/well, and extracts were incubated with the medication above treatment for 48 hours using Roche’s RNA extraction kit. Following RNA extraction, cDNA was produced by reverse transcription using Roche’s cDNA synthesis kit and PCR thermal cycler according to the manufacturer’s instructions. Control and minoxidil group cells underwent real-time quantitative polymerase chain reaction examination. The results were utilized after the reaction to compute the target gene expression in each sample using the 2−ΔΔCt calculation technique. The following primer sequences were used:
AR:5′-ACAGAGGGAAAAAGGGCCGA-3′ (forward),5′-TACCAGGCACTTTCCTTGCTT-3′ (reverse); CYP17A1:5′-GCGATACCCTTACGGTTGTTG-3′ (forward),5′- TATGGCCCCATCTATTCGGTT-3′ (reverse); CYP19A1:5′-GCAAAGCACCCTAATGTTGAAGA-3′ (forward),5′-CGAGTCTGTGCATCCTTCCAATA-3′ (reverse), GADPH:5′-TGAAGGTCGGAGTCAACGGAT-3′ (forward),5′-CCTGGAAGATGGTGATGGGATT −3′ (reverse).
Western Blot
Add a suitable amount of RIPA lysate and protease inhibitor (100:1) to each well based on cell density, then lyse for 15 minutes on ice or at 4°C to recover cellular proteins. Electrophoresis of 10% sodium dodecyl sulfate-polyacrylamide gel separated the proteins, which were then transferred to a nitrocellulose membrane. The membranes were then sealed with 5% skim milk powder for 1 hour before being incubated overnight at 4°C with primary antibodies [AR (Abcam, ab133273), CYP17A1 (Abcam, ab125022), CYP19A1 (Abcam, ab18995), and β-actin (Santa Cruz Biotechnology, sc-47778)]. After shaking with PBST (phosphate buffered solution) the next day, add the appropriate secondary antibody (both 1:1000) in PBS configuration and incubate the secondary antibody for 30 minutes at room temperature on a light-protected shaker. Image J software calculated protein expression levels relative to β -actin.
Enzyme-Linked Immunosorbent Assay (ELISA)
After each group of cell specimens was treated accordingly, the cell supernatants were collected for the determination of E2 (Estradiol, Elabscience, E-EL-0150c) and DHT (Dihydrotestosterone, Elabscience, E-EL-0031c) using ELISA kits according to the manufacturer’s recommendations. Levels. Quantification was performed using GraphPad software.
Immunofluorescence Assay
Cells were fixed for 20 minutes in 4% paraformaldehyde, washed, and permeabilized for 10 minutes in Immunostaining Permeabilization Buffer with Saponin (Beyotime, P0095-500mL). The portions were closed for 1 hour at room temperature with a closure solution. After blocking, slices were incubated at 4 °C overnight with primary antibodies (both 1:500). Primary antibodies were as follows: AR (Abcam, ab133273), CYP17A1 (Abcam, ab125022), CYP19A1 (Abcam, ab18995). After washing with PBS (5 minutes 3 times) the following day, a secondary antibody conjugated with Alexa Fluor 488 (Invitrogen, A32731) was incubated. After washing in PBS, the pictures were examined under a microscope with 4′,6-diamidino-2′-phenylindole dihydrochloride (DAPI) (Beyotime, C1006).
Result
The Predicted Targets of Minoxidil and Their Network
Based on the 2D and 3D chemical structures (Figure 1A), we predicted the proteins targeted by minoxidil on different databases. The top 100 were selected based on the likelihood score index (Figure 1B). The three main types of these targets were Enzyme (20%), Kinase (23%), and Protease (14%) (Figure 1C). Key potential targets with more interactions (degree index) with other targets were screened using Cytoscape software to construct the PPI network (Figure 2).
