Back to Journals » International Journal of Nanomedicine » Volume 21
Nanomaterials in Targeted Immunotherapy for Chronic Spontaneous Urticaria: Focusing on Immune Cell Modulation and Precision Treatment
Authors Yao W, Peng C, He Y, Yang G, Luo D, Zhang Z ![]() , Chen L
, Chen L ![]() , Huang X
, Huang X
Received 5 October 2025
Accepted for publication 31 December 2025
Published 13 January 2026 Volume 2026:21 572074
DOI https://doi.org/10.2147/IJN.S572074
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Professor Dong Wang
Wei Yao,1,2,* Chaoxia Peng,1,2,* Yufeng He,3 Guihong Yang,1,2 Dan Luo,1,2 Zhongsong Zhang,4,5 Long Chen,4– 6 Xianqiong Huang1,2
1Department of Dermatology, Daping Hospital, Army Medical University, Chongqing, 400042, People’s Republic of China; 2Research Center for Skin Tissue Engineering of Chongqing Higher Education Institutions, Daping Hospital, Army Medical University, Chongqing, 400042, People’s Republic of China; 3Department of Dermatology, 77th Group Army Hospital, Leshan, Sichuan, 614000, People’s Republic of China; 4The Second Affiliated Hospital of Chengdu Medical College, Nuclear Industry 416 Hospital, Chengdu, Sichuan, 610000, People’s Republic of China; 5Non-Coding RNA and Drug Discovery Key Laboratory of Sichuan Province, School of Basic Medical Sciences, Chengdu Medical College, Chengdu, Sichuan, 610550, People’s Republic of China; 6Department of Immunology, School of Basic Medical Sciences, Chengdu Medical College, Chengdu, Sichuan, 610500, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Long Chen, Email [email protected] Xianqiong Huang, Email [email protected]
Abstract: Chronic spontaneous urticaria (CSU) is an immune-mediated skin disorder marked by recurrent hives and angioedema, affecting a large proportion of the global population. Traditional immunotherapy often fails, particularly in difficult-to-treat cases, highlighting the need for more precise, immune cell-targeted treatments. Nanomaterials offer unique advantages, such as size tuning, surface functionalization, and controlled release, allowing precise targeting of immune cell receptors. This is crucial for enhancing treatment efficacy and minimizing systemic side effects. This review explores the potential of nanomaterials in CSU immunotherapy, focusing on their ability to modulate immune cells—such as mast cells, eosinophils, basophils, and T lymphocytes—to restore immune balance and alleviate symptoms. However, challenges remain in the clinical application of nanomaterials, including concerns about immunotoxicity, long-term safety, and the need for stronger evidence of efficacy in human trials. Additionally, while AI in optimizing nanomaterial design shows promise, it is still in early development and requires further exploration to enhance its practical applications. Current nanomaterial applications in allergic diseases, such as food allergies and asthma, show promise but differ from CSU in immune mechanisms like mast cell-eosinophil interactions and neuroimmune pathways. This approach is novel in that nanomaterials could provide a more precise, targeted solution for CSU, overcoming the limitations of current therapies. Future research should focus on optimizing nanomaterial formulations, integrating multi-omics analyses, and developing AI-driven strategies for personalized treatment. These advances could pave the way for safer, more effective immunotherapies in CSU and other immune-mediated diseases.
Keywords: chronic spontaneous urticaria, nanomaterials, immune cells, mast cells, drug delivery systems, immunotherapy
Introduction
Chronic spontaneous urticaria (CSU) is a prevalent immune-mediated skin disorder manifesting as recurrent wheals, angioedema, and intense pruritus, with a lifetime prevalence of 0.5–1%. It significantly impairs patients’ quality of life and imposes substantial economic and psychological burdens.1,2 Approximately 40–50% of patients respond inadequately to first-line H1-antihistamines, and even with second-line biologics such as omalizumab (an anti-IgE monoclonal antibody), nearly 30% remain refractory or experience relapse upon treatment discontinuation, highlighting an urgent unmet clinical need for more effective and durable therapies.3–7
In recent years, nanomaterials have emerged as a transformative platform in immunotherapy for allergic diseases, owing to their unique advantages, including tunable nanoscale size (<100 nm), high surface-to-volume ratio, versatile surface functionalization, targeted delivery, and controlled or stimuli-responsive cargo release.8–10 These properties enable precise modulation of immune cells by targeting specific receptors (eg, FcεRI, Siglec-8, MRGPRX2), inhibiting degranulation, reprogramming Th2-skewed responses, and inducing long-term immune tolerance with minimal systemic toxicity.11–13 Promising preclinical results have been reported in asthma and food allergy models, such as gold nanoparticle conjugates that block FcεRI crosslinking on mast cells and liposomal allergen-encapsulated carriers that promote regulatory T-cell induction.14,15 Despite these advances, the vast majority of nanomedicine research in allergy remains confined to asthma and food allergy, with only sporadic and fragmented studies addressing CSU.16,17
Crucially, CSU exhibits distinct pathophysiological features—notably autoimmunity-driven mast cell activation via IgG autoantibodies, MRGPRX2-mediated pseudo-allergic pathways, intense mast cell–eosinophil crosstalk, and a prominent neuroimmune axis—that differ substantially from the predominantly IgE/allergen-driven mechanisms in asthma and food allergy. These unique features demand CSU-specific nanomaterial designs capable of simultaneously inhibiting multiple activation pathways, disrupting pathogenic cell–cell interactions, and modulating neuroimmune signaling. To date, no comprehensive review has systematically evaluated the potential, challenges, and tailored design principles of nanomaterials for CSU immunotherapy. This review fills this critical gap by focusing exclusively on nanomaterial-based strategies for chronic spontaneous urticaria. We critically analyze the limited yet highly instructive CSU-specific studies, classify inorganic, organic, and carbon-based nanoplatforms according to their interactions with key effector cells (mast cells, eosinophils, basophils, and T lymphocytes), and highlight emerging concepts such as multivalent ligand presentation, synergistic pathway blockade, and AI-guided surface optimization. Furthermore, we address translational hurdles including immunotoxicity, long-term biocompatibility, and regulatory considerations. By synthesizing current evidence and proposing CSU-tailored nano-engineering roadmaps, this work aims to accelerate the development of next-generation precision nanotherapies for refractory CSU and provide a blueprint for nanomaterial applications in complex autoimmune/allergic skin diseases.
Immune Cells in CSU
In the pathogenesis of CSU, the complex interactions between key immune cells are mainly driven by the Th2 dominant pathway to sustain inflammatory responses. For example, mast cells act as central orchestrators, while eosinophils, basophils, and T lymphocytes contribute through mediator release, cellular crosstalk, and amplification of autoimmunity and inflammation.18,19 These cells not only interact with each other via cytokines and chemokines but also respond to autoantibodies and external stimuli, sustaining wheal formation, angioedema, and pruritus (Figure 1). Understanding their individual and collective roles is essential for identifying therapeutic targets. In the following sections, we detail each cell type’s mechanisms in CSU and exploring their potential targets as treatments for CSU (Table 1).
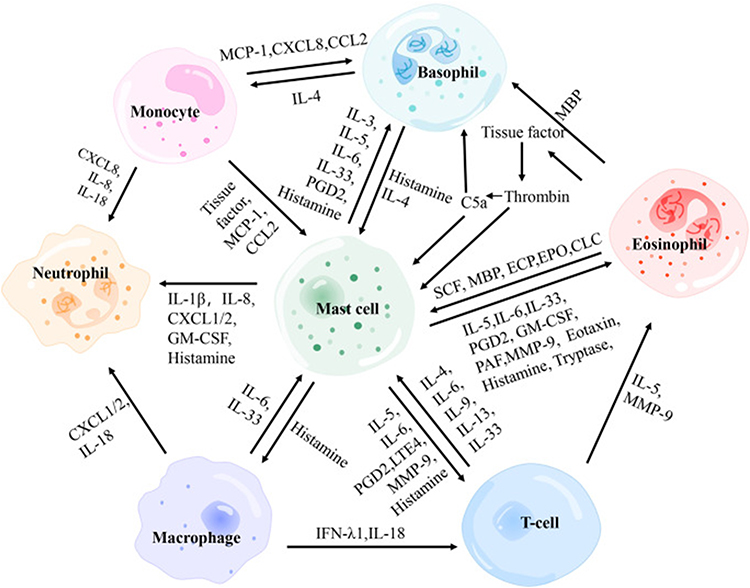 |
Figure 1 Interactions between immune cells in CSU. The diagram shows monocytes, basophils, eosinophils, mast cells, neutrophils, macrophages, and T cells communicating via cytokines and mediators (eg, IL-4, IL-5, IL-6, IL-33, histamine, PGD2). Arrows indicate signaling pathways and interactions contributing to urticaria pathophysiology. Adapted from Zhou, Bingjing et al. The Role of Crosstalk of Immune Cells in Pathogenesis of Chronic Spontaneous Urticaria. Frontiers in immunology vol. 13 879754. 31. Copyright © 2022 by authors.18 |
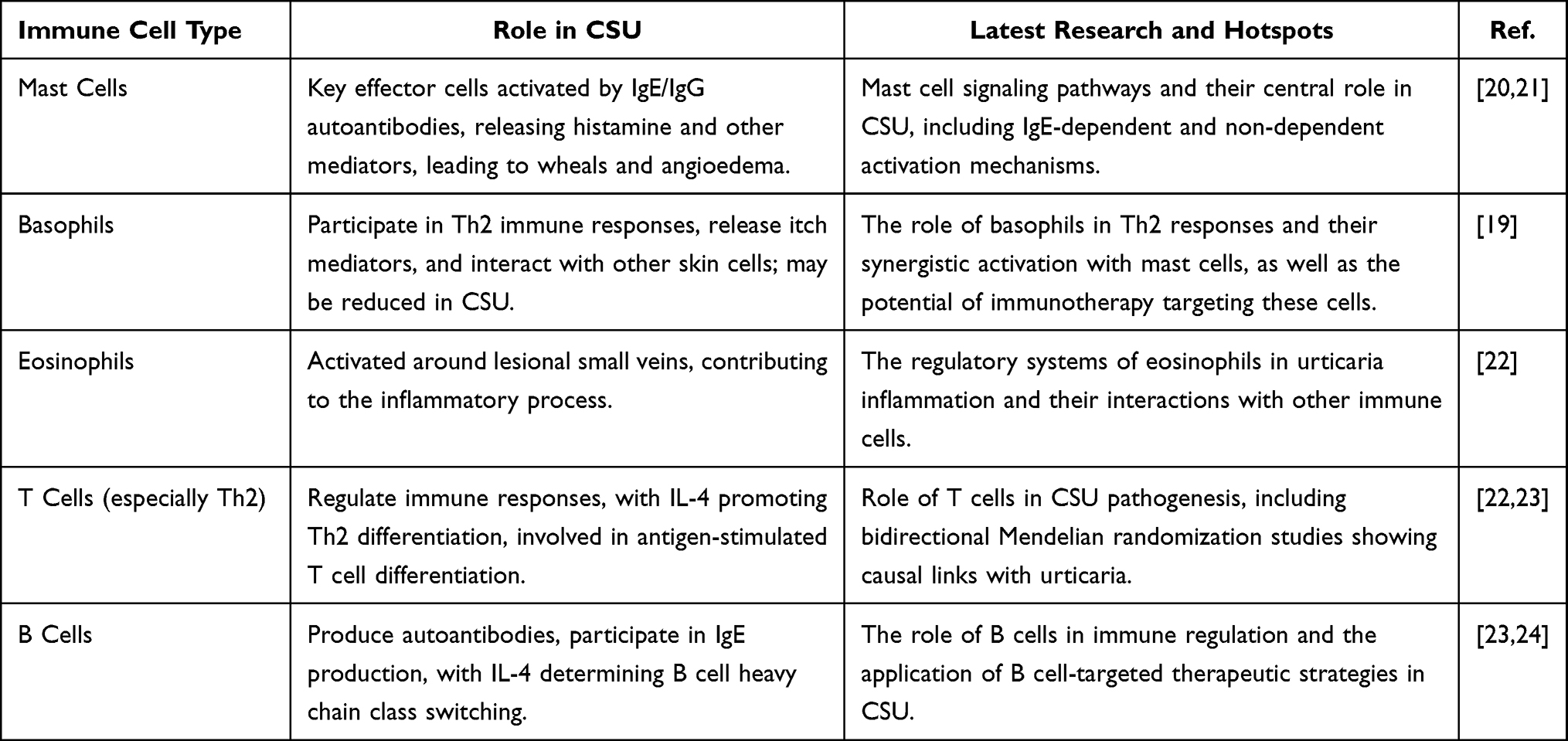 |
Table 1 Role of Immune Cells in CSU: Current Research and Insights |
Mast Cells
Mast cells are critical in the pathogenesis of CSU, acting as effector cells that drive the disease through activation and degranulation, releasing histamine and other mediators that cause wheals, angioedema, and pruritus.25,26 In CSU, mast cell activation occurs through IgE-dependent and IgE-independent mechanisms, with histopathology showing infiltrates of lymphocytes, eosinophils, neutrophils, basophils, and degranulated mast cells.19 In the IgE-dependent pathway (type I autoallergy), IgE autoantibodies target autoantigens like thyroid peroxidase, activating mast cells via FcεRI, triggering degranulation and mediator release.19 In type I endotype CSU, IgE activation amplifies inflammation, as seen in tests like the autologous serum skin test.19 In contrast, IgE-independent mechanisms dominate in the type IIb autoimmune endotype, where IgG autoantibodies bind to IgE or FcεRI, activating mast cells and leading to complement activation.5 In about 10% of type IIb CSU patients, autoantibodies contribute to mast cell degranulation, causing persistent symptoms unresponsive to antihistamines.11 Severe CSU may involve alternative pathways, such as thrombin-triggered protease-activated receptors and complement activation via C5a, further enhancing mast cell activity and vascular permeability.27,28 Additionally, MRGPRX2 receptors on mast cells respond to neuropeptides and host defense peptides, promoting non-immunologic degranulation, which is especially relevant in CSU unresponsive to omalizumab.25 A feedback loop, such as the PAF vicious circle, perpetuates mast cell activation and chronic inflammation.18 Mast cells also interact with eosinophils and basophils through cytokines and chemokines, amplifying the inflammatory response.25 These mechanisms highlight the complex role of mast cells, with endotype classification predicting treatment outcomes, as type IIb patients show reduced response to omalizumab due to reliance on non-IgE pathways.29
Eosinophils
Eosinophils play a significant role in CSU inflammation, being recruited to the skin alongside mast cells and basophils, contributing to symptoms such as wheals and itch.30 Histologic studies show eosinophils in CSU lesions, with their activated granules indicating active involvement in dermal inflammation. Eosinopenia, due to eosinophil recruitment into affected skin, is common during disease flares.30 Immunologic destruction of eosinophils in the blood, possibly driven by autoimmune processes, has been suggested as another cause of eosinopenia.31 Eosinophils amplify inflammation through cytokine cross-talk with mast cells, promoting mediator release, thrombin generation, and mast cell degranulation.32 In CSU lesions, eosinophils release vasoactive mediators that enhance tissue edema.33 Elevated eosinophil counts in nonlesional skin suggest a primed inflammatory state, contributing to the chronic nature of CSU.34 Eosinophils interact with neutrophils, basophils, macrophages, and T cells to sustain inflammation, with their role in T2-dominant inflammation linked to pathways like autoallergy and neuroimmune dysregulation.35 Targeting eosinophils with anti-IL-5 antibodies like mepolizumab has shown potential in reducing CSU symptoms, but the ARROYO trial showed that benralizumab-induced eosinophil depletion did not significantly improve symptoms, highlighting the complexity of their role in CSU.33 These findings suggest the need for endotype-specific approaches, where eosinophil biomarkers may predict responses to treatments like dupilumab.32
Basophils
In CSU, basophils play a vital role by infiltrating the skin and releasing mediators like histamine, contributing to skin edema, flares, and pruritus lasting over six weeks.36 Basophil activation in CSU is often triggered by autoantibodies, such as IgG against IgE or FcεRI, leading to degranulation and exacerbating symptoms through increased vascular permeability and itch.37 They also interact with the coagulation and complement systems, where anaphylatoxins like C3a and C5a activate protease-activated receptor 1 (PAR1) on basophils, promoting inflammation.26 Peripheral basopenia during active disease phases correlates with disease activity, reflecting basophil migration to skin lesions.38,39 In a mouse model of urticaria-like inflammation, basophil infiltration into the skin was observed within one to two days post-challenge, with reduced peripheral basophil counts.38 Basophils in CSU exhibit altered FcεRI-mediated degranulation, with heightened sensitivity to non-IgE stimuli like C5a, leading to histamine release even in non/low responders to anti-IgE antibodies.40 They also release Th2 cytokines (IL-4, IL-13, IL-31), which facilitate T-cell activation and intensify pruritus, and leukotriene C4 (LTC4), contributing to wheal formation.41 Basophil activation is further enhanced by priming factors like IL-3, IL-5, GM-CSF, and nerve-growth factor, increasing histamine release and promoting neuro-immune interactions.42 Basophils express receptors like H4, NK1, and MRGPRX2, amplifying itch through shared signaling with neurons.43 Therapies targeting IgE, such as omalizumab, normalize basophil counts and function, serving as a biomarker for disease remission. These mechanisms highlight basophils’ role in CSU’s inflammatory network, interacting with mast cells and eosinophils to prime the skin for recurrent symptoms.32
T Lymphocytes
In CSU, Th2 cells infiltrate the perivascular regions of the skin, promoting inflammation through cytokines like IL-4, IL-5, and IL-13, which enhance IgE production and mast cell activation, leading to histamine release and pruritus.29 Th2-driven inflammation is further amplified by elevated Th2-initiating cytokines, such as IL-33, IL-25, and TSLP, with IL-33 localizing to mast cells and IL-25 expressed by mast cells and eosinophils.18 Th2 cells also interact with mast cells via IL-6 and IL-4, promoting mast cell proliferation and chemotaxis, while mast cells release PGD2 and LTE4 to support Th2 activation.18,44 Histamine from mast cells acts on Th2 cells via the H4 receptor, boosting Th2 cytokine secretion while suppressing Th1 cytokines, reinforcing the Th2-dominant milieu in CSU.45 Th2 cells crosstalk with ILC2s, enriched in CSU lesions, leading to a feedback loop of IL-5 and IL-13 production, further exacerbating inflammation.46 Elevated serum levels of Th2 cytokines, like IL-31 and IL-33, correlate with disease activity and pruritus intensity. Th2 dominance in CSU biopsies is marked by increased GATA-3+ Th2 cells and Th17 skewing, with Th2 contributing to autoreactivity, such as IgE autoantibodies against thyroid peroxidase.47 In chronic cases, Th2-related cytokines (IL-10, IL-13) are associated with autoreactivity and allergen sensitivity.48 Omalizumab treatment does not significantly alter Th2 cytokine profiles, suggesting Th2 mechanisms may persist independently of IgE blockade in some patients.49,50 Th2-mediated mechanisms, along with Th17/Treg imbalance, may synergize to drive chronic inflammation, as seen in elevated IL-17 and IL-21 levels, promoting mast cell proliferation in severe CSU cases.50
Immune Cell Surface Receptors as Therapeutic Targets
The pathogenesis of CSU is driven by complex interactions among immune cells, with surface receptors playing critical roles in mediating inflammatory responses.11 Targeting these receptors offers a promising strategy for developing precise and effective immunotherapies. Key receptors, including Siglec-8, FcεRI, MRGPRX2, and the histamine H4 receptor (H4R), regulate activation and degranulation of mast cells, eosinophils, basophils, and T cells, contributing to CSU symptoms such as wheals, angioedema, and pruritus.51–53 By leveraging nanomaterials to modulate these receptors, novel therapeutic approaches can achieve selective immune suppression and enhanced treatment outcomes.54,55 The mechanisms of these receptors are summarized in Table 2, while the following sections focus on their therapeutic implications, clinical development status, and nanomaterial integration.
