Back to Journals » Infection and Drug Resistance » Volume 14
Molecular Characterization of Class 1, 2 and 3 Integrons in Serratia spp. Clinical Isolates in Poland – Isolation of a New Plasmid and Identification of a Gene for a Novel Fusion Protein
Authors Celejewski-Marciniak P, Wolinowska R, Wróblewska M
Received 26 June 2021
Accepted for publication 9 September 2021
Published 4 November 2021 Volume 2021:14 Pages 4601—4610
DOI https://doi.org/10.2147/IDR.S325943
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Suresh Antony
Piotr Celejewski-Marciniak,1 Renata Wolinowska,2 Marta Wróblewska1,3
1Department of Dental Microbiology, Medical University of Warsaw, Warsaw, Poland; 2Department of Pharmaceutical Microbiology, Centre for Preclinical Research, Medical University of Warsaw, Warsaw, Poland; 3Department of Microbiology, Central Clinical Hospital, University Clinical Centre, Medical University of Warsaw, Warsaw, Poland
Correspondence: Marta Wróblewska; Piotr Celejewski-Marciniak
Department of Dental Microbiology, Medical University of Warsaw, 1a Banacha street, Warsaw, 02-097, Poland
Tel +48225991777
; +48665101188
Email [email protected]; [email protected]
Purpose: Gram-negative rods of the genus Serratia play an increasing role as etiological agents of healthcare-associated infections (HAI) in humans. These bacteria are characterized by natural and acquired resistance to several groups of antibacterial agents. The aim of the study was to characterize class 1, 2 and 3 integrons in the clinical isolates of Serratia spp. in Poland.
Methods: The study comprised 112 clinical strains of Serratia, isolated from patients hospitalized in Poland in 2010– 2012. Identification of strains was confirmed using MALDI-TOF MS (matrix-assisted laser desorption/ionization time-of-flight mass spectrometry) system. Detection of class 1, 2 and 3 integrase DNA sequence was performed by multiplex-PCR. Amplicons obtained in the PCR reactions were purified and then sequenced bidirectionally.
Results: Among the analyzed strains, Serratia marcescens was a predominant species (103/112, 92.0%). All three classes of integrase DNA sequence were detected in the analyzed strains of Serratia spp. DNA sequence of class 3 integron, besides integrase gene, revealed three gene cassettes (dfrB3, blaGES-7,blaOXA/aac(6ʹ)-Ib-cr). BLAST analysis of DNA sequence revealed that class 3 integron was carried on 9448 bp plasmid which was named pPCMI3 – whole sequence of its DNA was submitted to GenBank NCBI (National Center for Biotechnology Information) – NCBI MH569711.
Conclusion: In this study, we identified a new plasmid pPCMI3 harboring class 3 integron. This is the first report of a gene oxa/aac(6ʹ)-Ib-cr coding for a novel fusion protein, which consists of OXA β-lactamase and acetyltransferase aac(6ʹ)-Ib-cr. In the analyzed strains, class 1 and 2 integrons were also detected. Among the strains with class 1 integron, nine contained cassette array 5ʹCS-aadA2-ORF-dfrA12-3ʹCS, and two – cassette array 5ʹCS-aacC1-ORF-ORF-aadA1-3ʹCS, which were not previously reported in Serratia spp.
Keywords: Serratia marcescens, gene cassettes, antibiotic resistance, carbapenemase, fusion protein
Introduction
Gram-negative rods of the genus Serratia are environmental bacteria which play an increasing role as etiological agents of healthcare-associated infections (HAI). These bacteria were previously classified in the family Enterobacteriaceae, but due to 2016 taxonomy change, the genus Serratia at present belongs to the newly formed family Yersiniaceae in the order Enterobacterales.1
Bacteria Serratia spp. are widely distributed in the external environment, mainly in water and soil, but also in plants and animals.2 Since the mid-1970s, they have been increasingly recognized as human pathogens. At present, 30 species of Serratia are recognized, out of which S. marcescens is the species most important in medicine, causing a wide range of diseases, particularly in neonates and small children as well as in hospitalized and/or immunocompromised adult patients.2–6 Serratia marcescens affects, most often, the urinary tract, skin and soft tissue, the central nervous system (CNS), the eye, or leads to pneumonia or sepsis.2,7,8
Antimicrobial resistance of S. marcescens is mediated by the resistance genes located on the bacterial chromosome and on the mobile genetic elements, eg, plasmids and integrons. A set of mobile genes (so-called “mobiliome”) enables horizontal transfer of genes contributing to the spread of antibiotic resistance between bacteria of the same or even different species.9
Integrons have a wide distribution among bacterial isolates obtained from clinical samples of hospitalized patients. One of the most crucial problems is activity of integrons leading to increased antibiotic resistance in clinical strains.10 Unil now, five classes of integrons have been characterized, based on analysis of the nucleotide sequence of the integrase gene, out of which the first three (class 1, 2 and 3) are involved in spreading the resistance gene cassettes.11
Class 1 integrons are the most prevalent type of integrons and have been frequently reported in clinical isolates of Gram-negative bacteria.12 The structure of class 1 integrons consists of two conserved regions, including 3′ conserved segment (3′ CS) and 5′ conserved segment (5′ CS), as well as internal gene cassettes which encode antimicrobial resistance genes. Class 1 integrons carry a variety of resistance gene cassettes, and most of them contain one of aadA genes, which confers resistance to streptomycin and spectinomycin. On Integrall website, which provides easy access to integron DNA sequences and genetic arrangements, 66 unique sequences obtained from Serratia spp. were submitted (date of release 05.08.2021).13 Most of them belong to class 1 integrons, there are two sequences of integrons class 3 and one of class 2. Among 95 cassettes carrying recognized antibiotic resistance genes within class 1 integrons, 47 (49.5%) contain resistance to aminoglycosides, 29 (30.1%) resistance to β-lactams, and 10 (10.5%) resistance to trimethoprim.