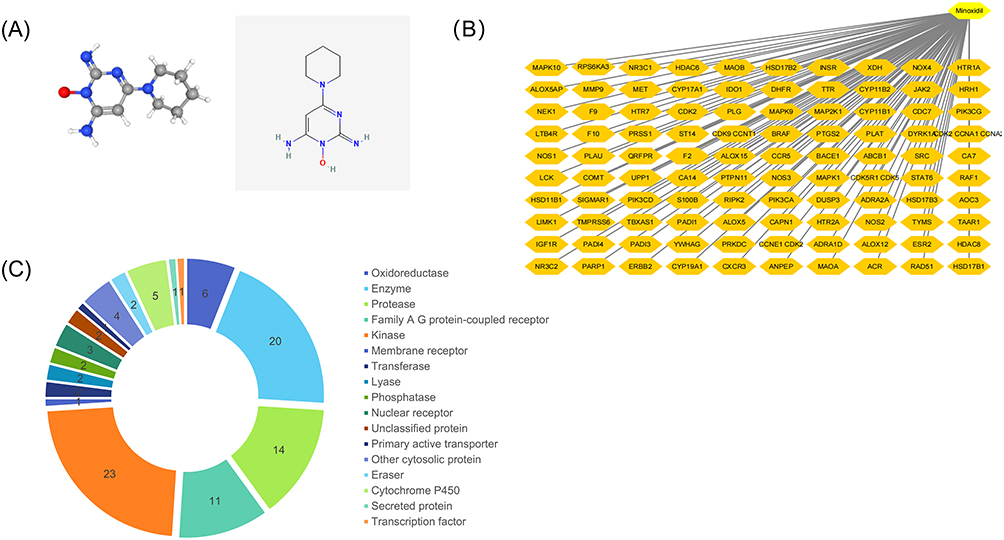 |
Figure 1 Targets predicted by the database involving minoxidil: (A) 3D and 2D chemical structures of minoxidil. (B) Top 50 potential targets of minoxidil. (C) Top 50 targets by category. |
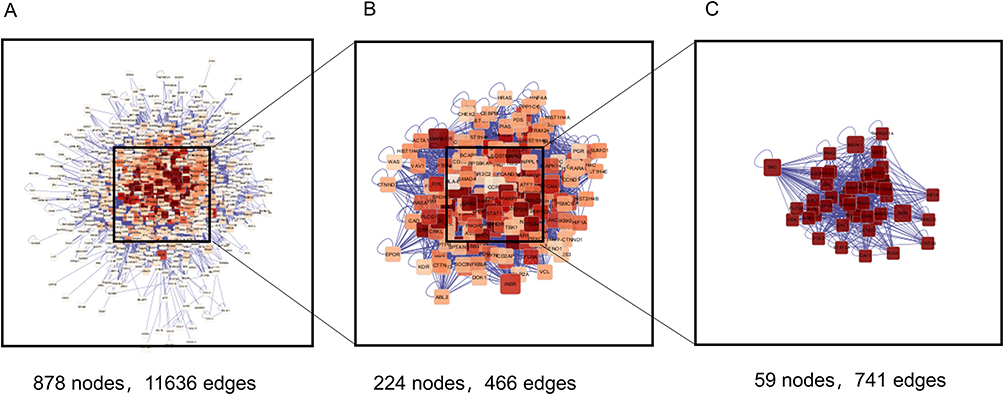 |
Figure 2 PPI network study using nine shared genes that are potential targets of androgenetic alopecia and minoxidil. (A) Amplified PPI network of 878 proteins. (B) 224 nodes after the first screening. (C) 59 nodes from the second screening. |
Gene Ontology and Pathway Enrichment of 878 Related Proteins
A total of 878 human genes involved in the mechanism of minoxidil treatment of androgenetic alopecia were enriched using GO and KEGG enrichment (Figure 3). Microbiological evaluation of these proteins was performed.
 |
Figure 3 878 gene enrichment analysis. (A) Top 10 significant changes in three different aspects of Biological Processes, Cellular Components, and Molecular Functions in GO analysis. (B) The first 20 significant changes in the KEGG pathway. |
According to GO enrichment: these proteins were mainly located in the focal adhesion, cell-substrate junction, membrane raft, and membrane microdomain (Figure 3A); as for the molecular functions, they were mainly concentrated in kinase binding, protein kinase binding, protein domain specific binding, protein kinase activity (Figure 3A); their biological processes were mainly focused on enzyme linked receptor protein signaling pathway, transmembrane receptor protein tyrosine kinase signaling pathway, response to the hormone, cellular response to hormone stimulation (Figure 3A).
KEGG enrichment analysis showed the top 20 pathways involved in minoxidil for androgenetic alopecia (Figure 3B), including the MAPK signaling pathway, Insulin signaling pathway, Estrogen signaling pathway, etc.
Molecular Docking
A total of 22 putative potential targets of minoxidil were analyzed using molecular docking (Supplementary Material 2), which offered a visual assessment of the interactions between minoxidil and its potential protein targets associated with AR. We concluded that values less than −10 indicated higher docking ability, which resulted in the screening of the top 15 targets (Table 1 and Figure 4). By combining the GO and KEGG results, we determined that AR, CYP17A1, and CYP19A1 were the most likely targets of minoxidil, with the highest binding power and spatial fit.