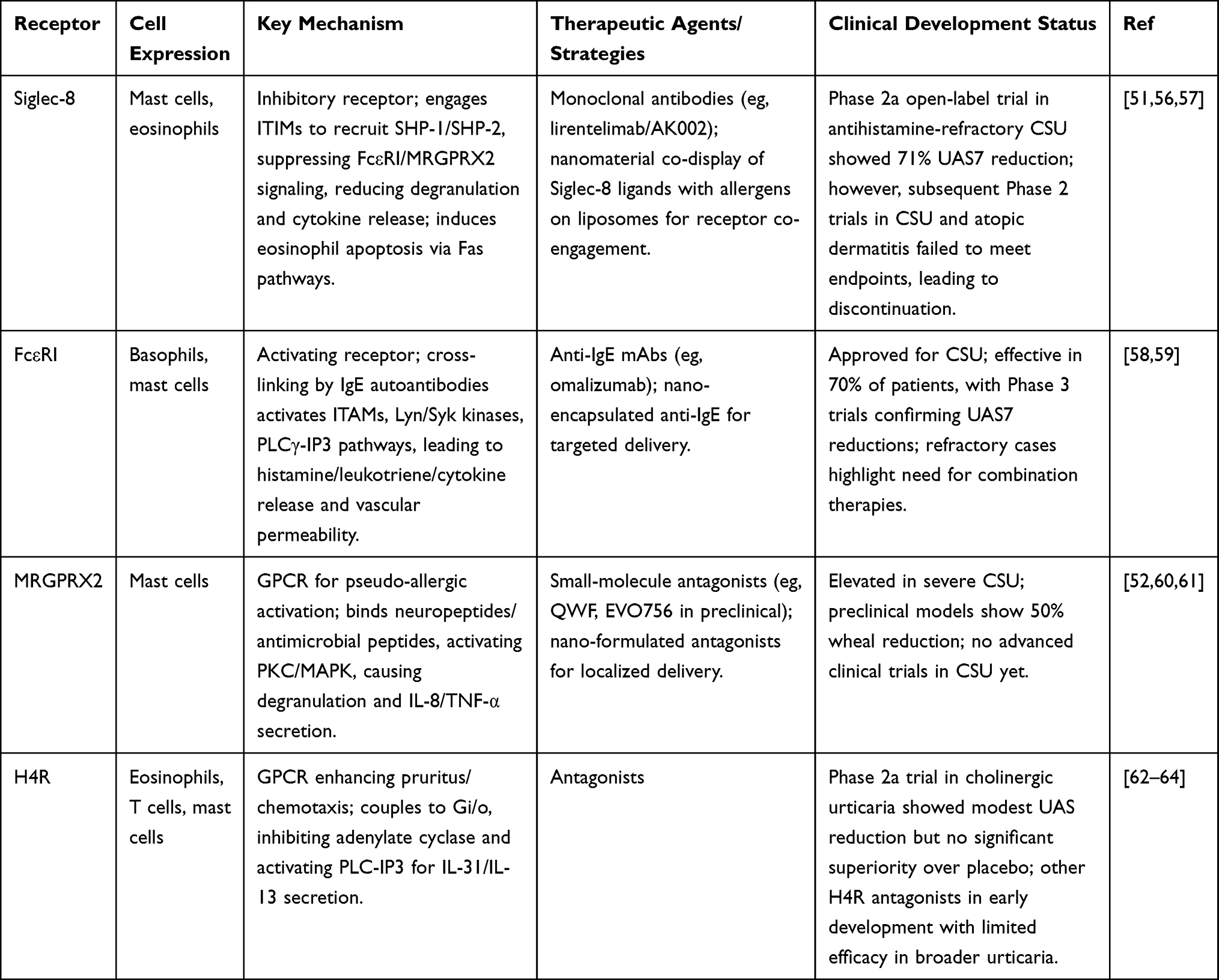 |
Table 2 Summary of Key Immune Cell Surface Receptors in CSU: Mechanisms, Therapeutic Targets, and Clinical Status |
In CSU, Siglec-8 is an inhibitory sialic acid binding immunoglobulin like lectin selectively expressed on mast cells and eosinophils. It recruits phosphatases such as SHP-1 and SHP-2 by binding to the immune receptor tyrosine based inhibitory motif (ITIM), thereby comprehensively inhibiting intracellular signaling pathways downstream of activated receptors such as Fc ε RI and MRGPRX2, thereby reducing degranulation, cytokine release, and inflammatory mediator production to allergens or stimulinon-specific stimuli.65 Specifically, ligation of Siglec-8 with monoclonal antibodies like lirentelimab (AK002) induces eosinophil apoptosis via Fas-mediated pathways and inhibits IgE-independent mast cell activation triggered by IL-33 or substance P, leading to decreased histamine and prostaglandin D2 release, as demonstrated in preclinical models where anti-Siglec-8 treatment reduced airway inflammation by 50–70% in humanized mice challenged with house dust mite extract.66 In a phase 2a open-label trial for antihistamine-refractory CSU, lirentelimab administration at doses up to 3 mg/kg monthly resulted in a 71% reduction in urticaria activity score (UAS7) by week 22, with skin biopsies showing diminished mast cell and eosinophil infiltration, underscoring Siglec-8’s role in modulating type 2 inflammation and pruritus through inhibition of key signaling molecules like Syk and Lyn.57 The high-affinity IgE receptor FcεRI, overexpressed on basophils and mast cells in CSU, initiates pathogenesis upon cross-linking by IgE autoantibodies or allergens, activating immunoreceptor tyrosine-based activation motifs (ITAMs) on its β and γ chains to phosphorylate kinases such as Lyn and Syk, which trigger phospholipase Cγ (PLCγ)-inositol trisphosphate (IP3) pathways, calcium mobilization, and subsequent release of histamine, leukotrienes, and cytokines that enhance vascular permeability and wheal formation.67 Anti-IgE therapies like omalizumab target FcεRI-bound IgE to downregulate receptor expression by depleting free IgE, disrupting receptor stabilization and reducing basophil sensitivity, as evidenced in a study of 110 CSU patients where 300 mg omalizumab every 4 weeks decreased FcεRI-positive basophils from 85% to 45% after 3 months, correlating with UAS7 scores dropping from 28 to 7 and improved anti-IgE responsiveness. In CSU with autoreactive IgE, such as against thyroid peroxidase, FcεRI cross-linking amplifies mast cell priming via IL-4 and IL-13 feedback loops, sustaining chronic inflammation, and omalizumab’s efficacy in 70% of patients highlights FcεRI’s central role in both type I and type II autoimmunity-driven urticaria subtypes.3
MRGPRX2, a G-protein-coupled receptor on mast cells, mediates IgE-independent pseudo-allergic activation in CSU. It binds neuropeptides such as substance P and antimicrobial peptides like LL-37, coupling with Gq/11 proteins to activate protein kinase C (PKC) and mitogen-activated protein kinase (MAPK) pathways. This results in rapid mast cell degranulation and the secretion of IL-8 and TNF-α, which exacerbate non-histaminergic itch and wheal persistence.68,69 Elevated MRGPRX2 expression in CSU lesional skin correlates with disease severity. In a cohort, serum MRGPRX2 levels were 2–3 times higher in patients with severe UAS7 ≥28 compared to controls. Stimulation with agonists such as compound 48/80 induced a 40–60% increase in histamine release from patient-derived mast cells, suggesting synergy with FcεRI priming in refractory cases.52 Pharmacological blockade of MRGPRX2 with small molecules like QWF inhibits degranulation in human mast cells, reducing wheal size by up to 50% in preclinical urticaria models, and supports its potential in antihistamine-resistant CSU where agonists like β-defensins from keratinocytes amplify inflammation.70 The histamine H4 receptor (H4R) on eosinophils, T cells, and mast cells in CSU enhances pruritus and chemotaxis by coupling to Gi/o proteins, inhibiting adenylate cyclase while activating PLC-IP3 to promote IL-31 and IL-13 secretion, thereby sustaining Th2-skewed responses and eosinophil recruitment.62 In a phase 2 trial for cholinergic urticaria, the H4R antagonist izuforant (LEO 152020) at 100 mg twice daily reduced provocation UAS by 4.5 points versus placebo after 4 weeks, associated with decreased IL-13 levels and eosinophil migration, illustrating H4R’s contribution to complement-independent inflammation in inducible urticaria subtypes.62 These receptor mechanisms provide a foundation for nanomaterial-based interventions in urticaria, where nanoparticles can be engineered for targeted delivery and multivalent ligand presentation to enhance inhibitory signaling, such as co-displaying allergens and sialic acid-based Siglec-8 ligands on liposomes to enforce receptor co-engagement and suppress FcεRI-mediated mast cell activation.14,51 For instance, in a murine anaphylaxis model mimicking urticaria-like responses, allergen-Siglec-8 ligand nanoparticles prevented IgE-FcεRI cross-linking-induced degranulation, reducing systemic histamine release by 80% and desensitizing mast cells to subsequent challenges via ITIM-mediated SHP-1 recruitment, highlighting nanomaterials’ potential for precise modulation of immune receptors without broad immunosuppression.14 Furthermore, hydrophobic nanoparticles can preferentially interact with innate immune cells to alter receptor clustering and signaling, as demonstrated in studies where surface-modified nanoparticles modulated FcεRI aggregation on basophils, reducing pro-inflammatory cytokine output by 40–60% in allergic models, paving the way for nano-formulated antagonists against MRGPRX2 or H4R in CSU therapy.71
Despite their potential, receptor-targeted strategies in CSU face significant limitations. Clinical efficacy varies due to disease heterogeneity; for instance, omalizumab succeeds in IgE-dependent cases but fails in 30% of refractory patients reliant on non-IgE pathways like MRGPRX2.72,73 Siglec-8 targeting with lirentelimab showed inconsistent results, with early promise overshadowed by Phase 2 failures, possibly due to incomplete suppression of multifaceted inflammation or patient selection biases.57,74 H4R antagonists exhibit limited benefits, as tissue drug levels may be insufficient for full receptor blockade, and anti-pruritic effects have not translated robustly across urticaria subtypes.62,75 Translational challenges include off-target effects, such as broad immune suppression risking infections, and immunotoxicity from nanomaterials (eg, unintended complement activation or cytokine storms).76 Nanomaterial integration adds hurdles like scalability, long-term biocompatibility, and regulatory approval for clinical use, with few CSU-specific trials advancing beyond preclinical stages due to high costs and variable biodistribution. Future success requires biomarker-driven patient stratification, combination therapies (eg, dual FcεRI/MRGPRX2 blockade), and AI-optimized nano-designs to overcome these barriers and enhance precision.77
An Overview About Immunological Interactions of Nanomaterials
Nanomaterials have emerged as promising platforms in the immunotherapy of CSU due to their unique physicochemical properties, which critically influence therapeutic efficacy by modulating interactions with immune cells and the biological environment.78 For instance, particle size plays a key role in cellular uptake; smaller sizes (eg, 20–100 nm) facilitate endocytosis in mast cells and basophils, thereby enhancing targeted delivery of therapeutic agents such as ascomycin while reducing systemic exposure and off-target effects, as evidenced in models where 60–70 nm PLGA nanoparticles effectively downregulated FcεRI expression.58 Surface charge governs cellular adhesion and immune activation, with positively charged nanoparticles (eg, cationic liposomes) promoting electrostatic binding to negatively charged membranes of eosinophils and T cells, thus supporting receptor engagement (eg, Siglec-8) and immune tolerance induction; however, excessive positive charge may activate complement pathways, requiring careful optimization to prevent pseudo-allergic reactions in CSU patients.56,79 Additionally, surface morphology, including roughness or porosity (eg, in mesoporous silica nanoparticles), impacts ligand density and release kinetics, enabling sustained administration of allergens or immunomodulators to dendritic cells for extended Th2 suppression, with research indicating that porous designs increase payload retention by 2–3 fold and decrease degranulation by 40–60% in basophil models.57,60 Collectively, these properties—size, charge, and morphology—enhance bioavailability, specificity, and safety in CSU immunotherapy, permitting precise modulation of inflammatory pathways without inducing broad immunosuppression. Meanwhile, nanomaterials have also provided direction for precise diagnosis and treatment of CSU patients.77
Nanomaterials in CSU Immunotherapy: Classification and Therapeutic Potential
Traditionally, nanomaterials can be divided into several categories based on their composition and structure, including inorganic nanomaterials (such as metal nanoparticles and silicon-based nanoparticles), organic nanomaterials (such as polymer nanoparticles and liposomes), and carbon-based nanomaterials (such as graphene and carbon nanotubes). These categories exhibit unique biocompatibility, immune regulatory ability, and targeted delivery potential in their immunological properties, making them particularly promising for immunotherapy of allergic diseases such as urticaria.9,55 Metal nanoparticles, such as gold and silver nanoparticles, typically possess antioxidant and anti-inflammatory properties, and can inhibit Th2-type immune responses by regulating immune cell activation, thereby alleviating mast cell degranulation in urticaria.9 Silicon-based nanoparticles are known for their high loading capacity and controllable release characteristics and can serve as allergen delivery carriers to promote immune tolerance induction.80 Polymer nanoparticles are widely used to target immune cells, such as dendritic cells and regulatory T cells, to reshape immune imbalances associated with urticaria owing to their biodegradability and surface modification flexibility.81 Liposome nanomaterials simulate cell membrane structure, can encapsulate immunomodulators, reduce systemic side effects, and target IgE-mediated allergic reactions in the treatment of urticaria.82 While carbon-based nanomaterials have limited applications, their high surface area and functional potential can enhance the efficacy of immune adjuvants and potentially be used for long-term immune regulation in urticaria (Figure 2, Table 3). Although the application of these nanomaterials in general allergic diseases such as asthma and food allergies has been widely reported, direct clinical evidence for CSU is still relatively limited, mainly relying on animal models and in vitro studies. This gap highlights the urgent need for CSU specific randomized controlled trials to validate its safety and efficacy.
 |
Figure 2 Types of nanoparticles and their immunological interactions in CSU immunotherapy. Upper panel: Classifications based on composition—Liposome, Polymeric, Spherical 3D, Metallic, Micelle, Gold NPs. Lower panel: Interactions with immune cells—Eosinophils with Metallic NPs (releasing IL-4, IL-10), Basophils with Gold NPs via FcεRI (releasing histamine, IL-4, IL-13), Mast cells with Spherical 3D NPs via FcεRI (releasing histamine, ROS). Adapted from González-Fernández, África, and Immacolata Maietta. The Forgotten Innate Immune Cells: Unraveling Their Prospective Interactions with Nanomaterials. International journal of nanomedicine vol. 20 8173–8189. Copyright © 2022 by authors.83 |
 |
Table 3 Comparative Advantages and Limitations of Nanomaterial Types in CSU Immunotherapy |
Furthermore, their applications in allergic diseases such as urticaria mainly include serving as immunotherapy carriers, immunomodulators, and diagnostic tools. The therapeutic targets focus on inhibiting the activation of mast cells and eosinophils, regulating Th1/Th2 balance, blocking the IgE Fc ε RI signaling pathway, and promoting the proliferation of regulatory immune cells.12,55 The prospects lie in improving treatment specificity and safety, thereby reducing dependence on traditional antihistamines. Table 4 summarizes the classification, applications, therapeutic targets, and prospects of nanomaterials.
