")
Back to Journals » Infection and Drug Resistance » Volume 13
Molecular Epidemiology of Myroides odoratimimus in Nosocomial Catheter-Related Infection at a General Hospital in China
Authors Yang S, Liu Q, Shen Z, Wang H, He L
Received 27 February 2020
Accepted for publication 14 June 2020
Published 25 June 2020 Volume 2020:13 Pages 1981—1993
DOI https://doi.org/10.2147/IDR.S251626
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Eric Nulens
Shuang Yang, Qian Liu, Zhen Shen, Hua Wang, Lei He
Department of Laboratory Medicine, Renji Hospital, School of Medicine, Shanghai Jiao Tong University, Shanghai, People’s Republic of China
Correspondence: Lei He; Hua Wang
Department of Laboratory Medicine, Renji Hospital, School of Medicine, Shanghai Jiao Tong University, No. 160 Pujian Road, Pudong New District, Shanghai, People’s Republic of China
Email [email protected]; [email protected]
Purpose: Catheter-related infection (CRI) is one of the most frequent causes of hospitalizations for immunocompromised patients. A major challenge is the increased prevalence of Myroides odoratimimus. The purpose of the present study was to evaluate the clinical features and molecular characteristics of M. odoratimimus collected from a general hospital in Shanghai, China.
Patients and Methods: From July 2015 to August 2016, a total of 22 isolates of M. odoratimimus were collected from inpatients respectively from the biliary and pancreatic surgery (6/22) and the urology department (16/22). Clonal relatedness among the isolates was assessed using pulsed-field gel electrophoresis (PFGE) and matrix-assisted laser desorption/ionization time-of-flight mass spectrometry (MALDI-TOF MS). Moreover, the antimicrobial susceptibility tests were carried out using the Clinical and Laboratory Standards Institute (CLSI) broth microdilution method. The presence of antibiotic resistance genes was screened using the polymerase chain reaction (PCR) assay. Additionally, protein structure prediction was analyzed using PSIPRED and RaptorX.
Results: PFGE differentiated these isolates into six possibly related clones from two different departments obtained during a distinct period, indicating clonal dissemination in the two departments. We compared the dendrograms of M. odoratimimus isolates obtained by MALDI-TOF MS with those obtained by PFGE and found that the coincidence rate between them was only 68.2%. All the M. odoratimimus isolates were highly resistant to most available antibiotics, including carbapenems. Furthermore, chromosome-encoded β-lactamases MUS-1 was confirmed by PCR in 6 of 22 Myroides odoratimimus isolates. Herein, we also reported a novel variant of blaMUS- 1 in the remaining 16 isolates, which encodes MUS-3 protein at position 60 (Valine to Alanine), differing from the structure of MUS-1.
Conclusion: The opportunistic and extensively antibiotic-resistant Myroides odoratimimus has a small range of epidemics in these two different departments. Clinicians should be aware that M. odoratimimus may induce a severe nosocomial outbreak of catheter-related infections, particularly in immunocompromised patients.
Keywords: Flavobacterium, Myroides odoratimimus, MUS-1, nosocomial, outbreak
Introduction
The Myroides genus was first isolated in 1923, originally classified as part of the genus Flavobacterium.1 In 1996, the new genus Myroides was established, in which two species named Myroides odoratus and Myroides odoratimimus were included.2 In recent years, three new species (pelagicus, profundi, and marinus) have been isolated from sea water.3–5 Myroides species are strictly aerobic and nonmotile, yellow-pigmented, oxidase-positive gram-negative rods with a characteristic fruity odor. They are ubiquitous in the environment and often found in soil and water.6 In the last two decades, Myroides species mostly behave as rare opportunistic pathogens for humans, which may cause an increasing number of infections such as soft tissue infection,7,8 necrotizing fasciitis,9 cellulitis,10–12 ventriculitis,13 and urinary tract infections (UTIs)6,14 in immunocompromised hosts. However, few cases have been recorded in immunocompetent individuals.7,8 The prevalence of Myroides odoratimimus is increasing. This may be due to inappropriate use of antibiotics15 or improved detection by reliable and practicable technologies, such as matrix-assisted laser desorption/ionization time-of-flight mass spectrometry (MALDI-TOF MS).16 Myroides odoratimimus has been gradually considered as an important nosocomial pathogen, which poses a serious health threat to immunocompromised patients and has the potential to cause a pandemic.17 In the past few decades, more and more catheter-related infection (CRI) cases caused by Myroides odoratimimus have been reported, including three main nosocomial outbreaks of UTI caused by M. odoratimimus. The first main outbreak occurred in the urology ward of a Turkish hospital, which lasted for three years, from 1994 to 1997.18 The second outbreak was in a Tunisian hospital from May to November 2010, also in the urology department.15 The third occurred in western Romania from June to August 2017.19 Here, we report on a nosocomial prevalence caused by M. odoratimimus in the Renji Hospital at the Shanghai Jiaotong University School of Medicine in China.
Patients and Methods
Bacterial Isolates and Data Collection
A total of 22 non-repetitive clinical isolates of Myroides odoratimimus were routinely obtained in clinical microbiology laboratory, Renji Hospital at the Shanghai Jiaotong University School of Medicine, China from July 2015 to August 2016. Of 22 isolates, 91.0% (20/22) were isolated from celiac drainage fluid, followed by urine (4.5%, 1/22) and bile (4.5%, 1/22), of inpatients undergoing aseptic abdominal surgery, admitted to two wards: the biliary and pancreatic surgery ward and the urology ward. According to the medical records, all subjects whose body fluid cultures were positive for Myroides odoratimimus were retrospectively reviewed based on the electronic patient hospital database and clinical microbiology laboratory databases. The patients without obvious infection symptoms were considered as M. odoratimimus colonization. Data, including demographic characteristics, underlying disease, diagnosis, type of surgery, ward floor, antimicrobial use, and outcome were collected.
Environmental Surveillance
During the same period, environmental surveillance samples were collected from various locations in the wards using a sterile swab (Copan, Italy), including the surface of the drainage tube, inner surfaces of sinks, commodes, bed rails, bedside tables, faucets, door handles, tap water, and patient’s or medical staff’s hands. Air sampling was performed with nutrient agar medium (Oxoid, UK) to determine the presence of Myroides spp.