Class 2 integrons are less common in Enterobacterales and are less diverse.14 They are associated with transposons which belong to Tn7 family.15 Crowley et al reported class 2 integrons in 9/30 (30.0%) clinical strains of S. marcescens, which contained the gene cassettes dfrA1, sat1 and aadA1, conferring resistance to trimethoprim, streptothricin, and streptomycin/spectinomycin, respectively.16
Class 3 integrons are rare and had been discovered only in a few species, including S. marcescens.9 Arakawa et al isolated class 3 integron from a carbapenem-resistant Serratia marcescens strain.17 Its module was characterized by Collis et al as consisting of the intI3 gene, attI3 site, and Pc promoter – in the same way as the class 1 integron module.18 Correia et al reported that an intI3 integrase gene contained in class 3 integron isolated from a strain of Klebsiella pneumoniae showed 98.8% identity to that of Serratia marcescens AK9373.19
The presence of integrons in S. marcescens isolated from clinical specimens in Poland has been documented in only one study.20 Thus, the aim of the present study was to characterize class 1, 2 and 3 integrons in the clinical isolates of Serratia spp. in Poland.
Materials and Methods
Collection of Clinical Isolates of Serratia spp.
The study comprised a total of 112 strains, which were isolated from patients hospitalized in 6 hospitals in two cities in Poland (Warsaw and Otwock) in 2010–2012. All isolates were initially tested by Vitek-2 Compact system, using GN-ID card dedicated to identification of Gram-negative bacteria. Antimicrobial susceptibility testing was performed simultaneously using AST-N-259 card for evaluation of MIC (minimal inhibitory concentration) value of 16 different antibiotics and other chemotherapeutics, including β-lactams, aminoglycosides, fluoroquinolones, tetracyclines, colistin and trimethoprim/sulfamethoxazole. MALDI-TOF MS was performed (MALDI Biotyper, Bruker) as a conclusive method for identification of Serratia spp. strains.21
DNA Extraction
Extraction of genomic DNA was performed by Genomic Mini kit (A&A Biotechnology) following the manufacturer’s instructions. The quantity of DNA was analyzed by gel electrophoresis and compared with GeneRuler DNA Ladder MIX (Thermo Fisher Scientific).
Detection of Integrase DNA Sequence
Detection of class 1, 2 and 3 integrase DNA sequence was performed by multiplex-PCR (polymerase chain reaction) in accordance with the protocol published by Su et al.22 DNA sequences of 6 primers used for investigation are shown in Table 1.
 |
Table 1 Primers Used for Detection of Integrase DNA Sequence22 |
DNA amplification was performed in DNA thermal cycler (S1000 Thermal Cycler, BIO-RAD) in a final volume of 50 µL, containing 5 µL of genomic DNA (500 ng) extract mixed with 5 µL of 10X Taq buffer (with ammonium sulfate) (Thermo Fisher Scientific), 5 µL of MgCl2 25 mM solution (Thermo Fisher Scientific), 1 µL of dNTPs 10 mM solution (Thermo Fisher Scientific), adequate volume of aqueous solution of primers (final concentration of each primer should amount to 40 pmol per reaction), 0.5 µL Taq DNA Polymerase (Thermo Fisher Scientific), deionized water up to 50 µL (Direct Q-3 UV Millipore). The conditions of amplification reaction were as follows: initial denaturation at 95°C for 3 minutes, followed by 30 cycles of DNA denaturation at 95°C for 30 seconds, primers annealing at 58°C for 1 minute, primer extension at 72°C for 1 minute and a final elongation at 72°C for 5 minutes. Positive and negative controls were included in PCR assays and Gene Ruler DNA Ladder MIX (Thermo Fisher Scientific) was used as a molecular size standard. PCR products were separated by DNA electrophoresis on 3% NuSieve 3:1 agarose gel (Lonza) in 1X TAE buffer, stained with ethidium bromide and visualized by VersaDoc Imaging System version 1000 (BIO-RAD).
Amplification of Variable Regions of Class 1 and 2 Integrons
Amplification of the variable region of class 1 and 2 integrons was performed by PCR reaction only for isolates known to contain respective 1 and 2 integrase sequences. DNA sequences of primers are shown in Table 2.