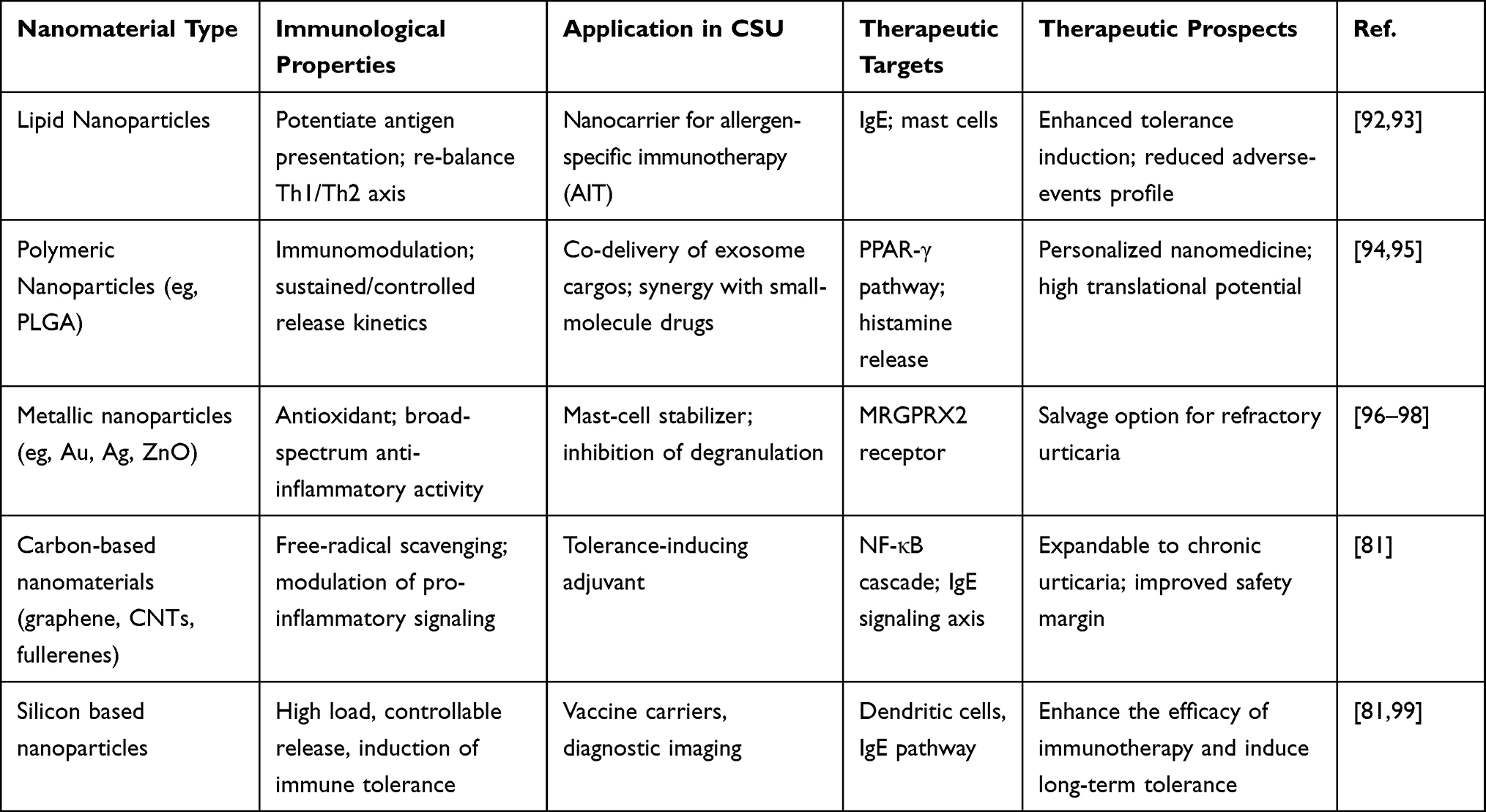 |
Table 4 Representative Nanomaterials Under Investigation for Targeted Intervention in CSU |
Specifically, different nanomaterials can affect the immune related effects of urticaria through various mechanisms, therefore nanomaterials have emerged as promising platforms in the immunotherapy of allergic diseases, including CSU.100 For example, inorganic nanoparticles such as gold and silica exhibit adjuvant-like effects, enhancing antigen presentation to dendritic cells and promoting tolerogenic responses in urticaria models.84 Furthermore, organic polymeric nanoparticles can be engineered to deliver allergens while minimizing systemic exposure, thereby reducing the risk of anaphylaxis associated with traditional subcutaneous immunotherapy for chronic urticaria.11 In addition, lipid-based nanomaterials facilitate targeted delivery to mast cells, inhibiting degranulation through Siglec-8 engagement and offering a novel mechanism to alleviate urticaria symptoms.8 While, carbon based nanomaterials can be functionalized into immunosensors to monitor urticaria immune markers such as serum IgE levels in real-time and regulate the local inflammatory microenvironment through photothermal effects. These mechanisms reflect the core role of size effects, surface charges, and biological functionalization of nanomaterials in immunotherapy for urticaria, enriching multi-level strategies from prevention to cure.12 Consequently, these nanomaterials not only improve the safety profile of allergen-specific immunotherapy but also enhance efficacy by reprogramming Th2-dominant immune responses toward tolerance in urticaria patients.12 Moreover, recent advancements in nanoparticle functionalization have demonstrated potential in modulating the tumor microenvironment analogies in allergic inflammation, suggesting broader applications for urticaria management.101 Therefore, integrating nanomaterials into clinical protocols could revolutionize urticaria treatment by providing sustained immune regulation and personalized therapeutic options. Up to now, the prospective outlook for nanomaterial-based therapies in urticaria immunotherapy is highly optimistic, with ongoing trials indicating reduced relapse rates and improved patient compliance.102 Despite the optimistic prospects of nanomaterials, potential risks must be considered, including immunotoxicity (such as cytokine storm induction), organ accumulation (especially long-term retention in the liver and kidneys), and PEG related allergic reactions (which may exacerbate CSU symptoms).103 For example, PEGylated liposomes may trigger complement activation, leading to pseudo allergic reactions, which is particularly relevant in allergic patients.104 These risks can be mitigated through surface modification and dose optimization, but the need for comprehensive toxicological evaluation in CSU clinical trials is emphasized.
Specific Targeting Mechanism of Nanomaterials on Immune Cells in CSU
At present, nanomaterials have emerged as powerful tools in the immunotherapy of CSU due to their ability to precisely target immune cells involved in the disease’s pathogenesis. In the following sections, we will explore the specific mechanisms by which various nanomaterials interact with these immune cells, emphasizing their therapeutic potential in CSU through targeted receptor engagement, immune tolerance induction, and reduced off-target effects (Table 5).
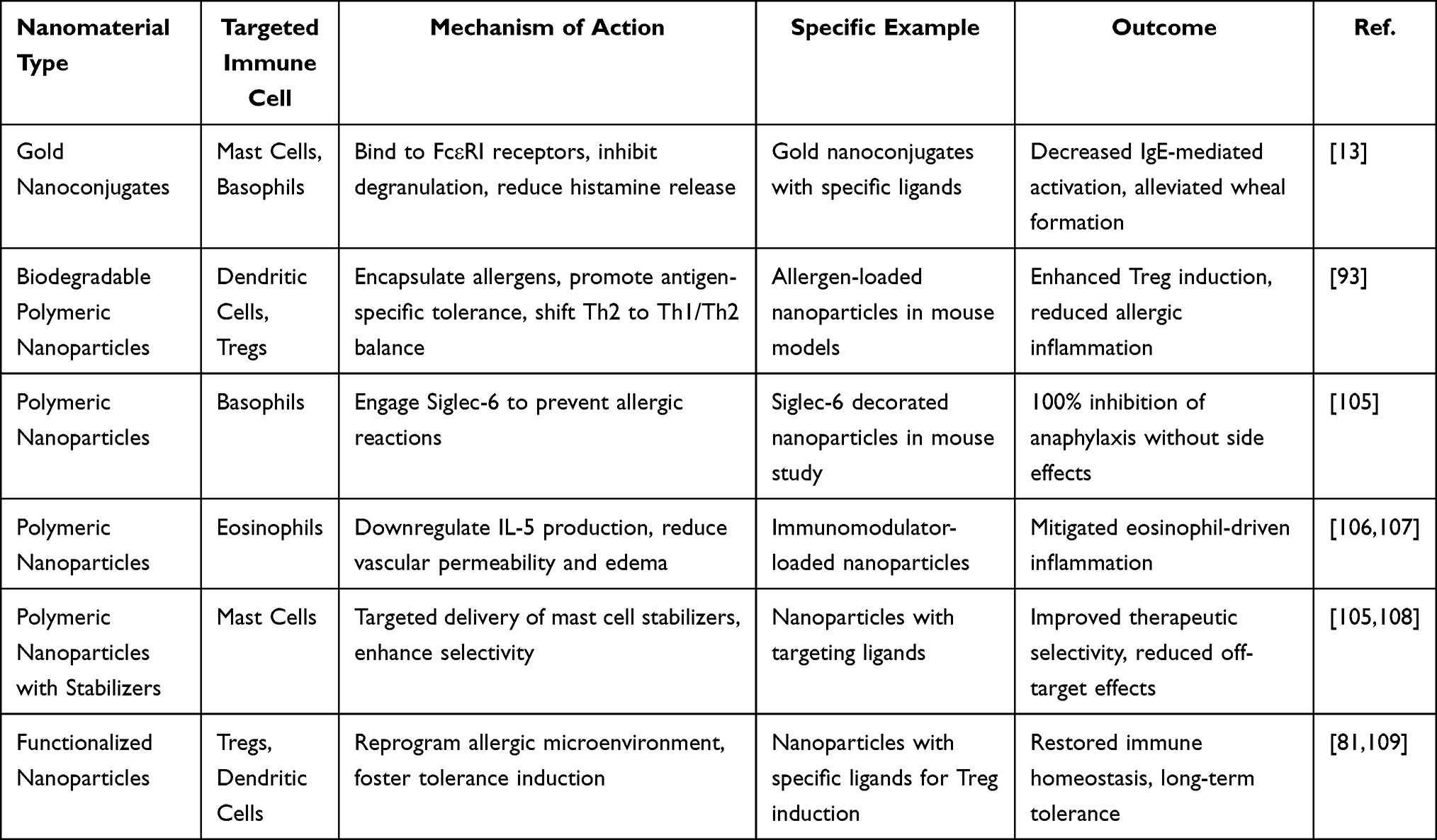 |
Table 5 Mechanisms of Nanomaterial-Mediated Immune Cell Targeting for CSU Immunotherapy |
Numerous studies have shown that nanomaterials can precisely target mast cells in urticaria using surface-functionalized nanoparticles.13 For example, Inna M. Yasinska et al used gold nanoconjugates decorated with specific ligands that bind to FcεRI receptors, inhibiting degranulation, reducing histamine release, and alleviating wheal formation in allergic models.13 These gold nanoconjugates have also been shown to effectively silence pro-allergic reactivity in basophils and mast cells, leading to reduced IgE-mediated activation. This interruption of the effector phase of hypersensitivity offers a promising therapeutic approach for CSU.13 Beyond mast cell targeting, biodegradable nanoparticles encapsulating allergens, as investigated in recent studies, target dendritic cells to promote antigen-specific tolerance, shifting the Th2-dominant response towards Th1/Th2 balance and preventing recurrent urticaria episodes through enhanced regulatory T cell induction.110,111 In a specific example, Saunders MN et al used prophylactic administration of such nanoparticles altered immune pathways in T cells, basophils, and mast cells, resulting in reduced allergic inflammation and long-term suppression of FcεRI signaling in mouse models of allergic diseases including urticaria-like symptoms.110 Another promising approach involves decorated nanoparticles designed to engage Siglec-6 on basophils. These nanoparticles have been shown to prevent severe allergic reactions, with a Northwestern University study reporting 100% success in inhibiting anaphylaxis in mice without side effects. This highlights their role in modulating basophil activation, which is central to urticaria pathogenesis.15 Additionally, several studies have identified polymeric nanoparticles loaded with immunomodulators that target eosinophils by downregulating IL-5 production. This mitigates vascular permeability and edema in urticaria, addressing the biological mechanism where eosinophil recruitment exacerbates chronic hives through cytokine amplification.12
These mechanisms collectively enable nanomaterials to modulate immune-related processes in urticaria, including cell degranulation, cytokine secretion, and antibody switching, by offering controlled drug release and enhanced bioavailability, thus restoring immune homeostasis without the need for systemic immunosuppression.78 Current research suggests nanoparticle-based strategies that combine targeting ligands with mast cell stabilizers to selectively target mast cell disorders, such as urticaria, improving therapeutic selectivity and minimizing off-target effects in preclinical evaluations.78 By leveraging the tunable properties of nanomaterials, such as size, charge, and functionalization, these treatments can precisely reprogram the allergic microenvironment in urticaria, promoting tolerance induction in key immune cells like Tregs and dendritic cells.16 Recent research highlights the promising potential of nanomaterial-based immunotherapy for urticaria. Ongoing investigations focus on improving efficacy, personalizing targeting, and minimizing adverse reactions, aiming to facilitate clinical translation in the near future.17,112
Administration Routes of Nanomaterials in CSU Immunotherapy
Following the discussion on the immunological interactions of nanomaterials, including how their physicochemical properties like size, charge, and morphology influence cellular uptake and precise targeting, along with their various classifications and mechanisms for engaging specific immune cells such as mast cells, eosinophils, basophils, and T lymphocytes, the route of administration plays a crucial role in maximizing their effectiveness for treating CSU.14,57 In managing CSU, where persistent inflammation stems from imbalanced Th2 pathways and excessive mast cell activity, selecting the appropriate delivery method directly affects drug absorption, access to affected immune cells, patient adherence, and treatment safety.113,114 Conventional approaches, like oral antihistamines or injectable biologics such as omalizumab, frequently encounter issues such as widespread side effects, inconsistent uptake, and poor reach to inflamed skin areas, often resulting in partial relief and frequent symptom recurrence.2,115 Nanomaterials overcome these hurdles through adaptable delivery options that leverage their customizable features—for example, improved mucosal absorption in intranasal administration or gradual release in oral systems—to enable focused immunomodulation, limit unintended impacts, and foster enduring immune tolerance without suppressing the entire immune system (Figure 3).51,116
 |
Figure 3 Nanoparticle administration routes for immunotherapy in allergic diseases. (A) Bispecific nanobody-coated nanoparticles targeting nasal and ocular epithelia to block IgE-mediated activation and inhibit effector cell responses. (B) Inhaled nanoparticles targeting dendritic cells in the lungs to modulate Th2-driven inflammation and promote immune tolerance. (C) Oral nanoparticles for food allergies, delivering anti-allergen and anti-cytokine treatments to reduce IgE responses and enhance regulatory T cell activity. Adapted from Zettl, Ines et al. Single-Domain Antibodies-Novel Tools to Study and Treat Allergies. International journal of molecular sciences vol. 25,14 7602. 11 Jul. 2024, doi:10.3390/ijms25147602. Copyright © 2024 by authors.117 |
Smaller nanoparticle dimensions (20–100 nm), for instance, support efficient absorption in sublingual or intranasal delivery, while tailored surface alterations improve binding to epithelial layers, facilitating direct interaction with receptors like FcεRI or Siglec on immune cells in the skin or mucous membranes.14 This synergy not only builds on the foundational benefits of nanomaterials but also aligns closely with CSU’s underlying biology, emphasizing localized skin inflammation that benefits from methods combining broad immune adjustment with targeted relief, ultimately leading to better results for patients resistant to standard treatments.5,115 To demonstrate how these routes can be applied practically in CSU or related allergic disorders (which share similar IgE-driven processes), Table 6 highlights selected nanomaterial examples, their delivery approaches, and associated advantages, based on insights from preclinical and clinical research.
