Back to Journals » Nature and Science of Sleep » Volume 18

Network Pharmacology-Based Elucidation of Gu’s Anshen Decoction in Modulating TNF Signaling to Ameliorate Insomnia
Received 20 November 2025
Accepted for publication 19 May 2026
Published 28 May 2026 Volume 2026:18 582222
DOI https://doi.org/10.2147/NSS.S582222
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Sarah L Appleton
Sanyuan Gu, Jingyu Xu
Leshan Vocational and Technical College, Leshan, 614000, People’s Republic of China
Correspondence: Jingyu Xu, Leshan Vocational and Technical College, No. 1336, Middle Section of Qingjiang Road, Shizhong District, Leshan, Sichuan Province, 614000, People’s Republic of China, Email [email protected]
Objective: To investigate the therapeutic mechanisms of Gu’s Anshen decoction (GAD) against insomnia (INS), focusing on the regulation of tumor necrosis factor (TNF)-mediated inflammatory pathways.
Methods: Bioactive compounds and potential targets of GAD were identified via network pharmacology and constructed herb–compound–target interaction network. INS-related targets derived from peripheral blood mononuclear cells in the Gene Expression Omnibus (GEO) database were intersected with GAD targets to establish a protein-protein interaction (PPI) network. Gene Ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) enrichment analyses were performed to identify the key pathways. Experimental validation employed a para-chlorophenylalanine (PCPA)-induced INS rat model, integrating behavioral tests, histopathology, and inflammatory cytokine quantification.
Results: Network pharmacology revealed multi-target, multi-pathway actions of GAD against INS, with the TNF signaling pathway identified as a critical axis. Animal experiments demonstrated that GAD significantly improved sleep architecture in INS rats, reduced serum and hippocampal levels of TNF-α and interleukin-1β (IL-1β), and downregulated phosphorylated NF-κB p65 (p-p65) expression, indicating suppression of TNF-driven neuroinflammation.
Conclusion: GAD ameliorates INS in a rat model by inhibiting TNF-mediated inflammatory signaling, offering a potential mechanistic basis for its therapeutic use in INS management.
Keywords: Gu’s Anshen decoction, insomnia, network pharmacology, TNF signaling pathway, neuroinflammation
Introduction
Insomnia (INS), characterized by persistent difficulties in sleep initiation, maintenance, or non-restorative sleep, is a prevalent global health concern affecting 10%–30% of the population, with incidence rates demonstrating a progressive upward trajectory.1 Beyond immediate consequences such as impaired concentration and emotional dysregulation, chronic INS is strongly linked to the pathogenesis of hypertension, diabetes mellitus, cognitive decline, and psychiatric disorders, posing substantial threats to both physical well-being and quality of life.2,3 Current pharmacological management primarily relies on benzodiazepines and non-benzodiazepine hypnotics. Although these agents provide rapid symptomatic relief, their prolonged use carries considerable risks of dependence, tolerance, and cognitive impairment, thereby limiting their therapeutic utility.4 Consequently, developing safer, mechanism-based interventions is an urgent priority in sleep medicine.
Traditional Chinese medicine (TCM) offers a holistic therapeutic paradigm based on syndrome differentiation and treatment.5 Herbal formulations exert multi-target effects through synergistic modulation of physiological networks, demonstrating durable efficacy with minimal adverse effects in managing sleep disorders.6 Several empirically developed Anshen decoctions have demonstrated clinical efficacy in ameliorating INS. For instance, a recent retrospective study showed that Tiaoshu Anshen decoction improved sleep disorders by regulating Qi dynamics and gastrointestinal function.7 Additionally, an animal experiment provided favorable evidence that Huaxiang Anshen decoction exerted sedative and hypnotic effects in para-chlorophenylalanine (PCPA)-induced INS mice.8 Gu’s Anshen decoction (GAD) is formulated based on TCM principles aiming to restore heart-kidney balance, calm the mind, and regulate Qi and blood.6 The formula comprises 19 herbs that work synergistically: Ziziphi Spinosae Semen, Polygalae Radix, and Platycladi Semen serve as principal herbs to nourish the heart and tranquilize the mind;9–11 Salviae Miltiorrhizae Radix et Rhizoma and Chuanxiong Rhizoma promote blood circulation and remove blood stasis, with anti-inflammatory and neuroprotective effects;12,13 Coptidis Rhizoma and Anemarrhenae Rhizoma clear heart fire and nourish yin;14,15 Ganoderma and Lilii Bulbus tonify Qi and calm the spirit;16,17 while mineral components such as Magnetitum, Chloriti Lapis, Margaritifera Concha, and Ostreae Concha are traditionally included to anchor the mind and improve sleep quality. This combination addresses both root causes and manifestations of the disorder, reflecting the holistic multi-target approach of TCM.6
Despite growing interest in TCM formulations for INS, the specific mechanisms underlying GAD’s therapeutic effects remain poorly understood. Although several constituent herbs, such as Coptidis Rhizoma, Salviae Miltiorrhizae Radix, and Ganoderma, have been individually associated with anti-inflammatory and immunomodulatory activities,12,14,16 whether GAD as a whole formula modulates inflammatory pathways to alleviate INS has not been systematically investigated. Emerging insights into the neuro-immuno-endocrine network have illuminated the pivotal role of inflammatory cascades in INS pathophysiology.18 Chronic sleep disruption activates central and peripheral immune responses, triggering the excessive release of pro-inflammatory cytokines, including tumor necrosis factor-alpha (TNF-α), interleukin-1β (IL-1β), and interleukin-6 (IL-6), which propagate neuroinflammatory cascades.19,20 The TNF signaling pathway, as a central inflammatory regulator, engages downstream effectors such as nuclear factor-kappa B (NF-κB) to drive neuronal injury, neurotransmitter dysregulation [eg., aberrant serotonin and γ-aminobutyric acid (GABA) signaling], and sleep-wake cycle disturbances, thereby representing a promising therapeutic target.21,22 However, whether GAD exerts its anti-INS effects through TNF pathway modulation remains unexplored. Addressing this knowledge gap is essential to elucidate the mechanistic basis of GAD and to support its broader clinical translation.
Network pharmacology, an interdisciplinary approach integrating systems biology, bioinformatics, and polypharmacology, provides a robust framework for deconvoluting the complex mechanisms of herbal formulations by constructing herb–compound–target–disease interaction networks.23 Using this methodology, we first employed computational strategies to identify GAD’s bioactive components and potential targets, subsequently intersecting these with INS-associated differentially expressed genes (DEGs) to construct protein-protein interaction (PPI) networks and perform functional enrichment analysis for pathway prediction. These predictions were experimentally validated using a PCPA-induced INS rat model through integrated behavioral, histopathological, and molecular assessments of TNF pathway regulation. This study aimed to elucidate the scientific basis of GAD’s anti-INS efficacy, providing empirical support for its clinical application, while advancing novel insights into inflammation-targeted therapies for INS. From a translational perspective, confirming that GAD acts through TNF signaling may inform the development of mechanism-based adjunctive or alternative treatments for INS, particularly in populations with inflammatory comorbidities.
Methods
Identification of Bioactive Components and Target Prediction for GAD
GAD comprises Lilii Bulbus, Ganoderma, Polygoni Multiflori Caulis, Albiziae Cortex, Margaritifera Concha, Coptidis Rhizoma, Ziziphi Spinosae Semen, Poria cum Radix Pini, Polygalae Radix, Magnetitum, Chloriti Lapis, Chuanxiong Rhizoma, Schisandrae Chinensis Fructus, Cinnamomi Cortex, Salviae Miltiorrhizae Radix et Rhizoma, Ostreae Concha, Platycladi Semen, Corydalis Rhizoma, and Anemarrhenae Rhizoma. Bioactive components were screened using the Traditional Chinese Medicine Systems Pharmacology Database and Analysis Platform (TCMSP; https://old.tcmsp-e.com/index.php) with established criteria of oral bioavailability (OB) ≥ 30% and drug-likeness (DL) ≥ 0.18. Potential therapeutic targets were retrieved from the TCMSP target module, supplemented with active ingredient data from the Herb database (http://herb.ac.cn/). To construct the herb-compound-target-disease network, the predicted targets were further intersected with INS-related DEGs obtained from the GEO dataset. Only herbs whose targets showed direct overlap with the INS-related gene set were retained for subsequent network visualization. A compound-target interaction network was constructed using Cytoscape software (version 3.10.2), where nodes represent bioactive compounds and potential targets and edges denote molecular interactions.
Identification and Analysis of INS-Related Target Genes
DEGs (P < 0.05, |logFC| > 1) between patients with INS (n = 17) and non-INS controls (n = 25) were obtained from the GSE208668 dataset in the Gene Expression Omnibus (GEO) database (https://www.ncbi.nlm.nih.gov/geo/), focusing on peripheral blood mononuclear cells (PBMCs). The intersection between these DEGs and the predicted drug targets was identified and subsequently visualized using Cytoscape 3.10.2 to elucidate potential therapeutic interactions. To further explore the biological functions and pathways associated with these overlapping genes, Gene Ontology (GO) enrichment and Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway analyses were performed using the bioinformatics platform (https://www.bioinformatics.com.cn/). The results were ranked based on the number of enriched genes per pathway.