Identification and Antimicrobial Susceptibility
All the 22 isolates were analyzed using the VITEK2 compact (BioMérieux, France) and MALDI-TOF MS (Bruker Daltonics, Bremen, Germany). Antimicrobial resistance profiles, including piperacillin–tazobactam (TZP), ceftazidime (CAZ), ceftriaxone (CRO), cefepime (FEP), aztreonam (ATM), ciprofloxacin (CIP), gentamicin (CN), and amikacin (AK) were determined by VITEK2 using gram-negative susceptibility cards (VITEK2 Compact AST GN13) according to Clinical and Laboratory Standards Institute (CLSI) guidelines as previously described.20 Then the minimum inhibitory concentrations (MICs) of imipenem (IPM), meropenem (MEM), trimethoprim–sulfamethoxazole (SXT), and levofloxacin (LEV) were determined using the commercial broth microdilution MIC method. The MIC results were interpreted following the Clinical and Laboratory Standards Institute standards (CLSI, 2018), according to the breakpoint for non-Enterobacteriaceae. Escherichia coli ATCC 25922 and Pseudomonas aeruginosa 27853 were used as quality control strains for the antibiotic susceptibility tests. The commercial broth microdilution MIC method is generally used to measure (semi-quantitatively) the in vitro activity of an antimicrobial agent against a bacterial isolate.21 This assay was repeated three times, and the results were reproducible.
Pulsed-Field Gel Electrophoresis (PFGE)
We performed molecular typing to characterize the clonal relatedness of the strains by using PFGE as previously described.22 Genomic DNA was extracted from bacterial cultures using EZ-10 spin column DNA isolation kits (Sangon, Shanghai, China) according to the manufacturer’s instructions. Agarose plugs were incubated with BssHII (New England Biolabs, UK). The DNA fragments were separated by electrophoresis on 1% SeaKem Gold agarose (Lonza, Rockland, ME, United States) in 0.5 × TBE buffer (45 mM Tris, 45 mM boric acid, 