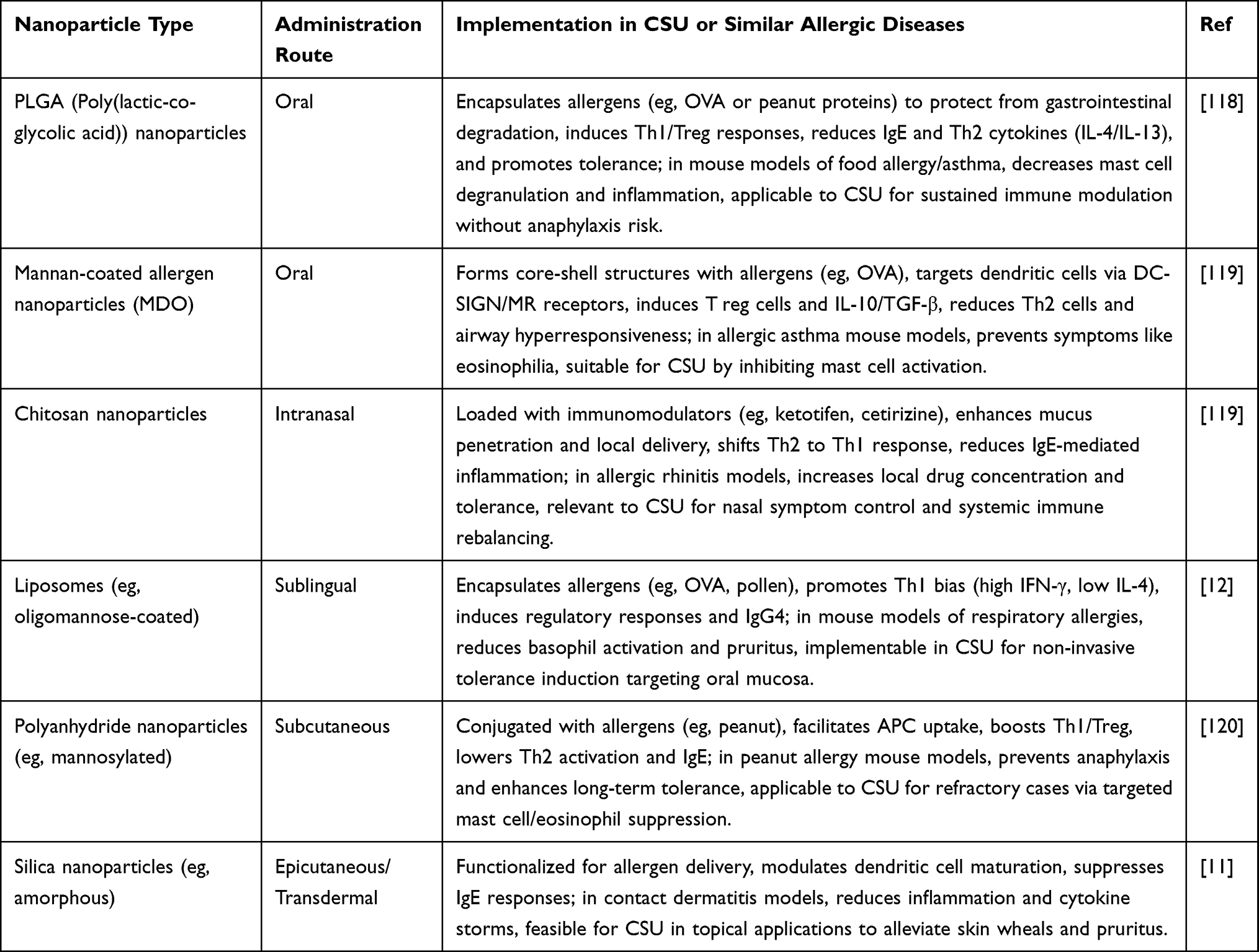 |
Table 6 Representative Administration Routes of Nanomaterials for CSU Immunotherapy |
Nanomaterials Targeting Immune Cells for The Treatment of CSU Immunotherapy
Targeted modulation of immune cells represents a critical strategy in CSU immunotherapy, as these cells orchestrate the complex inflammatory cascades underlying disease pathogenesis. Unlike conventional systemic treatments that often lack specificity and cause widespread immunosuppression, nanomaterial-based approaches enable precise intervention at the cellular level, selectively reprogramming pathogenic immune responses while preserving normal immune function. This targeted approach is particularly important in CSU, where multiple immune cell types—including mast cells, eosinophils, basophils, and T lymphocytes—interact dynamically to perpetuate chronic inflammation and tissue damage (Figure 4). By directly engaging these key cellular mediators, nanomaterials can interrupt disease-driving pathways with enhanced efficacy and reduced adverse effects.
 |
Figure 4 Nanomaterials targeting immune cells in urticaria immunotherapy. Nanoparticles interact with neutrophils (modulating migration and drug delivery), monocytes/macrophages (inhibiting production and recruitment), T/B lymphocytes (inducing tolerance and depleting autoreactive cells), dendritic cells (promoting tolerogenic phenotype), and NK cells (enhancing cytotoxicity). They also neutralize cytokines, scavenge ROS/RNS, inhibit MPO, and eliminate cell-free DNA. Adapted from Liu, Jin et al. The interaction between nanoparticles and immune system: application in the treatment of inflammatory diseases. Journal of nanobiotechnology vol. 20,1 127. 12 Copyright © 2022 by authors.121 |
Nanoparticle Platforms for Enhanced Mast Cell Targeting in CSU
Various nanoparticle platforms have been engineered to modulate mast cell function, with poly(propylene sulfone) (PPSU) nanoparticles being a notable example. These nanoparticles interact with mast cells by facilitating controlled adsorption of bioactive proteins like anti-Siglec-6 and anti-FcεRIα antibodies. Hydrophobic patches on the protein surfaces induce site-specific dipole relaxation in PPSU assemblies for non-covalent anchoring, without disrupting hydrogen bonding or protein structure.15 This interaction enables co-localized engagement of FcεRIα and Siglec-6 on mast cell surfaces, inhibiting IgE-mediated activation and degranulation by suppressing markers like CD107a and CD63. This potentially alleviates urticaria symptoms by reducing histamine release and inflammatory mediator secretion.15 Another important platform involves gold nanoparticles (AuNPs) conjugated with the calcineurin inhibitor ascomycin and stem cell factor (SCF) as a targeting ligand. These nanoparticles interact with mast cells through amine-coupling to reduced glutathione spacers on the gold surface, allowing specific binding and internalization that block pro-allergic signaling pathways and IgE-dependent degranulation. This mechanism inhibits mast cell hyperactivity in urticaria without systemic side effects.13 Beyond antibody- and ligand-based approaches, sesquiterpene-loaded poly(lactic-co-glycolic acid)/polyvinyl alcohol (PLGA/PVA) hybrid nanoparticles (60–70 nm) are readily internalized by mast cells through endocytosis or lipid bilayer traversal. These nanoparticles release their payload as peroxisome proliferator-activated receptor gamma (PPARγ) ligands, downregulating gene expression of FcεRI subunits (α, β, γ) and the stem cell factor receptor Kit. This diminishes antigen responsiveness and shifts the mast cell phenotype toward reduced tryptase expression.122 This gene modulation inhibits mast cell degranulation upon IgE crosslinking with FcεRI, potentially suppressing urticaria by attenuating allergic inflammation and mediator release, without compromising cell viability. Collectively, these examples demonstrate how nanoparticle platforms enhance mast cell targeting through multivalent ligand conjugation, optimization of size, morphology, and surface chemistry to improve pharmacokinetics and selectivity, while protecting drug payloads from degradation. This enables more effective inhibition of mast cell-driven pathologies like urticaria with minimized off-target interactions.78
The Role of Nanomaterials in Eosinophil Regulation
Eosinophil-targeted nanomaterial platforms have shown significant therapeutic potential by utilizing various mechanisms to regulate eosinophil function and reduce allergic inflammation.123 Liposomes co-displaying allergen and Siglec-8 ligands interact with eosinophils by binding to the inhibitory Siglec-8 receptor, selectively expressed on these cells. This suppresses IgE-mediated activation and induces eosinophil apoptosis, potentially alleviating allergic inflammation in conditions like urticaria by reducing eosinophil survival and effector functions.14,51 Building upon this receptor-targeting approach, bio-mimetic nanoparticles modulate eosinophil responses in models of allergic airway disease by mimicking natural cellular interactions, resulting in decreased eosinophil infiltration and attenuation of Th2-driven inflammation, thus offering a mechanism for inflammation relief in urticaria by inhibiting eosinophil recruitment and cytokine release.124,125 Complementary immunomodulatory strategies include curcumin-loaded nano-formulations combined with allergens. These formulations reduce eosinophil counts in nasal lavage fluid and downregulate Th2 cytokines like IL-4 and IL-5, inhibiting eosinophil activation. This provides a pathway for alleviating allergic symptoms in urticaria through targeted delivery and enhanced bioavailability.126 Similarly, peptide-based lipid nanodiscs, functioning as synthetic high-density lipoproteins, suppress eosinophil recruitment and chemotaxis via targeted receptor interactions and signaling modulation, leading to decreased migration and adhesion, which could translate to urticaria treatment by limiting eosinophil-mediated histamine release and tissue inflammation.127 Indirect modulation approaches have also shown promise.128 Dendritic cell-targeted gold nanoparticles act as biocompatible carriers for sublingual immunotherapy, modulating allergic responses by indirectly influencing eosinophil activation through enhanced antigen presentation and tolerance induction. This may reduce eosinophil degranulation and inflammatory mediator secretion in urticaria.128
Organic sheet-like nanosheets, designed as inorganic-free nanomaterials, address allergic airway inflammation by interacting with eosinophils to reduce their activation. This leads to decreased inflammatory cell infiltration and cytokine storms, suggesting their potential role in urticaria by promoting inflammation resolution through eosinophil phenotype modulation.129 Furthermore, nanoparticle-conjugated immunomodulators enhance the effects of allergen-specific therapy by facilitating pulmonary delivery and modulating eosinophil responses in the lungs, leading to decreased airway hyperresponsiveness and eosinophilia, with implications for urticaria management via systemic or localized suppression of eosinophil-driven allergic cascades.130,131 Additionally, PLGA-encapsulated mesenchymal stem cell exosomes exhibit immunomodulatory effects by reducing eosinophil infiltration in nasal mucosa. This is achieved through the delivery of anti-inflammatory factors that inhibit eosinophil activation and goblet cell hyperplasia, providing a novel nanomaterial-based strategy for inflammation relief in allergic diseases, including urticaria.132 These nanomaterial platforms improve eosinophil targeting through optimized size, surface functionalization, and controlled release profiles, enhancing pharmacokinetics and minimizing off-target effects. This enables precise regulation of eosinophil activation inhibition and overall allergic inflammation alleviation in urticaria, without compromising host immunity.123,133
The Role of Nanomaterials in the Regulation of Eosinophils
Recent advances in targeted basophil modulation show that functionalized nanoparticle platforms can precisely regulate IgE-mediated allergic responses through multiple complementary mechanisms.134 AuNPs functionalized with calcineurin inhibitors like ascomycin and targeting ligands such as stem cell factor interact synergistically with basophils by binding to surface receptors and delivering inhibitors. This suppresses IgE-mediated signaling and reduces pro-allergic mediator release, potentially alleviating urticaria symptoms through decreased histamine and cytokine secretion.13 This targeted approach inhibits basophil activation without systemic toxicity. The AuNPs’ biocompatibility and specific conjugation prevent off-target effects while promoting controlled release of the inhibitor to reduce allergic inflammation in urticaria.13 Fullerene nanomaterials (C60) preincubated with basophils inhibit IgE-dependent mediator release by regulating allergic signaling cascades. This suppresses degranulation and may alleviate urticaria-associated wheal formation through reduced vasoactive mediator levels.135 Poly(lactic-co-glycolic acid) (PLGA) nanoparticles encapsulating allergens for immunotherapy reduce basophil degranulation by shielding allergens from IgE recognition on basophil surfaces. This shifts immune responses toward tolerance and inhibits mediator release in allergic conditions like urticaria.136,137 Liposomes as nanoparticle carriers in allergen-specific immunotherapy enhance the tolerance-inducing effect. They modulate basophil responses by delivering immunomodulators that downregulate FcεRI-mediated activation, resulting in decreased histamine release and improved inflammation resolution in urticaria models.55,138 For diagnostic and therapeutic applications, dendrimeric antigen-silica nanoparticle composites enhance basophil activation test sensitivity for diagnostics but, in therapeutic contexts, can be engineered to inhibit basophil reactivity by optimizing surface chemistry to prevent epitope crosslinking and subsequent mediator liberation in urticaria immunotherapy.139,140 However, not all nanomaterial designs yield beneficial effects.141 Certain lipid nanoparticles (LNPs) may inadvertently enhance basophil activation through anti-PEG antibody induction and complement pathway stimulation, potentially exacerbating mediator release in susceptible individuals with urticaria, highlighting the need for careful nanomaterial design to avoid such proinflammatory effects.9 Chitosan-based nanoparticles loaded with histamine release inhibitors like ketotifen target basophils to suppress degranulation through prolonged mucosal retention and sequential drug release. This provides a mechanism to inhibit allergic cascades in urticaria without mucosal irritation.55 Mesoporous silica nanoparticles conjugated with IgE aptamers specifically target basophils to deliver payloads that disrupt diselenide bridges and inhibit activation. This reduces tryptase and histamine release, potentially controlling urticaria symptoms.11
The Role of Nanomaterials in The Regulation of T Lymphocytes
Therapeutic modulation of T lymphocyte responses is a critical frontier in CSU immunotherapy, as these cells drive Th2-dominant inflammatory cascades in chronic allergic reactions.16 By redirecting T cell differentiation toward regulatory phenotypes and restoring Th1/Th2 balance, nanoparticle-based interventions aim to address the underlying immune dysregulation, rather than merely suppressing symptoms.16 Biodegradable poly(lactic-co-glycolic acid) (PLGA) nanoparticles encapsulating antigens target antigen-presenting cells like dendritic cells, inducing antigen-specific tolerance. This suppresses Th2-mediated responses and may alleviate urticaria by reducing IgE production and inflammatory cytokine release.142,143 These PLGA nanoparticles shift the Th1/Th2 balance by enhancing regulatory T cell (Treg) differentiation and inhibiting Th2 cell activation, offering an innovative immunotherapy pathway for urticaria with minimal systemic effects.142 Gold nanoparticles conjugated with aryl hydrocarbon receptor (AhR) ligands and antigens complement this tolerogenic approach by modulating T cell responses via dendritic cell uptake. This induces tolerogenic phenotypes, expanding FoxP3+ Tregs and downregulating Th2 cytokines like IL-4 and IL-5, thus restoring Th1/Th2 equilibrium in allergic conditions like urticaria.121 pMHCII-coated nanoparticles specifically target autoreactive T cells, triggering TR1-like cell expansion and IL-10 secretion to suppress Th2-driven inflammation. This offers a targeted strategy for modulating T lymphocyte immunity in urticaria without broad immunosuppression.121,144 Surface modification strategies have further enhanced dendritic cell interactions. Mannose-modified PLGA nanoparticles loaded with allergens enhance dendritic cell phagocytosis and IL-10 production while reducing IL-6. This induces a tolerogenic shift, biasing T cell polarization away from Th2 dominance toward Th1 and Treg responses, potentially mitigating urticaria flares through balanced immune modulation.81 In a distinct approach, cellulose nanofibers as nanomaterials inhibit dendritic cell maturation and weaken Th1/Th17 responses while inducing Tregs, thereby facilitating a Th1/Th2 rebalancing that could inhibit mast cell activation indirectly in urticaria via altered T cell cytokine profiles.81 Liposomal nano-delivery systems with adjustable charges optimize dendritic cell targeting to suppress Th2 cell proliferation and enhance Treg-mediated tolerance. This offers a versatile platform for urticaria treatment by preventing excessive Th2 cytokine-driven histamine release.81,134
Nanomaterials Targeting Immune Cell Surface Receptors: Functional Design and Therapeutic Delivery System
Immune cell surface receptors play a key role in urticaria development. The high-affinity IgE receptor (FcεRI) on mast cells, basophils, and eosinophils, along with inhibitory receptors like Siglec-6 and Siglec-8, trigger IgE-mediated activation, degranulation, and the release of inflammatory substances like histamine and cytokines.14,145 When these receptors malfunction, they worsen allergic inflammation in urticaria, making them ideal targets for treatments that control overactive cells and restore immune balance without suppressing the entire immune system.145 Nanomaterials with specific surface molecules interact with these receptors, enabling precise drug delivery and immune modulation in allergic diseases.15 For example, in food allergies, nanoparticles block Th2-driven responses and promote regulatory T cells, reducing symptoms.93 These nanomaterial methods have effectively prevented severe allergic reactions, such as anaphylaxis, by deactivating mast cells and eosinophils through receptor-targeted blocking, reducing substance release and systemic inflammation in lab models.15,93 Similarly, nanoparticle-based allergen immunotherapy has enhanced treatment for allergic rhinitis and asthma by targeting dendritic cell receptors, shifting immune responses toward tolerance, reducing Th2 cytokine dominance, and improving long-term desensitization.17 New nanomaterial designs with anti-inflammatory drugs regulate innate immune cell receptors in atopic dermatitis, suppressing chronic inflammation and barrier issues through controlled release.146,147 These applications suggest that by using nanomaterials to target immune cell surface receptors, we can develop new treatments for urticaria that may achieve better results in blocking allergic reactions and resolving wheal formation with better safety.148