 |
Table 2 Primers Used for Amplification of the Variable Region of Class 1 and 2 Integron DNA Sequence |
DNA amplification was performed in DNA thermal cycler (S1000 Thermal Cycler, BIO-RAD) in a final volume of 50 µL, containing 1 µL of genomic DNA (1 ng) extract (100X diluted in deionized water) mixed with 10 µL of 5X iProof buffer (BIO-RAD), 1 µL of dNTPs 10 mM solution (Thermo Fisher Scientific), adequate volume of aqueous solution of primers (final concentration of each primer should amount to 300 pmol per reaction), 0.5 µL iProof High-Fidelity DNA Polymerase (BIO-RAD), deionized water up to 50 µL (Direct Q-3 UV Millipore). The conditions of the amplification reaction were as follows: initial denaturation at 98°C for 1 minute, followed by 35 cycles of DNA denaturation at 98°C for 30 seconds, primers annealing (temperature in accordance to data shown in Table 2) for 15 seconds, primer extension at 72°C for 2 minutes and a final elongation at 72°C for 10 minutes. Positive and negative controls were included in PCR assays, Gene Ruler DNA Ladder MIX (Thermo Fisher Scientific) was used as a molecular weight standard. PCR products were separated by DNA electrophoresis on 1% agarose gel (Sigma Aldrich) in 1X TAE buffer, stained with ethidium bromide and visualized by VersaDoc Imaging System version 1000 (BIO-RAD).
Sequencing of PCR Products and Sequence Analysis
Amplicons obtained in the PCR reactions before sequencing were purified by ChargeSwitch PCR Clean-UP kit (Thermo Fisher Scientific) in accordance with the manufacturer’s instructions. Purified PCR products were sequenced bidirectionally by the Laboratory of DNA Sequencing and Oligonucleotide Synthesis in the Institute of Biochemistry and Biophysics of the Polish Academy of Sciences using sequencer 373/x1 DNA Analyzer (Applied Biosystems). The chromatogram analysis and alignment were performed by Clone Manager. Homology analysis was carried out using the BLAST program provided by the National Center for Biotechnology Information (http://www.ncbi.nlm.nih.gov).
Sequencing of Plasmid Carrying Class 3 Integron by Next-Generation Sequencing (NGS)
Genomic DNA of strain carrying class 3 integrase gene was sequenced on the Genome Sequencer GS FLX Titanium. The results of sequencing were matched by Newbler program. Further work for determination of the final DNA sequence was performed by Seqman program (DNAStar).
Results
Identification and Antimicrobial Susceptibility
Among 112 tested strains, more than 90% (103 isolates) were identified as S. marcescens, 7 strains as S. liquefaciens, 1 as S. fonticola, and 1 as S. ureilytica. Summary of the antimicrobial susceptibility patterns of the analyzed strains is shown in Figure 1. Complete results of identification and antimicrobial susceptibility tests are shown in the Appendix A attached to this article.
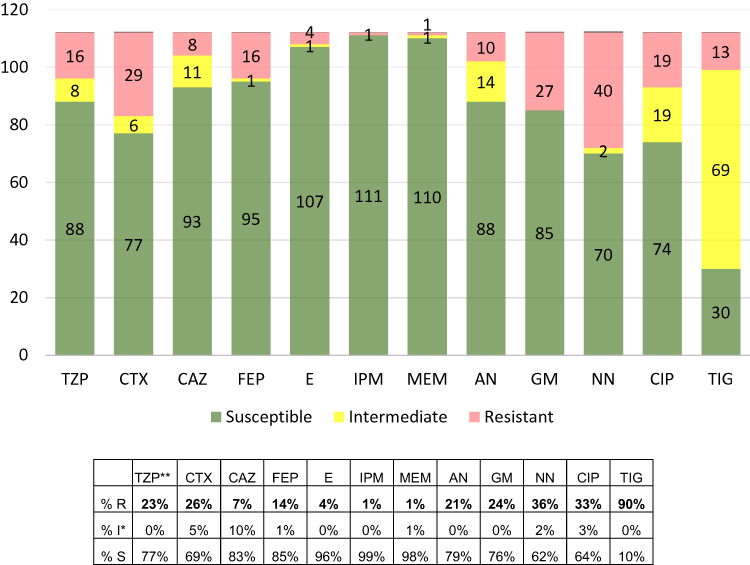 |
Figure 1 The antibiotic resistance patterns of Serratia spp. (n=112) measured by MIC value according to the EUCAST (European Committee on Antimicrobial Susceptibility Testing) recommendations.22 Abbreviations: TZP, piperacillin/tazobactam; CTX, cefotaxime; CAZ, ceftazidime; FEP, cefepime; E, ertapenem; IPM, imipenem; MEM, meropenem; AN, amikacin; GM, gentamicin; NN, tobramycin; CIP, ciprofloxacin; TIG, tigecycline. Notes: *According to the decision of the EUCAST in July 2019 (implemented in January 2020) at present letter “I” means “susceptible with increased exposure”. **n=104. |
Detection of Class 1, 2 and 3 Integrase
In the strains analyzed in this study, the presence of all three classes of integrase was confirmed. Class 1 was the most common type present in 26 isolates, which represented more than 23% of all tested strains. Class 2 was confirmed in 10 strains, 9 of them simultaneously carried class 1 integrase DNA sequence. Just one strain contained a single class 2 integrase DNA sequence. Only one isolate carried class 3 integrase DNA sequence. Results are shown in the attached Appendix A.