1.0 mM EDTA; pH 8.0), using the CHEF Mapper XA PFGE system (Bio-Rad, United States) at 6 V/cm and 14 °C, with alternating pulses at a 120 angle in a 5–40 s pulse time gradient for 19 h. We analyzed the generated DNA fingerprints as recommended by Tenover et al.22 The results of the PFGE homology analysis were compared with clustering analyses based on a specific mass peak using principal component analysis (PCA). Analysis was performed using the MALDI-Biotype 2.0 software.23,24
Detection of Antibiotic Resistance Genes
We used standard PCR conditions and appropriate primers, as listed in Table 3, to detect the presence of carbapenemase-encoding genes, including blaKPC, blaOXA, blaVIM, blaIMP, blaMUS, blaTUS-1, blaNDM, and blaB.25–27 The resulting DNA sequences and their corresponding amino acid sequences were analyzed using the DNAMAN 6.0 program and blasted with genetic databases available on the website (http://www.ncbi.nlm.nih.gov/BLAST/).
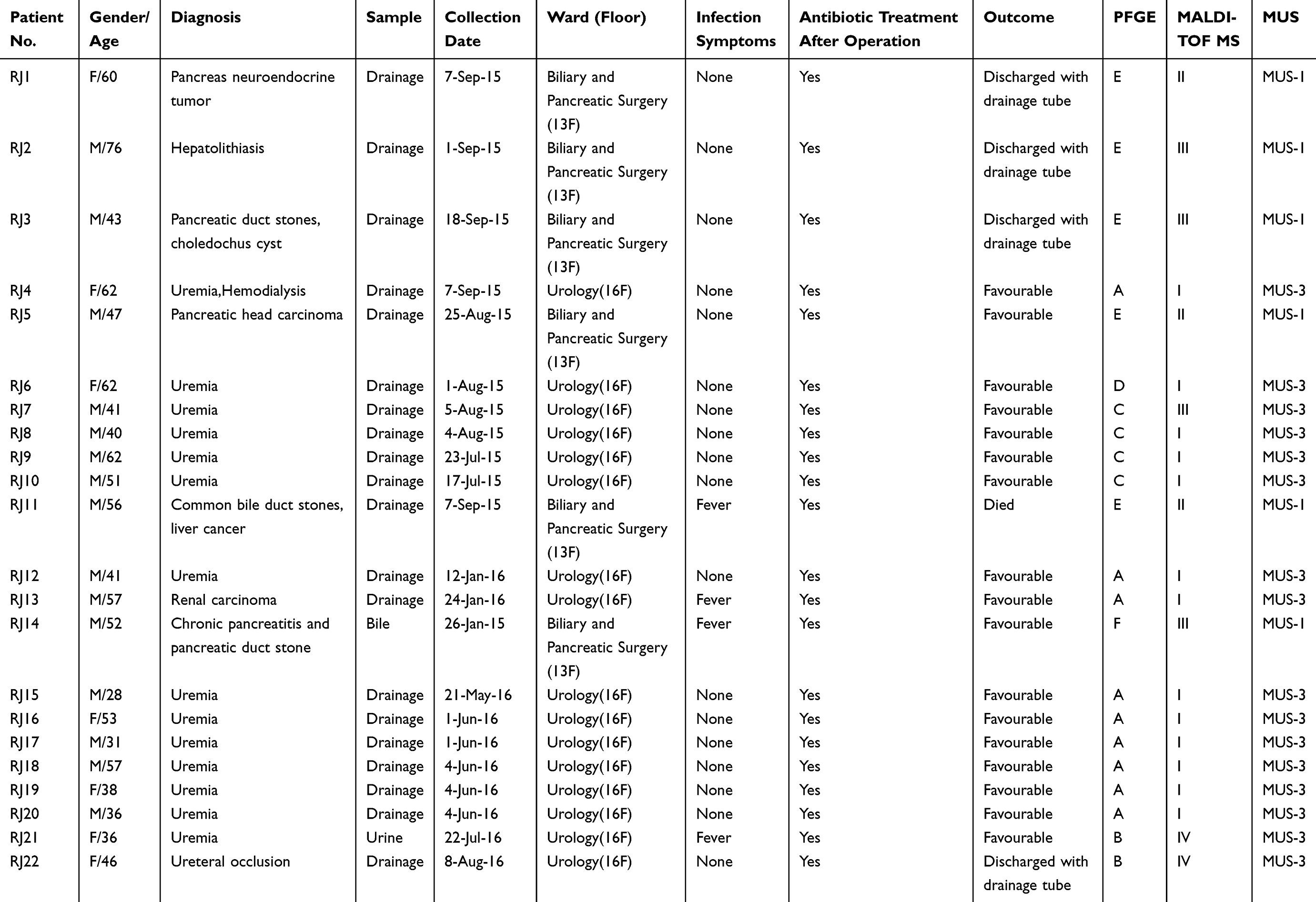 |
Table 1 Demographic Data of Patients Included in This Study |
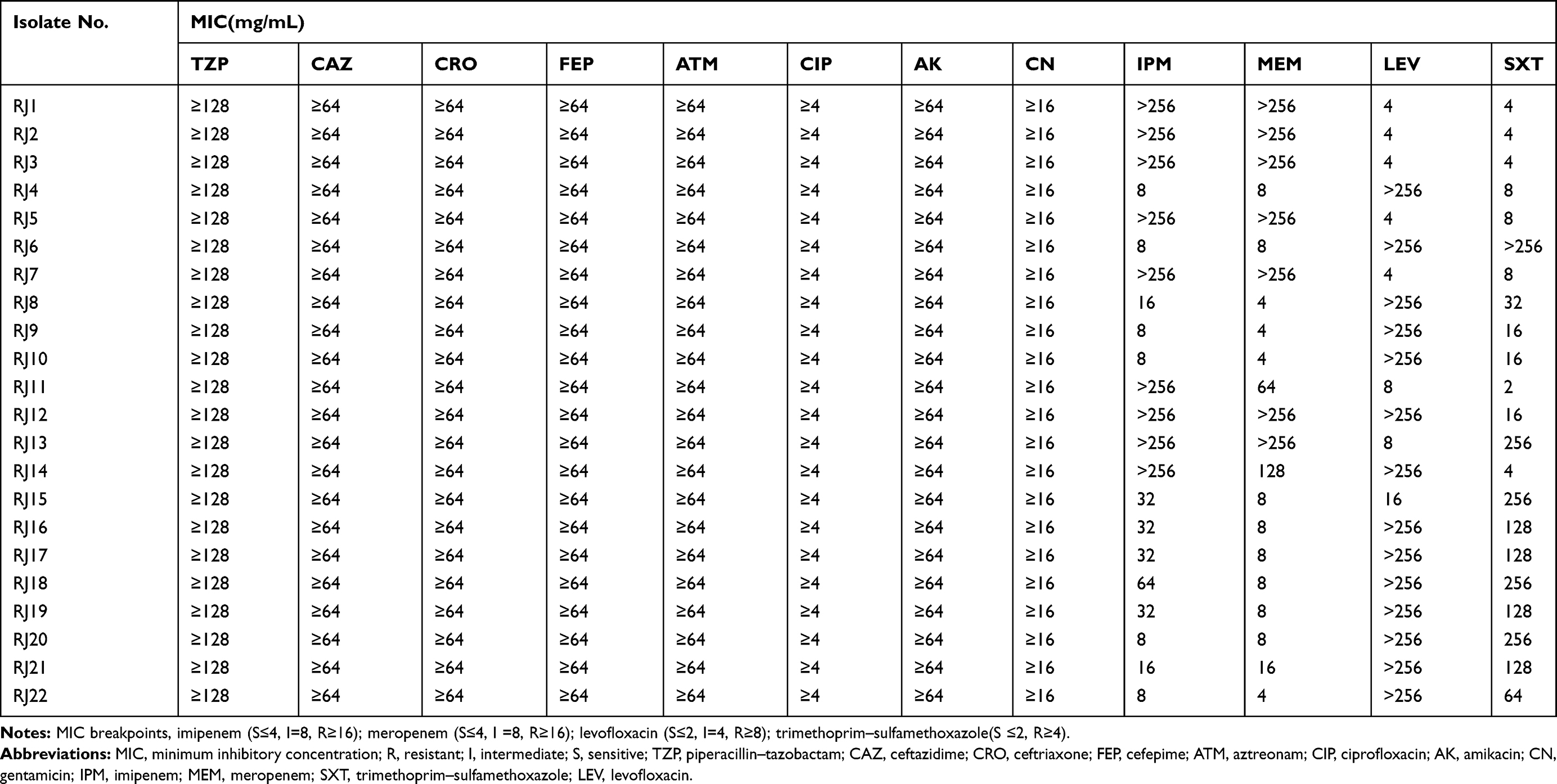 |
Table 2 Summary of MIC Results Among the 22 M. odoratimimus Isolates |
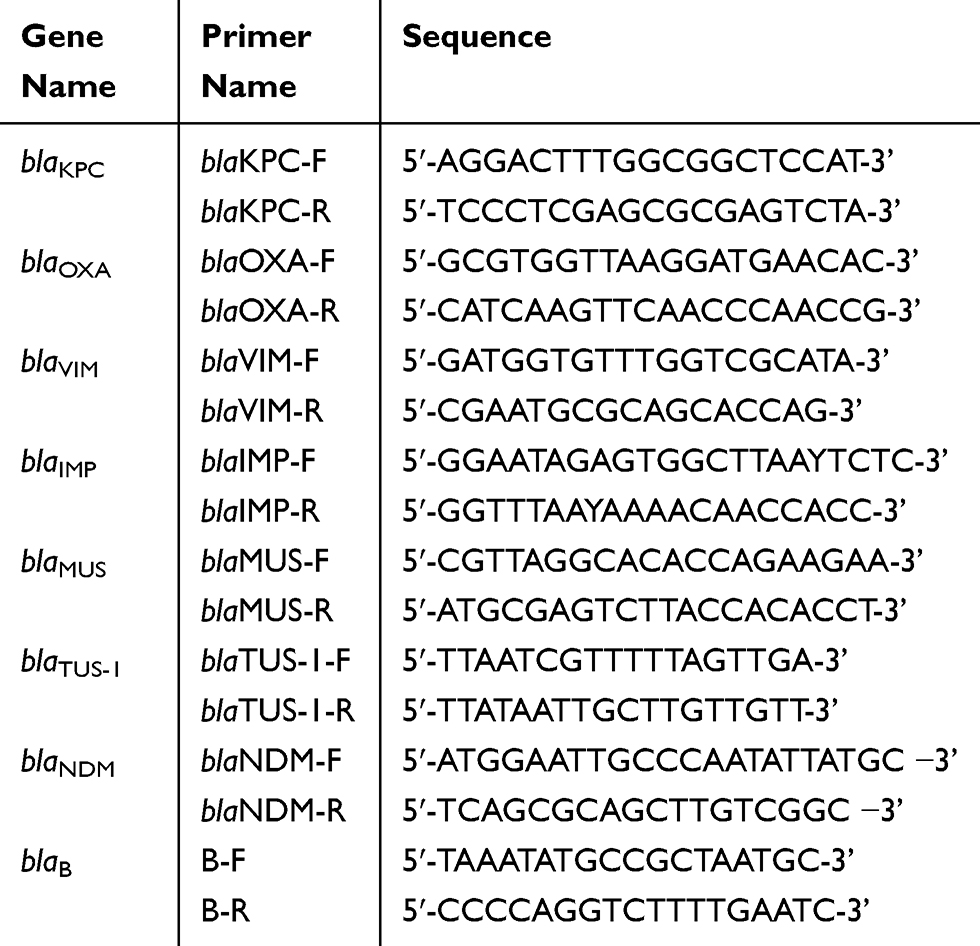 |
Table 3 Primers for the Carbapenemase Gene Sequencing Analysis |
Protein Secondary Structure Prediction by PSIPRED and Tertiary Structure Modeling Analysis by RaptorX
To predict whether a point mutation could have an effect at the protein level, the Mus amino acid substitution was identified with PSIPRED (http://bioinf.cs.ucl.ac.uk/psipred/).28
The Mus protein modeling was analyzed with RaptorX (http://raptorx.uchicago.edu/) to position the mutation on the tertiary structure.29
Statistical Analysis
Data were analyzed using SPSS TM software (IBM corp., USA) version 23.0. Chi-squared tests were used to estimate differences between groups. Statistical significance was regarded as P values < 0.05.