Building on receptor-targeting principles, several nanoparticle platforms have been developed for urticaria treatment. Poly(propylene sulfone) nanoparticles with anti-Siglec-6 antibodies target Siglec-6 on mast cells using non-covalent anchoring that preserves protein structure. This blocks IgE-mediated degranulation, suppressing urticaria symptoms by reducing substance release.149,150 This design enhances selective delivery to mast cells, reducing off-target effects and promoting inflammation resolution in urticaria by engaging inhibitory receptors to reduce cell activation.149 In a different approach, mannan-coated allergen nanoparticles target dendritic cell receptors like DC-SIGN and the mannose receptor through sugar molecules. This aids antigen presentation and induces regulatory T cell tolerance. This could reduce urticaria by modulating Th2-driven immune responses and decreasing IgE production.119 More broadly, engineered nanomaterials with specific properties like surface charge and ligand density interact with immune cell receptors to guide immune stimulation or suppression, enabling targeted therapy for urticaria by changing distribution and receptor signaling in effector cells. Several immunotherapy platforms have integrated these targeting strategies. Nanoparticle-based allergen immunotherapy platforms, such as polyesters and liposomes with specific molecules, engage immune cell surface receptors to disrupt Th2 cell dominance and improve tolerance. This potentially reduces urticaria flares by lowering allergen-specific IgE binding and effector cell activation.17 Similarly, biodegradable nanoparticles containing allergens like galactose-alpha-1,3-galactose target antigen-presenting cell receptors to increase immune tolerance. This reduces IgE-mediated reactions and offers a new delivery system for urticaria management through controlled adaptive immunity.110,151 Alternatively, nanoparticle-allergen complexes designed for receptor targeting on mast cells and basophils block degranulation through receptor crosslinking and signal disruption. This prevents histamine release in urticaria without suppressing the entire immune system.136 For T cell targeting, nanoparticles with antibodies against CD90 or CD45 on T cells enable precise delivery of immune-controlling drugs to restore Th1/Th2 balance. This modulates urticaria immunotherapy by suppressing pro-allergic cytokines and improving regulatory responses.152 Additionally, nanoparticles designed to interact with pattern recognition receptors on innate immune cells control inflammatory processes, providing a treatment delivery system that reduces urticaria symptoms through controlled receptor engagement and reduced inflammatory effects.103,121
Future Perspectives and Emerging Strategies
Clinical Translation
Although conventional treatment strategies for CSU, such as antihistamines (eg, H1-receptor antagonists like cetirizine), biologics (eg, omalizumab targeting IgE), and immunosuppressants (eg, cyclosporine), provide symptomatic relief, they face significant challenges including limited efficacy in refractory cases (eg, only 40–50% complete response rate to omalizumab in type IIb autoimmune CSU), development of resistance over time (eg, tachyphylaxis with prolonged antihistamine use), systemic side effects (eg, sedation, gastrointestinal disturbances, or increased infection risk with immunosuppressants), and poor patient adherence due to frequent dosing requirements and incomplete symptom control.153,154 These limitations often result in persistent disease burden, high relapse rates (up to 60% post-treatment), and the need for escalated therapies, underscoring the demand for more targeted approaches.2,155 At present, although the development of CSU immunotherapy based on nanomaterials continues, there remains a need for innovative strategies to bridge the gap between preclinical prospects and clinical translation.156,157 While current applications, such as targeted modulation of mast cells and eosinophils via functionalized nanoparticles, have demonstrated efficacy in alleviating Th2-driven inflammation, several challenges persist, including immunotoxicity, variable patient responses, and limited long-term safety data.158 To address these limitations, we propose a clearer clinical translation roadmap that outlines the sequential steps from preclinical studies, animal models, Phase I/II, and Phase III clinical trials. This roadmap could ensure systematic evaluation, beginning with laboratory validation of nanomaterials and progressing through controlled animal trials, followed by human clinical studies in progressive phases to monitor safety, efficacy, and long-term effects.159 Regulatory frameworks such as FDA and EMA guidelines are critical for nanomedicines, especially with respect to long-term safety monitoring.160 The development of nanotherapies must adhere to specific safety and efficacy endpoints established for Phase I/II trials, while the FDA and EMA provide guidance on nanomaterial toxicity, biocompatibility, and post-market surveillance.161 Addressing ethical concerns, such as equitable access to treatment and informed consent, particularly in vulnerable CSU populations, is essential for translating these therapies into practice.162 For instance, biodegradable PLGA nanoparticles exhibit low immunogenicity in recent airway allergy studies, with no systemic toxicity in long-term exposures.124,163 However, lipid nanoparticles in mRNA vaccines have been linked to urticaria flares, thereby emphasizing the need for optimized formulations to avoid MRGPRX2 activation.164,165 Moreover, rigorous evaluations, including ISO-compliant cytotoxicity tests, show >85% safety scores for functionalized variants, yet chronic accumulation risks persist for metallic nanoparticles, potentially exacerbating eosinophilia. Additionally, long-term immunological impacts balance substantial benefits against risks.166,167 Specifically, nanomaterials induce durable tolerance, as demonstrated by 2025 nanoparticle vaccines that prevent severe allergic reactions via sustained Treg expansion and IgE reduction. Benefits include 60–90% relapse prevention over 12 months in models, but risks like immune exhaustion or paradoxical inflammation necessitate monitoring.168,169 Furthermore, a risk-benefit matrix, quantifies high efficacy (benefit: 8.5/10) against moderate toxicity (risk: 3.5/10) for polymeric systems, yielding net positives in probabilistic models (eg, 80% favorable outcomes for Siglec-targeted nanoparticles in refractory CSU).12 Ethical and regulatory considerations are critical for translation. In particular, frameworks in the EU and US emphasize GLP toxicology and phase-specific endpoints for nanomedicines, addressing persistence risks through post-market surveillance.162,170 Consequently, ethics demand equitable access and consent for long-term exposures, especially in vulnerable CSU populations.76 Lipid nanoparticles in mRNA vaccines have been implicated in urticaria via PEG-induced complement pathways, providing key lessons for CSU nanomaterial safety.171 For instance, delayed systemic urticarial reactions post-mRNA vaccination have been reported, often treatable with antihistamines but indicative of non-life-threatening hypersensitivity, highlighting the need for alternative surface modifications in CSU-targeted nanomaterials to avoid similar flares in sensitive patients.172 Furthermore, chronic urticaria onsets following repeated mRNA immunizations, particularly with Moderna mRNA-1273 occurring on average 10 days post-vaccination, serve as a cautionary example of how nanoparticle formulations might exacerbate underlying autoimmune endotypes in CSU, advocating for pre-clinical screening of MRGPRX2 activation and anti-PEG antibody responses in future designs.173 Additionally, cohort analyses associate mRNA COVID-19 vaccination with increased urticaria risk, including persistent features over time, emphasizing translational gaps and the importance of long-term monitoring in nanomedicine trials, with parallels to CSU’s refractory cases where biologics like omalizumab fail, suggesting biomarker-guided patient stratification to exclude high-risk individuals.174
Additionally, a key challenge in CSU therapy is differentiating between Type I and Type IIb endotypes. The importance of stratifying patients based on their immune profiles, such as distinguishing omalizumab responders from non-responders, is crucial for guiding personalized nanotherapy.175,176 By using endotype-driven models, we can tailor the use of nanomedicines, enhancing efficacy and minimizing adverse effects. This approach aligns with emerging trends in precision medicine, ensuring that patients receive the most suitable therapy based on their immune system characteristics.77,113 To contextualize the translational value of nanomedicine in CSU, it is essential to compare these strategies with existing biologics like omalizumab, dupilumab, and benralizumab.177 While biologics have shown effectiveness in certain patient populations, nanomedicine offers the potential for more targeted, less invasive interventions. Nanomaterials may have advantages in terms of cost-efficiency, especially considering the high cost of biologics, and they may offer better control over side effects due to their precision in targeting immune cells.178 However, the comparison of safety, efficacy, and cost between nanomedicine and biologics is an essential area of ongoing research, as nanotherapy is still in its early stages.179
Future Perspectives
To address these limitations, the use of artificial intelligence (AI), multi-omics analyses, and biomarkers can enhance personalized medicine in nanomedicine.180 These strategies not only aim to optimize nanomaterial design but also to enhance safety profiles and therapeutic precision, thereby paving the way for more effective CSU management. First, AI-guided optimization of nanomaterials represents a pivotal frontier in enhancing their immunological interactions.181 Machine learning algorithms can predict nanoparticle properties, such as size, surface ligand density, and release kinetics, to maximize targeting efficiency toward CSU-specific immune cells like mast cells and basophils.182 For instance, AI models have been employed to simulate nano-immune interfaces, enabling the design of multivalent ligand-displaying nanoparticles that co-engage inhibitory receptors (eg, Siglec-8) and FcεRI, thereby suppressing degranulation with minimal off-target effects.183 Recent studies in nanomedicine emphasize the use of deep learning for high-throughput screening of nanomaterial formulations, which could accelerate the development of CSU-tailored therapies by reducing experimental iterations and predicting patient-specific responses based on endotype classifications.184 Furthermore, integrating AI with computational modeling of protein-nanoparticle interactions may facilitate the creation of adaptive nanomaterials that respond dynamically to inflammatory cues, such as elevated IL-4 or histamine levels in CSU flares.185
In addition, biomarker-driven safety assessments play a crucial role in mitigating the risks associated with nanomaterial use, particularly in immune-mediated disorders such as CSU.186 Biomarkers like serum IgE levels, MRGPRX2 expression on mast cells, and eosinophil-derived proteins (eg, major basic protein) serve as valuable tools for personalized dosing and real-time monitoring of nano-immune interactions.187 Emerging strategies include the use of diagnostic nanoparticles, also known as “theranostics”, which combine therapeutic delivery with imaging capabilities. These nanoparticles enable non-invasive tracking of biodistribution and immune activation, offering an innovative approach for monitoring treatment effects.188 For example, recent advancements in silica-based nanoparticles functionalized with fluorescent probes have made it possible to target biomarkers specifically in allergic models, shedding light on potential exacerbations like complement activation or cytokine storms.189 As a result, prospective clinical trials can utilize these assessments to stratify CSU patients, ensuring safer administration of nanomaterials, especially in refractory cases that do not respond to biologics like omalizumab.189 This approach aligns with ongoing nanomedicine research, which emphasizes regulatory compliance, where biomarker validation through phase I/II studies is essential to address concerns over long-term immunotoxicity, such as unintended Th1/Th2 shifts or nanoparticle accumulation in lymphoid tissues.190
Finally, multi-omics studies provide a comprehensive framework to unravel the complex nano-immune dynamics in CSU, integrating genomics, proteomics, and metabolomics to better understand the mechanisms underlying therapeutic efficacy and potential risks.191 By analyzing omics data from nanomaterial-exposed immune cells, researchers can identify novel pathways, such as epigenetic modifications in Tregs or metabolic reprogramming in eosinophils, that play a crucial role in tolerance induction.192 Recent studies demonstrate the utility of single-cell RNA sequencing in mapping nano-induced transcriptomic changes, revealing how lipid nanoparticles might enhance Treg proliferation while suppressing basophil hyperactivity.192 To this end, future strategies could involve omics-informed nanomaterial redesign, including the incorporation of CRISPR-edited payloads for gene silencing of pro-inflammatory cytokines (eg, IL-13), thereby promoting durable immune tolerance in CSU.193 Additionally, combining multi-omics with AI analytics could enable the prediction of exacerbation risks, such as MRGPRX2-mediated pseudo-allergic reactions, allowing for proactive adjustments in nanomaterial composition for vulnerable patient subsets.194
Beyond these core strategies, emerging hybrid approaches, such as nanomaterial-vaccine conjugates and bio-mimetic systems, hold significant promise for proactive CSU prevention.120 For instance, allergen-encapsulated nanoparticles, which mimic extracellular vesicles, could induce preemptive tolerance in high-risk individuals, drawing from recent successes in food allergy models.93 However, addressing exacerbation risks remains critical, and therefore, rigorous preclinical testing in humanized mouse models is necessary to simulate the heterogeneity of CSU.195 Overall, these forward-looking perspectives emphasize a multidisciplinary paradigm, where collaboration between nanotechnologists, immunologists, and clinicians will drive the translation of nanomaterials from bench to bedside. This approach aims to revolutionize CSU immunotherapy, offering enhanced specificity, reduced relapse rates, and improved patient quality of life.9,196 We believe that more effective methods will emerge in the future to harness nanomaterials for the precise treatment of CSU patients. Meanwhile, to date, fundamental and clinical research exploring the interaction between nanomaterials and urticaria, particularly CSU, remains extremely limited. Future studies are urgently needed to validate these proposed targeting strategies in CSU-specific animal models and, ultimately, in clinical settings.
Conclusion
In conclusion, nanomaterials represent a transformative platform for precision immunotherapy in CSU, with substantial promise in achieving targeted modulation of key immune cells, including mast cells, eosinophils, basophils, and T lymphocytes. Their inherent tunability—encompassing particle size, surface functionalization with ligands such as Siglec-8 or FcεRI inhibitors, and controlled drug release profiles—facilitates precision targeting of pathogenic pathways like FcεRI signaling and Th2-dominant inflammation, thereby enhancing bioavailability, inducing antigen-specific immune tolerance through regulatory T cell expansion, and enabling personalization based on CSU endotypes (eg, Type I autoallergy versus Type IIb autoimmunity). These attributes position nanomaterials as potential superior alternatives to existing biologics, such as omalizumab, particularly in refractory cases where they could offer reduced dosing frequency, minimized systemic immunosuppression, and improved symptom control, as evidenced by preclinical models demonstrating 60–90% inhibition of degranulation and mediator release. Nevertheless, significant limitations must be acknowledged, including safety concerns such as potential immunotoxicity from complement activation or pseudo-allergic reactions triggered by components like PEGylated lipids, translational gaps arising from the predominance of in vitro and animal studies over human trials, and a notable lack of CSU-specific clinical evidence, which currently relies heavily on extrapolations from broader allergic disease models. Addressing these challenges will require rigorous advancements in AI-driven design for predictive toxicity modeling, multi-omics integration to delineate nano-immune interactions, and focused clinical investigations to validate efficacy in stratified patient cohorts, ultimately paving the way for safer, more effective interventions that extend beyond CSU to other allergic disorders and enhance overall patient outcomes.
Abbreviations
CSU, Chronic Spontaneous Urticaria; Th2, T-helper 2 cells; FcεRI, High-affinity IgE receptor; MRGPRX2, Mas-related G protein-coupled receptor X2; Tregs, Regulatory T cells; IgE, Immunoglobulin E; UAS7, Urticaria Activity Score 7; AI, Artificial Intelligence; PLGA, Poly(lactic-co-glycolic acid); hERG, Human Ether-à-go-go-Related Gene; GFP, Green Fluorescent Protein; IL, Interleukin; PAF, Platelet-Activating Factor; TLC, Thin Layer Chromatography; PGA, Prostaglandin A; PDL1, Programmed Death Ligand 1; BPC, Biocompatible Polymer Coating; ACT, Antigen Crosslinking Therapy; FAP, Fibroblast Activation Protein; SNPs, Single Nucleotide Polymorphisms; NK, Natural Killer (cells); ROS, Reactive Oxygen Species; DCs, Dendritic Cells.
Data Sharing Statement
No new data has been generated, all references are cited in the manuscript.
Consent for Publication
All the authors consented for publication.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This study did not receive any funding.
Disclosure
The authors declare that there are no competing interests associated with the manuscript.