Characterization of Variable Regions of Class 1 and 2 Integrons
Variable regions of class 1 and 2 integrons were amplified by iProof polymerase which has a high thermostability and ability for fast synthesis of long DNA products of PCR reactions. Compilation of gene cassettes characterized in tested strains of Serratia spp. is shown in Table 3. Samples no. 64, 91, 96 and 99 did not show any products of PCR reactions.
 |
Table 3 Compilation of Gene Cassettes in Class 1 Integrons |
All 10 isolates containing class 2 integrons were identical and they had the same gene cassette arrangement: aadA1, sat2 and dfrA1 (Figure 2), which are responsible for resistance to aminoglycosides, streptomycin and trimethoprim, respectively.
 |
Figure 2 Diagram of integrons. Blue arrows mark open reading frames. Green boxes mark regulatory items: Pc- promotor and attI2 or attI3 – attachment site. (A) Class 2 integron: integrase (IntI2 – 977 bp) and antibiotic resistance gene cassettes (aadA1 – 852 bp, Sat2 – 583 bp, dfrA1– 473 bp). (B) Class 3 integron: integrase (IntI3 – 1040 bp) and antibiotic resistance gene cassettes (dfrB3 – 236 bp, blaGES-7 – 863 bp, blaOXA/aac(6ʹ)-Ib-cr – 554 bp). |
Characterization of Class 3 Integron
Strain 89 was of particular interest due to the presence of class 3 integron, therefore a search for plasmid DNA carrying the integron was performed. Genomic DNA of strain 89 was used as a template in NGS sequencing. BLAST analysis of DNA sequence revealed that class 3 integron is carried on 9448 bp plasmid which has been named pPCMI3. Whole DNA sequence of plasmid pPCMI3 was submitted to GenBank NCBI (NCBI MH569711).
DNA sequence of class 3 integron, besides integrase gene, harbors three gene cassettes (dfrB3, blaGES-7, blaOXA/aac(6ʹ)-Ib-cr) (Figure 2). The aac(6′)-Ib-cr is a variant of aac(6ʹ)-Ib with two amino acid substitutions (Trp87Arg and Asp164Tyr) allowing it to acetylate and reduce the activity of ciprofloxacin.23
Discussion
Gram-negative rods of the genus Serratia are members of the order Enterobacterales, the significance of which, in medicine remains underestimated. The most important species within the genus Serratia is S. marcescens, due to a high frequency of isolations from humans and resistance to many antimicrobial agents. Nowadays, S. marcescens is unquestionably regarded as an etiological agent of opportunistic infections in humans, particularly in neonates and small children, but also in severely immunocompromised adults.2–5
Samonis et al showed that more than 88% of Serratia spp. strains isolated from patients in Cretan hospitals between 2010 and 2015 were identified as S. marcescens.24 In our study as many as 90% of analyzed strains of the genus Serratia were identified as S. marcescens.
Almost 29% of strains tested in this study (32 isolates) were recovered from the lower respiratory tract, followed by 26 (23.2%) isolates from wounds, and 25 (22.3%) strains recovered from urine. Ferreira et al reported that respiratory tract was the major source of S. marcescens isolates from patients hospitalized in intensive care units.25 Analysis of over 18,000 of the clinical isolates previously classified in the family Enterobacteriaceae and Pseudomonas aeruginosa, collected from 85 US hospitals, showed domination of strains recovered from urine (34.5%), followed by the respiratory tract (28%), and isolates from wounds (18%).26 Liou et al analyzed 403 nonduplicate S. marcescens strains – isolates from respiratory samples comprised 39.0%, followed by the urinary tract samples (22.3%).27 These differences should be ascribed to various profiles of hospital wards that were the sources of analyzed strains.
According to EUCAST (European Committee on Antimicrobial Susceptibility Testing) criteria (version no. 11), all tested isolates of Serratia spp. were reported as resistant to colistin, despite the fact that strains nr 31, 51, 78, 98, 107, 108, 109, 114 and 115 examined in this study showed the MIC value meeting the criteria of “susceptible”.28 Strains of Serratia spp. are characterized by intrinsic resistance to amoxicillin/clavulanic acid, first and second generation cephalosporins due to the presence of a chromosomal AmpC beta-lactamase enzyme. These bacteria are also intrinsically resistant to colistin through the genes (eg, arnBCADTEF) that encode the LPS (lipopolysaccharide)-modifying enzymes (addition of pEtN and L-Ara4N to LPS).29 Therefore, regardless of the MIC value of mentioned antibiotics, all Serratia spp. strains have to be reported as “resistant” to these antibiotics because of the lack of clinical effect.
Among third generation cephalosporins, cefotaxime was the least effective against Serratia spp. strains. Less than 70% of tested strains were susceptible to this antibiotic in contrast to ceftazidime (83% of susceptible strains) and cefepime (85% of susceptible strains), which were the most effective cephalosporins (in our study the MIC values of carbapenems showed that only 5 (4%) strains were not susceptible to ertapenem), 1 (1%) to meropenem, and 1 (1%) to imipenem (Figure 1). Liou et al revealed that only 3 from 403 strains (0.7%) isolated in Taiwan between 2002–2010 were resistant to carbapenems.27 Number of strains resistant to amikacin is convergent with data from Polish hospitals where around 10% of tested strains were resistant to amikacin.30,31 Of note, in our study 33% of strains were characterized by resistance to ciprofloxacin. Similar rate (39.6% of resistant strains) was reported by Liou et al and these authors postulated the need for continued surveillance of antimicrobial resistance of Serratia spp. clinical isolates, particularly for fluoroquinolones.27 Hornsey et al reported that in therapy of infections caused by S. marcescens, the use of tigecycline is limited because of up-regulation of the SdeXY-HasF efflux pump which is associated with reduced susceptibility to this antibiotic and also to ciprofloxacin.32 In our study 90% of strains of Serratia spp. were resistant to tigecycline. In a study by Magiorakos et al, 21 isolates (19%) were resistant to at least 3 different groups of antibiotics and chemotherapeutics and were reported as multidrug-resistant (MDR) S. marcescens strains.33 Most of them (19 strains) harbored at least one integrase DNA sequence.