Results
Bacterial Isolates
We isolated 22 consecutive non-repetitive Myroides odoratimimus strains from immunocompromised patients with severe underlying diseases who had undergone abdominal surgery, such as a pancreatectomy or kidney transplantation (Table 1). As preventive intraperitoneal drainage was commonly performed in these patients, 20 out of the 22 strains were isolated from abdominal drainage fluid, and only two strains were isolated from bile and urine. Of the 22 patients, 16 (72.7%) were treated at the urology department on the 16th floor, and the remaining 6 (27.3%) were treated at the biliary and pancreatic surgery department on the 13th floor. The majority of patients (68.2%) were male, and the mean age of the included patients was 48.9 years, of which 17 (77.3%) were 28–60 years old. No Myroides odoratimimus strains were isolated from any of the environmental surveillance samples, indicating that the above clinical isolates were likely not derived from the environment.
The bacterium was isolated from Columbia blood agar after 16–18h of incubation in aerobic conditions. The colonies appeared round and yellow-pigmented and had a fruity odor. The organism was initially identified using conventional biochemical reactions by the VITEK2 AES as Myroides isolates, but this analysis could not accurately distinguish the bacteria at the species level. We used MALDI-TOF MS to further confirm the bacterial identification as Myroides odoratimimus (matching score > 2.2, sequence homology > 99%) using the Microflex LT System and analyzing data on the Biotyper 2.0 software (Bruker Daltonics, Germany) for all the 22 isolates.16
Dendrogram Analysis by PFGE and MALDI-TOF MS
According to the PFGE results, the 22 isolates belonged to six distant pulsotypes (types A-F) with a cut-off of 80% similarity (Figure 1). The dominant cluster was cluster A (40.9%, n = 9/22), followed by cluster B (9.1%, n = 2/22), which were all obtained from the urology ward during the period from September 2015 to August 2016. Cluster C (18.2%, n = 4/22) and cluster D (4.5%, n = 1/22) also came from the urology ward but were obtained between July and August 2015. Cluster E (22.7%, n = 5/22) was considered to be highly clonally-related to cluster F (4.5%, n = 1/22), both of which were isolated from the biliary and pancreatic surgery ward. MALDI-TOF MS, on the other hand, generated a dendrogram with four clusters with a cut-off value of 3 using PCA cluster analysis (Figure 2). The strains with similar PFGE patterns were divided into different branches by the MALDI-TOF MS dendrogram. Comparing the MALDI-TOF MS data with PFGE as the gold standard, 68.2% (15/22) of strains were classified with high homology into the same cluster.
 |
Figure 1 PFGE image of the M. odoratimimus strains. The 22 isolates belonged to six distant PFGE groups (types A–F) by using a cut-off of 80% similarity (Δurology department, □biliary and pancreatic surgery department). |
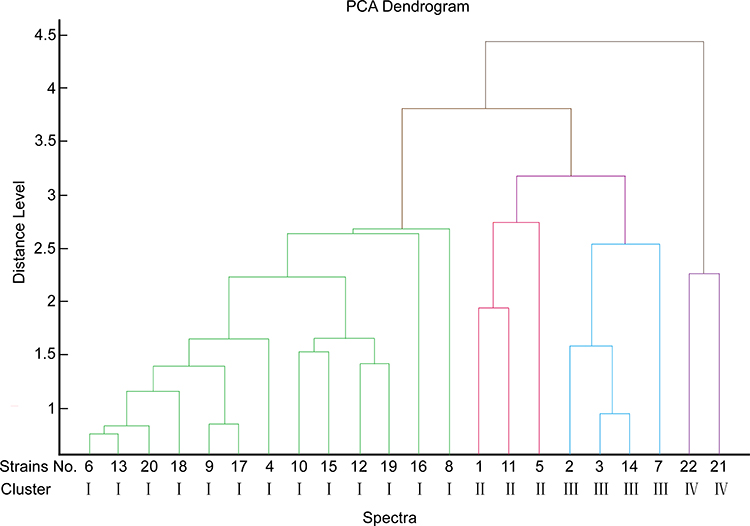 |
Figure 2 MALDI-TOF MS dendrograms for 22 isolates. Using PCA cluster analysis to construct a dendrogram, MALDI-TOF MS generated four clusters (types I, II, III, and IV) with a cut-off value of 3. |
Antibiotic Susceptibility Test
The results of antibiotic susceptibility testing revealed that all the Myroides odoratimimus isolates were resistant to piperacillin–tazobactam (MIC ≥ 128 µg/mL), ceftazidime (MIC ≥ 64 µg/mL), ceftriaxone (MIC ≥ 64 µg/mL), cefepime (MIC ≥ 64 µg/mL), aztreonam (MIC ≥ 64 µg/mL), amikacin (MIC ≥ 64 µg/mL), gentamicin (MIC ≥ 16 µg/mL), and ciprofloxacin (MIC ≥ 4 µg/mL). Those isolates also showed high rates of resistance to imipenem (72.7%), meropenem (54.5%), levofloxacin (77.3%), and trimethoprim/sulfamethoxazole (95.5%). The MIC results among M. odoratimimus isolates are shown in Table 2. Notably, the six clinical isolates from the biliary and pancreatic surgery ward possessed a significantly higher level of resistance to meropenem compared with the 16 isolates obtained at the urology ward (P<0.05) (Figure 3).