PPI Network Analysis
PPI data for all GAD targets were retrieved from the Search Tool for Recurring Instances of Neighboring Genes (STRING) database (https://string-db.org/). Network topology analysis was subsequently performed using Cytoscape, and target connectivity was quantified using the degree centrality parameter.
Molecular Docking
The three-dimensional structures of key bioactive compounds were retrieved from PubChem, whereas the crystal structures of the core target proteins were acquired from the Protein Data Bank (PDB; https://www.rcsb.org/). Protein structures were preprocessed using PyMOL to eliminate water molecules, metal ions, and small-molecule ligands, followed by identification of active binding pockets. Molecular docking simulations were performed using AutoDock Vina (version 1.1.2), involving preparation of PDBQT-format files. The binding energies, which are quantitative indicators of the complex stability, were calculated. The conformation exhibiting the lowest binding energy was visualized using PyMOL to analyze specific intermolecular interactions.
Animal Experiments
This study was approved by the Animal Ethics Committee of Hunan Evidence-based Biotechnology Co. Ltd. (approval no. XZ250711). All animal procedures were performed in accordance with the National Institutes of Health Guide for the Care and Use of Laboratory Animals and reported in compliance with the ARRIVE guidelines. Twenty-four male Sprague-Dawley (SD) rats (6-week-old, body weight 200–230 g) were obtained from the Shanghai SLAC Laboratory Animal Co., Ltd. Animals were housed under standardized conditions with a 12-h light/dark cycle, ambient temperature maintained at 22 ± 2°C, and relative humidity controlled at 50 ± 5%. Throughout the one-week acclimatization period, all rats were provided with free access to diet and water.
Animal Grouping and Modeling
Twenty-four male SD rats were randomly allocated to four experimental groups (n = 6): Control, INS, INS + saline, and INS + GAD. The sample size was determined based on previous studies using similar PCPA-induced INS rat models24,25 and in accordance with the 3Rs (Replacement, Reduction, Refinement) principles for animal experimentation. To establish the INS model, all groups except the control group received daily intraperitoneal injections of PCPA (300 mg/kg, 0938, Tocris Bioscience, Minneapolis, MN, USA) for three consecutive days, consistent with validated protocols.26 On day 4, the INS + GAD group received oral gavage administration of the GAD solution, whereas the INS + saline group received equivalent volumes of normal saline, both continuing for six days. Twenty-four hours after the final treatment, the rats were anesthetized with 2% isoflurane27 and blood was collected via the abdominal aorta.28 Subsequently, euthanasia was performed by intraperitoneal injection of an overdose of sodium pentobarbital (200 mg/kg; P3761; Sigma-Aldrich, Darmstadt, Germany), followed by rapid brain tissue extraction for subsequent analyses.
Pharmacological Intervention
The base formulation of GAD comprised the following crude herbs: Lilii Bulbus (15 g, U1000043, Beijing Tongrentang, China), Ganoderma (30 g, U1000066, Beijing Tongrentang), Polygoni Multiflori Caulis (30 g, 100223478731, Suzetang, Anhui, China), Albiziae Cortex (30 g, 100216388725, Suzetang), Margaritifera Concha (30 g, 411535, Beijing Tongrentang), Coptidis Rhizoma (6 g, 237190, Beijing Tongrentang), Ziziphi Spinosae Semen (30 g, Q2100143, Beijing Tongrentang), Poria cum Radix Pini (30 g, 100203890881, Taitaotang, Sichuan, China), Polygalae Radix (10 g, 100200770549, Taitaotang), Magnetitum (30 g, 205868, Beijing Tongrentang), Chloriti Lapis (30 g, 10106386905022, Jingwan, Bozhou, China), Chuanxiong Rhizoma (30 g, 100215682393, Suzetang), Schisandrae Chinensis Fructus (10 g, 7507088, Beijing Tongrentang), Cinnamomi Cortex (6 g, U1000059, Beijing Tongrentang), Salviae Miltiorrhizae Radix et Rhizoma (15 g, 100013741708, Beijing Tongrentang), Ostreae Concha (30 g, 100288478124, Suzetang), Platycladi Semen (15 g, 100213906368, Beijing Tongrentang), Corydalis Rhizoma (15 g, 100289087328, Suzetang), and Anemarrhenae Rhizoma (15 g, 100216346447, Suzetang). Following aqueous decoction and vacuum concentration, a stock solution of 6 g crude herb/mL was produced. The rat equivalent dose was calculated as 45.56 g crude herb/kg (7.59 mL/kg) based on body surface area conversion (human-to-rat factor: 6.3, standardized for 60 kg adults). Mineral components, including Magnetitum, Chloriti Lapis, Margaritifera Concha, and Ostreae Concha, predominantly insoluble mineral complexes, contributed minimally to the soluble constituents during decoction.
Open Field Test
Locomotor activity was assessed using an open field apparatus. Each rat was placed in the center of the arena and allowed to explore freely for 5 min. Total distance traveled and vertical rearing frequency were recorded and analyzed by an investigator blinded to the treatment groups.
Pentobarbital-Induced Sleep Test
The pentobarbital-induced sleep assay was performed according to established protocols27 by an experimenter blinded to the group allocation. Each rat received an intraperitoneal injection of sodium pentobarbital (42 mg/kg). Sleep latency (defined as the time interval from injection to loss of righting reflex) and sleep duration (measured as the period between righting reflex disappearance and recovery) were recorded in seconds. Rats that failed to attain a sleep state within 15 min post-injection were excluded from the subsequent analysis. No animals were excluded due to death or other complications during the experimental period.
Hematoxylin and Eosin (HE) Staining
The post-fixed brain tissues were immersed in 4% paraformaldehyde (158127; Sigma-Aldrich) for 24 h. After fixation, the specimens were subjected to sequential dehydration, clearing, and paraffin embedding prior to sectioning. Deparaffinization was achieved by incubating the sections in xylene (534056; Sigma-Aldrich) for 5 min followed by rehydration with ethanol (E7023; Sigma-Aldrich). Nuclei were stained with hematoxylin solution (HHS16, Sigma-Aldrich) for 5 min, differentiated in 1% acid alcohol, and blued in 0.2% ammonia water (1 min). Cytoplasmic staining was performed using eosin solution (HT110132; Sigma-Aldrich) for 60s, after which the sections were dehydrated using ethanol. Finally, cleared sections were mounted using Organo/Limonene Mount™ medium (O8015; Sigma-Aldrich) and examined under a Nikon Eclipse E200 microscope (Nikon Corporation, Tokyo, Japan).
Nissl Staining
The brain tissue sections were fixed in 4% paraformaldehyde at room temperature for 15 min. Subsequently, the sections were stained with Nissl staining solution (C0117; Beyotime Biotechnology, Shanghai, China) for 5 min to visualize the neuronal Nissl substance. After mounting, a histological evaluation was performed using a Nikon Eclipse E200 microscope.
Western Blot (WB) Analysis
Brain tissue homogenates were digested with trypsin and lysed in radioimmunoprecipitation assay (RIPA) buffer (R0278, Sigma-Aldrich) on ice for 30 min, with vortexing every 5 min. Lysates were centrifuged at 12,000 rpm for 10 min at 4°C, and the supernatants were collected. Protein concentration was quantified using a BCA assay kit (232207; Thermo Fisher Scientific, Waltham, MA, USA). Equal amounts of protein were electrophoretically separated on sodium dodecyl sulfate-polyacrylamide gels and subsequently transferred onto PVDF membranes (88518; Thermo Fisher Scientific). After blocking with 5% non-fat milk for 1 h, the membranes were incubated overnight at 4°C with the following primary antibodies: serotonin receptor 1A (5-HT1A; 1:1,000, Ab227165, Abcam, Cambridge, MA, USA), GABA A receptor (GABAA-R; 1:2,000, DF8548, Affinity Biosciences, Jiangsu, China), TNF-α (1:1,000, 17590-1-AP, Proteintech, Wuhan, China), TNF receptor 1 (TNFR1; 1:1,000, PA5-95585, Invitrogen, Waltham, MA, USA), phosphorylated NF-κB p65 (p-p65; 1:1,000, AF2006, Affinity Biosciences), total NF-κB p65 (p65; 1:1,000, AF5006, Affinity Biosciences), and glyceraldehyde-3-phosphate dehydrogenase (GAPDH; 1:5,000, AF7021, Affinity Biosciences). Membranes were then probed with horseradish peroxidase-conjugated goat anti-rabbit secondary antibody (1:10,000, 31460, Thermo Fisher Scientific) in 5% non-fat milk for 1 h at room temperature. Protein bands were visualized using an enhanced chemiluminescence substrate (32106, Thermo Fisher Scientific), and band densitometry was performed using ImageJ software and normalized to GAPDH expression.
Biochemical Analysis
The serum levels of IL-1β, IL-6, and TNF-α in rats were quantified in duplicate using commercial enzyme-linked immunosorbent assay (ELISA) kits according to the manufacturer’s protocols: IL-1β (900-K91K, Invitrogen), IL-6 (ERA31RB, Invitrogen), and TNF-α (SRTA00, Bio-Techne, Minnesota, USA).