Among 26 tested strains carrying the class 1 integrase DNA sequence, for 22 strains PCR products were obtained as a result of amplification of the variable region of class 1 integrons. All of them had at least one aminoglycoside resistance gene cassette, and all (except strain no. 41) showed resistance to gentamicin and tobramycin (Appendix A).
In this study we detected four different cassettes containing resistance genes to aminoglycosides, only one cassette with a resistance gene to β-lactams and one cassette with resistance genes to trimethoprim (Table 3). Among 39 cassettes containing genes with a known function, 28 (71.8%) comprised cassettes determining resistance to aminoglycosides, 9 (23.0%) carried genes of resistance to trimethoprim, and only 2 (5.1%) were cassettes with the genes of resistance to β-lactams. We observed similar proportions of cassettes as reported by Xia et al who analyzed different types of gene cassette arrays of integrons from clinical bacterial isolates in China, 2000–2014.34
Nielsen et al analyzed coexistence of gene cassettes linked to bacterial resistance to antimicrobials and integrase class 1 sequence in almost 16,000 nucleotide sequences of bacterial genomes.35 These authors showed that the most common mechanism of resistance linked to integron class 1 were gene cassettes coding for aminoglycoside nucleotidyltransferase.35 Among 22 variable regions of integrons class 1 characterized in this study, only three (no. 16, 18 and 44) did not contain a gene cassette coding for aminoglycoside nucleotidyltransferase. Ten strains carried cassette aadA1, and nine – cassette aadA2.
Among the analyzed strains of Serratia spp., there were 5 which carried only a single cassette (aadA1), 2 strains – carried two cassettes 5ʹCS-aac(6ʹ)-Ib-blavim-4-3ʹCS, 8 strains had three cassettes 5ʹCS-aadA2-ORF-dfr12-3ʹCS, and 5 strains – carried 5ʹCS-aacC1-ORF-ORF-aadA1-3ʹCS. It is often reported that cassette aac(6ʹ)-Ib is the first or the only one in the array. It is also typical for cassette aadA1 to be the lone one, or the last in the array.36
Cassette array 5ʹCS-aadA2-ORF-dfr12-3ʹCS is atypical, not reported in Serratia spp. so far, however it was reported in K. pneumoniae strains isolated in Russia [KY862013.1] and China [DQ323053]. Similarly, cassette array 5ʹCS-aacC1-ORF-ORF-aadA1-3ʹCS is rarely detected – it was isolated in Russia [KR028107.1], Ireland [AJ784787] and Australia [EF015496], solely in the strains of Acinetobacter baumannii.
Two strains (number 16 and 18) carried two identical gene cassettes encoding metallo-β-lactamase blaVIM-4 and aminoglycoside 6ʹ-N-acetyltransferase aac(6ʹ)-Ib (Table 3). BLAST analysis of these two sequences revealed their identity with integron sequences found in Hungarian strains of S. marcescens [NCBI JF905459.1], K. pneumoniae and K. oxytoca.37 In Poland, VIM-4 carbapenemase was originally observed in P. aeruginosa strains isolated from hospitalized children.38 Comparison of amino acid sequences of integrons no. 16 and 18 with VIM-4 from P. aeruginosa revealed a single substitution in the repeat region. Valine present in protein of P. aeruginosa was substituted by glycine in S. marcescens isolates no. 16 and 18 (Val168Gly). Between 2006 and 2012 Izdebski et al identified 20 S. marcescens strains producing VIM/IMP-carbapenemase, including 11 isolates carrying blaVIM-4 gene cassette.20 All of them possess In238 variant of class 1 integron containing the following array of gene cassettes: 5ʹCS-aacA4-blaVIM-4-3ʹCS which are identical to DNA sequence of integrons no. 16 and 18. We can speculate with high probability that Izdebski analyzed isolates identical to no. 16 and 18, but no sufficient data (antimicrobial resistance, source of strains) are available to conclude it definitely.
In view of the nonsense mutation in 179 codon Int2 sequence, class 2 integrons are less varied than class 1 integrons.14 Isolates no. 16, 18, 22, 27, 28, 43, 54, 61, 66 and 85 had identical nucleotide sequence of gene cassettes located in class 2 integrons (Figure 2). Separate analysis focusing on occurrence and characteristics of class 2 integrons was not conducted on Polish clinical isolates. Kotlarska et al analyzed E. coli isolated from wastewater and reported that three of four analyzed class 2 integrons were identical to integrons investigated in this paper.39
Nowadays class 3 integrons undergo a dynamic worldwide expansion, however it could not be explored quickly by PCR reaction due to the lack of 3ʹ conserved region in DNA sequence. For S. marcescens strain no. 89 with confirmed presence of DNA sequence of class 3 integrase, plasmid DNA sequencing was performed because previously class 3 integrons were reported throughout plasmid DNA.17,40–42 The sequence of pPCMI3 plasmid, which is 9448 bp in size, was submitted to NCBI GenBank (NCBI MH569711).