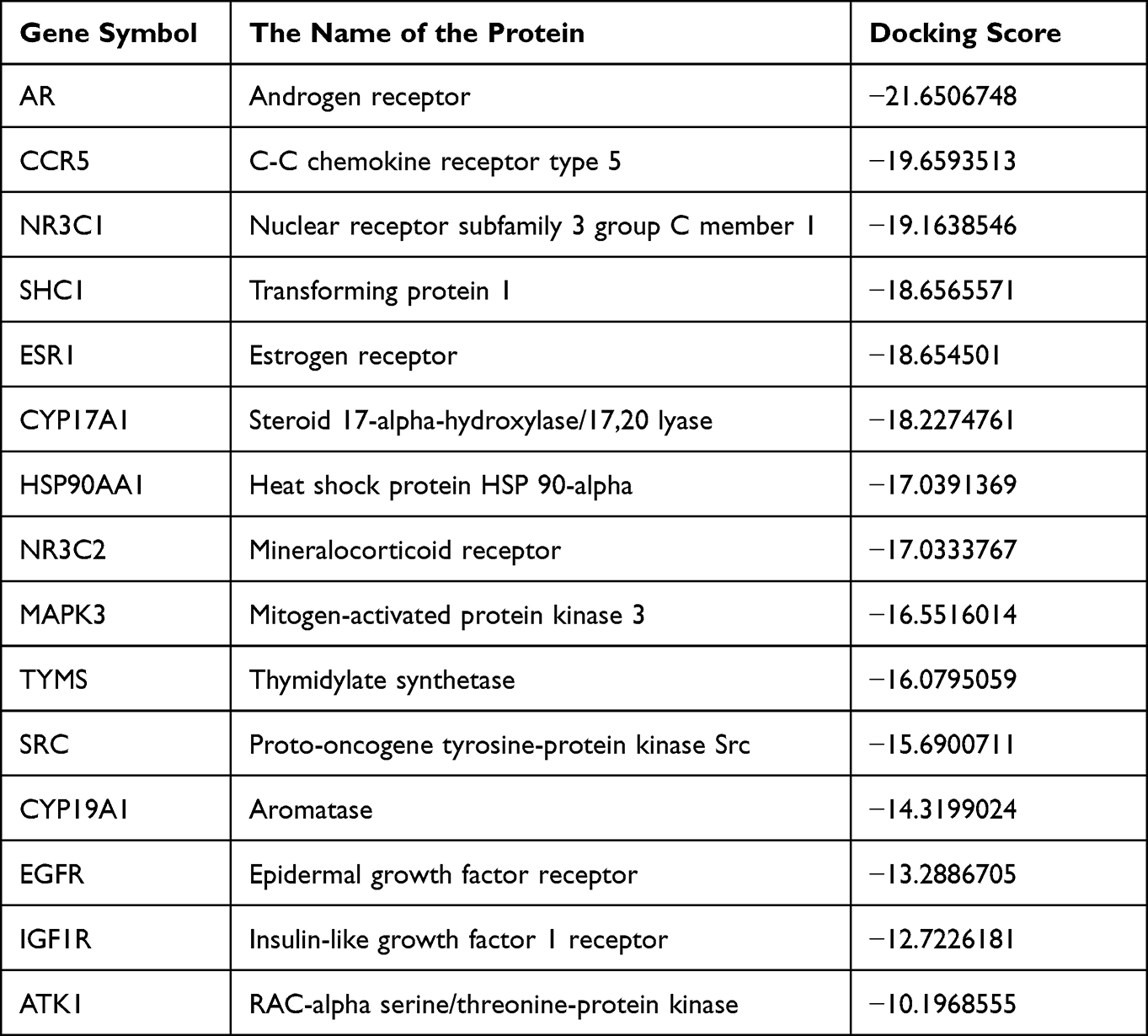 |
Table 1 Top 15 Targets for Androgenetic Alopecia Based on Docking Scores |
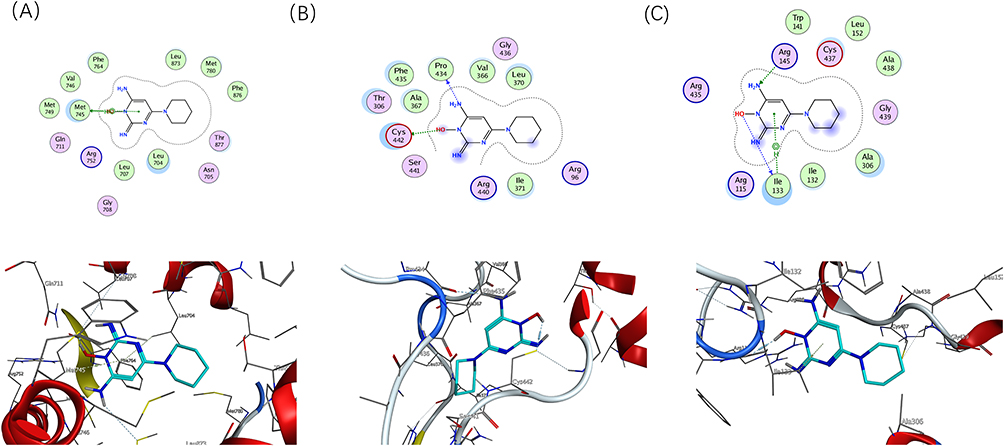 |
Figure 4 Molecular model of minoxidil binding to its predicted protein targets. Proteins 【(A) AR, (B) CYP17A1, (C) CYP19A1】are shown interacting with minoxidil molecules, represented by green bar models. The lines represent residues at the binding site. The blue dashed lines represent hydrogen bonds and the green dashed lines represent p-p interactions. |
Experimental Validation
Minoxidil Reduces Expression of AR
ARs were found in dermal papillae rather than epithelial cells in hair follicles, indicating that the latter were the primary targets of androgens in hair follicles. Based on the molecular docking results, stimulation of DPC with minoxidil followed by RT-PCR and Western Blot assay confirmed a significant decrease in AR mRNA expression in DPC in the minoxidil group (Figure 5A), as well as a significant decrease in AR protein expression (Figure 5B), indicating that minoxidil may act by inhibiting AR to reduce androgen sensitivity.
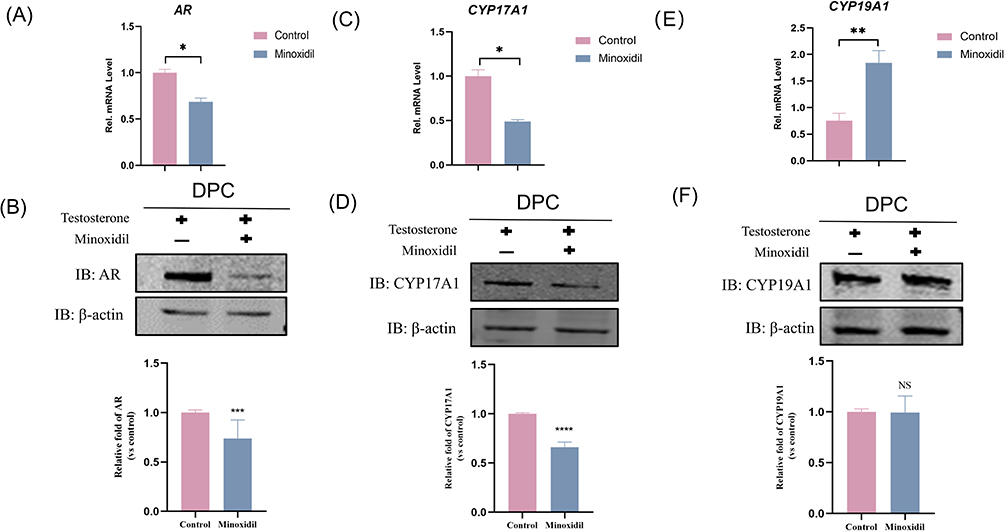 |