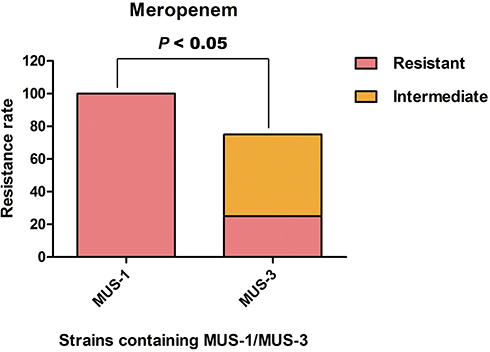 |
Figure 3 The level of resistance to meropenem between the clinical isolates of MUS-1 from the biliary and pancreatic surgery ward and MUS-3 from the urology ward (P < 0.05). |
Identification of Antibiotic Resistance Genes
Carbapenemase-encoding genes were further screened by PCR using carbapenemase gene primers. The nucleotide sequence analysis was negative for blaKPC, blaVIM, blaIMP, blaTUS-1, blaNDM, blaOXA, and blaB variants. However, the chromosomal blaMUS gene was amplified in all the 22 isolates. The six isolates from the biliary and pancreatic surgery ward carried the blaMUS-1 gene, while a novel missense single mutation from the 178th base transition (T178C) in blaMUS was identified in the 16 isolates from the urology ward. The above gene mutation produces a new MUS-1 enzyme with an amino acid mutation at the 60th position (V60A), differing from the original MUS-1 enzyme. Based on the definition used to name the novel variant enzyme MUS-2,27 our new enzyme variant was designated MUS-3 after the Blast Search of the Lahey database of β-lactamases (http://www.laced.uni-stuttgart.de/), and deposited in GenBank (https://www.ncbi.nlm.nih.gov/pathogens/submit-beta-lactamase) under the accession number MT364441. The new blaMUS gene was also named as blaMUS-3.
In silico Structure Prediction of Point Mutation Effect
To visualize the potential effect of the V60A mutation in the MUS-3 enzyme on the MUS structure without crystallographic data, we analyzed wild-type and mutant V60A alleles with the PSIPRED secondary structure prediction tool and the RaptorX tertiary structure modeling tool for in silico structure prediction, as previously performed.30,31 A short β-strand was predicted to form at the mutation position in the center of the amino acid chain (Figure 4). This modification is prone to result in differential enzyme activity in the MUS V60A mutant protein. Moreover, since the observed valine-to-alanine substitution is linked to the secondary and tertiary structure of the protein, the active site of the MUS enzyme may be located in the proximity of this mutation.
 |
Figure 4 The secondary and tertiary structure prediction of MUS-1 wild-type and V60A-mutant proteins using bioinformatics tools. (A) Secondary structure prediction of the MUS wild-type protein by PSIPRED. (B) Secondary structure prediction of the MUS-3 (V60A) mutant protein by PSIPRED. A beta-strand is predicted to form at the mutation position (red triangle). (C) Tertiary structure prediction of the MUS protein by RaptorX, with the mutation marked inside (black oval). |
Discussions
In our present work, we analyzed sampled from 22 patients with Myroides spp. infection/colonization who had been hospitalized long-term due to several underlying diseases and complicated abdominal surgery. After the operation, drainage tubes were routinely used to prevent ascites formation and to reduce the incidence of abdominal infection. Multiple antibiotics had also been used to prevent opportunistic infection. Therefore, the patients were at risk of acquiring multidrug-resistant infections.
The 22 non-repetitive Myroides odoratimimus strains were obtained from patients in two surgery departments between July 2015 and August 2016. The majority was obtained from the urology department (72.6%), while 27.3% of strains were obtained from the biliary and pancreatic surgery department. From the electronic medical records and communications with physicians, we deducted that four patients were diagnosed with M. odoratimimus infections with symptoms such as fever, local swelling, and pain. These patients were treated with imipenem and levofloxacin, but one of them had a poor prognosis. The remaining patients who were identified as M. odoratimimus colonization did not receive any drug treatment. After extubating, patients either left or stayed in outpatient service. However, weakened hosts have an increased likelihood of developing an infection following colonization. Spanik et al reported four cases of catheter-related bacteremia caused by Myroides for whom the infection symptoms disappeared after removal of the catheter.32 These findings are very similar to our case, and we speculate its virulence seems to be relatively low. Susceptibility data revealed M. odoratimimus was highly resistant to almost all antibiotics, including aminoglycosides and cephalosporins, and showed decreased susceptibility to carbapenems, sulfamethoxazole–trimethoprim, and fluoroquinolones. In our research, we found that 72.7% of M. odoratimimus isolates were resistant to imipenem and 54.5% to meropenem. This suggests that meropenem may be more effective than imipenem for the treatment of M. odoratimimus infections, which is consistent with previous research.9,33
Starting from these preliminary clinical and experimental findings, in the second phase of this study, we molecularly typed all the 22 strains by PFGE and MALDI-TOF MS. We identified a total of 6 PFGE types in the 22 Myroides odoratimimus isolates using a cut-off of 80% similarity.
This analysis revealed that four related clusters A, B, C, and D were isolated from the urology ward on the 16th floor, while clusters E and F were obtained from the biliary and pancreatic surgery ward on the 13th floor. This suggests the potential intrahospital clonal transmission within these two departments. We are considering other strategies to prove this assumption further in future work. PFGE is the current “gold standard” fingerprinting method used to identify epidemic strains in clinical microbiological laboratories.34,35 Nevertheless, there are some limitations of PFGE, for instance, it is time-consuming, and DNA restriction patterns can vary slightly between technicians or between time and place.22,36 Thus, this method is suitable for regional surveillance in which the isolates have to be compared in one laboratory. MLST is emerging as an alternative DNA fingerprinting technique, particularly when information regarding evolutionary history is required. MLST is not as discerning as PFGE. The sequence types (ST) need to be defined and can be exchanged between laboratories worldwide.37 However, MLST is the standard method for epidemiological investigation with large-scale international comparisons.38 Whole genome sequencing (WGS) could be an alternative for molecular epidemiological research of bacteria.39 WGS could obtain the entire sequence of a bacterial genome in a short turnaround time at an affordable cost. This greatly increases the amount of information that can be used to compare bacterial strains and, therefore, improves the discrimination of bacterial typing. The most frequently used WGS-based bacterial typing methods are based on single-nucleotide polymorphism (SNP) detection and core genome MLST (cgMLST).40 However, here, both MLST and cgMLST are not applicable for genotyping M. odoratimimus since this species is a rare opportunistic pathogen in humans, and there is minimal information of ST types for this species. SNP analysis, which could exploit the whole genome positions, has the advantage that it discriminates closely-related isolates and, consequently, can perform detailed epidemiological research into clonal transmission in M. odoratimimus.41,42 At present, the most important limitation for applying WGS-based methods in hospital epidemiological monitoring is the lack of established guidelines to determine bacterial correlation, similar to the guidelines for PFGE.43 On the other hand, MALDI-TOF MS provides PCA-based dendrograms, which were evaluated in a few bacterial species.23,44 In the present investigation, we also used MALDI-TOF MS for the fingerprinting of 22 M. odoratimimus strains and evaluated the results from the two used methods. The result of the homology analysis by MALDI-TOF MS was consistent with that of PFGE by 68.2% (15/22). This shows that the PCA in MALDI-TOF MS has insufficient ability to analyze the clonality of M. odoratimimus. Therefore, even though MALDI-TOF MS typing is simple, fast, and cost-effective,23 PFGE is more appropriate to define clonal relationships. PFGE uses genetic characteristics for strain classification, while MALDI-TOF MS adopts proteomics for phenotype analysis. On top of that, the operating procedures of MALDI-TOF MS have not yet been standardized, and even culture medium types could result in low reproducibility for identification scores. Given the above issues that could influence MALDI-TOF MS dendrograms, we argue that, at present, MALDI-TOF MS should not be substituted for PFGE to define clonal relationships.