Statistical Analysis
All statistical analyses were performed using GraphPad Prism software (Version 9.0.0, GraphPad Inc., San Diego, CA, USA), and continuous data were expressed as mean ± standard deviation (SD). Data were first tested for normality using the Shapiro–Wilk test and for homogeneity of variance using Levene’s test before performing parametric analyses. Intergroup comparisons between two cohorts were assessed using the t-test, while one-way or two-way analysis of variance (ANOVA) was applied for multi-group analyses followed by Tukey’s post hoc test. Statistical significance was defined as P < 0.05.
Results
Identification of Bioactive Components and Target Prediction for GAD
Pharmacochemical profiling of GAD was conducted using TCMSP.29 Based on the screening criteria of OB ≥ 30% and DL ≥ 0.18, 226 bioactive compounds were identified across 10 constituent herbs: Lilii Bulbus (6 compounds), Ganoderma (61), Coptidis Rhizoma (14), Ziziphi Spinosae Semen (9), Chuanxiong Rhizoma (7), Schisandrae Chinensis Fructus (8), Salviae Miltiorrhizae Radix et Rhizoma (65), Platycladi Semen (5), Corydalis Rhizoma (49), and Anemarrhenae Rhizoma (15). Deduplication yielded 221 compounds in total.
These compounds were mapped to 2,739 putative targets with herb-specific contributions as follows: Lilii Bulbus (84 targets), Ganoderma (54), Coptidis Rhizoma (250), Ziziphi Spinosae Semen (46), Chuanxiong Rhizoma (42), Schisandrae Chinensis Fructus (30), Salviae Miltiorrhizae Radix et Rhizoma (931), Platycladi Semen (43), Corydalis Rhizoma (1103), and Anemarrhenae Rhizoma (182). Target deduplication consolidated 263 unique biological targets. To construct the herb-compound-target-disease network, these targets were further intersected with INS-related DEGs from the GEO dataset. Only herbs whose targets showed direct overlap with the INS-related gene set were retained for network visualization. Consequently, nine herbs were included in the final network: Lilii Bulbus, Ganoderma, Ziziphi Spinosae Semen, Chuanxiong Rhizoma, Schisandrae Chinensis Fructus, Salviae Miltiorrhizae Radix et Rhizoma, Platycladi Semen, Corydalis Rhizoma, and Anemarrhenae Rhizoma. Coptidis Rhizoma, despite its compound and target richness, was excluded at this stage because its predicted targets showed no significant overlap with the INS-related DEGs identified in this study. A herb-compound-target interaction network was constructed using Cytoscape 3.10.2, comprising 434 nodes and 2,614 edges (Figure 1).
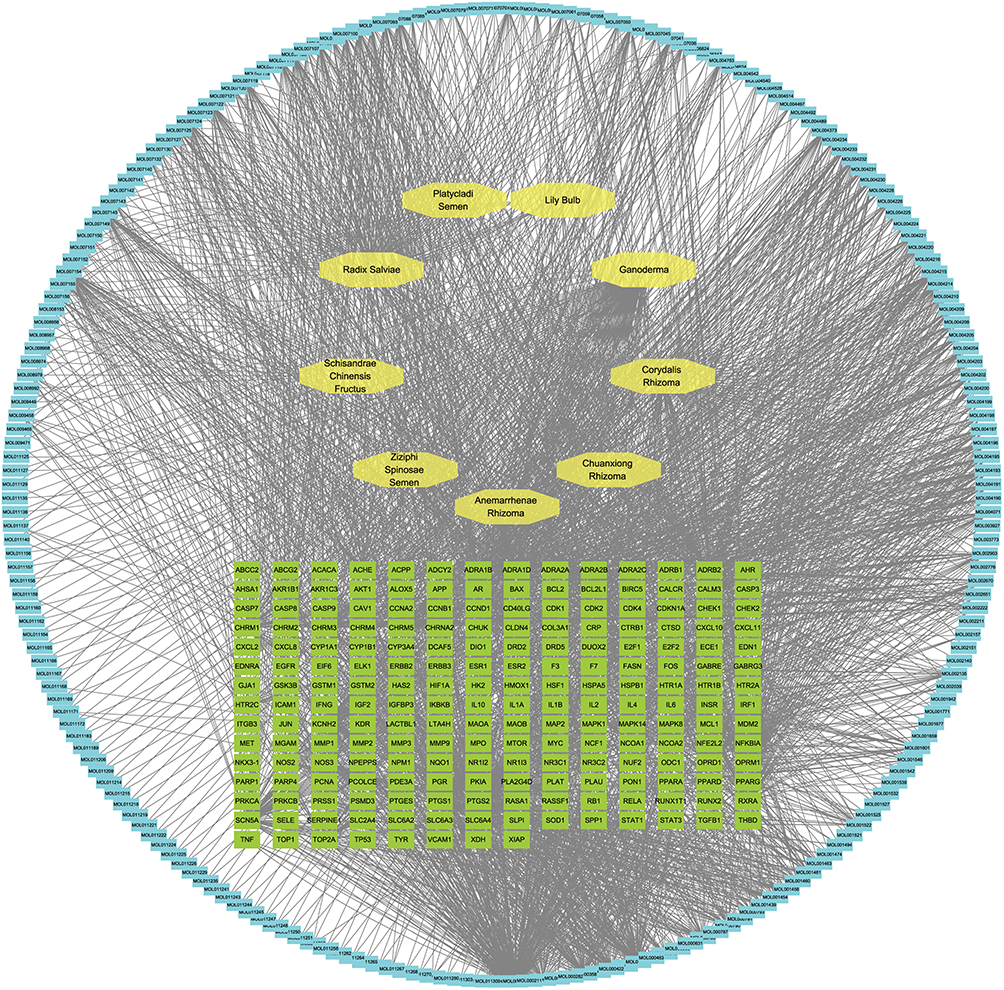 |
Figure 1 Herb-compound-target interaction network of GAD. |
Functional Enrichment Analysis of INS Therapeutic Targets Modulated by GAD
DEGs (P < 0.05) were obtained from the GEO dataset GSE208668 in PBMCs of patients with and without INS and intersected with the downstream targets of GAD compounds (Figure 2A and B). To elucidate the molecular mechanisms underlying INS-related genes, KEGG pathway and GO functional enrichment analyses were performed on the screened DEGs. The results revealed significant enrichment of immune-inflammatory and cellular stress response pathways. KEGG analysis demonstrated high enrichment of INS drug targets in inflammatory and stress-related signaling pathways, most prominently in TNF signaling (hsa04668; enrichment factor = 7.73), advanced glycation end-product (AGE)-receptor for AGE (RAGE) signaling (hsa04933; 6.88), and C-type lectin receptor pathways (hsa04625; 6.77) (Figure 2C). These pathways involve inflammatory mediators (eg., IL-6 and IL-1β), transcription factors (JUN and FOS), and downstream regulatory cascades, suggesting that INS pathogenesis involves TNF- and RAGE-associated neuroinflammation and metabolic stress that disrupt central neural function.
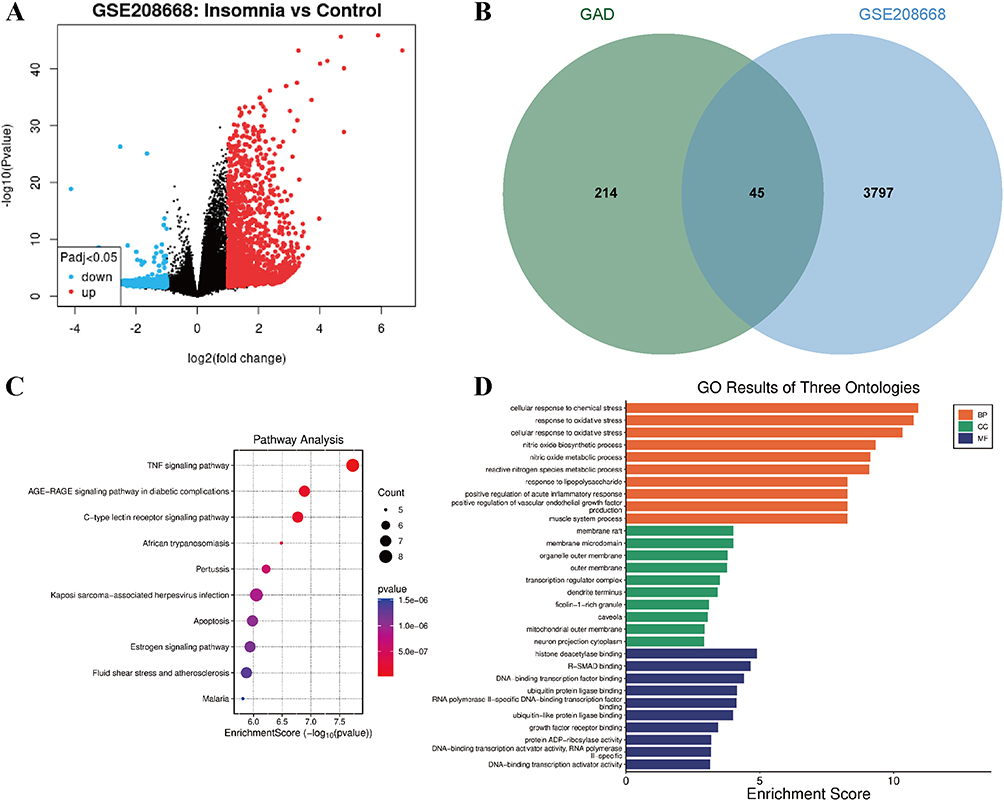 |
Figure 2 Profiling of INS-associated pathways and functional enrichment modulated by GAD. (A) Volcano plot of INS-associated DEGs from the GSE208668 dataset. (B) Venn diagram illustrating the intersection between GAD compound targets and INS-related DEGs. (C) Bubble plot of KEGG pathway enrichment analysis. (D) Bar graph of GO functional enrichment terms. |
Additionally, the estrogen signaling and apoptosis pathways were significantly enriched, indicating the potential roles of sex hormones and neuronal survival in INS regulation. GO biological process (BP) analysis further corroborated these findings, with DEG-drug target intersections enriched in “cellular response to chemical stress” (adjusted P = 2.16 × 10−8), “response to oxidative stress” (adjusted P = 2.16 × 10−8), “nitric oxide biosynthetic/metabolic process”, and “regulation of acute inflammatory response”. These results imply that oxidative damage and inflammatory activation in INS compromise neural function. GO cellular component (CC) analysis highlighted enrichment in membrane rafts, mitochondrial outer membranes, and transcriptional regulatory complexes, suggesting that neural membranes and mitochondrial dysfunction contribute to INS pathogenesis. Molecular function (MF) enrichment emphasized the regulatory roles of membrane microdomain proteins and the signal transduction machinery (Figure 2D). Collectively, functional enrichment profiling revealed inflammation, oxidative stress, apoptosis, and metabolic dysregulation as the core BPs in INS pathophysiology.