Figure 5 Effects of minoxidil-induced CYP19A1, CYP17A1, and AR expression levels in patients with androgenetic alopecia. (A, C and E) Relative expression of CYP19A1, CYP17A1, and AR detected by PCR analysis. (B, D and F) Relative expression of CYP19A1, CYP17A1, and AR detected by protein blotting analysis. Columns labeled ****p < 0.0001, ***p < 0.001, **p < 0.01, *p < 0.05 show statistically significant differences between control and experimental groups. NS p > 0.05 indicate that there is no statistically significant difference between the control and minoxidil groups. |
Minoxidil Regulates Hormone-Producing Related Enzymes
CYP17A1 and CYP19A1 are cytochrome P450 family members that play essential roles in steroidogenic responses, cholesterol metabolism, and drug metabolism.19 The effect of minoxidil on the expression of hormonogenesis-related enzymes CYP17A1 and CYP19A1 mRNA and protein in DPC was investigated. The results showed that minoxidil significantly lowered CYP17A1 gene transcript levels (Figure 5C) and CYP17A1 protein levels (Figure 5D). Although CYP19A1 gene transcript levels increased by 1.84-fold (Figure 5E), there was no significant difference in CYP19A1 protein expression (Figure 5F). Immunofluorescence assay was similar to the above results, and it was seen that the expression of AR and CYP17A1 were all reduced in the cells of minoxidil group, and the expression of CYP19A1 was still not enough to have a significant difference (Figure 6).