Plasmid pPCMI3 harbors class 3 integron with three gene cassettes. The first of these, dfrB3, which is responsible for resistance to trimethoprim,43 and the second gene cassette encoding β-lactamase GES-7, had been already described in the DNA sequences of integrons.44–46 The last one is a gene cassette coding the fusion protein OXA/aac(6ʹ)-Ib-cr, it consists of β-lactamase OXA responsible for resistance to β-lactams and acetyltransferase aac(6ʹ)-Ib-cr responsible for resistance to aminoglycosides and fluoroquinolones. Gene aac(6ʹ)-Ib-cr has been detected in many clinical strains of bacteria.47,48 Fusion oxa/aac(6ʹ)-Ib has also been described in the structure of plasmid classified in the group IncQ carrying integron class 3.49 Whereas fusion oxa/aac(6ʹ)-Ib-cr was detected for the first time in this study.
Piotrowska et al developed molecular characterization and comparative genomics of IncQ-3 plasmids isolated from raw wastewater of “Czajka” Wastewater Treatment Plant in Warsaw, Poland.45 Plasmid p115_p2 hosted in Raoultella sp. and isolated from raw wastewater has identical size as plasmid pPCMI3 and, moreover, BLAST analysis revealed high level homology (more than 99.9%) between them. The occurrence of IntI3 in pPCMI3, an IncQ plasmid, provides possibilities for widespread dissemination, an ability to replicate in diverse hosts and carrying of diverse antibiotic resistance genes conferring resistance to different antibiotics and chemotherapeutics, eg, carbapenems, quinolones, β-lactams, sulphonamides, and tetracyclines.50
In conclusion, although strains of Serratia spp. are opportunistic pathogens which cause nosocomial infections, they still remain outside the mainstream interest of scientists. The results obtained in this study constitute a comprehensive analysis of the integrons in the clinical isolates of Serratia spp. in Poland. Among tested isolates of Serratia spp., 28 (25%) harbored at least one integron DNA sequence with antimicrobial resistance gene cassettes. Carbapenems remain a group of antibiotics active against the the highest number of tested isolates, however their use should be restricted to avoid emergence of resistance. These observations indicate the need for ongoing research to monitor the prevalence of integrons in the clinical isolates of Serratia spp.
Ethics Statement
The clinical samples were part of the routine hospital laboratory procedure and therefore approval of the institutional review board or ethics committee was not applicable.
Funding
The study was supported by the grant funded by the National Science Centre of Poland. (No. DEC-2011/01/N/NZ7/05336).
Disclosure
We declare that there are no conflicts of interest associated with this study.
References
1. Adeolu M, Alnajar S, Naushad S, et al. Genome-based phylogeny and taxonomy of the ‘Enterobacteriales’: proposal for Enterobacterales ord. nov. divided into the families Enterobacteriaceae, Erwiniaceae fam. nov., Pectobacteriaceae fam. nov., Yersiniaceae fam. nov., Hafniaceae fam. nov., Morganellaceae fam. nov., and Budviciaceae fam. nov. Int J Syst Evol Microbiol. 2016;66:5575–5599.
2. Moehring R, Mahlen S. Infections due to Serratia species; 2020. Available from: www.uptodate.com/contents/infections-due-to-serratia-species.
3. Morillo A, Gonzalez V, Aguayo J, et al. A six-month Serratia marcescens outbreak in a neonatal intensive care unit. Enferm Infect Microbiol Clin. 2016;34:645–651. doi:10.1016/j.eimc.2016.01.006
4. Attman E, Korhonen P, Tammela O, et al. A Serratia marcescens outbreak in a neonatal intensive care unit was successfully managed by rapid hospital hygiene interventions and screening. Acta Paediatr. 2018;107:425–429. doi:10.1111/apa.14132
5. Escribano E, Saralegui C, Moles L, et al. Influence of a Serratia marcescens outbreak on the gut microbiota establishment process in low-weight preterm neonates. PLoS One. 2019;14(5):e0216581. doi:10.1371/journal.pone.0216581
6. List of prokaryotic names with standing in nomenclature (LPSN). Genus Serratia. Available from: https://lpsn.dsmz.de/genus/serratia.
7. Khanna A, Khanna M, Aggarwal A. Serratia marcescens – a rare opportunistic nosocomial pathogen and measures to limit its spread in hospitalized patients. J Clin Diagn Res. 2013;7(2):243–246.
8. Kim EJ, Park WB, Yoon J-K, et al. Outbreak investigation of Serratia marcescens neurosurgical site infections associated with a contaminated shaving razors. Antimicrob Res Infect Control. 2020;9:64. doi:10.1186/s13756-020-00725-6
9. Sandner-Miranda L, Vinuesa P, Cravioto A, et al. The genomic basis of intrinsic and acquired antibiotic resistance in the genus Serratia. Front Microbiol. 2018;9:828. doi:10.3389/fmicb.2018.00828
10. Deng Y, Bao X, Ji L, et al. Resistance integrons: class 1, 2 and 3. integrons. Ann Clin Microbiol Antimicrob. 2015;14:45. doi:10.1186/s12941-015-0100-6
11. Kaushik M, Kumar S, Kapoor RK, et al. Integrons in Enterobacteriaceae: diversity, distribution and epidemiology. Int J Antimicrob Agents. 2018;51:167–176. doi:10.1016/j.ijantimicag.2017.10.004
12. Pormohammad A, Pouriran R, Azimi H, et al. Prevalence of integron classes in Gram-negative clinical isolated bacteria in Iran: a systematic review and meta-analysis. Iran J Basic Med Sci. 2019;22:118–127.