References
1. Zuberbier T, Abdul Latiff AH, Abuzakouk M, et al. The international EAACI/GA2LEN/EuroGuiDerm/APAAACI guideline for the definition, classification, diagnosis, and management of urticaria. Allergy. 2022;77(3):734–26. doi:10.1111/all.15090
2. Zuberbier T, Ensina LF, Giménez-Arnau A, et al. Chronic urticaria: unmet needs, emerging drugs, and new perspectives on personalised treatment. Lancet. 2024;404(10450):393–404. doi:10.1016/S0140-6736(24)00852-3
3. Maurer M, Khan DA, Elieh Ali Komi D, Kaplan AP. Biologics for the use in chronic spontaneous urticaria: when and which. J Allergy Clin Immunol Pract. 2021;9(3):1067–1078. doi:10.1016/j.jaip.2020.11.043
4. Asero R, Ferrucci S, Tedeschi A, Cugno M. Biologics for chronic spontaneous urticaria: toward a personalized treatment. Expert Rev Clin Immunol. 2022;18(12):1297–1305. doi:10.1080/1744666X.2022.2138347
5. Bernstein JS, Bernstein JA, Lang DM. Chronic spontaneous urticaria: current and emerging biologic agents. Immunol Allergy Clin North Am. 2024;44(4):595–613. doi:10.1016/j.iac.2024.07.001
6. Tan S, Chen Z, Lin T. Global, regional, and National burden of urticaria (1990–2021), its potential risk factors, and projections to 2046. BMC Public Health. 2025;25(1):2709. doi:10.1186/s12889-025-23321-y
7. Gonçalo M, Gimenéz-Arnau A, Al-Ahmad M, et al. The global burden of chronic urticaria for the patient and society. Br J Dermatol. 2021;184(2):226–236. doi:10.1111/bjd.19561
8. Lenders V, Koutsoumpou X, Sargsian A, Manshian BB. Biomedical nanomaterials for immunological applications: ongoing research and clinical trials. Nanoscale Adv. 2020;2(11):5046–5089. doi:10.1039/d0na00478b
9. Croitoru GA, Pîrvulescu DC, Niculescu AG, et al. Nanomaterials in immunology: bridging innovative approaches in immune modulation, diagnostics, and therapy. J Funct Biomater. 2024;15(8):225. doi:10.3390/jfb15080225
10. Han J, Mao K, Yang Y-G, Sun T. Impact of inorganic/organic nanomaterials on the immune system for disease treatment. Biomater Sci. 2024;12(19):4903–4926. doi:10.1039/d4bm00853g
11. Pohlit H, Bellinghausen I, Frey H, Saloga J. Recent advances in the use of nanoparticles for allergen-specific immunotherapy. Allergy. 2017;72(10):1461–1474. doi:10.1111/all.13199
12. Krishna SS, Farhana SA, A TP, et al. Modulation of immune response by nanoparticle-based immunotherapy against food allergens. Front Immunol. 2023;14:1229667. doi:10.3389/fimmu.2023.1229667
13. Yasinska IM, Calzolai L, Raap U, et al. Targeting of basophil and mast cell pro-allergic reactivity using functionalised gold nanoparticles. Front Pharmacol. 2019:10. doi:10.3389/fphar.2019.00333
14. Duan S, Arlian BM, Nycholat CM, et al. Nanoparticles displaying allergen and siglec-8 ligands suppress IgE-FcεRI-mediated anaphylaxis and desensitize mast cells to subsequent antigen challenge. J Immunol. 2021;206(10):2290–2300. doi:10.4049/jimmunol.1901212
15. Du F, Rische CH, Li Y, et al. Controlled adsorption of multiple bioactive proteins enables targeted mast cell nanotherapy. Nat Nanotechnol. 2024;19(5):698–704. doi:10.1038/s41565-023-01584-z
16. Rad LM, Arellano G, Podojil JR, O’Konek JJ, Shea LD, Miller SD. Engineering nanoparticle therapeutics for food allergy. J Allergy Clin Immunol. 2024;153(3):549–559. doi:10.1016/j.jaci.2023.10.013
17. Abo-Zaid MA, Elsapagh RM, Sultan NS, Mawkili W, Hegazy MM, Ismail AH. Allergy treatment: a comprehensive review of nanoparticle-based allergen immunotherapy. Front Biosci. 2025;30(3):26550. doi:10.31083/FBL26550
18. Zhou B, Li J, Liu R, Zhu L, Peng C. The role of crosstalk of immune cells in pathogenesis of chronic spontaneous urticaria. Front Immunol. 2022;13:879754. doi:10.3389/fimmu.2022.879754
19. Saini SS, Asero R, Cugno M, Park HS, Oliver ET. Pathogenesis of chronic spontaneous urticaria with or without angioedema. J Allergy Clin Immunol Pract. 2025;13(9):2221–2228. doi:10.1016/j.jaip.2025.07.025
20. Criado PR, Criado RFJ, Miot HA, Abdalla BMZ, Marchioro HZ, Bonamigo RR. Chronic spontaneous urticaria: update on pathogenesis and therapeutic implications. Anais Brasileiros de Dermatologia. 2025;100(5):501198. doi:10.1016/j.abd.2025.501198
21. Kolkhir P, Giménez-Arnau AM, Kulthanan K, Peter J, Metz M, Maurer M. Urticaria. Nat Rev Dis Primers. 2022;8(1):61. doi:10.1038/s41572-022-00389-z
22. Chen Y, Chen X, Zhang Z. Association between immune cells and urticaria: a bidirectional mendelian randomization study. Front Immunol. 2024;15:1439331. doi:10.3389/fimmu.2024.1439331
23. Kolkhir P, Fok JS, Kocatürk E, et al. Update on the treatment of chronic spontaneous urticaria. Drugs. 2025;85(4):475–486. doi:10.1007/s40265-025-02170-4
24. Kaplan AP, Ferrer M. An algorithm for the diagnosis, pathogenesis and treatment of chronic spontaneous urticaria, 2024 update. Allergy. 2024;79(9):2567–2569. doi:10.1111/all.16113
25. Elieh-Ali-Komi D, Metz M, Kolkhir P, et al. Chronic urticaria and the pathogenic role of mast cells. Allergol Int. 2023;72(3):359–368. doi:10.1016/j.alit.2023.05.003
26. Bernstein JA, Maurer M, Saini SS. BTK signaling-a crucial link in the pathophysiology of chronic spontaneous urticaria. J Allergy Clin Immunol. 2024;153(5):1229–1240. doi:10.1016/j.jaci.2023.12.008
27. Asero R. Mechanisms of histamine release from mast cells beyond the high affinity IgE receptor in severe chronic spontaneous urticaria. Immunol Lett. 2024;265:1–4. doi:10.1016/j.imlet.2023.11.008
28. Yanase Y, Takahagi S, Ozawa K, Hide M. The role of coagulation and complement factors for mast cell activation in the pathogenesis of chronic spontaneous urticaria. Cells. 2021;10(7):1759. doi:10.3390/cells10071759
29. Giménez-Arnau AM, DeMontojoye L, Asero R, et al. The pathogenesis of chronic spontaneous urticaria: the role of infiltrating cells. J Allergy Clin Immunol Pract. 2021;9(6):2195–2208. doi:10.1016/j.jaip.2021.03.033
30. Altrichter S, Frischbutter S, Fok JS, et al. The role of eosinophils in chronic spontaneous urticaria. J Allergy Clin Immunol. 2020;145(6):1510–1516. doi:10.1016/j.jaci.2020.03.005
31. Kolkhir P, Church MK, Altrichter S, et al. Eosinopenia, in chronic spontaneous urticaria, is associated with high disease activity, autoimmunity, and poor response to treatment. J Allergy Clin Immunol Pract. 2020;8(1):318–325.e5. doi:10.1016/j.jaip.2019.08.025
32. Kaplan A, Lebwohl M, Giménez-Arnau AM, Hide M, Armstrong AW, Maurer M. Chronic spontaneous urticaria: focus on pathophysiology to unlock treatment advances. Allergy. 2023;78(2):389–401. doi:10.1111/all.15603
33. A S, Am GA, Ja B, et al. Benralizumab does not elicit therapeutic effect in patients with chronic spontaneous urticaria: results from the phase IIb multinational randomized double-blind placebo-controlled ARROYO trial. Br J Dermatol. 2024;191(2). doi:10.1093/bjd/ljae067
34. Kay AB, Ying S, Ardelean E, et al. Elevations in vascular markers and eosinophils in chronic spontaneous urticarial weals with low-level persistence in uninvolved skin. Br J Dermatol. 2014;171(3):505–511. doi:10.1111/bjd.12991
35. Konstantinou GN, Riedl MA, Valent P, Podder I, Maurer M. Urticaria and angioedema: understanding complex pathomechanisms to facilitate patient communication, disease management, and future treatment. J Allergy Clin Immunol Pract. 2023;11(1):94–106. doi:10.1016/j.jaip.2022.11.006
36. Yanase Y, Matsubara D, Takahagi S, Tanaka A, Ozawa K, Hide M. Basophil characteristics as a marker of the pathogenesis of chronic spontaneous urticaria in relation to the coagulation and complement systems. Int J Mol Sci. 2023;24(12):10320. doi:10.3390/ijms241210320
37. Bracken SJ, Abraham S, MacLeod AS. Autoimmune theories of chronic spontaneous urticaria. Front Immunol. 2019;10:627. doi:10.3389/fimmu.2019.00627
38. Kishimoto I, Ma N, Takimoto-Ito R, et al. Decreased peripheral basophil counts in urticaria and mouse model of oxazolone-induced hypersensitivity, the latter suggesting basopenia reflecting migration to skin. Front Immunol. 2022;13:1014924. doi:10.3389/fimmu.2022.1014924
39. Jiao Q, Lohse K, Rauber-Ellinghaus MM, et al. Basophil and eosinophil recruitment to skin lesions is linked to features of autoimmune chronic spontaneous urticaria including basopenia. J Eur Acad Dermatol Venereol. 2025;39(2):e155–e157. doi:10.1111/jdv.20204
40. Matsubara D, Yanase Y, Ishii K, et al. Basophils activation of patients with chronic spontaneous urticaria in response to C5a despite failure to respond to IgE-mediated stimuli. Front Immunol. 2022;13:994823. doi:10.3389/fimmu.2022.994823
41. Wiebe D, Limberg MM, Gray N, Raap U. Basophils in pruritic skin diseases. Front Immunol. 2023;14:1213138. doi:10.3389/fimmu.2023.1213138
42. Ruppenstein A, Limberg MM, Loser K, Kremer AE, Homey B, Raap U. Involvement of neuro-immune interactions in pruritus with special focus on receptor expressions. Front Med. 2021;8:627985. doi:10.3389/fmed.2021.627985
43. Vena GA, Cassano N, Di Leo E, Calogiuri GF, Nettis E. Focus on the role of substance P in chronic urticaria. Clin Mol Allergy. 2018;16:24. doi:10.1186/s12948-018-0101-z
44. McHale C, Mohammed Z, Deppen J, Gomez G. Interleukin-6 potentiates FcεRI-induced PGD2 biosynthesis and induces VEGF from human in situ-matured skin mast cells. Biochim Biophys Acta. 2018;1862(5):1069–1078. doi:10.1016/j.bbagen.2018.01.020
45. Nikolouli E, Mommert S, Dawodu DM, et al. The stimulation of TH2 cells results in increased IL-5 and IL-13 production via the H4 receptor. Allergy. 2024;79(8):2186–2196. doi:10.1111/all.16182
46. Fangal VD, Kılıç A, Mirzakhani H, et al. Vitamin D exerts endogenous control over TH2 cell fate and immune plasticity. iScience. 2025;28(4):112117. doi:10.1016/j.isci.2025.112117
47. Can Bostan O, Damadoglu E, Sarac BE, et al. Cytokine profiles of chronic urticaria patients and the effect of omalizumab treatment. Dermatol Pract Concept. 2023;13(4):e2023272. doi:10.5826/dpc.1304a272
48. Moy AP, Murali M, Nazarian RM. Identification of a Th2- and Th17-skewed immune phenotype in chronic urticaria with Th22 reduction dependent on autoimmunity and thyroid disease markers. J Cutan Pathol. 2016;43(4):372–378. doi:10.1111/cup.12673
49. Hoşgören-Tekin S, Eyüboğlu İP, Akkiprik M, Giménez-Arnau AM, Salman A. Inflammatory cytokine levels and changes during omalizumab treatment in chronic spontaneous urticaria. Arch Dermatol Res. 2024;316(6):261. doi:10.1007/s00403-024-02966-6
50. Yang X, Chen L, Wang S, Wu Y, Zhou X, Meng Z. The correlation between Th17/Treg immune dysregulation and the disease severity in chronic spontaneous urticaria patients. Immun Inflamm Dis. 2023;11(7):e920. doi:10.1002/iid3.920
51. Kiwamoto T, Kawasaki N, Paulson JC, Bochner BS. Siglec-8 as a drugable target to treat eosinophil and mast cell-associated conditions. Pharmacol Ther. 2012;135(3):327–336. doi:10.1016/j.pharmthera.2012.06.005
52. Cao TBT, Cha HY, Yang EM, Ye YM. Elevated MRGPRX2 levels related to disease severity in patients with chronic spontaneous urticaria. Allergy Asthma Immunol Res. 2021;13(3):498–506. doi:10.4168/aair.2021.13.3.498
53. Dunford PJ, O’Donnell N, Riley JP, Williams KN, Karlsson L, Thurmond RL. The histamine H4 receptor mediates allergic airway inflammation by regulating the activation of CD4+ T cells. J Immunol. 2006;176(11):7062–7070. doi:10.4049/jimmunol.176.11.7062
54. Lu W, Cao J, Lv Y, Li M, Tang Y, Feng Y. Progress and prospects of transdermal treatment of allergic skin diseases with natural drugs based on nanotechnology. AAPS Pharm Sci Tech. 2025;27(1):21. doi:10.1208/s12249-025-03244-y
55. Li K, Jin J, Yang Y, et al. Application of nanoparticles for immunotherapy of allergic rhinitis. Int J Nanomed. 2024;19:12015–12037. doi:10.2147/IJN.S484327
56. Bochner BS. Siglec-8 on human eosinophils and mast cells, and Siglec-F on murine eosinophils, are functionally related inhibitory receptors. Clin Exp Allergy. 2009;39(3):317–324. doi:10.1111/j.1365-2222.2008.03173.x
57. Altrichter S, Staubach P, Pasha M, et al. An open-label, proof-of-concept study of lirentelimab for antihistamine-resistant chronic spontaneous and inducible urticaria. J Allergy Clin Immunol. 2022;149(5):1683–1690.e7. doi:10.1016/j.jaci.2021.12.772
58. Dema B, Suzuki R, Rivera J. Rethinking the role of immunoglobulin E and its high-affinity receptor: new insights into allergy and beyond. Int Arch Allergy Immunol. 2014;164(4):271–279. doi:10.1159/000365633
59. Hide M, Igarashi A, Yagami A, et al. Efficacy and safety of omalizumab for the treatment of refractory chronic spontaneous urticaria in Japanese patients: subgroup analysis of the phase 3 POLARIS study. Allergol Int. 2018;67(2):243–252. doi:10.1016/j.alit.2017.10.001
60. Subramanian H, Gupta K, Ali H. Roles of Mas-related G protein-coupled receptor X2 on mast cell-mediated host defense, pseudoallergic drug reactions, and chronic inflammatory diseases. J Allergy Clin Immunol. 2016;138(3):700–710. doi:10.1016/j.jaci.2016.04.051
61. Azimi E, Reddy VB, Lerner EA. Brief communication: MRGPRX2, atopic dermatitis and red man syndrome. Itch. 2017;2(1):e5. doi:10.1097/itx.0000000000000005
62. Grekowitz E, Metz M, Altrichter S, et al. Targeting histamine receptor 4 in cholinergic urticaria with izuforant (LEO 152020): results from a phase IIa randomized double-blind placebo-controlled multicentre crossover trial. Br J Dermatol. 2024;190(6):825–835. doi:10.1093/bjd/ljae038
63. Ling P, Ngo K, Nguyen S, et al. Histamine H4 receptor mediates eosinophil chemotaxis with cell shape change and adhesion molecule upregulation. Br J Pharmacol. 2004;142(1):161–171. doi:10.1038/sj.bjp.0705729
64. Hofstra CL, Desai PJ, Thurmond RL, Fung-Leung WP. Histamine H4 receptor mediates chemotaxis and calcium mobilization of mast cells. J Pharmacol Exp Ther. 2003;305(3):1212–1221. doi:10.1124/jpet.102.046581
65. Korver W, Wong A, Gebremeskel S, et al. The inhibitory receptor siglec-8 interacts with FcεRI and globally inhibits intracellular signaling in primary mast cells upon activation. Front Immunol. 2022;13:833728. doi:10.3389/fimmu.2022.833728
66. Schanin J, Gebremeskel S, Korver W, et al. A monoclonal antibody to Siglec-8 suppresses non-allergic airway inflammation and inhibits IgE-independent mast cell activation. Mucosal Immunol. 2021;14(2):366–376. doi:10.1038/s41385-020-00336-9
67. Nagata Y, Suzuki R. FcεRI: a master regulator of mast cell functions. Cells. 2022;11(4):622. doi:10.3390/cells11040622
68. Metz M. No allergy, but mast cells are involved: MRGPRX2 in chronic inflammatory skin diseases. J Eur Acad Dermatol Venereol. 2025;39(3):451–452. doi:10.1111/jdv.20541