Myroides spp. are multidrug-resistant microorganisms, which have been reported to have a decreased susceptibility to β-lactams, including carbapenems, in many previous studies. However, the molecular mechanism behind this carbapenem resistance is not clear.33,45 Mammeri et al first claimed that the resistance to β-lactams in M. odoratus and M. odoratimimus was partly due to the production of the chromosome-encoded β-lactamases TUS-1 and MUS-1, respectively. These are members of the B1 subclass family, according to the classification by Ambler.46,47 MUS-2 is a novel variant of MUS-1, reported in M. odoratimimus isolated from livestock animals in 2015, which has 98.78% amino acid sequence homology with MUS-1 and harbors three substitutions of amino acids, including V60A, H133 T, and T224I compared with MUS-1.27 In our study, blaMUS-1 was detected from the clinical strains isolated from the biliary and pancreatic surgery department. We confirmed that decreased susceptibility to carbapenem resulted from the production of a chromosome-encoded metallo-β-lactamase. It is noteworthy that we detected another novel variant of blaMUS-1 in the 16 strains isolated from our urology department, leading to a single amino acid substitution (V60A), which is identical to one of the substitutions from blaMUS-2. Thus, we designated MUS-3 after the Blast Search of the Lahey database of β-lactamases and deposited it in GenBank. The new variant was named blaMUS-3. The amino acid sequence homology between MUS-3 and MUS-1 is 99.59%. On the other hand, the six clinical isolates with MUS-1, obtained from the biliary and pancreatic surgery ward, possessed a significantly higher level of resistance to meropenem than the isolates with MUS-3 from the urology ward. Furthermore, based on the protein secondary structure prediction by PSIPRED and tertiary structure modeling analysis by RaptorX, we found that the single amino acid mutation (V60A) could affect the active center of the MUS protein. Given the individual differences in drug resistance phenotype and genotype, we deduct that there was clonal spread of M. odoratimimus within the two surgical departments in our hospital. Some research has confirmed that the β-lactamase gene, especially chromosome-encoded MUS, was responsible for the decreased susceptibility to carbapenems.46,48 However, Mammeri et al cloned the blaMUS-2 in E. coli and found that it displays much lower levels of resistance to β-lactamase.46 Thus, it is apparent that MUS could only partly explain the intrinsic resistance of M. odoratimimus to carbapenem and this resistance mechanism should be investigated further.
In addition, it is noteworthy that there were two peak periods of M. odoratimimus detection. Eleven strains and eight strains were isolated from July to September 2015 and from May to August in 2016, respectively. We speculate that the current epidemic was due to contamination of the water system in the hospital in summer, and spread through poor hand hygiene, although environmental screens failed to reveal the source.49,50 As Myroides spp. is often isolated from sewage, we consider that the surface of the drainage tubes, inner surfaces of sinks, commodes, bed rails, bedside tables, faucets, door handles, and tap water are the major locations for environmental sampling. In the current study, thoroughly environmental surveillance was carried out in the biliary and pancreatic and urology ward, including air sampling, object surface, patient’s or medical staff’s hands, since Myroides odoratimimus clone transmission only appeared in these two wards. However, expanding environmental monitoring to the operating room in future studies might be useful since it is a potential high-risk place for being exposed to bacteria.
One of the limitations of this study is the small sample size, Myroides odoratimimus is rarely isolated in patients, making studies with larger sample sizes difficult to perform. We did obtain reliable statistical information of PFGE and MALDI-TOF MS based on the 22 clinical samples in this study. However, given the small sample size, it is difficult to form any definitive conclusions at present. Further studies with larger sample sizes are imperative to confirm our data.
Conclusion
In summary, over the past decade, an increasing number of infections caused by M. odoratimimus has been recognized. M. odoratimimus are environmental organisms and implicated as pathogens of nosocomial infection. These bacteria cause opportunistic infections in immunocompromised patients, especially in catheter-related individuals. Physicians should recognize that bacteria can spread in a hospital and even cause nosocomial outbreaks. Due to the organism’s multiple drug resistance, appropriate treatment options are limited. To find a more effective antibiotic treatment, extended drug sensitivity tests should be carried out. The mechanisms of drug resistance in M. odoratimimus are complex and have not been clarified. More research is needed to confirm the mechanisms behind drug resistance.
Abbreviations
CRI, catheter-related infection; PFGE, pulsed-field gel electrophoresis; MALDI-TOF MS, matrix-assisted laser desorption/ionization time-of-flight mass spectrometry; CLSI, Clinical and Laboratory Standards Institute; PCR, polymerase chain reaction; UTIs, urinary tract infections; TZP, piperacillin–tazobactam; CAZ, ceftazidime; CRO, ceftriaxone; FEP, cefepime; ATM, aztreonam; CIP, ciprofloxacin; CN, gentamicin; AK, amikacin; MIC, minimum inhibitory concentrations; IPM, imipenem; MEM, meropenem; SXT, trimethoprim–sulfamethoxazole; LEV, levofloxacin; PCA, principal component analysis; T, Thymine; C, Cytosine; V, Valine; A, Alanine; MLST, multilocus sequence typing; ST, sequence types; WGS, whole genome sequencing; SNP, single-nucleotide polymorphism; cgMLST, core genome MLST.
Compliance with Ethical Standards
This study was approved by the Ethics Committee of Renji Hospital in accordance with the 1964 Helsinki Declaration and its amendments or comparable ethical standards. The number of the ethics approval is RA-2019-198. The written informed consent was waived due to the noninterventional and retrospective nature of the study, that the collection of M. odoratimimus isolates was a part of the routine hospital laboratory procedures, all patient data were analyzed in anonymity, this retrospective study did not directly interfere with any patient and there was no adverse effect on the rights of patients.
Disclosure
The authors declare no conflict of interest.