 |
Figure 6 Immunofluorescence assay: Minoxidil induced a decrease in CYP17A1, AR expression in patients with androgenetic alopecia, but the difference in CYP19A1 expression was not significant. |
Changes in hormone content also showed enzyme changes laterally, and we used the Elisa assay to detect E2 and DHT changes in the supernatant of cells after drug stimulation. The E2 level in the minoxidil group was determined to be 13.22 pg/mL, while the E2 level in the control group was 12.01 pg/mL. The E2 level increased by 1.21 pg/mL in the minoxidil group compared to the control group (Figure 7A), which was statistically significant. The DHT level in the experimental group was 267.43 pg/mL, and 346.86 pg/mL in the control group. The minoxidil group’s DHT level fell by 79.43 pg/mL compared to the control group (Figure 7B), which was statistically significant. CYP19A1 activity was measured by calculating the percentage of T (Testosterone) conversion to E2, and CYP19A1 activity was significantly increased in the minoxidil group (Figure 7C).
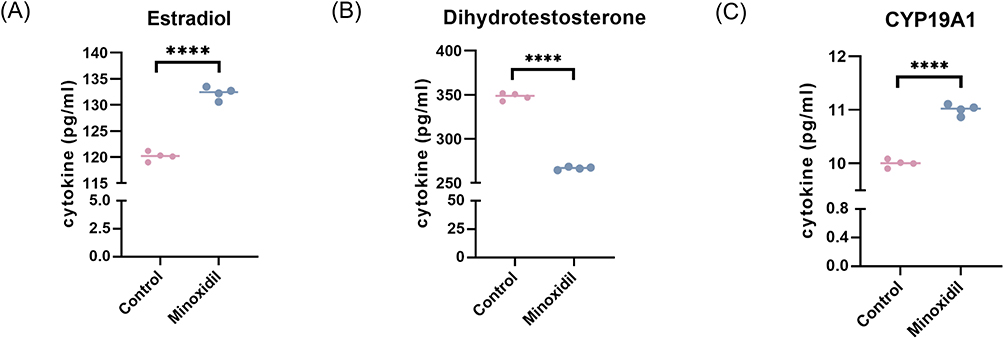 |
Figure 7 Minoxidil-induced conversion of testosterone to estradiol in patients with androgenetic alopecia and reduction of dihydrotestosterone (A and B) estradiol dihydrotestosterone levels in the control group and the minoxidil group. (C) Changes in CYP19A1 (Aromatase) activity. Columns labeled ****p < 0.0001 show statistically significant differences between control and experimental groups. |
Discussion
Even though minoxidil has been used to treat hair loss for over 30 years, the mechanism by which minoxidil promotes hair growth remains to be fully elucidated. This study was to explore the mechanism of action of minoxidil on AGA from a network pharmacology standpoint. The strategy involved drug target prediction and network analysis, followed by cellular studies to validate the prediction.
Combined with the network pharmacology section, according to GO and KEGG, it was known that the targets of minoxidil action were concentrated in some enzyme and hormone-related pathways, and molecular docking provides the target. We demonstrated experimentally that minoxidil may treat AGA by targeting CYP17A1, AR, and CYP19A1.
AR expression was significantly higher in the DPCs of AGA patients’ balding scalps. ARs are candidate genes for the x chromosome’s primary AGA risk locus.20,21 The combination of ARs and androgens activated hair growth regulation. This resulted in shortening and progressive atrophy of thinning hairs in the early stages of hair growth and ultimately inhibition of hair growth.22,23 Under the action of minoxidil, gene and protein expression of ARs was considerably reduced in the minoxidil group compared to the control group. Hsu et al previously observed similar results in androgen-sensitive LNCaP cells that express AR receptors endogenously.18 Minoxidil may prevent hair follicle miniaturization by reducing AR in DPC and inhibiting DHT binding to them.