Identification of Core Therapeutic Targets for GAD in INS Management
To delineate the core targets mediating GAD’s anti-INS effects, a herb-compound-disease interaction network was constructed and analyzed using the cytoHubba plugin in Cytoscape. The top three bioactive compounds were MOL000098 (quercetin), MOL000358 (β-sitosterol), and MOL000449 (stigmasterol) (Figure 3A). PPI networks of GAD-INS intersecting targets were generated using the STRING database and visualized using Cytoscape 3.10.2 (Figure 3B). CytoHubba identified the top 10 hub targets using the degree algorithm (IL-6, IL-1β, HIF1A, JUN, PTGS2, PTEN, HSP90AA1, FOS, CCND1, and IL-10) and the Maximal Clique Centrality (MCC) algorithm (JUN, HIF1A, IL-6, IL-1β, PTEN, PTGS2, IL-10, FOS, CXCL8, and CCND1) (Figure 3C and D). Deduplication yielded 13 core targets, all of which were significantly enriched in the top 10 KEGG pathways (Figure 3E). Notably, five targets, IL-6, IL-1β, JUN, PTGS2, and FOS, demonstrated specific enrichment in the TNF signaling pathway (Figure 3F).
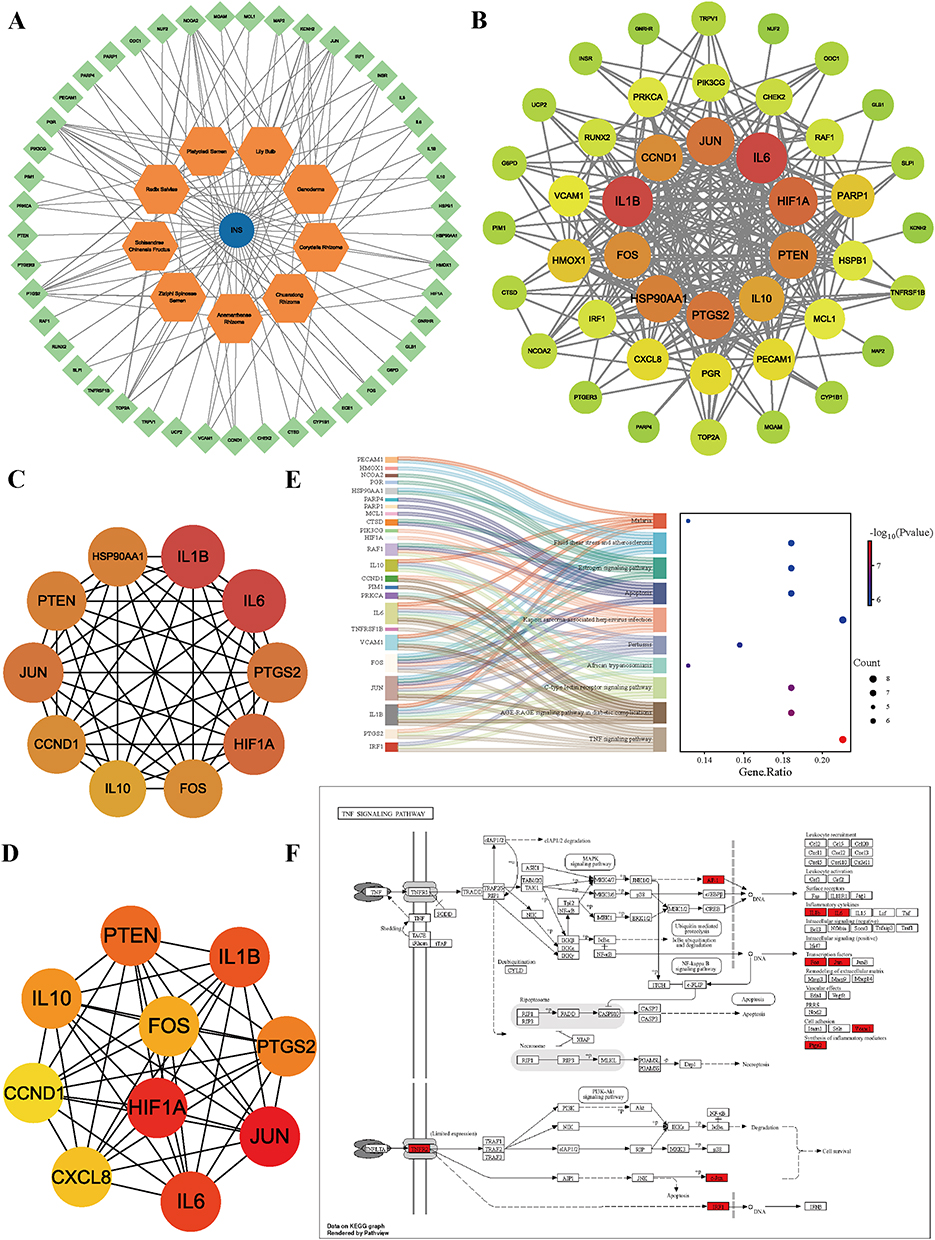 |
Figure 3 Mapping of core therapeutic targets and pathway enrichment for GAD in INS treatment. (A) Herb-compound-disease interaction network. (B) Optimized PPI network of GAD-INS intersecting targets. (C) Degree-based ranking of top 10 hub targets. (D) MCC-based ranking of top 10 hub targets. (E) Enrichment of core targets in the AGE-RAGE signaling pathway. (F) Sankey diagram illustrating the mapping of core targets to the top 10 enriched KEGG pathways. |
GAD Ameliorates Behavioral and Neuropathological Manifestations in INS Model Rats
To validate the therapeutic efficacy of GAD, an INS rat model was established and subjected to a multidimensional assessment. Behavioral analyses revealed that the INS and INS + saline groups exhibited significantly increased vertical rearing frequency and total locomotion in the open field test compared to the controls, indicating hyperarousal and locomotor hyperactivity consistent with successful modeling. GAD intervention substantially reduced both vertical and overall activities in the INS + GAD cohort compared to the INS group, demonstrating pronounced anxiolytic effects (Figure 4A and B). In pentobarbital-induced sleep tests, the INS and INS + saline groups showed prolonged sleep latency and shortened sleep duration compared with the controls, confirming sleep architecture disruption. GAD treatment significantly shortened latency and extended total sleep duration (vs. INS), indicating improved sleep quality (Figure 4C and D).