References
1. Hugo CJ, Bruun B, Jooste PJ. The genera empedobacter and myroides. In: The Prokaryotes. Vol. 7.
2. VANCANNEYT M, SEGERS P, TORCK U, et al. Reclassification of Flavobacterium odoratum (Stutzer 1929) Strains to a New Genus, Myroides, as Myroides odoratus comb. nov. and Myroides odoratimimus sp. nov. Int J Syst Evol Microbiol. 1996;46(4):926–932.
3. Yoon J, Maneerat S, Kawai F, Yokota A. Myroides pelagicus sp. nov., isolated from seawater in Thailand. Int J Syst Evol Microbiol. 2006;56(Pt 8):1917–1920. doi:10.1099/ijs.0.64336-0
4. Zhang XY, Zhang YJ, Chen XL, et al. Myroides profundi sp. nov., isolated from deep-sea sediment of the southern Okinawa Trough. FEMS Microbiol Lett. 2008;287(1):108–112. doi:10.1111/j.1574-6968.2008.01299.x
5. Cho SH, Chae SH, Im WT, Kim SB. Myroides marinus sp. nov., a member of the family Flavobacteriaceae, isolated from seawater. Int J Syst Evol Microbiol. 2011;61(Pt 4):938–941. doi:10.1099/ijs.0.024067-0
6. Holmes B, Snell JJ, Lapage SP. Flavobacterium odoratum: a species resistant to a wide range of antimicrobial agents. J Clin Pathol. 1979;32(1):73–77. doi:10.1136/jcp.32.1.73
7. Benedetti P, Rassu M, Pavan G, Sefton A, Pellizzer G. Septic shock, pneumonia, and soft tissue infection due to Myroides odoratimimus: report of a case and review of Myroides infections. Infection. 2011;39(2):161–165. doi:10.1007/s15010-010-0077-1
8. Maraki S, Sarchianaki E, Barbagadakis S. Myroides odoratimimus soft tissue infection in an immunocompetent child following a pig bite: case report and literature review. Braz J Infect Dis. 2012;16(4):390–392. doi:10.1016/j.bjid.2012.06.004
9. Crum-Cianflone NF, Matson RW, Ballon-Landa G. Fatal case of necrotizing fasciitis due to Myroides odoratus. Infection. 2014;42(5):931–935. doi:10.1007/s15010-014-0626-0
10. Bachmeyer C, Entressengle H, Khosrotehrani K, et al. Cellulitis due to Myroides odoratimimus in a patient with alcoholic cirrhosis. Clin Exp Dermatol. 2008;33(1):97–98. doi:10.1111/j.1365-2230.2007.02590.x
11. Bachman KH, Sewell DL, Strausbaugh LJ. Recurrent cellulitis and bacteremia caused by Flavobacterium odoratum. Clin Infect Dis. 1996;22(6):1112–1113. doi:10.1093/clinids/22.6.1112
12. Meyer A, Dang H, Roland W. Myroides spp. cellulitis and bacteremia: a case report. IDCases. 2019;18:e00638. doi:10.1016/j.idcr.2019.e00638
13. Macfarlane DE, Baum-Thureen P, Crandon I. Flavobacterium odoratum ventriculitis treated with intraventricular cefotaxime. J Infect. 1985;11(3):233–238. doi:10.1016/S0163-4453(85)93228-1
14. Paek J, Shin JH, Shin Y, et al. Myroides injenensis sp. nov., a new member isolated from human urine. Antonie Van Leeuwenhoek. 2015;107(1):201–207. doi:10.1007/s10482-014-0317-y
15. Ktari S, Mnif B, Koubaa M, et al. Nosocomial outbreak of Myroides odoratimimus urinary tract infection in a Tunisian hospital. J Hosp Infect. 2012;80(1):77–81. doi:10.1016/j.jhin.2011.09.010
16. Schröttner P, Rudolph WW, Eing BR, Bertram S, Gunzer F. Comparison of VITEK2, MALDI-TOF MS, and 16S rDNA sequencing for identification of Myroides odoratus and Myroides odoratimimus. Diagn Microbiol Infect Dis. 2014;79(2):155–159. doi:10.1016/j.diagmicrobio.2014.02.002
17. Hu S, Jiang T, Zhou Y, Ming D, Gao H, Wang M. Genomic analysis of the multi-drug-resistant clinical isolate Myroides odoratimimus PR63039. Mol Genet Genomics. 2017;292(1):133–144. doi:10.1007/s00438-016-1261-5
18. Yağci A, Cerikçioğlu N, Kaufmann ME, et al. Molecular typing of Myroides odoratimimus (Flavobacterium odoratum) urinary tract infections in a Turkish hospital. Eur J Clin Microbiol Infect Dis. 2000;19(9):731–732. doi:10.1007/s100960070001
19. Licker M, Sorescu T, Rus M, et al. Extensively drug-resistant Myroides odoratimimus – a case series of urinary tract infections in immunocompromised patients. Infect Drug Resist. 2018;11:743–749. doi:10.2147/IDR.S161069
20. Tian Y, Zheng B, Wang B, Lin Y, Li M. Rapid identification and multiple susceptibility testing of pathogens from positive-culture sterile body fluids by a combined MALDI-TOF mass spectrometry and vitek susceptibility system. Front Microbiol. 2016;7:523. doi:10.3389/fmicb.2016.00523
21. Matuschek E, Åhman J, Webster C, Kahlmeter G. Antimicrobial susceptibility testing of colistin - evaluation of seven commercial MIC products against standard broth microdilution for Escherichia coli, Klebsiella pneumoniae, Pseudomonas aeruginosa, and Acinetobacter spp. Clin Microbiol Infect. 2018;24(8):865–870. doi:10.1016/j.cmi.2017.11.020
22. Tenover FC, Arbeit RD, Goering RV, et al. Interpreting chromosomal DNA restriction patterns produced by pulsed-field gel electrophoresis: criteria for bacterial strain typing. J Clin Microbiol. 1995;33(9):2233–2239. doi:10.1128/JCM.33.9.2233-2239.1995
23. Griffin PM, Price GR, Schooneveldt JM, et al. Use of matrix-assisted laser desorption ionization-time of flight mass spectrometry to identify vancomycin-resistant enterococci and investigate the epidemiology of an outbreak. J Clin Microbiol. 2012;50(9):2918–2931. doi:10.1128/JCM.01000-12
24. Wang J, Zhou N, Xu B, et al. Identification and cluster analysis of Streptococcus pyogenes by MALDI-TOF mass spectrometry. PLoS One. 2012;7(11):e47152. doi:10.1371/journal.pone.0047152
25. Poirel L, Walsh TR, Cuvillier V, Nordmann P. Multiplex PCR for detection of acquired carbapenemase genes. Diagn Microbiol Infect Dis. 2011;70(1):119–123. doi:10.1016/j.diagmicrobio.2010.12.002