Although androgens play a role in the etiology of AGA, circulating androgen levels were standard in almost all AGA patients.24 Localized hormone levels had a more substantial impact on the hair follicle, and androgen-metabolizing enzymes found in the hair follicle may play a role.25,26
Through the PPI network and molecular docking, we screened 2 enzymes, CYP17A1 and CYP19A1, in the hormone conversion process. The anterior and posterior DHT production routes required CYP17A1.27,28 Minoxidil significantly suppresses the CYP17A1 gene and protein expression, which may decrease DHT production (Figure 8), resulting in a therapeutic effect on hair loss. Similarly, a study found that a good clinical study of topical ketoconazole for hair loss may be related to the inhibition of CYP17A1.29,30 CYP17A1 may also be a target of interest in alopecia areata treatment and could be used to validate subsequent hair loss drug development.
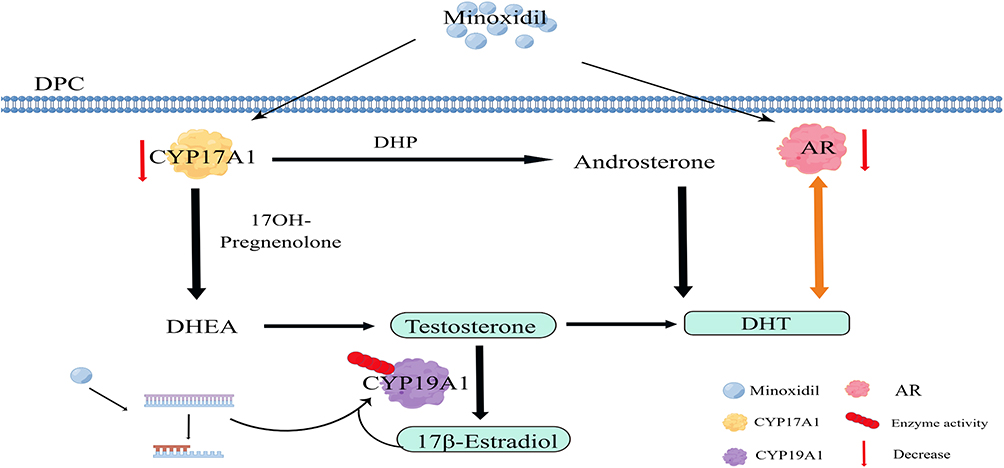 |
Figure 8 Multi-target mechanism of minoxidil for the treatment of androgenetic alopecia. Minoxidil significantly inhibits CYP17A1 and AR expression, thus reducing DHT production. Minoxidil may stimulate a mutation in the CYP19A1 gene, altering enzyme activity and facilitating the conversion of Testosterone to 17β-Estradiol, increasing 17β-Estradiol production. Abbreviations: DPC, dermal papilla cell; DHP, dihydroprogesterone; AR, androgenic receptors; DHEA, dehydroepiandrosterone; DHT, dihydrotestosterone; 17OH-pregnenolone: 17-Hydroxypregnenolone. |
According to our study, minoxidil did not significantly increase CYP19A1 expression but increased CYP19A1 activity; as shown in Figure 8, CYP19A1 can convert T to E2 and thus increase estrogen production. The altered activity may be due to drug stimulation of CYP19A1 gene mutations31 or positive estrogen feedback stimulation.32 There is some clinical evidence that CYP19A1 activity may be related to the pathogenesis of AGA; for example, one adverse effect of aromatase inhibitor medication is hair loss,33 which also causes women to exhibit male pattern hair loss.34 Combined with the results of changes in CYP19A1 activity and changes in E2 content, CYP19A1 may stimulate hair growth by increasing estrogen concentrations. It was also confirmed in an in vitro study that increasing estrogen concentrations stimulates hair growth.32 However, genetic studies suggest that higher estrogen levels may have the opposite effect.35 Systemic circulating estrogen levels’ role in hair growth is controversial.
We thus performed cellular experiments focusing on local studies of hair follicles. We found that estrogen levels increased in the minoxidil group relative to the control group after minoxidil use, but not by much. Because minoxidil can boost estrogen while decreasing DHT, it may enhance hair growth by adjusting the ratio of local estrogen concentration to DHT concentration. This sparked further discussion, as most existing research evaluated androgens in female pattern hair loss (FPHL) and discovered that many were non-hyperandrogenic,36 but few studies measured circulating estrogens in patients at the same time. Our next focus should be determining whether hair loss is related to the ratio of systemic estrogen to DHT concentration, which requires more research.