69. Ye D, Zhang Y, Zhao X, et al. MRGPRX2 gain-of-function mutation drives enhanced mast cell reactivity in chronic spontaneous urticaria. J Allergy Clin Immunol. 2025;156(1):159–170. doi:10.1016/j.jaci.2025.03.007
70. Macphee CH, Dong X, Peng Q, et al. Pharmacological blockade of the mast cell MRGPRX2 receptor supports investigation of its relevance in skin disorders. Front Immunol. 2024;15:1433982. doi:10.3389/fimmu.2024.1433982
71. Moyano DF, Goldsmith M, Solfiell DJ, et al. Nanoparticle hydrophobicity dictates immune response. J Am Chem Soc. 2012;134(9):3965–3967. doi:10.1021/ja2108905
72. Hernroth B, Krång AS, Baden S. Bacteriostatic suppression in norway lobster (Nephrops norvegicus) exposed to manganese or hypoxia under pressure of ocean acidification. Aquat Toxicol. 2015;159:217–224. doi:10.1016/j.aquatox.2014.11.025
73. Maurer M, Rosén K, Hsieh HJ, et al. Omalizumab for the treatment of chronic idiopathic or spontaneous urticaria. N Engl J Med. 2013;368(10):924–935. doi:10.1056/NEJMoa1215372
74. Li S, Yang L, Zhao Z, Yang X, Lv H. A polyurethane-based hydrophilic elastomer with multi-biological functions for small-diameter vascular grafts. Acta Biomater. 2024;176:234–249. doi:10.1016/j.actbio.2024.01.006
75. Agache I, Song Y, Alonso-Coello P, et al. Efficacy and safety of treatment with biologicals for severe chronic rhinosinusitis with nasal polyps: a systematic review for the EAACI guidelines. Allergy. 2021;76(8):2337–2353. doi:10.1111/all.14809
76. Csóka I, Ismail R, Jójárt-Laczkovich O, Pallagi E. Regulatory considerations, challenges and risk-based approach in nanomedicine development. Curr Med Chem. 2021;28(36):7461–7476. doi:10.2174/0929867328666210406115529
77. Lang DM, Sheikh J, Joshi S, Bernstein JA. Endotypes, phenotypes, and biomarkers in chronic spontaneous urticaria: evolving toward personalized medicine. Ann Allergy Asthma Immunol. 2025;134(4):408–417.e3. doi:10.1016/j.anai.2024.10.026
78. Rische CH, Thames AN, Krier-Burris RA, O’Sullivan JA, Bochner BS, Scott EA. Drug delivery targets and strategies to address mast cell diseases. Expert Opin Drug Deliv. 2023;20(2):205–222. doi:10.1080/17425247.2023.2166926
79. Li T, Tolksdorf F, Sung W, et al. Arginine-based cationic liposomes accelerate T cell activation and differentiation in vitro. Int J Pharm. 2022;623:121917. doi:10.1016/j.ijpharm.2022.121917
80. Johnson L, Aglas L, Punz B, et al. Mechanistic insights into silica nanoparticle–allergen interactions on antigen presenting cell function in the context of allergic reactions. Nanoscale. 2023;15(5):2262. doi:10.1039/d2nr05181h
81. Lin G, Wang J, Yang YG, Zhang Y, Sun T. Advances in dendritic cell targeting nano-delivery systems for induction of immune tolerance. Front Bioeng Biotechnol. 2023;11:1242126. doi:10.3389/fbioe.2023.1242126
82. Kratzer B, Hofer S, Zabel M, Pickl WF. All the small things: how virus‐like particles and liposomes modulate allergic immune responses. Eur J Immunol. 2020;50(1):17–32. doi:10.1002/eji.201847810
83. González-Fernández Á, Maietta I. The forgotten innate immune cells: unraveling their prospective interactions with nanomaterials. Int J Nanomed. 2025;20:8173–8189. doi:10.2147/IJN.S517249
84. Scheiblhofer S, Machado Y, Feinle A, Thalhamer J, Hüsing N, Weiss R. Potential of nanoparticles for allergen-specific immunotherapy - use of silica nanoparticles as vaccination platform. Expert Opin Drug Deliv. 2016;13(12):1777–1788. doi:10.1080/17425247.2016.1203898
85. Roach KA, Stefaniak AB, Roberts JR. Metal nanomaterials: immune effects and implications of physicochemical properties on sensitization, elicitation, and exacerbation of allergic disease. J Immunotoxicol. 2019;16(1):87–124. doi:10.1080/1547691X.2019.1605553
86. Palazzolo S, Bayda S, Hadla M, et al. The clinical translation of organic nanomaterials for cancer therapy: a focus on polymeric nanoparticles, micelles, liposomes and exosomes. Curr Med Chem. 2018;25(34):4224–4268. doi:10.2174/0929867324666170830113755
87. Zhu X, Li S. Nanomaterials in tumor immunotherapy: new strategies and challenges. Mol Cancer. 2023;22(1):94. doi:10.1186/s12943-023-01797-9
88. Krishnamurthy S, Vaiyapuri R, Zhang L, Chan JM. Lipid-coated polymeric nanoparticles for cancer drug delivery. Biomater Sci. 2015;3(7):923–936. doi:10.1039/c4bm00427b
89. Orecchioni M, Bedognetti D, Sgarrella F, Marincola FM, Bianco A, Delogu LG. Impact of carbon nanotubes and graphene on immune cells. J Transl Med. 2014;12:138. doi:10.1186/1479-5876-12-138
90. Wang X, Podila R, Shannahan JH, Rao AM, Brown JM. Intravenously delivered graphene nanosheets and multiwalled carbon nanotubes induce site-specific Th2 inflammatory responses via the IL-33/ST2 axis. Int J Nanomed. 2013;8:1733–1748. doi:10.2147/IJN.S44211
91. Wang Y, Li J, Li X, Shi J, Jiang Z, Zhang CY. Graphene-based nanomaterials for cancer therapy and anti-infections. Bioact Mater. 2022;14:335–349. doi:10.1016/j.bioactmat.2022.01.045
92. Cao W, Xia T. mRNA lipid nanoparticles induce immune tolerance to treat human diseases. Med Rev. 2023;3(2):180–183. doi:10.1515/mr-2023-0008
93. Saunders MN, Rad LM, Williams LA, et al. Allergen-encapsulating nanoparticles reprogram pathogenic allergen-specific Th2 cells to suppress food allergy. Adv Healthc Mater. 2025;14(5):e2400237. doi:10.1002/adhm.202400237
94. Liu M, Thijssen S, Hennink WE, Garssen J, van Nostrum CF, Willemsen LEM. Oral pretreatment with β-lactoglobulin derived peptide and CpG co-encapsulated in PLGA nanoparticles prior to sensitizations attenuates cow’s milk allergy development in mice. Front Immunol. 2022;13:1053107. doi:10.3389/fimmu.2022.1053107
95. Laddha UD, Kshirsagar SJ. Formulation of PPAR-gamma agonist as surface modified PLGA nanoparticles for non-invasive treatment of diabetic retinopathy: in vitro and in vivo evidences. Heliyon. 2020;6(8):e04589. doi:10.1016/j.heliyon.2020.e04589
96. Ebrahimi M, Asadi M, Akhavan O. Graphene-based nanomaterials in fighting the most challenging viruses and immunogenic disorders. ACS Biomater Sci Eng. 2022;8(1):54–81. doi:10.1021/acsbiomaterials.1c01184
97. Aldossari AA, Shannahan JH, Podila R, Brown JM. Influence of physicochemical properties of silver nanoparticles on mast cell activation and degranulation. Toxicol In Vitro. 2015;29(1):195–203. doi:10.1016/j.tiv.2014.10.008
98. Bawazir M, Amponnawarat A, Hui Y, Oskeritzian CA, Ali H. Inhibition of MRGPRX2 but not FcεRI or MrgprB2-mediated mast cell degranulation by a small molecule inverse receptor agonist. Front Immunol. 2022;13:1033794. doi:10.3389/fimmu.2022.1033794
99. Punz B, Johnson L, Geppert M, et al. Surface functionalization of silica nanoparticles: strategies to optimize the immune-activating profile of carrier platforms. Pharmaceutics. 2022;14(5):1103. doi:10.3390/pharmaceutics14051103
100. Longo A, Longo V, Colombo P. Nanoparticles in allergen immunotherapy. Curr Opin Allergy Clin Immunol. 2021;21(6):576–582. doi:10.1097/ACI.0000000000000782
101. Gu Y, Zhao Y, Zhang Z, et al. An antibody-like polymeric nanoparticle removes intratumoral galectin-1 to enhance antitumor T-Cell responses in cancer immunotherapy. ACS Appl Mater Interfaces. 2021;13(19):22159–22168. doi:10.1021/acsami.1c02116
102. Zemelka-Wiacek M, Jutel M. AIT 2023: current innovation and future outlook. Allergol Select. 2023;7:219–228. doi:10.5414/ALX02379E
103. Mayorga C, Perez-Inestrosa E, Rojo J, Ferrer M, Montañez MI. Role of nanostructures in allergy: diagnostics, treatments and safety. Allergy. 2021;76(11):3292–3306. doi:10.1111/all.14764
104. van den Hoven JM, Nemes R, Metselaar JM, et al. Complement activation by PEGylated liposomes containing prednisolone. Eur J Pharm Sci. 2013;49(2):265–271. doi:10.1016/j.ejps.2013.03.007
105. Korver W, Benet Z, Wong A, et al. Regulation of mast cells by overlapping but distinct protein interactions of Siglec-6 and Siglec-8. Allergy. 2024;79(3):629–642. doi:10.1111/all.16004
106. Wang J, Xian M, Cao H, et al. Prophylactic and therapeutic potential of magnolol-loaded PLGA-PEG nanoparticles in a chronic murine model of allergic asthma. Front Bioeng Biotechnol. 2023;11:1182080. doi:10.3389/fbioe.2023.1182080
107. Celebi Sozener Z, Ozdel Ozturk B, Cerci P, et al. Epithelial barrier hypothesis: effect of the external exposome on the microbiome and epithelial barriers in allergic disease. Allergy. 2022;77(5):1418–1449. doi:10.1111/all.15240
108. Oyarzun-Ampuero FA, Brea J, Loza MI, Torres D, Alonso MJ. Chitosan-hyaluronic acid nanoparticles loaded with heparin for the treatment of asthma. Int J Pharm. 2009;381(2):122–129. doi:10.1016/j.ijpharm.2009.04.009
109. Hong J, Xiao X, Gao Q, et al. Co-delivery of allergen epitope fragments and R848 inhibits food allergy by inducing tolerogenic dendritic cells and regulatory T cells. Int J Nanomed. 2019;14:7053–7064. doi:10.2147/IJN.S215415
110. Saunders MN, Rival CM, Mandal M, et al. Immunotherapy with biodegradable nanoparticles encapsulating the oligosaccharide galactose-alpha-1,3-galactose enhance immune tolerance against alpha-gal sensitization in a murine model of alpha-gal syndrome. Front Allergy. 2024:5. doi:10.3389/falgy.2024.1437523
111. Smarr CB, Yap WT, Neef TP, et al. Biodegradable antigen-associated PLG nanoparticles tolerize Th2-mediated allergic airway inflammation pre- and postsensitization. Proc Natl Acad Sci U S A. 2016;113(18):5059–5064. doi:10.1073/pnas.1505782113
112. Di Gioacchino M, Petrarca C, Gatta A, et al. Nanoparticle-based immunotherapy: state of the art and future perspectives. Expert Rev Clin Immunol. 2020;16(5):513–525. doi:10.1080/1744666X.2020.1762572
113. Segú-Vergés C, Gómez J, Terradas-Montana P, et al. Unveiling chronic spontaneous urticaria pathophysiology through systems biology. J Allergy Clin Immunol. 2023;151(4):1005–1014. doi:10.1016/j.jaci.2022.12.809
114. Casale TB. Novel biologics for treatment of chronic spontaneous urticaria. J Allergy Clin Immunol. 2022;150(6):1256–1259. doi:10.1016/j.jaci.2022.06.027
115. Wedi B. Biologic and small molecule therapies in chronic spontaneous urticaria: an update. Curr Opin Allergy Clin Immunol. 2025;25(5):418–425. doi:10.1097/ACI.0000000000001095
116. Friedrich RP, Cicha I, Tietze R, et al. Intranasal delivery of nanoparticles. Nanomedicine. 2022;17(2):71–76. doi:10.2217/nnm-2021-0418
117. Zettl I, Bauernfeind C, Kollárová J, Flicker S. Single-domain antibodies-novel tools to study and treat allergies. Int J Mol Sci. 2024;25(14):7602. doi:10.3390/ijms25147602
118. Souza Rebouças J D, Esparza I, Ferrer M, Sanz ML, Irache JM, Gamazo C. Nanoparticulate Adjuvants and Delivery Systems for Allergen Immunotherapy. J Biomed Biotechnol. 2012;2012:474605. doi:10.1155/2012/474605
119. Li S, Toriumi H, Takahashi D, et al. Safe and efficient oral allergy immunotherapy using one-pot-prepared mannan-coated allergen nanoparticles. Biomaterials. 2023;303:122381. doi:10.1016/j.biomaterials.2023.122381
120. Johnson L, Duschl A, Himly M. Nanotechnology-based vaccines for allergen-specific immunotherapy: potentials and challenges of conventional and novel adjuvants under research. Vaccines. 2020;8(2):237. doi:10.3390/vaccines8020237
121. Liu J, Liu Z, Pang Y, Zhou H. The interaction between nanoparticles and immune system: application in the treatment of inflammatory diseases. J Nanobiotechnology. 2022;20(1):127. doi:10.1186/s12951-022-01343-7
122. Arizmendi N, Qian H, Li Y, Kulka M. Sesquiterpene-loaded co-polymer hybrid nanoparticle effects on human mast cell surface receptor expression, granule contents, and degranulation. Nanomaterials. 2021;11(4):953. doi:10.3390/nano11040953
123. Sharma P, Dhanjal DS, Chopra C, et al. Targeting eosinophils in chronic respiratory diseases using nanotechnology-based drug delivery. Chem Biol Interact. 2022;365:110050. doi:10.1016/j.cbi.2022.110050
124. Scalise MC, Mutlu S, Ferrié C, et al. Modulation of allergic airways disease employing bio-mimetic nanoparticles with TLR agonists. Front Allergy. 2025;6:1633293. doi:10.3389/falgy.2025.1633293
125. Jin H, Li J, Zhang M, et al. Berberine-loaded biomimetic nanoparticles attenuate inflammation of experimental allergic asthma via enhancing IL-12 expression. Front Pharmacol. 2021;12:724525. doi:10.3389/fphar.2021.724525
126. Shahgordi S, Sankian M, Yazdani Y, et al. Immune responses modulation by curcumin and allergen encapsulated into PLGA nanoparticles in mice model of rhinitis allergic through sublingual immunotherapy. Int Immunopharmacol. 2020;84:106525. doi:10.1016/j.intimp.2020.106525
127. Rani A, Balandin D, Ravi R, et al. Peptide-based lipid nanodiscs suppress eosinophil recruitment and chemotaxis. J Control Release. 2025;385:114060. doi:10.1016/j.jconrel.2025.114060
128. Sadeghi M, Koushki K, Mashayekhi K, et al. DC-targeted gold nanoparticles as an efficient and biocompatible carrier for modulating allergic responses in sublingual immunotherapy. Int Immunopharmacol. 2020;86:106690. doi:10.1016/j.intimp.2020.106690
129. Tu Z, Lin J, Li Z, et al. Tackling allergic airway inflammation with organic sheet-like nanoplatforms by targeted elimination of epithelial small extracellular vesicles. Adv Sci. 2025:e04197. doi:10.1002/advs.202504197
130. Ballester M, Jeanbart L, de Titta A, et al. Nanoparticle conjugation enhances the immunomodulatory effects of intranasally delivered CpG in house dust mite-allergic mice. Sci Rep. 2015;5:14274. doi:10.1038/srep14274
131. Fang L, Nikfarjam N, Gharagozlou M, et al. Pulmonary delivery of levamisole nanoparticles as an immunomodulator affecting Th and a potential ADAM10 inhibitor to ameliorate severe allergic asthma. ACS Biomater Sci Eng. 2022;8(10):4566–4576. doi:10.1021/acsbiomaterials.2c00843
132. Shahzad KA, Wang Z, Li X, Li J, Xu M, Tan F. Immunomodulatory effect of PLGA-encapsulated mesenchymal stem cells-derived exosomes for the treatment of allergic rhinitis. Front Immunol. 2024;15:1429442. doi:10.3389/fimmu.2024.1429442
133. Di Gioacchino M, Di Giampaolo L, Mangifesta R, Gangemi S, Petrarca C. Exposure to nanoparticles and occupational allergy. Curr Opin Allergy Clin Immunol. 2022;22(2):55–63. doi:10.1097/ACI.0000000000000818
134. Peng X, Ge Y, Li W, et al. Targeting Lewis X oligosaccharide-modified liposomes encapsulated with house dust mite allergen Der f 2 to dendritic cells inhibits Th2 immune response. Eur J Pharm Sci. 2023;190:106570. doi:10.1016/j.ejps.2023.106570