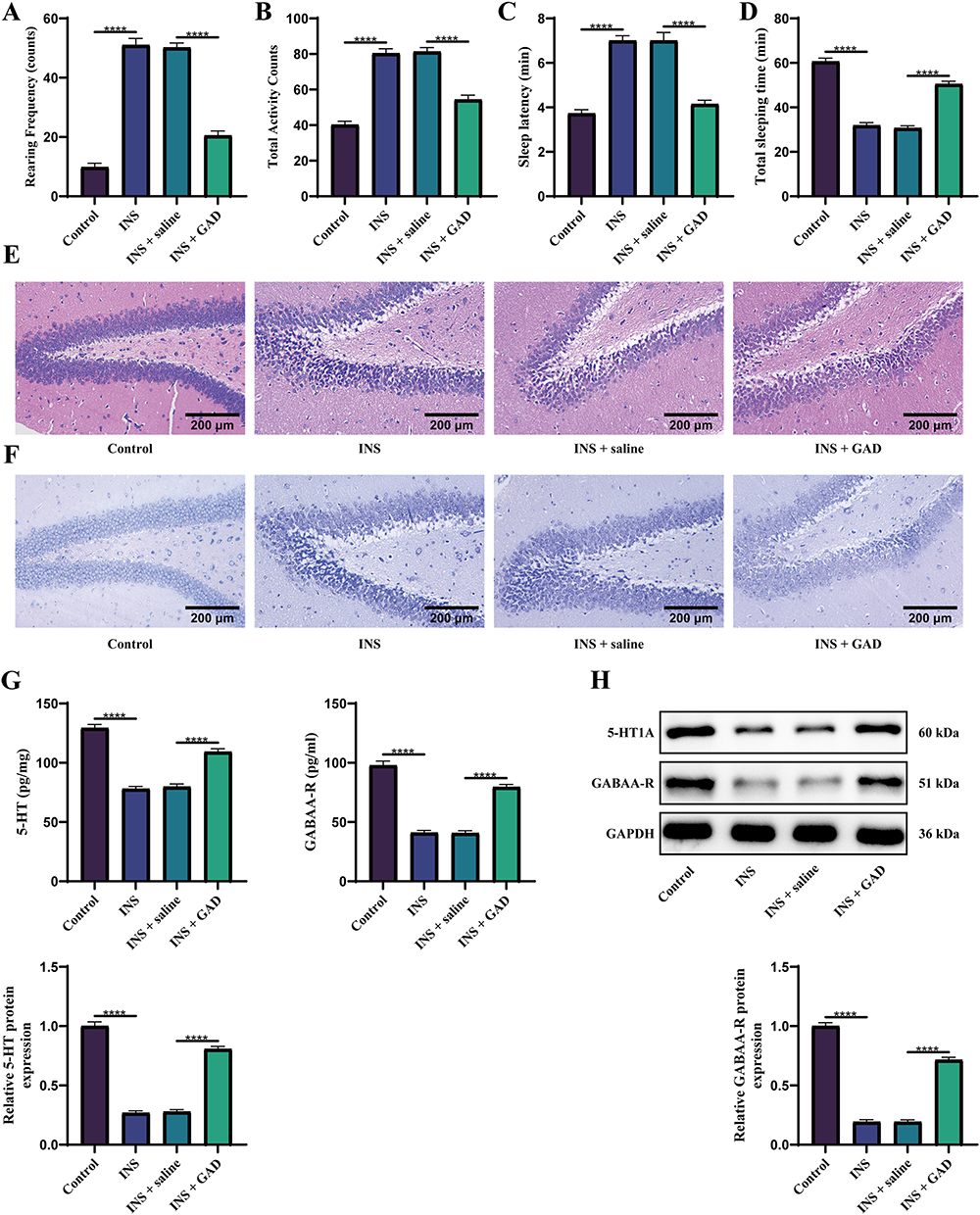 |
Figure 4 GAD reverses behavioral and neuropathological deficits in the INS rat model. (A and B) Open field test. (C and D) Pentobarbital-induced sleep test. (E) HE staining of the hippocampal region. (F) Nissl staining of neurons. (G) ELISA for hippocampal neurotransmitter levels (5-HT, GABA). (H) WB for protein expression of 5-HT1A receptor and GABAA-R. (n = 6). ****P < 0.0001. For three or more groups, one-way ANOVA was applied, followed by Tukey’s post hoc test. |
The histopathological evaluation corroborated these findings. HE staining demonstrated that in the INS and INS + saline groups, hippocampal neurons were disarranged with increased staining and exhibited varying degrees of atrophy, whereas GAD restored near-normal cytoarchitecture (Figure 4E). Nissl staining revealed neuronal loss in the INS group accompanied by nuclear condensation and fragmentation. After GAD intervention, neuronal number was partially restored, and cellular morphology significantly improved (Figure 4F). Neurotransmitter profiling elucidated GAD’s mechanisms. ELISA detected reduced hippocampal serotonin (5-hydroxytryptamine, 5-HT) and GABA levels in the INS models, which were reversed by GAD treatment (Figure 4G). WB confirmed the downregulation of 5-HT1A and GABAA-R receptors in the INS group, with GAD normalizing their expression (Figure 4H). Collectively, GAD mitigates INS-associated behavioral deficits, attenuates hippocampal neurodegeneration, and exerts neuroprotective sedative effects through monoaminergic and GABAergic modulation.
GAD Exerts Anti-Inflammatory and Anti-INS Effects via TNF Pathway Modulation
To elucidate the mechanisms underlying GAD’s efficacy, serum inflammatory cytokine levels and hippocampal TNF-α/NF-κB signaling were assessed. ELISA revealed significantly elevated serum levels of TNF-α, IL-1β, and IL-6 in the INS and INS + saline groups compared to the controls, indicating systemic inflammation in INS. The GAD intervention substantially reduced the levels of these cytokines (vs. INS), demonstrating potent systemic anti-inflammatory activity (Figure 5A). WB analysis further confirmed the upregulated hippocampal expression of TNF-α, TNFR1, and p-p65 in the INS models, confirming TNF pathway hyperactivation. GAD treatment significantly downregulated these proteins (vs. INS), indicating suppression of TNF-driven neuroinflammation (Figure 5B). Immunofluorescence staining further confirmed that GAD inhibited p-p65 nuclear translocation in hippocampal neurons (Figure 5C). Collectively, GAD may exert anti-inflammatory effects and improve INS by inhibiting TNF pathway activation.
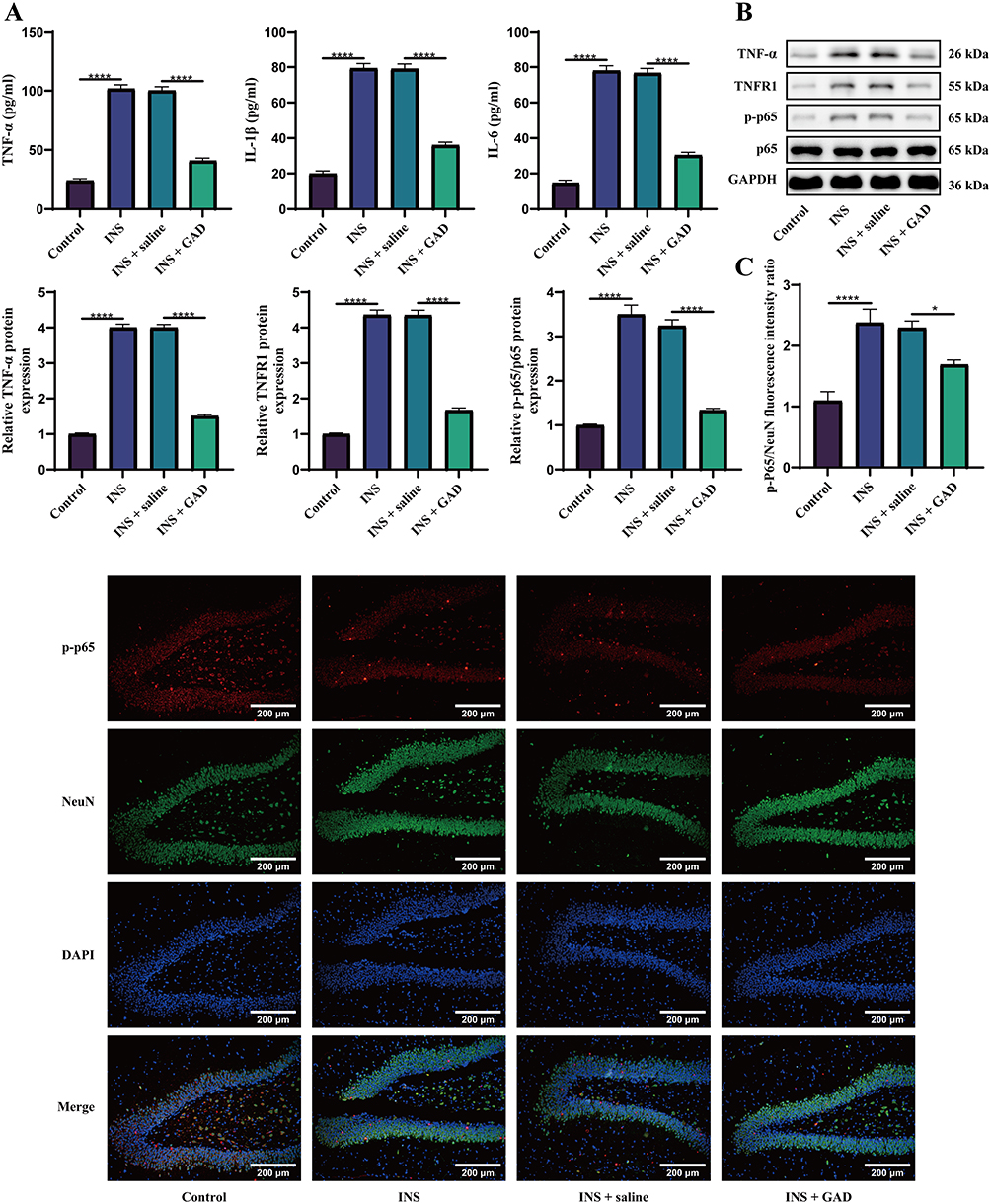 |
Figure 5 GAD ameliorates INS via TNF pathway inhibition. (A) ELISA for serum levels of TNF-α, IL-1β, and IL-6. (B) WB analysis for hippocampal protein expression of TNF-α, TNFR1, and p-p65. (C) Immunofluorescence staining of p-p65 (red) and NeuN (green) in the hippocampus. Nuclei were stained with DAPI (blue). (n = 6). *P < 0.05, ****P < 0.0001. For three or more groups, one-way ANOVA was applied, followed by Tukey’s post hoc test. |
Molecular Docking Validation
Molecular docking simulations were performed between the top three bioactive compounds (quercetin, stigmasterol, and β-sitosterol) and 11 core targets (IL-6, IL-1β, HIF1A, JUN, PTGS2, PTEN, HSP90AA1, FOS, CCND1, IL-10, and CXCL8) identified through network pharmacology. Following intersection analysis of compound-specific and core disease targets (Figure 6A), binding affinities were quantified using binding energy calculations. Per established criteria,30 binding energy < −4.25 kcal/mol indicated moderate binding; < −5.0 kcal/mol signified high affinity; < −7.0 kcal/mol denoted robust binding. The docking heatmap revealed that all 11 compound-target pairs exhibited binding energy < −4.25 kcal/mol, with 2 pairs achieving binding energy < −7.0 kcal/mol (Figure 6B). The spatial configurations of the seven pairs that formed characteristic hydrogen bonds were visualized (Figure S1), demonstrating strong binding complementarity between GAD’s bioactive constituents and INS-associated targets.