Conclusion
Minoxidil may act by altering hormonal and enzymatic pathways. Our study finds two new targets (CYP17A1, CYP19A1) of minoxidil and demonstrates that minoxidil inhibits AR. These targets may provide new directions for drug research.
Data Sharing Statement
Data sharing is not applicable to this article as no new data were created or analyzed in this study.
Acknowledgments
- Basic Public Welfare Research Project of Zhejiang (NO: LY23H110001).
- Science and Technology Major Project of Zhejiang Province and the State Administration of Traditional Chinese Medicine (NO: GZY-ZJ-KJ-23035).
- Health Science and Technology Major Project of Hangzhou (NO: Z20220040).
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Disclosure
The authors declare no conflict of interest.
References
1. Martinez-lopez A, Montero-Vilchez T, Sierra-Sánchez Á, Molina-Leyva A, Arias-Santiago S. Advanced medical therapies in the management of non-scarring alopecia: areata and Androgenic Alopecia. Int J Mol Sci. 2020;21(21):8390.
2. Starace M, Orlando G, Alessandrini A, Piraccini BM. Female androgenetic alopecia: an update on diagnosis and management. Am J Clin Dermatol. 2020;21(1):69–84.
3. Nestor MS, Ablon G, Gade A, Han H, Fischer DL. Treatment options for androgenetic alopecia: efficacy, side effects, compliance, financial considerations, and ethics. J Cosmet Dermatol. 2021;20(12):3759–3781.
4. Bao L, Zong H, Fang S, et al. Randomized trial of electrodynamic microneedling combined with 5% minoxidil topical solution for treating androgenetic alopecia in Chinese males and molecular mechanistic study of the involvement of the Wnt/β-catenin signaling pathway. J Dermatolog Treat. 2022;33(1):483–493. doi:10.1080/09546634.2020.1770162
5. Yum S, Jeong S, Kim D, et al. Minoxidil induction of VEGF is mediated by inhibition of HIF-prolyl hydroxylase. Int J Mol Sci. 2017;19(1):53. doi:10.3390/ijms19010053
6. Suchonwanit P, Thammarucha S, Leerunyakul K. Minoxidil and its use in hair disorders: a review. Drug Des Devel Ther. 2019;13:2777–2786.
7. Pekmezci E, Turkoğlu M, Gökalp H, et al. Minoxidil downregulates interleukin-1 alpha gene expression in HaCaT cells. Int J Trichology. 2018;10(3):108–112. doi:10.4103/ijt.ijt_18_17
8. Gupta AK, Talukder M, Venkataraman M, Bamimore MA. Minoxidil: a comprehensive review. J Dermatolog Treat. 2022;33(4):1896–1906.
9. Nogales C, Mamdouh ZM, List M, Kiel C, Casas AI, Schmidt HH. Network pharmacology: curing causal mechanisms instead of treating symptoms. Trends Pharmacol Sci. 2022;43(2):136–150.
10. Ma H, Xu F, Liu C, et al. A network pharmacology approach to identify potential molecular targets for cannabidiol’s anti-inflammatory activity. Cannabis Cannabinoid Res. 2021;6(4):288–299. doi:10.1089/can.2020.0025
11. Otasek D, Morris JH, Bouças J, Pico AR, Demchak B. Cytoscape automation: empowering workflow-based network analysis. Genome Biol. 2019;20(1):185.
12. Stelzer G, Rosen N, Plaschkes I, et al. The GeneCards suite: from gene data mining to disease genome sequence analyses. Curr Protoc Bioinformatics. 2016;54(1):1.30.1–1.3. doi:10.1002/cpbi.5
13. Martin A, Ochagavia ME, Rabasa LC, et al. BisoGenet: a new tool for gene network building, visualization and analysis. BMC Bioinform. 2010;11(1):91. doi:10.1186/1471-2105-11-91
14. Azuaje FJ, Zhang L, Devaux Y, et al. Drug-target network in myocardial infarction reveals multiple side effects of unrelated drugs. Sci Rep. 2011;1(1):52. doi:10.1038/srep00052
15. Rizvi SM, Shakil S, Haneef M. A simple click-by-click protocol to perform docking: autoDock 4.2 made easy for non-bioinformaticians. Excli J. 2013;12:831–857.