135. Ryan JJ, Bateman HR, Stover A, et al. Fullerene nanomaterials inhibit the allergic response. J Immunol. 2007;179(1):665–672. doi:10.4049/jimmunol.179.1.665
136. Di Felice G, Colombo P. Nanoparticle–allergen complexes for allergen immunotherapy. Int J Nanomed. 2017;12:4493–4504. doi:10.2147/IJN.S134630
137. Kostadinova AI, Middelburg J, Ciulla M, et al. PLGA nanoparticles loaded with beta-lactoglobulin-derived peptides modulate mucosal immunity and may facilitate cow’s milk allergy prevention. Eur J Pharmacol. 2018;818:211–220. doi:10.1016/j.ejphar.2017.10.051
138. Troelnikov A, Perkins G, Yuson C, et al. Basophil reactivity to BNT162b2 is mediated by PEGylated lipid nanoparticles in patients with PEG allergy. J Allergy Clin Immunol. 2021;148(1):91–95. doi:10.1016/j.jaci.2021.04.032
139. Mayorga C, Perez-Inestrosa E, Molina N, Montañez MI. Development of nanostructures in the diagnosis of drug hypersensitivity reactions. Curr Opin Allergy Clin Immunol. 2016;16(4):300–307. doi:10.1097/ACI.0000000000000282
140. Gil-Ocaña V, Jimenez IM, Mayorga C, et al. Multiepitope dendrimeric antigen-silica particle composites as nano-based platforms for specific recognition of IgEs. Front Immunol. 2021;12:750109. doi:10.3389/fimmu.2021.750109
141. Tenchov R, Sasso JM, Zhou QA. PEGylated lipid nanoparticle formulations: immunological safety and efficiency perspective. Bioconjug Chem. 2023;34(6):941–960. doi:10.1021/acs.bioconjchem.3c00174
142. Paris JL, de la Torre P, Flores AI. New therapeutic approaches for allergy: a review of cell therapy and bio- or nano-material-based strategies. Pharmaceutics. 2021;13(12):2149. doi:10.3390/pharmaceutics13122149
143. Podojil JR, Genardi S, Chiang MY, et al. Tolerogenic immune-modifying nanoparticles encapsulating multiple recombinant pancreatic β Cell proteins prevent onset and progression of type 1 diabetes in nonobese diabetic mice. J Immunol. 2022;209(3):465–475. doi:10.4049/jimmunol.2200208
144. Dang BTN, Duwa R, Lee S, et al. Targeting tumor-associated macrophages with mannosylated nanotherapeutics delivering TLR7/8 agonist enhances cancer immunotherapy. J Control Release. 2024;372:587–608. doi:10.1016/j.jconrel.2024.06.062
145. Asero R, Calzari P, Vaienti S, Cugno M. Therapies for Chronic spontaneous urticaria: present and future developments. Pharmaceuticals. 2024;17(11):1499. doi:10.3390/ph17111499
146. Tian Y, Wu X, Sun X, Rong L, Han K. Engineered strategies for the allergic disease therapy. Interd Med. 2025;3:e20240115. doi:10.1002/INMD.20240115
147. Lee JS, Oh E, Oh H, et al. Tacrolimus-loaded chitosan-based nanoparticles as an efficient topical therapeutic for the effective treatment of atopic dermatitis symptoms. Int J Biol Macromol. 2024;273(Pt 1):133005. doi:10.1016/j.ijbiomac.2024.133005
148. Ci Z, Wang H, Luo J, et al. Application of nanomaterials targeting immune cells in the treatment of chronic inflammation. Int J Nanomed. 2024;19:13925–13946. doi:10.2147/IJN.S497590
149. Keshavarz Shahbaz S, Varasteh AR, Koushki K, et al. Sublingual dendritic cells targeting by aptamer: possible approach for improvement of sublingual immunotherapy efficacy. Int Immunopharmacol. 2020;85:106603. doi:10.1016/j.intimp.2020.106603
150. Liu Q, Wang X, Liu X, et al. Use of polymeric nanoparticle platform targeting the liver to induce treg-mediated antigen-specific immune tolerance in a pulmonary allergen sensitization model. ACS Nano. 2019;13(4):4778–4794. doi:10.1021/acsnano.9b01444
151. Liu Q, Wang X, Liu X, et al. Antigen- and epitope-delivering nanoparticles targeting liver induce comparable immunotolerance in allergic airway disease and anaphylaxis as nanoparticle-delivering pharmaceuticals. ACS Nano. 2021;15(1):1608–1626. doi:10.1021/acsnano.0c09206
152. Lee NK, Kim SN, Park CG. Immune cell targeting nanoparticles: a review. Biomater Res. 2021;25(1):44. doi:10.1186/s40824-021-00246-2
153. Maurer M, Casale TB, Saini SS, et al. Dupilumab in patients with chronic spontaneous urticaria (LIBERTY-CSU CUPID): two randomized, double-blind, placebo-controlled, phase 3 trials. J Allergy Clin Immunol. 2024;154(1):184–194. doi:10.1016/j.jaci.2024.01.028
154. Kolkhir P, Bonnekoh H, Metz M, Maurer M. Chronic Spontaneous Urticaria: a Review. JAMA. 2024;332(17):1464–1477. doi:10.1001/jama.2024.15568
155. Sánchez J, Pite H, Gómez RM, et al. Chronic spontaneous urticaria remission definition and therapy stepping down: world allergy organization position paper. J Allergy Clin Immunol. 2025;155(3):1050–1056.e2. doi:10.1016/j.jaci.2024.11.039
156. Mohkam M, Sadraeian M, Lauto A, et al. Exploring the potential and safety of quantum dots in allergy diagnostics. Microsyst Nanoeng. 2023;9:145. doi:10.1038/s41378-023-00608-x
157. Shah SA, Oakes RS, Jewell CM. Advancing immunotherapy using biomaterials to control tissue, cellular, and molecular level immune signaling in skin. Adv Drug Deliv Rev. 2024;209:115315. doi:10.1016/j.addr.2024.115315
158. Wang T, Yin L, Ma Z, Zhang Y. Chlorogenic acid-loaded mesoporous silica nanoparticles modified with hexa-histidine peptides reduce skin allergies by capturing nickel. Molecules. 2022;27(4):1430. doi:10.3390/molecules27041430
159. Paris JL, Vallet-Regí M. Mesoporous silica nanoparticles for co-delivery of drugs and nucleic acids in oncology: a review. Pharmaceutics. 2020;12(6):526. doi:10.3390/pharmaceutics12060526
160. Musazzi UM, Franzè S, Condorelli F, Minghetti P, Caliceti P. Feeding next-generation nanomedicines to Europe: regulatory and quality challenges. Adv Healthc Mater. 2023;12(30):e2301956. doi:10.1002/adhm.202301956
161. Miao D, Song Y, De Munter S, et al. Photothermal nanofiber-mediated photoporation for gentle and efficient intracellular delivery of macromolecules. Nat Protoc. 2025;20(7):1810–1845. doi:10.1038/s41596-024-01115-7
162. Souto EB, Blanco-Llamero C, Krambeck K, et al. Regulatory insights into nanomedicine and gene vaccine innovation: safety assessment, challenges, and regulatory perspectives. Acta Biomater. 2024;180:1–17. doi:10.1016/j.actbio.2024.04.010
163. Wang YL, Lee YH, Chou CL, Chang YS, Liu WC, Chiu HW. Oxidative stress and potential effects of metal nanoparticles: a review of biocompatibility and toxicity concerns. Environ Pollut. 2024;346:123617. doi:10.1016/j.envpol.2024.123617
164. Lee Y, Jeong M, Park J, Jung H, Lee H. Immunogenicity of lipid nanoparticles and its impact on the efficacy of mRNA vaccines and therapeutics. Exp Mol Med. 2023;55(10):2085–2096. doi:10.1038/s12276-023-01086-x
165. Newton HS, Zhang J, Donohue D, et al. Multicolor flow cytometry-based immunophenotyping for preclinical characterization of nanotechnology-based formulations: an insight into structure activity relationship and nanoparticle biocompatibility profiles. Front Allergy. 2023;4:1126012. doi:10.3389/falgy.2023.1126012
166. Schmitt RR, Mahajan SD, Pliss A, Prasad PN. Small molecule based EGFR targeting of biodegradable nanoparticles containing temozolomide and Cy5 dye for greatly enhanced image-guided glioblastoma therapy. Nanomedicine. 2022;41:102513. doi:10.1016/j.nano.2021.102513
167. Gao X, Wang X, Li S, Saif Ur Rahman M, Xu S, Liu Y. Nanovaccines for advancing long-lasting immunity against infectious diseases. ACS Nano. 2023;17(24):24514–24538. doi:10.1021/acsnano.3c07741
168. Deng Y, Zheng H, Li B, et al. Nanomedicines targeting activated immune cells and effector cells for rheumatoid arthritis treatment. J Control Release. 2024;371:498–515. doi:10.1016/j.jconrel.2024.06.010
169. Deng J, Wang J, Shi J, et al. Tailoring the physicochemical properties of nanomaterials for immunomodulation. Adv Drug Deliv Rev. 2022;180:114039. doi:10.1016/j.addr.2021.114039
170. Rodríguez-Gómez FD, Monferrer D, Penon O, Rivera-Gil P. Regulatory pathways and guidelines for nanotechnology-enabled health products: a comparative review of EU and US frameworks. Front Med. 2025;12:1544393. doi:10.3389/fmed.2025.1544393
171. Kocatürk E, Muñoz M, Elieh-Ali-Komi D, et al. How infection and vaccination are linked to acute and chronic urticaria: a special focus on COVID-19. Viruses. 2023;15(7):1585. doi:10.3390/v15071585
172. Magen E, Yakov A, Green I, Israel A, Vinker S, Merzon E. Chronic spontaneous urticaria after BNT162b2 mRNA (Pfizer-BioNTech) vaccination against SARS-CoV-2. Allergy Asthma Proc. 2022;43(1):30–36. doi:10.2500/aap.2022.43.210111
173. Schwab J, Foglierini M, Pescosolido E, et al. Features of chronic urticaria after COVID-19 mRNA vaccine over time. Commun Med. 2024;4(1):254. doi:10.1038/s43856-024-00656-y
174. Dudukina E, Brâuner EV, Christiansen CB, et al. Associations between mRNA COVID-19 vaccination and urticaria: a nationwide registry-based cohort study in Denmark. Scand J Public Health. 2025;53(5):533–543. doi:10.1177/14034948251333901
175. Brás R, Esteves Caldeira L, Bernardino A, Costa C. Anti-TPO IgG/Total IgE ratio: biomarker for omalizumab response prediction in chronic spontaneous urticaria. Int Arch Allergy Immunol. 2023;184(9):866–869. doi:10.1159/000532021
176. Sella JA, Ferriani MPL, Melo JML, et al. Type I and type IIb autoimmune chronic spontaneous urticaria: using common clinical tools for endotyping patients with CSU. J Allergy Clin Immunol Glob. 2023;2(4):100159. doi:10.1016/j.jacig.2023.100159
177. Chu AWL, Oykhman P, Chu X, et al. Comparative efficacy and safety of biologics and systemic immunomodulatory treatments for chronic urticaria: systematic review and network meta-analysis. J Allergy Clin Immunol. 2025:S0091–6749(25)00646–3. doi:10.1016/j.jaci.2025.06.004
178. Patel D, Shelbaya A, Cheung R, Aggarwal J, Park SH, Coindreau J. Cost-effectiveness of early treatment with originator biologics or their biosimilars after methotrexate failure in patients with established rheumatoid arthritis. Adv Ther. 2019;36(8):2086–2095. doi:10.1007/s12325-019-00986-7
179. Zhao Z, Zheng Y, Song X, et al. Biological and target synthetic treatments for chronic spontaneous urticaria: a systematic review and network meta-analysis. Clin Transl Allergy. 2025;15(5):e70052. doi:10.1002/clt2.70052
180. Rosenblum D, Peer D. Omics-based nanomedicine: the future of personalized oncology. Cancer Lett. 2014;352(1):126–136. doi:10.1016/j.canlet.2013.07.029
181. Chou W, Canchola A, Zhang F, Lin Z. Machine Learning and Artificial Intelligence in Nanomedicine. Wiley Interdiscip Rev Nanomed Nanobiotechnol. 2025;17(4):e70027. doi:10.1002/wnan.70027
182. Bahrami Y, Bolideei M, Mohammadzadeh S, et al. Applications of artificial intelligence and nanotechnology in vaccine development. Int J Pharm. 2025;684:126096. doi:10.1016/j.ijpharm.2025.126096
183. Aljabali AAA, Obeid MA, Gammoh O, et al. Nanomaterial-driven precision immunomodulation: a new paradigm in therapeutic interventions. Cancers. 2024;16(11):2030. doi:10.3390/cancers16112030
184. Wu X, Wang J, Shi W, Ding Y, Li Y. High-throughput screening accelerated by machine learning for the morphology of silica nanoparticles with high cell permeability. Nanoscale. 2025;17(33):19353–19362. doi:10.1039/d5nr00587f
185. Li H, Nithin C, Kmiecik S, Huang SY. Computational methods for modeling protein-protein interactions in the AI era: current status and future directions. Drug Discov Today. 2025;30(6):104382. doi:10.1016/j.drudis.2025.104382
186. Chattopadhyay S, Goswami A, Sil M. Nanobiotechnology: traditional re-interpreting personalized medicine through targeted therapies and regenerative solutions. Naunyn Schmiedebergs Arch Pharmacol. 2025;398(8):10025–10048. doi:10.1007/s00210-025-04038-6
187. Tan IJ, Podwojniak A, Parikh A, Cohen BA. Precision dermatology: a review of molecular biomarkers and personalized therapies. Curr Issues Mol Biol. 2024;46(4):2975–2990. doi:10.3390/cimb46040186
188. Wu Y, Yang Y, Zhao W, et al. Novel iron oxide-cerium oxide core-shell nanoparticles as a potential theranostic material for ROS related inflammatory diseases. J Mater Chem B. 2018;6(30):4937–4951. doi:10.1039/c8tb00022k
189. Napp J, Markus MA, Heck JG, et al. Therapeutic fluorescent hybrid nanoparticles for traceable delivery of glucocorticoids to inflammatory sites. Theranostics. 2018;8(22):6367–6383. doi:10.7150/thno.28324
190. Mostovenko E, Young T, Muldoon PP, et al. Nanoparticle exposure driven circulating bioactive peptidome causes systemic inflammation and vascular dysfunction. Part Fibre Toxicol. 2019;16(1):20. doi:10.1186/s12989-019-0304-6
191. Wang X, Zhang J, Hu Y, et al. Multi-omics analysis reveals the unexpected immune regulatory effects of arsenene nanosheets in tumor microenvironment. ACS Appl Mater Interfaces. 2022;14(40):45137–45148. doi:10.1021/acsami.2c10743
192. Zhang T, Lei T, Yan R, et al. Systemic and single cell level responses to 1 nm size biomaterials demonstrate distinct biological effects revealed by multi-omics atlas. Bioact Mater. 2022;18:199–212. doi:10.1016/j.bioactmat.2022.03.026
193. Eslami M, Fadaee Dowlat B, Yaghmayee S, et al. Next-generation vaccine platforms: integrating synthetic biology, nanotechnology, and systems immunology for improved immunogenicity. Vaccines. 2025;13(6):588. doi:10.3390/vaccines13060588
194. Shaw JR, Caprio N, Truong N, et al. Inflammatory disease progression shapes nanoparticle biomolecular Corona-mediated immune activation profiles. Nat Commun. 2025;16(1):924. doi:10.1038/s41467-025-56210-4
195. Proetzel G, Wiles MV, Roopenian DC. Genetically engineered humanized mouse models for preclinical antibody studies. BioDrugs. 2014;28(2):171–180. doi:10.1007/s40259-013-0071-0
196. Othman HO, Issa KD, Ali DS, et al. Innovative nanomaterials in precision oncology: carbon quantum dots driving cancer immunotherapy. OMICS. 2025;29(7):293–308. doi:10.1089/omi.2025.0043
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
How Efficient are Alendronate-Nano/Biomaterial Combinations for Anti-Osteoporosis Therapy? An Evidence-Based Review of the Literature
Klara J, Lewandowska-Łańcucka J
International Journal of Nanomedicine 2022, 17:6065-6094
Published Date: 6 December 2022
Nanomaterial-Enhanced Immunotherapy: Advancing T-Cell-Based Treatments for Bladder Cancer
Chen J, Fu Y, Zhang Z, Zhao J, Zuo J, Ye X, Xiong Q, Nie Z, Dong H, Shi H, Tan Z, Wang C, Chen B, Wang Z, Li X, Chen P, Wang H, Fu S
International Journal of Nanomedicine 2025, 20:15235-15275
Published Date: 18 December 2025
Nanomaterial-Based Precision Drug Delivery for Advanced Nephrology Therapy: A Systematic Review
Feng J, Zhang R, Chen X, Jia M, Li L, Zhang L, Fu Y
International Journal of Nanomedicine 2026, 21:583744
Published Date: 25 March 2026