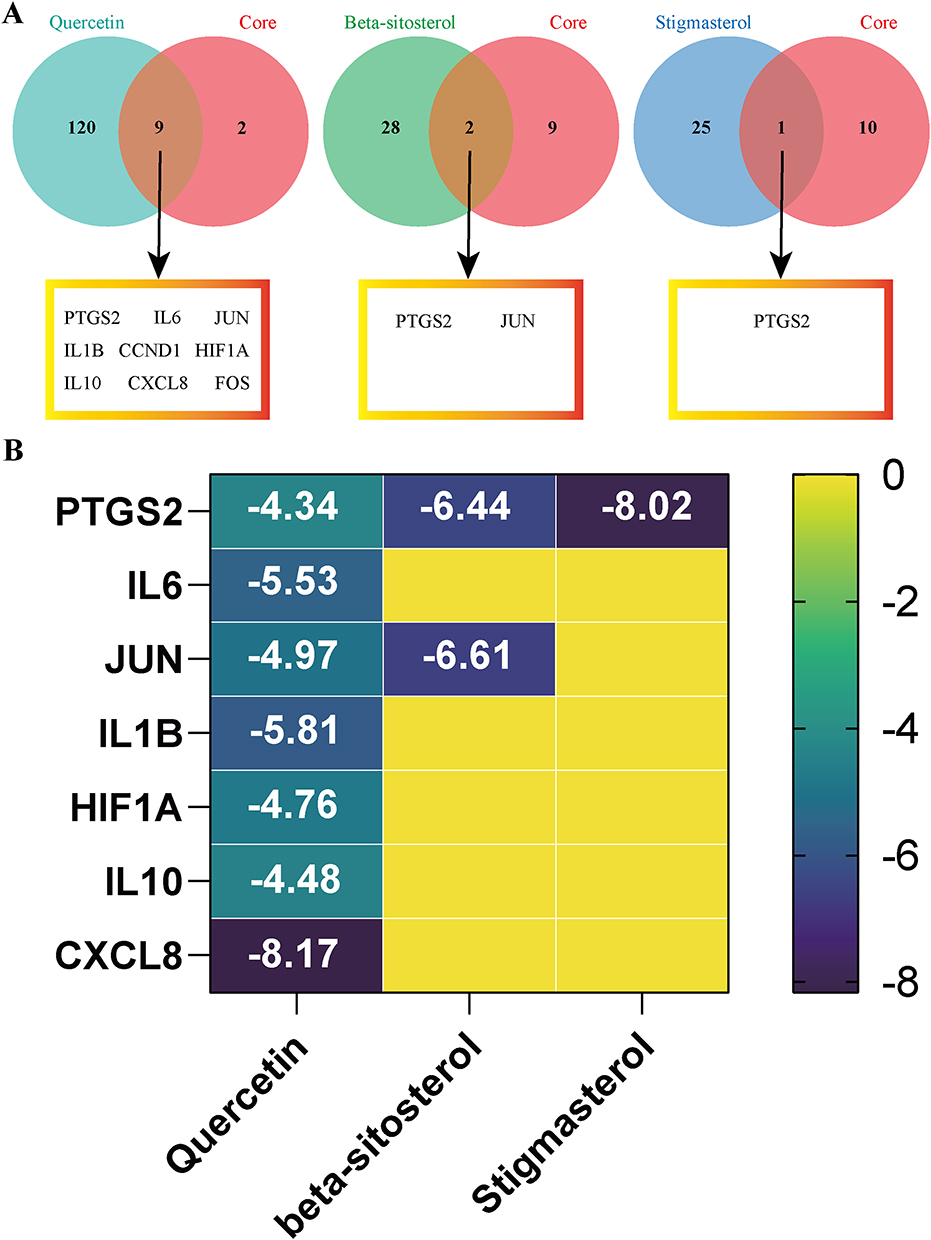 |
Figure 6 Molecular docking validates high-affinity binding of GAD compounds to core INS targets. (A) Venn diagrams illustrating the intersections between targets of the top three bioactive compounds (quercetin, β-sitosterol, stigmasterol) and core INS-associated targets. (B) Binding affinity heatmap. |
Discussion
Despite the empirical use of GAD in clinical INS management, its mechanism of action remains unclear, particularly regarding neuroinflammatory modulation, which limits its standardization and broader acceptance. To address this knowledge gap, this study systematically deciphered the therapeutic mechanisms of GAD in INS through integrated network pharmacology and experimental validation, demonstrating that GAD ameliorates sleep disturbances by suppressing TNF signaling-mediated neuroinflammation. Our findings provide empirical support for GAD’s therapeutic potential in INS and suggest novel anti-inflammatory strategies that may inform future INS management.
In this study, we employed a network pharmacology approach to systematically predict the potential targets and pathways of GAD, identifying the TNF signaling pathway as a critical axis. This bioinformatics-driven hypothesis was then experimentally validated using a PCPA-induced INS rat model. The rationale for investigating the anti-inflammatory effects of GAD is supported by two lines of evidence: first, several constituent herbs, including Coptidis Rhizoma, Salviae Miltiorrhizae Radix, and Ganoderma, have documented anti-inflammatory and immunomodulatory properties;12,14,16 second, a growing body of literature has established the pivotal role of neuroinflammation, particularly TNF signaling, in INS pathogenesis.19,20 Thus, our study design follows a hypothesis-generating (network pharmacology) and hypothesis-testing (animal experiment) paradigm, which is widely adopted in modern TCM mechanism research.
The therapeutic efficacy of herbal formulas stems from the multi-component synergies that target biological networks.31 GAD, a 19-botanical TCM formula including Coptidis Rhizoma, has been empirically used in INS management. Critically, Coptidis Rhizoma, recognized for purging Xin (heart) fire in TCM theory, constitutes a pharmacological principal component of anti-INS formulas.32 We identified 221 bioactive compounds and 263 potential targets, constructing a herb compound-target network reflecting its polypharmacological complexity. Core constituents, such as 3-demethylcolchicine, stigmasterol, and β-sitosterol, exhibit documented anti-inflammatory and neuroprotective properties,33 aligning with GAD’s pharmacodynamic basis. Moreover, our PPI and enrichment analyses revealed significant convergence of GAD targets and INS-associated DEGs in inflammatory pathways, particularly TNF and AGE-RAGE signaling, with hub targets such as IL-6 and IL-1β modulating TNF cascades. This finding implicates inflammatory network regulation as central to GAD’s anti-INS efficacy, positioning TNF signaling as a pivotal node. Such mechanistic insights corroborate established bidirectional relationships between sleep and inflammatory cytokines (eg., IL-6, TNF), where circadian rhythms modulate cytokine production, and in return, some cytokines induce sleep.20 The network pharmacology predictions not only provide a theoretical foundation for subsequent experimental validation but also exemplify how systems biology bridges TCM complexity and mechanistic clarity. Prior evidence also supports this approach, showing that Shuangren-Anshen capsules, a structurally analogous formula, exerted sedative-hypnotic effects in mouse models via similar pharmacology.34
It is important to note that the TNF pathway does not operate in isolation but engages in extensive crosstalk with other inflammatory signaling cascades. Among these, the AGE-RAGE signaling pathway emerged as significantly enriched in our analysis, suggesting potential synergistic interactions with TNF-mediated neuroinflammation in INS pathogenesis. AGE-RAGE signaling is known to amplify inflammatory responses through NF-κB activation and promote oxidative stress, both contributing to neuronal dysfunction and sleep disruption.35 Notably, RAGE activation can sustain TNF-α production, creating a positive feedback loop that exacerbates neuroinflammation.36 The convergence of GAD targets on both TNF and AGE-RAGE pathways implies that GAD may exert broader anti-inflammatory effects by interrupting this crosstalk, thereby providing more comprehensive neuroprotection. Although our experimental validation focused on the TNF axis, these findings open avenues for future investigations into whether GAD modulates AGE-RAGE signaling components, such as RAGE, high mobility group box 1, or downstream oxidative stress markers, to potentiate its therapeutic effects. Additionally, while estrogen signaling was not a primary focus of this study, given its known modulatory role in neuroinflammation and higher INS prevalence in perimenopausal women,37 exploring potential interactions between GAD, estrogen-related pathways, and inflammatory cascades represents a promising direction for future research. Such investigations could further elucidate GAD’s multi-pathway regulatory capacity and support its application in diverse INS populations.