13. Integrall. Available from: http://integrall.bio.ua.pt.
14. Mazel D. Integrons: agents of bacterial evolution. Nat Rev Microbiol. 2006;4:608–620. doi:10.1038/nrmicro1462
15. White PA, McIver CJ, Rawlinson WD. Integrons and gene cassettes in the Enterobacteriaceae. Antimicrob Agents Chemother. 2001;45:2658–2661. doi:10.1128/AAC.45.9.2658-2661.2001
16. Crowley D, Cryan B, Lucey B. First detection of a class 2 integron among clinical isolates of Serratia marcescens. Br J Biomed Sci. 2008;65(2):86–89. doi:10.1080/09674845.2008.11732803
17. Arakawa Y, Murakami M, Suzuki K, et al. A novel integron-like element carrying the metallo-β-lactamase gene blaIMP. Antimicrob Agents Chemother. 1995;39:1612–1615. doi:10.1128/AAC.39.7.1612
18. Collis CM, Kim M-J, Partridge SR, Stokes HW, Hall RM. Characterization of the class 3 integron and the site-specific recombination system it determines. J Bacteriol. 2002;184:3017–3026. doi:10.1128/JB.184.11.3017-3026.2002
19. Correia M, Boavida F, Grosso F, et al. Molecular characterization of a new class 3 integron in Klebsiella pneumoniae. Antimicrob Agents Chemother. 2003;47:2838–2843. doi:10.1128/AAC.47.9.2838-2843.2003
20. Izdebski R, Baraniak A, Żabicka D, et al. VIM/IMP carbapenemase-producing Enterobacteriaceae in Poland: epidemic Enterobacter hormaechei and Klebsiella oxytoca lineages. J Antimicrob Chemother. 2018;73:2675–2681. doi:10.1093/jac/dky257
21. Singhal N, Kumar M, Kanaujia PK, et al. MALDI-TOF mass spectrometry: an emerging technology for microbial identification and diagnosis. Front Microbiol. 2015;6:791. doi:10.3389/fmicb.2015.00791
22. Su J, Shi L, Yang L, et al. Analysis of integrons in clinical isolates of Escherichia coli in China during the last six years. FEMS Microbiol Lett. 2006;254:75–80. doi:10.1111/j.1574-6968.2005.00025.x
23. Robicsek A, Strahilevitz J, Jacoby GA, et al. Fluoroquinolone-modifying enzyme: a new adaptation of a common aminoglycoside acetyltransferase. Nat Med. 2006;12:83–88. doi:10.1038/nm1347
24. Samonis G, Vardakas KZ, Maraki S, et al. Resistance phenotypes and susceptibility of contemporary Serratia isolates in the university hospital of Crete, Greece. Infect Dis. 2017;49:847–853. doi:10.1080/23744235.2017.1361546
25. Ferreira RL, Rezende GS, Damas MSF, et al. Characterization of KPC-producing Serratia marcescens in an intensive care unit of a Brazilian tertiary hospital. Front Microbiol. 2020;11:956. doi:10.3389/fmicb.2020.00956
26. Sader HS, Castanheira M, Duncan LR, et al. Antimicrobial susceptibility of Enterobacteriaceae and Pseudomonas aeruginosa isolates from United States medical centers stratified by infection type: results from the International Network for Optimal Resistance Monitoring (NFORM) surveillance program, 2015–2016. Diagn Microbiol Infect Dis. 2018;92:69–74.
27. Liou BH, Duh RW, Lin YT, et al. Taiwan Surveillance of Antimicrobial Resistance (TSAR) Hospitals. A multicenter surveillance of antimicrobial resistance in Serratia marcescens in Taiwan. J Microbiol Immunol Infect. 2014;47:387–393. doi:10.1016/j.jmii.2013.04.003
28. European Committee on Antimicrobial Susceptibility Testing (EUCAST). Clinical breakpoints – bacteria v 11.0. Available from: https://eucast.org/ast_of_bacteria.
29. El-Sayed Ahmed MAE, Zhong LL, Shen C, Yang Y, Doi Y, Tian GB. Colistin and its role in the era of antibiotic resistance: an extended review (2000–2019). Emerg Microbes Infect. 2020;9(1):868–885. doi:10.1080/22221751.2020.1754133
30. Dybowski BA, Zapała P, Bres-Niewada E, et al. Catheter-associated bacterial flora in patients with benign prostatic hyperplasia: shift in antimicrobial susceptibility pattern. BMC Infect Dis. 2018;18:1–7. doi:10.1186/s12879-018-3507-9
31. Michno M, Sydor A, Wałaszek M, et al. Microbiology and drug resistance of pathogens in patients hospitalized at the nephrology department in the south of Poland. Pol J Microbiol. 2018;67:517–524. doi:10.21307/pjm-2018-061
32. Hornsey M, Ellington MJ, Doumith M, Hudson S, Livermore DM, Woodford N. Tigecycline resistance in Serratia marcescens associated with up-regulation of the SdeXY-HasF efflux system also active against ciprofloxacin and cefpirome. J Antimicrob Chemother. 2010;65:479–482. doi:10.1093/jac/dkp475
33. Magiorakos AP, Srinivasan A, Carey RB, et al. Multidrug-resistant, extensively drug-resistant and pandrug-resistant bacteria: an international expert proposal for interim standard definitions for acquired resistance. Clin Microbiol Infect. 2011;18:268–281. doi:10.1111/j.1469-0691.2011.03570.x
34. Xia W, Xu T, Qin T, et al. Characterization of integrons and novel cassette arrays in bacteria from clinical isolates in China, 2000–2014. J Biomed Res. 2016;30(4):292–303.