26. Dallenne C, Da Costa A, Decré D, Favier C, Arlet G. Development of a set of multiplex PCR assays for the detection of genes encoding important β-lactamases in Enterobacteriaceae. J Antimicrob Chemother. 2010;65(3):490–495. doi:10.1093/jac/dkp498
27. Al-Bayssari C, Gupta SK, Dabboussi F, Hamze M, Rolain JM. MUS-2, a novel variant of the chromosome-encoded β-lactamase MUS-1, from Myroides odoratimimus. New Microbes New Infect. 2015;7:67–71. doi:10.1016/j.nmni.2015.06.007
28. Jones DT. Protein secondary structure prediction based on position-specific scoring matrices. J Mol Biol. 1999;292(2):195–202. doi:10.1006/jmbi.1999.3091
29. Källberg M, Wang H, Wang S, et al. Template-based protein structure modeling using the RaptorX web server. Nat Protoc. 2012;7(8):1511–1522. doi:10.1038/nprot.2012.085
30. Bhatnager R, Dang AS. Comprehensive in-silico prediction of damage associated SNPs in Human Prolidase gene. Sci Rep. 2018;8(1):9430. doi:10.1038/s41598-018-27789-0
31. Kinyanyi D, Obiero G, Obiero GFO, Amwayi P, Mwaniki S, Wamalwa M. Structural and functional prediction of African swine fever virus protein-B263R reveals features of a TATA-binding protein. PeerJ. 2018;6:e4396. doi:10.7717/peerj.4396
32. Spanik S, Trupl J, Krcmery V. Nosocomial catheter-associated Flavobacterium odoratum bacteraemia in cancer patients. J Med Microbiol. 1998;47(2):183. doi:10.1099/00222615-47-2-183
33. Gunzer F, Rudolph WW, Bunk B, et al. Whole-genome sequencing of a large collection of Myroides odoratimimus and Myroides odoratus isolates and antimicrobial susceptibility studies. Emerg Microbes Infect. 2018;7(1):61. doi:10.1038/s41426-018-0061-x
34. van Belkum A, Tassios PT, Dijkshoorn L, et al. Guidelines for the validation and application of typing methods for use in bacterial epidemiology. Clin Microbiol Infect. 2007;13(Suppl 3):1–46. doi:10.1111/j.1469-0691.2007.01786.x
35. Gordillo ME, Singh KV, Baker CJ, Murray BE. Typing of group B streptococci: comparison of pulsed-field gel electrophoresis and conventional electrophoresis. J Clin Microbiol. 1993;31(6):1430–1434. doi:10.1128/JCM.31.6.1430-1434.1993
36. van Belkum A. DNA fingerprinting of medically important microorganisms by use of PCR. Clin Microbiol Rev. 1994;7(2):174–184. doi:10.1128/CMR.7.2.174
37. Ruiz-Garbajosa P, Bonten MJ, Robinson DA, et al. Multilocus sequence typing scheme for Enterococcus faecalis reveals hospital-adapted genetic complexes in a background of high rates of recombination. J Clin Microbiol. 2006;44(6):2220–2228. doi:10.1128/JCM.02596-05
38. Maiden MC, Bygraves JA, Feil E, et al. Multilocus sequence typing: a portable approach to the identification of clones within populations of pathogenic microorganisms. Proc Natl Acad Sci U S A. 1998;95(6):3140–3145. doi:10.1073/pnas.95.6.3140
39. Kao RR, Haydon DT, Lycett SJ, Murcia PR. Supersize me: how whole-genome sequencing and big data are transforming epidemiology. Trends Microbiol. 2014;22(5):282–291. doi:10.1016/j.tim.2014.02.011
40. de Been M, Pinholt M, Top J, et al. Core genome multilocus sequence typing scheme for high- resolution typing of Enterococcus faecium. J Clin Microbiol. 2015;53(12):3788–3797. doi:10.1128/JCM.01946-15
41. Dekker JP, Frank KM. Next-generation epidemiology: using real-time core genome multilocus sequence typing to support infection control policy. J Clin Microbiol. 2016;54(12):2850–2853. doi:10.1128/JCM.01714-16
42. Hu S, Cao L, Wu Y, et al. Comparative genomic analysis of Myroides odoratimimus isolates. Microbiologyopen. 2019;8(2):e00634. doi:10.1002/mbo3.634
43. Adler A, Lifshitz Z, Gordon M, et al. Evolution and dissemination of the Klebsiella pneumoniae clonal group 258 throughout Israeli post-acute care hospitals, 2008–13. J Antimicrob Chemother. 2017;72(8):2219–2224. doi:10.1093/jac/dkx135
44. Berrazeg M, Diene SM, Drissi M, et al. Biotyping of multidrug-resistant Klebsiella pneumoniae clinical isolates from France and Algeria using MALDI-TOF MS. PLoS One. 2013;8(4):e61428. doi:10.1371/journal.pone.0061428
45. Lorenzin G, Piccinelli G, Carlassara L, et al. Myroides odoratimimus urinary tract infection in an immunocompromised patient: an emerging multidrug-resistant micro-organism. Antimicrob Resist Infect Control. 2018;7:96. doi:10.1186/s13756-018-0391-4
46. Mammeri H, Bellais S, Nordmann P. Chromosome-encoded β-lactamases TUS-1 and MUS-1 from Myroides odoratus and Myroides odoratimimus (formerly Flavobacterium odoratum), new members of the lineage of molecular subclass B1 metalloenzymes. Antimicrob Agents Chemother. 2002;46(11):3561–3567. doi:10.1128/AAC.46.11.3561-3567.2002
47. Ambler RP. The structure of beta-lactamases. Philos Trans R Soc Lond B Biol Sci. 1980;289(1036):321–331.
48. Hummel AS, Hertel C, Holzapfel WH, Franz CM. Antibiotic resistances of starter and probiotic strains of lactic acid bacteria. Appl Environ Microbiol. 2007;73(3):730–739. doi:10.1128/AEM.02105-06
49. Vaz-Moreira I, Varela AR, Pereira TV, Fochat RC, Manaia CM. Multidrug resistance in quinolone-resistant gram-negative bacteria isolated from hospital effluent and the municipal wastewater treatment plant. Microb Drug Resist. 2016;22(2):155–163. doi:10.1089/mdr.2015.0118
50. Maneerat S, Nitoda T, Kanzaki H, Kawai F. Bile acids are new products of a marine bacterium, Myroides sp. strain SM1. Appl Microbiol Biotechnol. 2005;67(5):679–683. doi:10.1007/s00253-004-1777-1
 © 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.