The PCPA model, which depletes cerebral serotonin (5-HT) and GABA levels, remains the gold standard for INS induction.38 In our study, GAD intervention significantly improved sleep behavior in rats with PCPA-induced INS. Reduced vertical rearing and locomotion in open-field tests confirmed sedative effects, whereas shortened sleep latency and prolonged duration in pentobarbital tests demonstrated direct hypnotic potentiation. These outcomes are consistent with previous reports on Anshen-class formulas,8,39 supporting GAD’s therapeutic potential in INS. Histopathological analyses further revealed GAD’s neuroprotective effects, manifested as attenuated hippocampal neuronal disorganization, reduced atrophy, and increased Nissl body density. Given that the hippocampus governs sleep-wake regulation,40 its structural preservation likely underpins GAD’s pro-somnolence effects. In addition, GAD upregulated hippocampal 5-HT/GABA levels and receptor expression (5-HT1A, GABAA-R), consistent with neurotransmitter modulation by constituent herbs such as Ziziphi Spinosae Semen,9 further demonstrating its multi-target synergy. Analogous mechanisms operate in structurally related formulas. Liuwei Anshen capsule, a clinically prescribed INS remedy, enhances spatial learning and memory in sleep-deprived rats while suppressing hippocampal apoptosis, microglial activation, and pro-inflammatory cytokines (TNF-α/IL-1β/IL-6).41
Aberrant activation of inflammatory responses is a core pathogenic mechanism underlying INS occurrence and progression.42 In this study, we found that the levels of pro-inflammatory cytokines such as TNF-α, IL-1β, and IL-6 in the serum and hippocampus of INS model rats were significantly increased, accompanied by upregulated expression of TNFR1 and p-p65, key molecules in the downstream NF-κB pathway.43 These findings suggest that excessive TNF pathway activation plays an important role in experimental INS. GAD treatment significantly reduced these inflammatory mediators and suppressed TNFR1/p-p65 expression, confirming inhibition of the TNF axis. As a pathway initiator, TNF-α binding to TNFR1 activates NF-κB, propagating inflammatory cascades that disrupt neuronal integrity and neurotransmitter equilibrium.44 Interruption of this cycle by GAD via TNF blockade likely achieves dual benefits: mitigating hippocampal neuroinflammation and restoring 5-HT/GABA function from cytokine interference.22 These findings align with the reported GABA mechanisms for INS amelioration, including GABAA-R modulation, neuroinflammation suppression, oxidative damage repair, and circadian melatonin regulation.45 Moreover, sleep disturbances are positively associated with TNF levels, suggesting that circadian dysregulation is intimately linked to increased immune-inflammatory activity.46 Collectively, this evidence establishes an “anti-inflammation-neuroprotection-sleep improvement” action chain for GAD and provides crucial experimental evidence for its mechanism of action. Furthermore, high-affinity binding between GAD compounds (such as 3-demethylcolchicine) and TNF pathway targets (such as IL-6 and IL-1β) supports direct target engagement, thus validating the network predictions at molecular resolution.
However, this study had several limitations. First, a single component or fraction playing a core role in GAD has not yet been identified, and further verification through component knockout or single-component experiments is required. Second, the INS-related target genes used for network pharmacology analysis were derived from PBMCs rather than central nervous system tissues. While peripheral blood transcriptomes have been shown to reflect neuroinflammatory processes in INS, as demonstrated by Piber et al,47 who found that INS is associated with increased production of pro-inflammatory cytokines (IL-6, TNF-α) and activation of NF-κB-related transcription factors in PBMCs, future studies employing brain tissue-specific datasets or single-cell sequencing approaches would provide more precise insights into CNS-specific mechanisms. Third, the molecular docking analysis was based on static binding energy calculations, which do not account for protein flexibility or dynamic conformational changes over time. While the predicted binding affinities provide useful preliminary insights, molecular dynamics simulations would be required to further validate interaction stability and to better understand the dynamic behavior of compound-protein complexes. Future studies should incorporate molecular dynamics simulations to refine and confirm these findings. Additionally, only some molecules of the TNF pathway were investigated, and more detailed downstream regulatory mechanisms, including crosstalk with other inflammatory pathways such as AGE-RAGE signaling and potential interactions with estrogen-related pathways, require further study. Moreover, animal experiment results need further verification in clinical studies to clarify the efficacy and dose-dependent effects of GAD in humans.
Conclusion
This study demonstrates that GAD ameliorates INS in a rat model through polypharmacological suppression of the TNF pathway, attenuation of neuroinflammation, preservation of neuronal integrity, and restoration of neurotransmitter balance. These findings provide a mechanistic foundation for understanding GAD’s therapeutic effects in INS and establish a multi-compound-single pathway paradigm for TCM mechanism research. Future studies should prioritize lead compound identification, pathway interactome mapping, and randomized controlled trials to evaluate the translational potential of GAD for INS therapy.
Data Sharing Statement
The data that support the findings of this study are available from the corresponding author upon reasonable request.
Ethics Statement
This study was approved by the Animal Ethics Committee of Hunan Evidence-based Biotechnology Co. Ltd. (approval no. XZ250711). Animal experiments were conducted at Hunan Biotechnology Co due to lack of sterile animal facility in our institution. All procedures involving animals were conducted in compliance with the ARRIVE guidelines.
Author Contributions
Sanyuan Gu: Conceptualization; Data curation; Investigation; Writing – original draft. Jingyu Xu: Methodology; Formal analysis; Project administration; Writing – review and editing. All authors gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Disclosure
The authors declare no competing interests.
References
1. Morin CM, Jarrin DC. Epidemiology of Insomnia: prevalence, Course, Risk Factors, and Public Health Burden. Sleep Med Clin. 2022;17(2):173–17. doi:10.1016/j.jsmc.2022.03.003
2. Riemann D, Benz F, Dressle RJ, et al. Insomnia disorder: state of the science and challenges for the future. J Sleep Res. 2022;31(4):e13604. doi:10.1111/jsr.13604
3. Ferini-Strambi L. Insomnia disorder. Minerva Med. 2025;116(4):309–322. doi:10.23736/S0026-4806.25.09690-9
4. Riemann D, Espie CA, Altena E, et al. The European Insomnia Guideline: an update on the diagnosis and treatment of insomnia 2023. J Sleep Res. 2023;32(6):e14035. doi:10.1111/jsr.14035
5. Ye Z, Lai H, Ning J, et al. Traditional Chinese medicine for insomnia: recommendation mapping of the global clinical guidelines. J Ethnopharmacol. 2024;322:117601. doi:10.1016/j.jep.2023.117601
6. Zhang B, Wang Q, Zhang Y, et al. Treatment of Insomnia With Traditional Chinese Medicine Presents a Promising Prospect. Phytother Res. 2025. doi:10.1002/ptr.8495
7. Zhou F, Du L, Yu L, Zhang D, Jin H, Li Z. Efficacy and mechanism of Tiaoshu Anshen Decoction in treating insomnia with spleen and stomach Qi dysfunction: a retrospective study. Medicine. 2024;103(44):e40160. doi:10.1097/MD.0000000000040160
8. Li X, He C, Shen M, et al. Effects of aqueous extracts and volatile oils prepared from Huaxiang Anshen decoction on p-chlorophenylalanine-induced insomnia mice. J Ethnopharmacol. 2024;319(Pt 3):117331. doi:10.1016/j.jep.2023.117331
9. Sun M, Li M, Cui X, et al. Terpenoids derived from Semen Ziziphi Spinosae oil enhance sleep by modulating neurotransmitter signaling in mice. Heliyon. 2024;10(5):e26979. doi:10.1016/j.heliyon.2024.e26979
10. Jiang N, Wei S, Zhang Y, et al. Protective Effects and Mechanism of Radix Polygalae Against Neurological Diseases as Well as Effective Substance. Front Psychiatry. 2021;12:688703. doi:10.3389/fpsyt.2021.688703