35. Nielsen TK, Browne PD, Hansen LH. Mobilization of antibiotic resistance genes differ by resistance mechanism. bioRxiv. 2021. doi:10.1101/2021.01.10.426126v1
36. Partridge SR, Kwong SM, Firth N, Jensen SO. Mobile genetic elements associated with antimicrobial resistance. Clin Microbiol Rev. 2018;31(4):e00088–17. doi:10.1128/CMR.00088-17
37. Kristof K, Toth A, Damjanowa I, et al. Identification of a blaVIM-4 gene in the internationally successful Klebsiella pneumoniae ST11 clone and in a Klebsiella oxytoca strain in Hungary. J Antimicrob Chemother. 2010;65:1303–1305. doi:10.1093/jac/dkq133
38. Patzer J, Toleman MA, Lalitagauri MD, et al. Pseudomonas aeruginosa strains harbouring an unusual blaVIM-4 gene cassette isolated from hospitalized children in Poland (1998–2001). J Antimicrob Chemother. 2004;53:451–456. doi:10.1093/jac/dkh095
39. Kotlarska E, Luczkiewicz A, Pisowacka M, et al. Antibiotic resistance and prevalence of class 1 and 2 integrons in Escherichia coli isolated from two wastewater treatment plants, and their receiving waters (Gulf of Gdansk, Baltic Sea, Poland). Environ Sci Pollut Res Int. 2015;22:2018–2030. doi:10.1007/s11356-014-3474-7
40. Poirel L, Naas T, Le Thomas I, et al. CTX-M-type extended-spectrum β-lactamase that hydrolyzes ceftazidime through a single amino acid substitution in the omega loop. Antimicrob Agents Chemother. 2001;45:3355–3361. doi:10.1128/AAC.45.12.3355-3361.2001
41. Barraud O, Casellas M, Dagot C, et al. An antibiotic-resistant class 3 integron in an Enterobacter cloacae isolate from hospital effluent. Clin Microbiol Infect. 2013;19:306–308. doi:10.1111/1469-0691.12186
42. Tchuinte PLS, Stalder T, Venditti S, et al. Characterisation of class 3 integrons with oxacillinase gene cassettes in hospital sewage and sludge samples from France and Luxembourg. Int J Antimicrob Agents. 2016;48:431–434. doi:10.1016/j.ijantimicag.2016.06.018
43. Huovinien P, Huovinen P. Resistance to trimethoprim-sulfamethoxazole. Clin Infect Dis. 2001;32:1608–1614. doi:10.1086/320532
44. Kotsakis SD, Miriagou V, Tzelepi E, et al. Comparative biochemical and computational study of the role of naturally occurring mutations at Ambler positions 104 and 170 in GES β-lactamases. Antimicrob Agents Chemother. 2010;54:4864–4871. doi:10.1128/AAC.00771-10
45. Piotrowska M, Dziewit L, Ostrowski R, et al. Molecular characterization and comparative genomics of IncQ-3 plasmids conferring resistance to various antibiotics isolated from a wastewater treatment plant in Warsaw (Poland). Antibiotics. 2020;9:613. doi:10.3390/antibiotics9090613
46. Jones-Dias D, Manageiro V, Ferreira E, et al. Architecture of class 1, 2, and 3 integrons from Gram negative bacteria recovered among fruits and vegetables. Front Microbiol. 2016;7:1400. doi:10.3389/fmicb.2016.01400
47. Jiang Y, Zhou Z, Qian Y, et al. Plasmid-mediated quinolone resistance determinants qnr and aac(6′)-Ib-cr in extended-spectrum β-lactamase-producing Escherichia coli and Klebsiella pneumoniae in China. J Antimicrob Chemother. 2008;61(5):1003–1006. doi:10.1093/jac/dkn063
48. Luque-González P, López-Cerero L, Díaz-de-alba P, Rodríguez-Martínez JM. Association of blaOXA-1, and aac(6ʹ)-Ib-cr with ST405 K. pneumoniae clone. Enferm Infecc Microbiol Clin. 2019;37(6):417–418. doi:10.1016/j.eimc.2018.06.001
49. Poirel L, Carattoli A, Bernabeu S, Bruderer T, Frei R, Nordmann P. A novel IncQ plasmid type harbouring a class 3 integron from Escherichia coli. J Antimicrob Chemother. 2010;65(8):1594–1598. doi:10.1093/jac/dkq166
50. Bonemann G, Stiens M, Puhler A, et al. Mobilizable IncQ-related plasmid carrying a new quinolone resistance gene, qnrS2, isolated from the bacterial community of a wastewater treatment plant. Antimicrob Agents Chemother. 2006;50:3075–3080. doi:10.1128/AAC.00378-06
 © 2021 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2021 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.