16. Giatromanolaki A, Fasoulaki V, Kalamida D, et al. CYP17A1 and androgen-receptor expression in prostate carcinoma tissues and cancer cell lines. Curr Urol. 2019;13(3):157–165. doi:10.1159/000499276
17. Han JH, Kwon OS, Chung JH, Cho KH, Eun HC, Kim KH. Effect of minoxidil on proliferation and apoptosis in dermal papilla cells of human hair follicle. J Dermatol Sci. 2004;34(2):91–98. doi:10.1016/j.jdermsci.2004.01.002
18. Hsu CL, Liu JS, Lin AC, Yang CH, Chung WH, Wu WG. Minoxidil may suppress androgen receptor-related functions. Oncotarget. 2014;5(8):2187–2197.
19. Heidarzadehpilehrood R, Pirhoushiaran M, Abdollahzadeh R, et al. A review on CYP11A1, CYP17A1, and CYP19A1 polymorphism studies: candidate susceptibility genes for Polycystic Ovary Syndrome (PCOS) and infertility. Genes. 2022;13(2):302.
20. Sawaya ME, Price VH. Different levels of 5alpha-reductase type I and II, aromatase, and androgen receptor in hair follicles of women and men with androgenetic alopecia. J Invest Dermatol. 1997;109(3):296–300. doi:10.1111/1523-1747.ep12335779
21. Y CEG, Tan JH, Bahta AW, et al. Differential expression between human dermal papilla cells from balding and non-balding scalps reveals new candidate genes for androgenetic alopecia. J Invest Dermatol. 2016;136(8):1559–1567.
22. Heymann WR. The inflammatory component of androgenetic alopecia. J Am Acad Dermatol. 2022;86(2):301–302. doi:10.1016/j.jaad.2021.11.013
23. Hu S, Li Z, Lutz H, et al. Dermal exosomes containing miR-218-5p promote hair regeneration by regulating β-catenin signaling. Sci Adv. 2020;6(30):eaba1685.
24. Orme S, Cullen DR, Messenger AG. Diffuse female hair loss: are androgens necessary? Br J Dermatol. 1999;141(3):521–523.
25. Chen WC, Zouboulis CC. Hormones and the pilosebaceous unit. Dermatoendocrinol. 2009;1(2):81–86. doi:10.4161/derm.1.2.8354
26. Zouboulis CC, Degitz K. Androgen action on human skin - from basic research to clinical significance. Exp Dermatol. 2004;13(Suppl 4):5–10.
27. O’Shaughnessy PJ, Antignac JP, Le Bizec B, et al. Alternative (backdoor) androgen production and masculinization in the human fetus. PLoS Biol. 2019;17(2):e3000002.
28. Fukami M, Homma K, Hasegawa T, Ogata T. Backdoor pathway for dihydrotestosterone biosynthesis: implications for normal and abnormal human sex development. Dev Dyn. 2013;242(4):320–329.
29. Patel V, Liaw B, Oh W. The role of ketoconazole in current prostate cancer care. Nat Rev Urol. 2018;15(10):643–651.
30. Marks DH, Prasad S, De Souza B, Burns LJ, Senna MM. Topical antiandrogen therapies for androgenetic alopecia and acne vulgaris. Am J Clin Dermatol. 2020;21(2):245–254.
31. Ashraf S, Rasool SUA, Nabi M, et al. CYP17 gene polymorphic sequence variation is associated with hyperandrogenism in Kashmiri women with polycystic ovarian syndrome. Gynecol Endocrinol. 2021;37(3):230–234. doi:10.1080/09513590.2020.1770724
32. Hoffmann R, Niiyama S, Huth A, Kissling S, Happle R. 17alpha-estradiol induces aromatase activity in intact human anagen hair follicles ex vivo. Exp Dermatol. 2002;11(4):376–380.
33. Gallicchio L, Calhoun C, Helzlsouer KJ. Aromatase inhibitor therapy and hair loss among breast cancer survivors. Breast Cancer Res Treat. 2013;142(2):435–443. doi:10.1007/s10549-013-2744-2
34. Rossi A, Iorio A, Scali E, et al. Aromatase inhibitors induce ‘male pattern hair loss’ in women. Ann Oncol. 2013;24(6):1710–1711.
35. Yip L, Zaloumis S, Irwin D, et al. Gene-wide association study between the aromatase gene (CYP19A1) and female pattern hair loss. Br J Dermatol. 2009;161(2):289–294. doi:10.1111/j.1365-2133.2009.09186.x
36. Futterweit W, Dunaif A, Yeh HC, Kingsley P. The prevalence of hyperandrogenism in 109 consecutive female patients with diffuse alopecia. J Am Acad Dermatol. 1988;19(5 Pt 1):831–836. doi:10.1016/S0190-9622(88)70241-8
 © 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.