11. Xie J, Li Y, Liang Y, Kui H, Wang C, Huang J. Integration of non-targeted metabolomics with network pharmacology deciphers the anxiolytic mechanisms of Platycladi Semen extracts in CUMS mice. J Ethnopharmacol. 2023;315:116571. doi:10.1016/j.jep.2023.116571
12. Yin C, Zhang M, Jin S, et al. Mechanism of Salvia miltiorrhiza Bunge extract to alleviate Chronic Sleep Deprivation-Induced cognitive dysfunction in rats. Phytomedicine. 2024;130:155725. doi:10.1016/j.phymed.2024.155725
13. Wang Y, Wu L, Wang H, et al. Ligusticum chuanxiong: a chemical, pharmacological and clinical review. Front Pharmacol. 2025;16:1523176. doi:10.3389/fphar.2025.1523176
14. Zhao Y, Ma Y, Xiong L, et al. Neuroprotective effect of the herbal pair Coptidis Rhizoma-Cinnamomi Cortex against 6-OHDA-induced Parkinson’s disease rats through promotion of autophagy via the PI3K/AKT/mTOR pathway. J Ethnopharmacol. 2026;355(Pt A):120582. doi:10.1016/j.jep.2025.120582
15. He B, Li Q, Jia Y, et al. A UFLC-MS/MS method for simultaneous quantitation of spinosin, mangiferin and ferulic acid in rat plasma: application to a comparative pharmacokinetic study in normal and insomnic rats. J Mass Spectrom. 2012;47(10):1333–1340. doi:10.1002/jms.3072
16. Zhao C, Zhang C, Xing Z, Ahmad Z, Li JS, Chang MW. Pharmacological effects of natural Ganoderma and its extracts on neurological diseases: a comprehensive review. Int J Biol Macromol. 2019;121:1160–1178. doi:10.1016/j.ijbiomac.2018.10.076
17. Park HR, Lee H, Cho WK, Ma JY. Pro-neurogenic effects of Lilii Bulbus on hippocampal neurogenesis and memory. Biomed Pharmacother. 2023;164:114951. doi:10.1016/j.biopha.2023.114951
18. Negah SS, Forouzanfar F. Dual Role of Fibroblast Growth Factor Pathways in Sleep Regulation. Endocr Metab Immune Disord Drug Targets. 2023;23(1):63–69. doi:10.2174/1871530322666220802161031
19. Zielinski MR, Gibbons AJ. Neuroinflammation, Sleep, and Circadian Rhythms. Front Cell Infect Microbiol. 2022;12:853096. doi:10.3389/fcimb.2022.853096
20. Veler H. Sleep and Inflammation: bidirectional Relationship. Sleep Med Clin. 2023;18(2):213–218. doi:10.1016/j.jsmc.2023.02.003
21. Zhang MM, Zhao JW, Li ZQ, Shao J, Gao XY. Acupuncture at Back-Shu point improves insomnia by reducing inflammation and inhibiting the ERK/NF-kappaB signaling pathway. World J Psychiatry. 2023;13(6):340–350. doi:10.5498/wjp.v13.i6.340
22. Varinthra P, Anwar S, Shih SC, Liu IY. The role of the GABAergic system on insomnia. Tzu Chi Med J. 2024;36(2):103–109. doi:10.4103/tcmj.tcmj_243_23
23. Lee M, Shin H, Park M, Kim A, Cha S, Lee H. Systems pharmacology approaches in herbal medicine research: a brief review. BMB Rep. 2022;55(9):417–428. doi:10.5483/BMBRep.2022.55.9.102
24. Du L, Yang D, Wu L, et al. Integration of Gut Microbiota, Serum Metabolomic, and Network Pharmacology to Reveal the Anti Insomnia Mechanism of Mongolian Medicine Sugemule-4 Decoction on Insomnia Model Rats. Drug Des Devel Ther. 2024;18:2617–2639. doi:10.2147/DDDT.S455600
25. Wang L, Qi X, Wang S, et al. Banxia-Yiyiren alleviates insomnia and anxiety by regulating the gut microbiota and metabolites of PCPA-induced insomnia model rats. Front Microbiol. 2024;15:1405566. doi:10.3389/fmicb.2024.1405566
26. Wang W, Wang Y, Guo Q, et al. Valerian essential oil for treating insomnia via the serotonergic synapse pathway. Front Nutr. 2022;9:927434. doi:10.3389/fnut.2022.927434
27. Kim S, Jo K, Hong KB, Han SH, Suh HJ. GABA and l-theanine mixture decreases sleep latency and improves NREM sleep. Pharm Biol. 2019;57(1):65–73. doi:10.1080/13880209.2018.1557698
28. Wang Z, Li D, Chen M, et al. A comprehensive study on the regulation of Compound Zaoren Granules on cAMP/CREB signaling pathway and metabolic disorder in CUMS-PCPA induced insomnia rats. J Ethnopharmacol. 2024;332:118401. doi:10.1016/j.jep.2024.118401
29. Tang L, Liu Y, Tao H, et al. Combination of Youhua Kuijie Prescription and sulfasalazine can alleviate experimental colitis via IL-6/JAK2/STAT3 pathway. Front Pharmacol. 2024;15:1437503. doi:10.3389/fphar.2024.1437503
30. Yu H, Wei X, Ding H, et al. Exploring the potential mechanisms of Mahuang Fuzi Xixin decoction in treating elderly bronchial asthma through network pharmacology, molecular docking, and molecular dynamics simulations. Medicine. 2024;103(41):e39921. doi:10.1097/MD.0000000000039921
31. Yang L, Wang H, Zhu Z, et al. Network Pharmacology-Driven Sustainability: AI and Multi-Omics Synergy for Drug Discovery in Traditional Chinese Medicine. Pharmaceuticals. 2025;18(7). doi:10.3390/ph18071074
32. Huang WY, Dong H. Coptidis Rhizoma-Contained Traditional Formulae for Insomnia: a Potential to Prevent Diabetes? Chin J Integr Med. 2018;24(10):785–788. doi:10.1007/s11655-018-3012-4
33. Aleem M. Phytochemistry and pharmacology of Celastrus paniculatus Wild.: a nootropic drug. J Complement Integr Med. 2023;20(1):24–46. doi:10.1515/jcim-2021-0251
34. Liu XX, Agudamu A, Zhang X, Wei Y, Song X, Li YL. Pharmacology of a traditional Chinese herb, Shuangren-Anshen capsule, in a hemorrhage mouse model by using an orthogonal array design. Genet Mol Res. 2016;15(3). doi:10.4238/gmr.15038806
35. Chen YC, Hsu PY, Su MC, et al. MicroRNA Sequencing Analysis in Obstructive Sleep Apnea and Depression: anti-Oxidant and MAOA-Inhibiting Effects of miR-15b-5p and miR-92b-3p through Targeting PTGS1-NF-kappaB-SP1 Signaling. Antioxidants. 2021;10(11). doi:10.3390/antiox10111854
36. Samur DN, Yildirim S, Maytalman E, Kalay M, Tanriover G, Ozbey G. Vortioxetine attenuates rotenone-induced enteric neuroinflammation via modulation of the TLR2/S100B/RAGE signaling pathway in a rat model of Parkinson’s disease. Neuropharmacology. 2025;271:110385. doi:10.1016/j.neuropharm.2025.110385
37. Zhang Y, Tan X, Tang C. Estrogen-immuno-neuromodulation disorders in menopausal depression. J Neuroinflammation. 2024;21(1):159. doi:10.1186/s12974-024-03152-1
38. Bao Y, Zhou H, Fu Y, Wang C, Huang Q. Zhumian Granules improves PCPA-induced insomnia by regulating the expression level of neurotransmitters and reducing neuronal apoptosis. J Ethnopharmacol. 2024;327:118048. doi:10.1016/j.jep.2024.118048
39. Zhang C, Wu Q, Tao X, et al. Sedative-hypnotic effects of Yiyin Anshen Granule on mice models of insomnia by regulating neurotransmitters, cytokines, and gut microbiota. J Pharm Biomed Anal. 2025;263:116949. doi:10.1016/j.jpba.2025.116949
40. Brodt S, Inostroza M, Niethard N, Born J. Sleep-A brain-state serving systems memory consolidation. Neuron. 2023;111(7):1050–1075. doi:10.1016/j.neuron.2023.03.005
41. Li LY, Liu SZ, Yu X, et al. Liuwei Anshen Capsule alleviates cognitive impairment induced by sleep deprivation by reducing neuroapoptosis and inflammation. J Ethnopharmacol. 2025;341:119311. doi:10.1016/j.jep.2024.119311
42. Liu L, Yang X, Yang C, et al. Associations between insomnia symptoms and inflammatory cytokines in adolescents with first-episode and recurrent major depressive disorder. J Affect Disord. 2024;350:110–117. doi:10.1016/j.jad.2024.01.031
43. Ding H, Li Y, Zhao C, et al. Rutin Supplementation Reduces Oxidative Stress, Inflammation and Apoptosis of Mammary Gland in Sheep During the Transition Period. Front Vet Sci. 2022;9:907299. doi:10.3389/fvets.2022.907299
44. Hung CF, Chiu WC, Chen JC, Chuang WC, Wang SJ. NRICM101 prevents kainic acid-induced seizures in rats by modulating neuroinflammation and the glutamatergic system. Int Immunopharmacol. 2024;140:112842. doi:10.1016/j.intimp.2024.112842
45. Zhu W, Huang L, Cheng H, et al. GABA and its receptors’ mechanisms in the treatment of insomnia. Heliyon. 2024;10(23):e40665. doi:10.1016/j.heliyon.2024.e40665
46. da Silveira Cruz-Machado S, Guissoni Campos LM, Fadini CC, Anderson G, Markus RP, Pinato L. Disrupted nocturnal melatonin in autism: association with tumor necrosis factor and sleep disturbances. J Pineal Res. 2021;70(3):e12715. doi:10.1111/jpi.12715
47. Piber D, Cho JH, Lee O, Lamkin DM, Olmstead R, Irwin MR. Sleep disturbance and activation of cellular and transcriptional mechanisms of inflammation in older adults. Brain Behav Immun. 2022;106:67–75. doi:10.1016/j.bbi.2022.08.004
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Siji Kangbingdu Mixture Ameliorates LPS-Induced Acute Lung Injury by Downregulating the TNF/IL-17 Signaling Pathways: A Comprehensive Study Using Transcriptomics, Network Pharmacology and Experimental Verification
Wang Y, Zhou H, Wang B, Liu Y, Long K, Zhang H
Journal of Inflammation Research 2025, 18:18267-18290
Published Date: 30 December 2025